Embed Size (px)
Citation preview

A Comparison of Leaf Physiology and Anatomy of Quercus (Section Erythrobalanus-Fagaceae)Species in Different Light EnvironmentsAuthor(s): P. M. S. Ashton and G. P. BerlynReviewed work(s):Source: American Journal of Botany, Vol. 81, No. 5 (May, 1994), pp. 589-597Published by: Botanical Society of AmericaStable URL: http://www.jstor.org/stable/2445734 .Accessed: 24/09/2012 19:48
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Botanical Society of America is collaborating with JSTOR to digitize, preserve and extend access to AmericanJournal of Botany.
http://www.jstor.org

American Journal of Botany 8 1(5): 589-597. 1994.
A COMPARISON OF LEAF PHYSIOLOGY AND ANATOMY OF Q UERCUS (SECTION ER YTHROBALANUS- FAGACEAE)
SPECIES IN DIFFERENT LIGHT ENVIRONMENTS'
P. M. S. ASHTON2 AND G. P. BERLYN School of Forestry and Environmental Studies, Yale University, New Haven, Connecticut 06511
Physiological and anatomical attribut-es of leaves were examined of three species of Quercus section Erythrobalanus. All three species occur in moist temperate deciduous forests of eastern North America. Seedlings of each species were grown in different light conditions for comparison. The attributes measured were net photosynthesis, stomatal conductivity, blade and cuticle thickness, stomatal density, thickness of upper and lower epidermis, and thickness of palisade mesophyll. The results generally demonstrate the close association between anatomical adaptations and efficiency of physiological processes; they also elucidate the distribution patterns of the three Quercus species across the forest topography in southern New England. The most drought-tolerant and light-demanding species, Q. velutina (Lam.), exhibited the greatest leaf anatomical plasticity, the highest net photosynthesis in the different light conditions, and the lowest stomatal area per unit area of leaf. The most drought-intolerant species, Q. rubra (L.), exhibited the least leaf anatomical plasticity, the lowest net photosynthesis in the different light conditions, and the highest stomatal area per unit area of leaf. Quercus coccinea (Muenchh.) usually exhibited values that were intermediate between Q. rubra and Q. velutina.
Physiological processes and anatomical adaptations of leaves differ dramatically between tree species. Physio- logical differences in rates of photosynthesis, transpira- tion, and stomatal conductivity of tree species have been related to their successional status, their age (juvenile vs. mature), and their crown position in the canopy (Board- mann, 1977; Hinckley et al., 1978; Lichtenthaler et al., 1981; Fetcher, Strain, and Oberbauer, 1983; Bahari, Pal- lardy, and Parker, 1985; Abrams and Knapp, 1986; Abrams, 1988; Strauss-Debenedetti and Bazzaz, 1991). Anatomical differences in stomatal density, leaf thickness, epidermal thickness, and palisade mesophyll thickness have been similarly described (Wylie, 1951, 1954; Jack- son, 1967a, b; Carpenter and Smith, 1975; Givnish, 1988; Lee et al., 1990).
Selecting seedlings of closely related species that occur within the same forest facilitates comparison of physio- logical and anatomical characteristics (Jurik, Chabot, and Chabot, 1982). Some comparative studies of seedlings in the tropical evergreen forest have demonstrated that closely related species that occur within the same forest type, but that differ in successional status and crown stratum of occupation, have physiological processes that appear to be unrelated to their anatomical adaptations (Strauss- Debenedetti, 1989). Other tropical studies of seedlings within evergreen forest have examined closely related spe- cies that occur within the same forest type but that are of the same successional status and crown stratum. Under these circumstances results demonstrate the close asso- ciation between anatomical adaptation and efficiency in physiological process of seedlings (Ashton and Berlyn, 1992). Also, few studies have examined for differences in seedling physiology and anatomy under different light
' Manuscript received 4 February 1993; revision accepted 18 No- vember 1993.
The authors thank David Smith, Bruce Larson, Patricia Tomlinson, and Judd Isebrands for helpful comments. This study was made possible by support from the Yale University Forests.
2 Author for correspondence.
qualities (Taylor and Davies, 1988; Lee et al., 1990; Ash- ton and Berlyn, 1992).
The objective of this study was to observe and measure the physiological processes and anatomical adaptations of seedlings belonging to closely related species of the genus Quercus section Erythrobalanus (Fagaceae) within a moist temperate deciduous forest of southern New En- gland, USA. Seedlings of three species (Q. coccinea [Muenchh.], Q. rubra [L.], Q. velutina [Lam.]) were ex- amined under different light quantities and qualities. The study species are mid- to late successional trees that dom- inate the canopy stratum of mixed-species forests of east- ern North America. In southern New England each of the three species occupy a different part of the forest topog- raphy. The lower slopes and swales have Q. rubra. These sites have deep soils and are mesic year-round. Quercus velutina occurs on bedrock ridges with shallow soils. Quer- cus coccinea has a puzzling pattern of occurrence, but predominates on dry sites that have deep but coarse soils. All species have wide overlapping ranges and can fre- quently, although not predominantly, occur on any of the aforementioned sites (Burns and Honkala, 1991).
The study tested the hypothesis that these species, which are considered ofthe same successional status (sensu Watt, 1947) and occupy similar crown strata within a forest, have distinct rates of photosynthesis and stomatal con- ductivity and that this reflects the anatomical adaptations of their leaves. The pattern of relationships between leaf anatomy and physiology may explain why similar species may occur together but occupy different ecological niches within a forest (Bourdeau, 1954; Abrams, 1990).
MATERIALS AND METHODS
Design of the environment shelters-Environment shel- ters were constructed to investigate seedling leaf physi- ology and anatomy under simulated forest light environ- ments that controlled the quality (changes in R:FR ratio) and quantity of light. These shelters were placed outdoors in a large open area at the field station of the Yale-Myers
589
590 AMERICAN JOURNAL OF BOTANY [Vol. 81
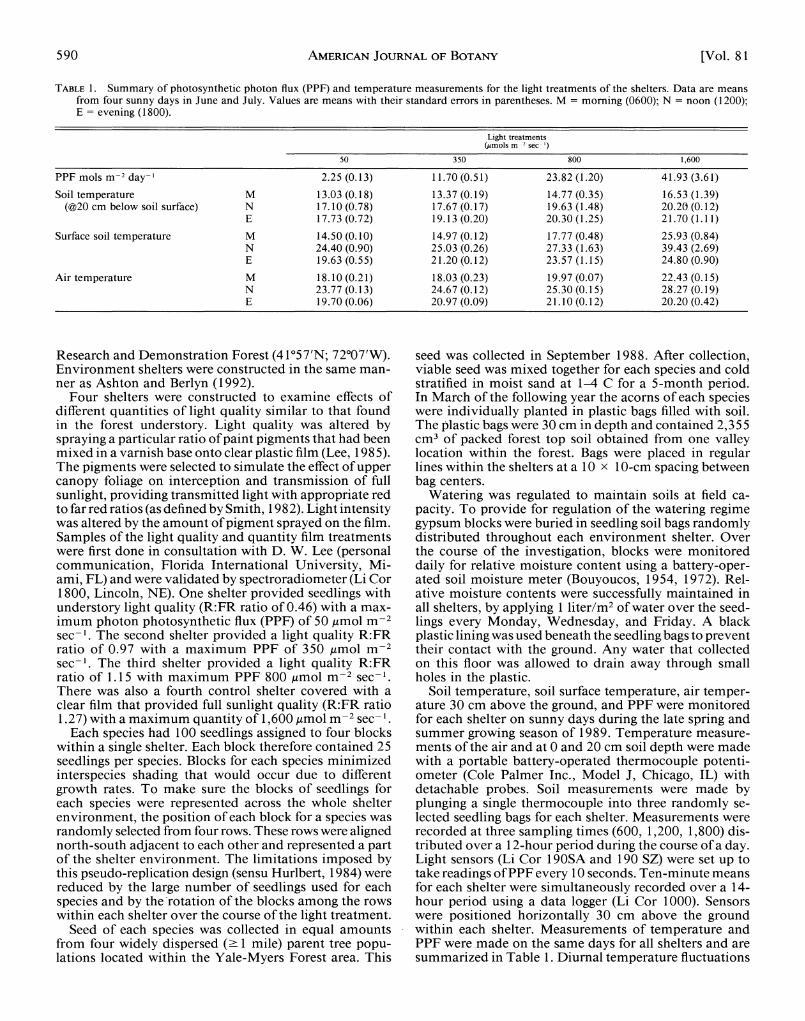
TABLE 1. Summary of photosynthetic photon flux (PPF) and temperature measurements for the light treatments of the shelters. Data are means from four sunny days in June and July. Values are means with their standard errors in parentheses. M = morning (0600); N = noon (1200); E = evening (1800).
Light treatments (umols m 2 sec ')
50 350 800 1,600
PPF mols m-2 day-' 2.25 (0.13) 11.70 (0.51) 23.82 (1.20) 41.93 (3.61) Soil temperature M 13.03 (0.18) 13.37 (0.19) 14.77 (0.35) 16.53 (1.39)
(@20 cm below soil surface) N 17.10 (0.78) 17.67 (0.17) 19.63 (1.48) 20.20 (0.12) E 17.73 (0.72) 19.13 (0.20) 20.30 (1.25) 21.70 (1.11)
Surface soil temperature M 14.50 (0.10) 14.97 (0.12) 17.77 (0.48) 25.93 (0.84) N 24.40 (0.90) 25.03 (0.26) 27.33 (1.63) 39.43 (2.69) E 19.63 (0.55) 21.20 (0.12) 23.57 (1.15) 24.80 (0.90)
Air temperature M 18.10 (0.21) 18.03 (0.23) 19.97 (0.07) 22.43 (0.15) N 23.77 (0.13) 24.67 (0.12) 25.30 (0.15) 28.27 (0.19) E 19.70 (0.06) 20.97 (0.09) 21.10 (0.12) 20.20 (0.42)
Research and Demonstration Forest (4 15 7'N; 72?07'W). Environment shelters were constructed in the same man- ner as Ashton and Berlyn (1992).
Four shelters were constructed to examine effects of different quantities of light quality similar to that found in the forest understory. Light quality was altered by spraying a particular ratio of paint pigments that had been mixed in a varnish base onto clear plastic film (Lee, 198 5). The pigments were selected to simulate the effect of upper canopy foliage on interception and transmission of full sunlight, providing transmitted light with appropriate red to far red ratios (as defined by Smith, 1982). Light intensity was altered by the amount of pigment sprayed on the film. Samples of the light quality and quantity film treatments were first done in consultation with D. W. Lee (personal communication, Florida International University, Mi- ami, FL) and were validated by spectroradiometer (Li Cor 1800, Lincoln, NE). One shelter provided seedlings with understory light quality (R:FR ratio of 0.46) with a max- imum photon photosynthetic flux (PPF) of 50 ,umol m-2 sec-1. The second shelter provided a light quality R:FR ratio of 0.97 with a maximum PPF of 350 ,umol m-2 sec-1. The third shelter provided a light quality R:FR ratio of 1.15 with maximum PPF 800 ,umol m-2 sec-1. There was also a fourth control shelter covered with a clear film that provided full sunlight quality (R:FR ratio 1.27) with a maximum quantity of 1,600 ,umol m-2 sec- 1.
Each species had 100 seedlings assigned to four blocks within a single shelter. Each block therefore contained 25 seedlings per species. Blocks for each species minimized interspecies shading that would occur due to different growth rates. To make sure the blocks of seedlings for each species were represented across the whole shelter environment, the position of each block for a species was randomly selected from four rows. These rows were aligned north-south adjacent to each other and represented a part of the shelter environment. The limitations imposed by this pseudo-replication design (sensu Hurlbert, 1984) were reduced by the large number of seedlings used for each species and by the rotation of the blocks among the rows within each shelter over the course of the light treatment.
Seed of each species was collected in equal amounts from four widely dispersed (> 1 mile) parent tree popu- lations located within the Yale-Myers Forest area. This
seed was collected in September 1988. After collection, viable seed was mixed together for each species and cold stratified in moist sand at 1-4 C for a 5-month period. In March of the following year the acorns of each species were individually planted in plastic bags filled with soil. The plastic bags were 30 cm in depth and contained 2,355 cm3 of packed forest top soil obtained from one valley location within the forest. Bags were placed in regular lines within the shelters at a 10 x 10-cm spacing between bag centers.
Watering was regulated to maintain soils at field ca- pacity. To provide for regulation of the watering regime gypsum blocks were buried in seedling soil bags randomly distributed throughout each environment shelter. Over the course of the investigation, blocks were monitored daily for relative moisture content using a battery-oper- ated soil moisture meter (Bouyoucos, 1954, 1972). Rel- ative moisture contents were successfully maintained in all shelters, by applying 1 liter/M2 of water over the seed- lings every Monday, Wednesday, and Friday. A black plastic lining was used beneath the seedling bags to prevent their contact with the ground. Any water that collected on this floor was allowed to drain away through small holes in the plastic.
Soil temperature, soil surface temperature, air temper- ature 30 cm above the ground, and PPF were monitored for each shelter on sunny days during the late spring and summer growing season of 1989. Temperature measure- ments of the air and at 0 and 20 cm soil depth were made with a portable battery-operated thermocouple potenti- ometer (Cole Palmer Inc., Model J, Chicago, IL) with detachable probes. Soil measurements were made by plunging a single thermocouple into three randomly se- lected seedling bags for each shelter. Measurements were recorded at three sampling times (600, 1,200, 1,800) dis- tributed over a 12-hour period during the course of a day. Light sensors (Li Cor 190SA and 190 SZ) were set up to take readings of PPF every 10 seconds. Ten-minute means for each shelter were simultaneously recorded over a 14- hour period using a data logger (Li Cor 1000). Sensors were positioned horizontally 30 cm above the ground within each shelter. Measurements of temperature and PPF were made on the same days for all shelters and are summarized in Table 1. Diurnal temperature fluctuations

May 1994] ASHTON AND BERLYN-LEAF PHYSIOLOGY AND ANATOMY OF QUERCUS 591
of the soil were the most moderated compared to surface and air temperatures. The highest temperatures recorded were for surface temperatures at midday. Higher soil and air temperatures were associated with shelters receiving greater PPF.
Physiology experiments-Gas exchange was measured during July 1989. Measurements determined the rate of net CO2 assimilation (net photosynthesis, PN) and sto- matal conductivity (g) of seedling leaf samples under con- ditions that approximated the environments in the ap- propriate shelters. Seedlings of each species were randomly selected from each of the four shelters, and were measured with a closed-system Li Cor 6200 infrared gas analyzer (Welles, 1986) with a 1-liter leaf chamber. The system was assembled in a well-ventilated room that attained a measured atmospheric CO2 level of 350 Al liter-1. The apparatus was periodically checked throughout the day, and calibrated according to Welles (1986) in the early morning, and in the afternoon before the start of mea- surements. Measurements were made on single attached leaves of seedlings that were well watered. To minimize ontogenetic differences (Hanson et al., 1988), leaves were all selected from the first flush of seedlings of the same age. Only undamaged leaves with no sign of scarring, disease, or herbivory were selected.
A light source (Sylvania metalarc lamp, 1000 BU-HOR) was used to obtain an illumination similar to the amount and quality provided by full sun (1,600 ,umol m-2 sec-'), with a projected area that allowed all selected seedlings for a light treatment to be positioned beneath. To emulate the light environment of the shelters, plastic films with the corresponding amounts of paint-spray pigment were placed immediately beneath the light source and above the seedlings. The light intensity was more finely adjusted by moving the metalarc lamp up or down and checking intensity beneath with a quantum sensor (Li Cor 190SA) that was placed at the level of the seedling leaf canopy. The heat generated by the light source was impeded by a glass plate that had been positioned immediately below the light source. The heat was removed by a series of extractor fans. The height of the light source varied be- tween 1 and 1.5 m above the seedling canopy.
Measurements were made on leaves of at least ten seed- lings over four pseudo-replicates of each species for each of the four light treatments. For each light treatment, measurements were taken on only those seedlings that had grown under that same light treatment in the envi- ronment shelters. To make a measurement, the chamber of the photosynthesis system was kept open over the se- lected leaf until chamber CO2 equaled atmospheric CO2 (350 Al liter-1). It was then sealed, and measurements started after a steady CO2 decline was observed on the monitor screen. Chamber temperatures were maintained at approximately 24-27 C, and relative humidity was kept between 50% and 70% during measurements. Each mea- surement of a leaf consisted of a set of three sequential readings.
To avoid bias from diurnal effects, sampling for all light treatments commenced at 8:00 a.m. and lasted until 6:00 p.m. Each species was sequentially measured at least five times over the course of this time period. Different seed-
lings for these measurements were randomly chosen with- in each species. During the period of the experiment, seedlings and their appropriate light treatment were run separately on 8 consecutive days, each treatment being repeated once.
Anatomy experiments-Leaves from seedlings were taken from two shelters and examined to identify ana- tomical traits that might relate to the physiology of each species. Measurements were designed to investigate sto- matal frequency, size, and distribution; dimensions of upper and lower epidermis, and of palisade mesophyll; and cuticle and leaf thickness. Sample leaves were taken from seedlings of each species growing in the shelters that simulated full-sun (sun leaves) and understory light (50 umol m-2 sec-') (shade leaves). Only fully expanded, undamaged leaves with no signs of scarring, disease, or herbivory were chosen for each stomate examination or sectioning. The leaves were ca. 2 months old and were randomly selected from those belonging to the first flush of a seedling.
To examine stomate density, size, and distribution, leaf peels were taken using a razor to skim the surface of the upper and lower epidermis. Each peel was placed on a slide and immersed in a drop of water underneath a cov- erslip and observed at x 200. The number of stomata was counted on both the upper and lower epidermis, and the lengths of ten stomata were taken using a x 10 filar mi- crometer eye-piece for a measure of size. Each peel was taken from a separate leaf and each leaf from a separate seedling. Five leaf peels each were used for each species and light treatment, and for each surface (upper and lower epidermis). For each peel five different fields of view were counted and lumped together as pseudoreplicates.
To examine and measure the dimensions and number of layers of epidermis and mesophyll, and to gauge the cuticle and leaf thickness, leaf cross-sections were pre- pared for microscopic examination. Three 0.5 x 1.0-cm strips were taken from the middle portion of the lamina across the midrib, each from a different seedling. Strips were immediately fixed in cold FAA (formalin: acetic acid: alcohol, formula of Berlyn and Miksche, 1976). The material was then dehydrated through a xylene series and embedded in wax. Cross sections were cut at 12 ,um with a rotary microtome. The tissue was stained with safranin and fast green following a modified procedure of Berlyn and Miksche (1976).
Slides were examined for each species and light treat- ment, with each slide representing a different leaf strip. For each slide, five measurements were made of length and breadth of upper and lower epidermis, and of palisade mesophyll. Cuticle and leaf thickness were measured on different sections of the slide, and in different places within each section, avoiding the region around the midrib. Mea- surements were made with a light microscope using a x 10 filar micrometer eye-piece and suitable objectives for the resolution required. Measures of the leaf thickness were made at x 400; cell dimensions were done at x 630; and cuticle thickness, at x 1,000.
Statistics -Analysis of variance was performed on each physiological measure (PN; g) using the ANOVA proce- dure of the Statistical Analysis System (SAS) (Ray, 1982).

592 AMERICAN JOURNAL OF BOTANY [Vol. 81
TABLE 2. Summary of physiological variables (untransformed) for three species of Quercusa in each of the light treatments. Values are means of 102 measurements taken from ten different leaves with their standard errors in brackets.
Light treatment (,umols m 2 sec ')
50 350 800 1,600 P
Net photosynthesis (PN) (,umols m-2 sec-') Quercus coccinea 1.56 (0.04)a 4.67 (0.09)a 5.27 (0.1 I)a 4.57 (0.19)b 2.93 Quercus rubra 1.53 (0.03)a 3.66 (0.09)b 3.66 (0.09)b 3.11 (0.13)c 2.03 Quercus velutina 1.45 (0.04)a 4.63 (0.08)a 5.02 (0.48)a 5.65 (0.24)a 3.89
Stomatal conductivity (g) (umol cm-2 sec-') Quercus coccinea 0.061 (0.005)a 0.089 (0.004)b 0.093 (0.005)a 0.116 (0.007)b 1.90 Quercus rubra 0.054 (0.006)a 0.075 (0.005)b 0.062 (0.004)b 0.079 (0.004)c 1.46 Quercus velutina 0.069 (0.006)a 0.120 (0.007)a 0.099 (0.006)a 0.183 (0.009)a 2.65
a Lowercase letters denote differences in log transformed data at the 5% level of significance among species within a light treatment. Species followed by the same lowercase letter within a treatment are not significantly different. Values are also given for physiological plasticity P (sun' 600/ shade50).
All data were log transformed prior to analysis. Analyses tested for differences among species, among light treat- ments, and for interactions among species and light treat- ment. All F statistics were significant (P < 0.001), and therefore differences among species were evaluated at the 5% significance level using Tukey's Studentized Range.
Data for the anatomical attributes were also analyzed by the ANOVA procedure of SAS (Ray, 1982). Analyses tested for differences among species, among light treat- ments, and for interactions among species and light treat- ment. Tukey's Studentized Range was used to determine differences among the treatments and species at the 5% significance level. Where appropriate, correlations were
obtained between anatomical and physiological attributes using the regression procedure of SAS (Ray, 1982). Phys- iological attributes were analyzed using log-transformed data.
RESULTS
Physiology - Results showed statistically significant dif- ferences in rate of net photosynthesis (PN) per unit of leaf area between species and light treatment (Table 2; Fig. 1). At low values of PPF (50 ,umol m-2 sec-1), Q. rubra, Q. coccinea, and Q. velutina had similar values of PN. At PPF amounts of 350 to 800 umol m-2 sec-', Q. velutina
C,,
o
E 4
cn ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~~~ .rubra
LU Q. coccinea 5 Q. velutina
>~2 cn 0
0)
0 5 0 3 50 8 00 1 60 0
PPF (ptmol m-2 s-1) Fig. 1. Mean net photosynthesis rates (PN) of the three Quercus species for the different light treatments.

TABLE 3. Summary of leaf anatomical variables for the three Quercusa species for full sun (sun leaves; 1,600 Amols m-2 sec-') and un- derstory shade (shade leaves; 50 Amols mr-2 sec- '). Values are given for anatomical plasticity P (sun/shade). Data are means from four different leaves. Values are means with their standard errors in parentheses.
Treatments
Species Shade Sun P
Blade thickness (,um) Quercus coccinea 84.079 (2.20)b 132.659 (2.39)a 1.58 Quercus rubra 90.152 (1.08)b 125.289 (5.54)a 1.39 Quercus velutina 93.952 (1.05)b 134.659 (9.26)a 1.43
Cuticle thickness (,um) Quercus coccinea 1.120 (0.05)d 3.051 (0.15)b 2.72 Quercus rubra 1.62 1 (0.13)cd 3.174 (0. I O)b 1.96 Quercus velutina 1.766 (0.08)c 3.860 (0.23)a 2.18
Stomatal length (rAm) Quercus coccinea 8.69 (0.41)b 8.65 (0.57)b 1.03 Quercus rubra 17.40 (1.23)a 17.39 (0.97)a 1.00 Quercus velutina 17.88 (l.l1 )a 17.90 (1.50)a 0.98
Stomatal density (no./mm2) Quercus coccinea 765 (40)b 1,032 (20)a 1.35 Quercus rubra 583 (19)d 695 (16)c 1.19 Quercus velutina 196 (8)f 293 (7)e 1.49
Stomate Area Index (SAI) Quercus coccinea 6,655 8,978 1.35 Quercus rubra 10,144 12,093 1.19 Quercus velutina 3,508 5,244 1.49
Lower epidermal cell thickness (,um) Quercus coccinea 7.123 (0.25)c 9.051 (0.23)a 1.27 Quercus rubra 9.368 (0.33)a 9.927 (0.17)a 0.98 Quercus velutina 8.793 (0.19)b 8.937 (0.23)ab 1.02
Upper epidermal cell thickness (,Am) Quercus coccinea 13.540 (0.64)d 18.930 (1.62)bc 1.37 Quercus rubra 20.644 (l.19)b 26.304 (0.87)a 1.27 Quercus velutina 17.720 (0.66)c 27.019 (0.83)a 1.52
Palisade cell thickness (,um) Quercus coccinea 19.659 (2.47)d 43.862 (1.34)a 2.23 Quercus rubra 20.414 (0.62)d 40.184 (1.32)a 1.97 Quercus velutina 24.404 (0.22)c 29.187 (1.53)b 1.19
a Lowercase letters beside the means denote difference among treat- ments and species at the 5% level. Species with treatments followed by the same lowercase letter are not significantly different. The Stomate Area Index (SAI) is the product of the mean stomate length (plem) and stomatal density.
and Q. coccinea had higher PN than Q. rubra. At amounts of PPF equal to that of full sun (1,600 ,umol m-2 sec-1) Q. velutina had higher PN than Q. coccinea, which in turn had greater PN than Q. rubra.
Results show that each of the three Quercus species had optimum PN rates at different PPF spectral qualities and intensities. The highest PN rates for Q. rubra were achieved at PPF beteen 350 Amol m-2 sec-t and 800 Amol m-2 sec- 1. For Q. coccinea, highest PN was at PPF of 800 ,umol m-2 sec-', and for Q. velutina PN was highest at PPF of 1,600 ,mol m-2 sec-'.
Fig. 2. Blade thickness (top), cuticle thickness (middle), and stomatal density (bottom) for sun and shade leaves of Quercus species.
LEAF THICKNESS
200 -
E Quercus coccinea
10Quercus rubra E 150 Quercus velutina
w 100
LL 50 _ w
SUN SHADE CUTICLE
w -J-
0
0
SUN SHADE
STOMATAL DENSITY
'4 1200
E
E
_ 800 s,S
60-
w
1200
I- 0 C,) 03i sst ^ Z rSUN SHAD

594 AMERICAN JOURNAL OF BOTANY [Vol. 81
2OjJm~~~~~~~~~S

May 1994] ASHTON AND BERLYN-LEAF PHYSIOLOGY AND ANATOMY OF QUERCUS 595
Trends were shown for stomatal conductivity (g) among species and across light treatments as those described for PN (Table 2), but there were some noticeable differences. All species increased g with increasing am'ounts of PPF. Highest rates of g were therefore associated with amounts of PPF that were equivalent to full sun (1,600 umol m-2 sec- 1). Significant differences were also shown among spe- cies within a light treatment. In all light treatments Q. velutina had higher rates of g than Q. coccinea, which in turn had higher rates of g than Q. rubra. Differences in g among species were more accentuated with higher PPF. Greatest physiological plasticity (sunl"600/shade50) was ex- hibited by Q. velutina, followed in order by Q. coccinea, and then by Q. rubra (Table 2).
Anatomy-All species produced significantly thicker leaf blades when grown in full sun as compared to shade (Table 3; Figs. 2, 3). No significant differences in thickness of leaf blades were found among species for a given light treatment, and this thickness was not correlated with PN or g. However, a correlation was found between relative water use (PN/g) and blade thickness (R2 = 0.60*) (* here- after denotes significance at 0.05 level).
There were significant differences in cuticle thickness among species and between leaves grown in full sun and those grown in shade (Table 3; Fig. 2). Cuticles of both shade and sun leaves of Q. velutina were significantly thicker than those of Q. coccinea, and the sun leaves of Q. rubra. In all species cuticle thicknesses of sun leaves were significantly thicker than those leaves grown in the shade. Although Q. velutina, the species with the greatest PN and g, had the thickest cuticle in comparison with the other species, the ranking with PN and g did not appear to hold when comparing Q. coccinea with Q. rubra, where the relationship with cuticle thickness was reversed.
Stomata for seedlings of Quercus species are located on the lower leaf surface only. There were significant differ- ences in the number of stomates per unit area among the species. Highest densities were obtained for Q. coccinea, followed in order by Q. rubra and Q. velutina. All Quercus species had significantly higher numbers of stomates per unit area in leaves that were exposed to full sun, compared to those exposed to shade. Stomatal density and relative water use (PN/g) among Quercus species were not well correlated.
The mean lengths of the stomates were not different between shade and sun leaves within each species. How- ever, stomate lengths of Q. coccinea were significantly smaller than Q. rubra and Q. velutina. An index of stomate area per unit area of leaf for each species (Table 3) was calculated by taking the product of the mean stomate length and the stomate density per unit area. This Stomate Area Index (SAI) is correlated with the physiological mea- surements. Correlations between SAI and PN among the Quercus species had an R2 = 0.98* for sun leaves. There was also a similar correlation among species between SAI and g (sun R2 = 0.99*; shade R2 = 0.99*), and SAI and PN/g (sun R2 = 0.80*; shade R2 = 0.97*).
Species and light treatments caused significant differ- ences in lower and upper epidermal cell thicknesses; these were more marked for upper epidermis than for lower epidermis. Thicknesses of upper epidermal cells for all species were significantly greater for sun leaves than for shade. For the lower epidermal cells only Q. coccinea was significantly thicker for sun leaves in comparison to shade leaves. In most all cases species had upper epidermal cell dimensions that were twice that of the lower epidermal cells. Quercus velutina and Q. rubra had thicker epidermal cells than Q. coccinea, except for the lower epidermal cells of sun leaves. No correlation was found between leaf epidermal cell dimensions and physiological measures for any species or light treatment.
Palisade cell lengths are significantly longer for sun leaves than shade leaves for all species. Quercus coccinea and Q. rubra had longer palisade cells than Q. velutina for leaves grown in the sun, but this was reversed for leaves grown in the shade. Palisade cell lengths were not well correlated with other anatomical or physiological attri- butes.
Anatomical measures of plasticity (P), exhibited by sun/ shade ratio (Table 3), in some cases showed close cor- relations with physiological measures of plasticity (P = sun" 600/shade50) (Table 2). Stomatal density plasticity was correlated with PN plasticity (R2 = 0.99*), and with g plasticity (R2 = 0.96*). Upper epidermal cell thickness was correlated with g (R2 = 0.99*), and with PN (R2 =
0.99*). Greatest plasticity of upper epidermal cell thick- ness, stomatal density, and the physiological measures (g, PN) was demonstrated by Q. velutina, and least was with Q. rubra. Also, plasticity of SAI was greatest for Q. velutina and least for Q. rubra. However, for stomate lengths little plasticity was exhibited between shade and sun leaves of each species. For blade and cuticle thicknesses, and lower epidermal cell thickness, greatest plasticity between shade and sun leaves was exhibited by Q. coccinea and least by Q. rubra.
DISCUSSION
Results clearly demonstrate correlation between certain anatomical adaptations and physiological processes of seedlings. Particularly evident is the relation of g and PN with SAI. This index is the most representative measure examined that can be associated with the sites where each species predominates. The species with the highest SAI, Q. rubra, is associated with the mesic sites of the valleys and lower slopes. This species, compared with the other species, has a low water use efficiency in full sun (Seidel, 1972; Abrams, 1990). The species with the lowest SAI, Q. velutina, occupies ridges that have shallow soils. This species has a high efficiency of water use in full sun. SAI may be a useful method of approximating stomate area per unit area of leaf and could be used to predict the drought tolerance of species. In this study SAI suggests Q. velutina to be the most drought tolerant and Q. rubra to be the most drought intolerant.
Fig. 3. Photomicrographs of leaf sections for Q. coccinea-sun (A), shade (B); Q. rubra-sun (C), shade (D); Q. velutina-sun (E), shade (F).

596 AMERICAN JOURNAL OF BOTANY [Vol. 81
The index also reflects light tolerance with Q. velutina, the species having the lowest SAI measure being the most light demanding, and Q. rubra, the species with the highest SAI measure being the most shade tolerant. These inter- pretations are reinforced by certain measures of physio- logical (PN, g) and anatomical (stomate frequency, upper epidermal cell thickness) plasticity. The most drought- tolerant species, Q. velutina, exhibited the highest level of plasticity, and Q. rubra, the most drought-prone and shade-tolerant species, showed the lowest plasticity.
No clear pattern could be found in the drought and light tolerance of the Quercus species that could be cor- related with their cuticle and leaf blade thickness. How- ever, there do appear to be trends in plasticity among these measures. Leaf blade and cuticle thickness show Q. coccinea to have greatest plasticity and Q. rubra to have the least. This flexibility of attributes would suggest a greater ability to adapt to heterogeneous environments. This might suggest why Q. coccinea can be found on the deep coarse soils that are more drought prone than those soils usually occupied by Q. rubra. In the other attributes that were measured, Q. coccinea generally exhibited val- ues that were intermediate between those of Q. rubra and Q. velutina and this is consistent with its intermediate status in drought tolerance. Therefore, Q. coccinea may not be able to compete successfully with Q. velutina when growing on the most drought-prone, shallow soils. This corresponds to its position within the forest topography in relation to its associates.
Although results clearly indicate why Q. rubra is re- stricted to the more mesic sites, they do not suggest why Q. rubra is more common on these sites when compared to Q. velutina and Q. coccinea. However, although PN of Q. coccinea and Q. velutina are significantly greater than Q. rubra under ever-moist conditions, we suggest that their rates of respiration are also higher. This would mean greater amounts of carbohydrate can be stored or used for growth rather than maintenance and repair in Q. rubra compared with the other Quercus spp.
Differences in cuticle and leaf blade thickness, and epi- dermal and palisade cell thickness among shade and sun leaves of Quercus produced similar results to those re- ported in many other studies for other temperate species (Wylie, 1951, 1954; Jackson 1967a, b; Carpenter and Smith, 1975; Taylorand Davies, 1988). Shade leaves have significantly thinner dimensions than sun leaves. The thicker anatomical dimensions promote higher efficiency in water use and lower evapotranspiration demands under high radiation.
Results support findings from another study done on an assemblage of species that occur, and occupy the same canopy stratum of the mature phase of stand development (Ashton and Berlyn, 1992). In that study we examined seedlings of four species of Shorea section Doona from mixed-dipterocarp forest in southwestern Sri Lanka. These species had closer and greater numbers of correlations between measured anatomical and physiological param- eters than the Quercus species in this study. This might be related to differences in degree of habitat specialization among species of Shorea section Doona as compared to Quercus section Erythrobalanus. Based on the hypothesis of Grubb (1977) concerning the partitioning of the re- generation niche, we speculate that closer and greater
numbers of correlations would be found between ana- tomical and physiological measures of related species that grow in environments that provide a more predictable, and more distinct variety of habitats. Our findings for this study therefore suggest that the greater overlap of anatomical and physiological attributes for Quercus spp., as compared to Shorea spp., ensure their survival in the more unpredictable forest environment of southern New England. It should be recognized that these studies have examined patterns in seedling anatomy and physiology and did not investigate changes in ontogeny with age. Findings for mature trees might be very different.
Further studies are investigating these relationships on other generic assemblages. Patterns among individual species that belong to a closely related assemblage, and patterns among the assemblages themselves, may prove useful to understanding species habitat specialization in different environments. With this knowledge it may be possible to use species assemblages as indicators of eco- system adaptibility and resilience to change.
LITERATURE CITED
ABRAMS, M. D. 1988. Comparative water relations of three succes- sional hardwood species in central Wisconsin. Tree Physiology 4: 263-273.
1990. Adaptations and responses to drought in Quercus species of North America. Tree Physiology 47: 227-238.
, AND W. K. KNAPP. 1986. Seasonal water relations of three gallery forest hardwood species in northeast Kansas. Forest Science 32: 687-696.
ASHTON, P. M. S., AND G. P. BERLYN. 1992. Leaf adaptations of some Shorea species to sun and shade. New Phytologist 121: 587-596.
BAHARI, Z. A., S. G. PALLARDY, AND W. C. PARKER. 1985. Photosyn- thesis, water relations, and drought adaptation in six woody species of oak-hickory forests in central Missouri. Forest Science 31: 557- 569.
BERLYN, G. P., AND J. P. MIKSCHE. 1976. Botanical microtechnique and cytochemistry. Iowa State University Press, Ames, IA.
BOARDMAN, N. K. 1977. Comparative photosynthesis of sun and shade plants. Annual Review of Plant Physiology 28: 355-377.
BOURDEAU, P. 1954. Oak seedling ecology determining segregation of species in piedmont oak-hickory forests. Ecological Monographs 24: 297-320.
Bouyoucos, G. J. 1954. New type electrode for plaster of Paris mois- ture blocks. Journal of Soil Science 78: 339-342.
1972. A new electrical soil-moisture measuring unit. Journal of Soil Science 114: 493.
BURNS, R. M., AND B. H. HONKALA. 1991. Silvics of North America, vol. 2, Hardwoods. USDA Forest Service Agriculture Handbook No. 654. U.S. Government Printing Office, Washington, DC.
CARPENTER, S. B., AND N. D. SMITH. 1975. A comparative study of leaf thickness among southern Appalachian hardwoods. Canadian Journal of Botany 59: 1393-1396.
FETCHER, N., B. R. STRAIN, AND S. F. OBERBAUER. 1983. Effects of light regimes on the growth, leaf morphology and water relations of seedlings of two species of tropical trees. Oecologia 58: 314-319.
GIVNISH, T. J. 1988. Adaptation to sun and shade: a whole plant perspective. Australian Journal of Plant Physiology 15: 63-92.
GRUBB, P. J. 1977. The maintenance of species-rich communities: the importance of the regeneration niche. Biological Review 52: 107- 145.
HANSON, P. J., J. G ISEBRANDS, R. E. DIXON, AND R. K. DIXON. 1988. Ontogenetic patterns of CO' exchange of Quercus rubra L. leaves during three flushes of shoot growth. I. Median flush leaves. Forest Science 34: 55-68.
HINCKLEY, T. M., R. G. ASLIN, R. R. AUBUCHON, C. L. METCALF, AND J. E. ROBERTS. 1978. Leaf conductance and photosynthesis in four species of the oak-hickory forest type. Forest Science 24: 73-84.

May 1994] ASHTON AND BERLYN-LEAF PHYSIOLOGY AND ANATOMY OF QUERCUS 597
HURLBERT, S. H. 1984. Pseudoreplication and the design of ecological field experiments. Ecological Monographs 54: 187-211.
JACKSON, L. W. R. 1967a. Effect of shade on leaf structure of deciduous tree species. Ecology 48: 498-499.
1967b. Relation of leaf structure to shade tolerance of dicot- yledonous tree species. Forest Science 13: 321-323.
JURIK, T. W., J. F. CHABOT, AND B. F. CHABOT. 1982. Effects of nutrients on leaf size, CO2 exchange, and anatomy in wild strawberry (Fragaria virginiana). Plant Physiology 70: 1044-1048.
LEE, D. W. 1985. Duplicating foliage shade for research on plant development. Hortscience 20: 28-30.
, R. A. BONE, S. L. TARSIS, AND D. STORCH. 1990. Correlates of leaf optical properties in tropical forest sun and extreme-shade plants. American Journal of Botany 77: 370-380.
LICHTENTHALER, H. K., C. BUSCHMANN, M. DOL, H. J. FIETZ, T. BACH, U. KOZEL, D. MEIER, AND U. RAHMSDORF. 1981. Photosynthetic activity, chloroplast ultrastructure and leaf characteristics of high- light and low-light plants and of sun and shade leaves. Photosyn- thesis Research 2: 115-141.
RAY, A. A. 1982. SAS user's guide: statistical analysis system. SAS Institute, Inc., Cary, NC.
SEIDEL, K. W. 1972. Drought resistance and internal water balance of oak seedlings. Forest Science 18: 34-40.
SMITH, H. 1982. Light quality, photoreception and plant strategy. Annual Review of Plant Physiology 33: 481-518.
STRAUSS-DEBENEDETTI, S. I. 1989. Responses to light in tropical Mo- raceae of different successional stages. Ph.D. thesis, Yale University. New Haven, CT.
, AND F. A. BAzzAz. 1991. Plasticity and acclimation to light in tropical Moraceae of different successional positions. Oecologia 87: 377-387.
TAYLOR, G., AND W. J. DAVIES. 1988. The influence of photosyn- thetically active radiation and simulated shadelight on the leaf growth of Betula and Acer. New Phytologist 108: 393-398.
WATT, A. S. 1947. Pattern and process in the plant community. Journal of Ecology 35: 1-22.
WELLES, J. 1986. A portable photosynthesis system. In E. D. Gensler [ed.], Advanced agricultural instruments, 231-236. Martinus Nijhoff, Amsterdam.
WYLIE, R. B. 1951. Principles of foliar organization shown by sun- shade leaves from ten different species of deciduous dicotyledonous trees. American Journal of Botany 36: 355-361.
1954. Leaf organization of some woody dicotyledons from New Zealand. American Journal of Botany 4: 186-192.






















