Embed Size (px)
Citation preview

A COMPARATIVE STUDY OF HEXOKINASE FROM YEAST AND ANIMAL TISSUES*
BY MILTON W. SLEIN,? GERTY T. CORI, AND CARL F. CORI
(From the Department of Biological Chemistry, Washington University School of Medicine, St. Louis)
(Received for publication, April 17, 1960)
Yeast hexokinase catalyzes the transfer of one phosphate group of adeno- sinetriphosphate (ATP) to fructose, glucose, and mannose, converting them to the respective g-esters. At concentrations which saturate the enzyme, the order in the rate of phosphorylation is fructose > glucose > mannose. The question has been investigated whether yeast hexo- kinase has one or more active centers for the utilization of these three hexoses. It was found that from equimolar mixtures of the pairs, fructose- glucose or fructose-mannose, the aldo sugars were much more rapidly re- moved than the keto sugar. There was also mutual inhibition between the pair mannose-glucose, the former sugar being a stronger inhibitor than the latter. The order of substrate affinity for the enzyme was mannose > glucose > fructose. In the case of fructose it seems probable that it is the furanose form (with a free hydroxyl on carbon 6) which is acted upon by the yeast enzyme.
Brain hexokinase also showed inhibition of fructose utilization by glu- cose or mannose. Fractionation of the brain enzyme did not result in a change of the relative rates of glucose and fructose utilization and the primary products formed were the respective 6-esters.
It was possible to obtain enzyme preparations from liver and muscle which acted on fructose but not on glucose and vice versa. The fructo- kinase of liver and muscle was not inhibited by glucose, and the primary reaction product, at least in the case of liver, appeared to be fructose-l- phosphate. Enzymes which act on fructose-l-phosphate have been found in liver and muscle, but not in brain.
Materials and Methods
Crystalline hexokinase was prepared from bakers’ yeast as previously described (1). Partially purified brain hexokinase was obtained by
* This work was supported by a grant from the Nutrition Foundation, Inc. From the thesis presented by M. W. Slein to the Board of Graduate Studies, Washington University, in partial fulfilment of the requirements for the degree of Doctor of Philosophy.
t Present address, Camp Detrick, Frederick, Maryland. 763
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

764 YEAST AND ANIMAL HEXOKINASE
(NH&S04 fractionation of aqueous extracts of acetone-dried homogenates of fresh beef or sheep brain cortex (2).
Phosphomannose isomerase was obtained from rabbit muscle in partially purified form by the procedure described in the preceding paper (3). Lohmann’s phosphohexose isomerase was partially purified from rabbit muscle extract according to an unpublished method of A. A. Green. Zwischenferment was prepared from brewers’ yeast by the methods of Warburg and Christian (4) and Negelein and Gerischer (5). It was separ- ated from hexokinase by repeated isoelectric precipitations at pH 4.8 and by allowing solutions to stand for several days at 0”.
The n-glucose used was Mallinckrodt’s analytical reagent. The D-
fructose (crystalline) and n-mannose were Pfanstiehl’s c.p. special grade. ATP was prepared from rabbit muscle by a modification of the method described by Lohmann (6). Triphosphopyridine nucleotide (TPN) was prepared from horse liver according to a modification of the method used by Warburg, Christian, and Griese for the isolation of TPN from red cells (7).
The rate of the hexokinase reaction was determined in three ways. The first procedure was the direct measurement of sugar disappearance in aliquots of the reaction mixture after precipitation of protein and phos- phorylated sugars with Ba(OH)2 and ZnSOI. Reducing sugars were esti- mated in the filtrates by the method of Nelson (8). Fructose was mea- sured by the procedure of Roe (9). Sugar disappearance was calculated by difference between initial and incubated samples. This method had rather limited usefulness for the determination of enzyme-substrate dis- sociation constants because a determination of initial rates, especially at low sugar concentrations, required that only a small fraction of the sub- strate be used up. Hence only a narrow range of sugar concentrations could be investigated.
Two other methods of much greater sensitivity and flexibility were based on the spectrophotometric determination of the reaction product of the hexokinase reaction, hexose-6-phosphate, by means of Zwischenferment and TPN.
In the “direct” method all auxiliary enzymes (Zwischenferment and one or two isomerases, as needed) were present in excess; the rate of reduction of TPN at 340 rnp, as observed in the Beckman spectrophotometer, was then a measure of the rate of the hexokinase reaction. Each micro- gram of sugar phosphorylated in a reaction mixture of 3 ml. in a 1 cm. cell corresponds to a log lo/l reading’ of 0.0115. In some cases discrepan-
1 The value of 6.22 X lo6 sq. cm. per mole for a cuvette of 1 cm. was used as the molecular extinction coefficient for reduced TPN (10).
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

M. W. SLEIN, Q. T. CORI, AND C. F. CORI 765
ties were noted between the rate of hexose utilization as measured spec- trophotometrically and by chemical analysis, the former method giving lower values than the latter. This was generally the result of not using a sufficient excess of auxiliary enzymes.
In the “indirect” spectrophotometric method a reaction mixture was prepared in multiples of the following: 0.5 ml. of 0.05 M veronal, pH 8, 0.1 ml. of 0.1 M MgCh, 0.1 ml. of 0.1 per cent insulin, 0.1 ml. of 0.013 M
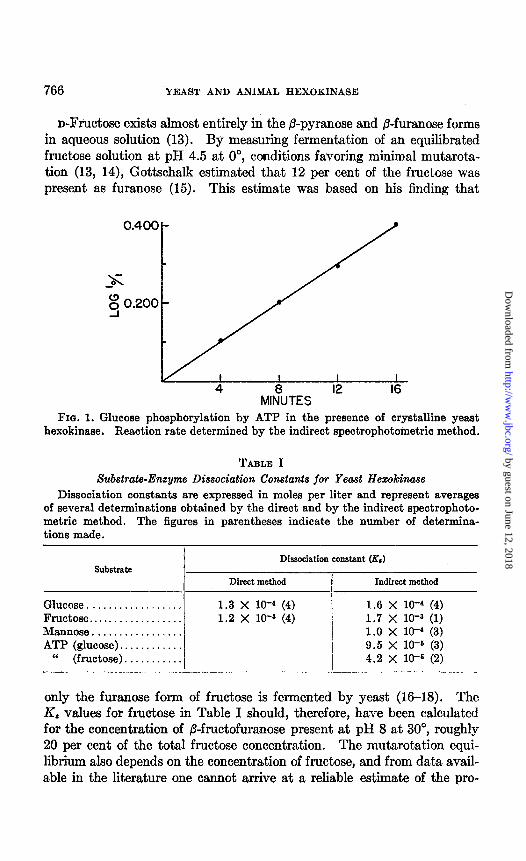
ATP, 0.3 ml. of HzO. Samples (1.0 ml.) were transferred to small tubes and 0.1 ml. of hexose solution (of desired concentration) was added. After temperature equilibration at 30”, the reaction was started by the addition of 0.1 ml. of a solution containing 0.05 to 0.3 y of crystalline yeast hexo- kinase stabilized with insulin (1). The reaction was stopped by the addi- tion of 0.5 ml. of 0.1 N HCI, an amount sufficient to inactivate the enzyme. After neutralization, an aliquot was transferred to a cuvette, and to this were added 0.5 ml. of 0.05 M veronal, pH 8, 0.2 ml. of Zwischenferment, and 0.1 ml. of Lohmann’s or phosphomannose isomerase.2 The volume was brought to 2.95 ml. and, after determining the initial optical density, with water in the reference cell, 0.05 ml. of TPN (0.3 PM) was added, and the reduction of TPN was followed. to the end-point which was usually reached within 10 minutes. Fig. 1 shows a time curve for hexokinase activity obtained by the indirect spectrophotometric method; the rate of the reaction remained constant. Even with the lowest concentrations of sugar or ATP used, experimental conditions could be so arranged with respect to hexokinase concentration and time of incubation that the measurements fell on the linear portion of the rate curve.
Results
Yeast HexoKzase-Dissociation constants obtained with crystalline yeast hexokinase and its substrates are given in Table I. The constants were calculated from linear plots of l/v cersus l/s, v being the initial rate and s the substrate concentration. The K, values for glucose and mannose represent the affinity of hexokinase for the (Y- and P-pyranose forms of these sugars, these being the prevalent forms in which these aldoses exist in solution. Gottschalk (11) and others have shown that the cr and /3 forms of glucose are fermented at the same rate by yeast. This is also true for mannose (12).
2 It is essential that these enzymes be free of hexokinase because the solution to be analyzed for hexose monophosphate contains free sugar and ATP. Similarly, phosphofructokinase must be absent; otherwise some of the monophosphate would be diverted from the Zwischenferment reaction by conversion to fructose diphosphate. Finally, the Zwischenferment preparation must be sufficiently purified so that it does not contain proteins which reoxidize reduced TPN.
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from
766 YEAST AND ANIMAL HEXOKINASE
D-Fructose exists almost entirely in the @pyranose and &furanose forms in aqueous solution (13). By measuring fermentation of an equilibrated fructose solution at pH 4.5 at 0”, conditions favoring minimal mutarota- tion (13, 14), Gottschalk estimated that 12 per cent of the fructose was present as furanose (15). This estimate was based on his finding that
4 ld”TES
12 16
FIN. 1. Glucose phosphorylation by ATP in the presence of crystalline yeast hexokinase. Reaction rate determined by the indirect spectrophotometric method.
TABLE I Substrate-Enzyme Dissociation Constants for Yeast Hexokinase
Dissociation constants are expressed in moles per liter and represent averages of several determinations obtained by the direct and by the indirect spectrophoto- metric method. The figures in parentheses indicate the number of determina- tions made.
I Dissociation constant (K.1 Substrate
Glucose .................. Fructose .................. Mannose ................. ATP (glucose). ...........
“ (fructose). ..........
Direct method
1.3 x 10-4 (4) 1.2 x 10-J (4)
Indirect method
1.6 x 10-d (4) 1.7 x 10-s (1) 1.0 x 10-d (3) 9.5 x 10-s (3) 4.2 X lO+ (2)
only the furanose form of fructose is fermented by yeast (16-18). The K, values for fructose in Table I should, therefore, have been calculated for the concentration of p-fructofuranose present at pH 8 at 30”, roughly 20 per cent of the total fructose concentration. The mutarotation equi- librium also depends on the concentration of fructose, and from data avail- able in the literature one cannot arrive at a reliable estimate of the pro-
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

M. W. SLEIN, G. T. CORI, AND C. F. CORI 767
portion of furanose present at each concentration used. The values in Table I have therefore been calculated for the total concentration of fructose present initially in each case.
Two types of experiments were carried out to test Gottschalk’s assump- tion that the exclusive fermentation of the furanose form of fructose by yeast is due to the specificity of hexokinase for this form. In both cases incubation was at pH 5.9 at O”, under which conditions the rate of mutaro- tation of fructose is only about 1.7 times as rapid as the minimal rate found at pH 4 (13). The reaction mixture contained 0.5 ml. of 0.2 M
acetate, pH 5.9, 0.1 ml. of 0.2 M MgCh, 0.5 ml. of 0.4 per cent hexose, and enzyme in a volume of 2.2 ml. The mixture was chilled in an ice bath for 5 minutes before 1 ml. was added to 0.1 ml. of 0.06 M ATP (or 0.1 ml. of water in the control sample) to start the reaction. After incubation, Ba(OH)2 and ZnSOI were added to control and to experimental samples and the filtrates analyzed for sugar.
TABLE II Proportionality of Fructose and Glucose Utilization to Hexokinase Concentration
under Conditions Giving Low Rates of Mutarotation The figures indicate micrograms of hexose used during incubation for 12 minutes
at pH 5.9, 0”. Approximately 900 y of hexose were j?resent initially.
Substrate
Fructose.............................. Glucose..............................
37
250 425
Hexokinase
68 7
230 735
In the experiment in Table II an equilibrated solution of glucose or fructose was added (about 900 y each) and the sugar utilization measured after 12 minutes of incubation at two enzyme concentrations. When glucose was the substrate, the amount of sugar used rose from 425 to 735 y when the enzyme concentration was doubled. With fructose the utiliza- tion amounted to 250 y and did not increase when the enzyme concentra- tion was doubled. This indicates that fructose utilization was limited by some non-enzymatic factor, presumably the rate of conversion of the pyranose to the furanose form.
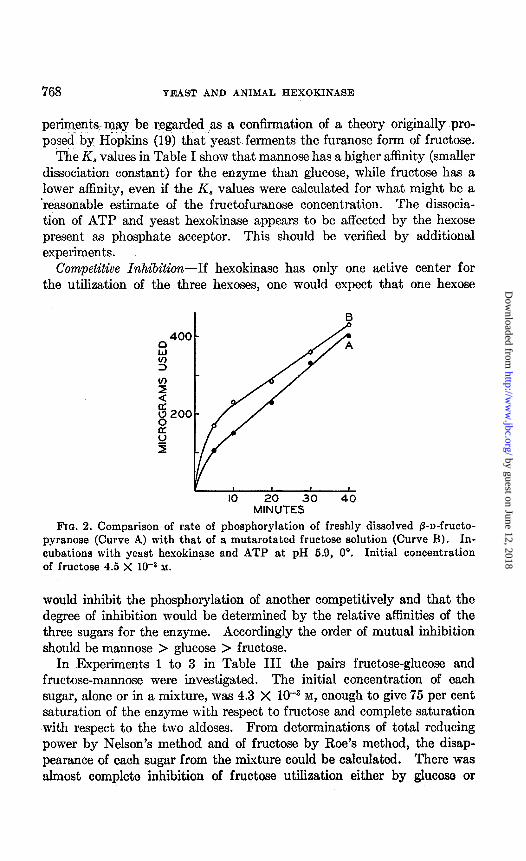
In another experiment, the rate of fructose utilization was measured under the above conditions, except that the reaction mixture contained either freshly dissolved crystalline @-n-fructopyranose or an equilibrated solution of fructose. The results given in Pig. 2 show that the initial rate of phosphorylation of freshly dissolved fructopyranose was signifi- cantly lower than that of an equilibrated solution of fructose. The ex-
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from
768 YFiAST AND ANIMAL HEXOKINASE
perimeqts-may be regarded as a confirmation of a theory originally pro- posed by Hbpkins (19) that yeast ferments the furanose form of fructose.
!I%; K, values in Table I show that mannose has a higher aflinity (smaller dissociation constant) for the enzyme than glucose, while fructose has a lower affinity, even if the K, values were calculated for what might be a ‘reasonable estimate of the fructofuranose concentration. The dissocia- tion of ATP and yeast hexokinase appears to be affected by the hexose present as phosphate acceptor. This should be verified by additional experiments.
Competitive Inhibition-If hexokinase has only one active center for the utilization of the three hexoses, one would expect that one hexose
IO 30 40 MlcJ-rES
Fro. 2. Comparison of rate of phosphorylation of freshly dissolved 8-D-frUCtO-
pyranose (Curve A) with that of a mutarotated fructose solut.ion (Curve B). In- cubations with yeast hexokinase and ATP at pH 6.0, 0”. Initial concentration of fructose 4.5 X lo+ M.
would inhibit the phosphorylation of another competitively and that the degree of inhibition would be determined by the relative affinities of the three sugars for the enzyme. Accordingly the order of mutual inhibition should be mannose > glucose > fructose.
In Experiments 1 to 3 in Table III the pairs fructose-glucose and fructose-mannose were investigated. The initial concentration of each sugar, alone or in a mixture, was 4.3 X 10V3 M, enough to give 75 per cent saturation of the enzyme with respect to fructose and complete saturation with respect to the two aldoses. From determinations of total reducing power by Nelson’s method and of fructose by Roe’s method, the disap- pearance of each sugar from the mixture could be calculated. There was almost complete inhibition of fructose utilization either by glucose or
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

M. W. SLEIN, Q. T. CORI, AND C. F. CORI 769
mannose, while fructose had little effect on the utilization of the two aldo sugars3
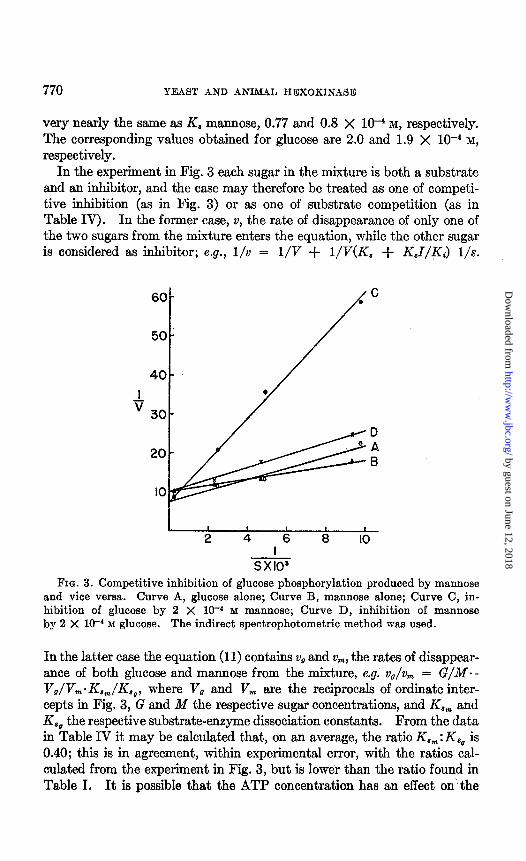
A detailed analysis was carried out for the pair glucose-mannose with the indirect spect.rophotometric method which permits initial rates to be measured, so that concentrations of reactants do not change appreciably during the period of measurements. In Fig. 3 the reciprocals of the initial velocity are plotted against the reciprocals of the substrate concentra- tion. The straight lines drawn for glucose alone (Curve A) or mannose alone (Curve B) intercept the ordinate at 7.5 and 10, respectively.4 The dissociation constants calculated from these intercepts and the slopes are 1.9 X lo4 M for glucose and 0.8 X 10V4 M for mannose. Curve C repre-
TABLE III lMutua1 Inhibitory Effects of Hexoses (Yeast Hexokinase)
The initial concentration of hexoses was 4.3 X 10m3 M (1800 y per 2.3 ml. of reac- tion mixture) and of ATP, 5 X W3 M. Incubations were with crystalline yeast hexokinase at 30”, pH 8, except in Experiment 2, where pH was 7. --
Expel?
1
2
3
-
t
--
-
Substrates
Fructose Glucose Fructose + glucose Fructose Glucose Fructose + glucose Fructose Mannose Fructose + mannose
_-
-
SiO
20 885
105 640
40
Y
570 570
550 465
170 140
T Per cent inhibition
Fructose
98
88
94
0
15
18
sents inhibition of glucose utilization by mannose and Curve D the reverse. The fact that these curves have intercepts essentially the same as those obtained in the absence of the inhibitory hexose indicates that the inhi- bition is competitive. When Ki mannose is calculated from Curve C, that is when mannose is considered as an inhibitor of glucose, the value is,
a The following substances, at a concentration of 100~ M, did not inhibit fructose utilization by yeast hexokinase: glucose-l-phosphate, glucose-g-phosphate, mannose- g-phosphate, galactose-l-phosphate, galactose.
4 This indicates that the ratio of mannose to glucose utilization is 0.75 at complete saturation with substrate. Considerably lower ratios were obt,ained previously with a manometric method (l), perhaps because at the high concentration used (0.03 &I) some impurity present in the mannose sample exerted an inhibitory effect of the enzyme.
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from
770 YEAST AND ANIMAL IIEXOKINASE
very nearly the same as K, mannose, 0.77 and 0.8 X lo--* M, respectively. The corresponding values obtained for glucose are 2.0 and 1.9 X lo-* M,
respectively. In the experiment in Fig. 3 each sugar in the mixture is both a substrate
and an inhibitor, and the case may therefore be treated as one of competi- tive inhibition (as in Fig. 3) or as one of substrate competition (as in Table IV). In the former case, 2r, the rate of disappearance of only one of the two sugars from the mixture enters the equation, while the other sugar is considered as inhibitor; e.g., l/v = l/V + l/V(K, + K,I/Ki) l/s.
I I * I L , 2 4 6 8 lo
I SXIO’
FIG. 3. Competitive inhibition of glucose phosphorylation produced by mannose and vice versa. Curve A, glucose alone; Curve B, mannose alone; Curve C, in- hibition of glucose by 2 X 10-4 M mannose; Curve D, inhibition of mannose by 2 X 1W4 M glucose. The indirect spectrophotometric method was used.
In the latter case the equation (11) contains v, and urn, the rates of disappear- ance of both glucose and mannose from the mixture, e.g. v,/v~ = G/M. - VdVm- K,,/.&,, where V, and V, are the reciprocals of ordinate inter- cepts in Fig. 3, G and M the respective sugar concentrations, and K,, and K,, the respective substrate-enzyme dissociation constants. From the data in Table IV it may be calculated that, on an average, the ratio K,,: K, is 0.40; this is in agreement, within experimental error, with the ratios cal- culated from the experiment in Fig. 3, but is lower than the ratio found in Table I. It is possible that the ATP concentration has an effect on the
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

M. W. SLEIN, G. T. CORI, AND C. F. CORI 771
sugar-enzyme dissociation constant; this had not been taken into consider- ation when these experiments were carried out. The possibility that mannose-6-phosphate inhibits the phosphorylation of glucose has been tested with negative results.
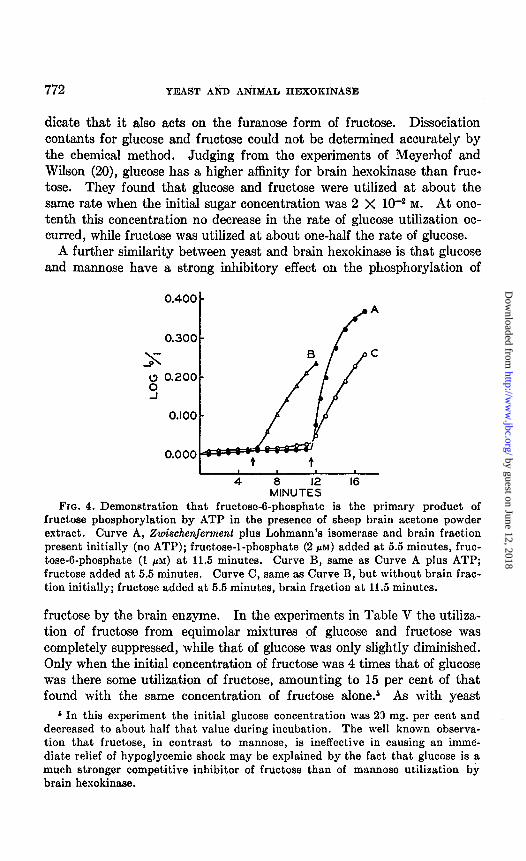
Brain Hexokinase-This enzyme utilizes fructose, glucose, and mannose at about the same relative rates as does yeast hexokinase. The primary product of glucose phosphorylation has been shown to be glucose-8phos- phate (2). The experiment in Fig. 4 indicates that the corresponding product with fructose is fructose-g-phosphate and not (as seemed possible) fructose-l-phosphate. Curve A shows that a system containing Zwischen- ferment, purified Lohmann’s isomerase, and brain hexokinase was unable to catalyze the conversion of fructose-l-phosphate to fructoseS-phos-
TABLE IV Glucose and Mannose As Competitive Substrates for Hexokinase
The same experiment as in Fig. 3, including two additional pairs of glucose- mannose. The concentration of glucose and mannose in the mixture is given in moles per liter. v glucose and v mannose correspond to the initial rate of disap- pearance of the two sugars from the mixture in terms of log IO/I readings. Ks,,,:Ks,,, the ratio of substrate-enzyme dissociation constants, calculated from the equation given in the text.
Glucose (M X lOa). ...... Mannose (M X lOa). ..... v glucose ................ s mannose. ............. G:M .................... V&II .................... Ks,,,:Ka,, ................
0.102 0.214 0.017 0.067 0.48
:I 0.254 0.398
0.205 0.410 0.205 0.205 0.214 0.214 0.107 0.428 0.028 0.048 0.042 0.019 0.056 0.046 0.040* 0.077 0.96 1.92 1.92 0.48 0.50 1.045 1.05 0.247 0.391 0.410 0.412 0.388
* Calculated, assuming the same ratio, vO:v,,,, as in the preceding column. The experimental value obtained (0.047) appears to be erroneous.
phate. That the system was otherwise complete is shown by the effect of addition of fructose-6-phosphate at 11.5 minutes; a rapid reduction of TPN occurred. Curve B represents the results obtained with the same system of enzymes as in Curve A, except that ATP was also present. Addition of fructose at 5.5 minutes resulted in a rapid reduction of TPN, showing that fructose had been converted to fructose-6-phosphate by brain hexo- kinase. That the brain fraction was the only significant source of hexo- kinase activity is shown by Curve C. In this case, the brain fraction was omitted initially and the addition of fructose at 5.5 minutes had no effect. When the brain fraction was added at 11.5 minutes, the rate of reduction of TPN proceeded at a rate comparable to that observed in Curve B.
The formation of fructose-6-phosphate by brain hexokinase would in-
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from
772 YEAST AND ANiIvUL HFXOKINASE
dicate that it also acts on the furanose form of fructose. Dissociation contants for glucose and fructose could not be determined accurately by the chemical method. Judging from the experiments of Meyerhof and Wilson (20), glucose has a higher affinity for brain hexokinase than fruc- tose. They found that glucose and fructose were utilized at about the same rate when the initial sugar concentration was 2 X 10” M. At one- tenth this concentration no decrease in the rate of glucose utilization oc- curred, while fructose was utilized at about one-half the rate of glucose.
A further similarity between yeast and brain hexokinase is that glucose and mannose have a strong inhibitory effect on the phosphorylation of
I I I I I
4 t&“TE
16
Fro. 4. Demonstration that fructose-6-phosphate is the primary product of fructose phosphorylation by ATP in the presence of sheep brain acetone powder extract. Curve A, Zwischmferment plus Lohmann’s isomerase and brain fraction present initially (no ATP); fructose-l-phosphate (2 PM) added at 5.5 minutes, fruc- tose-6-phosphate (1 PM) at 11.5 minutes. Curve B, same as Curve A plus ATP; fructose added at 5.5 minutes. Curve C, same as Curve B, but without brain frac- tion initially; fructose added at 5.5 minutes, brain fraction at 11.5 minutes.
fructose by the brain enzyme. In the experiments in Table V the utiliza- tion of fructose from equimolar mixtures of glucose and fructose was completely suppressed, while that of glucose was only slightly diminished. Only when the initial concentration of fructose was 4 times that of glucose was there some utilization of fructose, amounting to 15 per cent of that found with the same concentration of fructose alone.5 As with yeast
6 In this experiment the initial glucose concentration was 23 mg. per cent and decreased to about half that value during incubation. The well known observa- tion that fructose, in contrast to mannose, is ineffective in causing an imme- diate relief of hypoglycemic shock may be explained by the fact that glucose is a much stronger competitive inhibitor of fructose than of mannose utilization by brain hexokinase.
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from
M. W. SLEIN, Q. T. COH, AND C. P. CORI 773
hexokinase, the product of glucose’phosphorylation, glucose-&phosphate, did not inhibit the phosphorylation of fructose by brain hexokinase.
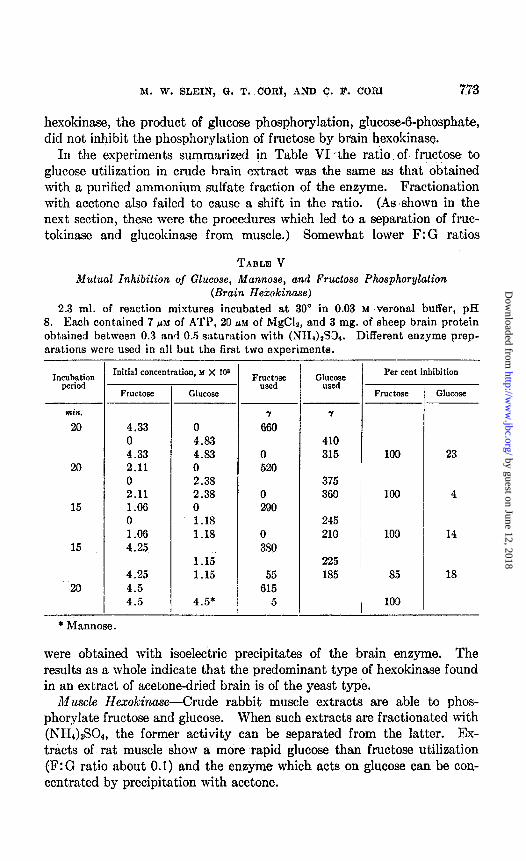
In the experiments summarized in Table VI -the ratio of fructose to glucose utilization in crude hrain extract was the same as that obtained with a purified ammonium sulfate fraction of the enzyme. Fractionation with acetone also failed to cause a shift in the ratio. (As shown in the next section, these were the procedures which led to a separation of fruc- tokinase and glucokinase from muscle.) Somewhat lower F:G ratios
TABLE V Mutual Inhibilion of Glucose, Mannose, and Fructose Phosphorylation
(Brain Hexokinose) 2.3 ml. of reaction mixtures incubated at 30” in 0.03 M Verona1 buffer, pH
8. Each contained 7 pin of ATP, 20 UM of MgC&, and 3 mg. of sheep brain protein obtained between 0.3 and 0.5 saturation with (.NH1hS3,. Different enzyme prep- arations were used in all but the first two experiments.
IllCCUU~tp
min.
20
20
15
15
20
T Initial concentration, y X 10s
Fructose
4.33 0 4.33 2.11 0 2.11 1.06 0 1.06 4.25
4.25 4.5 4.5
ClUCOSJ2
0 4.83 4.33 0 2.38 2.38 0 1.18 1.18
1.15 1.15
4.5*
6:O
0 520
0 290
0 380
55 615
5
Y
410 315
375 360
245 210
225 185
Per cent inhibition
Fructose
100 23
100
100
85
4
14
18
100
* Mannose.
were obtained with isoelectric precipitates of the brain enzyme. The results as a whole indicate that the predominant type of hexokinase found in an extract of acetone-dried brain is of the yeast type.
Muscle Hexo/&zse-Crude rabbit muscle extracts are able to phos- phorylate fructose and glucose. When such extracts are fractionated with (NH&Sod, the former activity can be separated from the latter. Ex- tracts of rat muscle show a more rapid glucose than fructose utilization (F:G ratio about 0.1) and the enzyme which acts on glucose can be con- centrated by precipitation with acetone.
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

774 YEAST AND ANIMAL HEXORINASE
The crude extract used for the experiments in Table VII was prepared by extracting ground rabbit muscle twice with 1 volume of cold 0.03 M KOH and once with 0.5 volume of distilled water. The filtrates obtained
TABLE VI
Ratio of Fructose to Glucose Utilization with Various Fractions of Sheep Brain Hexokinase
An aqueous extract of acetone-dried brain was the “crude” fraction; others were prepared from this extract. The initial concentrations of sugar (glucose or fructose) and ATP were 4 X 10-a M. Incubated in 0.05 M Verona1 buffer at pH 8 at 30”. .___
Fraction No: of experiments
btio of utilization of fructose to
glucose
Crude............................................ 7
Dialyzed crude................................... 3 20yo acetone...................................... 1 25-40yo acetone................................... 1 Ppt., pH 5.2.. . . . . . . . . . . . . 3 Supernatant fluid of ppt., pH 5.2.. . . . . . 1 Ppt., 0.3-0.5 saturation (NH&SOI. . . . . . . . . . . . . . . .4
I
_-
-
1.5 1.5 1.5 1.5 1.1 1.4 1.5
TABLE VII Utilization of Fructose and Glucose by Rabbit Muscle Extracts
The initial concentrations of sugar (glucose or fructose) and ATP were 3 X lOma 3~; MgCl* was 8 X 10ea M. Fluoride (0.06 M) was present in the reaction mixture with crude extracts. Incubations were in 0.04 M Verona1 buffer, pH 8. The figures represent micromoles of sugar used per gm. of protein
*T-2?
-.
1 2 3 4 5
i- -
.-
-
Crude extract
.~ Fructose
38 32 25 30
4
-
--
-
Glucose
37 35 83 86 69
--
--
-
0.41-0.5 saturated (NH&SO4 fraction
Fructose
262 929 591 694 694
-
. _ Ghcose
- --.-
8
0 45 0 0
--
.-
-
:r 30 minutes at 30”.
Protein per 100 gm. muscle
Crude extract
Em.
2.05 3.04 2.22 2.25 2.12
-
.-
-
@m)rSO4 fraction
m.
0.170 0.076 0.101 0.169 0.149
by straining the muscle suspension through gauze were combined (crude extract) and treated with saturated (NH&S04 adjusted to pH 7.8. The precipitate which formed at 0.41 saturation was filtered off in the cold and discarded. To the filtrate was added (NH&SOa solution to 0.5 sat-
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from
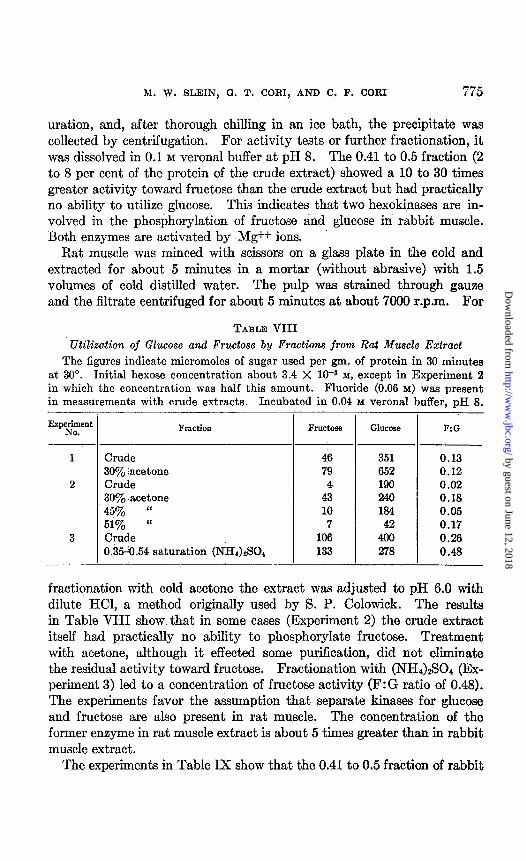
M. W. SLEIN, 0. T. CORI, AND C. F. CORI 775
uration, and, after thorough chilling in an ice bath, the precipitate was collected by centrifugation. For activity tests or further fractionation, it was dissolved in 0.1 M veronal buffer at pH 8. The 0.41 to 0.5 fraction (2 to 8 per cent of the protein of the crude extract) showed a 10 to 30 times greater activity toward fructose than the crude extract but had practically no ability to utilize glucose. This indicates that two hexokinases are in- volved in the phosbhorylation of fructose and glucose in rabbit muscle. Both enzymes are activat,ed by Mg++ ions.
Rat muscle was minced with scissors on a glass plate in the cold and extracted for about 5 minutes in a mortar (without abrasive) with 1.5 volumes of cold distilled water. The pulp was strained through gauze and the filtrate centrifuged for about 5 minutes at about 7000 r.p.m. For
TABLE VIII Utilization of Glucose and Fructose by Fractions from Rat Muscle Extract
The figures indicate micromoles of sugar used per gm. of protein in 30 minutes at 30”. Initial hexose concentration about 3.4 X 10-a M, except in Experiment 2 in which the concentration was half this amount. Fluoride (0.06 M) was present in measurements with crude extracts. Incubated in 0.04 M Verona1 buffer, pH 8.
1 Crude 46 351 0.13 30yo acetone 79 652 0.12 Crude 4 190 0.02 30% acetone 43 240 0.18 450/O it 10 184 0.05 51% “ 7 42 0.17 Crude 106 400 0.26 0.35-0.54 satdration (NH&SO* 133 278 0.48
Fraction I Glucose F:G
fractionation with cold acetone the extract was adjusted to pH 6.0 with dilute HCl, a method originally used by S. P. Colowick. The results in Table VIII show that in some cases (Experiment 2) the crude extract itself had practically no ability to phosphorylate fructose. Treatment with acetone, although it effected some purification, did not eliminate the residual activity toward fructose. Fractionation with (NH&SO4 (Ex- periment 3) led to a concentration of fructose activity (F:G ratio of 0.48). The experiments favor the assumption that separate kinases for glucose and fructose are also present in rat muscle. The concentration of the former enzyme in rat muscle extract is about 5 times greater than in rabbit muscle extract.
The experiments in Table IX show that the 0.41 to 0.5 fraction of rabbit
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from
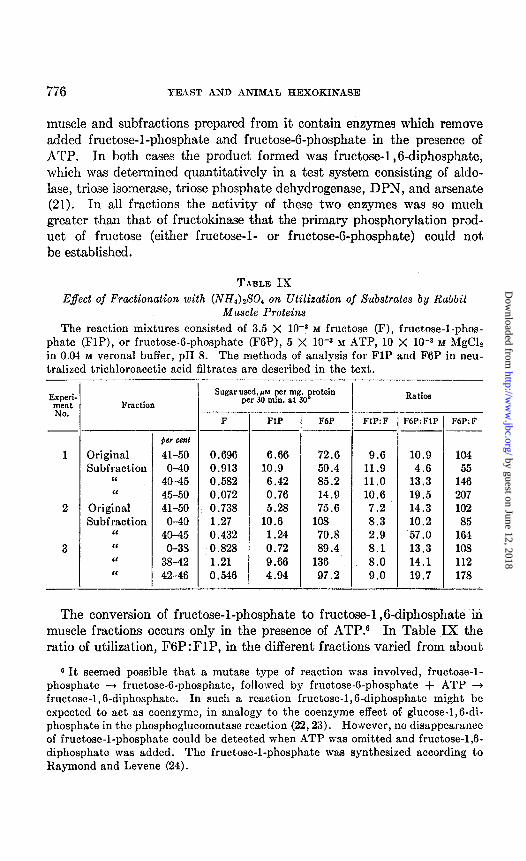
776 YEAST AND ANIMAL IIEXOKINASE
muscle and subfractions prepared from it contain enzymes which remove added fructose-l-phosphate and fructose-6-phosphate in the presence of ATP. In both cases the product formed was fructose-l ,6-diphosphate, which was determined quantitatively in a test system consisting of aldo- lase, triose isomerase, triose phosphate dehydrogenase, DPN, and arsenate (21). In all fractions the activity of these two enzymes was so much greater than that of fructokinase that the primary phosphorylation prod- uct of fructose (either fructose-l- or fructose-G-phosphate) could not be established.
TABLE IX Effect of Fractionation with (NH4)$04 on Utilization of Substrates by Rabbit
Muscle Proteins The reaction mixtures consisted of 3.5 X 10-a M fructose (F), fructose-l-phos-
phate (FlP), or fructose-6-phosphate (F6P), 5 X 10-a M ATP, 10 X 10-a M iUgClz in 0.04 M Verona1 buffer, pH 8. The methods of analysis for FlP and F6P in neu- tralized trichloroacetic acid filtrates are described in the text.
Fraction Experi-
ment No.
Original Subfraction
“ ‘C
Original Subfraction
“ I‘ “ “
- I
gcr cm1
41-50 040
40-45 45-50 41-50
O-40 40-45
o-35 38-42 42-46
Sugar used, w per mg. per 30 mm. at 30
protein
F FlP
0.696 6.66 0.913 10.9 0.582 6.42 0.072 0.76 0.738 5.28 1.27 10.6 0.432 1.24 0.828 0.72 1.21 9.66 0.546 4.94
- __
-
F6P FlP:F F6P: FlP --
F6P: F
72.6 9.6 10.9 104 50.4 11.9 4.6 55 85.2 11.0 13.3 146 14.9 10.6 19.5 207 75.6 7.2 14.3 102
108 8.3 10.2 85 70.8 2.9 57.0 164 89.4 8.1 13.3 108
136' 8.0 14.1 112 97.2 9.0 19.7 178
-
- Ratios
The conversion of fructose-l-phosphate to fructose-l ,6-diphosphate in muscle fractions occurs only in the presence of ATP.G In Table IX the ratio of utilization, FGP:FlP, in the different fractions varied from about
6 It seemed possible that a mutase type of reaction was involved, fructose-l- phosphate -+ fructose-6-phosphate, followed by fructose-6-phosphate + ATP -+ fructose-l, 6-diphosphate. In such a reaction fructose-l, 6-diphosphate might be expected to act as coeneyme, in analogy to the coenayme effect of glucose-1,6-di- phosphate in the phosphoglucomutase reaction (22,23). However, no disappearance of fructose-l-phosphate could be detected when ATP was omitted and fructose-1,6- diphosphate was added. The fructose-l-phosphate was synthesized according to Raymond and Levene (24).
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from
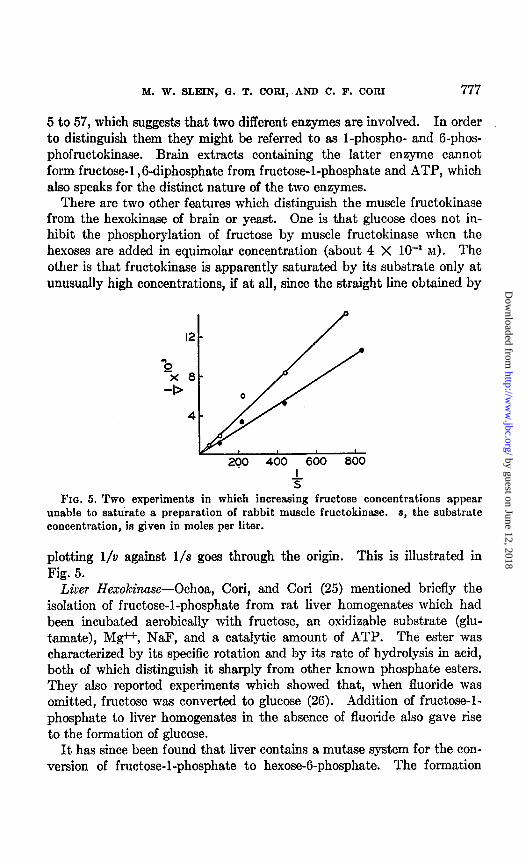
M. W. SLEIN, G. T. CORI, AND C. F. CORI 777
5 to 57, which suggests that two different enzymes are involved. In order to distinguish them they might be referred to as 1-phospho- and G-phos- phofructokinase. Brain extracts containing the latter enzyme cannot form fructose-l ,6-diphosphate from fructose-l-phosphate and ATP, which also speaks for the distinct nature of the two enzymes.
There are two other features which distinguish the muscle fructokinase from the hexokinase of brain or yeast. One is that glucose does not in- hibit the phosphorylation of fructose by muscle fructokinase when the hexoses are added in equimolar concentration (about 4 X 10e3 M). The other is that fructokinase is apparently saturated by its substrate only at unusually high concentrations, if at all, since the straight line obtained by
200 400 600 800 I
FIG. 5. Two experiments in which increasing fructose concentrations appear unable to saturate a preparation of rabbit muscle fructokinase. s, the substrate concentration, is given in moles per liter.
plotting l/u against l/s goes through the origin. This is illustrated in Fig. 5.
Liver Hexokinase-Ochoa, Cori, and Cori (25) mentioned briefly the isolation of fructose-l-phosphate from rat liver homogenates which had been incubated aerobically with fructose, an oxidizable substrate (glu- tamate), Mg*, NaF, and a catalytic amount of ATP. The ester was characterized by its specific rotation and by its rate of hydrolysis in acid, both of which distinguish it sharply from other known phosphate esters. They also reported experiments which showed that, when fluoride was omitted, fructose was converted to glucose (26). Addition of fructose-l- phosphate to liver homogenates in the absence of fluoride also gave rise to the formation of glucose.
It has since been found that liver contains a mutase system for the con- version of fructose-l-phosphate to hexose-6-phosphate. The formation
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

778 YEAST AND ANJ.MAL HEXOKINASE
of fructose-l-phosphate has been demonstrated in l’at liver homogenates under anaerobic conditions with ATP as phosphate donor, and the enzyme which catalyzes this reaction has been characterized further. It is a fructokinase which resembles that found in muscle, since it does not act on glucose and since its activity is not inhibited by glucose. Finally, liver contains a glucokinase, as shown in Table X. The activity of this enzyme cannot be measured accurately in liver homogenates because of the presence of glucose-6-phosphatase which is incompletely inhibited by fluoride. Another source of error is the formation of maltose and other
TABLE X
Phosphorylation of Fructose and Glucose by Rat Liver Homogenate Incubated for 20 minutes at 30’ in the presence of 0.06 M NaF, 4 X 10-s M hexose,
5 X 10-s M ATP, and 8 X 10-a M MgClt in potassium-phosphate buffer, pH 7.6. The figures represent micrograms of hexose utilized per 2 to 2.5 ml. of reac- tion mixture containing 1 ml. of homogenate (equivalent to 200 mg. of liver). The livers of fasted rats were perfused with salt solution prior to homogenization in 0.9 per cent sodium chloride solution.
Experiment No. Fructose
1 2 3 4 5
;r
8t gt
230 375 320 375
0 0 0
55
-
.-
-
Glucose
65 0
25 100 140 75
102 125 150
* 0.25 to 0.45 fraction with (NH&SOc; fructose used before fractionation, 130 and 250 y, respectively.
t Rabbit.
reducing substances from glycogen by diastatic activity. As has been pointed out previously (27), diastatic activity is particularly marked in rat liver and cannot be completely eliminated by previous perfusion of the liver. For these reasons the values recorded in Table X are minimal values for glucokinase activity. Experimental details concerning these findings will be reported in a subsequent paper.
SUMMARY
1. Substrate-enzyme dissociation constants were determined for crystal- line yeast hexokinase with a spectrophotometric method. The order of
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

M. R. SLEIN, G. T. COIU, AND C. F. C0R.I 779
affinity for the enzyme was mannose > glucose > fructose, while the order of maximal velocity, on saturation with substrate, was fructose > glucose > mannose. Evidence was presented that the enzyme reacts with that form of fructose in solution which contains a free hydroxyl group on carbon 6 (furanose form, roughly 20 per cent of total fructose concentration in these experiments). The ATP-enzyme dissociation constant was found to vary with the type of hexose present as phosphate acceptor.
2. With brain and yeast hexokinase there was nearly 100 per cent in- hibition of fructose utilization by an equimolar concentration of either glucose or mannose, while fructose had little inhibitory effect on the utilization ,of the two aldo sugars. There was also mutual inhibition between the pair mannose-glucose, the former sugar being a stronger inhibitor than the latter. From the fact that Ki mannose was the same as K, mannose and Kd glucose the same as K, glucose, it was concluded that they act as substrates and inhibitors at the same center of the enzyme.
3. The first product of fructose phosphorylation with brain hexokinase was shown to be fructose-6-phosphate. Various fractionation procedures did not result in a significant change in the ratio of fructose to glucose utilization. It appears that the predominant type of hexokinase in brain is of the yeast type.
4. The fructokinase of liver and muscle which does not act on glucose is not inhibited by this sugar. Enzymes which act on fructose-l-phosphate (probably the first phosphorylation product of fructose by liver fructo- kinase) have been found in liver and muscle but not in brain. Liver and muscle were also shown to contain a glucokinase.
BIBLIOGRAPHY
1. Berger, L., Slein, M. W., Colowick, S. P., and Cori, C. F., J. Gen. Physiol., 29, 379 (1946).
2. Colowick, S. P., Cori, G. T., and Slein, M. W., J. Biol. Chem., 168, 533 (1947). 3. Slein, M. W., J. Biol. Chem., 186, 753 (1950). 4. Warburg, O., and Christian, W., Biochem. Z., 264,438 (1932). 5. Negelein, E., and Gerischer, W., Biochem. Z., 234, 289 (1936). 6. Lohmann, K., Biochem. Z., 233, 460 (1931). 7. Warburg, O., Christian, W., and Griese, A., Biochem. Z., 282, 1.57 (1935). 8. Nelson, N., J. Biol. Chem., 163,375 (1944). 9. Roe, J. H., J. Biol. Chem., 107, 15 (1934).
10. Horecker, B. L., and Kornberg, A., J. Biol. Chem., 176,385 (1943). 11. Gottschalk, A., Australian J. Exp. Biol. and Med. SC., 22,291 (1944). 12. Gottschalk, A., Biochem. J., 41,276 (1947). 13. Isbell, H. S., and Pigman, W. W., J. Res. Nat. Bur. Standards, 20,773 (1938). 14. Nelson, J. M., and Beegle, F. M., J. Am. Chem. Sot., 41, 559 (1919). 15. Gottschalk, A., Australian J. Exp. Biol. and Med. SC., 21, 139 (1943). 16. Gottschalk, A., Australian J. Exp. Biol. and Med. SC., 21, 133 (1943).
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

780 YEAST AND ANIMAL HEXOKINASE
17. Gottschalk, A., Australian J. Exp. Biol. and Med. SC., 23, 261 (1945). 18. Gottschalk, A., Biochem. J., 41,478 (1947). 19. Hopkins, R. H., Biochem. J., 26, 245 (1931). 20. Meyerhof, O., and Wilson, J. R., Arch. Biochem., 19, 532 (1948). 21. Cori, G. T., Slein, M. W., and Cori, C. F., J. Biol. Chem., 173, 605 (1948). 22. Leloir, L. F., Trucco, R. E., Cardini, C. E., Paladini, A., and Caputto, R., Arch.
Biochem., 19, 339 (1948). 23. Sutherland, E. W., Cohn, M., Posternak, T., and Cori, C. F., J. Biol. Chem.,
180, 1235 (1949). 24. Raymond, A. L., and Levene, P. A., J. Biol. Chem., 83, 619 (1929). 25. Cori, C. F., Biol. Symposia, 6, 131 (1941). 26. Cori, C. F., in A symposium on respiratory enzymes, Madison (1942). 27. Cori, G. T., Cori, C. F., and Schmidt, G., J. Biol. Chem., 129, 629 (1939).
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

CoriMilton W. Slein, Gerty T. Cori and Carl F.
ANIMAL TISSUESHEXOKINASE FROM YEAST AND
A COMPARATIVE STUDY OF
1950, 186:763-779.J. Biol. Chem.
http://www.jbc.org/content/186/2/763.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/186/2/763.citation.full.haccessed free atThis article cites 0 references, 0 of which can be by guest on June 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























