Embed Size (px)
Citation preview

·霍乱· 图 8 表 3 参 16 占 6 面 发排稿
霍乱弧菌双精氨酸转运系统基因共转录和同源性分析
朱召芹 陈海丽 周海健 景怀琦 闫梅英 宰淑蓓 蔡金凤 胡芸文 阚飙
DOI:10.3760/cma.j.issn.0253-9624.2015.02.
基金项目:国家“十二五”科技重大专项(2012ZX10004-221、2013ZX10004221-004);
国家自然科学基金青年基金(31200108);上海市卫生局青年基金(20124467);2013年上
海市公共卫生临床中心启动基金
作者单位:201508 上海市公共卫生临床中心病原体应急检测与生物安全部(朱召芹、
陈海丽、胡芸文),医学检验科(宰淑蓓、蔡金凤);中国疾病预防控制中心传染病预防控
制所(周海健、景怀琦、闫梅英、阚飙)
通信作者:阚飙,Email:[email protected]
【摘要】目的 研究霍乱弧菌双精氨酸转运系统(twin-arginine translocation system,
Tat)基因簇各个基因的共转录情况,并确定参与Tat转录基因簇构成。方法 选择霍乱弧菌
El Tor型菌株N16961和tatABC基因缺失株N169-dtat进行Tat基因簇及上下游基因,即上游蛋
白质生物合成ubi aarF基因和下游细胞色素C551过氧化物酶cyt551基因进行转录分析。选择
霍乱弧菌及其他弧菌属菌株,通过PCR产物测序和序列比对,对Tat转运系统的同源性进行比
较,进行Tat基因簇各基因聚类分析。结果 Tat基因簇4个基因(tatA、tatB、tatC和 tatE)
均可以共转录。Ubi aarF基因可以与tatA及tatB共转录。Cyt551基因与上游的4个基因均不
共转录;通过N16961 Tat基因缺失株总RNA逆转录分析发现,虽然ubi aarF与tatA、tatB的
共转录由于基因缺失被阻断,但该基因仍可以被分别转录。不同型别的霍乱弧菌和其他弧菌
属的菌株tatA、tatB、tatC、tatE和tatABC基因聚类分析结果表明,Tat基因簇的种属变异
性很小,是一段很保守的序列。结论 霍乱弧菌ubi aarF基因与Tat基因簇可能共同受调控因
子的作用;Tat基因在弧菌属中属于保守结构。
【关键词】弧菌,霍乱;基因;序列同源性
Analysis of twin-arginine translocation system gene homology and
transcription in Vibrio species Zhu Zhaoqin*, Chen Haili, Zhou Haijian, Jing

Huaiqi, Yan Meiying, Zai Shubei, Cai Jinfeng, Hu Yunwen, Kan Biao. *Pathogen Diagnosis and
Biosafety department, Shanghai Public Health Clinical Center, Fudan University, Shanghai
201508, China
Corresponding author: Kan Biao,Email: [email protected]
【Abstract】Objective To determine the function of twin-arginine translocation
system(Tat)and gene cluster in Vibrio strains and to analyze the homology of tat gene
cluster among different Vibrio spp. strains based on N16961 and tatABC mutant
strains N169-dtat. Methods Different serotypes of biotype strains of Vibrio spp. were
selected to detect the transcription of 4 genes of Tat transport system and upstream ubi
aarF gene and downstream cyt551 gene by the total RNA reverse transcription and
homologicity of the gene cluster by sequencing analysis. Results Our results showed
that the 4 genes of tat cluster (tatA, tatB, tatC, and tatE) were intragenic and
co-transcribed. We found that ubi aarF gene could be co-transcribed with tatA, tatB,
but not with tatC. The electron transport chain and energy metabolism-related genes,
cytochrome C551 peroxidase gene, and 4 genes located at upstream of tatABC operon
were not transcribed with tatABC. Although the co-transcription between ubi aarF
and tatAB was blocked in N169-dtat strain, they were still transcribed separately.
Homologous analysis of genes of tat cluster in different types of V. cholerae showed
that tat gene cluster was a very conservative. Conclusion The ubi and aarF gene
might be co-transcribed with genes of tat cluster in V. cholerae, which and the close
relationship showed that they might play a key function in V. cholerae.
【Key words】Vibrio cholerae; Genes; Sequence homology
双精氨酸转运系统(twin-arginine translocation system,Tat)是 1997 年 Settles 等
[1]发现的存在于植物类囊体膜中的蛋白质运输系统,运输底物为完全折叠蛋白质并结合一定
的氧化还原因子[2],与经典的分泌途径转运系统(Sec 转运系统)不同。目前,已经发现 Tat
系统主要基因在许多细菌中存在,包括大肠埃希菌、幽门螺杆菌、鼠疫耶尔森菌、沙门菌、
铜绿假单胞菌、结核分枝杆菌、霍乱弧菌、流感嗜血杆菌、葡萄球菌等 [3-5]。
我们前期通过对基因同源比较分析,推定霍乱弧菌存在Tat转运系统的主要基因。但是,
霍乱弧菌Tat转运系统的操纵子和转录起始位点还未知,其结构基因与上下游基因的共转录

关系需要明确,同时,弧菌属中是否具有Tat系统、其序列保守性及与弧菌不同种分化背景
下的遗传进化关系也值得分析。
材料与方法
1.菌株:为了确定Tat基因簇的转录,选择不同血清型和生物型的霍乱弧菌代表株进行
Tat基因簇4个基因的共转录分析[15]。首先确定Tat基因簇各基因之间是否发生共转录,进一
步确定霍乱弧菌的Tat基因簇的功能单元组成,除此之外,还要根据Tat基因簇的位置,确定
上下游基因,即与细菌蛋白生物合成与能量代谢有关的蛋白质的生物合成ubi aarF基因
(VC0085)和细胞色素C551过氧化物酶cyt551基因(VC0089)是否参与共转录,寻找Tat基因簇
和上下游基因之间可能存在的关联转录。同时选择不同霍乱弧菌代表株(包括O1古典型、
O139、O1 El Tor、O22、非凝集弧菌等)和拟态弧菌(V. mimicus)、弗尼斯弧菌(V. furnissii)、
河弧菌(V. fluviolis)、副溶血弧菌(V. parahaemolyticus)、创伤弧菌(V. vulnificus)
等其他弧菌,进行Tat基因簇4个基因比较,初步分析弧菌属携带Tat基因状况及Tat基因簇的
序列遗传差异。所用的菌株见表1。
表1 实验用菌株的菌属特征、血清型、毒素产生情况及来源描述
编码 菌属 特征 霍乱毒素 来源
A VP20(河弧菌) / - 本实验室保存
B Vm7(拟态弧菌) / - 本实验室保存
C Vv1(创伤弧菌) / - 本实验室保存
D Vms3(麦氏弧菌) / - 本实验室保存
E Vfns1(弗尼斯弧菌) / - 本实验室保存
F1 ICDC-VP133(副溶血弧菌) / - 秦皇岛
F2 ICDC-VP24(副溶血弧菌) / - 印度
G 569B(霍乱弧菌) O1 古典生物型 + 本实验室保存
H MO45(霍乱弧菌) O139 + 本实验室保存
I Wujiang-2(霍乱弧菌) O1 埃尔托生物
型
+ 本实验室保存
J JS32(霍乱弧菌) O1 埃尔托生物
型
- 本实验室保存
K 4-7(霍乱弧菌) O1 埃尔托生物 - 本实验室保存
型
L 19-22(霍乱弧菌) O1 埃尔托生物
型
- 本实验室保存
M N53(霍乱弧菌) 非凝集弧菌 - 本实验室保存
N 169-68(霍乱弧菌) O22 - 本实验室保存
O B4(霍乱弧菌) O139 - 本实验室保存
P 94001(霍乱弧菌) 0139 - 本实验室保存
N16961 N16961(霍乱弧菌) O1 埃尔托生物
型
+ 本实验室保存
N169-dtat tatABC 变异 / + 本实验室构建
注:“+”表示产生霍乱毒素,“–”表示不产生霍乱毒素;“/”表示未进行血清分型
2.试剂:胰蛋白胨、酵母提取物购自上海普迈生物科技有限公司;rTaq聚合酶购自宝
生物工程(大连)有限公司;染色体提取试剂盒、细菌总 RNA 提取试剂盒 RNeasy Mini Kit
购自凯杰生物技术(上海)有限公司;Super ScripTM Ⅲ购自上海普迈生物科技有限公司。
表 2 霍乱弧菌双精氨酸转运系统基因共转录实验用引物
引物名称 序列(5'~3') 靶基因 基因预测
名称
长度(bp) 与前一基因间
隔(bp)
82-P1 GAATCTGGGCTGCGCGAAGGG VC0082
83-P1 CATTCTCGATTTGGGCGGCGG VC0083
84-P1 CTATCTGGCATTAAACCTCTC VC0084 623
85-P1 GTTTCGCGACAGTTTCTTCCA VC0085 ubi aarF 1 634
85-P2 CCAAGCCTTCGACATACAGCA VC0085
86-P1 CTTATCATTGCAGTGATCGTG VC0086 tatA 248 60
86-P2 GACGCATTGGTTTTGGCCTGC VC0086
87-P1 TCGCTATTGTGGCGCTGGTGG VC0087 tatB 401 4
87-P2 TGCGACGCGCTATCGAGGCGA VC0087
87-P3 AGCGATTACCCCATGCGATTC VC0087
87-P4 TCACTGCTCGGCGTTGCGGCG VC0087
88-P1 CAACCTTTGATCAGCCATTTG VC0088 tatC 752 89
88-P2 ATAAAAACGAGCGAAGAACAA VC0088
89-P1 GAGTTTGAAAAGACCTTAGTG VC0089 cyt551 1 001 312
89-P2 GCCACTATCTTACCGTTTTCA VC0089
32-P1 GCTACCGTCAGTCACTTCTTT VC0532 DNA bin 692

32-P2 GTAGCACTCGGTCACCACTCC VC0532
33-P1 CTCTTAATCTTGGGGTGTATT VC0533 tatE/tatA
2
236
33-P2 CTCTTGCTCAATAGCGAAGCT VC0533
34-P1 GTACCACGCGATCTAACCACT VC0534 dedA 675
34-P2 ATCCAGAATTAGCCAGCAAGT VC0534
3.总 RNA 提取:按照试剂盒的操作步骤提取。电泳和 PCR 验证是否有 RNA 的降解和(或)
DNA 的污染,确定 RNA 的浓度和纯度。提取样品的总 RNA 后,根据 RNA 的凝胶电泳图来判断
RNA 的质量。RNA 的质量判断标准是有清晰的 23 S 和 16 S 条带,无降解,且肉眼观察 23 S
条带亮度约是 16 S 的 1~2倍。细菌 23 S 和 16 S 大小分别约为 2.9 kb 和 1.5 kb;同时选
用 1~2 对引物进行 PCR 扩增,以排除 DNA 污染。
4.PCR 检测:10 µmol/L 上下游引物各 1 µl,总 RNA 10 µl,dNTP 4 µl,加水补至 20 µl。
65 ℃放置 5 min 后冰上放置 1 min 以上,向以上体系中继续加入:5×缓冲液 4 µl、0.1 mol/L
二硫苏糖醇(DTT) 和 Super ScripTMⅢ各 1 µl。50 ℃ 1 h 后 70 ℃ 15 min。向以上体系
中加入 RNase H,37 ℃作用 30 min,以去除多余的 RNA。10×缓冲液、dNTP、10 µmol/L
上下游引物、Taq 酶、模板各 2 µl,补三蒸水(TDW)至 20 µl。PCR 扩增参数:94 ℃ 5 min,
94 ℃ 35 s,50 ℃ 35 s,72 ℃ 2 min,35 个循环。
5.数据分析:使用分子进化遗传分析软件 Mega 5.0 采取 Clustalw 方法进行基因进化
树和蛋白序列比对分析,构建进化树。共转录分析用 Photoshop 软件进行展示。
结 果
1.N16961 全序列上 Tat 转运系统基因簇的位置关系:通过已测序的霍乱弧菌标准株
N16961 基因组与大肠埃希菌 tatABCD 操纵子同源比较分析,推定霍乱弧菌 N16961 存在 Tat
转运系统的主要基因。与其他革兰阴性菌相比,El Tor 霍乱弧菌 N16961 的 Tat 系统的基因
结构是独特的。tatA(VC0086)、tatB(VC0087)和 tatC(VC0088)位于Ⅰ号染色体上,
与大肠埃希菌不同的是,在霍乱弧菌 N16961Ⅰ号染色体上没有发现与大肠埃希菌 tatE同源
性高的基因片段,与大肠埃希菌 tatE基因同源性高的 tatA2 基因(VCA0533)位于霍乱弧菌
的小染色体上(Ⅱ号染色体);tatABC 基因簇附近没有 tatD 基因。与细菌蛋白生物合成与
能量代谢有关的蛋白质的生物合成 ubi aarF 基因 (VC0085)和细胞色素 C551 过氧化物酶
cyt551基因(VC0089)位于 tatABC基因簇的上下游,见表 2。
2.霍乱弧菌 El Tor 菌株 N16961 Tat基因簇共转录分析:以 TDW 和用于逆转录同管 RNA

作为阴性对照。以野生株 N16961 为模板,各基因内部均正常转录(图 1);以野生株 N16961
为模板进行共转录分析,Ubia arF 与下游的 tatA 和 tatB 共转录,Cyt551 与 tatB 和 tatC
均不共转录(图 2);以 N169-dtat 为模板进行共转录分析,因 tatABC 基因簇缺失,所以
tatABC基因不共转录,同时也与下游的 Cyt551不共转录(图 3)。但 Ubi aarF 和 Cyt551
基因没有受 tatABC基因缺失影响,还可以正常转录。从图 1~3 和表 3可以看出:(1)cyt551
基因与上游的 4 个基因均不共转录;(2)tatA、tatB、tatC 基因共转录;(3)Ubi aarF 基
因可以与 tatA、tatB共转录。图 3 是以 N169-dtat 提取的总 RNA 进行逆转录的结果,可以
看出 VC0085 虽然与 tatA、tatB 的共转录被阻断,但仍可以自身转录。因 VC0085 参与了共
转录,因此,需再设计引物察看上游 2~3 个基因是否也参与共转录,以期找到真正的转录
起始。VC0084 和 VC0090 与他们之间的序列都不共转录,图中未做标示。
表 3 埃尔托生物型霍乱弧菌 N16961 双精氨酸转运系统基因簇共转录分析
上游引物所在
的基因
下游引物所在
的基因
引物组合 模板 cDNA N16961 染色体
(阳性对照)
ubi aarF cyt551 85-P1,89-P2 N16961 - +
ubi aarF tatA 85-P1,86-P2 N16961 + +
tatA tatC 86-P1,88-P2 N16961 + +
tatA tatB 86-P1,87-P2 N16961 + +
tatB tatC 87-P1,88-P2 N16961 + +
tatC cyt551 88-P1,89-P2 N16961 - +
tatA cyt551 86-P1,89-P2 N16961 - +
tatB cyt551 87-P1,89-P2 N16961 - +
ubi aarF tatB 85-P1,87-P2 N16961 + +
ubi aarF tatC 85-P1,88-P2 N16961 +
DNA-bin tatE 32-P1,33-P2 N16961 - +
DNA-bin dedA 32-P1,34-P2 N16961 - +
tatE dedA 33-P1,34-P2 N16961 - +
VC0082 ubiaarF 82-P1,85-P2 N16961 - +
VC0082 tatA 82-P1,86-P2 N16961 - +
VC0082 tatB 82-P1,87-P2 N16961 - +

VC0082 tatC 82-P1,88-P2 N16961 - +
VC0083 ubiaarF 83-P1,85-P2 N16961 - +
VC0083 tatA 83-P1,86-P2 N16961 - +
VC0083 tatB 83-P1,87-P2 N16961 - +
VC0083 tatC 83-P1,88-P2 N16961 - +
VC0084 ubiaarF 84-P1,85-P2 N16961 - +
VC0084 tatA 84-P1,86-P2 N16961 - +
VC0084 tatB 84-P1,87-P2 N16961 - +
VC0084 TatC 84-P1,88-P2 N16961 - +
VC0084 VC0084 84-P1,84-P2 N16961 + +
ubiaarF ubiaarF 85-P1,85-P2 N16961 + +
dtatA dtatA 86-P1,86-P2 N16961 + +
dtatB dtatB 87-P1,87-P2 N16961 + +
dtatC dtatC 88-P1,88-P2 N16961 + +
cyt551 cyt551 89-P1,89-P2 N16961 + +
dtatE dtatE 33-P1,33-P2 N16961 + +
VC0084 VC0084 84-P1,84-P2 N169-dtat + +
ubiaarF ubiaarF 85-P1,85-P2 N169-dtat + +
dtatA dtatA 86-P1,86-P2 N169-dtat - -
dtatB dtatB 87-P1,87-P2 N169-dtat - -
dtatC dtatC 88-P1,88-P2 N169-dtat - -
cyt551 cyt551 89-P1,89-P2 N169-dtat - -
dtatE dtatE 33-P1,33-P2 N169-dtat + +
DNA-bin DNA-bin 32-P1,32-P2 N169-dtat + +
dedA dedA 34-P1,34-P2 N169-dtat + +
注:“+”表示 cDNA 检测有目的条带;“-”表示 cDNA 检测没有发现目的条带

tatA、tatB、tatC和 tatA2(tatE)4 个基因均可以基因内转录;tatABC基因簇上游基因 ubi aarF和下游基
因 cyt551可以基因内转录;tatA2 的上游基因 VC0032 和 VC0034 也可以基因内转录
图 1双精氨酸转运系统基因簇以野生株 N16961 为模板的基因内转录分析
ubi aarF基因可以与 tatA和 tatB两个基因共转录,但不能与 tatC基因共转录;tatA和 tatB基因可以共
转录,tatA、tatB和 tatC基因也可以供转录;tatB和 tatC基因均不能与 cyt551 基因共转录
图 2双精氨酸转运系统基因簇以野生株 N16961 为模板的基因间共转录分析
ubi aarF和 cyt55 基因可以基因内转录,但与 tatA、tatB和 tatC 之间的基因没有共转录
图 3双精氨酸转运系统基因簇以 tatABC基因缺失株 N169-dtat 为模板的共转录分析
3.不同弧菌 Tat 系统基因序列比对结果:总体上霍乱弧菌各血清型和生物型的同源性较
高,Tat基因簇的种属变异性很小,是一段很保守的序列。相同血清型和生物型的霍乱弧菌
的 tat 基因簇聚类分析并没有完全聚集在一起,提示霍乱弧菌 tat 系统的功能与霍乱弧菌的
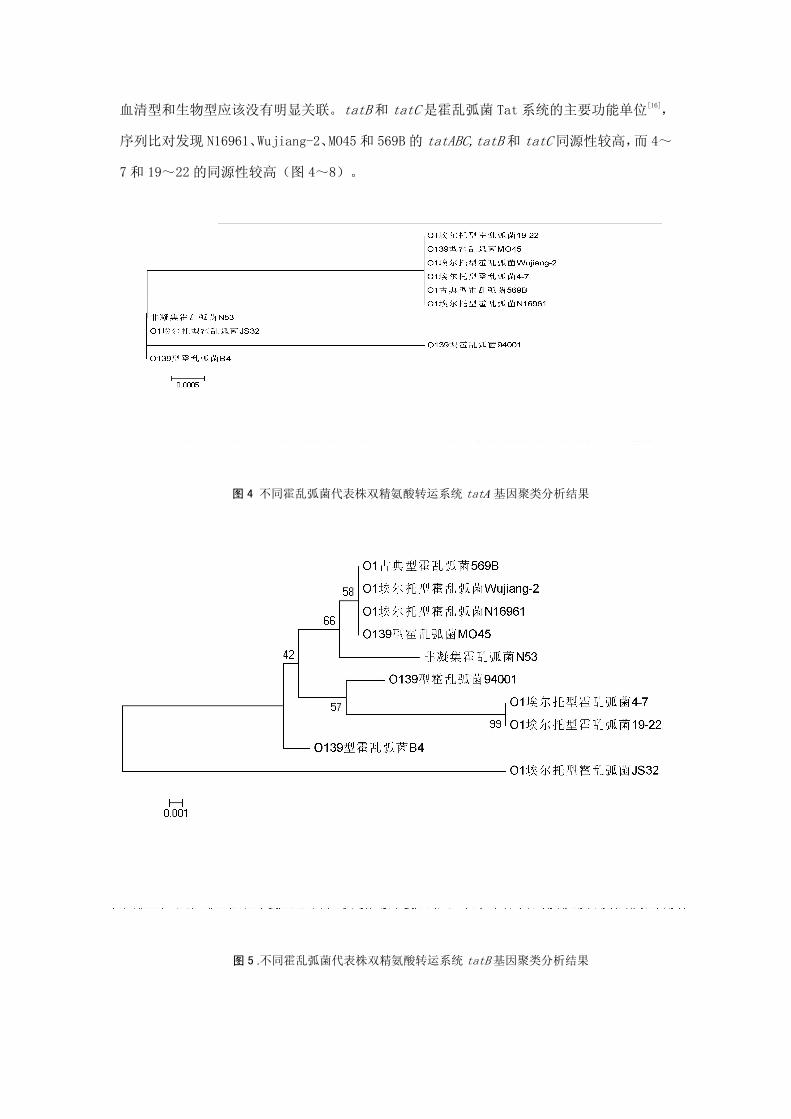
血清型和生物型应该没有明显关联。tatB和 tatC是霍乱弧菌 Tat 系统的主要功能单位[16],
序列比对发现 N16961、Wujiang-2、MO45 和 569B 的 tatABC,tatB和 tatC同源性较高,而 4~
7和 19~22 的同源性较高(图 4~8)。
图 4 不同霍乱弧菌代表株双精氨酸转运系统 tatA基因聚类分析结果
图 5 .不同霍乱弧菌代表株双精氨酸转运系统 tatB基因聚类分析结果

图 6 不同霍乱弧菌代表株双精氨酸转运系统 tatC 基因聚类分析结果
图 7 不同霍乱弧菌代表株双精氨酸转运系统 tatABC基因聚类分析结果

图 8 不同霍乱弧菌代表株双精氨酸转运系统 tatE基因聚类分析结果
讨 论
Tat 系统是存在于许多病原菌中有重要毒力机制的特殊蛋白转运系统。Tat 系统不仅在
细菌生理功能方面,而且在某些病原微生物的致病性中也发挥作用[2-7],还参与了很多重要
的机制,例如细胞膜的形成、生物膜的形成、电子运输链的排列、在很多环境压力下的生存
以及运动性等[5]。Tat 运输系统的底物在功能上通常是构成氧化呼吸链,尤其是厌养条件下
呼吸链的重要组成成分,为细菌能量代谢所必需[2]。Tat 系统的缺陷可能会导致包括生长、
运动性和一些毒性因子分泌在内的多种表型缺陷,因此,Tat 系统是细菌毒性决定因子[3,8-14]。
前期通过分析霍乱弧菌是否转运 Tat 运输底物三甲胺-N-氧化酶 TMAO,发现霍乱弧菌中
存在功能性的 Tat 系统。tatABC 基因簇缺失株和野生株在运动性和鞭毛合成及在需氧和厌
氧培养条件下的生长状况都没有发现差别。然而,Tat基因缺失株的生物膜形成能力下降,
而且在乳鼠肠道内的黏附定居能力及与 HT-29 细胞体外黏附能力都下降。这些结果表明,Tat
机制在对霍乱弧菌的致病性中也发挥重要的作用。该系统与霍乱弧菌的生物膜形成、黏附定
居和毒力有关[3]。但由于霍乱弧菌 Tat基因簇与大肠埃希菌有差异,比如 Tat基因簇分布在
不同的染色体,没发现 tatD 基因,转录起始位点未知等,同时,霍乱弧菌与其他弧菌的 Tat
系统的亲缘关系如何及作用机制是否相似等问题都值得进一步探讨。是否其他非致病性弧菌
的 Tat 系统也存在类似的机制需要进一步探索。
本研究发现,Tat 基因簇各基因(tatA、tatB 和 tatC)均可以内转录,而且可以相互
共转录。由于 ubi aarF (VC0085)可以与 Tat 基因簇共转录,因此,进一步检测上游 2~3
个基因,寻求转录起始位点。研究发现,ubi aarF 基因可以与 tatA和 tatB共转录,通过
N16961 Tat 基因缺失株总 RNA 逆转录分析发现,虽然 ubi aarF 与 tatA、tatB 的共转录被

阻断,但该基因仍可以自身转录。VC0085 与 Tat基因簇可以共转录,而上游的 VC0084 与 Tat
基因簇之间的序列都不共转录,已在结果描述上增加这句,推测 ubi aarF (VC0085)基因可
能是霍乱弧菌 Tat 转录起始位点。cyt551 基因与上游的 4 个基因均不共转录。提示 ubi aarF
与 tatABC 基因簇的功能或对环境的反应是相关联的,这与 E. coli的 Tat 转运系统 tatABCD
基因簇形成一个单独操纵子进行转录有不同。
不同血清型的霍乱弧菌 tatA、tatB、tatC、tatE基因聚类分析结果表明,Tat基因簇
的种属内变异性很小,是一段很保守的序列。霍乱弧菌中不同菌株间的 Tat 系统各基因比较,
显示了不同群型(古典生物型和埃尔托生物型)、产毒非产毒菌株之间的差异,同时,未发
现 Tat基因簇位于可水平转移的成分中,提示 Tat 基因簇与不同群型菌株的基因组分化是同
步的。Tat基因簇可能是弧菌本身固有的,是发挥关键功能的区域。
参考文献
[1] Settles AM, Yonetani A, Baron A, et al. Sec-independent protein translocation by the maizeHcf106 protein[J]. Science, 1997, 278(5342): 1467-1470.[2] Palmer T, Berks BC. The twin-arginine translocation(Tat)protein export pathway[J]. Nat Rev,Microbiol, 2012, 10(7): 483-496.[3] Zhang L, Zhu Z, Jing H, et al.. Pleiotropic effects of the twin-arginine translocation system onbiofilm formation, colonization, and virulence in Vibrio cholera[J]. BMC Microbiol, 2009, 9:114.[4] Wu LF, Ize B, Chanal A, et al. Bacterial twin-arginine signal peptide-dependent proteintranslocation pathway: evolution and mechanism[J]. J Mol Microbiol Biotechnol, 2000, 2(2):179-189.[5] Ochsner UA, Snyder A, Vasil AI, et al. Effects of the twin-arginine translocase on secretion ofvirulence factors, stress response, and pathogenesis[J]. Proc Natl Acad Sci U S A, 2002, 99(12):8312-8317.[6] Ding Z, Christie PJ. Agrobacterium tumefaciens twin-arginine-dependenttranslocation is important for virulence,flagellation,and chemotaxis but not type Ⅳ secretion[J].J Bacteriol, 2003, 185(3): 760-771.[7] Voulhoux R, Ball G, lze B, et al. Involvement of the twin-arginine translocation system inprotein secretion via the type Ⅱ pathway[J]. EMBO J, 2001, 20(23): 6735-6741.[8] Sargent F, Bogsch EG, Stanley NR, et al. Overlapping functions of components of a baterialSec-independent protein export pathway[J]. EMBO J, 1998, 17(13): 3640-3650.[9] Berks BC. A common export pathway for proteins binding complex redox cofactors?[J]. MolMicrobiol, 1996, 22(3): 393–404.
[10] Simon R, Priefer U, Piihler A. A broad host range mobilization system for in vivo genetic
engineering: transposon mutagenesis in gram-negative bacteria[J]. Bio Technology, 1983, 1:
784-791.
[11] Müller M. Twin-arginine-specific protein export in Escherichia coli[J]. Res Microbiol, 2005,156(2): 131-136.

[12] Palmer T, Berks BC. Moving folded proteins across the bacterial cell membrane[J].Microbiology, 2003,149(Pt 3): 547-556.[13] Lavander M, Ericsson SK, Bröms JE, et al. The twin arginine translocation system is essentialfor virulence of Yersinia pseudotuberculosi[J]. Infect Immuni, 2006, 74(3):1768-1776.
[14] 张丽娟,高守一,景怀琦, 等. 霍乱弧菌 Tat 蛋白运输系统基因簇的确定与功能阻断分
析[J].遗传学报,2002, 29(10): 936-940.
[15] 王鸣, 李孝权,莫自耀, 等. 2001—2005年广州地区霍乱弧菌主要致病相关基因特征分
析[J].中华预防医学杂志, 2006, 40(4): 257-261.
[16] 朱召芹, 景怀琦, 陈海丽, 等. El Tor霍乱弧菌双精氨酸转运系统功能单位的分析研究
[J]. 微生物与感染, 2012, 7(2): 99-104.
(收稿日期:2014-07-17)
(本文编辑:吕相征)

![Ä .T )ÅÆ...- 70 - Ä .T )ÅÆ .T )ÅÆ tu ¿À%Mn@ o k#$%&' 5 9:&] cd õö ¢£ xy ¿7L lm &X½ 9 a A AL7 . 2014 26 ³½ä \] 9:& ¸ J - Á ¢£ :& a&]o>" d ¯ 9 { ¸ J - Á Y N](https://img.dokumen.tips/doc/110x75/60cc04352ee93205654adaa7/-t-70-t-t-tu-mn-o-k-5-9.jpg)


![Â)ÃYÄ ÅÆ/ÇNÈ!É - IUMA - ULPGC · Â)ÃyÄ ÅÆ/ÇnÈ!É ÊdÇtË\Ì7ÍÎ\ÃgÏ*Í@Ð'Å!ÌpÑ7Ç]ÌÌbÍ@ÒrÇ Î Æ/Ì7Î\ÃyÎ+Ë\Ì7ÍÑjÃrÇ Î Ó Ô Õ¬ÖwÓ Ôd×ØÇ]ÒyÌ](https://img.dokumen.tips/doc/110x75/5ba15c2009d3f26b6b8c335b/aayae-aacnee-iuma-aayae-aacnee-edctei7iiagiidaipn7ciibiorc.jpg)



![5 PowerPoint 2010 · 5 245 ˛˚ ˜ˇˆPowerPoint 2010 5-4 PowerPoint 20100]VWX 5-5 åæ/ÉCfiæ Ú fl Ya[ åæ/ fl Yb[P ª fl 5-6 åæ//Pª fl åæ/ÉCƒ](https://img.dokumen.tips/doc/110x75/5f37cdc70738bb36b27a8168/5-powerpoint-5-245-oepowerpoint-2010-5-4-powerpoint-20100vwx-5-5-ci.jpg)



![· Þ23¯ Åæ ¼ U qj ÍU bwp§Ýzå j Øw¯ Å µ©ßï ` o X i ^ M {¢ Ð ï Ä U ù s M q ¡ UpV d wp] « Xi^ M {£ Þ23¯ Åæ ¼ U qj ÍU](https://img.dokumen.tips/doc/110x75/5c02549b09d3f20a538df19e/-b23-aae-u-qj-iu-bwpyza-j-ow-a-ssi-o-x-i-m-d-i.jpg)










![Â)ÃYÄ ÅÆ/ÇNÈ!É - IUMA - ULPGCjrsendra/Docencia/Electronica_Basica/download/exam... · Â)ÃyÄ ÅÆ/ÇnÈ!É ÊdÇtË\Ì7ÍÎ\ÃgÏ*Í@Ð'Å!ÌpÑ7Ç]ÌÌbÍ@ÒrÇ Î Æ/Ì7Î\ÃyÎ+Ë\Ì7ÍÑjÃrÇ](https://img.dokumen.tips/doc/110x75/5e71d43c3825b25628497edb/fy-n-iuma-jrsendradocenciaelectronicabasicadownloadexam.jpg)





