Embed Size (px)
Citation preview

2008, Sankey, J.T. and S. Baszio, eds. Vertebrate Microfossil Assemblages Their Role in Paleoecology and Paleobiogeography. Indiana University Press.
Vertebrate Paleoecology from microsites, Talley Mountain, upper Aguja Formation (Late
Cretaceous), Big Bend National Park, Texas
Julia T. Sankey
Department of Physics and Geology, California State University, Stanislaus
Turlock, California 95382. [email protected]
1. Abstract
The Late Cretaceous vertebrates from Big Bend National Park, Texas are some of the
southern most from the Western Interior of North America. Considerably less is known about
these southern vertebrates compared to those from northern areas like Alberta and Montana.
Numerous vertebrate fossils were collected from microsites in the Aguja Formation (late
Campanian), adding new information about the distinct southern Late Cretaceous ecosystems.
Many vertebrates common in Big Bend are rare or absent from northern areas, reflecting
paleoclimatic and paleoenvironmental differences, with periodic aridity occurring earlier in Big
Bend than in northern areas.
Key Words
1

Aguja, Cretaceous, Campanian, Big Bend National Park, Texas, vertebrates, dinosaurs,
mammals, paleoclimate, magnetostratigraphy.
2. Introduction
2.1. Southern Biogeographic Province
During the late Campanian to early Maastrichtian, Big Bend was part of a southern
dinosaur biogeographic province, the “Kritosaurus” fauna (LEHMAN 1997), within the Western
Interior of North America (Fig. 1). The southern province, from southern Colorado to the south,
was characterized by the Normapolles palynoflora, and had a warm climate with open-canopy
woodlands (WHEELER & LEHMAN 2000; LEHMAN & WHEELER 2001). In contrast, the northern
province, from southern Colorado to the north, was characterized by the Aquillapollenites
palynoflora, and had a temperate and more humid climate, with a more closed-canopy forest
(NICHOLS & SWEET, 1993). Rainfall and temperature differences were the main factors that
produced these north-south biogeographic patterns (BAGHAI 1996; LEHMAN 1997). (For a
differing opinion on Late Cretaceous provinces, see SULLIVAN & LUCAS 2003.) Big Bend in the
late Campanian to mid-Maastrichtian was at ~35 degrees N paleo-latitude (ROBINSON-ROBERTS
& KIRSCHBAUM 1995), and the Western Interior Seaway was nearby (ATCHLEY et al. 2004).
(((Figure 1 near here)))
Considerably less is known about the southern biogeographic province compared to the
long-studied northern areas. Big Bend is critical in understanding the southern province because
it contains some of the southernmost Late Cretaceous vertebrate fossils in North America. There
are some important differences between the two. First, because Big Bend was relatively distant
from uplands in the late Campanian, the area had slower sedimentation rates compared to
2

northern areas, resulting in condensed stratigraphic and faunal records in Big Bend (LEHMAN
1991). Second, increased aridity, due to retreat of the Western Interior Seaway and uplift of
western mountains, occurred in Big Bend earlier (late Campanian) than in the north and was an
important influence on its paleocommunities. Evidence for aridity in Big Bend comes from
dinosaur bonebeds from the upper Aguja Formation that formed during periodic droughts severe
enough to cause marshes to dry up (DAVIES & LEHMAN 1989). Further evidence is from the
numerous carbonate-cemented and pedogenic nodule rich conglomeratic channel lag deposits in
the upper Aguja and Javelina Formations (SANKEY 1998; SCHIEBOUT et al. 1997).
This paleoenvironmental and paleoecological picture for Big Bend during the late
Campanian differs from contemporaneous northern areas such as Alberta. For example, the mid
to late Campanian-aged Dinosaur Park Formation of Alberta is an extremely fossil-rich unit that
was deposited under high rates of sedimentation and fossil burial by a high-sinuosity fluvial
system within an extensive mesic coastal lowland and under high subsidence rates in the Alberta
foreland basin (EBERTH & HAMBLIN 1993). In contrast, the Aguja Formation is considerably less
fossil-rich and had considerably slower rates of sedimentation and fossil burial due to lower rates
of subsidence within the Tornillo Basin. Additionally, although Alberta also experienced
periodically dry conditions, it was not until the late Maastrichtian. The Maastrichtian is known
for having the coolest and most variable climates of the Cretaceous (BARRERA 1994), with the
exception of two short Greenhouse events in the mid and late Maastrichtian (NORDT et al. 2003).
2.2. Importance of Microsites
Big Bend is a good example of how important microsite studies can be for areas that do
not have abundant fossils. Information about the Late Cretaceous vertebrate faunas from
3

southern North America and from west Texas in particular would be missing a great deal of
information without the fossils recovered from microsites. In Big Bend, large fossil fragments
and bones are easily seen, so turtles, crocodylians, and large dinosaurs are well documented from
this area. However, almost no information about Late Cretaceous mammals would be known
from Big Bend without the collection of microsites, especially by screenwashing (ROWE et al.
1992; SANKEY & GOSE 2001). Collection of microsites in Big Bend has also produced important
records of Late Cretaceous lizards, and these lizards are considerably different to northern ones
(ROWE et al. 1992; MILLER 1997; NYDAM et al. in press). Additionally, most of our knowledge
about Big Bend theropod dinosaurs (tyrannosaurids and small theropods), including dinosaur
eggshells and juveniles, comes from microsites (SANKEY 2001; 2005; WELSH 2004; 2005;
WELSH & SANKEY this volume).
Collection of microsites is also important because large sample sizes are produced which
allow detailed paleoecological analyses. For example, from Late Cretaceous microsites in
Alberta, paleocommunities changed as a response to changes in paleoenvironments (BRINKMAN
1990; PENG et al. 2001; SANKEY et al. 2005a). Collection of Late Cretaceous microsites in Big
Bend contributes important paleoecological information and allows similar paleoecological
analyses to be done for this southern area.
2.3. Aguja Formation
The Aguja Formation is a widespread sedimentary unit in Big Bend (Fig. 2). It is an
eastward thinning deposit composed of 135 to 285 meters of sandstones interbedded with shale
and lignite. Environments of deposition vary from marine, paralic, to inland floodplain (LEHMAN
1985). The upper shale member represents the last of the pre-Laramide tectonic sedimentation in
4

the area (LEHMAN 1991), and records the final marine regression of the Western Interior Sea
from west Texas (Regression 8 of KAUFFMAN 1977). The lower part of this member contains
carbonaceous mudstones, thin beds of lignite, and large siderite ironstone concretions
representing distributary channels, levees, crevasse splays, and poorly drained interdistributary
marshes and bays. The upper part of this member, with variegated mudstones and sandstones
containing conglomeratic lags of paleo-caliche nodules, represents fluvial environments within a
deltaic coastal floodplain and inland floodplain. A variety of well-developed paleosols formed on
the inland floodplain of the upper Aguja and overlying Javelina Formations, and their stage of
development has been linked to sea level fluctuations in the nearby Western Interior Seaway
(ATCHLEY et al. 2004).
Typical large vertebrates from the lower part of the upper Aguja Formation are the giant
crocodilian Deinosuchus riograndensis and the horned dinosaur Chasmosaurus mariscalensis;
the hadrosaur Kritosaurus sp. is more abundant higher in the upper Aguja Formation (LEHMAN
1985). All are known from quarries yielding associated skeletons. The age of the microsites
reported here was constrained by paleomagnetic analysis of the fine-grained sedimentary rocks.
Magnetostratigraphic correlations to the base of chronozone 32 (Fig. 2), constrain the microsites
to approximately 71 to 75 Ma (late Campanian to early Maastrichtian (SANKEY 1998; SANKEY &
GOSE 2001). Similar work constrains the upper Aguja Formation to the early Maastrichtian
(LEHMAN 1985, 1989, 1990; STANDHARDT 1986).
(((Figure 2 near here)))
2.4. Objectives of Study
The purpose of this study was to significantly increase our knowledge of this southern
fauna by collecting large samples of vertebrates from microsites from the upper Aguja Formation
5

in the Talley Mountain area. In particular, the goals were to: 1) determine the abundance of each
vertebrate group within each microsite; 2) determine the paleoecological trends through the
sampled section; and 3) provide a more detailed paleoecological reconstruction of the upper
Aguja Formation.
3. Methods
3.1. Fossil Collection and Preparation
From the Talley Mountain Area, fossils were collected from five horizons of carbonate-
cemented sandstone conglomerates containing numerous clasts of clay, pedogenic nodules, small
bones and teeth, and large fragments of wood and bones. Collected rocks (1753 kg) were
disaggregated by soaking in a 10-25% solution of acetic acid and screened through fine-mesh
screens with 1 mm maximum openings (methods described in SANKEY 1998). Matrix was sorted
with a dissecting microscope, and all identifiable fossils, even small fragments of teeth, were
counted (Table 1) and percentages were calculated (Fig. 4).
3.2. Curation
All fossils are curated in the LSU Museum of Natural Science Vertebrate Paleontology
Collections. VL, refers to vertebrate locality and V, refers to the particular vertebrate fossil
specimen number. A complete LSUMG catalog number for a vertebrate fossil includes both of
these numbers, separated by a colon; for example, 488:5566. Often only the second number is
used. Additionally, all specimens have been collected within Big Bend National Park, and have
been assigned NPS catalog numbers. Specimen and locality information will be available to
qualified researchers by written request to the author.
6

3.3. Fossil Identifications
Fossil identifications were made in part by comparisons to collections at Louisiana State
University Museum of Natural Science Vertebrate Paleontology Collections, Baton Rouge;
Royal Tyrrell Museum of Paleontology, Drumheller, Alberta; University of Alberta Laboratory
for Vertebrate Paleontology, Edmonton; University of California Museum of Paleontology,
Berkeley; University of Texas, Texas Memorial Museum’s Vertebrate Paleontology Laboratory,
Austin; University of Oklahoma, Oklahoma Museum of Natural History, Norman; and Weber
State University, Department of Geosciences, Ogden, Utah. Literature used to identify the fossils
were: ESTES 1964, BRYANT 1989, and WELTON & FARISH 1993 for fish; ESTES 1964, ESTES
1981, STANDHARDT 1986, BRYANT 1989, GAO & FOX 1996, MILLER 1997, and TOMLINSON 1997
for amphibians and reptiles; CURRIE et al. 1990, BASZIO 1997, and SANKEY et al. 2002 for
dinosaurs; and ROWE et al. 1992 and WEIL 1992 for mammals.
3.4. Abbreviations
The following abbreviations are used: LSUMG (Louisiana State University Museum of
Natural Science Geology Collections, Baton Rouge); RTMP (Royal Tyrrell Museum of
Paleontology, Drumheller, Alberta); UALVP (University of Alberta, Lab for Vertebrate
Paleontology, Edmonton).
4. Results
4.1. Microsites
7

Five microsites spanning 20 meters of stratigraphic section were collected from the
Talley Mountain area. These microsites are stream channel deposits of carbonate-cemented
sandstone conglomerates containing numerous clasts of clay, pedogenic nodules, small bones
and teeth, and large fragments of wood and bones. Large samples of vertebrates (3,349
identifiable specimens) were collected. 38 taxa were identified, including fish (68%), sharks and
rays (14%), crocodylomorphs (11%), dinosaurs (3%), amphibians (2%), turtles (not counted),
squamates (1%), and mammals (1%) (figs. 3 & 4). The stratigraphically lowest microsite is
adjacent to the AMNH quarry yielding the giant crocodylian Deinosuchus riograndensis
(COLBERT & BIRD 1954) and to the bonebeds of the horned dinosaur Chasmosaurus
mariscalensis and the hadrosaur Kritosaurus sp. (LEHMAN 1982). Deposits become more
terrestrial up section, from interdistributary marsh-bay facies at the base to inland floodplain
facies at the top, and aquatic vertebrates (fish, sharks, and rays) decrease in abundance upsection,
while terrestrial vertebrates (dinosaurs and mammals) increase in abundance.
4.2. Relative Abundance of Taxa
Fish
Two shark taxa indicate the influence of marine environments in the lowest microsites.
First, the mitsukurinid shark, Scapanorhynchus texanus (Fig. 3.4-3.5) is present, but rare (less
than 1%; Fig. 4; Table 1). Although modern mitsukurinids live in waters of 200 to 700 m depth,
they can enter shallower water at night (CAPPETTA 1987). The second shark, Hybodus sp. (Fig.
3.1), is also present, but rare in the lowest site (less than 1%; Fig. 4; Table 1). S. texanus has been
reported from the lower Aguja Formation (LEHMAN 1985), but this is the first record of Hybodus
sp. for the Aguja Formation.
8

Several taxa indicate brackish to freshwater environments. The first, Lissodus selachos
(Fig. 3.2-3.3) is the most abundant shark in the sites, but decreases in abundance upsection (82%
from the lower site compared to 1% from the highest site; Fig. 4; Table 1). L. selachos is
considered to be a brackish water shark, but both ESTES (1964) and CAPPETTA (1987) report that
it probably also could live in fresh water. It is also common in the estuarine Terlingua fauna
(ROWE et al. 1992) and is present in a freshwater deposit from the uppermost Aguja Formation
(STANDHARDT 1986). The other taxa of sharks and rays from the sites are considerably less
common than L. selachos. For example, Squatirhina americana (Fig. 3.9) is rare in the sites (less
than 2%), but is considered a shallow marine to brackish water shark (WELTON & FARISH 1993).
The sawfish Ischyrhiza avonicola (Fig. 3.8) is also rare (less than 1%). The paleoecology of
Ischyrhiza avonicola was probably similar to modern sawfish (ESTES 1964), which are bottom
dwellers that feed on fish and shellfish and live in near-shore areas in or around the mouths of
rivers in warm to tropical seas. Although one modern Central American species of sawfish lives
in fresh water (BRYANT 1989), most live in brackish and estuarine conditions (WELTON &
FARISH 1993). Onchopristis dunklei (Fig. 3.6-3.7) is also rare (less than 3%). The paleoecology
of Onchopristis dunklei was probably similar to modern sawfish. (These specimens extend the
range of this taxon into the late Campanian). Ptychotrygon sp. (Fig. 3.10-3.11) is also rare (less
than 1%). The paleoecology of Ptychotrygon sp. was also probably similar to modern sawfish.
Dasyatid stingrays (Fig. 3.13) are rare (less than 4%). Stingrays are bottom dwellers; they feed
on crustaceans, molluscs, and small fish and live in warm, shallow, coastal waters in the
subtropics and tropics. They can enter estuaries and rivers, and some species live in freshwater
(BUDKER 1971; WELTON & FARISH 1993). The gar, Lepisosteus occidentalis (Fig. 3.14-3.17), is
the most common fossil from the microsites and from the Aguja Formation in general (LEHMAN
9

1985; STANDHARDT 1986; ROWE et al. 1992). It makes up 65% of all fossils from the lowest site,
but only 29% from the highest. Its paleoecology was probably similar to the modern alligator
gar, L. spatula, which lives in brackish to fresh water in large streams along the Gulf Coast
(WILEY 1976; BRYANT 1989).
(((Figure 3 near here)))
(((Table 1 near here)))
Amphibians
There are additional taxa that indicate aquatic environments, and possibly more
freshwater environments. For example, two taxa of amphibians were identified. Both
Albanerpeton (Fig. 3.22), which was considered by ESTES (1964) to be primarily aquatic, and
Scapherpeton (Fig. 3.23) are rare (5% and less; Fig. 4; Table 1). Amphibians have also been
reported from the Terlingua fauna (ROWE et al. 1992) and from the uppermost Aguja Formation
(STANDHARDT 1986).
Turtles
Other aquatic indicators are the trionychids turtles, which live in aquatic environments
(TOMLINSON 1997). Fragments of indeterminate trionychid turtles were recovered from the
Talley Mountain microsites, but were not counted because numbers are dependent on degrees of
fragmentation. The trionychid "Aspideretes" is the most common turtle from the upper shale
member of the Aguja Formation (LEHMAN 1985; ROWE et al. 1992; TOMLINSON 1997; SANKEY
2006), but this level of identification was not possible with the Talley Mountain material.
Crocodylians
10

Crocodylians are more abundant aquatic taxa in the microsites, at 10 to 25% of the
assemblage (Fig. 4; Table 1). Goniopholids (Fig. 3.32-3.33) and Brachychampsa (Fig. 3.34-3.37)
are present in the microsites. Goniopholis cf. G. kirtlandicus was reported from the Terlingua
fauna (ROWE et al. 1992). There are several teeth that cannot be assigned to taxa. However, they
indicate a possibly more diverse crocodylian assemblage than is indicated by the taxonomic list
(Table 1). The giant crocodylian, Deinosuchus (=Phobosuchus) riograndensis is present in the
fauna. However, these teeth (Fig. 3.30-3.31) were surface collected from the area near the
microsites and not recovered by screening the microsites. In order to keep collection
methodology consistent, these specimens were not included in the specimen counts (Table 1). D.
riograndensis was the largest crocodylian from the Aguja Formation and is common from the
middle shale and base of the upper shale members (LEHMAN 1985). The type specimen of D.
riograndensis was described from the Talley Mountain field area (COLBERT & BIRD 1954).
Lizards
Although the taxa indicating terrestrial environments are in the minority, there is a
diverse range of taxa represented. The lizards, according to ROWE et al. (1992) and MILLER
(1997), represent unique taxa compared to northern assemblages. Within the Talley Mountain
microsites, lizard fossils are rare (2% or less; Fig. 4; Table 1). Chamops sp. (Fig. 3.26) is a new
record for Big Bend and Peneteius sp. nov. (Fig. 3.27-3.29) is a southern range extension
(NYDAM et al. in press).
Dinosaurs
11

Small dinosaur teeth and fragments of teeth provide important information on the
dinosaur community in Big Bend. From the Talley Mountain microsites, they represent 2 to 8%
of the assemblage (Fig. 4). Dinosaurs include hadrosaurid, ceratopsian, pachycephalosaurid,
tyrannosaurid, Saurornitholestes cf. S. langstoni, Richardoestesia isosceles, and two
undetermined theropod taxa (SANKEY 2001). Many of the dinosaur teeth are small and may
represent hatchlings or young individuals, evidence that many of these taxa nested in the area
(see WELSH & SANKEY this volume). Although pieces of eggshells have been collected from the
upper Aguja Formation near Rattlesnake Mountain, no complete eggs or nests have been found
in Big Bend (SANKEY et al. 2005b; WELSH 2004, 2005; WELSH & SANKEY this volume).
Mammals
Mammal fossils are another important, but small part of the fauna (1 to 5%; Fig. 3.59-
3.64; Fig. 4; Table 1). They are important because certain species can provide valuable dating
constraints. Mammals in the fauna include the multituberculates Cimolomys sp., Mesodma sp.,
cf. Cimexomys, cf. Paracimexomys and the marsupial Alphadon cf. A. halleyi and are assigned to
the Judithian Land Mammal Age (SANKEY 1998; SANKEY & GOSE 2001).
(((Figure 4 near here)))
5. Discussion
5.1. Significance of Big Bend Microsites
Because large samples of vertebrates were collected in the Talley Mountain area from
five microsites spanning 20 meters of section, paleoenvironmental changes could be tracked
during a short span of time. From this local section, deposits become more terrestrial upsection,
12

with interdistributary marsh-bay facies at the base and inland floodplain facies at the top. The
aquatic vertebrates, such as fish (gar), which represent fresh to brackish water environments and
shark and rays, which represent brackish to marine water environments, decrease in abundance
upsection, from 79% from the lowest site (within the interdistributary marsh-bay facies) to 61%
from the highest site (within the inland floodplain facies), while terrestrial vertebrates, such as
dinosaurs and mammals, increase in abundance upsection.
Although microfossil sites provide large sample sizes and diverse taxa, two biases in the
samples are observed. First, there is a bias against large fossils, such as bones from turtle,
dinosaur, and large crocodylians, due to size-sorting during transport within the streams. Second,
there is a bias against fragile fossils, such as from amphibians and lizards, due to destruction
during transport.
Sampling and analysis of microsites from the Aguja Formation have helped flesh out the
Late Cretaceous ecosystems in Big Bend, which prior to this work was primarily based on
isolated finds of large vertebrates such as the crocodylian Deinosuchus riograndensis and
dinosaurs Chasmosaurus mariscalensis and Kritosaurus sp. The vertebrate assemblage from the
Talley Mountain microsites is similar to vertebrates from the Terlingua microsite, also from the
lower part of the upper Aguja Formation (ROWE et al. 1992), which confirms the uniqueness of
the Aguja Formation fauna.
5.2. Southern Biogeographic Province
Late Cretaceous vertebrate faunas from southern parts of North America are still poorly
known compared to northern assemblages. However, their composition and change during the
Late Cretaceous is important to document. Big Bend is one of the southernmost terrestrial
13

records for the Late Cretaceous of North America. Therefore, finding and recovering numerous
Late Cretaceous vertebrates from microsites in Big Bend has contributed important information
about this unique fauna.
The fossils reported here from the Tally Mountain microsites significantly increase our
knowledge for this southern area. Vertebrates from Big Bend are taxonomically distinct at the
species or genus level from contemporaneous northern faunas, confirming the uniqueness of this
southern fauna. Additionally, preliminary results indicate that dinosaurs in Big Bend were less
diverse than contemporaneous northern areas. The late Campanian Big Bend dinosaur
assemblage was more similar to the late Maastrichtian southern Alberta (Scollard Formation)
dinosaur assemblage in the following ways: Richardoestesia isosceles was more abundant than
R. gilmorei; Troodon and Dromaeosaurus were absent or rare; and pachycephalosaurids were
present to common (SANKEY 2001; SANKEY et al. 2005b).
Differences between the southern and northern areas and their vertebrate faunas were
primarily due to differences in climate, with periodic aridity occurring earlier in Big Bend than in
northern areas. The Aguja Formation dinosaur bonebeds probably resulted from periodic
droughts, severe enough to cause the marshes to dry up (DAVIES & LEHMAN 1989) and
paleocaliche to form in soils. Post-drought flooding, possibly even flash floods, would have
eroded and transported soil and vertebrate remains, quickly depositing them within streams,
forming conglomeratic beds with an appreciable fossil content, which are microsites (SANKEY
1998; SANKEY & GOSE 2001). This paleoenvironmental picture differs from contemporaneous
northern areas such as Alberta. During the late Campanian in Alberta, the Dinosaur Park
Formation was deposited within an extensive mesic coastal lowland (EBERTH & HAMBLIN 1993),
and Alberta did not experience aridity until the late Maastrichtian.
14

5.3. Future Focus: Late Cretaceous Climate Change
Understanding the detailed effects of climate change on the Late Cretaceous terrestrial
ecosystems is important in order to separate the terrestrial from extraterrestrial factors involved
in the Cretaceous/Tertiary extinctions (SANKEY 2006). Beginning in the late Campanian (~74
Ma), a cooling trend and corresponding drop in sea level and retreat of the Western Interior
Seaway caused changes in paleoenvironments and paleocommunities in the Western Interior of
North America. Then in the Maastrichtian, two short greenhouse events occurred, corresponding
to minor extinctions before the final K/T mass extinctions (NORDT et al. 2003). What were the
effects of these climatic changes on the vertebrate paleocommunities? Can we use climatically
sensitive vertebrate groups, for example turtles and crocodilians, to track these changes? Future
work in Big Bend will track paleocommunity changes corresponding with climatic and
environmental changes through the section by making further collections from microsites and by
analyzing associated paleosol carbonates. By studying the Big Bend area, we may identify a
relationship between vertebrates and climate such that we can use the vertebrates to interpret
climate in other areas.
6. Summary and Conclusions
Discovery and collection of microsites from the upper Aguja Formation (late Campanian)
in Big Bend National Park, Texas have produced numerous vertebrate fossils, significantly
improving our knowledge of the vertebrates from this extreme southern part of North America.
The vertebrates differ from contemporaneous northern faunas at the species or genus level,
confirming the uniqueness of the southern vertebrate fauna. Differences between the southern
15

and northern areas and vertebrate faunas are primarily due to differences in climate, with warmer
and drier climates occurring in the south starting in the late Campanian, but not until the late
Maastrichtian in the north. The southern vertebrate faunas were lower in diversity compared to
contemporaneous northern assemblages due to these climatic differences. Similar results would
be expected from other faunas further to the south, such as those from Mexico.
7. Acknowledgments
This work was part of Sankey’s Ph.D. dissertation (1998) at Louisiana State University
(LSU) in Baton Rouge, and she thanks J. Schiebout (major advisor), A. Bouma (minor advisor),
J. Hazel, J. Lorenzo, J. Wrenn, and G. Wise (committee members) for advice. Financial support
to Sankey is appreciated from the LSU Museum of Natural Science and Department of Geology
and Geophysics; the Dinosaur Society; the Jurassic Foundation; the U.S.-Canada Fulbright
Program; and the Haslem and Nelson Funds (South Dakota School of Mines and Technology,
Rapid City). The author appreciates discussions with D. Brinkman, S. Baszio, W. Clemens, R.
Cifelli, P. Currie, J. Eaton, D. Eberth, R. Fox, J. Gardner, W. Langston, T. Lehman, R. Nydam,
T. Rowe, J. Schiebout, B. Standhardt, and A. Weil. Access to the following fossil collections is
appreciated: Laboratory for Vertebrate Palaeontology, University of Alberta, Edmonton; Royal
Tyrrell Museum of Palaeontology, Drumheller, Alberta; the Vertebrate Paleontology Laboratory,
Texas Memorial Museum, University of Texas at Austin; Oklahoma Museum of Natural History,
University of Oklahoma, Norman; Department of Geosciences, Weber State University, Ogden,
Utah; and Museum of Paleontology, University of California, Berkeley. Fossils were collected in
Big Bend National Park under permit #BIBE-N-50.001. Jean Sankey helped with the field
work. Assistance is appreciated from Big Bend personnel P. Koepp, V. Davilla, and M. Fleming.
16

Lab work by volunteers and assistants from LSUMG (T. Aherns, C. Moore, J. Ramcharan, M.
Schneider, and S. Wachter) is appreciated. M. Eggart and L. Pond (LSU) drew the figures.
8. References
ATCHLEY, S.C.; NORDT, L.C.; DWORKIN, S.I. 2004. Eustatic control on alluvial sequence
stratigraphy: a possible example from the Cretaceous-Tertiary transition of the Tornillo
Basin, Big Bend National Park, West Texas, U.S.A. - Journal of Sedimentary Research
74(3): 391-404.
BAGHAI, N. 1996. An analysis of palynomorphs from Upper Cretaceous sedimentary rocks with
emphasis on the Aguja Formation, Big Bend National Park, Brewster County, Texas. –
Ph.D. Dissertation, University of Texas, Austin, 495 pp.
BARRERA, E. 1994. Global environmental changes preceding the Cretaceous-Tertiary boundary:
early-late Maastrichtian transition. - Geology 22: 877-880.
BASZIO, S. 1997. Investigations on Canadian dinosaurs: systematic paleontology of isolated
dinosaur teeth from the Latest Cretaceous of South Alberta. - Courier Forschungsinstitut
Senckenberg 196: 33-77.
BRINKMAN, D. 1990. Paleoecology of the Judith River Formation (Campanian) of Dinosaur
Provincial Park, Alberta, Canada: evidence from vertebrate microfossil localities. –
Palaeogeography, Palaeoclimatology, Palaeoecology 78: 37-54.
BRYANT, L.J. 1989. Non-dinosaurian lower vertebrates across the Cretaceous-Tertiary boundary
in northeastern Montana. - University of California Publications in Geological Sciences
134: 1-107.
BUDKER, P. 1971. The Life of Sharks. - Columbia University Press, New York, 222 p.
CAPPETTA, H. 1987. Chondrichthyes II: Mesozoic and Cenozoic Elasmobranchii. – Handbook of
17

Paleoichthyology, 3B: Gustav Fischer, New York, 193 p.
COLBERT, E.H. & BIRD, R.T. 1954. A gigantic crocodile from the Upper Cretaceous beds
of Texas. - American Museum Novitates 1688: 1-22.
CURRIE, P.J.; RIGBY, JR., J.K.; & SLOAN, R.E. 1990. Theropod teeth from the Judith River
Formation of southern Alberta, Canada. - In: CARPENTER, K. & CURRIE, P.J., eds.,
Dinosaur Systematics: Approaches and Perspectives: 107-125. Cambridge University
Press, Cambridge.
DAVIES, K. & LEHMAN, T.M.. 1989. The WPA Quarries. - In: BUSBEY III, A.B. & LEHMAN,
T.M., eds, Vertebrate Paleontology, Biostratigraphy, and Depositional Environments,
Latest Cretaceous and Tertiary, Big Bend Area, Texas: 32-42. Guidebook, Field Trips
Nos. 1a, b, c. Society of Vertebrate Paleontology 49th Annual Meeting, Austin, Texas.
EBERTH, D. & HAMBLIN, A.P. 1993. Tectonic, stratigraphic, and sedimentologic significance
of a regional discontinuity in the upper Judith River Group (Belly River wedge) of
southern Alberta, Saskatchewan, and northern Montana. - Canadian Journal of Earth
Sciences 30: 174-200.
ESTES, R. 1964. Fossil vertebrates from the late Cretaceous Lance Formation, eastern
Wyoming. - University of California Publications in Geological Sciences 49: 1-180.
ESTES, R. 1981. Gymnophiona, Caudata. - Handbuch der Palaeoherpetologie 2. Fischer Verlag,
Stuttgart, 115 p.
GAO, K. & FOX, R.C. 1996. Taxonomy and evolution of Late Cretaceous lizards (Reptilia:
Squamata) from Western Canada. - Bulletin of Carnegie Museum of Natural History 33:
1-107.
GRADSTEIN, F.M.; AGTERBERG, F.P.; OGG, J.G.; HARDENBOL, J; VENN, P.V.; THIERRY, J.;
18

HUANG, Z. 1995. A Triassic, Jurassic and Cretaceous time scale, - In: BERGGREN, W.A.;
KENT, D.V.; AUBRY, M.P.; & HARDENBOL, J., eds., Geochronology, Time Scale and
Global Stratigraphic Correlation: 95-126. Society for Sedimentary Geology Special
Publication.
KAUFFMAN, E.G. 1977. Geological and Biological Overview: Western Interior Cretaceous
Basin. - The Mountain Geologist 14: 75-99.
LEHMAN, T.M. 1982. A ceratopsian bone bed from the Aguja Formation (upper Cretaceous),
Big Bend National Park, Texas. - M.A. thesis, University of Texas, Austin, 209 p.
LEHMAN, T.M. 1985. Stratigraphy, sedimentology, and paleontology of Upper Cretaceous
(Campanian-Maastrichtian) sedimentary rocks in Trans-Pecos, Texas. - Ph.D.
dissertation, University of Texas, Austin, 299 pp.
LEHMAN, T.M. 1989. Chasmosaurus mariscalensis, sp. nov., a new ceratopsian dinosaur from
Texas. - Journal of Vertebrate Paleontology 9: 137-162.
LEHMAN, T.M. 1990. Paleosols and the Cretaceous/Tertiary transition in the Big Bend region of
Texas. - Geology 18: 362-364.
LEHMAN, T.M. 1991. Sedimentation and tectonism in the Laramide Tornillo Basin of West
Texas. - Sedimentary Geology 75: 9-28.
LEHMAN, T.M. 1997. Late Campanian dinosaur biogeography in the Western Interior of North
America. - In: WOLBERG, D.A. & STUMP, E., eds, Dinofest International, Proceedings of
a Symposium Sponsored by Arizona State University: 223-240. Special Publication,
Academy of Natural Sciences, Philadelphia.
LEHMAN, T.M. & WHEELER, E.A. 2001. A fossil dicotyledonous woodland forest from the
upper Cretaceous of Big Bend National Park, Texas. - Palaios 16: 102-108.
19

MILLER, M.S. 1997. Squamates of the Campanian Terlingua local fauna, Brewster County,
Texas. - M.A. thesis, University of Texas, Austin. 98 pp.
NICHOLS, D.J. & SWEET, A.R. 1993. Biostratigraphy of Upper Cretaceous non-marine
palynofloras in a north-south transect of the Western Interior Basin. - Special Paper,
Geological Association of Canada 39: 539-584.
NORDT, L; ATCHLEY, S; & DWORKIN, S. 2003. Terrestrial evidence for two greenhouse events
in the Latest Cretaceous. - Geological Society of America Today: 4-9.
NYDAM, R.L.; EATON, J.G.; & SANKEY, J.T. In Press. New taxa and revisions of
polyglyphanodontine lizards (Sqaumata: Scincomorpha) from the upper Cretaceous of
North America. - Journal of Paleontology (28 pp, 6 figs, 1 app).
PENG, J.; RUSSELL, A.P.; & BRINKMAN, D.B. 2001. Vertebrate microsite assemblages (exclusive
of mammals) from the Foremost and Oldman Formations of the Judith River Group
(Campanian) of southeastern Alberta: an illustrated guide. - The Provincial Museum of
Alberta, Natural History Occasional Paper 24: 1-54.
ROBINSON-ROBERTS, L & KIRSCHBAUM, M. 1995. Paleogeography of the Late Cretaceous of
the western interior of middle North America – coal distribution and sediment
accumulation. - U.S. Geological Survey Professional Paper 1561: 115 p.
ROWE, T.; CIFELLI, R.L; LEHMAN, T.M.; & WEIL, A. 1992. The Campanian Terlingua Local
fauna, with a summary of other vertebrates from the Aguja Formation, Trans-Pecos,
Texas. - Journal of Vertebrate Paleontology 12: 472-493.
SANKEY, J.T. 1998. Vertebrate paleontology and magnetostratigraphy of the upper Aguja
Formation (late Campanian), Talley Mountain area, Big Bend National Park, Texas.
Ph.D. dissertation. - Louisiana State University, Baton Rouge, LA. 263 pp.
20

SANKEY, J.T. 2001. Late Campanian southern dinosaurs, Aguja Formation, Big Bend, Texas.
- Journal of Paleontology 75(1): 208-215.
SANKEY, J.T. 2005. Late Cretaceous dinosaurs, eggs, babies, droughts, and fires in Big Bend
National Park, Texas. - The Geological Society of America, Cordilleran Section, San
Jose, California, April 29- May 1, 2005.
SANKEY, J.T. 2006. Turtles of the upper Aguja Formation (late Campanian), Big Bend
National Park, Texas. - In: LUCAS, S. & SULLIVAN, R., eds, Late Cretaceous Vertebrates
from the Western Interior: pp. 235-243. New Mexico Museum of Natural History and
Science Bulletin 35.
SANKEY, J.T.; BRINKMAN, D.B.; FOX, R.C.; & EBERTH, D.A. 2005a. Patterns of distribution of
mammals in the Dinosaur Park Formation and their paleobiological significance. – In:
CURRIE, P.J. & KOPPELHUS, E., eds., Dinosaur Provincial Park, Alberta: 436-449. Indiana
University Press.
SANKEY, J.T.; BRINKMAN, D.B.; GUENTHER, M; & CURRIE, P.J. 2002. Small theropod and bird
teeth from the Judith River Group (late Campanian), Alberta.- Journal of Paleontology
76(4): 751-763.
SANKEY, J.T. & GOSE, W.A. 2001. Late Cretaceous mammals and magnetostratigraphy, Big
Bend, Texas. - Occasional Papers of the Museum of Natural Science, Louisiana State
University No. 77: 1-16.
SANKEY, J.T.; STANDHARDT, B.R.; & SCHIEBOUT, J.A. 2005. Theropod teeth from the upper
Cretaceous (Campanian-Maastrichtian), Big Bend National Park, Texas. – In:
CARPENTER, KEN, ed., Carnivorous Dinosaurs: 127-152. Indiana University Press
(Bloomington).
21

SCHIEBOUT, J.A.; TING, S.; & SANKEY, J.T. 1997. Microvertebrate Concentrations in Pedogenic
Nodule Conglomerates: Recognizing the Rocks and Recovering and Interpreting the
Fossils. - Palaeontologia Electronica 1(2): 1-54.
http://wwwodp.tamu.edu/paleo/1998_2/schiebt/issue2.htm
STANDHARDT, B.R. 1986. Vertebrate paleontology of the Cretaceous/Tertiary transition of Big
Bend National Park, Texas. - Ph.D. dissertation, Louisiana State University, Baton
Rouge, 298 pp.
SULLIVAN, R.M. & LUCAS, S.G. 2003. The Kirtlandian, a new land-vertebrate “age” for the
Late Cretaceous of Western North America. - New Mexico Geological Society
Guidebook, 54th Field Conference, Geology of the Zuni Plateau, p. 369-377.
TOMLINSON, S.L. 1997. Late Cretaceous and Early Tertiary turtles from the Big Bend
region, Brewster county, Texas. - Ph.D. dissertation, Texas Tech University, Lubbock,
195 p.
WEIL, A. 1992. The Terlingua local fauna: stratigraphy, paleontology, and multituberculate
systematics. - M.A. thesis, University of Texas, Austin, 120 pp.
WELSH, E. 2004. Oodiversity in the Aguja Formation at Big Bend National Park, Texas.
- Program and Proceedings, The Nebraska Academy of Sciences: 49.
WELSH, E. 2005. Eggshells and baby dinosaurs in the upper Aguja Formation of Big Bend
National Park, Texas: - Journal of Vertebrate Paleontology Abstracts of Papers 25(3):
128A.
WELSH, E. & SANKEY, J.T. This volume. First dinosaur eggshells from Texas: Aguja Formation
(late Campanian), Big Bend National Park. - In: SANKEY, J.T. & BASZIO, S. eds, The
Unique Role of Vertebrate Microfossil Assemblages in Paleoecology and
22

Paleobiogeography: XXX-XXX. Indiana University Press (Bloomington).
WELTON, J.G. & FARISH, R.F. 1993. - The Collector's Guide to Fossil Sharks and Rays from
the Cretaceous of Texas. Before Time, Lewisville, Texas, 204 p.
WHEELER, E.A. & LEHMAN, T.M. 2000. Late Cretaceous woody dicots from the Aguja and
Javelina Formations, Big Bend National Park, Texas, USA. - IAWA Journal 21(1): 83-
120.
WILEY, E.O. 1976. The phylogeny and biogeography of fossil and recent gars (Actinopterygii:
Lepisosteidae). - University of Kansas Museum of Natural History Publication 64: 1-111.
ZIEGLER, A.M. & ROWLEY, D.B. 1998. The vanishing record of epeiric seas, with emphasis on
the Late Cretaceous “Hudson Seaway”, - In: CROWLEY, T.J., & BURKE, K., eds., Tectonic
Boundary Conditions for Climatic Reconstructions. Oxford University Press, Oxford.
23

FIGURE 1-- Late Cretaceous paleogeography. 1, Maastrichtian paleogeographic reconstruction of North America (redrawn from ZIEGLER & ROWLEY 1998). Star is location of Big Bend National Park, Texas. Arrows show close land connections during retreat of the Western Interior Seaway in the Maastrichtian. 2, Big Bend National Park, west Texas with Aguja Formation outcrops stippled. Locations of Talley Mountain and Terlingua field areas are shown by arrows. Both field areas have microsites that have yielded numerous vertebrates from the upper Aguja Formation (late Campanian).
24

FIGURE 2--Stratigraphy of the Aguja Formation, Big Bend National Park, Texas. 1, Lithostratigraphy of the Aguja Formation modified from ROWE et al. (1992), showing positions of the Talley Mountain (SANKEY 1998) and Terlingua (ROWE et al. 1992) sites. Stratigraphic position of the Talley Mountain microsites and WPA dinosaur quarries 1 and 2 (Work Progress Administration) from LEHMAN (1985; Plate III and written comm., 1998). Formal members of the Aguja Formation are capitalized; informal members are not. 2, Magnetostratigraphic correlations of upper Aguja Formation from Talley Mountain area (from SANKEY & GOSE 2001).
25

26

FIGURE 3 -- Vertebrate taxa from the Talley Mountain sites. 1, Hybodus sp. (LSUMG 489:5627). 2-3, Lissodus selachos; 2, LSUMG 491:5965; 3, LSUMG 489:5692. 4-5, Scapanorhynchus texanus; 4, LSUMG 489:5626; 5, LSUMG 489:5666. 6-7, Onchopristis dunklei (LSUMG 489:5667). 8, Ischyrhiza avonicola (LSUMG 491:5977). 9, Squatirhina americana (LSUMG 489:5689). 10-11, Ptychotrygon sp. (LSUMG 489:5670). 12, Chondrichthyan dermal denticle (LSUMG 140:6163). 13, Dasyatidae (LSUMG 489:5648). 14, Lepisosteus complete tooth (LSUMG 488:5521). 15-16, Lepisosteus tooth caps; 15, LSUMG 140:6127; 16, LSUMG 140:6128. 17, Lepisosteidae scale (LSUMG 489:5732). 18-19, Indeterminate fish vertebrae (LSUMG 140:6144). 20-21, Phyllodontid tooth plates (LSUMG 489:5577). 22, Albanerpeton sp. maxilla, lingual view (LSUMG 491:5958). 23, Scapherpeton sp. dentary, lingual view (LSUMG 489:5694). 24, Glyptosaurinae indet., maxilla, lingual view (LSUMG 140:6106). 25, Teiidae-"Species B" (Miller, 1997), tooth (LSUMG 489:5656). 26, Chamops sp. mandible, labial view (LSUMG 140:6104). 27-29, Peneteis sp. nov., tooth, lateral and occlusal views (LSUMG 492:6253). 30-31, Deinosuchus riograndensis teeth; 30, LSUMG 489:5672; 31, LSUMG, 488:5484. 32-33, Goniopholid indet. Teeth; 32, LSUMG 489:5673; 33, LSUMG 489:5608. 34-37, cf. Brachychampsa sp. Teeth; 34, LSUMG 491:5621; 35, LSUMG 140:6125; 36-37, LSUMG 489:5658. 38-40, Crocodylian indeterminate type B teeth; 38, LSUMG 140:5564; 39-40, 489:5697. 41-42, Crocodylian indet. type A teeth; 41-42, LSUMG 488:5496. 43-44, Crocodylian indet. Scutes; 43, LSUMG 489:5582; 44, LSUMG 140:6136. 45, Hadrosaurid tooth, hatchling or juvenile (LSUMG 489:5538, occlusal view). 46, Ceratopsian tooth fragment (LSUMG 488:6230, occlusal view). 47, Tyrannosaurid tooth fragment, close-up of denticles (LSUMG 489:5580). 48-49, Pachycephalosauridae, lateral view and close-up (LSUMG 140:6117). 50-51, Undetermined family and genus, lateral view and close-up of denticles (LSUMG 488:5483); 52, Saurornitholestes cf. S. langstoni, lateral view (LSUMG 140:6139); 53-54, Saurornitholestes cf. S. langstoni unshed tooth, lateral view and close-up of denticles (LSUMG 489:5659); 55-56, Richardoestesia isosceles n. sp., lateral view and close-up of denticles (LSUMG 489:6238). 57-58, Undetermined family and genus, lateral view and close-up of denticles (LSUMG 489:6239). 59, Cimolomys sp. LM1 (LSUMG 489:5681). 60, Mesodma sp. anterior fragment, Lm1 (LSUMG 491:5779). 61, cf. Cimexomys rm1 (LSUMG 140:6121). 62, cf. Paracimexomys RM2 (LSUMG 491:5778). 63, cf. Paracimexomys anterior fragment, PM3 (LSUMG 489:5684). 64, Alphadon cf. A. halleyi LM3 (LSUMG 492:6252).
27
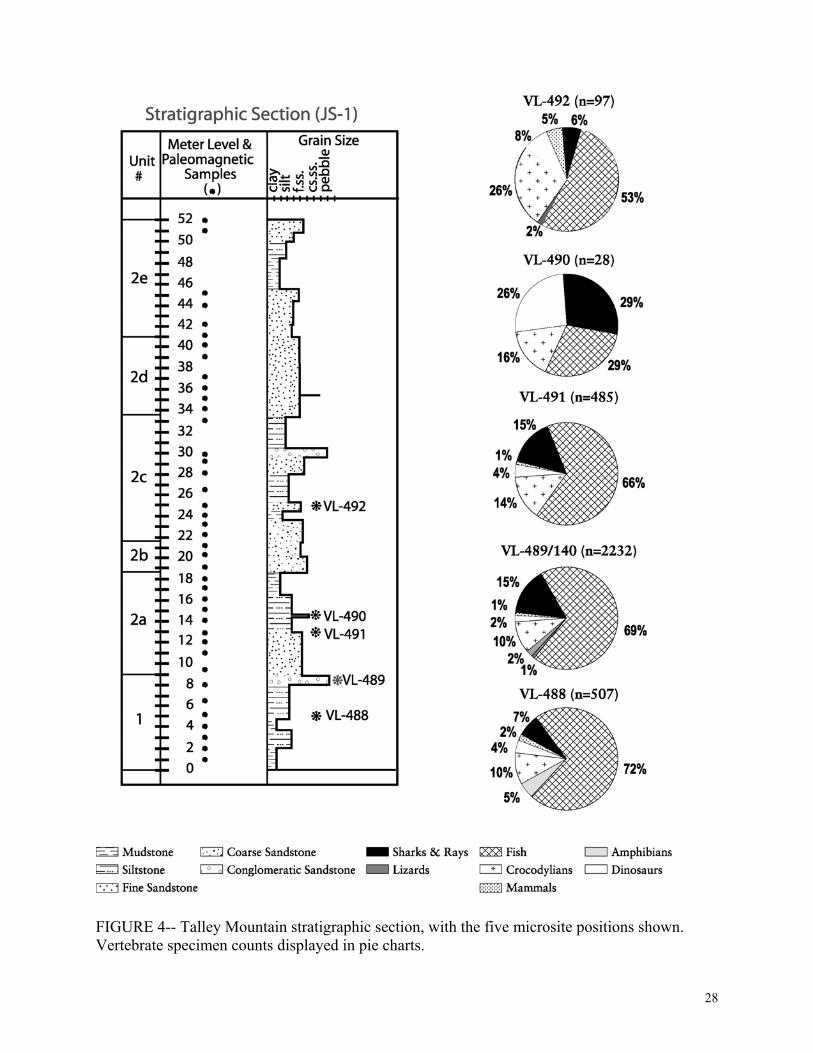
FIGURE 4-- Talley Mountain stratigraphic section, with the five microsite positions shown. Vertebrate specimen counts displayed in pie charts.
28
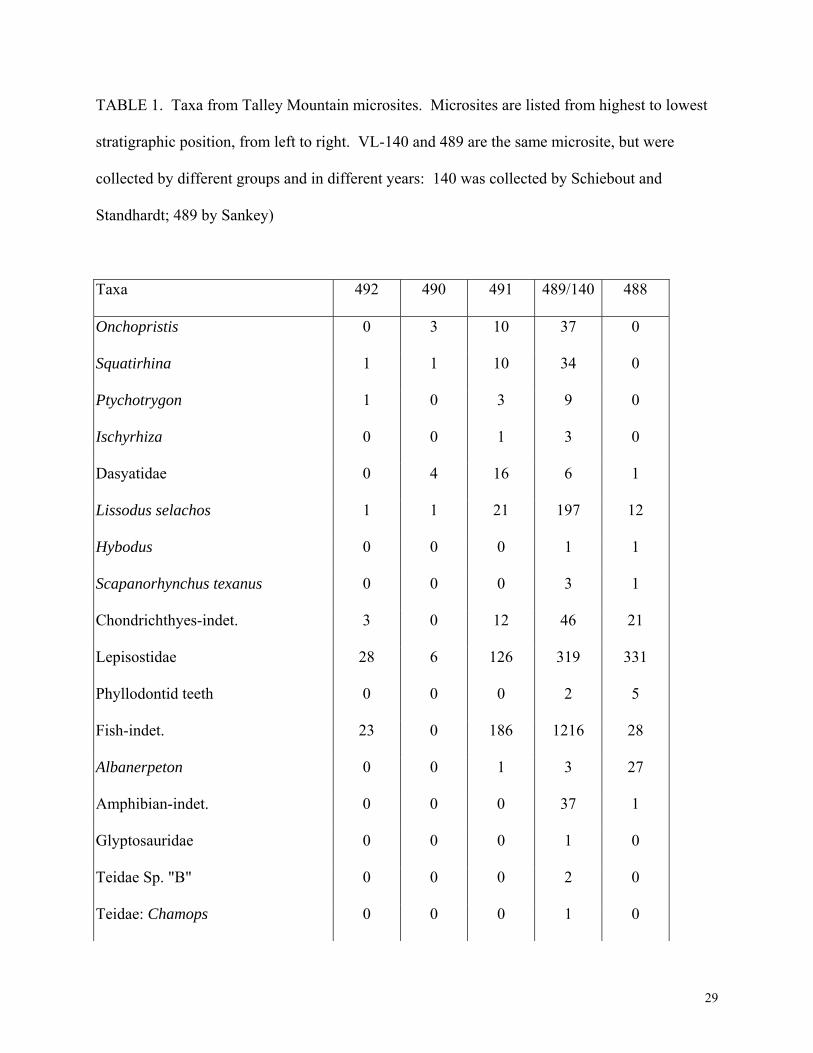
TABLE 1. Taxa from Talley Mountain microsites. Microsites are listed from highest to lowest
stratigraphic position, from left to right. VL-140 and 489 are the same microsite, but were
collected by different groups and in different years: 140 was collected by Schiebout and
Standhardt; 489 by Sankey)
Taxa 492 490 491 489/140 488
Onchopristis 0 3 10 37 0
Squatirhina 1 1 10 34 0
Ptychotrygon 1 0 3 9 0
Ischyrhiza 0 0 1 3 0
Dasyatidae 0 4 16 6 1
Lissodus selachos 1 1 21 197 12
Hybodus 0 0 0 1 1
Scapanorhynchus texanus 0 0 0 3 1
Chondrichthyes-indet. 3 0 12 46 21
Lepisostidae 28 6 126 319 331
Phyllodontid teeth 0 0 0 2 5
Fish-indet. 23 0 186 1216 28
Albanerpeton 0 0 1 3 27
Amphibian-indet. 0 0 0 37 1
Glyptosauridae 0 0 0 1 0
Teidae Sp. "B" 0 0 0 2 0
Teidae: Chamops 0 0 0 1 0
29

Peneteius sp. nov. 1 0 0 0 0
Scincomorph 0 0 0 1 0
Lizard-indet. 1 0 0 23 1
Croc indet. B 0 3 29 105 13
Croc indet. A 0 0 8 11 4
Brachychampsa 7 0 28 50 23
Gonopholidae 0 0 0 4 1
Crocodylian-indet. 18 2 10 46 10
Hadrosauridae 1 0 1 6 0
Ornithischian-indet. 2 0 0 4 1
Tyrannosauridae 0 0 4 2 0
R. isosceles 1 0 0 6 2
R. gilmorei 0 0 0 1 0
Pachycephalosauridae 0 0 0 1 0
cf. Richardoestesia 0 0 0 2 0
Saurornitholestes-indet. 0 0 0 2 1
S. langstoni 1 0 9 5 0
Theropod indet. 0 8 5 10 16
Dino. frags. 3 0 0 6 0
cf. Aves 0 0 0 4 0
Alphadon cf. A. halleyi 1 0 0 1 0
Marsupialia-indet. 1 0 2 8 1
Cimolomys sp. nov. 0 0 0 1 0
30
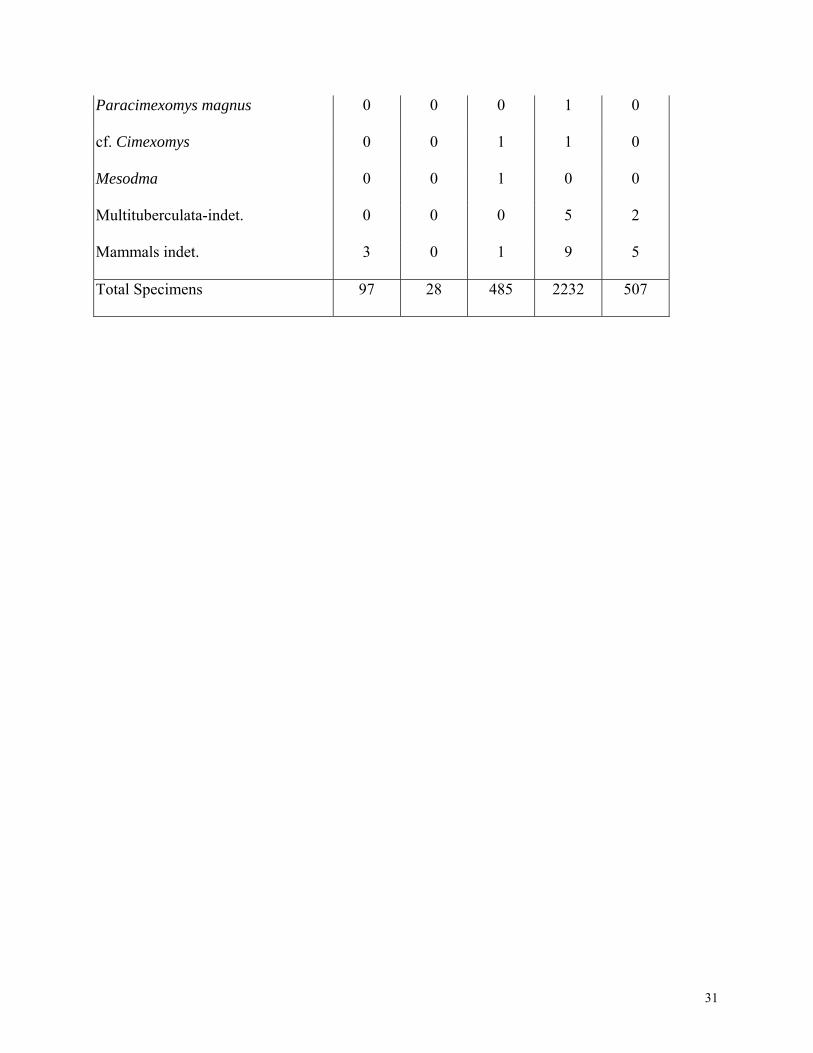
Paracimexomys magnus 0 0 0 1 0
cf. Cimexomys 0 0 1 1 0
Mesodma 0 0 1 0 0
Multituberculata-indet. 0 0 0 5 2
Mammals indet. 3 0 1 9 5
Total Specimens 97 28 485 2232 507
31





























