Embed Size (px)
Citation preview

17. Lecture WS 2006/07
Bioinformatics III 1
V17 Modelling signalling cascades
Cells respond to external stimuli using a limited number of signalling pathways that
are activated by plasmamembrane receptors, such as G protein-coupled receptors
(GPCRs) and receptor tyrosine kinases (RTKs).
These pathways do not simply transmit, but they also process, encode and integrate
internal and external signals.
Distinct spatio-temporal activation profiles of the same repertoire of signalling
proteins may´result in different gene-expression patterns and diverse physiological
responses pivotal cellular decisions, such as cytoskeletal reorganization, cell-
cycle checkpoints and cell death (apoptosis), depend on the precise temporal
control and relative spatial distribution of activated signal transducers.
B.N. Kholodenko, Nature Rev. Mol. Cell. Biol. 7, 165 (2006)

17. Lecture WS 2006/07
Bioinformatics III 2
receptor tyrosine kinasesRTK-mediated signalling pathways have a central role in the regulation of embryogenesis,
cell survival, motility, proliferation, differentiation, glucose metabolism and apoptosis.
Malfunction of RTK signalling is a leading cause of important human diseases involving e.g.
developmental defects, cancer, chronic inflammatory syndromes and diabetes.
Upon stimulation, RTKs undergo dimerization (e.g. the epidermal growth factor receptor.
EGFR) or allosteric transitions (insulin receptor) that result in the activation of the intrinsic
tyrosine-kinase activity.
Subsequent phosphorylation of multiple tyrosine residues on the receptor transmits a
biochemical signal to numerous cytoplasmic proteins, thereby triggering their mobilization to
the cell surface.
The resulting cellular responses occur through complex biochemical circuits of protein–
protein interactions and covalent-modification cascades.

17. Lecture WS 2006/07
Bioinformatics III 3
Cycle and cascade motifs
A universal motif that is
found in cellular networks is
the cycle that is formed by
two or more interconvertible
forms of a signalling protein.
This protein is modified by
two opposing enzymes, such
as a kinase and a
phosphatase for phospho
proteins, or a guanine
nucleotide exchange factor
(GEF) and a GTPase-
activating protein (GAP) for
small G proteins.

17. Lecture WS 2006/07
Bioinformatics III 4
Feedback loops induce complex dynamicsFeedback is one of the most fundamental concepts in biological control. An increase in the number of interconnecting cycles in a cascade or positive feedback further increases the sensitivity of the target to the input signal.
Positive feedback amplifies the signal, whereas negative feedback attenuates it. However, feedback loops not only change steady-state responses, but also favour the occurrence of instabilities. When a steady state becomes unstable, a system can jump to another stable state, start to oscillate or exhibit chaotic behaviour.
Positive feedback can cause bistability. Furthermore, positive feedback, alone or in combination with negative feedback, can trigger oscillations; for example, the Ca2+ oscillations that arise from Ca2+-induced Ca2+ release and the cell-cycle oscillations.Such positive-feedback oscillations generally donot have sinusoidal shapes and are referred to as relaxation oscillations, operating in a pulsatory manner: a part of a dynamic system is bistable, and there is a slow process that periodically forces the system to jump between ‘off ’ and ‘on’ states, generating oscillations.

17. Lecture WS 2006/07
Bioinformatics III 5
complex dynamics
Complex dynamic properties have traditionally been associated with cascades of cycles.
Yet, even single cycles can exhibit complex dynamics, such as bistability and relaxation
oscillations.
For instance, multisite protein modification not only increases ultrasensitivity, but it could
potentially lead to bistability.
Based on the reported kinetic data, it has been proposed that a single MAPK cascade level
— for example, the dual phosphorylation ERK cycle — can exhibit bistability and hysteresis
within a certain parameter range. This prediction that awaits experimental verification.
A simple one-site modification cycle can turn into a bistable switch by four different
regulatory mechanisms, in which one of the protein forms stimulates its own production or
inhibits its consumption, thereby creating a destabilizing control loop.
An extra (stabilizing) feedback loop that affects the rate of synthesis or degradation of a
converting enzyme can render this bistable switch into a relaxation oscillator (32 distinct
feedback designs result that can give rise to oscillations).

17. Lecture WS 2006/07
Bioinformatics III 6
receptor tyrosine kinasesFeedback designs that can turn a universal signalling cycle into a bistable switch and relaxation oscillator. A simple cycle can turn bistable in 4 distinct ways: either a protein M or its phosphorylated form Mp stimulates its own production (positive feedback) by product activation or substrate inhibition of the kinase (Kin) or phosphatase (Phos) reactions. Each of the 4 rows of feedback designscorresponds to a different bistable switch, provided that the kinase and the phosphatase abundances are assumed constant and only a single feedback (within the M cycle) is present. Sixteen relaxation-oscillation designs are generated by extra negative feedback brought about by negative or positive regulation of the synthesis or degradation rates of the kinase protein or phosphatase protein by M or Mp. Designs a*–h* are mirror images of designs a–h. Although synthesis and degradation reactions are shown for both the kinase and the phosphatase proteins, the protein concentration that is not controlled by feedback from the M cycle is considered constant, therefore it results in only two differential equations for each diagram. All the feedback regulations are described by simple Michaelis–Menten-type equations. The remaining 16 relaxation-oscillation designs are shown on the next slide and can require some degree of cooperativity within feedback loops.
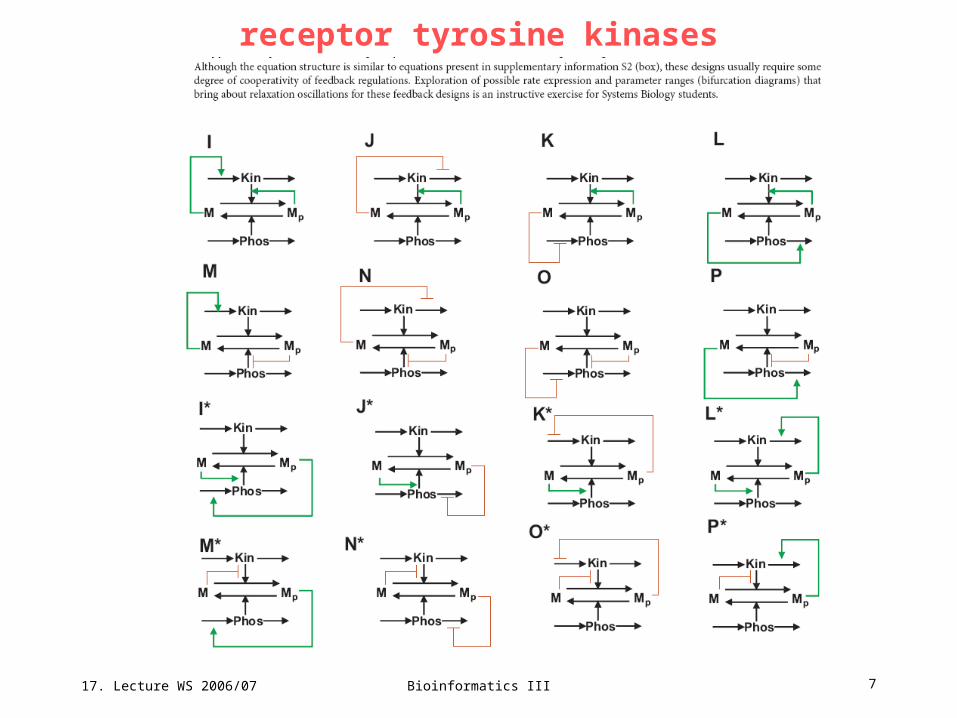
17. Lecture WS 2006/07
Bioinformatics III 7
receptor tyrosine kinases

17. Lecture WS 2006/07
Bioinformatics III 8
cell-cycle machineryThe signalling dynamics can become multi-stable when two or more bistable cycles form a
cascade, such as a MAPK cascade. The biological outcome of multistability is the ability to
control multiple irreversible transitions, for instance, sequential transitions in the cell cycle.
Central components of the cell-cycle machinery are cyclin-dependent kinases (such as CDK1/ CDC2), the sequential activation and inactivation of which govern cell-cycle transitions. The activity of CDK1/CDC2 is low (off) in the G1 phase and has to be high (on) for entry into mitosis (M phase). Tyson et al., Curr.Opin.Cell.Biol. 15, 221 (2003)

17. Lecture WS 2006/07
Bioinformatics III 9
Cell cycle control system
Tyson et al., Curr.Pin.Cell.Biol. 15, 221 (2003)
The G1/S module is a toggle switch, based on mutual inhibition between
Cdk1-cyclin B and CKI, a stoichiometric cyclin-dependent kinase inhibitor.

17. Lecture WS 2006/07
Bioinformatics III 10
Cell cycle control system
Tyson et al., Curr.Pin.Cell.Biol. 15, 221 (2003)
The G2/M module is a second toggle switch, based on mutual activation between
Cdk1-cyclinB and Cdc25 (a phosphotase that activates the dimer) and mutual
inhibition between Cdk1-cyclin B and Wee1 (a kinase that inactivates the dimer).
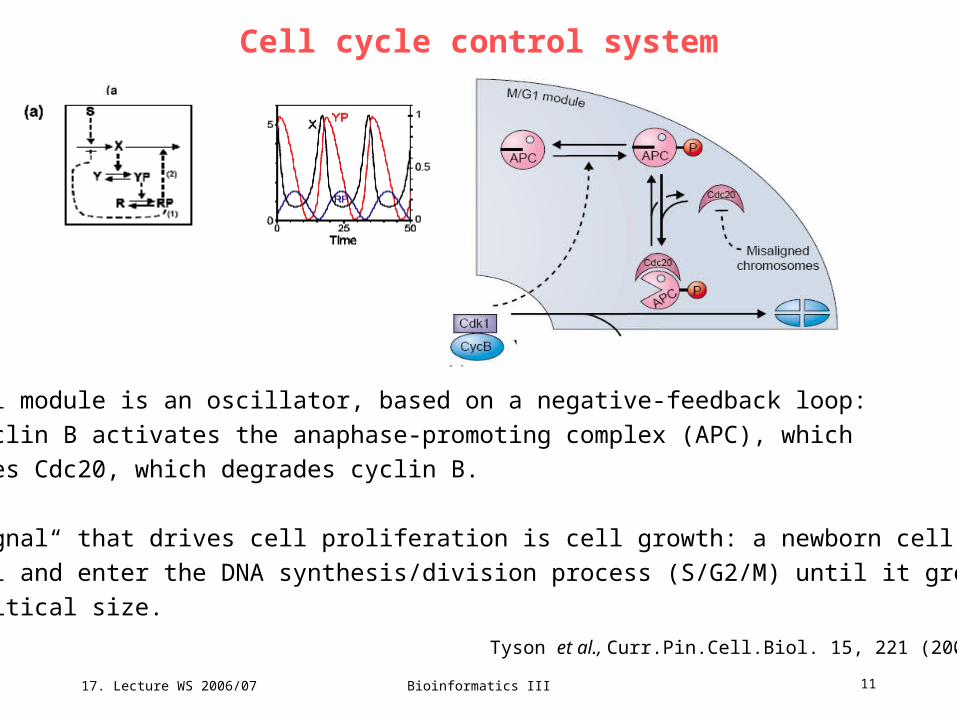
17. Lecture WS 2006/07
Bioinformatics III 11
Cell cycle control system
Tyson et al., Curr.Pin.Cell.Biol. 15, 221 (2003)
The M/G1 module is an oscillator, based on a negative-feedback loop:
Cdk1-cyclin B activates the anaphase-promoting complex (APC), which
activates Cdc20, which degrades cyclin B.
The „signal“ that drives cell proliferation is cell growth: a newborn cell cannot
leave G1 and enter the DNA synthesis/division process (S/G2/M) until it grows
to a critical size.

17. Lecture WS 2006/07
Bioinformatics III 12
cell-cycle machineryHysteresis and bistability were recently shown to occur in the activation/ inactivation of
CDK1/CDC2, an observation that confirmed a theoretical prediction by Novak and Tyson 10
years ago.
Bistability in the CDK1/CDC2 cycle arises from positive and double-negative feedback loops in
the reactions. CDK1/CDC2 activates its activator (the phosphatase CDC25) and inactivates its
inhibitors (the kinases Wee1 and Myt1).
Negative feedback from the anaphase-promoting complex (APC) turns the CDK1/CDC2
bistable switch into a relaxation oscillator that drives the cell cycle.
Intriguingly, CDC25 and Wee1 can be phosphorylated on multiple sites and can therefore
potentially exhibit bistability, which implies that the entire CDK/cyclin system can display
multiple steady states — this prediction is awaiting experimental verification.
Sequential bifurcations of multiple steady states provide more flexibility in the control of the cell
fate and allow for several checkpoints in the cell cycle.

17. Lecture WS 2006/07
Bioinformatics III 13
Cell cycle control system
The signal-response curve is
a plot of steady-state activity
of Cdk1-cyclin B as a
function of cell size.
Progress through the cell cycle is
viewed as a sequence of bifurcations.
A very small newborn cell is attracted
to the stable G1 steady state.
As it grows, it eventually passes the
saddle-point bifurcation SN3 where
the G1 steady state disappears.
The cell makes an irreversible
transition into S/G2 until it grows so
large that the S/G2 steady state
disappears, giving way to an infite
period oscillation (SN/IP).
Tyson et al., Curr.Pin.Cell.Biol. 15, 221 (2003)
Cyclin-B-dependent kinase activity soars, driving the cell into mitosis, and then plummets, as cyclin B is degraded by APC–Cdc20. The drop in Cdk1–cyclin B activity is the signal for the cell to divide, causing cell size to be halved from 1.46 to 0.73, and the control system is returned to its starting point, in the domain of attraction of the G1 steady state.

17. Lecture WS 2006/07
Bioinformatics III 14
signalling pathway: specificity?
The picture of interconnected signalling networks has replaced the earlier
concepts of discrete linear pathways, which relate extracellular signals to the
expression of specific genes, raising questions about the specificity of signal-
response events.
In fact, the protein complement that mediates signal transduction downstream of
RTKs is similar for all the RTK-mediated pathways.
Both GPCRs and RTKs activate kinase and phosphatase cascades, such as
mitogen-activated protein kinase (MAPK) cascades, that induce the expression of
nuclear transcription factors.
For any individual receptor pathway, there is no single protein or gene that is
responsible for signalling specificity. Instead, specificity is determined by the
temporal and spatial dynamics of downstream signalling components.

17. Lecture WS 2006/07
Bioinformatics III 15
receptor tyrosine kinases
A classical example is the distinct biological outcome of the PC12 cell-line
stimulation with EGF and nerve growth factor (NGF).
EGF induces transient MAPK activation, which results in cell proliferation,
whereas a sustained MAPK activation by NGF changes the cell fate and induces
cell differentiation.
However, the factors that control the kinetics of MAPK cascades are intricate.
MAPK cascades can generate bistable dynamics (in which two stable ‘on’ and ‘off’
steady states coexist), abrupt switches and oscillations, and their responses
depend dramatically on their subcellular localization and their recruitment to
scaffold proteins.

17. Lecture WS 2006/07
Bioinformatics III 16
Epidermal growth factor receptor signaling pathway
The EGFR signaling pathway is one of the most important pathways that regulate
growth, survival, proliferation, and differentiation in mammalian cells.
International consortium has assembled a comprehensive pathway map including
- EGFR endocytosis followed by its degradation or recycling,
- small GTPase-mediated signal transduction such as MAPK cascade, PIP
signaling, cell cycle, and GPCR-mediated EGFR transactivation via intracellular
Ca2+ signalling.
Map includes 211 reactions and 322 species taking part in reactions.
Species: 202 proteins, 3 ions, 21 simple molecules, 73 oligomers, 7 genes, 7 RNAs.
Proteins: 122 molecules including 10 ligands, 10 receptors, 61 enzymes (including 32 kinases), 3 ion
channels, 10 transcription factors, 6 G protein subunits, 22 adaptor proteins.
Reactions: 131 state transitions, 34 transportations, 32 associations, 11 dissociations, 2 truncations.
Oda et al. Mol.Syst.Biol. 1 (2005)

17. Lecture WS 2006/07
Bioinformatics III 17
Oda et al. Mol.Syst.Biol. 1 (2005)

17. Lecture WS 2006/07
Bioinformatics III 18
Architecture of signaling network: bow-tie structure
Oda et al.
Mol.Syst.Biol. 1 (2005)

17. Lecture WS 2006/07
Bioinformatics III 19
Network control
Several system controls define the overall behavior of the signaling network:
- 2 positive feedback loops
- Pyk2/c-Src activates ADAMs, which shed pro-HB-EGF so that the
amount of HB-EGF will be increased and enhance the signalling
- active PLC/ produces DAG which results in the cascading activation
of protein kinase C (PKC), phospholipase D, and PI5 kinase.
- 6 negative feedback loops
- inhibitory feed-forward paths
There are also a few positive and negative feedback loops that affect ErbB
pathway dynamics.
Oda et al. Mol.Syst.Biol. 1 (2005)

17. Lecture WS 2006/07
Bioinformatics III 20
Process diagram
Oda et al. Mol.Syst.Biol. 1 (2005)

17. Lecture WS 2006/07
Bioinformatics III 21
Modification and localization of proteins
Oda et al. Mol.Syst.Biol. 1 (2005)

17. Lecture WS 2006/07
Bioinformatics III 22
Precise association states between EGFR and adaptorsOda et al. Mol.Syst.Biol. 1 (2005)
Ellipsis in drawing association states of proteins using an ‘address’. (A) Precise association states between EGFR and adaptors. Three adaptor proteins, Shc, Grb2, and Gab1, bind to the activated EGFR via its autophosphorylated tyrosine residues. Shc binds to activated EGFR and is phosphorylated on its tyrosine 317. Grb2 binds to activated EGFR either directly or via Shc bound to activated EGFR. Gab1 also binds to activated EGFR either directly or via Grb2 bound to activated EGFR, and is phosphorylated on its tyrosine 446, 472, and 589.

17. Lecture WS 2006/07
Bioinformatics III 23
Cells of living organism sense their
environment and respond to
environmental stimuli.
Cellular signaling mechanisms govern how information
from the environment is decoded, processed and transferred to the appropriate
locations within the cell.
Signaling through the receptor tyrosine kinase (RTK) family of receptors regulates
a wide range of biological phenomena, including cell proliferation and
differentiation.
Integrated PW-DMC Model of Epidermal Growth Factor Receptor Trafficking and Signal Transduction
Diagram showing the compartments involved in
receptor trafficking and the receptor movement
pathways within the cell.
Resat et al. Biophys Journal 85, 730 (2003)

17. Lecture WS 2006/07
Bioinformatics III 24
Integrated Model of Epidermal Growth Factor Receptor Trafficking and Signal Transduction
Signaling pathways of various RTKs are reasonably well characterized.
Common features:
- receptor self-phosphorylation on tyrosine residues
- subsequent interaction with molecules containing SH2 and phospho-Tyr
residues.
The signal from the receptor is transmitted to downstream effector molecules
through a series of protein-protein interactions, such as the MAP kinase cascade.
Resat et al. Biophys Journal 85, 730 (2003)

17. Lecture WS 2006/07
Bioinformatics III 25
Integrated Model of Epidermal Growth Factor Receptor Trafficking and Signal Transduction
The EGF receptor can be activated by the
binding of any one of a number of different
ligands.
Each ligand stimulates a somewhat different
spectrum of biological responses.
The effect of different ligands on EGFR
activity is quite similar at a biochemical level
the mechanisms responsible for their
differential effect on cellular responses are
unkown.
After binding of any of its ligands, EGFR is
rapidly internalized by endocytosis.
Resat et al. Biophys Journal 85, 730 (2003)

17. Lecture WS 2006/07
Bioinformatics III 26
Integrated Model of Epidermal Growth Factor Receptor Trafficking and Signal Transduction
Different EGFR ligands vary in their ability to bind to EGFR as a function of
receptor microenvironment such as intravesicular pH.
After endocytosis, receptor-ligand complexes pass through several different
compartments that vary in their intravesicular milieu.
Receptor movement among cellular compartments („receptor trafficking“) can
exert a significant effect on the activity of the complexes.
The different intracellular compartments also vary in their access to some of the
substrates of the EGFR kinase.
This coupled relationship between substrate access and ligand-dependent
activity in different endocytic compartments suggests that trafficking could
function to „decode“ the information unique to each ligand.
Resat et al. Biophys Journal 85, 730 (2003)

17. Lecture WS 2006/07
Bioinformatics III 27
3 functions of trafficking
(1) controlling the magnitude of the signal
(2) controlling the specificity of the response
(3) controlling the duration of the response.
Understanding the relative contribution of these 3 aspects for any given
combination of cells, conditions, and ligands is very difficult
use computational models!
Next lecture: stochastic dynamic simulations of EGF pathway.
Resat et al. Biophys Journal 85, 730 (2003)

17. Lecture WS 2006/07
Bioinformatics III 28
temporal dynamics of signalling networks
simplified scheme of signalling routes starting at EGFR

17. Lecture WS 2006/07
Bioinformatics III 29
Spatial dimension of signalling networks
Activation of cell-surface receptors and their downstream targets leads to the
spatial relocation of multiple proteins within the cell.
During evolution, cells have developed not only means to control the temporal
dynamics of signalling networks, but also mechanisms for precise spatial sensing
of the relative localization of signalling proteins.
The regulation of signalling within the cellular space is pivotal for several
physiological processes, such as cell division, motility and migration.
Here, we will discuss how the basic principles of the control of reaction rates,
diffusive movement and directed transport underlie sophisticated signalling
mechanisms that provide spatial cues for cell division and transmit signals to
distant cellular targets.

17. Lecture WS 2006/07
Bioinformatics III 30
Regulation of signalling by membrane recruitmentReceptor stimulation triggers the mobilization of cytosolic adaptor proteins and enzymes to
cellular membranes. Subsequent phosphorylation results in the assembly of signalling
complexes on receptors, scaffolds and cytoskeletal elements. These spatial relocations are
effective control mechanisms that switch on signalling pathways.
A classic example is the control of the Ras/ MAPK cascade through the membrane
recruitment of son of sevenless (SOS) and RasGAP (which are a GEF and a GAP for the
small GTPase Ras, respectively), which is mediated by RTKs (e.g. by EGFR) and membrane-
bound scaffolds.
Although it has been previously proposed that the role of this recruitment is to increase
diffusion-limited rates (first-encounter rates), it has been recently shown that the function of
membrane localization is to amplify the number of complexes that are formed between the
signalling partners. SOS and RasGAP bound to EGFR are confined to a small volume near
the membrane that results in a 102–103-fold increase in the apparent affinity of these catalysts
for Ras. Computational simulations corroborate this theory; they show that in the absence of
the membrane recruitment, the cytoplasmic concentrations of SOS and RasGAP would have
to increase 102–103-fold to account for the observed rates of Ras activation/deactivation.
the spatial organization of the Ras circuit is crucial for the effective control of Ras activity.

17. Lecture WS 2006/07
Bioinformatics III 31
Localization determines signalling outputsThe localization of signalling proteins to distinct subcellular regions, such as internal
membranes and membrane microenvironments (including lipid rafts) modulates signalling
outputs.
Specific anchoring subunits direct the catalytic subunits of kinases and phosphatases, such as
cyclic AMP (cAMP)-dependent protein kinase, protein kinase C and serine/threonine protein
phosphatases PP1, PP2A and PP2B, to different cellular regions.
The general mechanism is to orientate broad-specificity enzymes towards specific targets and
physically separate them from undesirable substrates.
Discrete subcellular distribution enhances the specificity and fidelity of phosphorylations and
dephosphorylations that are catalysed by these kinases and phosphatases.
Qualitatively different patterns of signalling are generated by receptors and downstream effectors
that are associated with endosomes or the plasma membrane.
Likewise, the same protein cascades operate in surprisingly dissimilar ways when they are
localized to different cellular compartments. The input–output sensitivity of the MAPK cascade is
different for signalling from the plasma membrane, the Golgi apparatus and the endosomes.

17. Lecture WS 2006/07
Bioinformatics III 32
Spatial gradients of signalling activitiesIn the late 1990s, the concept of protein-activity gradients within a cell was proposed.
This concept has recently matured, when FRET-based biosensors enabled discoveries of
intracellular gradients of the active form of the small GTPase Ran and the phosphorylated
form of stathmin oncoprotein 18 (Op18/stathmin), which regulate the polymerization of the
microtubules.
Spatial gradients of protein activities organize signalling around cellular structures, such as
membranes, chromosomes and scaffolds, and provide positional cues for important
processes, including cell division.
During mitosis, the microtubule network changes from the radial architecture that stems from
the centrosome to a bipolar spindle. How this remarkable rearrangement occurs is not
completely understood.
It has been recently suggested that spatial gradients of several molecules that influence
microtubule dynamics, including Op18/stathmin and RanGTP, which interacts with the
nuclear-transport receptor importin-β, guide microtubule–kinetochore positioning during the
mitotic-spindle assembly.

17. Lecture WS 2006/07
Bioinformatics III 33
Spatial gradients of signalling activities
The basic prerequisite for signalling gradients is the spatial segregation of opposing reactions (for instance, kinase and phosphatase) in a universal protein-modification cycle. For a protein that is phosphorylated by a membrane-bound kinase and dephosphorylated by a cytosolic phosphatase, a gradient of the phosphorylated protein could exist — high concentration close to the membrane and low concentration within the cell. Based on measured values of protein diffusivity and kinase and phosphatase activities, it was estimated that phosphoprotein gradients might be large within the intracellular space. Spatial gradients of this kind have also been postulated for chemotaxis proteins in small bacterial cells.

17. Lecture WS 2006/07
Bioinformatics III 34
Future directionsQuantitative models that generate novel, experimentally testable hypotheses will have an
increasingly important role in post-genomic biology.
Future models will integrate data on the distinct spatio-temporal dynamics of signalling from
different cellular compartments and provide new insights into the connection between external
stimuli and the signalling outcome in terms of gene-expression responses.
Challenges of the combinatorial complexity of signalling networks and experimental
uncertainty in parameter values will be addressed by modular approaches, and stochastic and
pattern-oriented modelling.
The goal of the pattern-oriented approach is to predict and explain dynamic patterns of
cellular responses to a multitude of external cues and perturbations.
An exceedingly large number of quantitative and qualitative data patterns will facilitate the
verification of the proposed molecular mechanisms and exclude models that are too simplistic
and uncertain. These systems-level, data-driven models will generate new knowledge and
provide strategies for the regulation of the cellular machinery.
Understanding the mechanisms that underlie the functions of signalling networks will support
the identification of the critical controlling factors that will be targets for pharmacological
interventions in the treatment of human diseases.

17. Lecture WS 2006/07
Bioinformatics III 35
additional slides (not used)

17. Lecture WS 2006/07
Bioinformatics III 36
temporal dynamics of signalling networks
EGFR is the beststudied RTK. Together with other members of the ErbB family it
has a pivotal role in carcinogenesis.
Phosphorylation of tyrosine residues on EGFR leads to the recruitment and
activation of EGFR adaptor proteins and enzymes.
These events initiate signal propagation through multiple interacting cascades,
including
phospholipase C-γ (PLCγ),
phosphatidylinositol 3-kinase (PI3K)-AKT/protein kinase B (PKB) and
extracellular signal-regulated kinase (ERK)/MAPK pathways.
The complex temporal responses of multiple downstream EGFR targets cannot be
explained by qualitative arguments.
Reliable and testable computational models are required to predict signalling
dynamics.

17. Lecture WS 2006/07
Bioinformatics III 37
Phosphoprotein gradients in MAPK cascades
Phosphoprotein gradients are hallmarks of kinase/phosphatase cascades, including the MAPK cascades. MAPK cascades contain three interconnected cycles of a MAPK, a MAPK kinase (MAPKK) and a MAPKK kinase (MAPKKK). In the MAPK/ERK cascade, the cascade consists of ERK, MEK and Raf. Upon RTK stimulation and Ras activation, the cytosolic Raf is recruited to the cell membrane, where it binds to and phosphorylates MEK on two serine residues. Phosphorylated MEK drifts into the cell interior, where it phosphorylates ERK on threonine and tyrosine residues. MEK is dephosphorylated in the cytoplasm; therefore, spatial gradients of phosphorylated MEK, and subsequently phosphorylated ERK, might occur. Calculations show that these gradients can be precipitous, decreasing the strength of the phosphorylation signal to the nucleus. If the cascade has more levels, then the phosphorylation signal would reach further into the cell, which indicates that one of the reasons that cascades exist might be to promote signal propagation. The cascades that are found in eukaryotes tend to have more levels than the cascades that exist in prokaryotes, an observation that can be related to larger distances of signal propagation in eukaryotes.





























