Embed Size (px)
Citation preview

1
ATLANTIC COD BI3063 H10J. Mork
Both with respect to potential harmful genetic effects on fish stocks from human activities, and the available management tools for remedying the situation, Hallerman mostly use examples from salmonids, and in particular anadromous Pacific salmonids.Among the targeted human activities, the fish farming industry and its effects has atracted particular attention.
The Danish population geneticist Dorte Bekkevold and collegues (Bekkevold et al. 2005) argue that the problems encountered in salmonid farming apply to cod farming as well (cf next slide).
Web site for downloading Bekkevold et al. (2005):http://icesjms.oxfordjournals.org/cgi/reprint/63/2/198

2
BI3063 H10J. Mork
ATLANTIC COD

3
BI3063 H10J. Mork
ATLANTIC COD

4
BI3063 H10J. Mork
ATLANTIC COD
In the following, we shall explore various aspects of cod and In the following, we shall explore various aspects of cod and gadoid genetics which have a bearing on whether these gadoid genetics which have a bearing on whether these species are vulnerable to various human activities (wild stock species are vulnerable to various human activities (wild stock exploitation, introgression by escaped farmed fish) in a way exploitation, introgression by escaped farmed fish) in a way similar to the salmon. similar to the salmon.

5
BI3063 H10J. Mork
ATLANTIC COD
Genetic effects from ranching; lessons come mainly from salmonid farming/ranching:
Changes in genetic composition of farmed stock over timeChanges in genetic composition of farmed stock over time
Agents:Agents:1. Random genetic drift2. Domestication selection (no predators, excess food, vaccines)3. Selection for performance traits reduces genetic variability4. Mating of close relatives
Effects:Effects:1. Reduced genetic variability2. Reduced shyness3. Loss of natural adaptivity4. Harmful inbreeding effects

6
BI3063 H10J. Mork
Knowledge on gadoid population structure exists for
Cod (Gadus morhua L.)
Haddock (Melanogrammus aeglefinus)
Whiting (Merlangus merlangius)
Saithe (Pollachius virens)
Norway pout (Trisopterus esmarkii)
Poor cod (Trisopterus minutus)
Blue whiting (Micromesistius poutassou)
Hake (Merluccius merluccius)
ATLANTIC COD

7
BI3063 H10J. Mork
ATLANTIC COD
(Source: Smith & Mork unpublished)

8
BI3063 H10J. Mork
ATLANTIC COD
Milestone studies on the genetic population structure of the Atlantic cod are:
Schmidt (1930) Meristics used in cod study throughout rangeTåning (1946) Showed meristics are affected by selection (temperature)Sick (1961) Cod and whiting haemoglobin polymorphismSick (1965 a,b) Cod HbI allele frequencies throughout rangeFrydenberg et al. (1965) Cod HbI allele frequencies Norwegian watersMøller (1968) Cod HbI, Tf and blood type Norwegian watersKarpov & Novikov (1980) Cod HbI affected by natural selection (temperature)Mork et al. (1985) Cod tissue allozymes througout rangeDahle (1992) Cod (NEAC& NCC) mtDNA RFLP Pogson (1995) Cod allozyme and PanI cDNA RFLP) throughout rangeArnason & Palsson (1996) Cod (NEAC & NCC) mt Cyt b sequences Arnason (1998) Cod mtDNA Cyt b sequences throughout rangeKarlson & Mork (2001) Showed PanI heavily affected by natural selectionKarlsson & Mork (2005) Showed microsat loci affected by natural selectionNielsen et al. (2005) Microsat hitchhike selection create false impression of isolation
Are 3rd codon position silent substitutions (e.g. at mtDNA Cyt b) the great white hope?

9
BI3063 H10J. Mork
ATLANTIC COD
Arnason, E., Palsson, S. 1996. Mitochondrial cytochrome b DNA sequence variation of Atlantic cod (Gadus morhua) from Norway. Molecular Ecology 5: 715-724.
Arnason, E. 1998. Mitochondrial cytochrome b DNA sequence variation of Atlantic cod Gadus morhua. In The implications of localized fishery stocks (Ed.) von Herbing, I.H., Kornfield, I., Tupper, M., Wilson, J. Natural Resource Agriculture and Engineering Service New York. pp 129-137.
Frydenberg, O., Møller, D., Nævdal, G., Sick, K. 1965. Haemoglobin polymorphism in Norwegian cod populations. Hereditas 53: 257-272.
Godø, O.R., Moksness, E. 1987. Growth and maturation of Norwegian coastal cod and northeast Arctic cod under different conditions. Fisheries Research 5: 235-242.
Godø, O.R. 1995. Transplantation-tagging experiments in preliminary studies of migration of cod off Norway. ICES Journal of Marine Science 52: 955-962.
Karlsson, S., Mork, J. 2001. Evidence for natural selection at the synaptophysin locus (Syp1) in a natural population of cod (Gadus morhua L.). ICES C.M.2001/L:11
Karpov, A.K., Novikov, G.G. 1980. Hemoglobin alloforms in cod, Gadus morhua (gadiformes, gadidae), their functional characteristics and occurrence in populations. Journal of Ichthyology (In Russian) 20(6): 45-50.
Mork, J., Ryman, N., Ståhl, G., Utter, F., Sundnes, G. 1985. Genetic variation in Atlantic cod (Gadus morhua) throughout its range. Canadian Journal of Fisheries & Aquatic Sciences 42: 1580-1587.
Cod genetics literature

10
BI3063 H10J. Mork
ATLANTIC COD
Møller, D., 1968. Genetic diversity in spawning cod along the Norwegian coast. — Hereditas60 1–32.Møller, D., 1969. The relationship between arctic and coastal cod in their immature stages illustrated by frequencies of genetic characters. — FiskDir. Skr. (Havunders.)15 220–233.
Nielsen, E.E., Hansen, M.M. & Meldrup, D. 2006. Evidence of microsatellite hitch-hiking selection in Atlantic cod (Gadus morhua L.): implications for inferring population structure in nonmodel organisms.Molecular Ecology 15 (11): 3219-3229.Nielsen, E. MacKenzies, B., Magnussen, E. and Meldrup, D. 2007. Historical analysis of Pan I in Atlantic cod (Gadus morhua): temporal stability of allele frequencies in the southeastern part of the species distribution.Can. J. Fish. Aquat. Sci. 64:(10): 1448-1455.
Pogson, G.H., Mesa, K.A., Boutilier, R.G. 1995. Genetic population structure and gene flow in the Atlantic cod Gadus morhua: a comparison of allozyme and nuclear RFLP loci. Genetics 139: 375-385. Pogson, G.H. 2001. Nucleotide polymorphism and natural selection at the Panthophysine (Pan I) locus in the Atlantic cod, Gadus morhua (L.). Genetics 157: 317-330.
Sick, K., 1961. Haemoglobin polymorphism in fishes. — Nature, Lond.192 894–896. Sick, K. 1965a. Haemoglobin polymorphism of cod in the Baltic and Danish Belt Sea. Hereditas 54: 19-48.Sick, K. 1965b. Haemoglobin polymorphism of cod in the North Sea and the North Atlantic Ocean. Hereditas 54: 49-69.
Rollefsen, G., 1933. The otoliths of the cod. — FiskDir. Skr. (Havunders.)4 (3), 1–14.
Wilkins, N. P., 1971. Haemoglobin polymorphism in cod, whiting and pollack in Scottish waters. — Rapp. P.-v. Réun. Cons. perm. int. Explor. Mer161 60–64.
Tåning, A.V. 1946: Stage of determination of vertebrae in teleostean fishes. Nature 157: 594-595.
Cod genetics literature cont'd

11
BI3063 H10J. Mork
ATLANTIC COD
Cod population genetic studies at Trondhjem Biological Station:
Mork. J., Giskeødegard, R., Sundnes, G. 1983. Haemoglobin polymorphism in Gadus morhua: genotypic differences in maturing age and within-season gonad maturation. Helgolander Meerescuntersuchungen 36: 313-322.
Mork, J., Ryman, N., Ståhl, G., Utter, F., Sundnes, G. 1985. Genetic variation in Atlantic cod (Gadus morhua) throughout its range. Canadian Journal of Fisheries & Aquatic Sciences 42: 1580-1587.
Mork. J., Sundnes, G. 1985a. 0-Group cod (Gadus morhua) in captivity: differential survival of certain genotypes. Helgolander Meerescuntersuchungen 39: 63-70.
Mork. J., Sundnes, G. 1985b. The haemoglobin polymorphism in cod (Gadus morhua): allele frequency variation between year classes in a Norwegian fjord stock. Helgolander Meerescuntersuchungen 39: 55-62. Mork, J., Giæver, M. 1999. Genetic structure of cod along the coast of Norway; results from isozyme studies. Sarsia 84: 157-168.
Karlsson, S., and Mork, J. 2003. Selection-induced variation at the. pantophysin locus (PanI) in a Norwegian fjord population of. cod (Gadus morhua L.). Molecular Ecology 12: 3265-3274.
Karlsson, S. & Mork, J. 2005. Deviation from Hardy–Weinberg equilibrium, and temporal instability in allele frequencies at microsatellite loci in a local population of Atlantic cod. ICES Journal of Marine Science: Journal du Conseil 2005 62(8):1588-1596.
Cod genetics literature cont'd

12
BI3063 H10J. MorkATLANTIC CODCase studies in cod

13
BI3063 H10J. Mork
Deviation from Hardy–Weinberg equilibrium, and temporal instability in allele frequencies at microsatellite loci in a local population of Atlantic cod
S. Karlsson & J. Mork
Abstract:A total of 1455 spawning cod, sampled from a local spawning area in Trondheimsfjord (Norway) between 1985 and 2002, was screened at the microsatellite loci Gmo132 and Gmo2. Samples from 15 spawning years comprising 29 consecutive cohorts were analysed. At the Gmo132 locus, but not at Gmo2, allele frequencies varied significantly among sampling years as well as cohorts, corresponding to FST-values of 0.004 and 0.006, respectively. Both loci showed examples of significant deviations from Hardy–Weinberg expectation within sampling years as well as cohorts, manifested as deficiencies of heterozygotes. Combining the p-values from the single tests (Fisher's method) revealed an overall significant p-value for deviation from the Hardy–Weinberg expectations at Gmo132 but not at Gmo2. Trend tests showed significant HW
deficiencies at both loci for annual samples but not for cohorts. Possible reasons for the deficiencies were discussed; inter
alia the existence of null alleles, or a form of pseudo Wahlund effect due to a patchy distribution of habitats for settling O-group cod in the Trondheimsfjord. It was noted that there might be a relationship between the relatively high temporal within-population variability of allele frequencies at Gmo132 and the fact that among microsatellite loci studied so far,
Gmo132 is the one that usually shows the highest genetic differentiation geographically in cod.
ATLANTIC CODCase studies in cod

14
BI3063 H10J. Mork
ATLANTIC COD
Evidence of microsatellite hitch-hiking selection in Atlantic cod (Gadus morhua L.): implications for inferring population structure in nonmodel organisms
Nielsen, E.E., Hansen, M.M. & Meldrup, D.
Abstract:Microsatellites have gained wide application for elucidating population structure in nonmodel organisms. Since they are generally noncoding, neutrality is assumed but rarely tested. In Atlantic cod (Gadus morhua L.), microsatellite studies have revealed highly heterogeneous estimates of genetic differentiation among loci. In particular one locus, Gmo 132, has demonstrated elevated genetic differentiation. We investigated possible hitch-hiking selection at this and other microsatellite loci in Atlantic cod. We employed 11 loci for analysing samples from the Baltic Sea, North Sea, Barents Sea and Newfoundland covering a large part of the species' distributional range. The 'classical' Lewontin–Krakauer test for selection based on variance in estimates of FST and (standardized genetic differentiation) revealed only one significant pairwise test (North Sea–Barents Sea), and the source of the elevated variance could not be ascribed exclusively to Gmo 132. In contrast, different variants of the recently developed ln Rθ test for selective sweeps at microsatellite loci revealed a high number of significant outcomes of pair-wise tests for Gmo 132. Further, the presence of selection was indicated in at least one other locus. The results suggest that many previous estimates of genetic differentiation in cod based on microsatellites are inflated, and in some cases relationships among populations are obscured by one or more loci being the subject to hitch-hiking selection. Likewise, temporal estimates of effective population sizes in Atlantic cod may be flawed. We recommend, generally, to use a higher number of microsatellite loci to elucidate population structure in marine fishes and other nonmodel species to allow for identification of outlier loci that are subject to selection.
Case studies in cod

15
BI3063 H10J. Mork
Abstract:We investigated temporal genetic differentiation at the pantophysin (Pan I) locus in four Atlantic cod (Gadus morhua) populations from the southeastern part of the species distribution: the Baltic Sea, the North Sea, the Faroe Plateau, and the Faroe Bank. Historical otolith collections enabled investigation of allele frequency variation over time periods up to 69 years employing Pan I primers specifically designed for partially degraded DNA. Small and nonsignificant temporal changes in Pan I allele frequencies were observed in the four populations. Simultaneous microsatellite analysis revealed similar temporal genetic stability with temporal FST values ranging from 0 to 0.006, suggesting limited demographic changes. Sea surface temperature, which has been suggested as the primary driver for the geographical distribution of Pan I alleles in cod, showed no long-term trend although temperature has increased since the mid-1990s. Our study demonstrates that populations in the southeastern part of the species range has been characterized by very high frequencies of the Pan IA allele for many decades, and accordingly, Pan I serves as a reliable marker for genetic stock identification on a macrogeographical scale.
Historical analysis of Pan I in Atlantic cod (Gadus morhua): temporal stability of allele frequencies in the southeastern part of the species distribution
Nielsen, E. E.; MacKenzie, B.R.; Magnussen, E. & Meldrup, D.
Canadian Journal of Fisheries and Aquatic Sciences, Volume 64, Number 10, 1 October 2007 , pp. 1448-1455(8)
ATLANTIC CODCase studies in cod

16
BI3063 H10J. Mork
BLUE WHITING

17
BI3063 H10J. Mork
Blue whiting population genetic studies at Trondhjem Biological Station:
Mork, J. & Giæver, M. 1995. Genetic variation at isozyme loci in blue whiting from the north-east Atlantic. Journal of Fish Biology 46: 462-468.
Giæver, M. & Mork, J. 1995. Further studies on the genetic population structure of the blue whiting (Micromesistius poutassou) in the north-east parts of the distribution range. ICES C.M. 1995/H:11.
Mork, J. & Giæver, M. 1993. The genetic population structure of the blue whiting (Micromesistius poutassou). ICES C.M. 1993/H:5.
Giæver & Stien, M. 1997. Population genetic substructure in blue whiting based on allozyme data. Journal of Fish Biology 52: 782-795.
Ryan, A.W., Mattiangeli, V. & Mork, J. 2005. Genetic differentiation of blue whiting (Micromesistius poutassou Risso) populations at the extremes of the species range and at the Hebrides-Porcupine Bank spawning grounds. ICES Journal of Marine Sciences, 62; 948-955).
BLUE WHITINGCase studies in blue whiting

18
BI3063 H10J. Mork
BLUE WHITINGCase studies in blue whiting
Giæver & Stien, M. 1997. Population genetic substructure in blue whiting based on allozyme data. Journal of Fish Biology 52: 782-795.

19
BI3063 H10J. Mork
HADDOCK

20
BI3063 H10J. Mork
HADDOCK
Material:Material:Haddock samples from the Russian boarer to Haddock samples from the Russian boarer to the west coast of Sweden.the west coast of Sweden.
MethodsMethodsStarch gel electrophoresis and histochemical Starch gel electrophoresis and histochemical staining of 10 polymorphic tissues enzyme loci.staining of 10 polymorphic tissues enzyme loci.
Main result:Main result: No statistically significant genetic differentiationNo statistically significant genetic differentiationin the investigated area. in the investigated area.
Case study in haddock

21
BI3063 H10J. Mork
SAITHE

22
BI3063 H10J. Mork
WHITING

23
BI3063 H10J. Mork
SAITHE & WHITING
Case study in saithe and whiting

25
BI3063 H10J. Mork
NORWAY POUTCase study in Norway pout

26
BI3063 H10J. Mork
HAKE

27
BI3063 H10J. Mork
HAKECase study in hake

28
BI3063 H10J. Mork
POOR COD

29
BI3063 H10J. Mork
VNTR variability in Atlantic poor cod (Trisopterus minutus minutus) throughout its range: single locus minisatellite data suggest reproductive isolation for the Faroe Bank
population
Mattiangeli, V., Galvin, P., Ryan, A., Mork, J., & Cross, T.
AbstractNine samples of Atlantic poor cod, Trisopterus minutus minutus, collected from the Bay of Biscay to Trondheimsfjord, Norway, were analysed using three minisatellite DNA loci, amplified using PCR and screened with an automated sequencer. One locus was found to be polymorphic. FST analysis using the polymorphic locus indicated that 2.2% of the total genetic diversity detected was due to differences among samples (FST = 0.022; P < 0.001). The only significant pair-wise heterogeneity was found between the Faroe Bank and each of the other samples. This is consistent with previous analyses of the same samples using 11 polymorphic allozyme loci.
Journal: Fisheries research ISSN 0165-7836
POOR CODCase study in poor cod

30
BI3063 H10J. Mork
Norteast arctic cod (NEAC) Norteast arctic cod (NEAC)
and and
Norwegian coastal cod (NCC)Norwegian coastal cod (NCC)
Special case: NEAC and NCC)

31
BI3063 H10J. Mork
Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)
Cod sampling sites throughout the distribution rangeCod sampling sites throughout the distribution range
AllozymesAllozymes

32
BI3063 H10J. Mork
Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)
Left: UPGMA Dendrogram leaving out not significant bifurcations. Left: UPGMA Dendrogram leaving out not significant bifurcations. Right: Comparison of allele frequencies at polymorphic loci in all localities.Right: Comparison of allele frequencies at polymorphic loci in all localities.
AllozymesAllozymes
33
BI3063 H10J. Mork
Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)
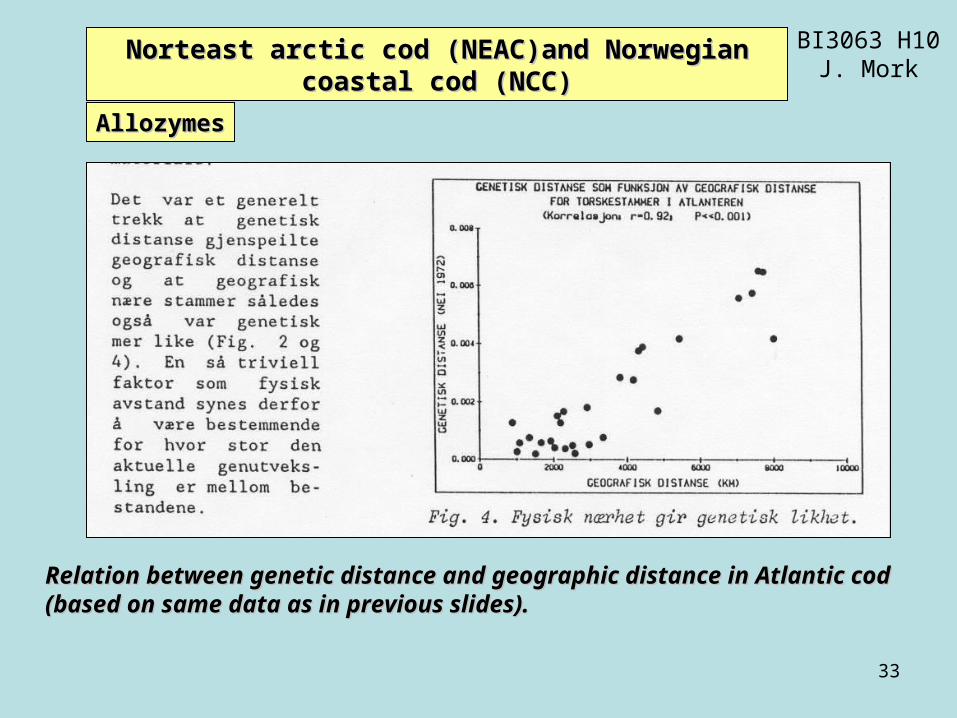
Relation between genetic distance and geographic distance in Atlantic codRelation between genetic distance and geographic distance in Atlantic cod(based on same data as in previous slides).(based on same data as in previous slides).
AllozymesAllozymes

34
BI3063 H10J. Mork
Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)
Left: Side-by-side comparison of allele frequencies at the 11 most polymorphic allozyme loci in Left: Side-by-side comparison of allele frequencies at the 11 most polymorphic allozyme loci in NEAC and NCC. Right: Small arrows point at the level of genetic distances between cod stocks NEAC and NCC. Right: Small arrows point at the level of genetic distances between cod stocks compared to usual levels at the population level.compared to usual levels at the population level.
AllozymesAllozymes

35
BI3063 H10J. Mork
Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)
FFST ST values for intra-specific values for intra-specific groupings in cod compared groupings in cod compared to other species: to other species: From left to right: Atlantic From left to right: Atlantic cod, atlantic salmon, brown cod, atlantic salmon, brown trout, rainbow trout, house trout, rainbow trout, house mouse, Drodophila, and mouse, Drodophila, and man.man.
The hatched upper part of The hatched upper part of the bars represent the Fthe bars represent the FSTST vales.vales.
AllozymesAllozymes

36
BI3063 H10J. Mork
Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)Norteast arctic cod (NEAC)and Norwegian coastal cod (NCC)
Karpov & Novikov (1980) Karpov & Novikov (1980) suggested that the south-north suggested that the south-north HbIHbI allele frequency cline allele frequency cline observed on both sides of the observed on both sides of the Atlantic can be explained by a Atlantic can be explained by a cline due to natural selection by cline due to natural selection by temperatures. temperatures.
This simultaneously suggested a This simultaneously suggested a new explanation for the observed new explanation for the observed HbIHbI allele frequency cline along allele frequency cline along the Norwegian coast NCC) and the Norwegian coast NCC) and into the Barents Sea (NEAC).into the Barents Sea (NEAC).
In the Trondheimsfjord cod, In the Trondheimsfjord cod, HbI HbI genotypic growth differences genotypic growth differences seem to concord with the seem to concord with the HbI HbI
genotypic differences in Ogenotypic differences in O22 affinity by temperature (Figure to affinity by temperature (Figure to the left).the left).
HaemoglobinsHaemoglobins

37
BI3063 H10J. Mork
Gadoid phylogenyGadoid phylogeny

38
NFFR project I 309.007NFFR project I 309.007
Phylogeny and genetic Phylogeny and genetic structure in common benthic structure in common benthic fish species of the east Atlantic fish species of the east Atlantic
AllozymesAllozymes

39
BI3063 H10J. Mork
Gadoid Gadoid phylogenyphylogeny
AllozymesAllozymes

40
BI3063 H10J. Mork
NFFR project I 309.007NFFR project I 309.007
Phylogeny and genetic structure Phylogeny and genetic structure in common benthic fish species in common benthic fish species of the east Atlantic of the east Atlantic
Gadoid Gadoid phylogenyphylogeny
AllozymesAllozymes

41
BI3063 H10J. Mork
Gadoid Gadoid phylogenyphylogeny
AllozymesAllozymes

42
BI 3063 J. Mork H08
Genetic and biologic stock management
(From Bakke & Johansen)
For comparison, the time since divergence between Atlantic cod and Pacific cod is probably around half a million years.
Gadoid Gadoid phylogenyphylogeny
mt DNA sequencesmt DNA sequences

43
BI3063 H10J. MorkGadoid Gadoid
phylogenyphylogenyAllozymesAllozymes

44
BI3063 H10J. Mork
Gadoid phylogenyGadoid phylogenyAllozymesAllozymes

45
BI3063 H10J. Mork
Gadoid phylogenyGadoid phylogeny
Comparing Comparing measures measures of genetic of genetic variability variability in gadoidin gadoidspecies.species.
AllozymesAllozymes

46
BI3063 H10J. Mork
Gadoid phylogenyGadoid phylogenyAllozymesAllozymes

47
BI3063 H10J. Mork
Gadoid phylogenyGadoid phylogeny
INTERPRETATION OF THE INTRA-SPECIFIC DENDROGRAMS ON PREVIOUS SLIDEINTERPRETATION OF THE INTRA-SPECIFIC DENDROGRAMS ON PREVIOUS SLIDE
AllozymesAllozymes

48
BI3063 H10J. Mork
Gadoid phylogenyGadoid phylogeny
List of P-values fromList of P-values fromtests of intra-specifictests of intra-specificgenetic differentiationgenetic differentiationbetween geographic between geographic areas in 5 gadoid areas in 5 gadoid species.species.
Only P-values from Only P-values from the loci that showed the loci that showed the the largest largest genetic genetic differences are differences are shown, and even they shown, and even they are all far from are all far from significance.significance.
AllozymesAllozymes

49
BI3063 H10J. Mork
Gadoid phylogenyGadoid phylogeny
Marianne Giæver at Trondhjem Biological Marianne Giæver at Trondhjem Biological Station wrote her doctoral Thesis on the Station wrote her doctoral Thesis on the genetic differentiation in three gadoid species; genetic differentiation in three gadoid species; cod, haddock, and blue whiting.cod, haddock, and blue whiting.
One of the main conclusions was that in all One of the main conclusions was that in all the species, the genetic differentiation the species, the genetic differentiation appeares to be caused by "Isolation by appeares to be caused by "Isolation by distance". distance".
Typically, it was the populations at the fringes of the species' range that were the genetically most divergent.
This is consistent with the meta-population concept.
AllozymesAllozymes

50
BI3063 H10J. Mork
Gadoid phylogenyGadoid phylogeny
With respect to genetic stock management, the above treatment of genetic With respect to genetic stock management, the above treatment of genetic studies and considerations suggest that the studies and considerations suggest that the gadoids are more resilient than gadoids are more resilient than anadromousanadromous salmonids to some human activities (e.g. over-exploitation, salmonids to some human activities (e.g. over-exploitation, genetic introgression by farmed fish escapes, and disease transfer from captive genetic introgression by farmed fish escapes, and disease transfer from captive stocks).stocks).
This is This is notnot to say that gadoids are to say that gadoids are immuneimmune to effects from over-exploitation and to effects from over-exploitation and mis-management. mis-management.
Probably, many species and populations are more vulnerable in times of Probably, many species and populations are more vulnerable in times of climate changes, like the situation is today. climate changes, like the situation is today.
There is still every reason to conduct stock management by the pre-cautionary There is still every reason to conduct stock management by the pre-cautionary principle, with respect to biological as well as genetic and evolutionary hazards. principle, with respect to biological as well as genetic and evolutionary hazards.
Summing upSumming up

51
BI3063 H10J. Mork
Gadoid phylogenyGadoid phylogeny






























