Embed Size (px)
Citation preview

Anti-CD4 and immune checkpoint antibody synergy
1
Robust antitumor effects of combined anti-CD4-depleting antibody and anti-PD-1/PD-L1 1
immune checkpoint antibody treatment in mice 2
3
Satoshi Ueha1, *, Shoji Yokochi1, 2, *, Yoshiro Ishiwata1, 2, *, Haru Ogiwara1, Krishant Chand1, 4
Takuya Nakajima1, Kosuke Hachiga1, 2, Shigeyuki Shichino1, Yuya Terashima1, Etsuko Toda1, 5
Francis HW Shand3, Kazuhiro Kakimi4, Satoru Ito1, 2, and Kouji Matsushima1 6
7 1Department of Molecular Preventive Medicine, Graduate School of Medicine, The University 8
of Tokyo; 2IDAC Theranostics, Inc.; 3Department of Pharmacology and Therapeutics, The 9
University of Melbourne; 4Department of Immunotherapeutics, The University of Tokyo 10
Hospital. 11 * S. Ueha, S. Yokochi and Y. Ishiwata contributed equally to this paper. 12
13
Running title: Anti-CD4 and immune checkpoint antibody synergy 14
15
Keywords: cancer treatment, CD4 depletion, immune checkpoint antibodies, CTL, Treg 16
17
Correspondence: Kouji Matsushima 18
Department of Molecular Preventive Medicine, Graduate School of Medicine, The University of 19
Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-0033 Japan 20
Telephone: 81-3-5841-3431; E-mail: [email protected] 21
22
Conflicts of interest: SY, SU, SI and KM own stock in IDAC Theranostics, Inc. We declare no 23
other conflicts of interest. 24
Financial support: This work was supported by the Japan Science and Technology Agency 25
CREST program; Grants-in-Aid for Scientific Research (C) 25460491 (to S.U.) and (B) 26
25293113 (to K.M.) from the Japanese Ministry of Education, Culture, Sports, Science and 27
Technology; and Health and Labor Science Research Grants for Research for Promotion of 28
Cancer Control (Applied Research for Innovative Treatment of Cancer). 29
30
Word count: 5,967/ 6,000 31
Figures: 6 main, 9 supplementary 32
Tables: 2 supplementary 33
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
2
ABSTRACT 34
Depletion of CD4+ cells in tumor-bearing mice has strong antitumor effects. However, the 35
mechanisms underlying these effects and the therapeutic benefits of CD4+ cell depletion relative 36
to other immunotherapies have not been fully evaluated. Here, we investigated the antitumor 37
effects of an anti-CD4-depleting monoclonal antibody (mAb) as a monotherapy or in 38
combination with immune checkpoint mAbs. In B16F10, Colon 26 or LLC subcutaneous tumor 39
models, administration of the anti-CD4 mAb alone had strong antitumor effects that were 40
superior to those elicited by CD25+ Treg depletion or other immune checkpoint mAbs, and 41
which were completely reversed by CD8+ cell depletion. CD4+ cell depletion led to the 42
proliferation of tumor-specific CD8+ T cells in the draining lymph node and increased 43
infiltration of PD-1+CD8+ T cells into the tumor, with a shift towards type I immunity within the 44
tumor. Combination treatment with the anti-CD4 mAb and immune checkpoint mAbs, 45
particularly anti-PD-1 or anti-PD-L1 mAbs, synergistically suppressed tumor growth and 46
greatly prolonged survival. To our knowledge, this work represents the first report of robust 47
synergy between anti-CD4 and anti-PD-1 or anti-PD-L1 mAb therapies. 48
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
3
INTRODUCTION 49
Immune checkpoint modulators such as those targeting cytotoxic 50
T-lymphocyte-associated antigen-4 (CTLA-4) and programmed cell death-1 (PD-1) have 51
attracted attention due to their extraordinary antitumor effects in patients with advanced 52
melanoma, lung cancer, and renal cancer (1, 2). A monoclonal antibody (mAb) against CTLA-4 53
(ipilimumab) that enhances both early T-cell activation and CTL function was approved for 54
treatment of patients with advanced melanoma in the USA in 2011. An anti-PD-1 mAb 55
(nivolumab) that protects activated T cells from exhaustion in peripheral tissues was approved 56
for treatment of patients with melanoma in Japan and in the US in 2014. In addition, other 57
mAbs against CTLA-4 (tremelimumab), PD-1 (pembrolizumab) and programmed death-ligand 58
1 (PD-L1, a ligand for PD-1) are currently undergoing clinical trials to evaluate their antitumor 59
efficacy. However, despite clear survival benefits in a subset of tumor patients, other groups of 60
patients are refractory to these single agent therapies. 61
62
Combination therapies comprising immune checkpoint modulators that have different 63
points of action, targeting for example the activation and expansion of T cells in lymphoid 64
tissues and the exhaustion and deletion of T cells in the effector site, represent promising 65
strategies for tumor immunotherapy (1). Synergistic anti-tumor effects in advanced melanoma 66
have been reported with a combination of anti-CTLA-4 and anti-PD-1 mAbs (3). The antitumor 67
efficacy of other combinations of regulators of lymphocyte activation and expansion (e.g. 68
Lymphocyte activation gene-3/LAG-3, OX40/CD134) and of lymphocyte exhaustion and 69
deletion (e.g. T-cell immunoglobulin mucin-3/TIM-3, 4-1BB/CD137, B- and T-lymphocyte 70
attenuator/BTLA, glucocorticoid-induced TNF-receptor/GITR) are currently under investigation. 71
Because immune checkpoint modulators play both positive and negative roles in the immune 72
inhibitory pathway with some redundancy, identification of optimal therapeutic combinations 73
remains a considerable challenge. 74
75
Another approach to immune checkpoint modulation involves depleting 76
immunosuppressive leukocyte populations such as forkhead box P3 (Foxp3)+CD25+ regulatory 77
T cells (Treg), Th2 cells, T regulatory (Tr) 1/3 cells (4), myeloid-derived suppressor cells 78
(MDSC) and indoleamine-2,3-dioxygenase (IDO)+ plasmacytoid DCs (pDC) (5-7). Several 79
groups have suggested that depletion of CD4+ cells, including Tregs, Th2 cells, Tr1/3 cells, and 80
a subpopulation of MDSCs and pDCs, results in strong antitumor effects in mouse models due 81
to the enhancement of CTL responses (8-12). These antitumor effects may be associated with 82
the modulation of multiple immune checkpoints caused by CD4+ cell depletion. However, the 83
relative advantage of CD4+ cell depletion over other immune checkpoint mAb-based treatments 84
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
4
remains unclear. Encouraged by the positive reports surrounding the benefits of anti-CD4 mAb 85
treatment in mice, and by the recent clinical data supporting anti-CTLA-4 and anti-PD-1 mAb 86
therapies, here we examine whether treatments that combine an anti-CD4 mAb and immune 87
checkpoint modulators produce synergistic antitumor activity. 88
89
Thus, in the present study we used comprehensive immunologic analyses to compare the 90
antitumor effects of an anti-CD4-depleting mAb with those of a variety of mAbs against 91
immune checkpoint molecules, including PD-1, PD-L1, PD-L2, CTLA-4, OX40, LAG-3, 92
TIM-3, BTLA and GITR, in mouse subcutaneous tumor models. We also investigated the 93
antitumor effects of treatments that combined an anti-CD4 mAb and antibodies against these 94
immune checkpoint molecules. We report that treatment with an anti-CD4 mAb alone induces 95
strong antitumor effects and expansion of tumor-specific CD8+ T cells, and that combination of 96
an anti-CD4 mAb with anti-PD-1 or anti-PD-L1 mAbs results in striking synergy in the 97
suppression of tumor growth.98
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
5
MATERIALS AND METHODS 99
Mouse. Seven-week-old female C57BL/6 and male BALB/c mice were purchased from Japan 100
SLC. Fluorescent ubiquitination-based cell cycle indicator (Fucci) double transgenic mice were 101
generated by crossbreeding FucciG1-#639 and FucciS/G2/M-#474 animals (obtained from Dr. A. 102
Miyawaki through the RIKEN BRC) as described previously (13). Mice transgenic for the 103
gp100 melanoma antigen-specific Pmel-1-TCR or the ovalbumin specific OT-I TCR were 104
purchased from The Jackson Laboratory. Each experimental group contained 8 mice except 105
where otherwise specified. All animal experiments were conducted in accordance with 106
institutional guidelines with the approval of the Animal Care and Use Committee of the 107
University of Tokyo. 108
109
Cell lines and tumor models. B16F10 and LLC were obtained from the American Type Culture 110
Collection. Colon 26 was obtained from the Cell Resource Center for Biomedical Research, 111
Institute of Development, Aging, and Cancer, Tohoku University. B16F10 cells expressing the 112
truncated form of human low-affinity nerve growth factor receptor (∆hLNGFR/hCD271) were 113
generated by retroviral transduction and 2 subsequent rounds of in vivo passaging 114
(Supplementary Fig. S1). B16F10 cells (5 × 105 per mouse), LLC cells (5 × 105 per mouse) and 115
Colon 26 cells (2 × 105 per mouse) were inoculated s.c. into the right flanks of C57BL/6 or 116
BALB/c mice. Tumor diameter was measured twice weekly and used to calculate tumor volume 117
(mm3) [(major axis; mm) × (minor axis; mm) 2 × 0.5236]. 118
119
In vivo antibody treatment. Anti-CD4 (clone GK1.5), anti-CD8 (clone YTS169.4), anti-PD-1 120
(clone J43), anti-PD-L1 (clone 10F.9G2), anti-PD-L2 (clone TY25), anti-OX40 (clone OX-86), 121
anti-CTLA-4 (clone 9D9), anti-LAG-3 (clone C9B7W), anti-BTLA (clone 6A6), anti-TIM-3 122
(clone RMT3-23), anti-GITR (clone DTA-1) and anti-CD25 (clone PC-61.5.3) mAbs were 123
purchased from BioXcell. Antibodies were injected intraperitoneally at a dose of 200 μg per 124
mouse. Anti-CD4 mAb (200 μg/mouse) was administered in a single dose or in successive doses 125
on days 5 and 9 after tumor inoculation. Immune checkpoint antibodies (200 μg/mouse) were 126
administered on days 4, 8, 14 and 18 after tumor-inoculation. Combination treatments with the 127
anti-CD4 mAb and anti-immune checkpoint antibodies were administered under the same 128
conditions as respective single agent protocols. 129
130
Immunohistological analysis. Immunofluorescent staining was performed as described 131
previously (14-16) using primary antibodies and the appropriate fluorophore-conjugated 132
secondary Abs as listed in Table S1, then photographed using a SP5 confocal microscope (Leica 133
Microsystems). 134
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
6
135
Flow cytometry. Intravascular leukocytes were stained by intravenous injection of 136
fluorophore-conjugated mAb (3 μg/mouse) against CD45 or CD45.2 3 min prior to collecting 137
tissues. Single-cell suspensions were prepared by enzymatic or mechanical dissociation of 138
tissues with or without subsequent density separation, as described previously (17, 18). 139
Flow-Count fluorospheres (Beckman Coulter) were used to determine cell numbers and 140
normalize cell concentrations prior to antibody staining. Cells were pretreated with Fc Block 141
(anti-mouse CD16/CD32 mAb; clone 2.4G2, BioXcell), then stained with mix of 142
fluorophore-conjugated anti-mouse mAbs as indicated in Table S1. Data were acquired on a 143
Gallios flow cytometer (Beckman Coulter) and analyzed using FlowJo software (version 9.7.5; 144
FlowJo, LLC). Non-viable cells were excluded from the analysis based on forward and side 145
scatter profiles and propidium iodide staining. 146
147
Quantitative reverse transcription real-time polymerase chain reaction. Total RNA was 148
extracted using a RNeasy Mini kit (Qiagen) and converted to cDNA using ReverTra Ace qPCR 149
RT Master Mix with gDNA Remover (Toyobo) according to the manufacturer's instructions. 150
Real-time quantitative PCR analysis was performed using THUNDERBIRD Probe qPCR Mix 151
or THUNDERBIRD SYBR qPCR Mix (Toyobo) and an ABI 7500 sequence detector system 152
(Life Technologies). The primers used for the PCR reaction are listed in Table S2. The 153
expression levels of each gene were normalized to Rps3 expression level for each sample. 154
155
Statistics. Unless otherwise stated, data are presented as mean ± SE. Statistical analyses were 156
performed using GraphPad Prism software (version 6.0e, GraphPad Software). For comparisons 157
between groups in the in vivo study we used one-way ANOVA with Dunnett’s post-hoc test. For 158
comparisons between the means of two variables we used paired Student’s t-tests. Comparisons 159
of survival data between groups were made using the log-rank test after Kaplan-Meier analysis. 160
A P-value < 0.05 was considered to be statistically significant. 161
162
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
7
RESULTS 163
An optimized anti-CD4 mAb treatment protocol exerts robust antitumor effects 164
We began by optimizing the protocol for anti-CD4 mAb administration in B16F10, 165
Lewis lung carcinoma (LLC) and Colon 26 tumor models. Mice bearing subcutaneous tumors 166
received a single intraperitoneal injection of 200 μg anti-CD4 mAb 2 days before (day –2) or 0, 167
3, 5 or 9 days after tumor inoculation. In all three models, administration of anti-CD4 mAb on 168
days 3 and 5 significantly suppressed tumor growth (Supplementary Fig. S2A–C). B16F10 169
tumor growth, but not LLC and Colon 26 tumor growth, was also inhibited by mAb 170
administration on days –2 and 0 (Supplementary Fig. S2A). However, the growth of LLC and 171
Colon 26 tumors was not significantly affected by mAb administration at days –2 and 0 172
(Supplementary Fig. S2B and C). Successive administration of the anti-CD4 mAb on days 5 and 173
9 resulted in the greatest inhibition of tumor growth in all three models (data not shown). Doses 174
of anti-CD4 mAb (3.1 or 12.5 μg/mouse) that were insufficient to cause CD4 lymphocyte 175
depletion had no inhibitory effect on tumor growth in the melanoma model (Supplementary Fig. 176
S2D and E). Based on these results, for subsequent studies we adopted a protocol of 177
administering the anti-CD4 mAb at a dose of 200 μg/mouse successively on days 5 and 9 after 178
tumor inoculation. 179
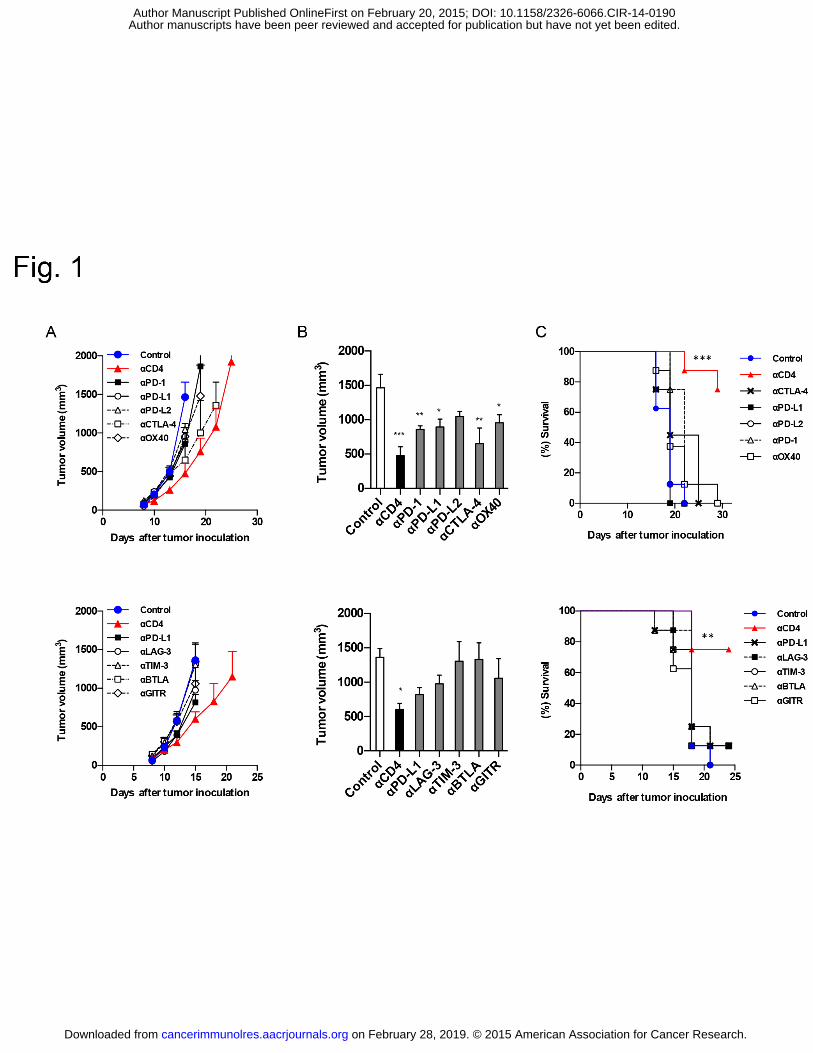
We next compared the antitumor effects of the anti-CD4 mAb against those of a 180
variety of immune checkpoint mAbs (PD-1, PD-L1, PD-L2, CTLA-4, OX40, LAG-3, TIM-3, 181
BTLA and GITR) in the B16F10 model, because melanoma is a major target of anti-immune 182
checkpoint antibody therapy. We found that twice-weekly injections of immune checkpoint 183
antibodies were sufficient to produce the same level of anti-tumor effect as achieved with daily 184
injections (data not shown). Among the mAbs tested, the anti-CD4 mAb was the most effective 185
single-agent treatment in terms of tumor growth inhibition and survival (Fig. 1A–C). 186
Collectively, these results confirm the potent antitumor effects of anti-CD4 mAb treatment in 187
mice and reveal a surprising advantage of anti-CD4 mAb treatment over immune checkpoint 188
mAb treatment. 189
190
Anti-CD4 mAb treatment depletes CD4+ T cells and pDCs 191
To determine which cells are depleted by anti-CD4 mAb therapy, we next examined 192
changes in cell populations with immunosuppressive potential following anti-CD4 mAb 193
administration at day 5 in mice bearing B16F10 tumors. Flow cytometric analysis revealed that 194
numbers of CD4+ T cells including Foxp3+CD25+ Tregs decreased 50- to 100-fold over days 2 195
to 9 following anti-CD4 mAb administration (7 to 14 days after tumor inoculation), as 196
compared to cell numbers in phase-matched untreated tumor-bearing mice (Supplementary Fig. 197
S3A–C). When LLC tumor-bearing mice were administered anti-CD4 mAb on days 5 and 9, 198
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
8
CD4+ T cells disappeared from the blood until at least day 15 after the first mAb administration 199
(Supplementary Fig. S3D). pDCs, a subset of which are positive for CD4 and have been 200
implicated in the suppression of antitumor immune responses (7), also decreased 3- to 10-fold 201
over days 2 to 9 following mAb treatment (Supplementary Fig. S3A–C). MDSC subpopulations, 202
including neutrophils and Ly-6Chi or Ly-6Clo monocytes, were not significantly affected by mAb 203
treatment (data not shown). These results indicate that CD4+ T cells (including Tregs) and pDCs 204
are the targets of anti-CD4 mAb therapy. 205
206
Anti-CD4 mAb treatment increases the number of tumor-infiltrating CD8+ T cells 207
We next investigated the effects of anti-CD4 mAb therapy on tumor-infiltrating CD8+ 208
T-cell populations. Intravascular staining (IVS) is a technique that allows circulating leukocytes 209
present in tissue blood vessels (which represent a proportion of total leukocytes recovered) to be 210
distinguished from cells actually infiltrating the parenchyma of tissues, including tumors (19). 211
In untreated B16F10 tumors, about 15% of CD8+ T cells were positive for IVS, and the 212
frequency of PD-1+CD137+ tumor-reactive cells (20) was about 10-fold lower in this population 213
than in the IVS-negative parenchymal cell population (Supplementary Fig. S4A and B). 214
Anti-CD4 mAb treatment significantly increased the frequency and number of IVS-CD45- CD8+ 215
T cells in the tumor (Fig. 2A and B). The increased number of CD8+ T cells in the tumors of 216
anti-CD4 mAb-treated mice was also evident in histological sections (Fig. 2C). Furthermore, the 217
IVS–CD8+ T cells induced by anti-CD4 mAb treatment contained a higher proportion of 218
PD-1+CD137+ tumor-reactive cells (Fig. 2D and E), had greater potential to produce IFNγ in 219
response to ex vivo PMA/ionomycin stimulation (Fig. 2F and G), and showed higher specific 220
killing activity against B16F10 tumor cells (Supplementary Fig. S5A–C), compared to T cells 221
from the untreated group. In the LLC and Colon 26 tumor models, anti-CD4 mAb-treated mice 222
displayed decreased tumor growth, systemically increased CD8+CD44hiPD-1+ T cells, and 223
upregulation of LAG-3, TIM-3, and CTLA-4 in tumor-infiltrating CD8+ T cells (Supplementary 224
Fig. S6A–D). Collectively, these results suggest that anti-CD4 mAb treatment enhances 225
antitumor CD8+ T-cell responses and induces a shift towards type I immunity within the tumor. 226
227
Anti-CD4 mAb treatment promotes expansion of tumor-specific CD8+ T cells in the 228
draining lymph node 229
To further investigate the effects of anti-CD4 mAb treatment on tumor-specific CD8+ 230
T-cell responses, we adoptively transferred melanoma antigen-specific Pmel-1 TCR transgenic 231
CD8+ T cells (21) into mice 1 day before inoculation with B16F10 tumors (day –1) 232
(Supplementary Fig. S7A and B). On day 14 after tumor inoculation, numbers of Pmel-1 CD8+ 233
T cells in the blood, draining lymph node (dLN), non-dLN (ndLN), spleen and tumor were 10- 234
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
9
to 100-fold higher in anti-CD4 mAb-treated mice compared to that in untreated mice 235
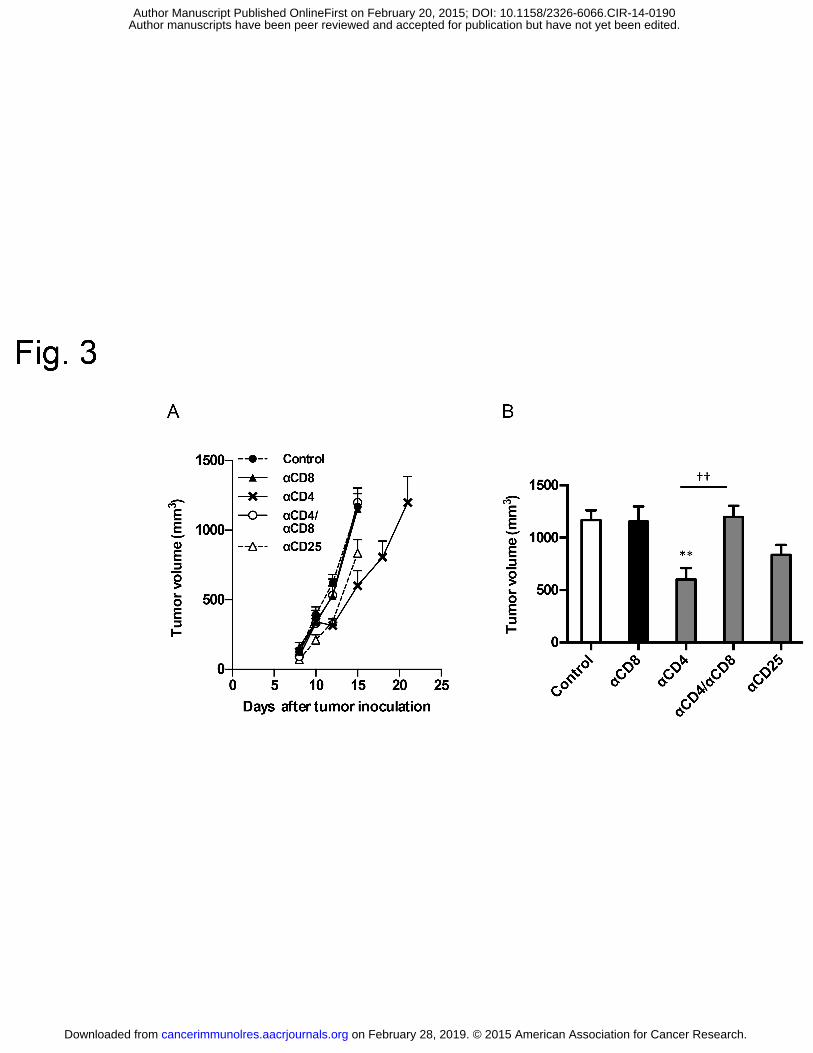
(Supplementary Fig. S7C and D). As tumors grew, Pmel-1 CD8+ T-cell numbers were 236
unchanged or decreased in untreated group mice, whereas Pmel-1 CD8+ T-cell numbers 237
increased in anti-CD4 mAb-treated mice (Supplementary Fig. S7E). To determine the site of 238
Pmel-1 CD8+ T-cell expansion, we administered BrdU one hour prior to collecting tissues. The 239
number of BrdU+ proliferating Pmel-1 CD8+ T cells in the dLN far outnumbered those in the 240
tumor, irrespective of anti-CD4 mAb treatment (Supplementary Fig. S7F and G). Importantly, 241
proliferating cell numbers decreased between days 9 and 14 in untreated mice, but increased in 242
anti-CD4 mAb-treated mice (Supplementary Fig. S7H). Similar CD4-depletion-induced 243
proliferation was also observed in endogenous polyclonal CD8+ T cells (data not shown). These 244
data suggest that anti-CD4 mAb treatment protects tumor-reactive CD8+ T cells from deletion, a 245
mechanism of peripheral tolerance in which the continuous and excessive exposure of 246
antigen-specific T cells to cognate antigens eventually results in the loss of the antigen-specific 247
T-cell clones. 248
To confirm the effects of anti-CD4 mAb treatment on the proliferation of CD8+ T cells, 249
we used fluorescent ubiquitination-based cell-cycle indicator (Fucci) double transgenic mice. In 250
Fucci mice, Fucci-orange (mKO2) and Fucci-green (mAG) are expressed reciprocally in the 251
G0/G1 and S/G2/M phases of the cell cycle, respectively (13, 18). In the B16F10 tumor model, 252
anti-CD4 mAb treatment significantly increased the proportion of mAG+ proliferating cells 253
among CD8+CD44hi T cells in both the dLN and non-dLN, compared to the proportion of these 254
cells in untreated control mice (Supplementary Fig. S7I and J). 255
To determine whether this CD4 depletion-induced proliferation was specific for 256
tumor-specific CD8+ T cells or was a tumor antigen-independent response such as homeostatic 257
proliferation (22), we adoptively transferred a CFSE-labeled mixture of Pmel-1, 258
ovalbumin-specific OT-I and polyclonal CD8+ T cells into B16 tumor-bearing or tumor-free 259
mice with or without anti-CD4 mAb treatment (Supplementary Fig. S8A). Pmel-1 but not OT-I 260
or polyclonal CD8+ T cells selectively proliferated in the dLN of B16 tumor-bearing mice 261
(Supplementary Fig. S8B–E). These results indicate that CD4 depletion-induced T-cell 262
expansion is specific for tumor-specific CD8+ T cells. Collectively, these results suggest that 263
anti-CD4 mAb treatment systemically increases the availability of tumor-specific CD8+ T cells 264
by enhancing their proliferation in the dLN in a tumor-associated antigen-dependent manner. 265
266
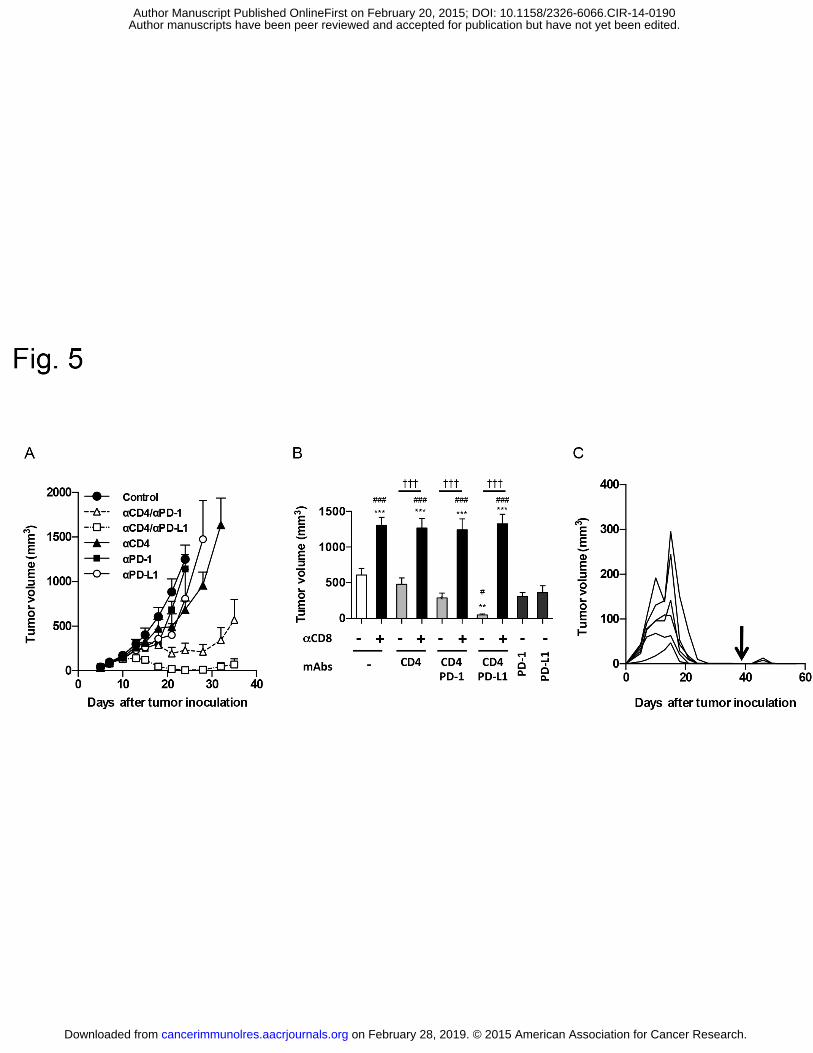
Enhanced CD8+ T-cell responses underlie the antitumor effects of anti-CD4 mAb 267
treatment 268
To determine whether enhanced CTL responses are responsible for the antitumor 269
effects of anti-CD4 mAb treatment, we administered the anti-CD4 mAb together with an 270
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
10
anti-CD8-depleting mAb. When the anti-CD8-depleting mAb was administered together with 271
the anti-CD4 mAb, the inhibitory effect of anti-CD4 mAb treatment on tumor growth was 272
completely reversed (Fig. 3A and B). We also investigated whether treatment with an 273
anti-CD25-depleting mAb, which is widely used to deplete Foxp3+CD25+ Tregs in mice (23), 274
could produce the same effect as anti-CD4 mAb treatment. Under our administration protocol, 275
tumor growth in the anti-CD25 mAb-treated group was almost equivalent to that observed in 276
untreated mice (Fig. 3A and B). These results suggest that the tumor-specific CD8+ T cells that 277
are induced by CD4 mAb treatment are responsible for the antitumor effects of the treatment, 278
and that anti-CD4 mAb treatment might deplete immunosuppressive populations more 279
efficiently than anti-CD25 mAb treatment. 280
281
Combination treatment with anti-CD4 and anti-PD-1 or anti-PD-L1 mAbs synergistically 282
enhances antitumor effects 283
Next, we examined whether synergistic antitumor effects could be achieved by 284
supplementing anti-CD4 mAb treatment with various immune checkpoint mAbs, particularly 285
those targeting the exhaustion and deletion phase of the immune response. We devised a 286
combination treatment protocol of anti-CD4 mAb with immune checkpoint antibodies as 287
depicted in Fig. 4A. Strikingly, combination treatment with anti-CD4 and anti-PD-L1 mAbs, 288
and to a lesser extent anti-CD4 and anti-PD-1 mAbs, resulted in dramatic synergistic inhibition 289
of tumor growth in the B16F10 melanoma model (Fig. 4B and C). Combination treatment with 290
anti-CD4 and anti-CTLA-4, anti-TIM-3, anti-BTLA and anti-GITR mAbs also had additive or 291
synergistic effects (Fig. 4B and C), but anti-PD-L2, anti-OX40 and anti-LAG-3 mAbs produced 292
no synergistic antitumor effect when combined with the anti-CD4 mAb (Fig. 4B and C). 293
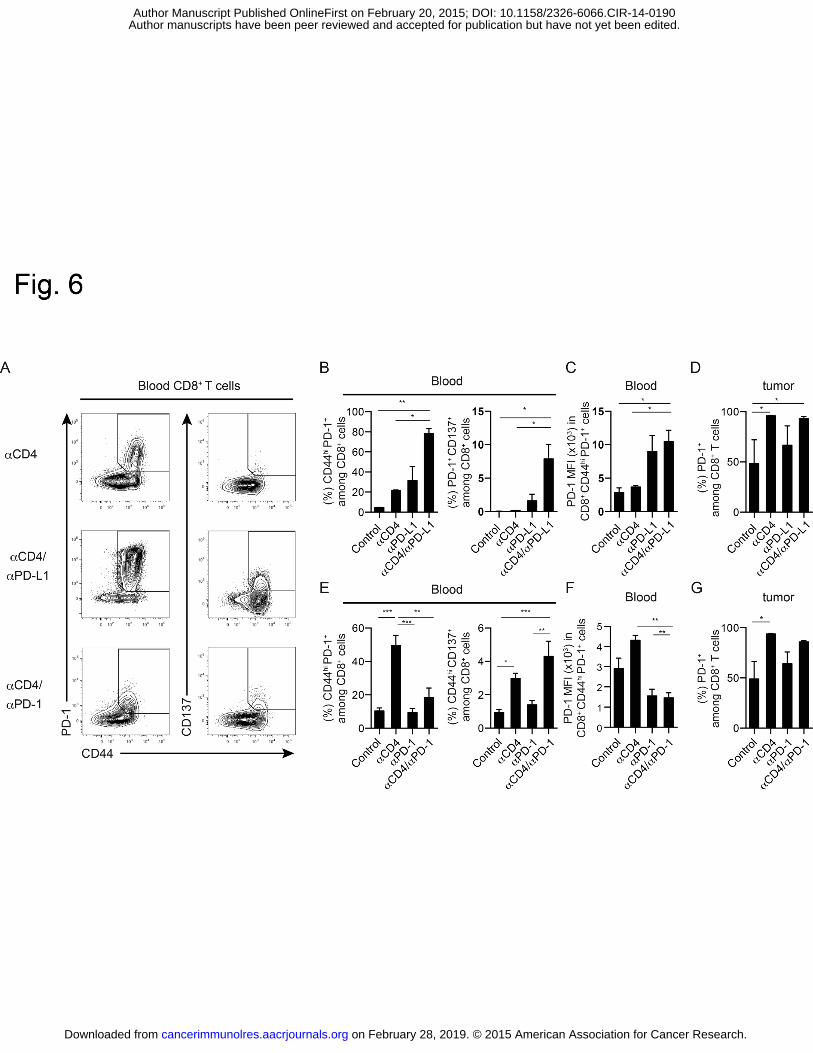
Survival was also prolonged by combination treatment with anti-CD4 and anti-PD-L1 mAbs 294
compared to anti-CD4 mAb monotherapy, but not by other combinations of anti-CD4 and 295
immune checkpoint mAbs (Fig. 4D). Importantly, depletion of CD8+ T cells completely 296
abrogated the tumor growth inhibition induced by the combination of anti-CD4 and anti-PD-1 297
or PD-L1 mAbs, indicating that CD8+ T cells play a critical role in the antitumor effects of the 298
combination treatment (Fig. 4E). 299
To determine whether the synergistic antitumor effects of anti-CD4 and anti-PD-1 or 300
anti-PD-L1 mAb treatment are common to other tumor types and mouse strains, we examined 301
the effect of combination treatment in the Colon 26 subcutaneous tumor model in BALB/c mice. 302
The anti-PD-1 or anti-PD-L1 mAb treatment alone did not inhibit tumor growth, whereas 303
combination treatment with anti-CD4 and anti-PD-1 or anti-PD-L1 mAbs resulted in strong 304
synergistic inhibition of tumor growth (Fig. 5A and B). These effects were completely reversed 305
by treatment with an anti-CD8 depleting mAb (Fig. 5B). Notably, we observed complete 306
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
11
remission in three of ten mice treated with the anti-CD4/anti-PD-1 mAb combination, and in six 307
of ten mice treated with the anti-CD4/anti-PD-L1 mAb combination. In addition, the six mice 308
that rejected the tumor in the anti-CD4/anti-PD-L1 mAb-treated group were resistant to 309
re-challenge with Colon 26 tumor cells at a dose 5 times higher than that used in the initial 310
inoculation (Fig. 5C). Collectively, these results indicate that combination treatment with 311
anti-CD4 and anti-PD-1 or anti-PD-L1 mAbs has a dramatic and robust antitumor effect that is 312
mediated by antitumor CD8+ T cells. 313
314
Blockade of the PD-1/PD-L1 signaling axis increases the number of PD-1+ tumor-reactive 315
CD8+ T cells in the circulation 316
Finally, we investigated the cellular and molecular mechanisms underlying the synergy 317
between anti-CD4 and anti-PD-1 or anti-PD-L1 mAbs in the B16F10 melanoma model. 318
Quantitative RT-PCR analysis of whole tumor tissue demonstrated that anti-CD4 mAb treatment 319
alone augmented expression of the antitumor cytokine genes Ifng and Tnf, the IFNγ-inducible 320
genes Cxcl10 and Cd274/PD-L1 (24, 25), and genes encoding the pro-apoptotic molecules Fasl, 321
Prf1/perforin, and Gzmb/Granzyme B, compared with the expression levels of these genes in 322
untreated tumors (Supplementary Fig. S9A and B). The upregulation of PD-L1 by anti-CD4 323
mAb treatment was also observed at the protein level (Supplementary Fig. S9C). However, no 324
additive or synergistic effects on gene expression were observed in groups receiving 325
combination treatment with anti-CD4 and anti-PD-1 or PD-L1 mAbs. Consistent with these 326
results, the proportion of IFNγ-producing and TNFα-producing cells within the 327
tumor-infiltrating CD8+ T-cell population was equivalent between mice receiving anti-CD4 328
mAb alone and mice receiving the combination of anti-CD4 and anti-PD-1 or anti-PD-L1 mAbs 329
(data not shown). 330
We next analyzed the effects of anti-PD-1 and anti-PD-L1 mAbs on the PD-1+CD8+ T 331
cells that increase in number in the systemic circulation in response to anti-CD4 mAb treatment. 332
We examined cell populations expressing the effector/memory T-cell marker CD44 and the 333
activation marker CD137. Combination treatment with anti-CD4 and anti-PD-L1 mAbs 334
increased the frequency of CD44hiPD-1+ cells amongst CD8+ T cells in the blood, dLN and 335
non-dLN, compared to that in mice receiving the anti-CD4 mAb alone (blood data shown in Fig. 336
6A and B). In blood CD8+ T cells, expression levels of PD-1 on cells within the CD44hiPD-1+ 337
population and the frequency of PD-1+CD137+ cells were significantly higher in mice that 338
received the combination of anti-CD4 and anti-PD-L1 mAbs compared to the corresponding 339
expression levels and frequency in mice that received the anti-CD4 mAb alone (Fig. 6A–C). In 340
contrast, combination treatment with anti-CD4 and anti-PD-1 mAbs decreased the frequency of 341
the CD44hiPD-1+ population among blood CD8+ T cells, and decreased the expression levels of 342
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
12
PD-1 on cells within the CD44hi PD-1+ population (Fig. 6A, E and F). However, the frequency 343
of the CD44hiCD137+ tumor-reactive cell population was higher in mice receiving the 344
combination of anti-CD4 and anti-PD-1 mAbs compared to mice receiving the anti-CD4 mAb 345
alone (Fig. 6A, E and F), suggesting that anti-PD-1 mAb treatment does not actually decrease 346
the number of tumor-reactive CD8+ T cells in the blood, but rather decreases the level of PD-1 347
expression on these cells. On the other hand, the frequency of PD-1+ cells among 348
tumor-infiltrating CD8+ T cells in anti-CD4 mAb-treated mice was not affected by treatment 349
with anti-PD-1 or anti-PD-L1 mAbs (Fig. 6D and G). 350
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
13
DISCUSSION 351
The recent success of anti-CTLA-4 and anti-PD-1 mAb therapies in the clinic has 352
highlighted the potential of immunotherapy for the treatment of cancer (2, 3, 26-29). However, 353
the development of immunotherapy for widespread clinical use remains in its early stages. 354
Extensive efforts have been directed toward enhancing endogenous antitumor immunity by 355
dampening the influence of immunosuppressive mechanisms. Treatment strategies have 356
included combinations of antibodies with other antibodies and with other immunotherapies or 357
anti-cancer therapeutics. In the present study, we demonstrate that antibody-mediated depletion 358
of CD4+ cells from tumor-bearing mice results in enhanced polyclonal PD-1+CD137+ 359
tumor-reactive and monoclonal tumor-specific Pmel-1 CD8+ T-cell responses, and strong 360
inhibition of tumor growth. Combination treatment with the anti-CD4 mAb and various immune 361
checkpoint mAbs, particularly anti-PD-1 and anti-PD-L1 mAbs, revealed striking synergy in 362
suppressing tumor growth and prolonging survival. 363
Several previous reports have described antitumor activity of anti-CD4 mAb treatment 364
in solid tumor models in C57BL/6 mice, including subcutaneous tumors induced by inoculation 365
with B16 melanoma cells (9, 11, 12), recurrent TC1 lung cancer cells (30), or embryo cells 366
expressing the adenovirus-derived E1A protein (10). Although the efficacy of immunotherapy in 367
mouse tumor models often depends on tumor type, taken together, these reports from 368
independent groups and our results from the present study suggest that anti-CD4 mAb treatment 369
is likely to have broad spectrum antitumor activity against solid tumors. Optimization of the 370
anti-CD4 mAb administration protocol revealed robust antitumor effects when mice received 371
the mAb on days 3 or 5, rather than when mice receive the mAb prior to tumor inoculation (day 372
–2). These results suggest that pretreatment is not necessary. However, priming and/or the 373
pre-existence of activated CD8+ T cells are important for effective anti-CD4 mAb therapy. 374
Although the mechanistic link between the timing of anti-CD4 antibody administration and the 375
efficacy of treatment remains to be elucidated, administration of the antibody to patients with 376
early-stage cancer or whose tumor burden has been reduced by surgical resection, irradiation or 377
chemotherapeutics is likely to be most beneficial. 378
A dose of anti-CD4 mAb sufficient to deplete most CD4+ cells was required in order 379
for antitumor effects to be observed. The CD4+ cell population includes Foxp3+ CD25+ Tregs, 380
Th2 cells, Tr1/3 cells (4) and IDO+ immunosuppressive pDCs (7). Considering that markedly 381
increased proliferation of tumor-specific CD8+ T cells was observed in the dLN, anti-CD4 mAb 382
treatment is likely to augment proliferation of tumor-reactive CD8+ T cells through the removal 383
of these CD4+ immunosuppressive cells from the dLN. In addition, anti-CD4 mAb treatment 384
increased the proportion of PD-1+CD137+ tumor-reactive cells and IFNγ-producing cells among 385
tumor-infiltrating CD8+ T cells in the B16F10 model, suggesting that anti-CD4 mAb treatment 386
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
14
augmented both the quantity and quality of tumor-specific CD8+ T-cell responses. We recently 387
demonstrated that IFNγ- and TNFα-induced cell-cycle arrest is an important mechanism 388
underlying the antitumor effects induced by tumor-specific CD8+ T-cell transfer (31). The shift 389
towards IFNγ-dominant type I immunity, which was evident in the strong induction of IFNγ and 390
TNFα in tumor-infiltrating CD8+ T cells after anti-CD4 mAb treatment, is likely to play a 391
central role in the antitumor effects that we observed (32). Notably, depletion of CD25+ Tregs 392
by administration of an anti-CD25 mAb on days 5 and 9 post tumor inoculation did not 393
reproduce the antitumor effect of anti-CD4 mAb treatment. Because some Foxp3+ Tregs have 394
low-to-negative CD25 expression, residual Foxp3+ CD25–/lo Tregs may have contributed to this 395
discrepancy. Moreover, the antitumor effects of anti-CD25 mAb treatment have been reported to 396
be optimal when the mAb is administered prior to tumor inoculation (33, 34), because when 397
administered after tumor inoculation the anti-CD25 mAb depletes not only Tregs but also other 398
activated lymphocytes expressing CD25. The involvement of Treg and other CD4+ 399
immunosuppressive populations in the suppression of CD8+ T cell-mediated antitumor 400
responses remains to be elucidated. 401
The synergy that occurs in combination treatment with anti-CD4 and anti-PD-1 or 402
anti-PD-L1 mAbs is likely due to the blockade of PD-1/PD-L1 signaling in PD-1+ activated 403
CD8+ T cells that are induced by anti-CD4 mAb treatment. We did not detect any synergistic 404
effect in terms of the quantity and quality of the tumor-infiltrating CD8+ T-cell response 405
promoted by anti-CD4 and anti-PD-1 or anti-PD-L1 mAb treatment. However, the frequency of 406
the PD-1+CD137+ and CD44hiCD137+ tumor-reactive populations increased among CD8+ T 407
cells in the blood upon blockade of the PD-1/PD-L1 signaling axis. Considering that T cells 408
continuously traffic between peripheral and secondary lymphoid tissues via the lymph-blood 409
circulation, the blockade of PD-1/PD-L1 signaling may prevent exhaustion or deletion of 410
tumor-reactive PD-1+CD8+ T cells in the tumor and allow them to migrate into the dLN, thus 411
sustaining antitumor CD8+ T-cell responses. In addition, anti-CD4 mAb treatment increased the 412
number of IFNγ-producing PD-1+ CD8+ T cells in the tumor, resulting in the upregulation of 413
IFNγ-inducible genes including PD-L1. Although the shift towards IFNγ-dominant type-1 414
immunity within the tumor contributes to the inhibition of tumor growth, it also promotes the 415
exhaustion or deletion of tumor-infiltrating PD-1+CD8+ T cells by enhancing PD-1/PD-L1 416
signaling. It is therefore likely that the synergy of the anti-CD4 and anti-PD-1 or anti-PD-L1 417
mAb combination treatment arises due to the blockade of this adverse negative feedback 418
mechanism. 419
We are in the process of developing a humanized anti-CD4 mAb with potent 420
antibody-dependent cell-mediated cytotoxicity (ADCC) as an anti-cancer therapeutic. Because 421
CD4+ T cells play important roles in both humoral and cellular immunity, the heightened risk of 422
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
15
infectious diseases that may be associated with transient CD4+ T-cell depletion should be 423
carefully evaluated in clinical trials. In addition, trials should seek to maximize clinical efficacy 424
and safety through rigorous optimization of the antibody administration protocol. In pre-clinical 425
studies in nonhuman primates, no serious adverse effects were detected after several weeks of 426
treatment with our humanized anti-human CD4 mAb that resulted in CD4+ T-cell depletion. In 427
addition, no severe adverse effects have been observed during phase-II clinical trials for T-cell 428
malignancy with long-term administration of other humanized anti-CD4 mAbs (35, 36). 429
Preexisting humoral immune mediators, such as immunoglobulin, plasma cells and memory B 430
cells, CD8+ T-cell responses, and unimpaired natural immunity are likely to provide basal 431
protection against infectious diseases during CD4+ T cell-depleting therapies. On the other hand, 432
consideration should also be given to the potential for the acute exacerbation of chronic diseases 433
associated with viral infection (e.g. hepatitis C and B) due to excessive activation of effector 434
and memory CD8+ T cells after CD4+ cell depletion. 435
In conclusion, our study represents the first report of robust antitumor effects of 436
combination treatment with an anti-CD4-depleting antibody and anti-PD-1 or anti-PD-L1 437
immune checkpoint antibodies in mice. We have also characterized the immunologic bases for 438
the synergy between these agents. Recent clinical trials suggest that anti-PD-1, anti-PD-L1, or 439
anti-CTLA-4 mAbs, or combinations of these agents, are not effective against all types of solid 440
tumors. Our findings suggest that combination treatment with an anti-CD4 mAb and immune 441
checkpoint mAbs, particularly anti-PD-1 or anti-PD-L1 mAbs, is likely to result in greater 442
clinical efficacy against a broader ranges of cancers. 443
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
16
ACKNOWLEDGMENTS 444
We thank A. Miyawaki, A. Sakaue-Sawano and the RIKEN BioResource Center for providing 445
FucciG1 and FucciS/G2/M mice; A. Hosoi for assistance with Pmel-1-B16F10 experiments; H. 446
Yamazaki, K. Tsuji, and K. Yoshioka for animal care; A. Yamashita, S. Aoki and S. Fujita for 447
expert technical assistance; and M. Otsuji, K. Takeda and S. Shibayama for helpful discussions. 448
449
FOOTNOTES 450
Author contributions: S.Y., S.U., Y.I., S.I. and K.M. designed research; S.Y., S.U., Y.I., H.O., 451
K.C., T.N., K.H., Y.T., E.T., A.H. performed research; S.Y., S.U., Y.I., H.O., K.C., S.S., K.K., S.I. 452
and K.M. analyzed data; S.Y., S.U., Y.I., K.C., F.H.W.S., K.K., S.I. and K.M. wrote the paper. 453
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
17
REFERENCES 454
1. Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat 455
Rev Cancer. 2012;12:252-64. 456
2. Topalian SL, Weiner GJ, Pardoll DM. Cancer immunotherapy comes of age. J Clin 457
Oncol. 2011;29:4828-36. 458
3. Wolchok JD, Kluger H, Callahan MK, Postow MA, Rizvi NA, Lesokhin AM, et al. 459
Nivolumab plus ipilimumab in advanced melanoma. N Engl J Med. 2013;369:122-33. 460
4. Whiteside TL. Disarming suppressor cells to improve immunotherapy. Cancer 461
Immunol Immunother. 2012;61:283-8. 462
5. Alizadeh D, Larmonier N. Chemotherapeutic targeting of cancer-induced 463
immunosuppressive cells. Cancer Res. 2014;74:2663-8. 464
6. Camisaschi C, De Filippo A, Beretta V, Vergani B, Villa A, Vergani E, et al. 465
Alternative activation of human plasmacytoid DCs in vitro and in melanoma lesions: 466
involvement of LAG-3. J Invest Dermatol. 2014;134:1893-902. 467
7. Matta BM, Castellaneta A, Thomson AW. Tolerogenic plasmacytoid DC. European 468
J Immunol. 2010;40:2667-76. 469
8. Nagai H, Hara I, Horikawa T, Fujii M, Kurimoto M, Kamidono S, et al. Antitumor 470
effects on mouse melanoma elicited by local secretion of interleukin-12 and their 471
enhancement by treatment with interleukin-18. Cancer Invest. 2000;18:206-13. 472
9. Nagai H, Hara I, Horikawa T, Oka M, Kamidono S, Ichihashi M. Elimination of 473
CD4(+) T cells enhances anti-tumor effect of locally secreted interleukin-12 on B16 mouse 474
melanoma and induces vitiligo-like coat color alteration. J Invest Dermatol. 475
2000;115:1059-64. 476
10. den Boer AT, van Mierlo GJ, Fransen MF, Melief CJ, Offringa R, Toes RE. CD4+ T 477
cells are able to promote tumor growth through inhibition of tumor-specific CD8+ T-cell 478
responses in tumor-bearing hosts. Cancer Res. 2005;65:6984-9. 479
11. Yu P, Lee Y, Liu W, Krausz T, Chong A, Schreiber H, et al. Intratumor depletion of 480
CD4+ cells unmasks tumor immunogenicity leading to the rejection of late-stage tumors. 481
The Journal of experimental medicine. 2005;201:779-91. 482
12. Choi BK, Kim YH, Kang WJ, Lee SK, Kim KH, Shin SM, et al. Mechanisms 483
involved in synergistic anticancer immunity of anti-4-1BB and anti-CD4 therapy. Cancer 484
Res. 2007;67:8891-9. 485
13. Sakaue-Sawano A, Kurokawa H, Morimura T, Hanyu A, Hama H, Osawa H, et al. 486
Visualizing spatiotemporal dynamics of multicellular cell-cycle progression. Cell. 487
2008;132:487-98. 488
14. Ueha S, Yoneyama H, Hontsu S, Kurachi M, Kitabatake M, Abe J, et al. CCR7 489
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
18
mediates the migration of Foxp3+ regulatory T cells to the paracortical areas of peripheral 490
lymph nodes through high endothelial venules. J Leukoc Biol. 2007;82:1230-8. 491
15. Ueha S, Murai M, Yoneyama H, Kitabatake M, Imai T, Shimaoka T, et al. 492
Intervention of MAdCAM-1 or fractalkine alleviates graft-versus-host reaction associated 493
intestinal injury while preserving graft-versus-tumor effects. J Leukoc Biol. 2007;81:176-85. 494
16. Shono Y, Ueha S, Wang Y, Abe J, Kurachi M, Matsuno Y, et al. Bone marrow 495
graft-versus-host disease: early destruction of hematopoietic niche after MHC-mismatched 496
hematopoietic stem cell transplantation. Blood. 2010;115:5401-11. 497
17. Sawanobori Y, Ueha S, Kurachi M, Shimaoka T, Talmadge JE, Abe J, et al. 498
Chemokine-mediated rapid turnover of myeloid-derived suppressor cells in tumor-bearing 499
mice. Blood. 2008;111:5457-66. 500
18. Shand FH, Ueha S, Otsuji M, Koid SS, Shichino S, Tsukui T, et al. Tracking of 501
intertissue migration reveals the origins of tumor-infiltrating monocytes. ProcNatl Acad Sci 502
U S A. 2014;111:7771-6. 503
19. Anderson KG, Mayer-Barber K, Sung H, Beura L, James BR, Taylor JJ, et al. 504
Intravascular staining for discrimination of vascular and tissue leukocytes. Nat Protoc. 505
2014;9:209-22. 506
20. Ye Q, Song DG, Poussin M, Yamamoto T, Best A, Li C, et al. CD137 accurately 507
identifies and enriches for naturally occurring tumor-reactive T cells in tumor. Clin Cancer 508
Res. 2014;20:44-55. 509
21. Overwijk WW, Theoret MR, Finkelstein SE, Surman DR, de Jong LA, Vyth-Dreese 510
FA, et al. Tumor regression and autoimmunity after reversal of a functionally tolerant state 511
of self-reactive CD8+ T cells. J Exp Med. 2003;198:569-80. 512
22. Surh CD, Sprent J. Homeostasis of naive and memory T cells. Immunity. 513
2008;29:848-62. 514
23. Sakaguchi S. Regulatory T cells: history and perspective. Methods Mol Biol. 515
2011;707:3-17. 516
24. Dong H, Strome SE, Salomao DR, Tamura H, Hirano F, Flies DB, et al. 517
Tumor-associated B7-H1 promotes T-cell apoptosis: a potential mechanism of immune 518
evasion. Nature Med. 2002;8:793-800. 519
25. Furuta J, Inozume T, Harada K, Shimada S. CD271 on melanoma cell is an 520
IFN-gamma-inducible immunosuppressive factor that mediates downregulation of 521
melanoma antigens. J Invest Dermatol. 2014;134:1369-77. 522
26. Couzin-Frankel J. Breakthrough of the year 2013. Cancer immunotherapy. Science. 523
2013;342:1432-3. 524
27. Hamid O, Robert C, Daud A, Hodi FS, Hwu WJ, Kefford R, et al. Safety and tumor 525
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
19
responses with lambrolizumab (anti-PD-1) in melanoma. The New England journal of 526
medicine. 2013;369:134-44. 527
28. Topalian SL, Drake CG, Pardoll DM. Targeting the PD-1/B7-H1(PD-L1) pathway to 528
activate anti-tumor immunity. Curr Opin Immunol. 2012;24:207-12. 529
29. Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermott DF, et al. 530
Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N Engl J Med. 531
2012;366:2443-54. 532
30. Predina J, Eruslanov E, Judy B, Kapoor V, Cheng G, Wang LC, et al. Changes in 533
the local tumor microenvironment in recurrent cancers may explain the failure of vaccines 534
after surgery. Proc Natl Acad Sci U S A. 2013;110:E415-24. 535
31. Matsushita H, Hosoi A, Ueha S, Abe J, Fujieda N, Tomura M, et al. Cytotoxic T 536
lymphocytes block tumor growth both by lytic activity and IFN-gamma-dependent cell cycle 537
arrest. Cancer Immunol Res. 2015;3:26-36. 538
32. Braumuller H, Wieder T, Brenner E, Assmann S, Hahn M, Alkhaled M, et al. 539
T-helper-1-cell cytokines drive cancer into senescence. Nature. 2013;494:361-5. 540
33. Onizuka S, Tawara I, Shimizu J, Sakaguchi S, Fujita T, Nakayama E. Tumor 541
rejection by in vivo administration of anti-CD25 (interleukin-2 receptor alpha) monoclonal 542
antibody. Cancer Res. 1999;59:3128-33. 543
34. Shimizu J, Yamazaki S, Sakaguchi S. Induction of tumor immunity by removing 544
CD25+CD4+ T cells: a common basis between tumor immunity and autoimmunity. J 545
Immunol. 1999;163:5211-8. 546
35. Kim YH, Duvic M, Obitz E, Gniadecki R, Iversen L, Osterborg A, et al. Clinical 547
efficacy of zanolimumab (HuMax-CD4): two phase 2 studies in refractory cutaneous T-cell 548
lymphoma. Blood. 2007;109:4655-62. 549
36. Rider DA, Havenith CE, de Ridder R, Schuurman J, Favre C, Cooper JC, et al. A 550
human CD4 monoclonal antibody for the treatment of T-cell lymphoma combines inhibition 551
of T-cell signaling by a dual mechanism with potent Fc-dependent effector activity. Cancer 552
Res. 2007;67:9945-53. 553
554
555
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
20
FIGURE LEGENDS 556
Figure 1. Antitumor effects of anti-CD4 mAb treatment. 557
Mice bearing B16F10 melanoma tumors were injected intraperitoneally with anti-CD4 mAb 558
(200 μg/mouse) on days 5 and 9 or anti-immune checkpoint mAbs on days 4, 8, 14 and 18 after 559
tumor inoculation. (A) Tumor growth curves. (B) Tumor volume on day 16 (upper panel) or day 560
15 (lower panel). (C) Survival following tumor inoculation (8 mice per group). (A, B) Data 561
represent mean ± SE of 8 mice per group. *, P < 0.05; **, P < 0.01; ***, P < 0.001 (compared to 562
control). 563
564
Figure 2. Anti-CD4 mAb treatment increases the number of tumor-infiltrating CD8+ T 565
cells. 566
Mice bearing B16F10 (A, B, D–G) or B16F10-∆hLNGFR (C) tumors were injected 567
intraperitoneally with anti-CD4 mAb on days 5 and 9, and tumor-infiltrating CD8+ T cells were 568
analyzed on day 14 after tumor inoculation. Control mice received an injection of vehicle only. 569
For flow cytometric analyses, mice were given an intravenous injection of anti-CD45.2 Ab 3 570
min prior to the collection of tissues to enable identification of cells in the blood compartment 571
(intravascular staining, IVS). (A) Flow cytometry plots of parenchymal leukocyte compartments 572
(CD45+ IVS-CD45.2–). (B) The number of parenchymal CD8+ T cells in tumor. (C) Distribution 573
of CD8+ T cells in the tumor. Green, CD8; red, ∆hLNGFR; blue, propidium iodide (PI). 574
Enlargements in white boxes show non-necrotic areas, yellow box shows necrotic area. Scale 575
bar represents 200 μm. (D) Flow cytometry plots and (E) frequencies of PD-1+ CD137+ 576
tumor-reactive cells among the parenchymal CD8+ T-cell population. (F) Flow cytometry plots 577
and (G) frequencies of IFNγ- and TNFα-producing cells among the parenchymal CD8+ T-cell 578
population following ex vivo re-stimulation with PMA and ionomycin. Data represent mean ± 579
SE of 4 mice per group and are representative of at least four independent experiments. 580
Numbers in flow cytometry plots indicate mean frequencies within live cells (A) or parental 581
populations (D and F). ***, P < 0.001 (compared to control). 582
583
Figure 3. CD8+ T cells play a pivotal role in the antitumor effects of anti-CD4 mAb 584
treatment. 585
Mice bearing B16F10 tumors were injected intraperitoneally with anti-CD4, anti-CD8 and/or 586
anti-CD25 mAbs (200 μg/mouse) on days 5 and 9 after tumor inoculation. (A) Tumor growth 587
curves. (B) Tumor volume on day 15 after tumor inoculation. Data represent mean ± SE of 8 588
mice per group. **, P < 0.05 (compared to control); ††, P < 0.01 (comparison as indicated). 589
590
Figure 4. Combination treatment with anti-CD4 and anti-PD-1 or anti-PD-L1 mAbs has 591
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Anti-CD4 and immune checkpoint antibody synergy
21
synergistic antitumor effects. 592
Mice bearing B16F10 tumors received anti-CD4 mAb, anti-immune checkpoint mAb, or a 593
combination of these, according to the treatment schedule shown in (A). (B) Tumor volume on 594
day 16 (left) or 15 (right). *, P < 0.05; **, P < 0.01; ***, P < 0.001 (compared to control); #, 595
P=0.021 (compared to αCD4); ††, P < 0.01; †††, P < 0.001 (comparisons as indicated). (C) 596
Tumor growth curves. (D) Survival plots representative of two independent experiments. *, P < 597
0.05; **, P < 0.01 ***, P < 0.001 ****, P < 0.0001 (compared to control); †, P < 0.05; ††, P < 598
0.01; †††, P < 0.001 (compared to αCD4). (E) Anti-CD8 mAb was administered together with 599
anti-CD4 mAb and tumor volumes were measured on day 16. **, P < 0.01 (compared to 600
control). Data represent mean ± SE of 8 mice per group. In the text as (data not shown) but 601
should insert into Figure 4. 602
603
Figure 5. Combination treatment with anti-CD4 and anti-PD-1 or anti-PD-L1 mAbs 604
induces long-term antitumor CD8+ T-cell memory. 605
Mice bearing Colon 26 tumors received anti-CD4, anti-PD-L1, anti-PD-1 or anti-CD8 mAbs or 606
a combination of these according to the treatment schedule shown in Fig. 4A. (A) Tumor growth 607
curves. (B) Tumor volume on day 18. **, P < 0.01; ***, P < 0.001 (compared to control); #, 608
P=0.029; ###, P < 0.001 (compared to αCD4); †††, P < 0.001 (comparisons as indicated). (C) 609
The six mice that achieved complete remission of Colon 26 tumors after anti-CD4 and 610
anti-PD-L1 treatment were re-challenged on day 39 with Colon 26 tumor cells at five-times the 611
cell number of the initial challenge. Arrow indicates day of re-challenge. *, P < 0.05; **, P < 612
0.01 (compared to control). (A and B) Data represent mean ± SE of 10 mice per group. 613
614
Figure 6. Anti-PD-L1 and anti-PD-1 treatments target PD-1+ CD8+ T cells that are induced 615
by anti-CD4 treatment. 616
Mice bearing B16F10 tumors were treated with anti-CD4, anti-PD-L1 or anti-PD-1 mAbs, or a 617
combination of these according to the treatment schedule shown in Fig. 4A. (A) Flow cytometry 618
plots of blood CD8+ T cells. (B and E) Proportions of CD44hi PD-1+ cells, PD-1+ CD137+ cells 619
or CD44hi CD137+ cells among blood CD8+ T cells on day 14. (C and F) Mean fluorescent 620
intensity (MFI) of PD-1 expression on CD8+ CD44hi PD-1+ cells in the blood. (D and G) 621
Proportions of PD-1+ cells among tumor-infiltrating CD8+ T cells. (B–D) show anti-PD-L1 mAb 622
experiments; (E–G) show anti-PD-1 mAb experiments. Data represent mean ± SE of 4 mice per 623
group and are representative of two independent experiments. *, P < 0.05; **, P < 0.01; ***, P < 624
0.001. 625
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190

Published OnlineFirst February 20, 2015.Cancer Immunol Res Satoshi Ueha, Shoji Yokochi, Yoshiro Ishiwata, et al. treatment in miceantibody and anti-PD-1/PD-L1 immune checkpoint antibody Robust anti-tumor effects of combined anti-CD4 depleting
Updated version
10.1158/2326-6066.CIR-14-0190doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerimmunolres.aacrjournals.org/content/suppl/2015/02/20/2326-6066.CIR-14-0190.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerimmunolres.aacrjournals.org/content/early/2015/02/20/2326-6066.CIR-14-0190To request permission to re-use all or part of this article, use this link
on February 28, 2019. © 2015 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 20, 2015; DOI: 10.1158/2326-6066.CIR-14-0190





























