Embed Size (px)
Citation preview

メルク バイオインサイトNo.62
特集:大腸菌でタンパク質を可溶性に発現させるツールについて
特集:大腸菌でタンパク質を可溶性に発現させるツールについて
可溶性に発現させる 3 つの工夫
大腸菌でタンパク質を発現させるとき,大腸菌の菌体内では動物細胞などと大きく異なり,極めて還元環境となっているため,S-S 結合などを必要とするタンパク質の正しいフォールディングが起こらないことが多く見られます.このため,封入体(インクルージョンボディー)になってしまったり,活性のないタンパク質になるといった問題が生じます.
大腸菌で,正しくフォールディングさせ可溶性を向上させる工夫には主に3つの方法があります.
1. 大腸菌内で可溶性の高いタンパク質と融合タンパク 質として発現させる.(NusAなど)
2. 還元環境を改善させるタンパク質と融合させて発現 させる.(Trx,GST など)
3. 分泌させて,中性環境に近いペリプラズム(大腸菌 内外膜間の空間)に発現させる.(シグナル配列, Dsbなど)
Novagen®では,これら 3つの工夫を実現するベクターとホストを持っています.今回の特集では,タンパク質の構造解析など多くの実績を持つ種々 pET ベクターと,菌体内部の還元環境を改善した株Origami™ シリーズなどのホストについてご紹介します.
NusA タンパク質は 1999 年に報告された 495 アミノ酸(55 kDa,SDS-PAGE では 69 k 位置)からなる転写終結関連因子として見出されたタンパク質で(Davis 1999 年),2000 年に Novagen® により pET NusA Fusion System( Nus•Tag)として初めて製品化されました.このシステムは NusA タンパク質の可溶性促進作用,すなわち,大腸菌内で単独発現させると不溶性だった標的タンパク質のアミノ末端に NusA タンパク質を融合させると可溶性が促進される報告にもとづいています. 販売開始以来 Nus•Tag は様々なハイスループット発現スクリーニング系で採用されています.プロテアーゼによってNus•Tag を切断除去した後に,活性を保持したタンパク質を得ることに成功したという報告が多数あり,さらに Nus•Tagと融合した状態で活性を保持するタンパク質の報告もあります. Nus•Tag は現在,大腸菌発現系で広く使用されている融合タグの 1 つです(Cabrita 2006 年).Nus•Tag による可溶性促進機構についても研究されています.
Nus•Tag 融合タンパク質の可溶性評価大腸菌組み換えタンパク質発現系にての Nus•Tag 融合タンパク質の可溶性を検討した論文報告から,幅広い分子量の異種標的タンパク質を多数用いた3つの実験結果を 表1 (p.3参照)にまとめました.Shih らが実施した実験(2002 年)の注目すべき点は,様々な生物種の標的タンパク質を用いた点です.Korf らが実施した実験(2005 年)において,Nus•Tag の可溶性は,30℃で誘導した場合よりも 20℃で誘導した場合で高い結果が得られています.さらに,8 種類の分子量の大きい
(70 kDa を超える)タンパク質を様々な融合タグに結合させ,可溶性について比較した結果として,発現誘導された可溶性タンパク質は,Nus•Tag を用いた場合は 7 種類であったのに対し,他の融合タグ 3 種類(GST,MBP,6 × His)を用いた場合は 4 種類だったとの報告もあります.Korf らによる他の所見として,通常は真核細胞の細胞内小器官,細胞膜,細胞骨格に存在するタンパク質の場合,他の融合タグと比較してNus•Tag を選択した場合もっとも可溶性が向上すると述べています.また,Kohl ら(2008 年)によると,タンパク質発
Nus•Tag システムの魅力ミニレビュー
Roger G. Harrison – University of Oklahoma

現を 20 ~ 25℃で誘導する場合,膜タンパク質などの可溶化が困難なタンパク質の可溶性促進にとって Nus•Tag は有用であると述べています.Korf らの所見と同じく,Kohl らは,30℃または 37℃でタンパク質発現を誘導した場合と比較して,25℃でタンパク質発現を誘導した場合に Nus•Tag 融合タンパク質の収量が増える可能性を示唆しています.
Nus•Tagを切断除去しても活性を保持するタンパク質,正しい折り畳み構造を維持するタンパク質Nus•Tag 融合タンパク質として発現させ,Nus•Tag を切断除去した後も活性を保持するタンパク質,もしくは正しい折り畳み構造を維持する標的タンパク質に関するいくつかの研究データを 表 2(p.3 参照)にまとめました.実験には主に分子量 20 kDa 以下の標的タンパク質を用いました.標的タンパク質の収量は,培養液1 L あたり 1.5 ~ 100 mg の範囲で,ケモカインおよびサイトカインに関しては,30 ~ 100 mg/L という高い収量が得られています.また,各タンパク質の発現および精製に関して,以下の通り,興味深い追加情報があります.
植物由来 9ホスホエノールピルビン酸 - カルボン酸キナーゼ(Ermolova2003年)
BDA(ブルーデキストラン・アガロース)色素アフィニティカラムを用い,タグ切断後の標的タンパク質をさらに精製しました.タグ切断前と比較して,タグ切断後の標的タンパク質では 50 倍高い触媒効率が得られました.
Xklp3a,Tep3Ag,E8R(DeMarco2004年)
タグ切断後,His タグ融合 TEV プロテアーゼ及び His タグ融合Nus•Tag を Ni2+ カラムを用いて,選択的に除去しました.アフィニティタグと樹脂を強く結合させたまま,標的タンパク質を未結合画分に回収しました.標的タンパク質 3 種類は,いずれも精製後に正しく折りたたまれていました.細胞膜結合 E8R ワクシニアウイルスタンパク質を精製し,Nus•Tag を除去したところ,Tris 緩衝液中で不溶性になり沈殿しました.しかし,0.02% ドデシルマルトシドおよび 150 mmol/L 塩化ナトリウムを添加することにより,Nus•Tag 除去後も E8R タンパク質は可溶性になりました.
シクロマルトデキストリナーゼ(Turner2005年)
シクロマルトデキストリナーゼはα - アミラーゼタンパク質ファミリーのひとつです.このファミリーに属するタンパク質を大腸菌内で活性型として発現させることが困難な事が知られています.シクロマルトデキストリナーゼを Nus•Tag 融合タンパク質として発現させ,エンテロキナーゼ(製品番号 69066-3)を用いて Nus•Tag を切断した結果,Nus•Tag を切断していない融合タンパク質と比較して 2 倍の活性レベルに達しました.この結果から,タンパク質融合によって酵素活性が減少することがわかりました.
ヒト由来ケモカイン(Magistrelli2005年)
すべての標的タンパク質を Novagen® Origami™ B(p.9 参照)株内で発現させ,ケモカインをコードする配列の C 末端に AviTag™(アビディティー)ビオチン化部位の配列を導入しました.単量体アビジン樹脂を用いたアフィニティクロマトグラフィによって精製後,Nus•Tag を切断,精製しました.Nus•Tag 融合タンパク質はいずれも活性を示しませんでしたが,タグ切断,精製したタンパク質は活性を示しました.
ヘムエリトリン(Karlsen2005年)
Nus•Tag を切断した後,ヘムエリトリンをゲルろ過クロマトグラフィーによってさらに精製しました.精製ヘムエリトリンを円偏光
二色性によって測定した結果α - ヘリックス構造含量は,モデリングによって予測したα - ヘリックス構造含量とほぼ一致しました.また精製タンパク質は,単量体で安定でした.
ヒト由来 IL-29(Li2006年)
Ni2+ カ ラ ム 用 い て His•Tag ア フ ィ ニ テ ィ ー 精 製 と 比 較 し て,S•Tag アフィニティー精製を用いた方が,高純度の融合タンパク質が得られました.アミノ末端 NusA/His•Tag®/S•Tag™ 融合タンパク質をビオチン化トロンビン(製品番号 69672-3)を用いて切断した後,ストレプトアビジン・アガロース(製品番号 69203-3)を用いて残存ビオチン化トロンビンを除去しました.ヒト不死化羊膜上皮細胞系(WISH 細胞)に感染させた疱性口内炎ウイルス
(VSV)に対して精製 IL-29 の抗ウイルス活性が観察されました.
ヒト由来 IFN- λ 2(Li2007年)
Nus•Tag を組換え型エンテロキナーゼ(製品番号 69066-3)で切断した後,残存エンテロキナーゼを Novagen® EKapture™ Agarose
(製品番号 69068)を用いて除去しました.ヒト不死化羊膜上皮細胞系(WISH 細胞)への疱性口内炎ウイルス(VSV)感染 24 時間前に添加した場合,精製 IFN- λ 2 は,ウイルス誘発変性作用からの WISH 細胞保護能を示しました.
活性を保持した Nus•Tag 融合タンパク質Nus•Tag 融合タンパク質から Nus•Tag を切断,除去し,活性型タンパク質を精製している報告が多数ありますが,Nus•Tag と融合した状態でも標的タンパク質が活性を保持しているとの報告も多数あります.そのような Nus•Tag 融合タンパク質として,
ScFv 触媒抗体 14D9(Zheng 2003 年),オワンクラゲ Aequorea victoria 由来の緑色蛍光タンパク質(Nallamsetty 2006 年),ヒト由来のジヒドロ葉酸還元酵素(Nallamsetty 2006 年),マテガイ
Ensis directus 由来のアルギナーゼキナーゼ(Compaan 2003 年),B. thuringiensis 由来の修飾δエンドトキシン(Kumar 2005 年),ヒト由来の BCMA 膜貫通型受容体(Guan 2006 年),植物由来のα- ジオキシゲナーゼ 1(Liu 2006 年),および熱帯熱マラリア原虫
Plasmodium falciparum 由来のβ - ケトアシル - アシル担体タンパク質合成酵素(Lack 2006 年)などです.
Nus•Tag の可溶性促進機構NusA タンパク質は,in vivo でシャペロニン GroEL の必須基質となることがわかっています(Houry 1999 年).GroEL(および補因子 GroES)は,あらゆる大腸菌増殖条件下で必須となる唯一のシャペロン系です.Douette ら(2005 年)は,可溶型 Nus•Tag-UCP1(非共役タンパク質 1:ミトコンドリア膜タンパク質)融合タンパク質の発現量について検討しました.その結果,16℃でGroEL の共過剰発現に反応し,可溶性が増加しました.本結果は,NusA タンパク質がパートナータンパク質の凝集を抑制し,シャペロン経路と相互作用することを示唆しています.
まとめNus•Tag は単独発現時には不溶性である様々な標的タンパク質に対し,大腸菌内で可溶性を促進することが示されています.様々なプロテアーゼを用いて Nus•Tag を切断除去した後も活性を保持するタンパク質,もしくは正しい折り畳み構造を維持する標的タンパク質の例が多数報告されています.Nus•Tag と結合した状態で活性を保持する標的タンパク質の例も多数報告されています.NusA タンパク質の優れた可溶性促進作用は,大腸菌シャペロン経路との相互作用に起因する可能性が考えられます.
ミニレビュー Nus•Tagシステムの魅力
2
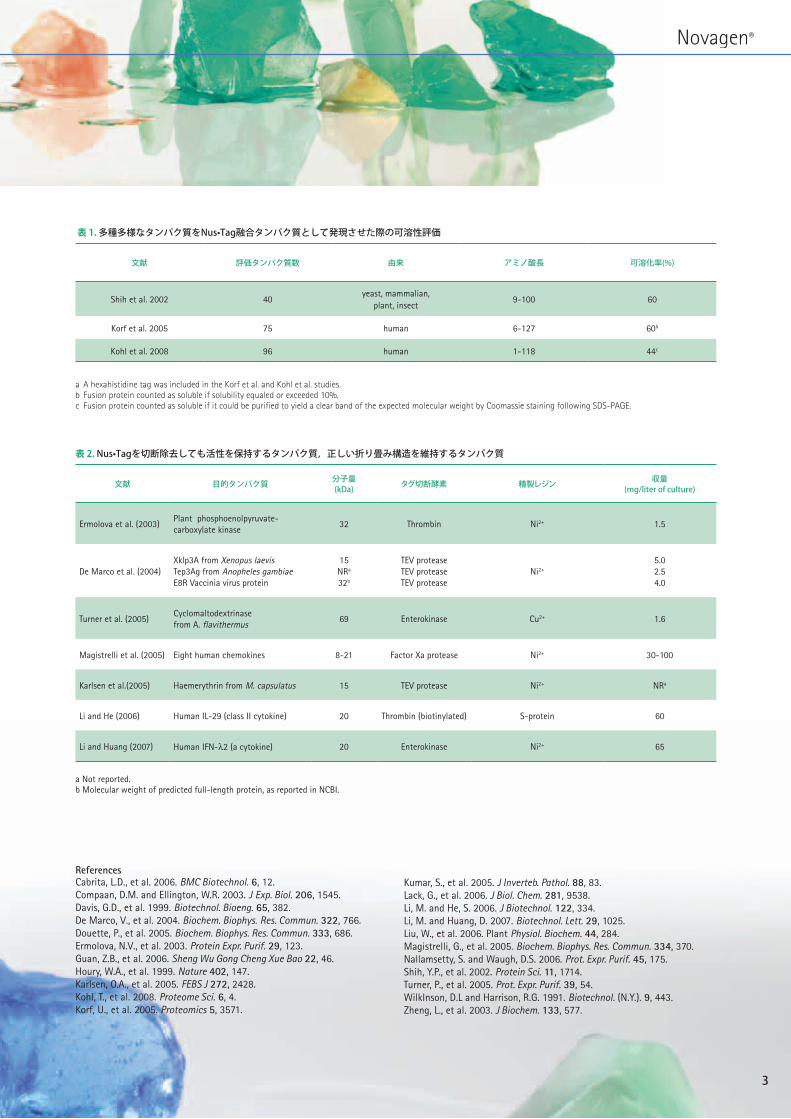
表 1.多種多様なタンパク質をNus•Tag融合タンパク質として発現させた際の可溶性評価
文献 評価タンパク質数 由来 アミノ酸長 可溶化率(%)
Shih et al. 2002 40yeast, mammalian,
plant, insect 9-100 60
Korf et al. 2005 75 human 6-127 60b
Kohl et al. 2008 96 human 1-118 44c
a A hexahistidine tag was included in the Korf et al. and Kohl et al. studies. b Fusion protein counted as soluble if solubility equaled or exceeded 10%.c Fusion protein counted as soluble if it could be purified to yield a clear band of the expected molecular weight by Coomassie staining following SDS-PAGE.
表 2.Nus•Tagを切断除去しても活性を保持するタンパク質,正しい折り畳み構造を維持するタンパク質
文献 目的タンパク質 分子量(kDa) タグ切断酵素 精製レジン 収量
(mg/literofculture)
Ermolova et al. (2003)Plant phosphoenolpyruvate-carboxylate kinase
32 Thrombin Ni2+ 1.5
De Marco et al. (2004)Xklp3A from Xenopus laevisTep3Ag from Anopheles gambiaeE8R Vaccinia virus protein
15NRa
32b
TEV proteaseTEV proteaseTEV protease
Ni2+
5.02.54.0
Turner et al. (2005)Cyclomaltodextrinase from A. flavithermus
69 Enterokinase Cu2+ 1.6
Magistrelli et al. (2005) Eight human chemokines 8-21 Factor Xa protease Ni2+ 30-100
Karlsen et al.(2005) Haemerythrin from M. capsulatus 15 TEV protease Ni2+ NRa
Li and He (2006) Human IL-29 (class II cytokine) 20 Thrombin (biotinylated) S-protein 60
Li and Huang (2007) Human IFN-λ2 (a cytokine) 20 Enterokinase Ni2+ 65
a Not reported.b Molecular weight of predicted full-length protein, as reported in NCBI.
ReferencesCabrita, L.D., et al. 2006. BMC Biotechnol. 6, 12.Compaan, D.M. and Ellington, W.R. 2003. J Exp. Biol. 206, 1545.Davis, G.D., et al. 1999. Biotechnol. Bioeng. 65, 382.De Marco, V., et al. 2004. Biochem. Biophys. Res. Commun. 322, 766.Douette, P., et al. 2005. Biochem. Biophys. Res. Commun. 333, 686.Ermolova, N.V., et al. 2003. Protein Expr. Purif. 29, 123.Guan, Z.B., et al. 2006. Sheng Wu Gong Cheng Xue Bao 22, 46.Houry, W.A., et al. 1999. Nature 402, 147.Karlsen, O.A., et al. 2005. FEBS J 272, 2428.Kohl, T., et al. 2008. Proteome Sci. 6, 4.Korf, U., et al. 2005. Proteomics 5, 3571.
Kumar, S., et al. 2005. J Inverteb. Pathol. 88, 83.Lack, G., et al. 2006. J Biol. Chem. 281, 9538.Li, M. and He, S. 2006. J Biotechnol. 122, 334.Li, M. and Huang, D. 2007. Biotechnol. Lett. 29, 1025.Liu, W., et al. 2006. Plant Physiol. Biochem. 44, 284.Magistrelli, G., et al. 2005. Biochem. Biophys. Res. Commun. 334, 370.Nallamsetty, S. and Waugh, D.S. 2006. Prot. Expr. Purif. 45, 175.Shih, Y.P., et al. 2002. Protein Sci. 11, 1714.Turner, P., et al. 2005. Prot. Expr. Purif. 39, 54.WilkInson, D.L and Harrison, R.G. 1991. Biotechnol. (N.Y.). 9, 443.Zheng, L., et al. 2003. J Biochem. 133, 577.
3
発現ベクターからのアプローチ
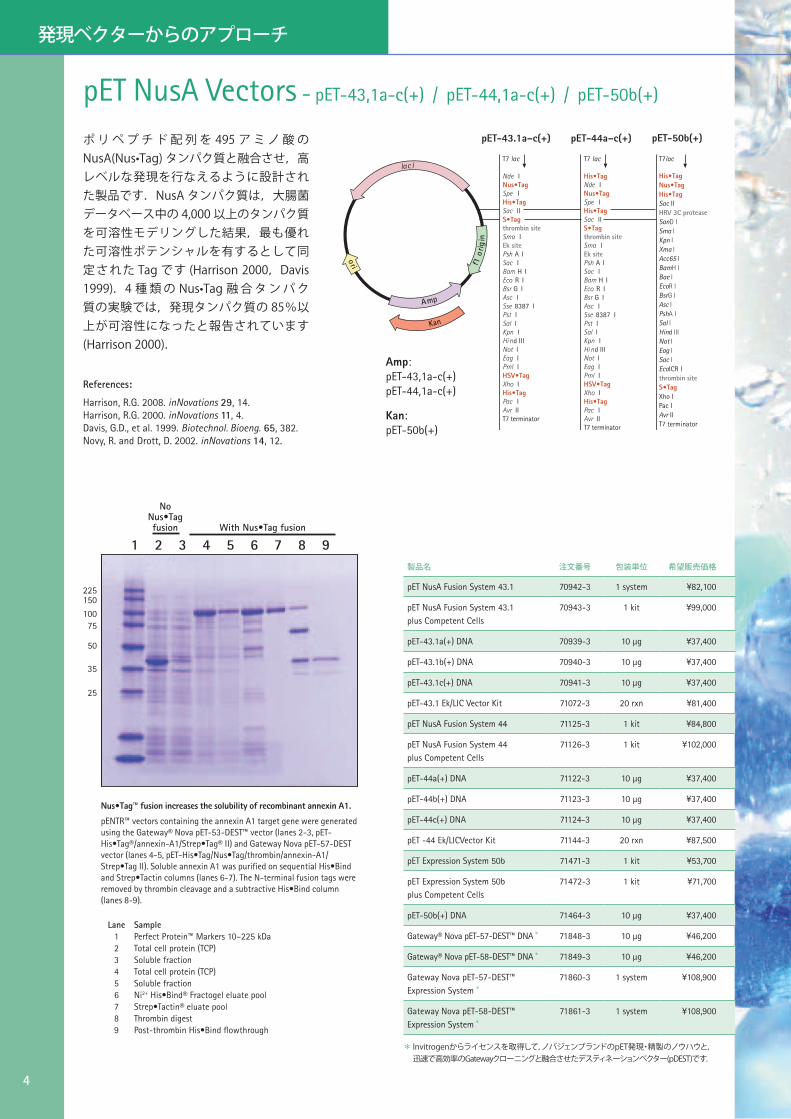
pET NusA Vectors - pET-43,1a-c(+) / pET-44,1a-c(+) / pET-50b(+)
His•Tag
ポ リ ペ プ チ ド 配 列 を 495 ア ミ ノ 酸 のNusA(Nus•Tag) タンパク質と融合させ,高レベルな発現を行なえるように設計された製品です.NusA タンパク質は,大腸菌データベース中の 4,000 以上のタンパク質を可溶性モデリングした結果,最も優れた可溶性ポテンシャルを有するとして同定された Tag です (Harrison 2000,Davis 1999).4 種 類 の Nus•Tag 融 合 タ ン パ ク質の実験では,発現タンパク質の 85%以上が可溶性になったと報告されています(Harrison 2000).
製品名 注文番号 包装単位 希望販売価格
pET NusA Fusion System 43.1 70942-3 1 system ¥82,100
pET NusA Fusion System 43.1plus Competent Cells
70943-3 1 kit ¥99,000
pET-43.1a(+) DNA 70939-3 10 µg ¥37,400
pET-43.1b(+) DNA 70940-3 10 µg ¥37,400
pET-43.1c(+) DNA 70941-3 10 µg ¥37,400
pET-43.1 Ek/LIC Vector Kit 71072-3 20 rxn ¥81,400
pET NusA Fusion System 44 71125-3 1 kit ¥84,800
pET NusA Fusion System 44plus Competent Cells
71126-3 1 kit ¥102,000
pET-44a(+) DNA 71122-3 10 µg ¥37,400
pET-44b(+) DNA 71123-3 10 µg ¥37,400
pET-44c(+) DNA 71124-3 10 µg ¥37,400
pET -44 Ek/LICVector Kit 71144-3 20 rxn ¥87,500
pET Expression System 50b 71471-3 1 kit ¥53,700
pET Expression System 50bplus Competent Cells
71472-3 1 kit ¥71,700
pET-50b(+) DNA 71464-3 10 µg ¥37,400
Gateway® Nova pET-57-DEST™ DNA * 71848-3 10 µg ¥46,200
Gateway® Nova pET-58-DEST™ DNA * 71849-3 10 µg ¥46,200
Gateway Nova pET-57-DEST™Expression System *
71860-3 1 system ¥108,900
Gateway Nova pET-58-DEST™Expression System *
71861-3 1 system ¥108,900
References:
Harrison, R.G. 2008. inNovations 29, 14.Harrison, R.G. 2000. inNovations 11, 4.Davis, G.D., et al. 1999. Biotechnol. Bioeng. 65, 382.Novy, R. and Drott, D. 2002. inNovations 14, 12.
Amp:pET-43,1a-c(+) pET-44,1a-c(+)
Kan:pET-50b(+)
* Invitrogenからライセンスを取得して,ノバジェンブランドのpET発現・精製のノウハウと, 迅速で高効率のGatewayクローニングと融合させたデスティネーションベクター(pDEST)です.
4
Lane Sample1 Perfect Protein™ Markers 10–225 kDa2 Total cell protein (TCP)3 Soluble fraction4 Total cell protein (TCP)5 Soluble fraction6 Ni2+ His•Bind® Fractogel eluate pool7 Strep•Tactin® eluate pool8 Thrombin digest9 Post-thrombin His•Bind flowthrough
25
35
50
75100
150225
With Nus•Tag fusion
No Nus•Tag fusion
Nus•Tag™ fusion increases the solubility of recombinant annexin A1.
pENTR™ vectors containing the annexin A1 target gene were generated using the Gateway® Nova pET-53-DEST™ vector (lanes 2-3, pET-His•Tag®/annexin-A1/Strep•Tag® II) and Gateway Nova pET-57-DEST vector (lanes 4-5, pET-His•Tag/Nus•Tag/thrombin/annexin-A1/Strep•Tag II). Soluble annexin A1 was purified on sequential His•Bind and Strep•Tactin columns (lanes 6-7). The N-terminal fusion tags were removed by thrombin cleavage and a subtractive His•Bind column (lanes 8-9).
1 2 3 4 5 6 7 8 9
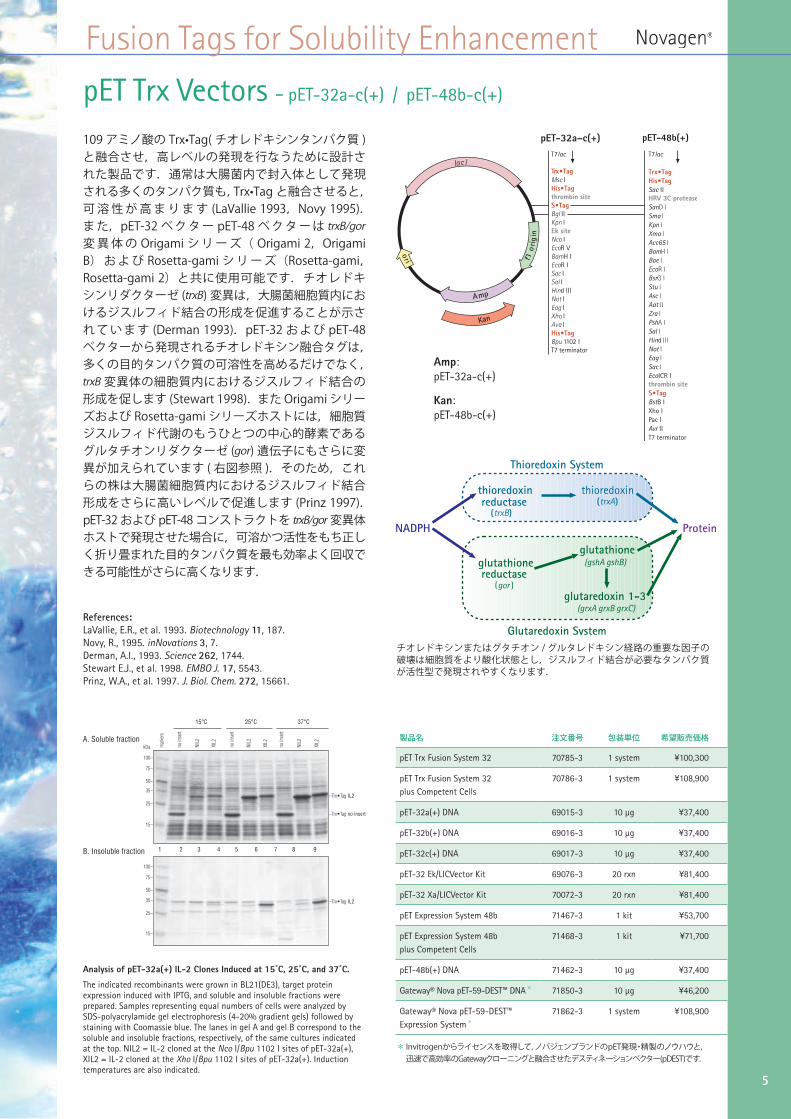
pET Trx Vectors - pET-32a-c(+) / pET-48b-c(+)
109 アミノ酸の Trx•Tag( チオレドキシンタンパク質 )と融合させ,高レベルの発現を行なうために設計された製品です.通常は大腸菌内で封入体として発現される多くのタンパク質も,Trx•Tag と融合させると,可 溶 性 が 高 ま り ま す (LaVallie 1993,Novy 1995).ま た,pET-32 ベ ク タ ー pET-48 ベ ク タ ー は trxB/gor変 異 体 の Origami シ リ ー ズ( Origami 2,Origami B) お よ び Rosetta-gami シ リ ー ズ(Rosetta-gami,Rosetta-gami 2)と共に使用可能です.チオレドキシンリダクターゼ (trxB) 変異は,大腸菌細胞質内におけるジスルフィド結合の形成を促進することが示され て い ま す (Derman 1993).pET-32 お よ び pET-48ベクターから発現されるチオレドキシン融合タグは,多くの目的タンパク質の可溶性を高めるだけでなく,trxB 変異体の細胞質内におけるジスルフィド結合の形成を促します (Stewart 1998).また Origami シリーズおよび Rosetta-gami シリーズホストには,細胞質ジスルフィド代謝のもうひとつの中心的酵素であるグルタチオンリダクターゼ (gor) 遺伝子にもさらに変異が加えられています ( 右図参照 ).そのため,これらの株は大腸菌細胞質内におけるジスルフィド結合形成をさらに高いレベルで促進します (Prinz 1997).pET-32 および pET-48 コンストラクトを trxB/gor 変異体ホストで発現させた場合に,可溶かつ活性をもち正しく折り畳まれた目的タンパク質を最も効率よく回収できる可能性がさらに高くなります.
製品名 注文番号 包装単位 希望販売価格
pET Trx Fusion System 32 70785-3 1 system ¥100,300
pET Trx Fusion System 32plus Competent Cells
70786-3 1 system ¥108,900
pET-32a(+) DNA 69015-3 10 µg ¥37,400
pET-32b(+) DNA 69016-3 10 µg ¥37,400
pET-32c(+) DNA 69017-3 10 µg ¥37,400
pET-32 Ek/LICVector Kit 69076-3 20 rxn ¥81,400
pET-32 Xa/LICVector Kit 70072-3 20 rxn ¥81,400
pET Expression System 48b 71467-3 1 kit ¥53,700
pET Expression System 48bplus Competent Cells
71468-3 1 kit ¥71,700
pET-48b(+) DNA 71462-3 10 µg ¥37,400
Gateway® Nova pET-59-DEST™ DNA * 71850-3 10 µg ¥46,200
Gateway® Nova pET-59-DEST™Expression System *
71862-3 1 system ¥108,900
A. Soluble fraction
B. Insoluble fraction
100–
75–
50–
35–
25–
15–
kDa
100–
75–
50–
35–
25–
15–
–Trx•Tag IL2
–Trx•Tag no insert
–Trx•Tag IL2
no in
sert
NIL2
XIL2
15°C 25°C 37°C
no in
sert
NIL2
XIL2
no in
sert
NIL2
XIL2
1 2 3 4 5 6 7 8 9
mar
kers
Analysis of pET-32a(+) IL-2 Clones Induced at 15˚C, 25˚C, and 37˚C.
The indicated recombinants were grown in BL21(DE3), target protein expression induced with IPTG, and soluble and insoluble fractions were prepared. Samples representing equal numbers of cells were analyzed by SDS-polyacrylamide gel electrophoresis (4-20% gradient gels) followed by staining with Coomassie blue. The lanes in gel A and gel B correspond to the soluble and insoluble fractions, respectively, of the same cultures indicated at the top. NIL2 = IL-2 cloned at the Nco I/Bpu 1102 I sites of pET-32a(+), XIL2 = IL-2 cloned at the Xho I/Bpu 1102 I sites of pET-32a(+). Induction temperatures are also indicated.
References:LaVallie, E.R., et al. 1993. Biotechnology 11, 187.Novy, R., 1995. inNovations 3, 7.Derman, A.I., 1993. Science 262, 1744.Stewart E.J., et al. 1998. EMBO J. 17, 5543.Prinz, W.A., et al. 1997. J. Biol. Chem. 272, 15661.
Amp:pET-32a-c(+)
Kan:pET-48b-c(+)
Fusion Tags for Solubility Enhancement
チオレドキシンまたはグタチオン / グルタレドキシン経路の重要な因子の破壊は細胞質をより酸化状態とし,ジスルフィド結合が必要なタンパク質が活性型で発現されやすくなります.
* Invitrogenからライセンスを取得して,ノバジェンブランドのpET発現・精製のノウハウと, 迅速で高効率のGatewayクローニングと融合させたデスティネーションベクター(pDEST)です.
5

pET GST Vectors - pET-41a-c(+) / pET-42a-c(+) / pET-49b(+)
融合タグとしてグルタチオン -S- トランスフェラーゼ (GST; GST•Tag) が組込まれています.この GST•Tagは,その融合タンパク質の合成を強化し,また場合によっては,可溶性も高めることが報告されています (Smith 1988).可溶性で正しく折り畳まれて発現した GST•Tag 融合タンパク質は,固定したグルタチオンを用いて精製可能です.そして,還元グルタチオンを含むバッファーを用いて,穏やかな溶出を行なえます.また,可溶な GST 融合タンパク質の定量は,トランスフェラーゼ活性の測定によって行なえます.他の市販の GST 融合ベクターとは対照的に,Factor Xa(IleGluGlyArg,pET-42 シリーズ ) およびエンテロキナーゼ (AspAspAspAspLys,pET-41 シリーズ ) のタンパク質切断部位は,ベクターに由来する配列を 100%完全に除去し,もともとの N 末端残基をもつ目的タンパク質を生成できるように設計されています.
発現ベクターからのアプローチ
References:Smith, D.B. and Johnson, K.S. 1988. Gene 67, 31.
pET Dsb Vectors - pET-39b(+) / pET-40b(+)
ペプチド配列に 208 アミノ酸の DsbA•Tag[pET-39b(+)]または 236 アミノ酸の DsbC•Tag[pET-40b(+)] を融合させ,高レベルな発現を行なうために設計された製品です.DsbA と DsbC はそれぞれ,ジスルフィド結合の形成と異性化を触媒するペリプラズム酵素です(Rietsch 1996,Sone 1997,Missiakas 1994,Zapun 1995,Raina 1997).細胞質中での可溶性を高めるよう設計された pET-32 Trx•Tag シリーズとは異なり,この DsbA•Tag および DsbC•Tag ベクターはペリプラズムへの局在化を促進します.ペリプラズムの非還元環境中では可溶性が向上し,適切な折り畳みが促進されます (Collins-Racie 1995).
References:Rietsch, A., et al. 1996. Proc. Natl. Acad. Sci. USA 93, 13048.Sone, M., et al. 1997. J. Biol. Chem. 272, 10349.Missiakas, D., et al. 1994. EMBO J. 13, 2013.Zapun, A., et al. 1995. Biochemistry 34, 5075.Raina, S. and Missiakas, D. 1997. Ann. Rev. Microbiol. 51, 179.Collins-Racie, L.A., et al. 1995. Biotechnology 13, 982.
製品名 注文番号 包装単位 希望販売価格
pET GST Fusion System 41 70559-3 1 kit ¥82,100
pET GST Fusion System 41plus Competent Cells
70560-3 1 kit ¥99,000
pET-41a(+) DNA 70556-3 10 µg ¥37,400
pET-41b(+) DNA 70557-3 10 µg ¥37,400
pET-41c(+) DNA 70558-3 10 µg ¥37,400
pET-41 Ek/LICVector Kit 71071-3 20 rxn ¥81,400
pET GST Fusion System 42 70564-3 1 kit ¥82,100
pET GST Fusion System 42plus Competent Cells
70565-3 1 kit ¥99,000
pET-42a(+) DNA 70561-3 10 µg ¥37,400
製品名 注文番号 包装単位 希望販売価格
pET-42b(+) DNA 70562-3 10 µg ¥37,400
pET-42c(+) DNA 70563-3 10 µg ¥37,400
pET Expression System 49b 71469-3 1 kit ¥53,700
pET Expression System 49bplus Competent Cells
71470-3 1 kit ¥71,700
pET-49b(+) DNA 71463-3 10 µg ¥37,400
Gateway® Nova pET-60-DEST™ DNA
71851-3 10 µg ¥46,200
Gateway® Nova pET-60-DEST™Expression System
71863-3 1 system ¥108,900
製品名 注文番号 包装単位 希望販売価格
pET Dsb Fusion System 39b 70789-3 1 system ¥49,100
pET Dsb Fusion System 39bplus Competent Cells
70790-3 1 system ¥65,300
pET-39b(+) DNA 70090-3 10 µg ¥37,400
pET Dsb Fusion System 40b 70791-3 1 system ¥49,100
pET Dsb Fusion System 40bplus Competent Cells
70792-3 1 system ¥65,300
pET-40b(+) DNA 70091-3 10 µg ¥37,4006

pET Signal Sequence Vectors - pET-20b(+) / pET-22b(+) / pET-25b(+) / pET-26b(+) / pET-27b(+)
目的タンパク質にペリプラズムへの輸送を促進するシグナル配列(ペリラリズムへ移行させるための pelB シグナルシークエンス)を融合するよう設計されたベクターです.ペリプラズムは非還元的環境でフォールディングに適した環境であり,ジスルフィド結合の形成を促進することから,ある種の目的タンパク質では可溶性と活性が高まります.通常,シグナル配列は輸送に伴って,シグナルペプチダーゼにより切断されます.
製品名 注文番号 包装単位 希望販売価格
pET Expression System 20b 70761-3 1 system ¥39,200
pET Expression System 20bplus Competent Cells
70762-3 1 system ¥55,700
pET-20b(+) DNA 69739-3 10 µg ¥34,100
pET Expression System 22b 70765-3 1 system ¥39,200
pET Expression System 22bplus Competent Cells
70766-3 1 system ¥55,700
pET-22b(+) DNA 69744-3 10 µg ¥34,100
pET Expression System 25b 70771-3 1 system ¥39,200
pET Expression System 25bplus Competent Cells
70772-3 1 system ¥55,700
製品名 注文番号 包装単位 希望販売価格
pET-25b(+) DNA 69753-3 10 µg ¥34,100
pET Expression System 26b 70773-3 1 system ¥39,200
pET Expression System 26bplus Competent Cells
70774-3 1 system ¥55,700
pET-26b(+) DNA 69862-3 10 µg ¥34,100
pET Expression System 27b 70775-3 1 system ¥39,200
pET Expression System 27bplus Competent Cells
70776-3 1 system ¥55,700
pET-27b(+) DNA 69863-3 10 µg ¥34,100
Fusion Tags for Solubility Enhancement
7

Protein Expression Strains一般的なタンパク質発現
BL21(DE3)BL21
タンパクの不溶化 /不活性の改善
ジスルフィド結合の効率が悪く,正常にフォールドされたタンパクが得られない場合
Origami™ 2Origami 2(DE3)Rosetta-gami™ 2Rosetta-gami 2(DE3)Rosetta-gami BRosetta-gami B(DE3)
高レベルの発現誘導により正常にフォールドされたタンパクが得られない場合
Tuner™Tuner(DE3)Rosetta-gami BRosetta-gami B(DE3)
毒性タンパクの発現
タンパクが全く発現しない / 大腸菌が増殖しない場合
Tuner™Tuner(DE3)NovaBlueNovaBlue(DE3)Any pLysS host
動物由来原料不使用
Veggie™BL21(DE3)Veggie™BL21(DE3)pLysS
完全長のタンパクが得られない
大腸菌固有のコドン使用頻度の問題
Rosetta™ Rosetta(DE3) Rosetta 2Rosetta 2(DE3)Rosetta-gami™ 2Rosetta-gami 2(DE3)Rosetta-gami BRosetta-gami B(DE3)RosettaBlue™RosettaBlue™(DE3)
プラスミドの安定化
インサート内の配列(繰り返し)の影響でプラスミドが不安定になる場合
BLR(DE3)HMS174HMS174(DE3)NovaBlueNovaBlue(DE3)
目的タンパクのラベリング
B834B834(DE3)
trxB/gor ホ ス トを用いての細胞質内でのタンパク質発現効率の改善を試みる
レアコドン 補充株の使用
lacY 欠失ホストを用いての IPTG濃度依存型の誘導制御を試みる
recA 欠失ホストの使用
Origami 2(DE3) Genotype : D(ara-leu)7697 DlacX74 DphoA PvuII phoR araD139 ahpC galE galK rpsL F’[lac+ lacIq pro] (DE3) gor522::Tn10 trxB (StrR, TetR)
Origami 2(DE3)pLysS Genotype : D(ara-leu)7697 DlacX74 DphoA PvuII phoR araD139 ahpC galE galK rpsL F’[lac+ lacIq pro] (DE3) gor522::Tn10 trxB pLysS (CamR, StrR, TetR)
基底発現の抑制
メチオニン 栄養要求変異株
ホストの使用
ホスト選択からのアプローチ
Expression of rRSC-c from a pET-22b construct under different conditions
BL21(DE3) and Origami(DE3) hosts carrying a pET-22b/RSC-c recombinant plasmid were grown to OD600 0.6-1.0 before induction with 1 mM IPTG. After addition of IPTG to the cultures, growth was continued either for 4 h at 37℃ or for 24 h at 20℃. Samples of the total (T), soluble (S), and insoluble (I) protein fractions were prepared and analyzed on 15% SDS-polyacrylamide gels followed by staining with Coomassie blue.
37°C, 4 h
20°C, 24 h
37°C, 4 h
20°C, 24 h
T T S I InductionConditions
– + + + IPTG
A BL21(DE3)
B BL21(DE3)
C Origami(DE3)
D Origami(DE3)
Figure 1. Expression of rRSC-c from a pET-22bconstruct under different conditionsBL21(DE3) and Origami(DE3) hosts carrying a pET-22b/RSC-cwere grown to OD600 0.6–1.0 before induction with 1 mM IPTG.After addition of IPTG to the cultures, growth was continuedeither for 4 h at 37°C or for 24 h at 20°C. Samples of the total(T), soluble (S), and insoluble (I) protein fractions from cellscontaining a pET-22b/RSC-c were prepared and analyzed on15% SDS-PAGE followed by staining with Coomassie blue.
Origami™ 2 Host StrainsK-12 由来でチオレドキシンリダクターゼ (trxB) とグルタチオンリダクターゼ (gor) に変異をもつホストです.2 種の還元酵素変異によって,細胞質をより酸化状態とし,ジスルフィド結合の形成を大幅に強化できます.その結果,必要なタンパク質が正しくおりたたまれた活性型で発現されやすくなります.カナマイシンもしくはアンピシリン耐性ベクターと併用できます.
8

※ Origami 系のホストは pET-32,pET48b との併用で更なる効果が期待できます.これはチオレドキシン融合タグが細胞質中でのジスルフィド結合をさらに助けるためです.また,リダクターゼ変異の影響で培養時間が長くかかり 18-24 時間程度必要です.
製品名 注文番号 包装単位 希望販売価格
Origami™ 2(DE3) Competent Cells 71345-3 0.4 mL ¥18,000
71345-4 1 mL ¥32,700
Origami™ 2(DE3) Singles™ Competent Cells
71408-3 11 rxn ¥21,600
71408-4 22 rxn ¥41,700
Origami™ 2(DE3)pLysS Competent Cells
71346-3 0.4 mL ¥18,000
71346-4 1 mL ¥32,700
Origami™ B (DE3) Competent Cells 70837-3 0.4 mL ¥17,200
70837-4 1 mL ¥31,200
origami™ B (DE3)pLysS Competent Cells
70839-3 0.4 mL ¥17,200
70839-4 1 mL ¥31,200
製品名 注文番号 包装単位 希望販売価格
Rosetta-gami™ 2(DE3) Competent Cells
71351-3 0.4 mL ¥18,000
71351-4 1 mL ¥32,700
Rosetta-gami™ 2(DE3)pLysS Competent Cells
71352-3 0.4 mL ¥18,000
71352-4 1 mL ¥32,700
Rosetta-gami™ B (DE3) Competent Cells
71136-3 0.4 mL ¥18,000
71136-4 1 mL ¥32,700
Rosetta-gami™ B (DE3)pLysS Competent Cells
71137-3 0.4 mL ¥18,000
71137-4 1 mL ¥32,700
Origami™ Technology
Tuner™ (DE3) Strain Competent CellsBL21 の lacZY 欠失変異体です.培養液中,全ての細胞のタンパク質発現レベルを調節できます.Tuner™ の lacY 変異が細胞内に取り込まれる IPTG 量を均一にし,IPTG 濃度依存的な誘導を行えます.IPTG 濃度を調節することで,非常に低いレベルから充分に誘導したレベルまでタンパク質発現を制御できます.低いレベルでの発現は,発現が難しいタンパク質の可溶性や活性を促進することがあります.
製品名 注文番号 包装単位 希望販売価格
Tuner™ (DE3) Competent Cells 70623-3 0.4 mL ¥17,200
70623-4 1 mL ¥31,200
Tuner™ (DE3) pLysS Competent Cells
70624-3 0.4 mL ¥17,200
70624-4 1 mL ¥31,200
Tuner(DE3) Genotype:F– ompT hsdSB(rB– mB
–) gal dcm lacY1 (DE3)Tuner(DE3)pLysS Genotype:F– ompT hsdSB(rB
– mB–) gal dcm lacY1 (DE3) pLysS (CamR)
Origami™ B Host StrainsIPTG 濃度調節することでタンパク質の発現レベルをコントロールできる BL21 の lacY 変異体(Tuner™)に,ジスルフィド結合の形成を助けるチオレドキシンリダクターゼとグルタチオンリダクターゼの変異を加えたホストです.BL21 のlon と ompT 欠損も受け継いでいますので,発現したタンパク質の安定性が上昇します.カナマイシン耐性株なのでカナマイシン耐性ベクターは使用できません.アンピシリン耐性ベクターと併用できます.Origami B(DE3) Genotype:F– ompT hsdSB(rB
– mB–) gal dcm lacY1 ahpC (DE3) gor522::Tn10 trxB (KanR, TetR)
Origami B(DE3)pLysS Genotype:F– ompT hsdSB(rB– mB
–) gal dcm lacY1 ahpC (DE3) gor522::Tn10 trxB pLysS (CamR, KanR, TetR)
Rosetta-gami™ 2 Host Strains大腸菌で発現したタンパク質中に存在するジスルフィド結合の形成を助ける Origami ™ 2( チオレドキシンリダクターゼ変異,グルタチオンリダクターゼ変異 ) に,大腸菌で存在量の少ないコドンに対応する tRNA(AUA, AGG, AGA, CUA, CCC, GGA, CGG) をコードする pRARE2 を導入し,生物種差に関係ないタンパク質発現を可能にしたホストです.recA, endA, lacIq 変異をもつ K-12 由来なので,細胞内でコンストラクトを安定に保持できます.カナマイシンもしくはアンピシリン耐性ベクターと併用できます.Rosetta-gami 2(DE3) Genotype:D(ara-leu)7697 DlacX74 DphoA PvuII phoR araD139 ahpC galE galK rpsL (DE3) F’[lac+ lacIq pro] gor522::Tn10 trxB pRARE2 (CamR, StrR, TetR)Rosetta-gami 2(DE3)pLysS Genotype:D(ara-leu)7697 DlacX74 DphoA PvuII phoR araD139 ahpC galE galK rpsL (DE3) F’[lac+ lacIq pro] gor522::Tn10 trxB pLysSRARE2 (CamR, StrR, TetR)
Rosetta-gami™ B Host StrainsIPTG 濃度調節することでタンパク質の発現レベルをコントロールできる BL21 の lacY 変異体(Tuner™)にジスルフィド結合の形成を助けるチオレドキシンリダクターゼ変異とグルタチオンリダクターゼ変異を加えた上,コドン補充プラスミドである pRARE2 を導入したホストです.Tuner™ , Origami™ , Rosetta™ の全ての特長を兼ね備えたドリームホストです.アンピシリン耐性ベクターと併用できます.Rosetta-gami B(DE3) Genotype:F– ompT hsdSB(rB
– mB–) gal dcm lacY1 ahpC (DE3) gor522::Tn10 trxB pRARE (CamR, KanR, TetR)
Rosetta-gami B(DE3)pLysS Genotype:F– ompT hsdSB(rB– mB
–) gal dcm lacY1 ahpC (DE3) gor522::Tn10 trxB pLysSRARE (CamR, KanR, TetR)
9

分子生物学・遺伝子工学用の顆粒状 / 粉末状培地シリーズのご紹介です.
高品質な大腸菌培養用培地シリーズ
Molecular Biology Media
TOPIX
製品名 注文番号 包装単位 希望販売価格
2xYT Broth 71755-3 500 g ¥11,400
71755-4 5 kg ¥87,100
製品名 注文番号 包装単位 希望販売価格
Veggie™ Peptone 71280-3 500 g ¥17,100
Veggie™ Yeast Extract 71279-3 500 g ¥22,300
製品名 注文番号 包装単位 希望販売価格
LB Broth Miller 71753-3 5 EasyPak ¥6,800
71753-5 500 g ¥8,700
71753-4 25 EasyPak ¥30,400
71753-6 5 kg 弊社照会
71753-7 25 kg 弊社照会
LB Agar Miller 71752-3 5 EasyPak ¥10,100
71752-5 500 g ¥16,500
71752-4 25 EasyPak ¥45,500
71752-6 5 kg ¥110,000
製品名 注文番号 包装単位 希望販売価格
LB Broth Lennox 71751-3 5 EasyPak ¥6,800
71751-5 500 g ¥10,300
71751-4 25 EasyPak ¥30,400
71751-6 5 kg ¥81,400
Terrific Broth 71754-3 500 g ¥10,300
71754-4 5 kg ¥81,400
71754-5 10 kg ¥147,400
LB Broth,LB Agar および Terrfic Broth は顆粒状培地です.粉末状培地に比べて秤量,調整の際に培地の飛散が少なく培地の溶解もスムーズです.
顆粒状培地
粉末状培地
EasyPak1 L 用の EasyPak パウチ包装の顆粒状 LB Broth ,LB Agar です.秤量などの煩雑な作業は不要です.1 L の水に EasyPakの内容物を加え,溶解後電子レンジにて加熱してください.オートクレーブ滅菌も可能です.
LB Broth MILLER1L あたりの LB Broth MILLER の成分は,酵母エキス 5g,カゼインペプトン 10g,塩化ナトリウム 10g です.
LB Agar MILLER1L あたりの LB Broth MILLER の成分は,酵母エキス 5g,カゼインペプトン 10g,塩化ナトリウム 10g,agar 12g です.
LB Broth LENNOX1L あたりの LB Broth LENNOX の成分は,酵母エキス 5g,カゼインペプトン 10g,塩化ナトリウム 5g です.
Terrific Broth1L あたりの Terrific Broth の成分は,トリプトン 12g,酵母エキス 24g,リン酸 2 カリウム 9.4g,リン酸 1 カリウム 2.2g です.
2×YT Broth1L あたりの 2xYT Broth の成分は,トリプトン 16g,酵母エキス 10g,塩化ナトリウム 5g です.
Veggie™ Peptone and Veggie™ Yeast ExtractVeggie 製品シリーズは,動物由来成分の使用を避けたい実験用途に理想的です.Veggie Peptone は,大豆のパパイン消化ペプトンで動物由来成分を含まないことを保証した培地成分であり,大腸菌増殖培地におけるトリプトンの直接的な代替品として使用できます.Veggie Yeast Extract は,大腸菌増殖培地における従来の酵母抽出物の直接的な代替品として使用可能な,動物由来物質を含まないことの保証された培地成分です.両製品とも品質試験によって,細胞の適切な増殖と維持が保証されています.
10
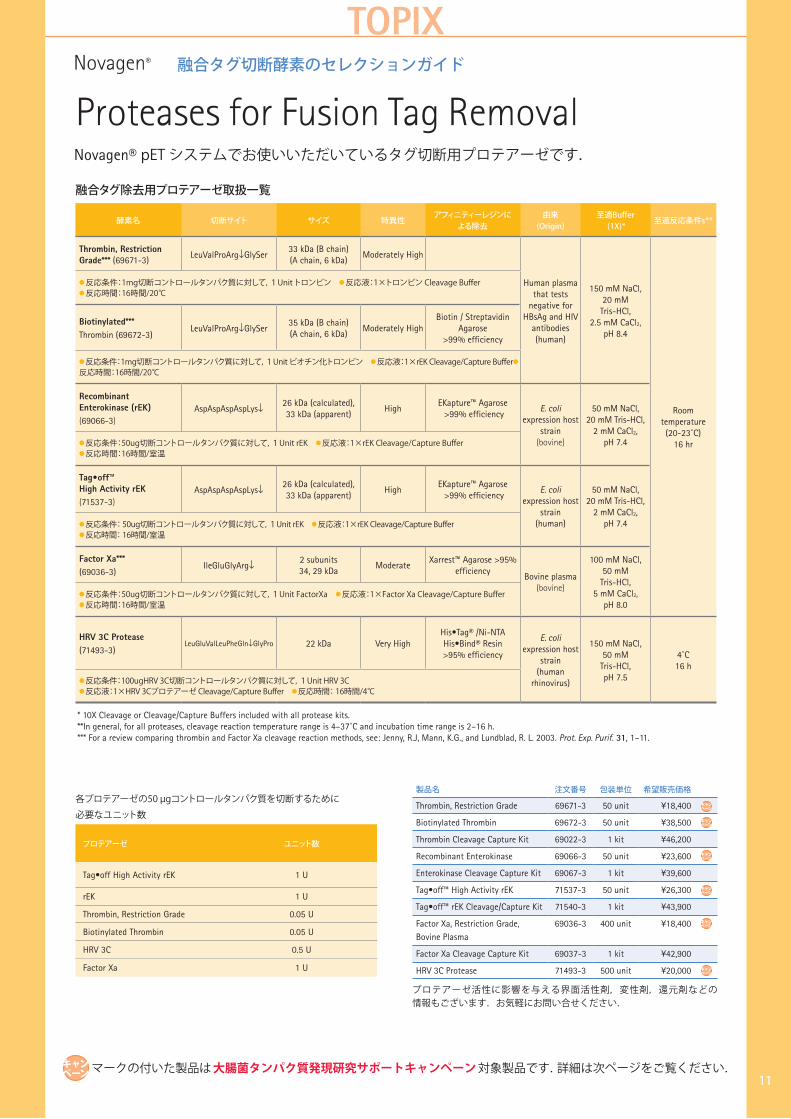
融合タグ切断酵素のセレクションガイド
Novagen® pET システムでお使いいただいているタグ切断用プロテアーゼです.
プロテアーゼ活性に影響を与える界面活性剤,変性剤,還元剤などの 情報もございます.お気軽にお問い合せください.
Proteases for Fusion Tag Removal
TOPIX
融合タグ除去用プロテアーゼ取扱一覧
酵素名 切断サイト サイズ 特異性アフィニティーレジンに
よる除去由来(Origin)
至適Buffer(1X)*
至適反応条件s**
Thrombin, Restriction Grade*** (69671-3)
LeuValProArg↓GlySer33 kDa (B chain)(A chain, 6 kDa)
Moderately High
Human plasma that tests
negative for HBsAg and HIV
antibodies(human)
150 mM NaCl,20 mM
Tris-HCl,2.5 mM CaCl2,
pH 8.4
Roomtemperature(20-23˚C)
16 hr
● 反応条件:1mg切断コントロールタンパク質に対して,1Unit トロンビン ● 反応液:1×トロンビン Cleavage Buffer● 反応時間:16時間/20℃
Biotinylated***Thrombin (69672-3)
LeuValProArg↓GlySer35 kDa (B chain)(A chain, 6 kDa)
Moderately HighBiotin / Streptavidin
Agarose >99% efficiency
● 反応条件:1mg切断コントロールタンパク質に対して,1Unit ビオチン化トロンビン ● 反応液:1×rEK Cleavage/Capture Buffer●
反応時間:16時間/20℃
Recombinant Enterokinase (rEK) (69066-3)
AspAspAspAspLys↓ 26 kDa (calculated), 33 kDa (apparent)
High EKapture™ Agarose >99% efficiency
E. coli expression host
strain (bovine)
50 mM NaCl,20 mM Tris-HCl,
2 mM CaCl2,pH 7.4● 反応条件:50ug切断コントロールタンパク質に対して,1Unit rEK ● 反応液:1×rEK Cleavage/Capture Buffer
● 反応時間:16時間/室温
Tag•off™ High Activity rEK(71537-3)
AspAspAspAspLys↓ 26 kDa (calculated), 33 kDa (apparent)
High EKapture™ Agarose >99% efficiency
E. coli expression host
strain (human)
50 mM NaCl,20 mM Tris-HCl,
2 mM CaCl2,pH 7.4● 反応条件: 50ug切断コントロールタンパク質に対して,1Unit rEK ● 反応液:1×rEK Cleavage/Capture Buffer
● 反応時間: 16時間/室温
Factor Xa***(69036-3)
IleGluGlyArg↓ 2 subunits 34, 29 kDa
ModerateXarrest™ Agarose >95%
efficiencyBovine plasma
(bovine)
100 mM NaCl,50 mM
Tris-HCl,5 mM CaCl2,
pH 8.0● 反応条件:50ug切断コントロールタンパク質に対して,1Unit FactorXa ● 反応液:1×Factor Xa Cleavage/Capture Buffer● 反応時間:16時間/室温
HRV 3C Protease (71493-3)
LeuGluValLeuPheGln↓GlyPro 22 kDa Very HighHis•Tag® /Ni-NTA His•Bind® Resin >95% efficiency
E. coli expression host
strain (human
rhinovirus)
150 mM NaCl,50 mM
Tris-HCl,pH 7.5
4˚C16 h
● 反応条件:100ugHRV 3C切断コントロールタンパク質に対して,1Unit HRV 3C ● 反応液:1×HRV 3Cプロテアーゼ Cleavage/Capture Buffer ● 反応時間: 16時間/4℃
* 10X Cleavage or Cleavage/Capture Buffers included with all protease kits.**In general, for all proteases, cleavage reaction temperature range is 4–37˚C and incubation time range is 2–16 h.*** For a review comparing thrombin and Factor Xa cleavage reaction methods, see: Jenny, R.J, Mann, K.G., and Lundblad, R. L. 2003. Prot. Exp. Purif. 31, 1–11.
各プロテアーゼの50 µgコントロールタンパク質を切断するために
必要なユニット数
プロテアーゼ ユニット数
Tag•off High Activity rEK 1 U
rEK 1 U
Thrombin, Restriction Grade 0.05 U
Biotinylated Thrombin 0.05 U
HRV 3C 0.5 U
Factor Xa 1 U
製品名 注文番号 包装単位 希望販売価格
Thrombin, Restriction Grade 69671-3 50 unit ¥18,400
Biotinylated Thrombin 69672-3 50 unit ¥38,500
Thrombin Cleavage Capture Kit 69022-3 1 kit ¥46,200
Recombinant Enterokinase 69066-3 50 unit ¥23,600
Enterokinase Cleavage Capture Kit 69067-3 1 kit ¥39,600
Tag•off™ High Activity rEK 71537-3 50 unit ¥26,300
Tag•off™ rEK Cleavage/Capture Kit 71540-3 1 kit ¥43,900
Factor Xa, Restriction Grade, Bovine Plasma
69036-3 400 unit ¥18,400
Factor Xa Cleavage Capture Kit 69037-3 1 kit ¥42,900
HRV 3C Protease 71493-3 500 unit ¥20,000
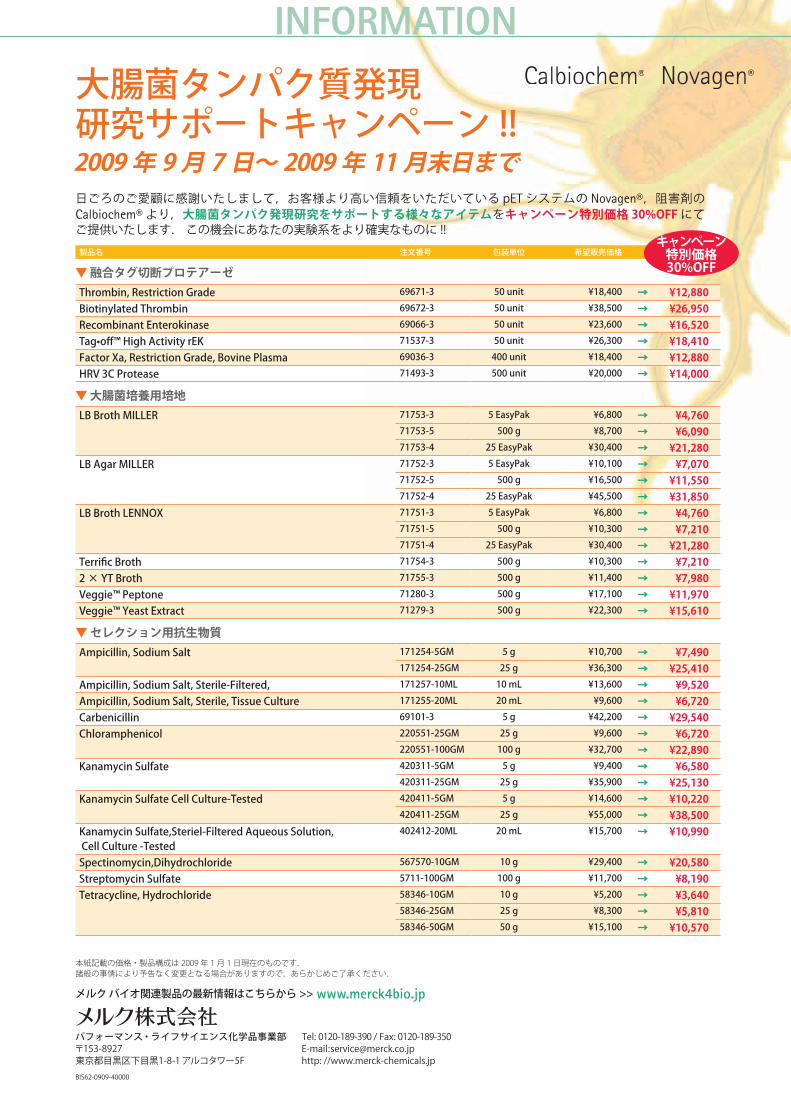
11★ マークの付いた製品は 大腸菌タンパク質発現研究サポートキャンペーン 対象製品です.詳細は次ページをご覧ください.
BIS62-0909-40000
メルクバイオ関連製品の最新情報はこちらから>>www.merck4bio.jp
パフォーマンス・ライフサイエンス化学品事業部 Tel: 0120-189-390 / Fax: 0120-189-350〒153-8927 E-mail:[email protected]東京都目黒区下目黒1-8-1 アルコタワー5F http: //www.merck-chemicals.jp
本紙記載の価格・製品構成は 2009 年 1 月 1日現在のものです.諸般の事情により予告なく変更となる場合がありますので,あらかじめご了承ください.
INFORMATION
日ごろのご愛顧に感謝いたしまして,お客様より高い信頼をいただいている pET システムの Novagen®,阻害剤のCalbiochem® より,大腸菌タンパク発現研究をサポートする様々なアイテムをキャンペーン特別価格 30%OFF にてご提供いたします. この機会にあなたの実験系をより確実なものに !!
製品名 注文番号 包装単位 希望販売価格
▼ 融合タグ切断プロテアーゼThrombin,RestrictionGrade 69671-3 50unit ¥18,400 → ¥12,880BiotinylatedThrombin 69672-3 50unit ¥38,500 → ¥26,950RecombinantEnterokinase 69066-3 50unit ¥23,600 → ¥16,520Tag•off™HighActivityrEK 71537-3 50unit ¥26,300 → ¥18,410FactorXa,RestrictionGrade,BovinePlasma 69036-3 400unit ¥18,400 → ¥12,880HRV3CProtease 71493-3 500unit ¥20,000 → ¥14,000
▼ 大腸菌培養用培地LBBrothMILLER 71753-3 5EasyPak ¥6,800 → ¥4,760
71753-5 500g ¥8,700 → ¥6,09071753-4 25EasyPak ¥30,400 → ¥21,280
LBAgarMILLER 71752-3 5EasyPak ¥10,100 → ¥7,07071752-5 500g ¥16,500 → ¥11,55071752-4 25EasyPak ¥45,500 → ¥31,850
LBBrothLENNOX 71751-3 5EasyPak ¥6,800 → ¥4,76071751-5 500g ¥10,300 → ¥7,21071751-4 25EasyPak ¥30,400 → ¥21,280
TerrificBroth 71754-3 500g ¥10,300 → ¥7,2102 × YTBroth 71755-3 500g ¥11,400 → ¥7,980Veggie™Peptone 71280-3 500g ¥17,100 → ¥11,970Veggie™YeastExtract 71279-3 500g ¥22,300 → ¥15,610
▼ セレクション用抗生物質Ampicillin,SodiumSalt 171254-5GM 5g ¥10,700 → ¥7,490
171254-25GM 25g ¥36,300 → ¥25,410Ampicillin,SodiumSalt,Sterile-Filtered, 171257-10ML 10mL ¥13,600 → ¥9,520Ampicillin,SodiumSalt,Sterile,TissueCulture 171255-20ML 20mL ¥9,600 → ¥6,720Carbenicillin 69101-3 5g ¥42,200 → ¥29,540Chloramphenicol 220551-25GM 25g ¥9,600 → ¥6,720
220551-100GM 100g ¥32,700 → ¥22,890KanamycinSulfate 420311-5GM 5g ¥9,400 → ¥6,580
420311-25GM 25g ¥35,900 → ¥25,130KanamycinSulfateCellCulture-Tested 420411-5GM 5g ¥14,600 → ¥10,220
420411-25GM 25g ¥55,000 → ¥38,500KanamycinSulfate,Steriel-FilteredAqueousSolution,CellCulture-Tested
402412-20ML 20mL ¥15,700 → ¥10,990
Spectinomycin,Dihydrochloride 567570-10GM 10g ¥29,400 → ¥20,580StreptomycinSulfate 5711-100GM 100g ¥11,700 → ¥8,190Tetracycline,Hydrochloride 58346-10GM 10g ¥5,200 → ¥3,640
58346-25GM 25g ¥8,300 → ¥5,81058346-50GM 50g ¥15,100 → ¥10,570
大腸菌タンパク質発現研究サポートキャンペーン !!2009 年 9 月 7 日〜 2009 年 11 月末日まで
キャンペーン特別価格 30%OFF





























