Embed Size (px)
Citation preview
h ' ' 'f MASGC-B-78-001
Ac. 3
MARINE MALADIES?
Worms, Germs, and Other SymbiontsFrom the Northern Gulf of Mexico
CRCDU7M COPYSea Grant Depositor
NATIONAL SEA GRANT DEPOSITORY \PELL LIBRARY BUILDING
URI NA8RAGANSETT BAY CAMPUSNARRAGANSETT. Rl 02882
%
Robin M. Overstreet
r ii
MISSISSIPPI—ALABAMASEA GRANT CONSORTIUM
MASGP—78—021

MARINE MALADIES?
Worms, Germs, and Other Symbionts
From the Northern Gulf of Mexico
by
Robin M. Overstreet
Gulf Coast Research Laboratory
Ocean Springs, Mississippi 39564
This study was conducted in cooperation with the U.S. Department of Commerce, NOAA,Office of Sea Grant, under Grant No. 04-7-158-44017 and National Marine Fisheries Service,under PL 88-309, Project No. 2-262-R. The Mississippi-AlabamaSea Grant Consortium furnished all of the publication costs. The U.S. Government is authorized to produceand distributereprints for governmental purposes notwithstanding any copyright notation that may appearhereon.

Copyright© 1978by Mississippi-Alabama Sea Gram Consortium and R.M. Overstrect
All rights reserved. No pari of this bookmay be reproduced in any manner without
permission from the author.
Primed by Blossman Printing, Inc.. Ocean Springs, Mississippi

CONTENTS
PREFACE 1
INTRODUCTION TO SYMBIOSIS 2
INVERTEBRATES AS HOSTS 5
THE AMERICAN OYSTER 5
Public Health Aspects 6
Dcrmo 7
Other Symbionts and Diseases 8
Shell-Burrowing Symbionts IIFouling Organisms and Predators 13
THE BLUE CRAB 15
Protozoans and Microbes 15
Mclazoans and their I lypeiparasites 18Misiellaneous Microbes and Protozoans 25
PENAEID SHRIMPS 28
Microbial Diseases 28
Protozoan Infections 32
Helminths 34
Miscellaneous Diseases and Attached Organisms 38
GRASS SHRIMP 10
Protozoans 41
Metazoans 42
VERTEBRATES AS HOSTS 46
FISHES 16
Microbes 47
Protozoans in General 50
Flagellates 50Amoebas 5-
Sporozoans 52Microsporidans 53Myxosporidans 54Ciliates 55Tapeworms JlMonogenean Flukes <"Digenean Flukes &/Roundworms ,-
Thorny-headetl Worms 79Crustaceans 80
MISCELLANEOUS HOSTS 85The American Alligator 85Birds 86Marine Mammals 88
MISCELLANEOUS SYMBIONTS 89FISH-KILLS AND MISCELLANEOUS DISEASES 92

TECHNICAL ASPECTS 96
SYMBIOSIS 96
THE AMERICAN OYSTER 98
THE BLUE CRAB 101
PENAEID SHRIMPS 104
GRASS SHRIMP 107
FISHES 107
Microbes 108
Protozoans 108
Cestodes Ill
Monogeneans 112
Digeneans 113
Nematodes 114
Acanthocephalans 114
Crustaceans 115
MISCELLANEOUS HOSTS 116
The American Alligator 116
Birds 116
Marine Mammals 118
MISCELLANEOUS SYMBIONTS 118
FISH-KILLS AND MISCELLANEOUS DISEASES 119
ACKNOWLEDGMENTS 121
LITERATURE CITED 122
GLOSSARY 137

PREFACE
This guidebook will inform those curiousabout parasites and other symbionts associated with marine and estuarine hosts in
the northern Gulf of Mexico. Designed as ateaching aid for students, fishermen, seafood consumers, beachcombers, and even
parasitologists, it should allow for better
understanding and appreciation of severalof the numerous shellfish and finfish symbionts. (A symbiotic organism is one thatinteracts with the host in or on which it
lives.) Even though most selected examplesare from Mississippi and other regions a-long the northern Gulf, the same or relatedspecies also occur along the Atlantic seaboard and elsewhere; present informationshould apply similarly for several but not allof those cases. Some symbionts significantlyaffect the size and production of a fisheryand consumption of the product, whereasothers mostly stimulate environmental oracademic interests. In the natural environ
ment, parasitic relationships seldom resultin harm to the host. Harm, however, often
becomes apparent when animals are concentrated and confined as they are duringculture or when they are otherwise stressed.This guide will discuss some of those cases.
This guidebook is divided into sectionsdiscussing the various hosts of parasites.Primary headings refer to basic host-types.The reader, however, must keep in mindthat groups of both symbionts and hostsoverlap. The same parasite may have adifferent stage of its life history infecting ashrimp, a fish, and a bird. Consequently,perusal of several sections may help thereader understand more about the symbionts of any particular host.
The guidebook has been written withboth the student without a strong biologicalbackground and the layman in mind, but itpresents additional separate notes for moreserious readers interested in technical as
pects. That technical section, in the latterhalf, directs the reader to referenced scienti
fic literature which either presents supporting data or reviews the topics under con-
Preface 1
sideration. An effort has been made to define
scientific terminology either in the text or ina glossary at the end of the guide. Terms presented in the glossary are indicated by anasterisk (*) the first and occasionally other
times they are used. Diagrams, illustrations,photographs, and legends provide furtherassistance.
Latin names have been included as well
as common names for three reasons. First,
some organisms have no common name.Second, a specific common name may referto more than one animal or one animal mayhave several common names. When most
people talk about the blue crab, they probably talk about Callinectes sapidus, butthey might also knowingly or unknowinglybe referring to Callinectes similis, Callinectes omatus, or a variety of other related
crabs. On the other hand, residents of
Mississippi call the spotted seatrout (Cyno-scion nebulosus) a speck or speckled troutwhereas people elsewhere may call it or arelated species by the name weakfish,squeteague, or any of several others. Third,knowing a Latin name often allows one tomore easily find other literature about theorganism.
A binomial name, that is the two-component Latinized name of an organism, consists of the capitalized generic name and thenoncapitalized specific name. One or severalspecies may occur in a genus, and differenttaxonomists, scientists who deal with thenomenclatural problems, may not all agreeto which genus a given species belongs.Nevertheless, names allow people to refer tospecific organisms. Some names reflect theanimal's characteristics, its locality, or itsfinder, whereas others are merely fabricated.
Readers interested in the hygienic aspectsof eating infected finfishes and shellfishesmay be comforted by the fact that cookingdestroys all potentially harmful agents inseafood products from the northern Gulf ofMexico. Perhaps this statement needs somequalification. A person could acquire gastric distress from eating cooked seafood ifstaphylococci toxin was present, if contam-

2 Marine Maladies?
inalion occurred after cooking, if the product was spoiled before cooking, or ifcondiments contained salmonellae, shi-
gellae, or some other infectious bacteria.Also, heavy metals and some other nonparasitic toxins are not destroyed bycooking. Products infected with parasitesshould not dismay a consumer. Seldom doesone get the opportunity to see some of thesepuzzling little invadersl If infected productsare cooked, all become edible, a few have less
appeal and flavor, and several taste better because of the added rich, juicy worm nestledamong the tissue. I am willing to admit,however, that most people preferring infected products do not realize that the"flavor bud" is a parasite.
INTRODUCTION TO
SYMBIOSIS
As is the case with many scientific endeavors, the same concept or item may bereferred to by different terms, and, conversely, different concepts or items can be calledthe same name. Seldom does one realize that
confusion exists until he progresses into adiscussion. What is a parasite? Does it haveto harm its host? If so, does it have to harm
all the different hosts in all stages of its life?Does it have to be smaller than its host? Can
it be the same speciesas the host?Dependingon how one prefers to answer the abovequestions, even a human baby can qualifyas a parasite. Before birth the fetus nourishesitself at the expense of its host (mother) andreleases toxic wastes into that host; occas
ionally at birth, it even causes the host'sdeath. After normal birth, it continues to
suck vital nutrients from the host. From
another viewpoint, even after maturing,that offspring may obtain its well beingfrom its parents or friends at their expense. By defining terms now, we shouldhave a clearer understanding of some of thedifferent types of relationships among animals. Three terms that must be understood
to aid in understanding parasites are
"symbiosis," "commensalism," and "mutualism."
"Symbiosis," which means "living together," accounts for a variety of long andshort term relationships that benefit one orboth parties. For purposes of simplicity, Iconsider symbiotic relations to be those specialized associations ranging from commensalism, where neither party is harmedand both could live without the other, to
mutualism, where both parlies benefit eachother and neither could live without the
other. A true parasite lies in between. Parasitism defines a one-way relationship inwhich one partner, the parasite, dependsupon and benefits from the other partner,the host. The host is usually the larger of anytwo associates and can be harmed by theparasite. Actually, much overlap in types ofrelationships exists, and, for most readers,what happens in a given relationship farsurpasses in importance the term someonemay want to tag on it.
"Commensalism" means "eating at thesame table." A sea anemone attached to the
mollusc an shell of a hermit crab derives
mobility and scraps of food from the crabwithout being metabolically dependent onit. The relationship, however, might not beone sided and may be truly mutual. In somecases, the anemone, by virtue of its stingingtentacles, contributes by warding off octopuses or other predators from the crab. Thedegree of benefit to each party depends onthe species of anemone and crab involved aswell as the environmental conditions and
the additional organisms which make upthe community*. In the northern Gulf ofMexico, a specific anemone (Calliactis tricolor) usually attaches to a shell, often of themoon snail, inhabited by a hermit crab(e.g., Pagurus longicarpus), as illustrated inFigure 1.
Some inhabited shells harbor hydroidsrather than anemones. Two species of hydroids (Podocoryne selena and Hydractiniaechinata) commonly use this movable substratum* in the northern Gulf of Mexico
(Figure 2). They probably benefit the crabbecause colonies of each possess protective

Figures 1 and 2. Hermit crabs with symbiontson theU molluscan shells. Top, twoindividuals of an anemone (Calllactlstricolor) on the moon snail Inhabited byPagurus longlcarpus. Bottom, a hydroidcovering the shell housing Paguruspollicarls.
stinging zooids* in addition to the nutritive, generative, and sensory ones found incolonies of related nonsymbiotic hydroids.
Commensalism can be expanded to include "phoresis" meaning "traveling together." The two parties in this relationship do not eat at the same table. A possibleexample of a phoront is a hydroid (Clytiasp.) that attaches to the posterior edge of a
live coquina's shell (Figure 3). Theseclams in Texas with the hydroid attachedcontained no internal parasitic flukes. Thisfinding suggests the hydroid prevented theparasitic infection*. If true, we could interpret the clam-hydroid relationship as mu-tualistic, a relationship to be discussedlater, because both parties benefit. If Clytiasp. is a phoront, it benefits only by utilizing
Symbiosis 3
Figure 3. A commensal hydroid (Clytia sp.)attached to a coquina clam (Donax roemerlprotracta). The hydroid benefits by having asubstratum along a sandy beach on which toattach, but neither clam nor hydroid harmsthe other.
the clam shell as something hard on whichto attach in the turbulent beach habitat. If,however, the clam benefits by being protected from infection by a parasite that destroysits reproductive tissue, that hydroid is somewhat mutualisticeven though both the clamand hydroid can live without the other.
Already the reader probably realizes thatwe do not always know what constitutes aparasite. When a symbiont depends entirelyon a host, occasionally harming it in theprocess, it is unequivocally a parasite. Harmcan result from boring into tissues, digesting tissues, displacing vital tissues, releasing toxic metabolic products, or competingfor nutrients. Some parasites even live apartfrom their hosts for a portion of their lives,whereas others always remain with theirhosts, but act as commensal until confront
ed with a particular stress. Tapeworms andspiny-headed worms are obvious parasitessince they have no gut and necessarily depend on their hosts to provide all theirnutrients in a state that can pass throughtheir body surfaces. Whether many othersymbionts are parasites requires a subjectivedecision.
The difference between a parasite and predator can also be confusing. Some animalskill their prey outright rather than dependon a living source of food, and these constitute predators. When animals feed on avariety of prey much smaller than themselves, they act as predators. However, somemicropredators and even large predatorsperiodically eat away part of specific species

4 Marine Maladies?
of prey without killing them, and thesecould be considered parasites. I will let thereader term such a relationship anything hedesires, but the more biochemically andphysiologically dependent a "predator" becomes on a given "prey," the more parasiticthe relationship becomes.
In a well-adapted host-parasite relationship, the host is not significantly harmed.This may sound like a contradiction, but asa parasite becomes more dependent on a specific host species, it becomes more vulnerable to extinction if it or anything elseseriously harms that host. It must be able toobtain its well being from the host withoutharming it because the parasite needs ahealthy host to survive. Strangely, however,a well-adapted parasite often benefits whenone of its larval stages affects the intermediate host in such a manner that the final
host can prey on that intermediate host moreeasily and thereby insure completion of theparasite's life cycle.
Usually for a parasite to harm its host, thehost is either stressed or weakened or the in
fection is either "accidental" or heavy. Anaccidental, or incidental, parasite afflictsa host other than its normal one. Excessive
numbers of an otherwise harmless parasitein the normal host can cause disease*.
Twenty or so hookworms in an individual'sintestine would have no esthetic appeal if heknew about them, but such an infection
would not constitute disease. When the
number of worms increases, especially in amalnourished patient, a disease becomesmanifest. The actual number of worms
needed to cause bloody stools, anemia, lossof appetite, a desire to eat soil, and othersymptoms of disease depends on both thespecies of hookworm and the individualperson (host) involved.
"Mutualism" requires that both mutualsdepend on each other. In the strict sense, thismust be a metabolic* dependency in whichneither party survives without the other.The combination of a termite with its inter
nal protozoans provides an example close tohome. Intestinal flagellates produce the
enzyme cellulase which digests the ingestedwood so that the termite can utilize it.
Neither animal can survive for long by itself,and the two together do much to benefit anecological community* (assuming a houseis not the food source!). In the ocean, numerous cases of mutualism occur. Many ofthese include specific algae associated withparticular invertebrates.
Symbiotic algae inhabit various tissuesor cells of invertebrates belonging to numerous genera in at least ten phyla*. Reef-building corals provide often-mentionedexamples. An assumption dictates that allthe invertebrate hosts would die without
their algae, but experimental studies haveshown that these otherwise greenish orbrownish (from chlorophyll*) corals andother cnidarians may survive when bleachedin darkness from shedding their symbionts.Nevertheless, the well-adapted relationshipsbenefit both parties, even though some animals may appear to exploit their algal mu-tualist excessively. In each association, thebenefits probably vary, and most have notbeen thoroughly investigated. The invertebrate host provides nutrients and shelter forthe alga. The alga provides the host oxygenand organic nutrients (by photosynthesis*);removes carbon dioxide, nitrogenous wastesand other metabolic products from hostcells; provides protective coloration forthe host; and in the case of corals, aids in
deposition of a calcified skeleton. Few ofthese algae have been cultured and properlyidentified. Consequently, for most associations, an alga's possible relationships withdifferent invertebrates, its metabolic de
mands on a host, and its ability to livewithout a host have not been firmly esta
blished.
Most conspicuous examples of invertebrate-alga mutualism occur in tropicalwaters, but some less conspicuous onesoccur in the northern Gulf. Perhaps somereaders will search for the typically greenishor brownish invertebrate hosts. One is the
sun jellyfish (Cassiopea xamachana) whichoccasionally can be found near Horn Island

where, when not swimming, it rests on thebottom with its underside facing up, exposing the algal symbionts.
Hosts from Florida which appear toutilize multiple symbionts are colonialzoanthid anthozoans (two species ofZoanthus, see Figure 4 for one of them).Their greenish-brown polyps with extrudedtentacles give the impression of a flower;the color is derived mostly from the internalgolden-brown zooxanthella. Rather thanhaving one symbiont like corals and otherinvestigated hosts, these cnidarians have avariety of symbionts. One form migratesextensively, perhaps depending on theamount of light present. Since the Floridianzoanthids readily feed when presented smallcrustaceans or pieces of fish, the symbiontsappear to have a function other than a solenutritive source. Some other animals
culture their food. Algae are grown inamphipod tubes by some amphipodinhabitants, and fungi grow on thepieces of leaves provided by leaf-cutting ants. The identification andinterrelationships among the zoanthids andsymbionts other than the zooxanthella(alga) which is similar in appearance to thatin most other cnidarian-algal associationshave not been established.
INVERTEBRATES ASHOSTS
THE AMERICAN OYSTER
The American oyster (Crassostreavirginica) supports an important fisheryin most regions where it occurs along theAtlantic and Gulf of Mexico coasts fromCanada to Mexico. The benthic* animalgrows rapidly in the warm water of thenorthern Gulf, reaching 8 centimeters (cm)(about 3 inches, see glossary for examplesof measurements) in as short a period as 4months, but typically taking a year or so.When on soft mud, it sinks and conse
quently grows thin and long to keep its
American Oyster 5
Figure 4. A colonial zoanthid anemonecommonly called a green sea mat. Theanimal normally utilizes internal plant andanimal symbionts for its well-being.
bill, or flared end, above the mud. On theother hand, on a firm uncrowded bottom,the oyster forms a broad cupped shell. Itgrows best in estuarine* water withreasonably constant salinity* between15 and 30 ppt.
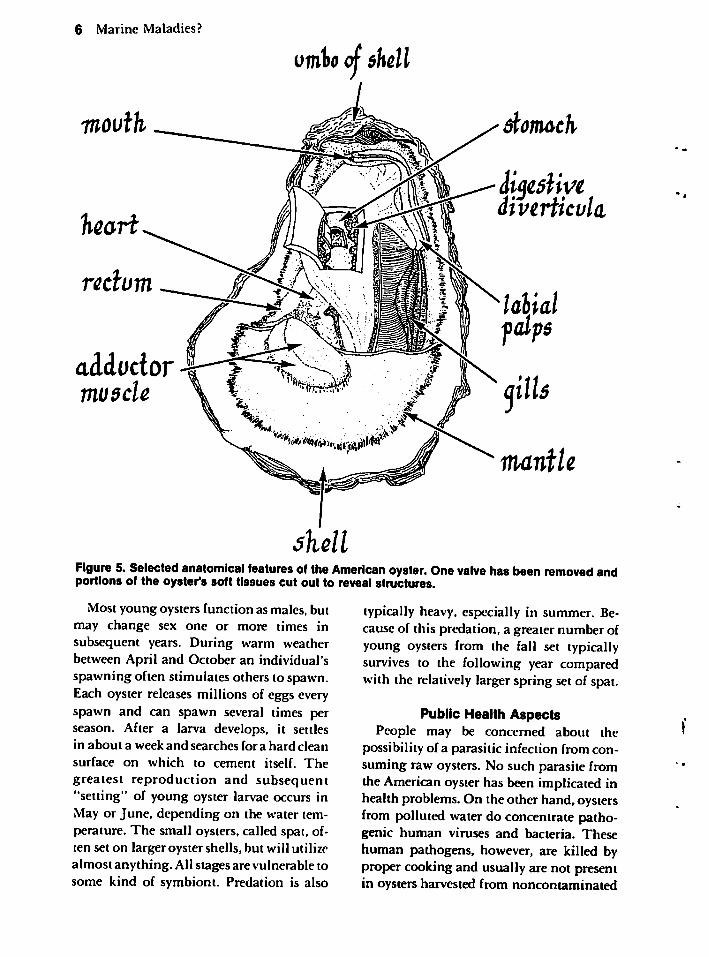
Figure 5 illustrates a partially shucked(opened) oyster. In a closed oyster, the twovalves join anteriorly immediately internalto the umbo, or beak, by a continuallyreplaced elastic hinge ligament. The largeadductor muscle, referred to as the "eye" byshuckers, holds the valves together and mustbe cut to open the oyster.
Special cells in the mantle secrete threeprimary layers of the shell. Externally, athin horny organic layer (periostracum)covers a middle prismatic layer of mineralwhich develops most substantially on theright flat valve. The inner shiny layer(calcific ostracum) usually constitutes thethickest stratum.
Features of the soft body are mostly self-explanatory. Dark bands ofsensory tentaclesborder mantle flaps. Ciliary rows of the gillsdirect up to 28 liters (about 30 quarts) ofwater per hour. The gills remove oxygenfrom and excrete wastes into that water.
They also accumulate food which, whenacceptable to the oyster, continues to beconveyed to the mouth by action of cilia(these structures will be described in a latersection on protozoans of fishes) on the gillsand palps.
6 Marine Maladies?
vmho of shell
adductormu$cle
shell
dlomach
iigestiyediverticala
iticnvtle
Figure 5. Selected anatomical features of the American oyster. One valve has been removed andportions of the oyster's soft tissues cut out to reveal structures.
Most young oysters function as males, butmay change sex one or more times insubsequent years. During warm weatherbetween April and October an individual'sspawning often stimulates others to spawn.Each oyster releases millions of eggs everyspawn and can spawn several times perseason. After a larva develops, it settlesin about a week and searches for a hard clean
surface on which to cement itself. The
greatest reproduction and subsequent"setting" of young oyster larvae occurs inMay or June, depending on the water temperature. The small oysters, called spat, often set on larger oyster shells, but will utilize
almost anything. All stages are vulnerable tosome kind of symbiont. Predation is also
typically heavy, especially in summer. Because of this predation, a greater number ofyoung oysters from the fall set typicallysurvives to the following year comparedwith the relatively larger spring set of spat.
Public Health AspectsPeople may be concerned about the
possibility of a parasitic infection from consuming raw oysters. No such parasite fromthe Americanoyster has been implicated inhealth problems. On the other hand, oystersfrom polluted water do concentrate pathogenic human viruses and bacteria. Thesehuman pathogens, however, are killed byproper cooking and usually are not presentin oysters harvested from nonconlaminated

•f *; yi^i j.* ja&^e'isv.
Figure 6. The middle gaping oyster cannotclose its valves tightly. Several disease-agents and environmental stresses cancause gaping, but this condition is acommon sign during the hot summerindicating that the oysters may have heavyinfections of dermo. These dying oystersbecome easy prey for a numberof predators,including organisms previously acting ascommensals.
waters. Certain inorganic contaminants,such as lead or mercury, and some algaltoxins are sometimes found in pollutedwaters, and these may be concentratedin oysters growing there. These potentialtoxins are not removed by cooking.
Even though not harmful to man, severalparasites do injure the oyster. Most of thesecause significant disease* in oysters onlywhen antagonized by additional stresses.Environmental stresses — such as hightemperature, low salinity, excessivesiltation, or pollutants —probablyall affectsome symbiotic relationships. When threatened, the soft-bodied oyster closes togetherits two thick shells, or valves. After becoming overpowered by predators orintolerable conditions, the oyster gapes(Figure 6) and soon after dies or is eaten.
Dermo
Many oysters in the northern Gulf ofMexico gape because of "dermo" acolloquial name for a disease caused byDermocystidium marinum (presentlyknown as Perkinsus marinus). This micro
organism, once thought to be a protozoan,then a fungus, and now a protozoan again,cannot be seen without a microscope;however, oysters dying in late summer witha shrunken appearance and a yellowish castprobably have dermo. For a long time such
American Oyster 7
mortalities in Louisiana had been blamed
on oil pollution until the oil industry contracted Texas A&M to investigate theproblem in the 1950's. Findings of thatstudy revealed many things aboutthe dermo organism, including howto detect it easily. When infected oystertissue is placed for a week or more in athioglycollate culture medium with antibiotics, the organisms enlarge and becomereadily distinguishable. These enlargedcells develop a wall which turns a bluish-black color when stained with an iodine
solution (Lugol's) (Figure 7). Clusters ofthese spores* can be detected without amicroscope. Most tissues of infectedindividuals harbor the parasite, but thegills, rectum, mantle, or adductor muscleprovide the best results.
Infections* by dermo have importanteconomic implications. The prevalence*(percentage) of infected oysters and thenumber of organisms per infected individual typically remain low during winterand increase considerably during summer.Periodically, the majority of adult oysterson a reef succumb to this pathogen duringhot summer months, producing massivekills, or epizootics. In order to avoid theloss of these oysters to consumers severalyears ago, William Demoran of theMississippi Marine ConservationCommission would sample oysters from
Figure 7. Enlarged spores of dermo(Dermocystidium marinum) that have beencultured in a special fluid (thioglycollatemedium) and stained with iodine. Themethod allows easy detection of thisorganism which kills many stressed oystersduring the hot summer.

8 Marine Maladies?
Lower Square Handkerchief Reef duringthe winter. If the sample revealed a highprevalence of dermo, the Commissionallowed most of the oysters from that vulnerable region to be harvested then,rather than wait for the expected summermortalities.
Salinity has often been considered thecritical factor regulating the disease sinceinfections rarely occur in water less than
15 ppt. Such is not always the case. In fact,biologists from Mississippi and Floridahave found over half of many samples fromwater less than 15 ppt to be infected. Oftenoysters have infections in water that is freshor quite low in salt content.
Apparently temperature acts as theprimary controlling factor. Acquisition ofmost infections probably occurs duringa period of high salinity, butdevelopment ismost pronounced when temperature isrelatively high. High temperature may decrease the host's ability to defend itselfagainst the organism, just as indiscriminantuse of steroids can cure one condition in a
person but allow another previouslycontrolled condition to become expressed.
Juvenile oysters do not seem to developdermo as readily or severely as older ones.Although host-mediated resistance* (alsosee humoral and cellular mechanisms inglossary) in juveniles has been suggested,thephenomenon is probably because youngoysters are in comparatively better healththan older ones and therefore better ableto fight off or control any challenginginfective agents. Active defense mechanismsused to fight the parasite are well known andchemical defenses may exist as well. Also,young oysters commonly grow so fast thatthey can reach adult size as fast as an infection can occur. This is especially true ofthe numerous late summer-spawned oystersthat grow through the winter when theparasite is limited by lower temperature.Oysters seem most susceptible to infectionand disease immediately after spawning.The extreme stress associated with spawning apparently greatly weakens the oysters
and their defenses against the parasite.Heavy oyster mortalities — an estimated
50% of the crop — occur annually on theaverage from the disease in Florida. Fortunately, spat and juveniles quicklyreestablish reefs, even after extensive die-
offs.
Dermocystidium marinum is consideredrather specific for the American oyster.However, a few other invertebrates harbor
similar and, in some cases, identical spores,but apparently do not undergo massmortalities. Actually, several related,poorly-understood species infect oysters andother invertebrates, and some of these can
cause mortalities in stressed hosts.
Other Symbionts and DiseasesSeveral other organisms in addition to
dermo-like agents cause disease in oysters.Most necessitate a microscope to see. Anunidentified "mycelial" (suspected fungus,seehypha in glossary) disease infects oystersfrom at least Texas and Louisiana
primarily in spring; infections result insome mortalities, but show no obvious
relationships with salinity.Other diseases of bacterial and fungal
origin also occur. One discussed here is"foot disease." That name is a misnomer
because the oyster's foot, an organ whichaids crawling of the larva before it attaches,becomes rapidly resorbedby the young spataftersetting.Apparentlycausedbya fungus,small rough greenish spots speckle the inside of the shell under the attachment of theadductor muscle. In severe cases, part of themuscle separates from the shell and a hornyelastic cyst forms. As it extends beyond thesite of attachment, the cyst acquires a hardcalcareous encasement. The site of
attachment necessarily changes continuallyto accommodate growth of the oyster andshell. Thought bysome to bemore prevalentin warm muddy waters, foot disease makesan oyster vulnerable to predation becausethe valves no longer close efficiently.
The cyst illustrated in Figure 8 may befoot disease, a rare condition in Mississippi

Figure 8. A hard horny growth secreted bythe oyster. This may exemplify "footdisease," a reaction by the oyster against afungus under the attachment of the adductormuscle. Low quality pearls form when theoyster secretes a protective coating aroundan irritant.
Sound, or some other similar condition. A
variety of hard growths can develop in anoyster. Pearls form when the oyster secretesa protective calcareous coating around anirritant such as a sand grain or a larval parasite, but these rarely have the qualityessential for good jewelry. "Mud pearls"may also be caused by penetration ofmud worms at the site where the adductor
muscle attaches.
Consumers occasionally worry abouteating pink oysters. This condition typically develops in oysters refrigerated for afew days. A yeast can cause this, and it is destroyed with even minimal cooking. Otherdiscolorations occur in live oysters. The adductor muscle of an oyster that has fed on aspecific diatom (a very small alga) "bleeds"a reddish pigment when cut. Certaindinoflagellates (other small algal forms)and heavy metals also cause discolorationin various tissues.
A microscope allows detection of spores*of a gregarine protozoan. Two similarspecies, one infecting mostly mantle tissueand the other infecting gill tissue, occurcommonly in Mississippi. Within the sporesare worm-like protozoans, and if eaten bycertain small crabs, the life cycle iscompleted. Some inconclusive experimental evidence suggests that the gregarinespores cause mortality in oysters. However,
American Oyster 9since the spores do not undergo reproduction like spores of Dermocystidiummarinum, they are not numerous, and theycause little pathological change in theoyster, the likelihood is greater for otherfactors being involved in the mortalities.
A bucephalid digenean (flatworm) alsoinfects oysters in the northern Gulf ofMexico. Some infected individuals gape,suggesting harm caused by the fluke or anassociated stress. Most infected oysters,however, become castrated, and their full
ness and tastiness often improves like that ofa gelded domestic animal. Theimprovement becomes noteworthy duringthe summer when the condition of
uninfected oysters diminishes because ofspawning activity. Even though eatingoysters harboring the larval stages of thisfluke may be esthetically displeasing, theconsumer cannot be harmed. The larvae
only infect specific fishes which in turnmust be consumed by other fishes to mature.Man digests both larval and adult stages.Life cycles of other digenetic flukes, ortrematodes as they are often called, will bediscussed in detail later.
Any oyster in the Gulf can occasionallyharbor a tapeworm larva, one or two larvalroundworms, or a variety of amoeboid,ciliated, or flagellated protozoans. Whenthe oyster undergoes stress, some of theprotozoans reproduce extensively andbecome implicated in disease. Quantification of the role of those protozoans inoyster disease and mortality must awaitfurther investigation.
A few larger symbionts, visible to thenaked eye, inhabit the oyster, but seldomharm it by themselves. Those most likelyto arouse curiosity are a small snail, peacrabs, and turbellarian flatworms.
A 5 millimeter (mm) long (see glossaryunder measurements), whitish snail(commonly known as the impressedodostome) congregates in numbers sometimes greater than 100 at the edge of anoyster shell. Reaching over the edge, a snailintermittently protrudes its proboscis into
10 Marine Maladies?
the oyster's mantle to feed on mucus andtissues. Not specific to the oyster, butapparently preferring it, the little gastropod also feeds on other molluscs and onpolychaete worms. It is part of the highsalinity oyster reef community*. Youngersnails replace the older ones in latesummer, more often inhabiting olderoysters.
Pea crabs associate with a variety ofinvertebrates. One species, popularlyreferred to as the oyster crab, commonlyinvades oysters in New England and onoccasion inhabits individuals along thenorthern Gulf, especially from high salinityreefs such as those off Pass Christian,
Mississippi. Small crabs less than 1 mmwide enter an oyster, usually settling onthe gill surface. Spat and year-old oystersappear to attract more crabs, even thoughthe crab survives best in year-old and olderoysters. The pinkish female crab growsalong with the oyster until it reaches over1 cm wide. In contrast, the male remainssmall and even has a hard stage whichallows it to leave one oyster to mate withfemales in others. Usually one or two females occupy an oyster, but many can betolerated.
Not feeding directly on the oyster, thecrab nevertheless irritates and erodes the
host's gills. To feed, it traps mucus and foodmasses in its walking legs, or it picks stringsof food directly off the host's gills with itspinchers. Young crabs probably filter foodfrom the water.
From a gourmet's standpoint, anydiminishment caused in an oyster'scondition is compensated for by theaddition of a tasty morsel. In New England,pea crabs get eaten both raw and cooked. Ifcooked, the bite-sized morsels can be
sauteed or deep-fat fried along with hermitcrabs and small fish.
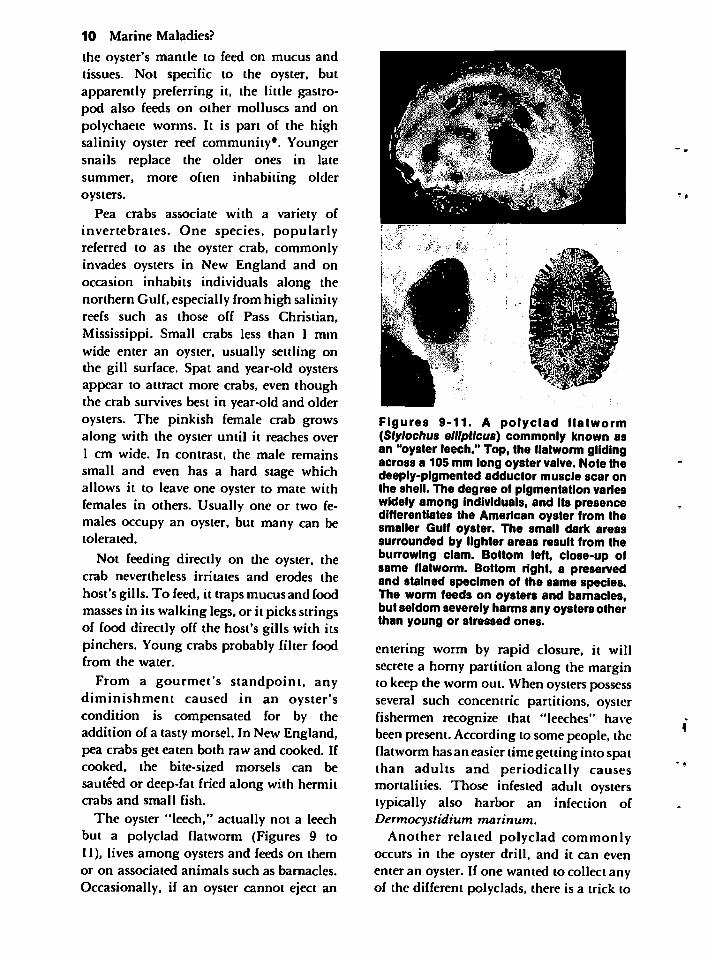
The oyster "leech," actually not a leechbut a polyclad flatworm (Figures 9 to11), lives among oysters and feeds on themor on associated animals such as barnacles.
Occasionally, if an oyster cannot eject an
Figures 9-11. A polyclad flatworm(Stylochus ellipiicus) commonly known asan "oyster leech." Top, the flatworm glidingacross a 105 mm long oyster valve. Note thedeeply-pigmented adductor muscle scar onthe shell. The degree of pigmentation varieswidely among individuals, and its presencedifferentiates the American oyster from thesmaller Gulf oyster. The small dark areassurrounded by lighter areas result from theburrowing clam. Bottom left, close-up ofsame flatworm. Bottom right, a preservedand stained specimen of the same species.The worm feeds on oysters and barnacles,but seldom severely harms any oysters otherthan young or stressed ones.
entering worm by rapid closure, it willsecrete a horny partition along the marginto keep the worm out. When oysters possessseveral such concentric partitions, oysterfishermen recognize that "leeches" havebeen present. According to some people, theflatworm has an easier time getting into spatthan adults and periodically causesmortalities. Those infested adult oysterstypically also harbor an infection ofDermocystidium marinum.
Another related polyclad commonlyoccurs in the oyster drill, and it can evenenter an oyster. If one wanted to collect anyof the different polyclads, there is a trick to

it. An easy way is to place oysters or crusheddrills in a bucket of water from the mollusc'shabitat and leave them for a day or so. Oncethe water fouls and loses its oxygen to decomposing hosts, the flatworms will crawlabout on the side of the bucket and can be
readily seen. Without a concertedcollectingeffort, many worms may be overlooked.Indeed, most fall off or dry up and are neverseen by those who relish oysters-on-the-half-shell. An organic solvent such as xylene willcause these and many other hiddensymbionts to leave their host, but use ofsuch methods will kill the organisms.
With a dish of specimens plus somehealthy hosts, some enterprising studentshould be able to design an experiment tostudy feeding and pathological effects onthe hosts, to investigate survival orreproductive capabilities of the wormsunder different conditions, and to win a
prize in a science fair. All species usuallyinhabit the mantle cavity of their hosts, butI have observed specimens inside a drill'sshell near the top of the spire. What are theydoing there? One species in a hermit crab'sshell eats the crab's eggs, but, duringother seasons, helps maintain the innersurface of the shell by feeding on foulingorganisms.
Shell-Burrowing SymbiontsA few associations between oyster and
symbiont involve species that burrow intothe shell. Even though confusion existsconcerning definitions, here I consider aburrowing animal one that excavates aspace for the purpose of living all or a partof its life. In contrast, a borer is one which
forms a hole in order to obtain food or
inject a substance.One burrower on high salinity reefs in
Mississippi (Diplothyra smithii)commonly goes by the name burrowing (orboring) clam (Figures 12 to 14). The foot ofa young individual secretes an etching orsoftening substance, allowing the serratedmargin of the clam's shell to rasp and scrapea tunnel. The clam does not burrow inde-
American Oyster 11
Figures 12-14. The burrowing clam(Diplothyra smlthli) in the oyster. Top,oysters show the diagnostic small roundholes of the clam as well as the raised siphon-chimneys that protect the burrow from silt.Specimens of the hooked mussel occur onboth margins of the middle oyster; the outercentral portion of that oyster's valve becameweakened and broke off because of clamburrows. Middle, valve has been purposefully broken to expose clam. The smallirregular holes are sponge burrows. Theconspicuous network of pits at the margin isactually an encrusting bryozoan. Bottom,adult burrowing clams removed from theirfinal excavation site.
finitely, only excavating as a juvenile.During summer months, spawning occursand the larval stage with its developed footfinds and penetrates into an oyster or, less
12 Marine Maladies?
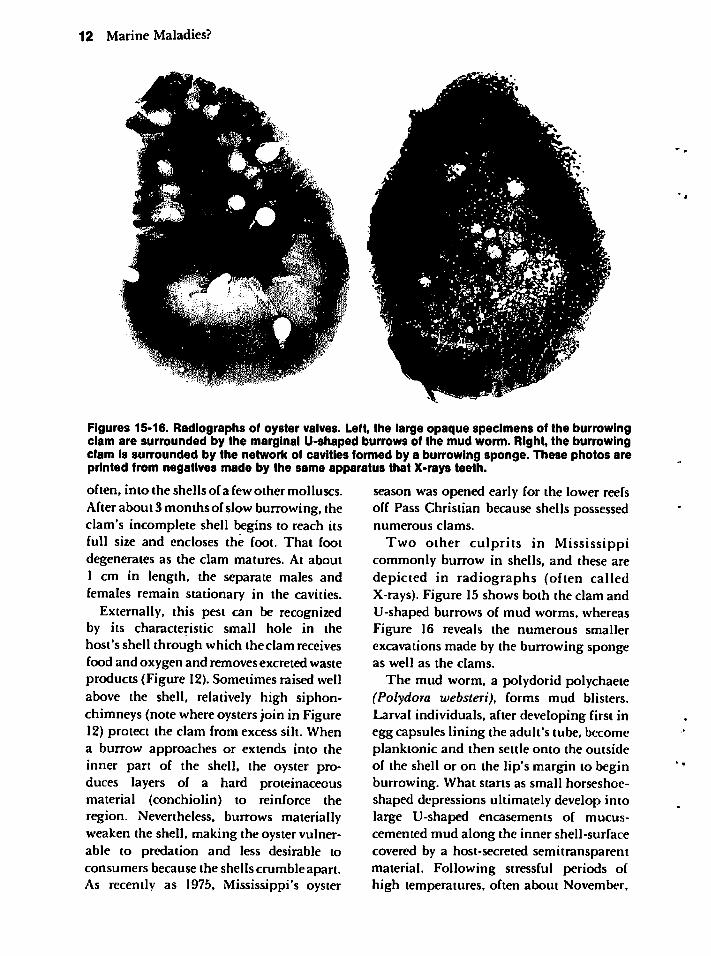
Figures 15-16. Radiographs of oyster valves. Left, the large opaque specimens of the burrowingclam are surrounded by the marginal U-shaped burrows of the mud worm. Right, the burrowingclam is surrounded by the network of cavities formed by a burrowing sponge. These photos areprinted from negatives made by the same apparatus that X-rays teeth.
often, into the shells ofa few other molluscs.
After about 3 months of slow burrowing, theclam's incomplete shell begins to reach itsfull size and encloses the foot. That foot
degenerates as the clam matures. At about1 cm in length, the separate males andfemales remain stationary in the cavities.
Externally, this pest can be recognizedby its characteristic small hole in thehost's shell through which the clam receivesfood and oxygen and removes excreted wasteproducts (Figure 12).Sometimes raised wellabove the shell, relatively high siphon-chimneys (note where oysters join in Figure12) protect the clam from excess silt. Whena burrow approaches or extends into theinner part of the shell, the oyster produces layers of a hard proteinaceousmaterial (conchiolin) to reinforce theregion. Nevertheless, burrows materiallyweaken the shell, making the oyster vulnerable to predation and less desirable toconsumers because the shells crumble apart.As recently as 1975, Mississippi's oyster
season was opened early for the lower reefsoff Pass Christian because shells possessednumerous clams.
Two other culprits in Mississippicommonly burrow in shells, and these aredepicted in radiographs (often calledX-rays). Figure 15 shows both the clam andU-shaped burrows of mud worms, whereasFigure 16 reveals the numerous smallerexcavations made by the burrowing spongeas well as the clams.
The mud worm, a polydorid polychaete(Polydora websteri), forms mud blisters.Larval individuals, after developing first inegg capsules lining the adult's tube, becomeplanktonic and then settle onto the outsideof the shell or on the lip's margin to beginburrowing. What starts as small horseshoe-shaped depressions ultimately develop intolarge U-shaped encasements of mucus-cemented mud along the inner shell-surfacecovered by a host-secreted semitransparentmaterial. Following stressful periods ofhigh temperatures, often about November,
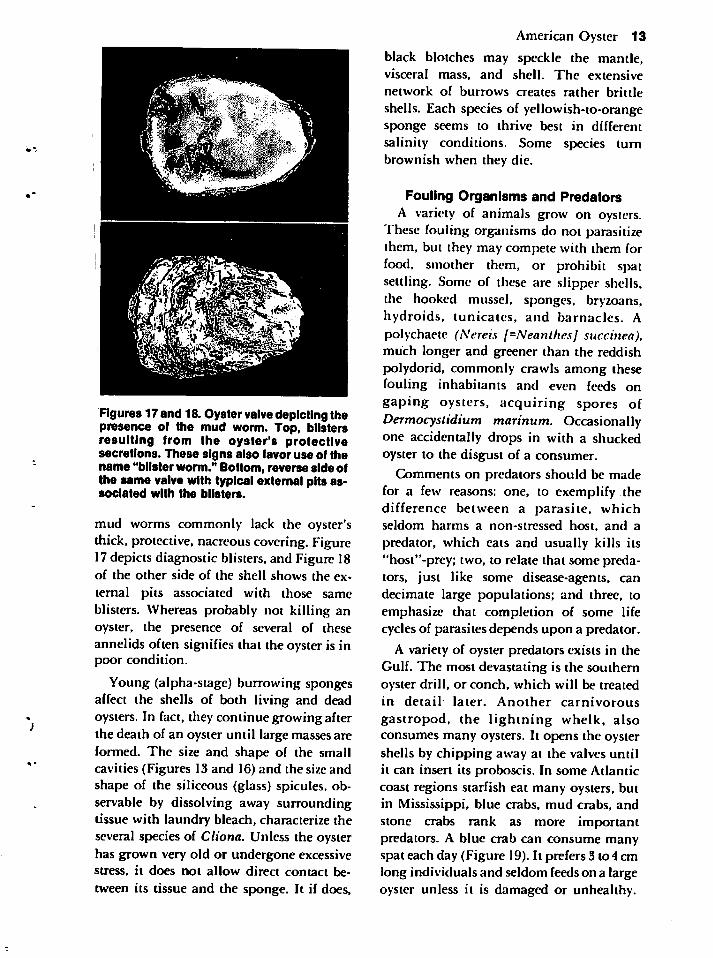
Figures 17 and 18. Oyster valve depicting thepresence of the mud worm. Top, blistersresulting from the oyster's protectivesecretions. These signs also favor use of thename "blister worm." Bottom, reverse side ofthe same valve with typical external pits associated with the blisters.
mud worms commonly lack the oyster'sthick, protective, nacreouscovering. Figure17depicts diagnostic blisters, and Figure 18of the other side of the shell shows the ex
ternal pits associated with those sameblisters. Whereas probably not killing anoyster, the presence of several of theseannelids often signifies that the oyster is inpoor condition.
Young (alpha-stage) burrowing spongesaffect the shells of both living and deadoysters. In fact, they continue growing afterthe death of an oyster until large masses areformed. The size and shape of the smallcavities (Figures 13 and 16) and the size andshape of the siliceous (glass) spicules, observable by dissolving away surroundingtissue with laundry bleach, characterize theseveral species of Cliona. Unless the oysterhas grown very old or undergone excessivestress, it does not allow direct contact be
tween its tissue and the sponge. It if does,
American Oyster 13
black blotches may speckle the mantle,visceral mass, and shell. The extensive
network of burrows creates rather brittle
shells. Each species of yellowish-to-orangesponge seems to thrive best in differentsalinity conditions. Some species turnbrownish when they die.
Fouling Organisms and PredatorsA variety of animals grow on oysters.
These fouling organisms do not parasitizethem, but they may compete with them forfood, smother them, or prohibit spatsettling. Some of these are slipper shells,the hooked mussel, sponges, bryzoans,hydroids, tunicates, and barnacles. Apolychaete (Nereis [=Neanthes] succinea),much longer and greener than the reddishpolydorid, commonly crawls among thesefouling inhabitants and even feeds ongaping oysters, acquiring spores ofDermocystidium marinum. Occasionallyone accidentally drops in with a shuckedoyster to the disgust of a consumer.
Comments on predators should be madefor a few reasons: one, to exemplify thedifference between a parasite, whichseldom harms a non-stressed host, and a
predator, which eats and usually kills its"host"-prcy; two, to relate that some predators, just like some disease-agents, candecimate large populations; and three, toemphasize that completion of some lifecycles of parasites depends upon a predator.
A variety of oyster predators exists in theGulf. The most devastating is the southernoyster drill, or conch, which will be treatedin detail- later. Another carnivorous
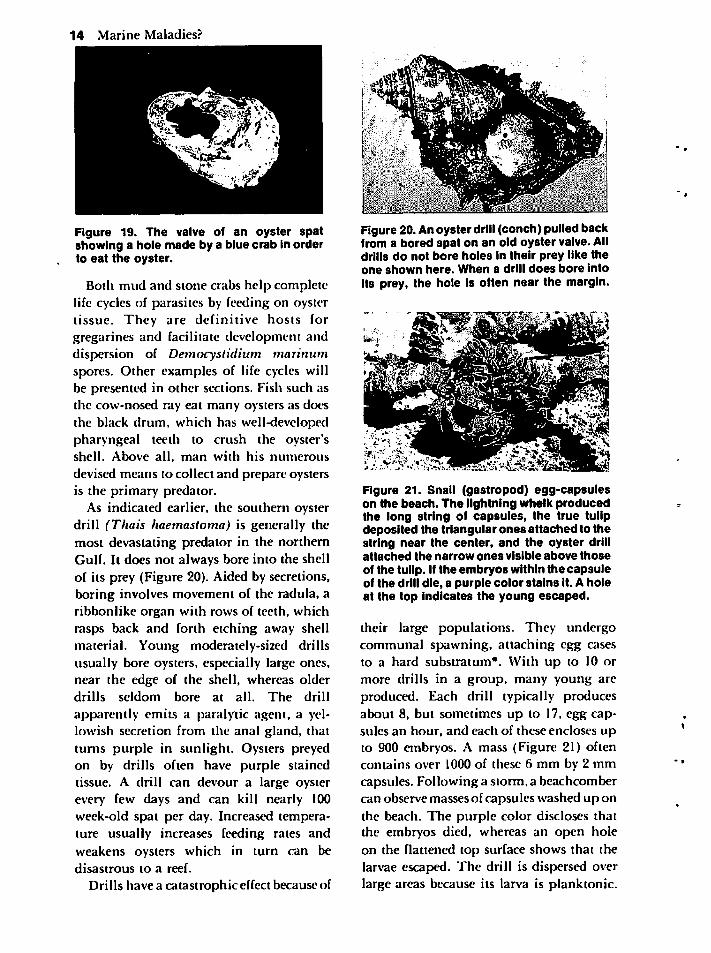
gastropod, the lightning whelk, alsoconsumes many oysters. It opens the oystershells by chipping away at the valves untilit can insert its proboscis. In some Atlanticcoast regions starfish eat many oysters, butin Mississippi, blue crabs, mud crabs, andstone crabs rank as more importantpredators. A blue crab can consume manyspat each day (Figure 19). It prefers 3 to4 cmlong individuals and seldom feeds on a largeoyster unless it is damaged or unhealthy.
14 Marine Maladies?
Figure 19. The valve of an oyster spatshowing a hole made by a blue crab in orderto eat the oyster.
Both mud and stone crabs help completelife cycles of parasites by feeding on oystertissue. They are definitive hosts forgregarines and facilitate development anddispersion of Democystidium marinumspores. Other examples of life cycles willbe presented in other sections. Fish such asthe cow-nosed ray eat many oysters as doesthe black drum, which has well-developedpharyngeal teeth to crush the oyster'sshell. Above all, man with his numerous
devised means to collect and prepare oystersis the primary predator.
As indicated earlier, the southern oysterdrill (Thais haemastoma) is generally themost devastating predator in the northernGulf. It does not always bore into the shellof its prey (Figure 20). Aided by secretions,boring involves movement of the radula, aribbonlike organ with rows of teeth, whichrasps back and forth etching away shellmaterial. Young moderately-sized drillsusually bore oysters, especially large ones,near the edge of the shell, whereas olderdrills seldom bore at all. The drill
apparently emits a paralytic agent, a yellowish secretion from the anal gland, thatturns purple in sunlight. Oysters preyedon by drills often have purple stainedtissue. A drill can devour a large oysterevery few days and can kill nearly 100week-old spat per day. Increased temperature usually increases feeding rates and
weakens oysters which in turn can bedisastrous to a reef.
Drills have a catastrophic effect because of
Figure 20.Anoyster drill (conch) pulled backfrom a bored spat on an old oyster valve. Alldrills do not bore holes in their prey like theone shown here. When a drill does bore intoits prey, the hole is often near the margin.
Figure 21. Snail (gastropod) egg-capsuleson the beach. The lightning whelk producedthe long string of capsules, the true tulipdeposited the triangular ones attached to thestring near the center, and the oyster drillattached the narrow ones visible above thoseof the tulip. If the embryos within the capsuleof the drill die, a purple color stains it. A holeat the top indicates the young escaped.
their large populations. They undergocommunal spawning, attaching egg casesto a hard substratum*. With up to 10 ormore drills in a group, many young areproduced. Each drill typically producesabout 8, but sometimes up to 17, egg capsules an hour, and each of these encloses upto 900 embryos. A mass (Figure 21) oftencontains over 1000 of these 6 mm by 2 mmcapsules. Following a storm, a beachcombercan observe masses of capsules washed up onthe beach. The purple color discloses thatthe embryos died, whereas an open holeon the flattened top surface shows that thelarvae escaped. The drill is dispersed overlarge areas because its larva is planktonic.

THE BLUE CRAB
As the most economically important crabfrom the Gulf and Atlantic states, the blue
crab (Callinectes sapidus) is caught primarily in chicken wire "crab pots" andtrawls by professional crabbers (Figure 22)and in drop nets or on a piece of twine bysports fishermen. Rather than beingattracted to poultry parts, pork scraps, orfish using the above means, the soft-shelledstage of the crab does not feed and can becollected with a dip net or in an artificialshelter. This special gourmet's delight remains in the soft stage for several hoursfollowing molting. Molting will bedescribed later, but of importance here tothose who have never eaten soft-shelled
crabs is that the entire crab minus gills,"apron*," and a narrow anterior portionprovides a rich broiled, fried, or barbequeddelicacy.
In most of the northern Gulf of Mexico,
the adult crab may spawn once or twiceduring the 9 months from spring throughfall as opposed to once in Chesapeake Baywhere the crab concentrates spawning toabout a 2 month period. Spawning occurs inhigh salinity regions, often near barrierislands in the Gulf. If not in high salinitywater, the first larval stage (nauplius) heldby the female and the next larval stage (zoea)die or do not develop properly. After developing in a highly saline habitat, the larvametamorphoses (into a megalops), migratesinto estuaries, and metamorphoses again (toan early crab stage). Juveniles thrive in theestuaries; they commonly inhabit the softmuddy bottoms of navigational channelsand marsh regions. They mature within a
year, at which time a male typicallyfertilizes and protects the female during herfinal molt. Insemination occurs once in her
life, even though the stored sperm mayproduce several "sponges" of eggs. Unlikethe female, the male typically remains inthe estuary and molts a few more times, insome cases for as long as 5 years. The adventof dropping temperatures, usually inconjunction with lowered salinities brought
Blue Crab 15
Figure 22. Commercial crabber and his sonemptying catch from crab pot into sortingtray.
on by winter northerly winds, usuallysignals females to leave the estuary. Atypicalconditions like the extended high salinitiesin Lake Borgne, Louisiana, during late1976 and early 1977 inhibited the migration,thereby causing a decline in the winter'scrab catch in Mississippi Sound.
I presented the above features of the crab'slife history because they dictate the kindsand amounts of symbionts that affect a crab.Molting represents an example. When acrab sheds its molt, it sheds manyectosvmbionts. Discussion of some
symbionts follows, not all of which areshed by molting.
Protozoans and Microbes
As true parasites, microsporidan protozoans infect a variety of host cells. Of atleast five different species infecting theblue crab, one (/Im<?.von michaelis) deservesspecial consideration because it harms themost crabs and its life cycle is the only onefrom the blue crab that has been established.
Because it is an internal infection, it is
not lost by molting, but it affects die crab'sbehavior.
"Sick crabs," the term used by somefishermen to designate individuals diseasedwith Ameson michaelis, occur commonly ina variety of habitats from Chesapeake Baythrough Louisiana. Heavy infections can be
recognized easily because muscle tissueacquires a chalky appearance in joints of

16 Marine Maladies?
Figure 23. Life cycle of a microsporidan (Ameson michaelis) in the blue crab. The cycle of this protozoan is direct; there is no true intermediate host. When a blue crab feeds on infected crab meator spores from another source, the long internal tubular filament of the spore turns inside out as itprotrudes, allowing the Infective nucleus and associated material to pass through it and infect ahost blood cell. Divisions of the microsporidan within the blood cell produce a continuous supplyof the parasite that reaches muscle tissue where there forms a string of developing spores. As aspore approaches maturity, there are first two Joined cells and then a single mature spore. Crabmeat infected with spores turns opaque and chalky. The muscle tissue surrounding the masses ofspores breaks down, and the crab becomes weakened and vulnerable to stress.
the appendages and the abdomen oftenturns grayish. Even though virtually absentfrom Mississippi Sound until spring of1978, sick crabs typically inhabit nearbyLake Pontchartrain and Lake Borgne inLouisiana. Generally less than 1% of thecrabs there show infections, but occasionally more than this number linger in warmlagoons and close to shore. Perhaps thisdisease has a great impact on young crabs. Inour studies, we have experimentally infectedboth young and old individuals withoutdifficulty. Naturally-infected crabs asnarrow as 2 cm across their carapace* havebeen seen, but small crabs avoid most
routine collecting procedures by people
who would recognize infections. Naturallyinfected crabs often fail to undergo migrations with noninfected counterparts, andthey tend to die more readily when stressed.
Before learning about the life history ofthe infectious agent, fishermen wouldsmash infected crabs and return them to the
water, allowing noninfected blue crabs tofeed on the tissue and acquire the disease.Education therefore helped the fishermen ina way similar to the situation when oysterfishermen from Chesapeake Bay and fromPuget Sound, Washington, learned that foreach starfish arm ripped off with a centralportion, another predator starfish regenerated to feed on their crop.

Figure 24. Highly magnified view of microsporidan spores (Ameson michaelis) amongmuscle fibers of blue crab.
A diagram (Figure 23) reveals anextremely magnified representation ofstages in the cycle. A mature spore (Figure24) measures about one-fourth that of a
human red blood cell (1.9 microns[;i] versus7.6 pi). Under appropriate conditions, aningested spore everts its long internaltube (polar filament), an action roughlyanalogous to blowing out the invertedfinger of a rubber glove. The nucleus andcytoplasm of this single celled animalsqueezes through the extruded pliablefilament and invades a host crab's blood
cell. Once there, the organism undergoesvegetative growth, producing cells which inturn produce strings of eight individualspores along muscle tissue. Ultimately,enormous numbers of these resistant sporesand their products destroy adjacent tissueand replace the crab's normal musculature.This parasite can be considered highly host-specific* because the muscle tissue of onlyCallinectes sapidus exhibits infections.Many symbionts, like the bacteriamentioned in the next paragraph, have awide, or loose, host-specificity* because theyassociate with a wide range of hosts.
Shell disease ("box burnt" crabs) represents a typically non-fatal microbialcondition usually initiated by a wound. Inany event, infected crabs usually have notmolted for a long period and have undergone stress. Starting as brownish areas withreddish brown depressed centers forming onthe exoskelton, these lesions develop intodeep necrotic* pits (Figure 25) that typicallydo not penetrate through the shell. Once the
Blue Crab 17
Figure 25. Claw of blue crab with shell-disease. This brownish lesion withdegenerated internal regions apparentlyresults from a bacterium (Benekla type I)which digests the chitin from the exo-skeleton. An infection is rarely fatal and islost during molting of the host.
thin external lipid (fat) layer of the shell(epicuticle) has been disrupted, the exposedchitin* becomes digested by specificbacteria and possibly also fungi. Probablyfewer crabs than lobsters die from the
disease, especially if lobsters are culturedin warm water. Death probably results fromsecondary infections. Nevertheless, crabslook esthetically displeasing, and confinement fosters disease. Hence, some crabbers
know the disease as "box burnt" because
they maintain crabs in floating cypress-boxcages waiting for their catch to molt.
Certain other bacteria harm both crabs
and people, when given the properconditions. One such organism(Vibrio parahaemolyticus) readily killscrabs and causes food poisoning in people.In crabs, species including Vibrioparahaemolyticus cause large jelly-likeblood clots in addition to whitish-colored
nodules to develop in the gills and elsewherefrom masses of aggregated blood cells andbacteria. Crabs from Mississippi watersoccasionally die from these infections. Inpeople, numerous cases of intestinal upsetassociated with eating seafood probablyalso result from this organism. Evenminimal heating, however, eliminates thisthreat. To avoid food poisoning in crab-preparing operations, continued careshould be taken not to permit fluid fromfresh crabs to contaminate previously-

18 Marine Maladies?
boiled crabs. Additonal difficult-to-
diagnose microbial diseases of crabs willappear following the section on metazoansand their hyperparasites*. Metazoa, not anatural group of organisms, designates thatan included organism is multicellular withtwo or more tissue layers.
Metazoans and their HyperparasitesLarval flukes (digeneans), unlike larval
tapeworms (cestodes) (Figures 26 and 27),infect many freshwater, estuarine, and marine crabs. Microphallids form a family ofdigeneans that utilize three hosts: l) a snailas a first intermediate host*, 2) a crustacean
as a second intermediate host, and 3) a bird,
mammal, or rarely a cold blooded vertebrateas a definitive host. At least six species infect
the blue crab. We will mention two, the first
(Levinseniella capitanea) because it can beseen easily among the gonads or thediverticula of the digestive gland if thecrab's carapace* is removed. Nearly 4 mmlong, the excysted worm ranks as the largest
Figures 26 and 27. Two views of same larvaltapeworm moving through muscle of lesserblue crab (Callinectes slmllls). Althoughapparently common in this crab, it has notbeen observed in the common blue crab
(Callinectes sapidus).
microphallid known. Most other speciesaverage less than 1 mm, appearing somewhat similar to a comma in diis booklet in
both size and shape. Also very unusual.Levinseniella capitanea has no gut ormuscular pharynx, principal features formost larval microphallids and used byalmost all adult digeneans for feeding. Thegut does not develop even after the larva inits spherical, yellowish cyst has been eatenby and matures in a raccoon.
The second species (Microphallusbasodactylophallus) can barely be seen incrabmeat (striated muscle tissue) without a
microscope, unless a urosporidan protozoanhas hyperparasitized it. It measures about0.45 mm (450 microns) long after removalfrom the spherical 0.35 mm cyst (about thesize of the following period).
Figure 28 illustrates the life history ofthis digenean. Any of four species of snailsfrom the shallow low salinity estuaries* mayrelease the infective free swimming larva
(cercaria). That free-living larva was produced in quantity within another asexualstage (sporocyst) which was itself producedby a similar larva that in turn haddeveloped from a germ ball within theinitial ciliated larva (miracidium). Theinitial larva occupied the egg which was fedon by the snail. In other words, from oneegg develops an enormous number of larvaeinfective to the blue crab and at least two of
the several species of fiddler crabs. Also, eac hresulting adult worm produces anenormous number of eggs. Because of dependency on environmental parameters andon need for specific hosts, these largenumbers of eggs and larvae are necessary toassure completion of the cycle for one ormore individuals and continued survival of
the species.
Each individual possesses both male andfemale organs, making it hermaphroditic.It can successfully fertilize itself, eventhough all tested species of digeneansprefer to utilize partners, if given theopportunity. These worms mature when anappropriate final host eats the infected crab,
Blue Crab 19
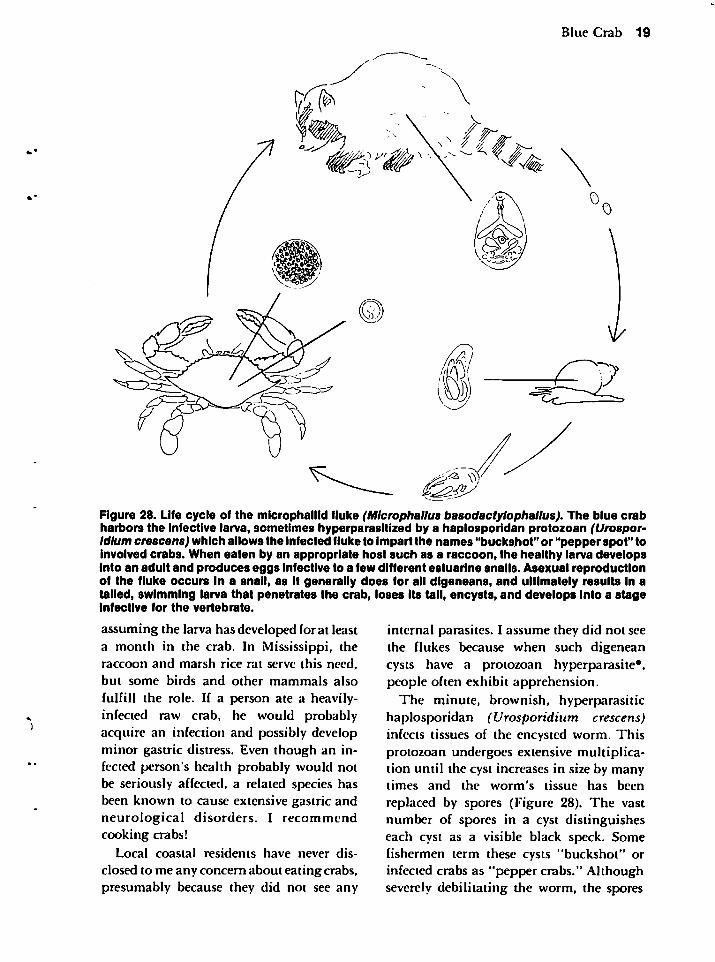
Figure 28. Life cycle of the microphaiiid fluke (Microphallus basodactylophallus). The blue crabharbors the infective larva, sometimes hyperparasitized by a haplosporidan protozoan (Urospor-Idlum crescens) which allows the infected fluke to impart the names "buckshot" or "pepper spot" toinvolved crabs. When eaten by an appropriate host such as a raccoon, the healthy larva developsinto an adult and produces eggs infective to a few different estuarine snails. Asexual reproductionof the fluke occurs in a snail, as it generally does for all digeneans, and ultimately results In atailed, swimming larva that penetrates the crab, loses its tail, encysts, and develops into a stageInfective for the vertebrate.
assuming the larva has developed for at leasta month in the crab. In Mississippi, theraccoon and marsh rice rat serve this need,
but some birds and other mammals also
fulfill the role. If a person ate a heavily-infected raw crab, he would probablyacquire an infection and possibly developminor gastric distress. Even though an infected person's health probably would notbe seriously affected, a related species hasbeen known to cause extensive gastric andneurological disorders. I recommendcooking crabs!
Local coastal residents have never dis
closed to me any concern about eating crabs,presumably because they did not see any
internal parasites. I assume they did not seethe flukes because when such digeneancysts have a protozoan hyperparasite*.people often exhibit apprehension.
The minute, brownish, hyperparasitichaplosporidan (Urosporidium crescens)infects tissues of the encysted worm. Thisprotozoan undergoes extensive multiplication until the cyst increases in size by manytimes and the worm's tissue has been
replaced by spores (Figure 28). The vastnumber of spores in a cyst distinguisheseach cyst as a visible black speck. Somefishermen term these cysts "buckshot" orinfected crabs as "pepper crabs." Althoughseverely debilitating the worm, the spores

20 Marine Maladies?
Figure 29. Buckshot, or pepper spot, in bluecrab meat. Dark, almost spherical objectsnear the pointer and at the top of the meatare fluke larvae (Microphallus basodactylophallus) infected with a protozoan(Urosporldlum crescens). The immensenumber of spores enlarges the fluke cystmany times. The piece of crab has beencooked; the exoskeleton for the swimmingleg occurs in the lower left of the photo.
harm neither the crab nor man. A piece ofcooked crab (Figure 29) reveals a couple ofclusters of buckshot. Other encysteddigenean larvae also harbor this or relatedspecies. Some also possess other kinds ofprotozoan hyperparasites including micro-sporidans, myxosporidans, flagellates, andopalinids.
The blue crab, a crustacean, has its own
crustacean symbionts. Those discussed here,however, appear much different from thecrab except for the early larval stage.They are barnacles and range between beingfouling organisms and true parasites.
One common barnacle (Balanus venustusniveus) found on a varietyof hard substrata*in the Gulf also establishes itself on the
carapace and legs of the blue crab. Another,the acorn barnacle (Chelonibia patula),exhibits more host-specificity*. It restrictsitself to the external surfaces of a small
group of crabs. Figure 30 shows it on theblue crab. Whether lacking or expressingspecificity towards the crab, both encrustedbarnacles cause their host hardship byproducing excess weight which sometimesconstitutes a considerable burden.
A gooseneck, or pedunculate, barnacle(Octolasmis muelleri) confines its presenceto the gill region of a few decapod species.
Figures 30-32. External bamacle symbiontson the blue crab. Top, the acorn barnacle(Chelonibia patula) on the carapace.Middle, a goose-neck barnacle (Octolasmismuelleri) on a gill filament propped up by adissecting pin. Bottom, underside of gillsshowing many medium-sized and smallOctolasmis muelleri. Note how few can beseen without lifting the gills.
including the blue crab (Figure 31). It needsa living crab for survival. Special cleaningappendages sweep gill surfaces freeof debrisand presumably of settling organisms. Oncean individual does become established, the
crab's cleaners apparently become lessefficient. Consequently, increasingamounts of debris and numbers of larvae
can settle on the gills. In fact, a crab canacquire over 1000such barnacles on the gillsand elsewhere in the gill chamber. When

small-and medium-sized barnacles
infest a host, over 700 can coat the underside of the gills without hardly beingapparent on the top surface of the gills(Figure 32).
The combination of these heavy infestations* and debris can make respirationdifficult for the crab because barnaclescompete for oxygen and decrease theamount of gill surface available for respiration. Healthy crabs are hardy; they live forlong periods out of the water when theirgills are moist, but die quickly immersedin stale water. Infested crabs act sluggishmost of the time and probably attract pre
dators.
The last barnacle to be discussed trulyparasitizes its host (Figures 33 to 35). Ifnot for being able to recognize its larvalstages, biologists would not have been ableto classify it as a barnacle. An externalportion of this rhizocephalan (Loxothyla-cus texanus) protrudes from under thecrab'sabdomen (Figure 33). This bulge, calledthe externa, contains a brood- pouch forlarvae and both male and female gonads. Acrab can have as many aseight externae, andthree are fairly common (Figure 35). Theremainder of the barnacle consists of rootlike structures penetrating through mosthost tissues(interna). Crabs becomeinfectedwhen young by a swimming larva (cypris).After a period of internal development, theinterna of the organism penetrates througha crab's soft abdominal joint when the crabmolts. Infection retards a host's gro\vth,leaving most externae-bearing individualsbetween 3 and 8 cm wide. Additionally,secondary sexual characteristics of infectedmales transform into those of females.Therefore, most infected crabs appear asminiature adult females, either with orwithout externae. The size-differencebetween an egg-bearing (sponge or berry)female and an infected crab is usually great.Figure34shows the size-difference betweenan uninfected immature female with heracutely pointed abdominal apron and asmaller infected crab with the transformed
Blue Crab 21
Figures 33-35. Sacculinid barnacle(Loxothytocus texanus) of the blue crab.Top, the protruding pouch (externa) underthe crab's abdomen contains larvae andgonads (ovary and testes). Long, narrowextensions invade most other tissue. Middle,top crabhas dark externa indicatinganolderinfection than that of the yellowish externa ofthe middle.crab. The larger lower crabreveals the small size of infected crabs. It isan immature female, wheras the aprons onthe infected crabs reveal the apron of amature crab. Infections also modify theshape of the male's abdomen into that of afemale. Bottom, crab with three developingexternae. Usually only one externa occursper crab, but occasionally over five may bepresent.
rounded apron normal for mature females.Castration of the crab may result frominvolvement with a structure called anandrogenic gland. Male crabs have this

22 Marine Maladies?
gland, and, if it is removed from a youngindividual, female characteristics, including ovarian tissue in the testes, develop.Perhaps destruction of this gland or someaction on its secretions, possiblyadditionally involving hormones producedby the barnacle's ovary, causes feminizationof the infected crab.
Crabs with externae can molt and losethat part of the parasite, but whether thatprocess is typical is unknown. Most biologists assume crabs with this and othermature rhizocephalans do not molt.
Infections, although prevalent in somehigh salinity bays and bayous in Louisiana,have been negligible until 1977 inMississippi Sound since an epizooticoutbreak in 1965. While aboard a shrimpboat in July 1977,1 noted over half the crabswe trawled in the Sound exhibited
infections. The apparent magnitude ofinfection in the population could be exaggerated because the small crabs would becontinually thrown back by shrimpers forothers to recapture, whereas large oneswould be saved for eating. Nevertheless,infections occurred commonly for severalweeks, and epizootics will undoubtedlyoccur again.
As with other cited barnacles, infectionsrelate directly with salinity and probablywith temperature. In inshore regions withmoderately high salinity water, crabs havea greater potential for infection. Infectionsusually coincide with breedingand maturation peaksof the blue crab, beingmostprevalent about May and October in Louisiana.
Unlike the other barnacles attached to the
crab, the adult rhizocephalan cannottolerate low salinity. Maturing externaeapparently do not protrude, and ones already protruding take on waterand rupture.Low salinity habitats characteristic ofMississippi estuaries certainly do notfavor barnacle infections.
The presence of "dwarf" or "button"crabs concerns some fishermen. Do thesesmall apparently sexually mature individuals that show up mostly during spring
Figure 36. Sacculinid barnacle parasite(Loxothylacus panopael) in the mud crab(Eurypanopeus depressus) fromChincoteague Bay, Virginia. Note differencebetween this species and the one in the bluecrab.
and early summer in Mississippi result froma distinctive genetic stock, fromenvironmental influence, or from aparasitic infection? It seems quite possiblethat these crabs possess prepatent*sacculinid infections, and, indeed, thatproblem needs attention.
Mud crabs, much smaller crabs than theblue crab, also possess a related rhizocephalan barnacle, and the externa of that species(Loxothylacuspanopaei)differs(Figure 36).
An understanding of molting aidsinterpretation of barnacle-crab associationsand other symbiotic relationships for crustaceans. Like most crustaceans, the blue crabhas a hard external skeleton also knownas a cuticle* or shell. In order to increase
in size, the crab must shed the old shell.When initiating this molting process(ecdysis), a new soft shell forms beneaththe hard one of this "peeler" stage.Enzymes begin to dissolve the shell, allowing the products to be resorbed into thebody-proper. Thus, amino acids, calcium,and other products can be salvaged forincorporation in the new cuticle. Theposterior part of the old shell then cracks,allowing the "buster" stage crab to wiggleout backwards. This amazing process iscontrolled by a balance of separately-produced molt-inhibiting and molt-

Figures 37 and 38. Blue crab molting. Top,molt, or ecdysis, including covering of gillsand lining of fore- and hindgut. Crabs thatback out of shells like this remain extremelysoft and pliable for about 4 or 5 hours.Bottom, soft-shelled crab. Note pliablenature of the twisted claw and depressedcarapace.
stimulating hormones. The results are thediscarded molt (Figure 37) and the helplesssoft-shelled crab enlarged by 20 to 60%of itsprevious size (Figure 38). The crab imbibesand otherwise incorporates water while stillsoft in order to achieve the expanded size.After about 5 hours, unless retarded or
halted in a refrigerator, the new shellhardens because the protein becomes tannedand calcium chloride is redeposited. Eventhough not as soft, this "paper-shell," or"buckram" crab will remain vulnerable to
predators for another day or so.As mentioned earlier, following mating, a
female rarely molts again. Consequently,the shell of a mature female offers a primesubstratum* on which barnacles and other
fouling and symbiotic organisms canattach. In addition to being able to shedsome symbionts by molting, the maturemale usually inhabits low salinity habitatsnot conducive for barnacles.
Blue Crab 23
Figure 39. Long orange ribbonworm(Carclnonemertes carclnophlla) pulled outfrom between gill lamellae. Other lightcolored regions on gills are additionalspecimens. The orange color indicates thatthe worm fed on the crab-host's eggs; thus,the crab had spawned. Also, note thebarnacles on the gills. Both the ribbonwormand the barnacle indicate that the crab hadinhabited high salinity water.
A ribbonworm (nemertean) (Carcinone-mertes carcinophila) approximately 1.5 cmin length often inhabits the space betweengill lamellae in crabs from high salinitywater (Figure 39). Since, in the northernGulf, most males do not accompany femalesmigrating to salty regions in order to spawn,fewer male than female crabs have
nemertean infestations* (see infection inglossary). This higher prevalence forfemales also acts in the best interest of the
nemertean because in order for the worm to
achieve sexual maturity and reproduce, itmust migrate from the gills to the egg massof the host. While there, it builds a mucus
tube for inhabitation. Crab eggs providenourishment for the worm, transforming
the worm's color from creamy to orange.Because of the large quantity of yolk material, crab eggs appear orange during thefirst 5 days of the larva's 14-daydevelopmental period. Since a crab can produce over 2million eggs and may generate those twicein a year, even a heavy infestation by thenemertean has little effect on crab pro
duction.
Some time after mating, the female wormleaves her collapsed tube with its attachedeggs. The resulting ciliated larva hatchesand stays in the tube, swims among the crab

24 Marine Maladies?
eggs, or leaves the host and undergoesmetamorphosis into a creeping stage thatinfests another crab. The blue crab is the
common host, but other portunids(swimming crabs) and occasionallynonrelated crabs harbor the worm. Because
of an attraction by both the nemertean larvaand the crab toward light, the two oftencome into contact and infestation occurs
readily.
As indicated above, worms change colorafter feeding on crab eggs. Consequently,since adults return to the host's gills, thepresence of orange worms encased betweenlamellae reveals that such a host crab has
spawned at least once. The worm is a biological indicator! Strangely though, thesetags lose some of their adult characteristicsand may not feed at all during the winter.
A leech (Myzobdella lugubris), unlike thenemertean and barnacles, conspicuouslyinhabits crabs in low salinity regions(Figures 40 and 41). Even though a case ofnumerous leeches associated with a few
dying crabs implicates them in harming thecrab, I doubt the leech causes lesions in
the crab. The leech often occurs on consider
able numbers of crabs in Mississippi andadjacent areas, but I have seen no evidencesuggesting a harmful relationship.
After seeing Humphrey Bogart coveredwith leeches in the African Queen andhearing tales of people having their bloodsucked while walking through tropical rainforests or wading through swamps, peoplehave a tendency to be disgusted by theintriguing jawless leeches that thrive in ourlocal fresh-to-marine habitats. Perhaps thethought of losing blood intensifies whenpeople realize that in 1863, doctors used 7million medicinal leeches (Hirudomedicinalis) on their patients in Londonhospitals and almost that same number inParis. The practice of bloodletting inEurope declined soon after, but occasionallysomeone will still pull one of these from aculturing jar to remove blood from a bruiseand alleviate swelling.
Even though many leeches do not suck
•••:i;;^\!0?"5:'
mm nfl*B '-.vat: iiii. ''fe-^;Wiiw
Figures 40-43. Leech (Myzobdella lugubris)on blue crab. Top, several specimens of theleech on the crab's anterior. Middle, severalspecimens on crab's posterior; mostindividuals deposit cocoons near the rearmargin. Bottom left, cocoons on carapace.Bottom right, enlarged cocoon. A singleyoung leech emerges from each cocoon; itdevelops in low salinity water and searchesfor a fish in order to obtain a blood meal.
blood, the crab leech does. Soon after hatch
ing, it obtains its first blood meal from afish. In Mississippi, a specimen of thesouthern flounder, striped mullet, or Gulfkillifish may have several individuals attached to its gills, opercles, nostrils, fins,or body surface. Other estuarine fish mayalso possess this or other species of leeches,but usually in lesser numbers. The whitecatfish acts as a good host in regions whereit enters brackish habitats. After receiving anadequate amount of blood, the engorgedleech transforms from a skinny reddishworm about I cm long to a robust greenish-brown worm which can stretch a distance

greater than 4 cm.The engorged crab leech typically
associates with vegetation and throughoutits life shows some ability to swim. Sincethe blue crab also associates with vegetation,the two come in contact. The leech then de
posits adhesive cocoons near the rear marginof the crab's carapace (Figures 42 and 43).Often many leeches infest one crab. Based onobservations of leeches attaching cocoons tofinger bowls, some individuals probably attach their cocoons on other hard surfaces,
also. In addition to providing a substratum*for cocoons, the crab offers a means of
dispersing the leeches.A single young leech develops within
each cocoon. It can swim after emerging,and, like its parent, prefers water withlittle or no salt; neither stage tolerates muchmore salt than 15 ppt. The young leech mustinfest a fish to continue its cycle.
A leechlike organism also infests the bluecrab in low salinity and freshwater habitatsof Alabama, Mississippi, and Louisiana.This branchiobdellid (Cambrincolamesochoreus) shows phylogenetic* affinities to both leeches and oligochaetes (thelatter being best known for earthworms andaquatic worms used to feed tropical fishes).About 2 to 3 mm long, this pink coloredworm infests both the gill chamber and theexternal shell-surfaces without causing itshost apparent harm. It feeds on small protozoans, algae, and detritus, and its presencesuggests that the host crab has been in freshor nearly fresh water for some time.
Miscellaneous Microbes and Protozoans
The crab harbors many more organismsthan those already discussed. Most necessitate a microscope to observe individually, but a few can be seen when present inlarge numbers or when the crab shows signsindicative of specific infections. An electronmicroscope is needed to confirm the presence of some organisms. I will brieflycomment on some of these microscopically-sized organisms, none of which has beenimplicated in human disease.
Blue Crab 25
At least four different viruses infect the
crab. Figures 44 and 45 provide an ultra-structural view of one of these. Until
recently, few viruses had been reported fromany invertebrate. Of the four in the crab,three cause fatalities when the hosts are keptcaptive. They occur in crabs along the eastern U.S. coast, and, if examined for in the
Gulf, some would probably be found. Littlehas been learned about any of them or theirrelationships with the crab host.
A severely pathogenic amoeba(Paramoeba pernicosa) causes mortalitiesin the blue crab from high salinity habitatsalong the eastern U.S. coast. It is treatedhere because of the potential for it to beintroduced or to be present already, butunrecognized, in the Gulf. The causativeamoeba occurs in connective tissues and
blood (hemal) spaces throughout the infected crab's body during all times of the year,but enters the blood and kills the host
primarily during summer. When amoebaeenter the blood, the ventral portion ofthe crab, especially a male, becomes grayish in color, and, if the disease advances tothis state, the crab always dies. Crabs withthis "gray crab disease" may have all theirblood cells destroyed and replaced by theamoeba. The nonclotting hemolymph* acquires the grayish tint from the organisms.By this stage of the disease, the host'sstored nutrients have been depleted, makingrecovery unlikely. Some crabs of all sizes,sexes, and stages, however, can overcomelight infections and survive. Hemocytes*engulf or encapsulate the protozoan, andother individuals are destroyed by humoral*mechanisms.
The presence of even low grade amoebicinfections has not been discovered in the
Gulf. If that absence results from geographical barriers and not from insufficient
examinations or from some biological orenvironmental reasons, it becomes
imperative not to introduce eastern crabsinto the Gulf. Crabs are already transportedin the opposite direction. Seafood companies routinely ship live Gulf crabs to the
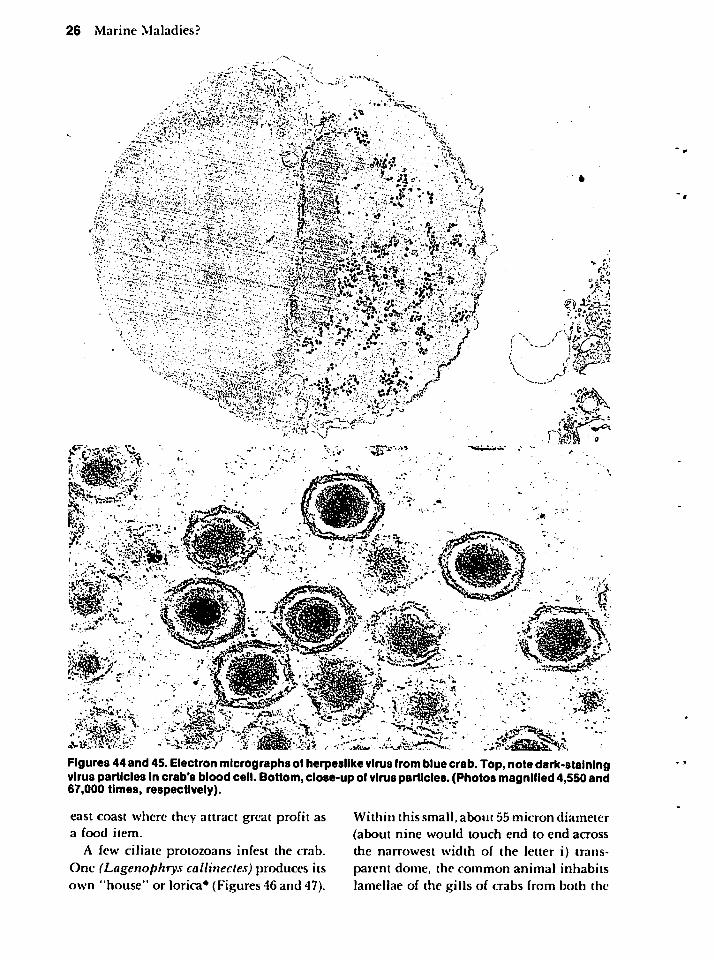
26
Marin
eM
ala
die
s?
"N
^
\<
**
1FV&
&•
,*-A4
PS
*-
I'-
"
•*^ii
%.
<*
"3P
-"
Figures44
and45.E
lectronm
icrographsofherpeslike
virusfrom
bluecrab.T
op,notedark-staining
virusparticles
incrab's
bloodcell.
Bottom
,close-u
pofvirus
particles.(Ph
oto
sm
agnified4,550
an
d6
7,0
00
times,
respectively).
eastco
astw
here
theyattract
great
pro
fitas
afo
od
item
.
Afew
ciliatep
roto
zoan
sinfest
thecrab.
On
e(L
agenop
hrys
callinectes)
producesits
ow
n"h
ou
se"o
rlorica*
(Figures
46an
d47).
With
inth
issm
all,a
bo
ut
55
mic
ron
dia
mete
r
(abo
ut
nin
ew
ou
ldto
uch
end
toen
dacro
ss
thenarrow
estw
idth
ofth
eletter
i)tran
sp
arent
dome,
the
com
mo
nan
imal
inh
abits
lamellae
ofth
eg
illso
fcrab
sfrom
bo
thth
e
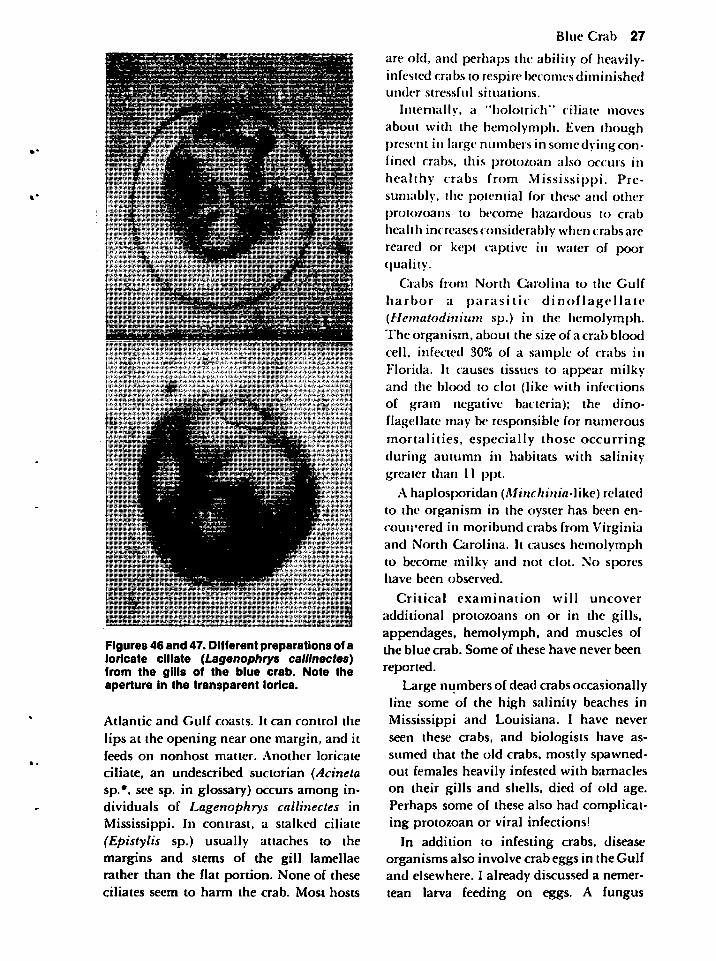
Figures 46 and 47. Different preparations of aloricate ciliate (Lagenophrys callinectes)from the gills of the blue crab. Note theaperture in the transparent lorica.
Atlantic and Gulf coasts. It can control the
lips at the opening near one margin, and itfeeds on nonhost matter. Another loricate
ciliate, an undescribed suctorian (Acinetasp.*, see sp. in glossary) occurs among individuals of Lagenophrys callinectes inMississippi. In contrast, a stalked ciliate(Epistylis sp.) usually attaches to themargins and stems of the gill lamellaerather than the flat portion. None of theseciliates seem to harm the crab. Most hosts
Blue Crab 27
are old, and perhaps the ability of heavily-infested crabs to respire becomes diminishedunder stressful situations.
Internally, a "holotrich" ciliate moves
about with the hemolymph. Even thoughpresent in large numbers in some dying confined crabs, this protozoan also occurs inhealthy crabs from Mississippi. Presumably, the potential for these and otherprotozoans to become hazardous to crab
health increases considerably when crabs arereared or kept captive in water of poorquality.
Crabs from North Carolina to the Gulf
harbor a parasitic dinoflagellate(Hematodinium sp.) in the hemolymph.The organism, about the size of a crab bloodcell, infected 30% of a sample of crabs inFlorida. It causes tissues to appear milkyand the blood to clot (like with infectionsof gram negative bacteria); the dinoflagellate may be responsible for numerousmortalities, especially those occurringduring autumn in habitats with salinitygreater than 11 ppt.
A haplosporidan (MincAtnia-like) relatedto the organism in the oyster has been encountered in moribund crabs from Virginiaand North Carolina. It causes hemolymphto become milky and not clot. No sporeshave been observed.
Critical examination will uncover
additional protozoans on or in the gills,appendages, hemolymph, and muscles ofthe blue crab. Some of these have never been
reported.
Large numbers of dead crabs occasionallyline some of the high salinity beaches inMississippi and Louisiana. I have neverseen these crabs, and biologists have assumed that the old crabs, mostly spawned-out females heavily infested with barnacleson their gills and shells, died of old age.Perhaps some of these also had complicating protozoan or viral infections!
In addition to infesting crabs, diseaseorganisms also involve crab eggs in the Gulfand elsewhere. I already discussed a nemertean larva feeding on eggs. A fungus

28 Marine Maladies?
(Lagenidium callinectes) invades them. Insalinities of 5 to 30 ppt, up to 25%of an eggmass may be infected. Infected eggs aresmaller and more opaque than healthy ones.The diseased portion of the sponge, whichis always at the periphery, appears brownish rather than yellowish-orange or grayishwhen the crab's larvae have developedenough to give the healthy part of thesponge a brown or blackish appearance.Infections become a real concern for
people rearing crustaceans, especially sincelarvae can become infected. On the other
hand, infections can aid natural populations; the fungus also infects the ova of thesymbiotic barnacle (Chelonibia patula).
PENAEID SHRIMPS
The valuable shrimp fishery in thenorthern Gulf of Mexico includes primarilythree species: the brown shrimp (Penaeusaztecus), the white shrimp (Penaeussetiferus), and the pink shrimp (Penaeusduorarum), the latter being much moreimportant in southern Florida than in thenorthern Gulf.
All species spawn in high salinity regions;their larvae develop in the offshore plankton* before metamorphosing into post-larvae and invading the estuarine* nurseryregions. Of all the shrimp, the pink shrimprequires the highest salinity water; it alsoprefers a sandy-shell habitat. Brown andwhite shrimp thrive in muddy-sand or siltyhabitats, where, in about 30 to 50 meters
of water, the brown shrimp can be caught inlarge numbers during night hours betweenJune and August. It remains commerciallyabundant down to 110 meters. On the other
hand, the white shrimp moves about duringthe day and occurs more commonly in waterless than 18 meters deep with salinity lessthan full strength seawater. Consequently,the white shrimp is available for small inshore trawlers, usually between August andNovember.
Postlarval shrimps invade the estuary atdifferent periods, with most brown shrimp
arriving about March and most whiteshrimp during the summer. As both becomejuveniles, the white shrimp can be foundfurther up bayous and rivers than brownshrimp; the white shrimp remains longer ininshore regions.
The number of these young shrimp thatwill enter the commercial fishery dependsnot on the number of spawning adults, buton the temperature, amount of freshwater,and other environmental conditions that
affect survival and growth. These factorsalso have a bearing on the parasites anddiseases of shrimp hosts.
Adults usually live 1 to 2 years, spawningtwo or more times. Some individuals, how
ever, survive a few additional years and theseshrimp usually possess quite an assortmentof symbionts. Shrimp of all ages and stagesharbor parasites, and the parasites occur inmany sites. Figure 48 of a brown shrimp indicates many of these sites occurring in allspecies.
The majority of parasites and diseaseconditions affecting shrimp, unlike thoseaffecting most marine organisms, have beeninvestigated in some detail. Still, a weekend fisherman or even a commercial
shrimper seldom recognizes many of theconditions. In fact, those people rearingshrimp and encountering heavy mortalitiesrarely know the causes. Although blamedfor many shrimp-kills, low oxygenconcentration levels may not always be thesole culprit. The shrimp may have to harbora symbiont in addition to being exposed tolow oxygen levels in order to succumb.
Because shrimp diseases are well documented, I will discuss several of the
conspicuous ones as well as some parasitesthat could be encountered from a highpercentage of shrimp if a curious individualdecided to dissect his specimens and use amicroscope.
Microbial Diseases
Unlike most viral diseases known from
crustaceans, one (Baculovirus penaei)involves the digestive gland of shrimp, and
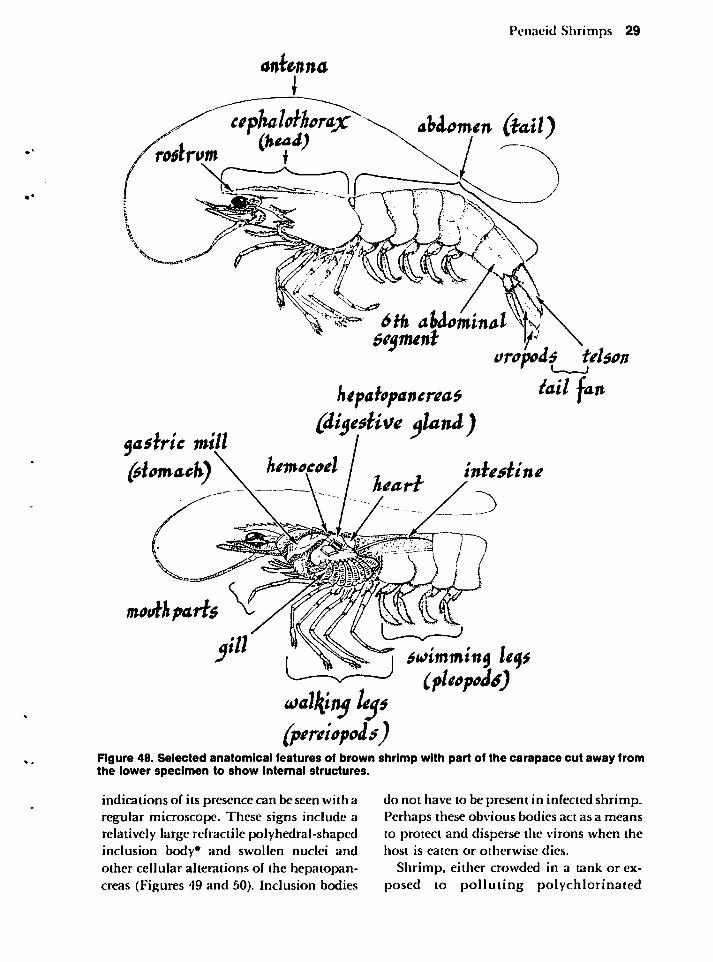
Penaeid Shrimps 29
Momen (fail)
eeymentoropo&s UUcn
htpahpantrea* iail fan(di$e$Uw aland)
hemocod I inhdineyasiric mill
nu>dkpi
(pereiopolsj
fu>immin$ Ufy(fUopids)
Figure 48. Selected anatomical features of brown shrimp with part of the carapace cut away fromthe lower specimen to show internal structures.
indications of its presence can be seen with aregular microscope. These signs include arelatively large refractile polyhedral-shapedinclusion body* and swollen nuclei andother cellular alterations of the hepatopan-creas (Figures 49 and 50). Inclusion bodies
do not have to be present in infected shrimp.Perhaps these obvious bodies act as a meansto protect and disperse the virons when thehost is eaten or otherwise dies.
Shrimp, either crowded in a tank or exposed to polluting polychlorinated
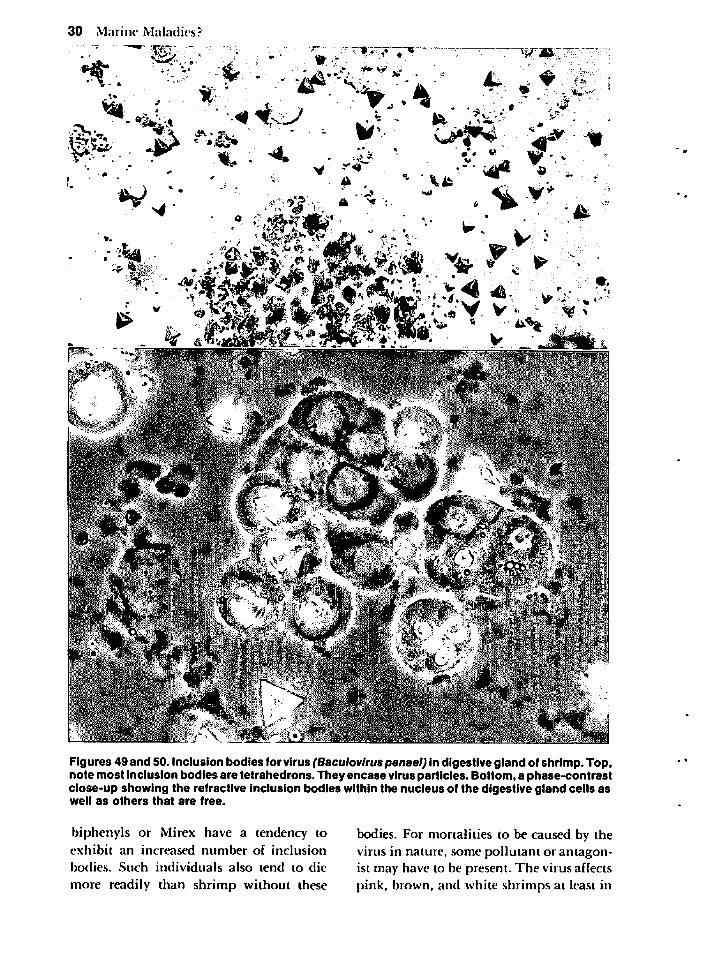
30 Marine Maladies?
mz r?» •
Z\
V
A '^ a".-,"...-:. •' V
v ms.M^jm&ft*
Figures 49 and 50. Inclusion bodies for virus (Baculoviruspenael) in digestive gland of shrimp. Top,note most inclusion bodies are tetrahedrons. They encase virus particles. Bottom, a phase-contrastclose-up showing the refractive inclusion bodies within the nucleus of the digestive gland cells aswell as others that are free.
biphenyls or Mirex have a tendency to bodies. For mortalities to be caused by theexhibit an increased number of inclusion virus in nature, some pollutant or antagon-bodies. Such individuals also tend to die ist may have to be present. The virus affectsmore readily than shrimp without these pink, brown, and white shrimps at least in

Figures 51 and 52. Brown shrimp with shell-disease. Bottom, disease-conditionoccurring in site of wound. Lesions in bothshrimp possessed a bacterium (Beneckeatype I) which digests chitin in the exo-skeleton and probably caused the disease.
Florida and Mississippi and probably infects many penaeids wherever they occur.
Chitinoclastic organisms includingbacteria cause shell disease, or black spotdisease, in shrimps (Figures 51 and 52) aswell as in the blue crab and other crusta
ceans. The lesion in Figure 52 illustratesa wound inflicted by a predator. Such infections are usually lost with a molt, andrarely does an affected shrimp die from thisalone. When specific bacteria (e.g., achitinoclastic variety of Vibrioanguillarum) involve pond-reared shrimps,death can occur. Indeed, low grade mortalities, apparently not resulting fromsecondary* infections, have plagued reared-shrimp production in Mexico.
Shrimps, just as the blue crab, host avariety of internal bacteria, some of whichcause extensive mortalities in pond-rearedhosts and others that do not affect the host.
The white opaque gill filaments of a whiteshrimp from Mississippi Sound shown inFigure 53 probably exemplify a bacterial
Penaeid Shrimps 31
Figure 53. A white shrimp with some white-colored gills probably caused by a bacterialinfection. Many host blood cells crowdedtogether in the affected gills.
infection. A massive hemocytic responseinvolved the area, producing the opaquecondition.
Black gills or "black gill disease" canresult from a bacterial infection, but more
often this multicaused disease results from
other agents, and a few of these will bediscussed later.
Externally, a filamentous bacterium(probably Leucothrix mucor) entangleslarval shrimp, and it, along with considerable trapped debris, probably inhibitsrespiration and otherwise burdens the host.As with most diseases, reared shrimp usuallyconstitute the primary victims. The same ora similar bacterium also occurs on a varietyof crustaceans, fishes, and plants.
Shrimp, as well as a variety of othermarine animals, can harbor Mycobacteriummarinum. This bacterium, related to the
tuberculosis agent of man, can cause a skinrash (dermatitis) on those susceptiblepeople who handle shrimp or other seafoodproducts. Reactions differ among people:some have hard pustules on their handswhereas others have blisters.
A variety of fungi infects shrimps, butmost have not been cultured or investigated.One that has been investigated (Fusariumsolani) shows variation in its virulence todifferent penaeid shrimps. A response by thehost hemocytes to spores and hyphae* in thegills can result in a deposition of darkpigment (melanin) producing "black gilldisease."

32 Marine Maladies?
Figures 54-57. Fungal black gill disease. Top left, brown shrimp with disease compared with uninfected individual. Top right, spores (macroconldia) of causative agent (Fusarlum solani). Bottomleft, host response by shrimp blood cells after 24 hours. Bottom right, increased pigmentation(melanization) apparent by 28 hours. The text discusses several other agents that cause black gilldisease.
Deposition of brown-to blackish melaninpigment occurs in shrimp that have beenwounded, have been exposed to foreignmaterials, and have fungal (see Figures 54to 57), protozoan, or helminth infections.In some cases, the tissue underlying the carapace* and not the actual gills is darkened.Perhaps waste products (nitrates orammonia) discharged through the gills oreven precipitation of materials (metallicsulfides) produces the condition.
Protozoan Infections
What's a cotton shrimp? Four species ofmicrosporidan protozoans infect commercial shrimps in the northern Gulf. Three ofthose infect and replace nearly all theabdominal muscle tissue of their hosts,
resulting in an extremely opaque conditionsomewhat similar in appearance to a cookedshrimp (Figure 58). Shrimpers refer to thesehosts as "cotton shrimp" or "milky
Figure 58. Tails of the brown shrimp; the topshrimp exhibits cotton shrimp disease.Three different species of microsporidanscause this milky or chalky appearance oftails of penaeids in the northern Gulf ofMexico. In this case, tiny spores of Amesonnelsonl surround and eventually replace themuscle bundles.
shrimp." The name "cotton shrimp" alsoreflects the texture of the cooked product.
The fourth microsporidan species(Agmosoma penaei, Figure 59) is more

Figure 59. Chalky appearance under cuticlereflects microsporidan (Agmasoma penaei)that invaded blood vessels under the cara
pace of the white shrimp. Usually, tissuealong the middle portion of the top of the tailexhibits infections by this protozoan. Aninsert shows some spores in the typicalpacket of eight. Others have broken loosefrom the membrane, and one spore (macro-spore) is typically much larger than theothers.
host-specific than the others and does notinfect the tail. Involving primarily thewhite shrimp, an infection appears mostdistinctly along the dorsal midline of thetail and under the carapace in the cephalo-thorax. Many people think that infectedgonads and intestinal tissue are ripe ovaready to be released. Consequently, someprefer the taste of these "ripe" shrimp overnoninfected shrimp.
More than one species of microsporidancan infect an individual shrimp. In fact,three species in a single shrimp can bevisually differentiated by their color andlocation if the tail is cut crossways.
Infected shrimp react poorly to stressconditions. Usually, when compared withtheir noninfected counterparts, they diemore readily when handled, are more vulnerable to predators, and do not normallypartake in migrations. Often many shrimpcaught inshore exhibit microsporidaninfections. In these cases the bulk of the
shrimp stock may have previously departedto other waters; however, a few cotton
shrimp do occur in offshore catches,suggesting that some infected individualsmigrate and survive.
Several protozoans other thanmicrosporidans have symbiotic relation-
Penaeid Shrimps 33
ships with penaeids. Also, all species ofshrimp in the northern Gulf harbor a cepha-line gregarine (Nematopsis penaeusor someother indistinguishable species). The motilestage (trophozoi te) can be seen attached to orgliding about in the intestine, sometimeswith as many as seven individuals joinedend to end in a chain or branched so as to
exhibit a forked posterior end (Figures 60and 61). Two attached individuals developinto a spherical cyst that embeds in theshrimp rectum (Figure 62); the cyst in turnproduces a stage infective for some yet unknown mollusc.
****?"
».
<•; * -
"J
Figures 60-62. A cephaline gregarine fromthe white shrimp. Top, an association ofthree Individuals in a chain gliding about inthe intestine. Middle, a forked association.Bottom, cysts (gamontocysts) embedded inrectum. These result from encystment ofassociations and can barely be seen as achalky-colored dot with unaided naked eyes.Stages in the rectum infect appropriatemolluscan hosts.

34 Marine Maladies?
For another similar appearing species(Nematopsis duorari) restricted to pinkshrimp, the shrimp acquires infections by-feeding on mucus trails of several commonbivalves including the Atlantic bay scallop,broad ribbed cardita, cross-barred venus,and sunray venus.
These and at least two more gregarinespecies that attach to feeding appendagesprobably cause no more than minor diseasein either the shrimp or the correspondingmolluscan hosts.
Ciliated protozoans occur in as muchvariety as do gregarines. On the gills,colonies of a stalked peritrich (Zootham-nium sp.) give a brownish cast to the gillswhen enough of these organisms attach.The attachment of the stalk to a gill examined with an electron microscope revealsno pathological alterations. The simpleattachment of a small colony can be seen inFigure 63. However, when a large number ofthe ciliate infests the gills, as it can whenhosts are crowded or placed under specificenvironmental conditions, shrimp oftendie. Death usually coincides with periods oflow concentration of the water's oxygen, acommon condition following several warmovercast days or following decomposition ofa large algal bloom. The ciliate apparentlycompetes for the available oxygen becauseshrimp with few colonies of the organisms
Figure 63. A colonial ciliate (Zoothamnlumsp.) on the gills of a commercial shrimp. Thedark fiber extending along the stalk's centerpermits the entire colony to contractsimultaneously. Note the cilia at the top ofthe bell which direct food particles into theorganism. The protozoan harms shrimp onlywhen certain environmental stresses arise.
often survive the stress whereas those with
many die.
A related species (Epistylis sp.) differsfrom Zoothamnium sp. primarily by lacking a central contractile fibril within thestalk. Thus, it cannot contract its stalk. The
fibrils (myonemes) of Zoothamnium sp.connect where the stalks branch, allowingsimultaneous contraction or expansion ofan entire colony and a wonderful showwhen seen under the microscope!
Unlike Zoothamnium sp., Epistylis sp.usually infests the appendages and exo-skeleton-proper rather than the gills. Bothspecies infest larval shrimp and presenttreatable threats to cultured shrimp. Theeffect on natural populations has not beenestablished.
Another unnamed ciliate encysts on or ingills. Common where distal gill filamentsbranch, this nonstalked apostome alsocauses black gills (Figures 64 to 67). Specksof melanin surround the ciliate. The life
cycle of this foettingeriid has not been established, and several technical papersconfuse it or a similar species with loricateforms like the one on the blue crab. External
loricae*. however, have apertures surrounded by lip-like structures and thereby providean easy means of differentiation.
Other protozoans can be seen by carefullyexamining shrimps from a variety ofhabitats. Relationships among some ofthese with the host remain obscure. An un
identified suctorian species of Ephelota onthe exoskeleton looks entirely different fromthe one on crab gills. An internal ciliate(Parauronema sp.) which can fill the entirehemocoel and a flagellate (TLeptomonassp.) which also occurs internally have beenimplicated in larval mortalities, possiblyin conjunction with the virus Baculoviruspenaei.
Helminths
Shrimps act as second intermediate hostsfor a few digeneans. For those species withspherical cysts enclosing the larval worms,infections are localized primarily in the tail,
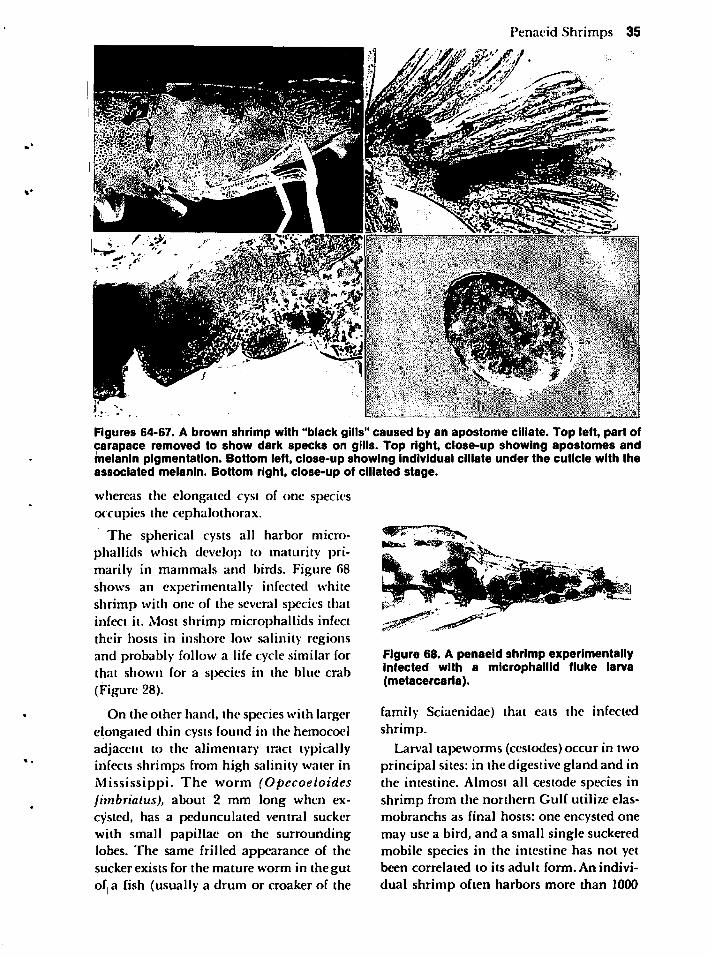
Penaeid Shrimps 35
Figures 64-67. A brown shrimp with "black gills" caused by an apostome ciliate. Top left, part ofcarapace removed to show dark specks on gills. Top right, close-up showing apostomes andmelanin pigmentation. Bottom left, close-up showing individual ciliate under the cuticle with theassociated melanin. Bottom right, close-up of ciliated stage.
whereas the elongated cyst of one speciesoccupies the cephalothorax.
The spherical cysts all harbor microphallids which develop to maturity primarily in mammals and birds. Figure 68shows an experimentally infected whiteshrimp with one of the several species thatinfect it. Most shrimp microphallids infecttheir hosts in inshore low salinity regions
and probably follow a life cycle similar forthat shown for a species in the blue crab(Figure 28).
On the other hand, the species with largerelongated thin cysts found in the hemocoeladjacent to the alimentary tract typicallyinfects shrimps from high salinity water inMississippi. The worm (Opecoeloidesfimbrialus), about 2 mm long when ex-cysted, has a pedunculated ventral suckerwith small papillae on the surroundinglobes. The same frilled appearance of thesucker exists for the mature worm in the gutof a fish (usually a drum or croaker of the
Figure 68. A penaeid shrimp experimentallyinfected with a microphaiiid fluke larva(metacercaria).
family Sciaenidae) that eats the infectedshrimp.
Larval tapeworms (cestodes) occur in twoprincipal sites: in the digestive gland and inthe intestine. Almost all cestode species inshrimp from the northern Gulf utilize elas-mobranchs as final hosts: one encysted onemay use a bird, and a small single suckeredmobile species in the intestine has not yetbeen correlated to its adult form. An indivi
dual shrimp often harbors more than 1000

36 Marine Maladies?
Figure 69. Larval (plerocercoid) tapewormfrom intestine of commercial shrimp. Over1000 individuals of this small unidentifiedlarva can attach by their single sucker to thegut of a single shrimp.
specimens of this larval intestinal cestode(Figure 69), and large numbers may harmthe shrimp.
Becausemost prevalent, most studied, andmost conspicuous in all penaeids from theGulf, one larval trypanorhynch, or tetra-rhynch as it is also called (Prochristianellahispida), will be treated in detail.
Crustacean-feeding stingrays (Dasyatissabina and Dasyatis sayi) harbor matureProchristianella hispida in theirspiral valve(an adaptation of the intestine to increase itssurface area). Figure 70 diagrams the lifecycle. Segments (proglottids) with partiallydeveloped eggs break free from the tapeworm and pass out with a ray's feces.Once inseawater the proglottids split open, liberating many eggs, each with two thin, sticky
Figure 70. Life cycle of a trypanorhynch tapeworm (Prochristianella hispida) in commercialshrimps. Stingrays harborthe adult worm in theirspiral valve. Terminal segments drop off the wormand release eggs that develop larvae when in seawater. The enclosed larva develops In cope-pods. Whether shrimp obtain their infections from eggs or from copepods has not been established, but rays probably acquire infections from various shrimps.

i"" in.
.-• '
alll* /ililflllill
-?*r ££**"'''•• w
dr "
Figures 71 and 72. Larval stages of atrypanorhynch cestode (Prochristianellahispida). Top, filamented eggs with larvaeclumped together. Bottom, blastocysts, eachwith enclosed larva (plerocercoid) in thedigestive gland of a penaeid shrimp. Aportion of the pigmented membranecovering the digestive gland has beenremoved on one half to expose digestivecaeca. Three whitish cysts are apparent onthe pigmented membrane.
filaments (Figure 71). A single hooked larva(oncosphere with six anchor-like hooks)within the egg develops to an infectivestage in about 4 days. Whether shrimp getinfected by eating food with entangled eggsor by eating infected copepods has not beenestablished. A larval form (procercoid toplerocercoid) develops in the hemocoel ofsome copepods and may progress similarlyin a variety of shrimp, or possibly a shrimpmay have to eat the infected copepod. A hardcapsular cyst surrounds the early plerocercoid within the fleshy procercoid-like stage(blastocyst) when in the copepod. This en-cystment has never been seen nor reportedfrom shrimp and might be a transitory developmental structure present only undercertain conditions. On the other hand, other
crustaceans such as the ghost shrimp, whichpossesses blastocysts larger than those in
Penaeid Shrimps 37
penaeid shrimps, may be the ray's mostutilized crustacean host. In order for the
cycle to be complete, the ray eats an infectedcrustacean. The blastocyst, but not theactual plerocercoid, can be seen in a shrimpwithout disturbing it if the cyst protrudesthrough the host's membrane covering thedigestive gland (Figure 72). Some individualblastocysts occur entirely within the glandand elsewhere within the hemocoel.
The adult trypanorhynch attaches to theray's gut by means of four eversible hookedtentacles. A tentacle turns itself inside out.
By the size, shape, and arrangement of thehooks, one characterizes the several speciesthat infect penaeids as well as other speciesthat parasitize a wide variety of crustaceans,molluscs, fishes, and occasionally otherhosts. These attachment structures remain
unchanged from the late larval to adultstage.
Larval roundworms (nematodes) alsoinfect penaeids and several other hosts.Most common in a variety of fishes andinvertebrates including shrimp is an ascari-doid (Thynnascaris type MA). Several individuals of this worm and a smaller
number of a smaller member in the
same genus (Thynnascaris type MB) occurcommonly either coiled and encysted or freein the digestive gland, among other tissuesof the cepahlothorax, or occasionally in thetail portion of the shrimp. The smallerspecies (type MB) may be a health hazard.Within an hour after being eaten by amouse, it can penetrate into or through thewall of the mouse's alimentary tract andcause a host-response. Some individualslive for a considerable time in the mammal,
whereas the host reacts against and quicklykills others. In any event, the potential existsfor a person to develop a severe inflammatory response after eating raw seafoodharboring the worm or to express a hypersensitivity reaction from worms eatensubsequent to the initial infection. Suchconditions, with symptoms similar to thosefor an ulcer, appendicitis, or cancer, have
affected people in Japan, the Netherlands,

38 Marine Maladies?
and other places where people consume rawseafood harboring related nematodes. Eithercooking or subzero freezing prevents humaninfections!
Life histories and identifications of the
larval nematodes have not been established.
Just as with the trypanorhynch, I do notknow if shrimp acquire infections by eatinglarvae directly, by eating copepods withinfective larvae, or by other means. Thefinal host must be a fish and several speciesof these nematodes exist in the Gulf of
Mexico and elsewhere.
One reason why so many individual hostsmay harbor the same species of larva is thatan individual worm may penetrate throughthe alimentary tract of a predator afterbeing eaten along with a previous host andrcencyst in the mesentery or elsewhere, remaining infective for another predator thatin turn eats and retains it. Not until ac
quired by a proper definitive host will theworm molt and develop to maturity.Parasitologists call these hosts in whichno development of a worm occurs "para-tenic" hosts. These hosts allow completionof a cycle in cases where the definitive hostrarely feeds on the true intermediate host.The worm may increase in size, but it doesnot develop additional features. A similarpattern of life cycle probably occurs formost related ascaridoid nematodes and these
will be diagrammed and discussed morelater.
Even though quite rare in penaeidshrimp, another larval nematode (Spiro-camallanus cricotus) completes its cycle in awide variety of fishes. Whether shrimps actually constitute a necessary link in thecycle has not been established, but theyprobably do not because other invertebratesalso host the larva of this dramatic worm. It
will be figured and discussed in more detailin the section on fish nematodes because it
is in fish where the large reddish adultworm is most easily visible and most
commonly found.
Even though other larval nematodes infect penaeids from other parts of the world.
the only other fairlycommon nematode thatI have seen in Gulf shrimp is what appearsto be a free-living* nematode (Leptolaimussp.). Leptolaimus sp. in shrimps fromLouisiana to Alabama appear able to livein and on shrimp without being digested.The tolerance by the shrimp and relationship between worm and shrimp requirefurther investigation; all related nematodespecies thrive without hosts in muddy-sandyhabitats, and this one probably also livesin such a habitat. Many strictly free-livingnematodes exist in water and soil, and
notably few of these possess any strongfamily ties to any of the symbiotic speciesfrom aquatic hosts.
Miscellaneous Diseases and Attached
OrganismsOne amazing disease can be characterized
by the opaque musculature in the tail ofaffected shrimp. Going by the technicalname "spontaneous necrosis," this progressive disease begins as a whitish areausually near the end or middle of the tail.Caused by an adverse stress such as lowoxygen concentration or rapid change ofthe water's temperature or salinity, thecondition progressively worsens, leading todeath unless the harsh conditions improve.The disease may occur frequently in shrimpheld in bait holding tanks and could be confused with cotton shrimp unless shrimpwith each of the two conditions could be
compared.
The above necrosis* is reversible. Shrimpprovided water of adequate quality will losethe opaque appearance unless the conditionhas developed beyond a certain thresholdpoint. Once the tip of the tail acquires atotally chalky white appearance, the shrimpusually dies. The really amazing phenomenon that biologists should pursue is howshrimp with a partially necrotic conditioncan, within a day or so, recover and appearnormal. Microscopic examination of affected opaque tissue reveals nearly totallydissociated material, but a few days later,the tissue appears to regain its normal

character. Investigation of this regeneratingtissue aided by the electron microscoj>eshould go a long way to explain the processof muscle regeneration, a transformationthat takes considerable time in man and
other vertebrates.
Another condition called "crampedshrimp" develops in individuals held inbait tanks and ponds. The tail can becompletely or partially flexed. It becomesso rigid that it cannot be straightenedout. Partially-cramped individuals swimabout in a humped fashion and may survivethe ordeal. Those laying on their sides seldom improve and usually die. An imbalanceof ions appears to cause cramping.
Occasionally a shrimp portrays a goldenappearance throughout all its tissue. Theyellowish-gold shrimp may appear eitheropaque or transparent, and the rarity ofaffected individuals reared in pondssuggests that the condition may representan hereditary phenomenon rather than oneresulting from dietary deficiency or some
other unnatural process.The crab leech may also occur on pena
eids, but not nearly as frequently as itutilizes the crab or grass shrimp.
A variety of symbionts and fouling organisms often attach to older shrimp.Usually those shrimp not molting for longperiods reveal the most spectacular casesbecause large sessile barnacles grow on thecarapace or elsewhere. Algae, hydroids. andoccasionally other organisms attach to andsometimes invade gills, eyes, or less vulnerable regions. Presumably these interferewith the best possible life style for a shrimp.
Confusion may also exist when separating parasites from normal anatomicalstructures. One of these structures, the podlike sperm capsule, or spermatophore, israrely seen in inshore shrimp. Most individuals spawn offshore. During mating, themale transfers his sperm capsule to the female where it becomes attached to the
female's external genital organ (thelycum),located between her fourth and fifth pair ofwalking legs, by means of various anchor-
Penaeid Shrimps 39
ing devices and a glutinous material. Sincethe brown shrimp has a closed thelycum,copulation apparently occurs between arecently molted, soft-shelled female and ahard-shelled male in contrast with the process in white shrimp which takes placebetween hard-shelled individuals because
the thelycum is open. In spite of the plates,bristles, spines, glue, and other devices, thecapsule easily dislodges. Nevertheless,fertilization of ova in these individuals
seems to take place.
The female white shrimp with theattached spermatophore in Figures 73 and74 shows a typical condition in offshoreshrimp. A fisherman, however, collectedthis particular shrimp from inshore waterwhere attachment is rare and, for that
matter, unproductive if eggs are releasedunless the salinity concentration is higherthan normal. The strange attached objectcould be misinterpreted as an externalparasite.
Some noninfectious diseases result from
Figures 73 and 74. A male's spermatophore(encapsulated sperm) attached to a 173 mmlong female white shrimp. Top, side view.Bottom, underside view.
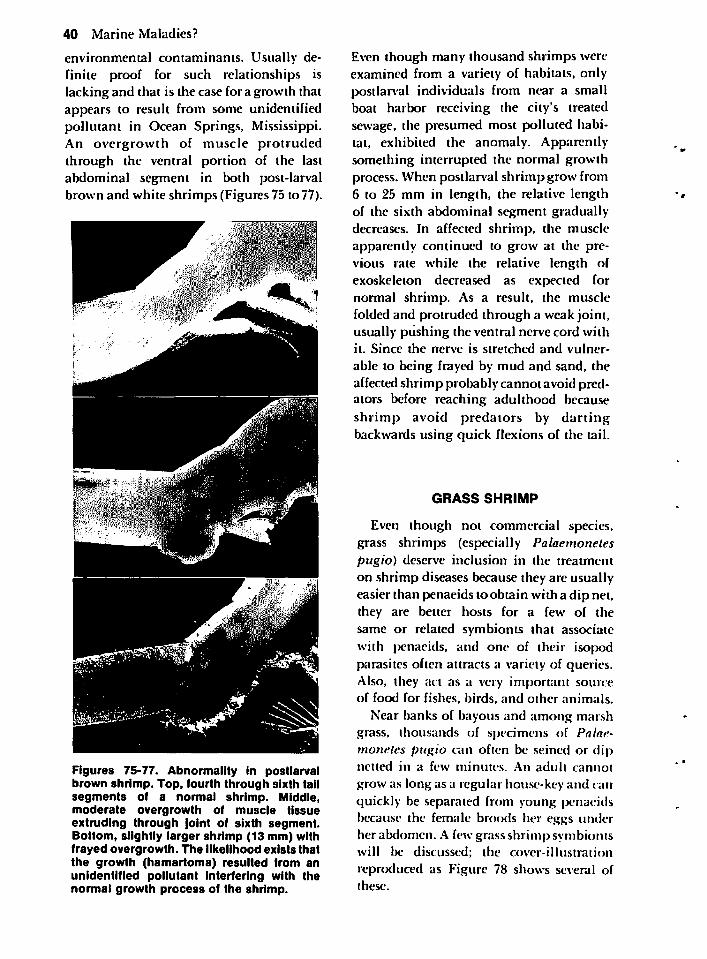
40 Marine Maladies?
environmental contaminants. Usually definite proof for such relationships islacking and that is the case for a growth thatappears to result from some unidentifiedpollutant in Ocean Springs, Mississippi.An overgrowth of muscle protrudedthrough the ventral portion of the lastabdominal segment in both post-larvalbrown and white shrimps (Figures 75 to 77).
Ml-;"%$&
Figures 75-77. Abnormality in postlarvalbrown shrimp. Top, fourth through sixth tailsegments of a normal shrimp. Middle,moderate overgrowth of muscle tissueextruding through joint of sixth segment.Bottom, slightly larger shrimp (13 mm) withfrayed overgrowth. The likelihood exists thatthe growth (hamartoma) resulted from anunidentified pollutant interfering with thenormal growth process of the shrimp.
Even though many thousand shrimps wereexamined from a variety of habitats, onlypostlarval individuals from near a smallboat harbor receiving the city's treatedsewage, the presumed most polluted habitat, exhibited the anomaly. Apparentlysomething interrupted the normal growthprocess. When postlarval shrimp grow from6 to 25 mm in length, the relative lengthof the sixth abdominal segment graduallydecreases. In affected shrimp, the muscleapparently continued to grow at the previous rate while the relative length ofexoskeleton decreased as expected fornormal shrimp. As a result, the musclefolded and protruded through a weak joint,usually pushing the ventral nerve cord withit. Since the nerve is stretched and vulner
able to being frayed by mud and sand, the
affectedshrimp probably cannot avoid predators before reaching adulthood becauseshrimp avoid predators by dartingbackwards using quick flexions of the tail.
GRASS SHRIMP
Even though not commercial species,grass shrimps (especially Palaemonetespugio) deserve inclusion in the treatmenton shrimp diseases because they are usuallyeasier than penaeids to obtain with a dip net,they are better hosts for a few of thesame or related symbionts that associate
with penaeids, and one of their isopodparasites often attracts a variety of queries.Also, they act as a very important sourceof food for fishes, birds, and other animals.
Near banks of bayous and among marshgrass, thousands of specimens of Palaemonetes pugio can often be seined or dipnetted in a few minutes. An adult cannot
grow as long as a regular house-key and canquickly be separated from young penaeidsbecause the female broods her eggs underher abdomen. A few grass shrimp symbiontswill be discussed; the cover-illustration
reproduced as Figure 78 shows several ofthese.

Grass Shrimp 41
Figure 78. A grass shrimp (Palaemonetes pugio) with a variety of its symbionts. On the tail fan andlegs, a ciliate (Lagenophrys lunatus) lives within a transparent encasement opened to the externalenvironment. Another ciliate attached by its stalks to the anterior claw. In the tail muscle, theencysted spherical fluke larva (Microphallus choanophallus) looks conspicuous in the illustrationand also in the living shrimp if a host is held up to the light. The three larger dark spheres constitute the same fluke cysts, but hyperparasitized by a haplosporidan protozoan (Urosporldlumcrescens). When Infecting a related fluke in the blue crab, the association has been termed"buckshot" or "pepper spot." A bopyrld isopod (Probopyrus pandallcola) occludes the anteriorside of the shrimp, and the common leech (Myzobdella lugubris) darts about on the top of theshrimp. The text covers these and several other symbionts from the grass shrimp.
Protozoans
The loricate ciliate (Lagenophryslunatus) encased on the tail fan, walkinglegs, and swimming legs in the illustratedshrimp provides an easier model to studythan the related ciliate species infesting thegills of the blue crab. It can be observedwithout harming the host and its development on a molting host can be followed.This protozoan species appears specific toshrimps of the family Palaemonidae (grassand river shrimp) in fresh and brackishwater.
Within the transparent lorica*, theperitrich projects its oral cilia throughthe aperture for feeding on scraps from thehost's food and on phytoplankton. Eventhough its body isattached to the opening of
the lorica, a gap exists that allows a swarmer(its telotroch larva) to emerge. The adultorganism can also contract, sealing off thataperture formed by the lorica's flexible collar extension. Microscopic examinationclearly shows the horseshoe-shapedmacronucleus and small micronucleus
diagnostic of many ciliates.Preceding the host's molting, the or
ganism, most prevalent on sites with thegreatest water flow, undergoes a special typeof asexual division producing twoswarmers(free swimming larvae). Loricas on freshmolts contain these swarmers, as well as
dividing (asexually reproducing), trophic(feeding), and conjugating (sexually reproducing) individuals. In other words,when the host molts, stages have already

42 Marine Maladies?
been produced to reinfest the same host oradditional hosts and thereby perpetuate thespecies.
Other ciliates also infest grass shrimp.One (Terebrospira chattoni) can be seen almost anywhere except on or adjacent to thegills. Small (50 to 100 u) elliptical cystsusually divided into compartmentalizedsectors may contain individuals or be empty.Related apostomes feed on molting(exuvial) fluid, but this species has becomeadapted to feeding on the endocuticle. Thetiny flattened ciliate can be found digestingthe cuticle* at the end of a tunnel leadingaway from one of the empty compartments.Is this a transition between an external and
internal symbiont?
In addition to ciliates, a variety of microsporidan protozoans also infect grassshrimp. One (Pleistophora linloni) differslittle from related species in penaeids, crabs,and even fishes. Large numbers of spores fillthe pansporoblast's chamber (Figure 79),and a heavy infection produces an opaquetail characteristic of "cotton shrimp."
Another species (Inodosporus spraguei)produces the same gross appearance, butthe spores are unusual. Each undischargedspore (with polar filament intact) containsthree or four long extensions ("tails") on itsbasal portion and a divided short one on theapical end. After an infected shrimp iseaten,tails on a spore voided with the predator's
Figure 79. A microsporidan (Pleistophorallntonl) from the tall of the grass shrimp.Members of this genus have several to manyhundred spores per chamber.
feces probably aid in tangling the spore tovegetation or trapping tidbits that mightultimately induce another grass shrimp toingest it. While developing, the tails maychannelize nutrients and ions to and from
the forming spore. Eight of these tailedspores occur within a pansporoblast.
Electron micrographs (Figures 80 and 81)show the organelles that these tiny (3 u longby 2 u wide) spores possess. The polar filament has its base (end of the diagonal narrow structure) attached to the apical end.Alterations in the plug at this junctureallow the tube to turn inside out and extrude
a considerable distance (about 40 u). Theprocess is aided by the lamellar (layered)structure (polarplast) at the apical end. Thesmall, round, peripheral structures arecross-sections of the uneverted coiled portion of the filament. The external tails do
not extrude; they develop simultaneouslywith the spores. The second micrographshows several early developing stages separated from mature ones by some unharmedhost muscle tissue. Each of the early stageswill divide, producing eight spores. Thetunnel-like arrangements adjacent to thematuring spores constitute various sectionsthrough the tails. Luck allowed a few sporesin this very thin section to reveal the placewhere the tails join the spores.
Whereas microphaiiid larval cysts(metacercariae) are hard to see in the penaeids because of a shrimp's large size and thepaucity of infections, cysts of Microphallus choanophallus are seldom absentin most stocks of Palaemonetes pugio. By-holding a grass shrimp up to a light or to thesky, many opaque specks, each representinga cyst, stand out conspicuously. Often,
Vrosporidium crescens hyperparasitizes*one or more of the cysts (Figure 78), makingthe cyst even more recognizable.
Metazoans
A bopyrid isopod (related to terrestrialorganisms called sowbugs, roly-poly bugs,or potato bugs) stands as a good example ofa parasite on grass shrimp that is not found
'c/
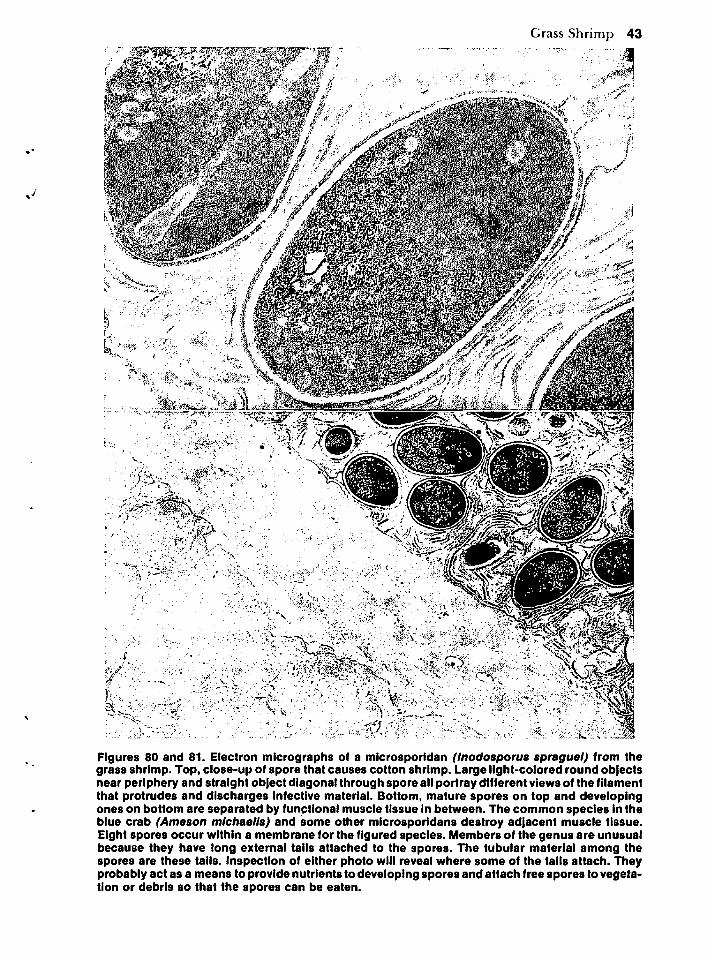
Grass Shrimp 43
Figures 80 and 81. Electron micrographs of a microsporidan (Inodosporus spraguel) from thegrass shrimp. Top, close-up of spore that causes cotton shrimp. Large light-colored round objectsnear periphery and straight object diagonal through spore all portray different views of the filamentthat protrudes and discharges infective material. Bottom, mature spores on top and developingones on bottom are separated by functional muscle tissue In between. The common species in theblue crab (Ameson michaelis) and some other microsporidans destroy adjacent muscle tissue.Eight spores occur within a membrane for the figured species. Members of the genus are unusualbecause they have long external tails attached to the spores. The tubular material among thespores are these tails. Inspection of either photo will reveal where some of the tails attach. Theyprobably act as a means to provide nutrients to developing spores and attach free spores to vegetation or debris so that the spores can be eaten.
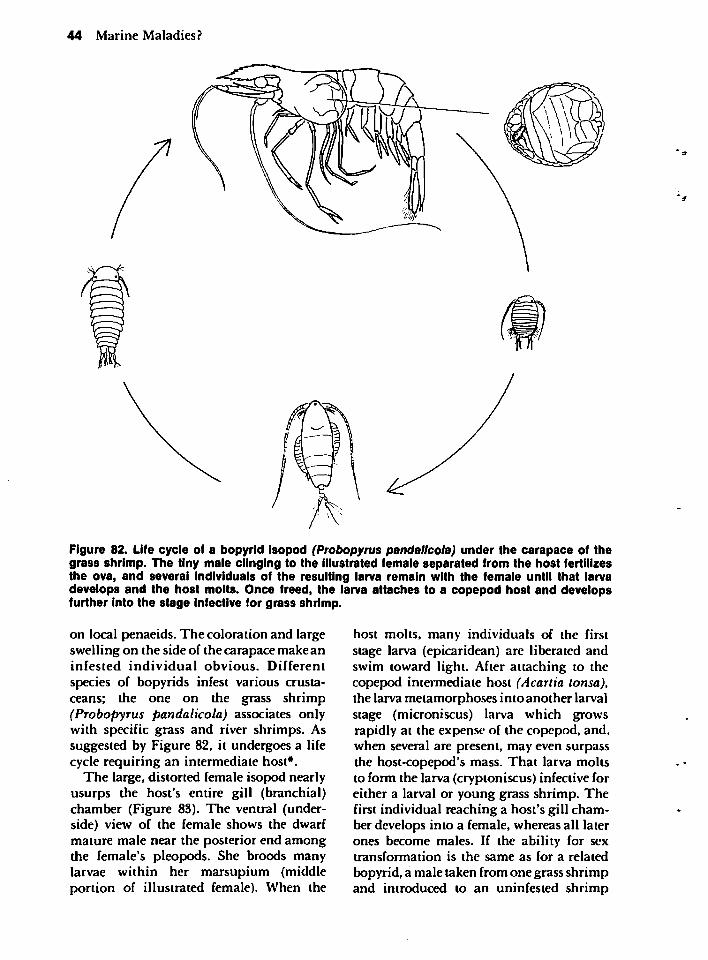
44 Marine Maladies?
Figure 82. Life cycle of a bopyrid isopod (Probopyrus pandallcola) under the carapace of thegrass shrimp. The tiny male clinging to the illustrated female separated from the host fertilizesthe ova, and several individuals of the resulting larva remain with the female until that larvadevelops and the host molts. Once freed, the larva attaches to a copepod host and developsfurther into the stage infective for grass shrimp.
on local penaeids. The coloration and largeswelling on the side of the carapace make aninfested individual obvious. Different
species of bopyrids infest various crustaceans; the one on the grass shrimp(Probopyrus pandalicola) associates onlywith specific grass and river shrimps. Assuggested by Figure 82, it undergoes a lifecycle requiring an intermediate host*.
The large, distorted female isopod nearlyusurps the host's entire gill (branchial)chamber (Figure 83). The ventral (underside) view of the female shows the dwarfmature male near the posterior end amongthe female's pleopods. She broods manylarvae within her marsupium (middleportion of illustrated female). When the
host molts, many individuals of the firststage larva (epicaridean) are liberated andswim toward light. After attaching to thecopepod intermediate host (Acartia tonsa),the larva metamorphoses into another larvalstage (microniscus) larva which growsrapidly at the expense of the copepod, and,when several are present, may even surpassthe host-copepod's mass. That larva moltsto form the larva (cryptoniscus) infective foreither a larval or young grass shrimp. Thefirst individual reaching a host's gill chamber develops into a female, whereas all laterones become males. If the ability for sextransformation is the same as for a related
bopyrid, a male taken from one grass shrimpand introduced to an uninfested shrimp

•0
Figure 83. Adult female bopyrid isopod(Probopyrus pandallcola) under carapace ofgrass shrimp.
would turn into a female! If several larvae
invade an uninfected shrimp simultaneously, all become females, but only one wouldeventually mature. Think of the experiments that could be conducted using thishost-symbiont system.
The ability for symbionts and hosts tosense their surroundings provides acontinual source of problems to investigate.The grass shrimp acts as a good host to studyfor harboring the "crab leech" (Myzobdellalugubris). Occasionally one finds an individual with two or more leeches attached
(Figure 84). However, if a person puts an infested and many noniniested shrimp in abowl and starts adding leeches to the bowl,he will observe that most will even
tually attach to one or a few shrimp. Asmany as 20 can reside on a single shrimp.
The leech probably does not utilize theshrimp as much as the blue crab for deposition of cocoons. It will use the shrimp,though, and after it does, the shrimp ofteneats the leech. Perhaps this results fromlethargy on the part of the spent* leech. Ihave seen the anterior third of an eaten leech
remain alive in a grass shrimp for an entireday, sucking on the digestive gland andmoving. The shrimp stayed alive!
A few freshwater crustaceans rate highlywith consumers or fishermen, and they harbor a variety of parasites. One of theseparasites from a crayfish will serve as anexample.
Cats and other mammals that feed on
crayfish (pronounced "crawfish" inLouisiana) from ditches may acquire a lungfluke (Paragonimus kellicotti). Figure 85shows the closely related oriental lung flukeof man (Paragonimus westermani) trans-
Grass Shrimp 45
:^V;,"V^f.--' -?£*-. •••:'-$*'.j'i ' •"
ii r* * •••"•%, /^•*•: ••••J& 4&f&, **''!*»!
#"'••j\\
;"v^* •• '" •rrl^lS? %- ••-'*' *
Figure 84. Three Individuals of a commonleech (Myzobdella lugubris) swaying abouton their grass shrimp host.
Figure 85. The oriental lung fluke (Para-gonlmus westermani) which is related to onethat encysts In pairs in the lungs of localcarnivores that feed on crayfish.
mitted by a few crabs in the Far East. Unlikemany intestinal flukes that infect localestuarine and terrestrial mammals, the locallung fluke can grow over 1 cm in length. Ithas provocative mate-finding behavior thatneeds to be critically investigated. Nevertheless, if two or occasionally moreindividual migrating larvae occur in thecat, they quickly join together and becomeencapsulated by the host in the lung withoutcausing much harm. On the other hand, if asingle worm infects a cat, it migrates aboutfor an extended period in the lung or lungcavity, presumably needing to come in contact with a mate and occasionally causing

46 Marine Maladies?
continuous damage to the host. Thus, ahost appears better off with several wormsthan with one. Like most investigateddigeneans, Paragonimus kellicotti appearsto cross-fertilize rather than fertilize itself.
According to veterinarians along theGulf Coast, this freshwater lung fluke is rarein pets except for outbreaks in Mobile,Alabama. Still, presence of the fluke shouldbe recognized. Pets that eat either dead orliving fish from the estuaries stand thechance of acquiring several differenthelminths.
VERTEBRATES AS HOSTS
FISHES
Because of the large number of fishes*present in the northern Gulf, I make noattempt to treat each species as thoroughlyas I covered symbionts of selected invertebrates. I picked only a few of the manyinvertebrates from the Gulf, but they are allcommercially important. A much largernumber of commercially important fin-fishes than shellfishes inhabits that region.The number of fishes one is likely toencounter commonly while shrimping andfishing probably would be about 100, butexerted effort should probably turn up over400 species. These fishes exhibit a widevariety of living and eating habits. Suchecological differences plus additional biochemical and physiological ones guaranteea variety of parasites.
In our Parasitology Section at the GulfCoast Research Laboratory, one of our investigations deals specifically withvarious aspects of parasites from theAtlantic croaker. From this single speciesin Mississippi, we have encountered about90 different species of parasites, about one-third of which occur commonly. Wide overlap of different hosts occurs for some speciesinfecting croaker, whereas others infect onlythe croaker or just a few host species.
Possibly a remark should be added forthose squeamish about parasites from fishin general. When one cleans a fish, he removes most of the parasites. Those that
remain in the flesh or under the skin may beeither easily visible or loo small to see withunaided eyes. In either case, most do notinfect man, but those that can die readilywhen cooked, even for short periods.
The following treatments should encourage an appreciation of some of thesevariously-adapted piscine symbionts. Mostwill be grouped by category of symbiont.The reader, however, should remember thatmany parasites utilize more than just a fishto complete their life histories, and some ofthose hosts have already been discussed. Inorder to help readers locate the site of a parasite, Figure 86 reveals many anatomical features of a bony fish in general and of theAtlantic croaker specifically. Because manysymbionts from elasmobranchs do not involve bony fishes (teleosts) and becauseHollywood has spurred recent interest insharks, elasmobranchs deserve some specialcomments.
About 800 species of elasmobranchs(sharks, skates, and rays) exist worldwide.Of these, at least 38 may occur during a partof the year in the northern Gulf of Mexicoand 26 are sharks. Many sharks can becaught near shore until the water temperature drops. Perhaps their prey movesoffshore or becomes otherwise unavailable
during winter. The bull shark, a commonspecies, exemplifies this by not turning upin nets or on fishermen's long lines* inPensacola, Florida, from November to mid-April. Sharks often aggravate bothcommercial and recreational fishermen bydamaging fishing gear and removing baitintended for other fishes. However, because
sharks are usually available, because of thethrill of catching a shark, and becausesharks taste good, many are caught andbrought ashore.
Different elasmobranchs feed on a varietyof fishes and invertebrates, including manycommercial species. This variation amongspecies is reflected by the variety of internalparasites they host. Large sharks tend tohave more individual specimens than stingrays, but the large rays, probably becausethey feed on a wider range of dietary itemsthan sharks, including crustaceans andmolluscans in addition to fish, usually hosta larger number of species.

Fishes 47
dorsal finsalll rackyrs
bum bladder caudal fin
$en$crybarbels I heart ITr^^f ff^<^«w/fci
urinary bladder
directed operculum t&eta °(axil cover)
Figure 86. Selected anatomical features of a bony fish (Atlantic croaker). Most fishes have similar,but modified, features.
organs of worms and therefore on the speciesof parasites present. Rather than possessinga simple tube for an intestine like mostteleosts, elasmobranchs have developed aspiral valve which increases the absorptivearea. This structure exists in a few different
forms. The concentric spiral valve incharcharhinid sharks (Figure 87) remindsme of a jelly roll whereas that in skates, rays,and other sharks looks more like a cork
screw.
Figure 87. Opening up the spiral valve of asmall blacktlp shark. This modified intestineincreases the amount of absorptive area andprovides an ideal habitat for tapeworms.
Many species of parasites, especially tapeworms, occur in one or a few species ofelasmobranchs. This specificity may relateprimarily to the kind of prey preferred by thedifferent hosts. Structures of the jaw andteeth usually govern the type of prey anelasmobranch can easily capture. Sharkswith sharp, pointed, or serrated teeth(charcharhinids) probably eat many fish,whereas sharks with flattened or small teeth
(mustelids and squalids) eat more crustaceans and molluscs. Modifications of the gutalso have some bearing on the attachment
Microbes
Lymphocystis exemplifies a viral diseaseof fishes which has external signs* that canbe readily observed (Figure 88). A specifictype of host cell (fibroblast*) becomes infected by the viral particles and may enlarge(hypertrophy) to a few hundred times itsoriginal size. Enlarged cells acquire a thickhyaline* capsule, and critical examinationof an affected fish will reveal these indivi
dual cells among crusty masses, or"tumorous growths." The disease cannot beassociated with cancer* (see tumor in
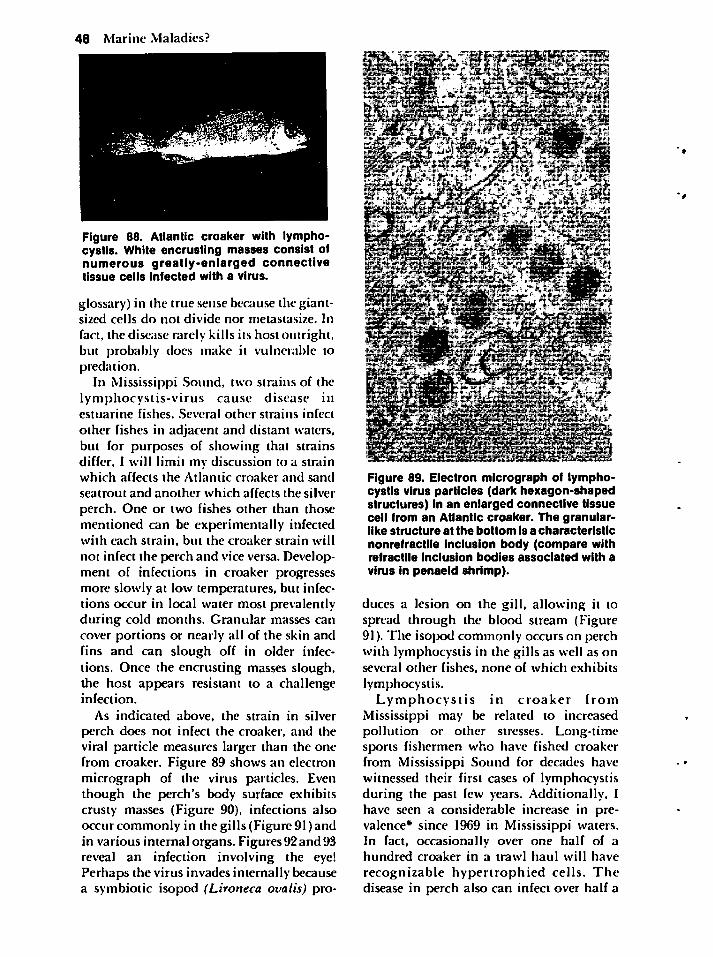
48 Marine Maladies?
Figure 88. Atlantic croaker with lymphocystis. White encrusting masses consist ofnumerous greatly-enlarged connectivetissue cells infected with a virus.
glossary) in the true sense because the giant-sized cells do not divide nor metastasize. In
fact, the disease rarely kills its host outright,but probably does make it vulnerable topredation.
In Mississippi Sound, two strains of thelymphocystis-virus cause disease inestuarine fishes. Several other strains infect
other fishes in adjacent and distant waters,but for purposes of showing that strainsdiffer, I will limit my discussion to a strainwhich affects the Atlantic croaker and sand
seatrout and another which affects the silver
perch. One or two fishes other than thosementioned can be experimentally infectedwith each strain, but the croaker strain will
not infect the perch and vice versa. Development of infections in croaker progressesmore slowly at low temperatures, but infections occur in local water most prevalentlyduring cold months. Granular masses cancover portions or nearly all of the skin andfins and can slough off in older infections. Once the encrusting masses slough,the host appears resistant to a challengeinfection.
As indicated above, the strain in silverperch does not infect the croaker, and theviral particle measures larger than the onefrom croaker. Figure 89 shows an electronmicrograph of the virus particles. Eventhough the perch's body surface exhibitscrusty masses (Figure 90), infections alsooccur commonly in the gills (Figure 91) andin various internal organs. Figures 92 and93reveal an infection involving the eye!Perhaps the virus invades internally becausea symbiotic isopod (Lironeca ovalis) pro-
Figure 89. Electron micrograph of lymphocystis virus particles (dark hexagon-shapedstructures) in an enlarged connective tissuecell from an Atlantic croaker. The granular-like structure at the bottom Is a characteristicnonrefractile inclusion body (compare withrefractile inclusion bodies associated with a
virus in penaeid shrimp).
duces a lesion on the gill, allowing it tospread through the blood stream (Figure91). The isopod commonly occurs on perchwith lymphocystis in the gills as well as onseveral other fishes, none of which exhibits
lymphocystis.Lymphocystis in croaker from
Mississippi may be related to increasedpollution or other stresses. Long-timesports fishermen who have fished croakerfrom Mississippi Sound for decades havewitnessed their first cases of lymphocystisduring the past few years. Additionally, Ihave seen a considerable increase in prevalence* since 1969 in Mississippi waters.In fact, occasionally over one half of ahundred croaker in a trawl haul will have
recognizable hypertrophied cells. Thedisease in perch also can infect over half a
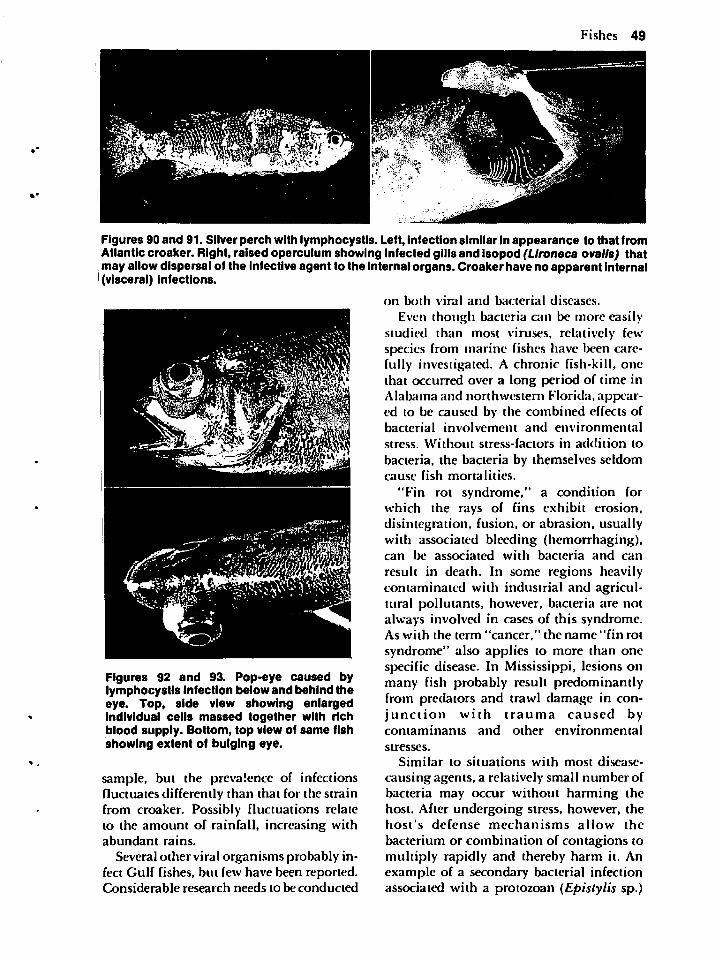
Fishes 49
Figures 90 and 91. Silver perch with lymphocystis. Left, infection similar in appearance to that fromAtlantic croaker. Right, raised operculum showing infected gills and isopod (LIroneca oralis) thatmay allow dispersal of the infective agent to the internal organs. Croaker have no apparent internal(visceral) infections.
on both viral and bacterial diseases.
Even though bacteria can be more easilystudied than most viruses, relatively fewspecies from marine fishes have been carefully investigated. A chronic fish-kill, onethat occurred over a long period of time inAlabama and northwestern Florida, appeared to be caused by the combined effects ofbacterial involvement and environmental
stress. Without stress-factors in addition to
bacteria, the bacteria by themselves seldomcause fish mortalities.
"Fin rot syndrome," a condition forwhich the rays of fins exhibit erosion,disintegration, fusion, or abrasion, usuallywith associated bleeding (hemorrhaging),can be associated with bacteria and can
result in death. In some regions heavilycontaminated with industrial and agricultural pollutants, however, bacteria are notalways involved in cases of this syndrome.As with the term "cancer," the name "fin rot
syndrome" also applies to more than onespecific disease. In Mississippi, lesions onmany fish probably result predominantlyfrom predators and trawl damage in conjunction with trauma caused bycontaminants and other environmental
stresses.
Similar to situations with most disease-
causing agents, a relatively small number ofbacteria may occur without harming thehost. After undergoing stress, however, thehost's defense mechanisms allow the
bacterium or combination of contagions tomultiply rapidly and thereby harm it. Anexample of a secondary bacterial infectionassociated with a protozoan (Epistylis sp.)
Figures 92 and 93. Pop-eye caused bylymphocystis Infection below and behind theeye. Top, side view showing enlargedindividual cells massed together with richblood supply. Bottom, top view of same fishshowing extent of bulging eye.
sample, but the prevalence of infectionsfluctuates differently than that for the strainfrom croaker. Possibly fluctuations relateto the amount of rainfall, increasing withabundant rains.
Several other viral organisms probably infect Gulf fishes, but few have been reported.Considerable research needs to be conducted

50 Marine Maladies?
Figures 94 and 95. Tidewater silverside withgas bubble disease. Top, side view. Bottom,top view emphasizing bulging nature of eyeand clearer view of a bubble. A variety offactors cause pop-eye and some arediscussed in the text.
will be discussed in the section on protozoans.
In most bacterial diseases, the presenceof bacteria has to be determined by culturingin order to diagnose a disease. The nonspecific "pop-eye" disease (technically called"exophthalmos") provides a good example.A variety of bacteria can cause this diseaserecognizable by a bulging eye. We,however, have already learned thatlymphocystis can cause eyes to bulge insilver perch, and many other factors alsocause a protrusion. Helminths cause it andso can mechanical injury or gas bubbles. Acase of "gas bubble" disease in a tidewatersilverside is illustrated by both top (dorsal)and side (lateral) views (Figures 94 and 95).The presence of gas does not rule outbacteria, but gas can be produced by over-aeration, malfunctioning organs, and othercauses. Because bacteria in finfishes that
could influence human health are the same
as those in shellfishes, fish from contaminated waters should also be cooked to insure a
safe product.
Protozoans in General
Many different groups of protozoansinfect fishes. The phylum* Protozoa bydefinition contains all single-celled animals, both parasitic and entirely free-living.Actually, many biologists presentlyworking with different nonrelated protozoans have decided that they should split upthe phylum into several phyla, therebyfurther emphasizing the differences amongnonrelated groups. Not until various stagesof selected species can be critically investigated with an electron microscope andmodern biochemical analyses, can manyscientists feel confident about the classifi
cation schemes. Many relationships stillneed clarification.
Even though single-celled, protozoanspossess a variety of specialized structuresthat aid in food gathering, locomotion,attachment, and protection. Some of thesestructures will become apparent in thefollowing sections. The sections also present a few of the numerous possibleexamples which portray a wide range infish-symbiont relationships. They includeflagellates, amoebas, sporozoans, microsporidans, myxosporidans, and ciliates.
FlagellatesA dinoflagellate acts as the most harmful
flagellate to captive marine fishes. Mostdinoflagellates occur in the plankton*,possess chlorophyll*, and do not parasitizehosts; they possess two flagella (read latercomments on cilia) which permit the linyorganism to spiral through the water. Onewraps around a subequatorial depression ofthe subspherical organism and the othertrails a long distance posteriorly. Actually,the parasitic species also produce a stagethat looks like a typical planktonicdinoflagellate mentioned above, and it isthat stage that allows proper classificationof the parasite. That stage, however, turnsinto opaque chalky blobs (Figures 96 and97) on the gills and skin of fishes.
A few different species of parasiticdinoflagellates occur in the Gulf of Mexico,but one (Amyloodinium ocellatum)happens to be especially pathogenic tocaptive fish. Careful examination reveals alarge nucleus, several food vacuoles, an
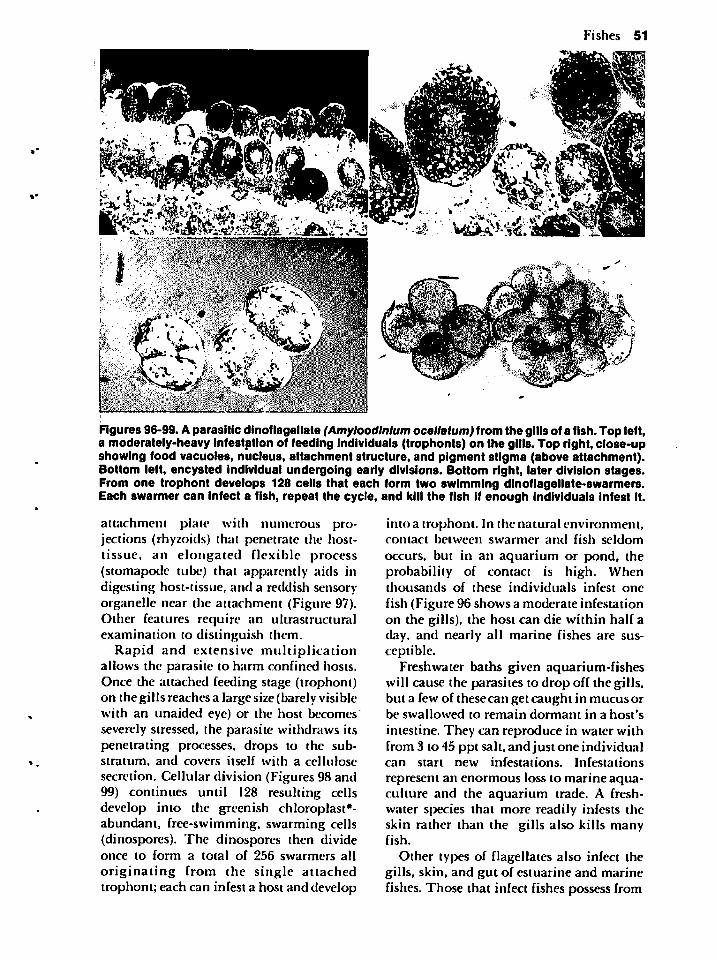
Fishes 51
Figures 96-99. A parasitic dinoflagellate (Amyloodlnlum ocellatum) from the gills of a fish. Top left,a moderately-heavy infestation of feeding Individuals (trophonts) on the gills. Top right, close-upshowing food vacuoles, nucleus, attachment structure, and pigment stigma (above attachment).Bottom left, encysted individual undergoing early divisions. Bottom right, later division stages.From one trophont develops 128 cells that each form two swimming dinoflagellate-swarmers.Each swarmer can infect a fish, repeat the cycle, and kill the fish if enough individuals Infest it.
attachment plate with numerous projections (rhyzoids) that penetrate the host-tissue, an elongated flexible process(stomapode tube) that apparently aids indigesting host-tissue, and a reddish sensoryorganelle near the attachment (Figure 97).Other features require an ultrastruciuralexamination to distinguish them.
Rapid and extensive multiplicationallows the parasite to harm confined hosts.Once the attached feeding stage (trophont)on the gills reaches a large size (barely visiblewith an unaided eye) or the host becomesseverely stressed, the parasite withdraws itspenetrating processes, drops to the substratum, and covers itself with a cellulosesecretion. Cellular division (Figures 98 and99) continues until I28 resulting cellsdevelop into the greenish chloroplast*-abundant, free-swimming, swarming cells(dinospores). The dinospores then divideonce to form a total of 256 swarmers all
originating from the single attachedtrophont; each can infest a host and develop
into a trophont. In the natural environment,contact between swarmer and fish seldom
occurs, but in an aquarium or pond, theprobability of contact is high. Whenthousands of these individuals infest one
fish (Figure 96 shows a moderate infestationon the gills), the host can die within half aday, and nearly all marine fishes are susceptible.
Freshwater baths given aquarium-fisheswill cause the parasites to drop off the gills,but a few of these can get caught in mucus orbe swallowed to remain dormant in a host's
intestine. They can reproduce in water withfrom 3 to 45 ppt salt, and just one individualcan start new infestations. Infestations
represent an enormous loss to marine aqua-culture and the aquarium trade. A freshwater species that more readily infests theskin rather than the gills also kills manyfish.
Other types of flagellates also infect thegills, skin, and gut of estuarine and marinefishes. Those that infect fishes possess from

52 Marine Maladies?
>
Figure 100. Trypanosome (Trypanoplasmabullocki) in blood of the southern flounder.Note the anteriorand posterior flagella. Also,note that fish red blood cells, unlike humanones, have nuclei.
one to eight flagella. Most occur in lownumbers in fish from natural waters, but can
rapidly intensify in stressed or cultivatedhosts. For most of those flagellates, theidentification and life histories are poorlyunderstood, but reproduction typicallytakes place on the host.
The blood of several fishes harbors a
different type of flagellate. The mostcommon blood flagellate in Mississippiestuaries (Trypanoplasma bullocki, Figure100) has two flagella, whereas some othershave one. Both flagella extending from thiselongated organism originate at its anteriorend. One borders a protruding undulatingmembrane and trails beyond it. Thisparasite, the body of which is longer thanthe host's blood cells, can best beappreciated when viewed alive. A drop ofblood from an infected southern flounder
placed under a coverslip reveals intenseundulating activity through a microscope.If a fish has trypanosomes, individuals canbe observed easily by allowing several dropsof blood to clot. The parasites swim about inthe clear serum*.
Unlike their mammalian counterpartswhich cause African sleeping-sickness,piscine trypanosomes rarely cause disease.One species has harmed hatchery-rearedrainbow trout in California.
Transmission of trypanosomes into fishoccurs when leeches obtain their blood
meal. In the case of Trypanoplasmabullocki in the southern flounder, sexualreproduction of the flagellate probably takesplace in a specific small leech (Calliobdella
vivida). In addition to being an invertebratehost, a leech can also be a passive vector* fortrypanosomes.
Amoebas
Commensal amoebas probably occur inintestines of Gulf fishes, but fishes have not
been adequately examined for them. At leastone species has invaded pond-reared freshwater fish in the southeastern U.S. and
caused serious disease. Implicatedmortalities may have required additionalinvolvement with bacteria or other
protozoans.
SporozoansA group of entirely parasitic protozoans
called Sporozoa have members that typicallyhave a spore* stage. Life cycles usuallyinclude alternation between one host in
which gametes* and asexually-producedindividuals are formed, and another host in
which gametes unite and spores with infective stages develop. Malaria-causing organisms belong to this group; fertilization and"spore" (nonresistant) formation occurs inmosquitoes, and the asexual phase takesplace in reptiles, birds, man, and othermammals.
Coccidians constitute the most studied
organisms with "resistant" spores (thosethat can withstand the elements when
passing from one host to another) because oftheir devastating effects on poultry andlivestock. Some species have intermediatehosts, whereas others bypass such hostscompletely and are transmitted by feedingon the spores. Figure 101 shows four sporesin a cyst (oocyst) taken from a lesion on thetail fin of a Gulf killifish in a Louisiana
marsh in Cameron Parish. Each sporecontains two organisms (sporozoites)presumably infective for other killifish.The number of spores and sporozoites percyst determines to what genus a speciesbelongs. The genus of the figured species(Eimeria sp.) also includes many speciesthat infect most farm-animals; however, itscyst wall is thinner because the host'saquatic environment is seldom threatened

Figure 101. Coccidian (Eimeria sp.) fromlesion on tail fin of the Gulf killifish. Eachthin-walled cyst contains four spores, andeach spore contains two infective organisms(sporozoites). Related members of this samegenus cause serious economic losses tountreated, infected, farm animals.
by desiccation. Unlike species in mostwarm-blooded vertebrates, all or most ofthose infecting fishes produce infectivestages while still in fish tissues and do notutilize an intermediate host.
Different piscine coccidians involvedifferent host tissues such as the testes of
menhaden and swim (air) bladder of cod.Medical doctors have misdiagnosed ashuman coccidian infections cases where
cysts were recovered in stools of peoplehaving eaten sardines and other fishesuncooked. These fish parasites do not harmman.
Related sporozoans (hemogregarines)infect peripheral red blood corpuscles ofelasmobranchs and bony fishes. Mostspecies presumably also infect blood cellsin bone marrow, liver, or elsewhere in thesame host, but these stages have not beenproperly described for a piscine hemogregarine; they have been for a turtle species.For some other species, however, asexualmultiplication occurs in peripheral blood.More prevalent in Mississippi fishes thantrypanosomes, hemogregarines may l>emore difficult to find in a cursory examination because thin blood smears must be
stained to observe the subspherical togreatly-elongated parasites (gametes*)which occupy a peripheral blcx>d cell(Figure 102). An individual southernflounder may have both a hemogregarine(Haemogregarina platessae) and a flagellate(Trypanoplasma bullocki). The same leechprobably transmits both parasites. Blood-
Fishes 53
Figure 102. Hemogregarine (Haemogre-garina platessae) in blood cells of southernflounder. Note that the parasite (a gametestage) is inside the red blood cell rather thanexternal to it. A pair of gametes fuse to form azygote, presumably in a leech.
Figure 103. Spot with tumorous growthscaused by a microsporidan protozoan(Pleistophora sp.). Insert shows microscopicview of spores.
feeders other than leeches may also transmitsome hemogregarines.
MicrosporidansAnother group of Protozoa, the Micro-
sporida, has been treated in some detailearlier when discussing the life cycle ofAmeson michaelis in the blue crab and the
spores of Inodosporus spraguei in grassshrimp. Several microsporidans infectfishes, but more appear to infect species inregions more temperate than the northernGulf. In the Northeast, winter flounder andshad are conspicuous hosts. In the Gulf,sciaenids (the croaker, spot, drums, andseatrouts) and a few other fishes host microsporidans, but cysts are usually small andinfections are much more prevalent incrustaceans and other invertebrates,including flukes. The conspicuous growth

54 Marine Maladies?
Figure 104. Myxosporidan from cyst in liverof striped mullet showing discharged polarfilaments.
on the spot in Figure 103 results from largemasses of a microsporidan (Pleistophorasp.). Some growths are hard, whereas othersare soft; all are infrequently seen.
MyxosporidansMyxosporidans occur much more
commonly than microsporidans in Gulffishes, and they comprise a quite unusualprotozoan group for several reasons. Froman evolutionary point of view,Myxosporida does not fit in with otherprotozoan groups because members havemore than a single cell. Depending on thespecies under consideration, the actualnumber of cells per spore differs, but aspore of most piscine species develops fromfive cells.
From an anatomical point of view,myxosporidans possess internal polarcapsules which evert an elongated filament(Figure 104) similarly to the various structures (stinging, adhesive, anchoring, andentangling) everted from capsules found injellyfishcs and other cnidarians.
From a host-symbiont point of view,myxosporidans can be divided into twoprimary types: those that infect lumensof hollow organs such as the gall bladder(coelozoic) and those that infect tissuessuch as muscles (histozoic). Rather thanoccurring in spaces between muscle cellslike a histozoic form in the strict sense, somespecies develop within muscle fibers(cytozoic).
Tumorous* growths caused by a histozoicmyxosporidan (Myxobolus lintoni) provide a dramatic example (Figure 105). Notprevalent in the Gulf, the disease, which
• ••,•*• .••'' • '••••. *••- •. , "".. • • • :.'<:•.*%'-. •. ..
*••••.• ^vV-i-Sffj*:'!-"'.''JM >.:••• J ,. • • •
l*fei r •
Bfl&^^gP. p,mr.s /'•'/•
;•• ,r : , lit'.:, ••.•.••.•:.•'••.•,. •;•:••:••(• EM*.:- .•:•::.'.'. ;i • .,
.
i- •• '•.•:•:• ..;•;»-...-. ... ..*-:,.•.. . .
Figure 105. Sheepshead minnow withtumorous growths caused by myxosporidan(Myxobolus lintoni). As in the microsporidaninfection from spot, the cysts containmillions of infective spores.
affects the sheepshead minnow at least fromMississippi to Texas, may be related topollution or other stresses. It has othersimilarities to cancer in addition to possiblybeing induced by a pollutant. Vegetativegrowths invade and replace healthy muscletissue and new growths spread to otherregions of the body. A related species(Myxobolus cerebralis) invades and erodescartilage supporting the central nervoussystem of salmonid fishes. Hatchery-rearedrainbow and brook trout seem most sus
ceptible to "whirling disease." the resultof heavy infections. Infected fish can bespotted quickly because their tail regionturns black; neurological control ofchromatophores* becomes impaired.
Several histozoic species infect muscle,gill, liver, skin, cartilage, and other tissue inGulf fishes. One worth mentioning becauseit can be readily seen in commercially important fishes without a microscope formscysts between fin rays of some seatroutsand related species. Figure 106 showsseveral cysts of this species (Henneguyasp.) in a spot. As with most species, millionsof spores fill each cyst. In members of thisgenus, the two joined spore-valves havelong posterior extensions (tails) (see insertof Henneguya sp. in Figure 106).
Coelozoic or cavity forms comprise alarger group of parasites, but fish must bedissected in order to detect them. Some large,flat, elliptical forms (trophozoites) can beseen through a gall bladder's wall, whereassmears of bile or other fluid must be micro
scopically examined to detect others. Gall
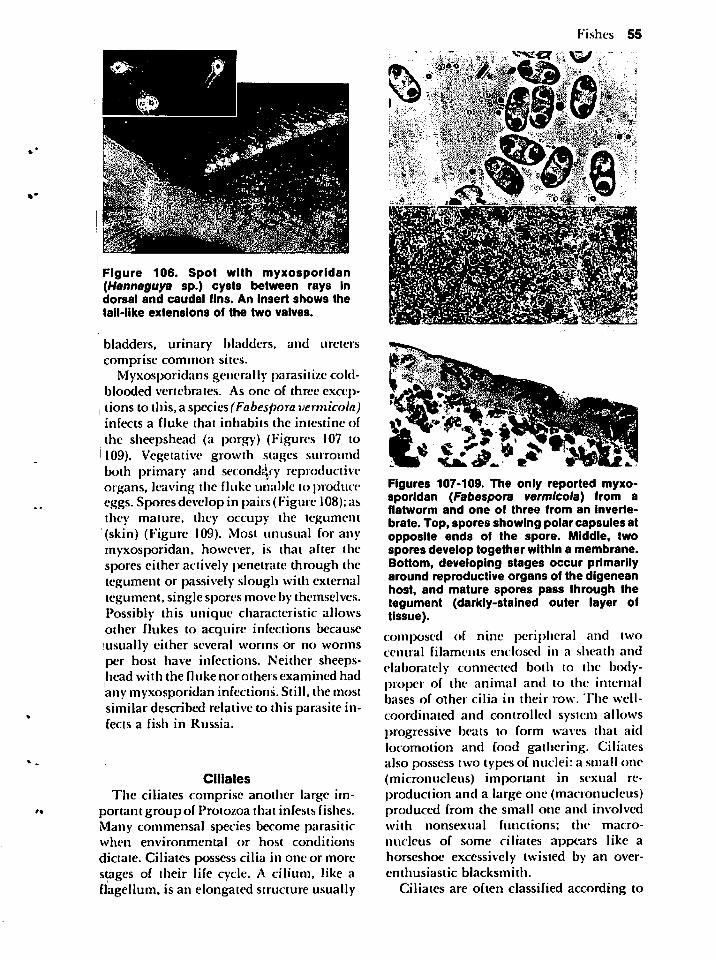
Figure 106. Spot with myxosporidan(Henneguya sp.) cysts between rays indorsal and caudal fins. An insert shows the
tail-like extensions of the two valves.
bladders, urinary bladders, and ureterscomprise common sites.
Myxosporidans generally parasitize coldblooded vertebrates. As one of three exceptions to this, a species (Fabespora vermicola)infects a fluke that inhabits the intestine of
the sheepshead (a porgy) (Figures I07 lo109). Vegetative growth stages surroundboth primary and secondary reproductiveorgans, leaving the fluke unable to produceeggs. Spores develop in pairs (Figure 108);asthey mature, they occupy the tegument(skin) (Figure 109). Most unusual for anymyxosporidan, however, is that after thespores either actively penetrate through thetegument or passively slough with externaltegument, single spores move by themselves.Possibly this unique characteristic allowsother flukes to acquire infections becauseusually either several worms or no wormsper host have infections. Neither sheepshead with the fluke nor others examined hadany myxosporidan infections. Still, the mostsimilar described relative to this parasite infects a fish in Russia.
Ciliates
The ciliates comprise another large important group of Protozoa that infests fishes.Many commensal species become parasiticwhen environmental or host conditions
dictate. Ciliates possess cilia in one or morestages of their life cycle. A cilium, like aflagellum, is an elongated structure usually
Fishes 55
&k4••*;£j
Figures 107-109. The only reported myxosporidan (Fabespora vermicola) from aflatworm and one of three from an inverte
brate. Top, spores showing polar capsules atopposite ends of the spore. Middle, twospores develop together within a membrane.Bottom, developing stages occur primarilyaround reproductive organs of the digeneanhost, and mature spores pass through thetegument (darkly-stained outer layer oftissue).
composed of nine peripheral and twocentral filaments enclosed in a sheath andelaborately connected both to the body-proper of the animal and to the internalbases of other cilia in their row. The well-
coordinated and controlled system allowsprogressive beats to form waves that aidlocomotion and food gathering. Ciliatesalso possess two types of nuclei: a small one(micronucleus) important in sexual reproduction and a large one (macronucleus)produced from the small one and involvedwith nonsexual functions; the macro-
nucleus of some ciliates appears like ahorseshoe excessively twisted by an over-enthusiastic blacksmith.
Ciliates are often classified according to

56 Marine Maladies?
their feeding apparatus. Peritrichs have along tubular buccal cavity lined with spiralling cilia leading toward the mouth, as wellas some stouter cilia in rows encircling thetop of the cavity. Many peritrichs attachpermanently to their host or other substratum by a stalk, disk, or lorica (discussedearlier with crustacean ciliates), whereasother motile ones may attach only temporal ily. All living peritrichs provideimpressive spectacles under a microscope.
Most coastal fishes in Mississippi harborat least one motile species of peritrich(members of Trichodina or a related genus).The ciliates occur primarily on the gills, butalso infest the skin. In contrast, most of
those that infest freshwater fishes, and thereare many, preferentially infest the skin.Normally few individuals infest a fish, butsome fishes typically host both more individuals and more species than others. TheFlorida pompano and striped mullet eachcommonly host at least two species. Juvenilepompano placed in a garbage can filled withaerated seawater and examined daily for gilland skin protozoans will acquire increasingly dense populations of trichodinids.Apparently reproducing extensively whentheir hosts become crowded and stressed,they may be undergoing a "normal survivalprocedure" in order to assure a fewindividuals an opportunity to infestadditional hosts should theirs die.
Trichodinids may attract curiosity because of the ring of interlocking teeth(denticles) on the ventral surface. Figures110 and 111 show these denticles for two
different species. One photograph displaysa live ciliate and the other shows a preservedone stained to accentuate the denticles.
Devastating as these denticles might appear,they do not harm the host; they support thedisk like a flexible skeleton. Ciliary movements permit an individual to glide over orhover close to its host. On the other hand,the peripheral border of the basal diskmembrane can pinch and even dislodge hosttissue. In this manner, the ciliate can harmits host, especially if the fish is young or hasdense populations on its gills. Pathologicalconditions usually involve hosts in rearingfacilities. Many estuarine and marineciliates have not been named, possibly
!J ••**.>;,>?l . * •" .< ^
• «... '.'••W!:
i' •; •mm
Figures 110 and 111. Trichodlnid ciliateparasites from estuarine fishes. Top, livespecimen. Bottom, stained (impregnationwith silver nitrate) specimen of anotherspecies to accentuate the ring of "teeth"(denticles) on the ventral surface. Theseciliates can cause mortalities in reared fish.
because of the detailed study necessary todelineate the patterns of cilia for the different stages.
Attached peritrichs have also beenimplicated in fish mortalities. One species(Epistylis sp.) has, within the last few years,infested an increasingly large number ofbluegill, bream, bass, and marine fishesthat invade fresh or nearly freshwater coastalMississippi bayous. The disease derives itsname, "red sore," from the bloody lesionsassociated with this long-stalked, colonialciliate.
Red sore disease portrays a good exampleof both a disease associated with pollutionand a disease resulting from interactionsbetween two organisms. Fish with extensivered sores have a systemic* bacterial infection(Aeromonas hydrophila) that can kill thehost. I believe the ciliate which is invadinghost tissue acts as the primary invader, allowing secondary entrance by the bacteriuminto the blood where it can proliferateextensively. The bacterium can also enterby other routes, even in fish without theciliate.

Some pollutants seem to enhance reproductive capabilities of Epistylis sp.Hundreds of colonies of this organismproduce a fuzzy growth which appears likecotton or fungi. Fish farmers find that anincrease in organic materials in their pondspromotes an increase in red sores. In somelow salinity and freshwater bayous thathave, over the past few years, increased theirconcentration of pollutants or decreaseddieir flow of water, the prevalence of redsore disease has increased dramatically. Infact, fishing in poorly-flushed regions can.during certain seasons, produce a preponderance of bluegill with red sores. Suchcatches have kept some fishermen fromreturning to particular favorite holes wherethey have fished for decades. Infections havekept some from fishing at all.
Holotrichous ciliates do not form a
natural (phylogenetically* related) group;most have simple uniform body cilia and nospecialized cilia around the mouth. The bestknown marine "holotrich," at least to
aquarists, is the marine counterpart(Cryptocaryon irritans) of the ireshwaterspecies that causes "ich," a shortened termused by tropical fish fanciers to designateIchthyophthirius multifiliis. Both the saltwater and freshwater species cause elevations on the gills and skin consisting of nestsof these ciliates eating away at tissuesjust under the outer layer of skin. Theyirritate the host and promote secondarybacterial infections. As in the life cycle of thedinoflagellate Amyloodinium ocellatum,the release of one feeding individualultimately gives rise to numerous infectiveorganisms, each adding to disorder in anaquarium or pond. Most aquarium-remedies affect the short-lived swarmingstage; therefore, treatment should continueuntil all feeding individuals (trophonts)leave the fish.
Cryptocaryon irritans affects a varietyof hosts, but is rare in nature. Most otherholotrichs do not share its parasitic modeof life; however, under appropriateconditions, some free-living species invadefish and cause damage. Reported cases haveinvolved freshwater species primarily, butconfined marine hosts possess the samevulnerability.
Fishes 57
Free-living ciliates in other groups canalso turn parasitic. A hypotrich (a speciesof Euplotes or closely-related genus)damaged gills and skin of several specimensof the sheepshead minnow collected fromHorn Island and maintained for a month in
an aquarium. The opercula and snout ofinfested fish developed bloody lesions, theirnormal swimming behavior modified intojerky and gasping motions, and they died,either from secondary infections or directlyfrom the ciliate.
TapewormsTapeworms (cestodes) form a group of
worms which exhibits two striking features.The worms have no alimentary canal, andthey have an elongated "segmented" body,thereby giving an impression of a measuring tape, hence their common name"tapeworm." As adults they inhabit tubularregions like an intestine with its highnutritional level, attaching to their hostwith a holdfast organ (scolex). This organmay be simple or ornamented with a varietyof hooks, suckers, and other structures, andit represents the major feature used forclassification. Each segment of the "tape"usually possesses one set of reproductiveorgans (a proglottid), and in most species afew segments are continuously beingformed.
In general, few marine bony fishes (teleosts) in the northern Gulf host adulttapeworms. Examples are a worm(Anantrum tortum) from the inshorelizardfish and those (ptychobothriids) fromneedlefishes and related species. The
Figure 112. An inshore lizardfish portrayingits viscera and the adult tapeworms(Anantrum tortum) from the intestine of asimilar-sized fish. Worms also occur inIllustrated fish.

58 Marine Maladies?
Figure 113. Life cycle of the tapeworm (Poecllanclstrlum caryophyllum) that causes "wormytrout." The adult develops in the bull shark (or other related sharks). Released segments from theadult disperse eggs, each with a larvathat infects a crustacean which in turn is eaten by a fish. Wheneaten by seatrout and related fishes, the larva develops further in flesh and becomes infectivefor sharks.
lizardfish worm looks more like a twisted
Christmas ribbon than a measuring tape.The normally twisted body possesses aseries of reproductive organs, but they arenot compartmentalized into segments.Without any specialized holdfast organ, thisworm (Figure 112) often maintains itsposition by winding in clumps of severalindividuals at bends in the intestine or bysticking an expanded or folded-over"scolex" into a pyloric caecum. The position in the gut appears related to externalwater temperature. As the temperature increases, an increased percentage of wormslocates near the anus. If a fresh lizardfish
comes into possession, see whether a relatively large creamy-colored worm occupiesthe intestine. Watch it writhe in a corkscrew
fashion within a mixture of one or more
parts full strength seawater and three parts
freshwater (the one-fourth seawater solutionapproximates the host's body fluidsand willmaintain most helminths for several hours).
Even though marine teleosts have fewadult cestodes, they harbor a rich variety oflarval ones. They act as intermediate hostsfor the stage (plerocercoid) that infects thefinal host. With a few exceptions, theselarvae mature in specific elasmobranchs.The perfect example of this causes "wormytrout." Indeed, when coastal residents,especially those who catch and fillet marineand estuarine fishes, are asked if they knowabout any parasites in fishes, most respondfirst and some respond exclusively aboutthis trout-worm (Poecilancistriumcaryophyllum). To regress a bit, fish harbormany parasites, but even the most pathogenic is rendered harmless by cooking. Thecommon one in the trout does not even need

Fishes 59
Figures 114-117. Larval stages of tapeworm (Poecllanclstrlum caryophyllum) that causes "wormytrout." Top left, egg with enclosed coracidium showing cilia and booklets. Top right, early development (procercoid) in copepod (Tlgriopus callfomlcus). Bottom left, a more-developed larval stage.Note two individuals are present. Bottom right, subsequent larval stage (plerocercoid in blastocyst) in flesh of spotted seatrout. These organisms have been lifted some from the flesh to makethem more obvious, but most fillets will have 80% of the worms obvious. Usually about two wormsinfect a fish, but cutting the caudal extension of a cyst gives the impression of many worms beingpresent.
heating to become harmless to people.Unfortunately, too many people feel disgust at these infections.
"Wormy trout" refers to the presenceof the entwined fleshy stage (blastocyst) thatharbors an encased infective larva (plerocercoid) at its swollen end. As seen in thediagram (Figure 113), sharks (the bullshark and related species) acquire infectionsby eating seatrout (or less often a few otherrelated fishes including redfish, croaker, anddrum). Within the intestine and spiral valve,the worm matures, reaching about 20 cmlong and producing eggs in attached proglottids. When the worm or a batch ofsegments passes into seawater, a ventralcleft in a segment splits, allowing eggs(Figure 114) to disperse. After about a week,a tiny swimming larva (coracidium) popsout of the capped egg. If eaten within 2 daysby an appropriate crustacean, another larvalstage (procercoid) develops (Figures 115and
116). This host in turn must be eaten by anappropriate fish, usually a seatrout (seeFigures 117and 118).The worm apparentlycan live in the trout for a few years. Mostinfected sharks measure longer than 135 cm(45$ feet) long, presumably because largersharks eat more seatrout.
Even though some fishermen think theircatch had 20 or so worms per fish, fewindividual fish have more than 3 or 4. The
elongated, twisted, fleshy extension may becut into several pieces, giving the illusionof many worms. Apparently, once a fishbecomes infected, it acquires resistance* tofuture infections. Examination of a largenumber of spotted seatrout suggests that theworms do not harm adult fish. However, nofish less than 14 cm and few less than 25 cm
long had infections, suggesting either thatyoung fish do not get infected or that the fishget killed. Larvae in small fish occasionallyencroach on vital organs, giving some sup-

60 Marine Maladies?
Figure 118. Plerocercoid of Poecllanclstrlumcaryophyllum removed from fleshyblastocyst. Note the four hooked tentaclesand the sheaths that contain the tentacleswhen they are not protruded. Constriction ofthe bulbs at the posterior of the attachmentorgan (scolex) protrudes them.
port to the latter possibility. Perhaps theworm is detrimental to good fishing. It isnot detrimental to good eating!
Another "fish tale" needs rectifying.Wormy trout occur year around becauseonce a host becomes infected, the\vormscanremain for a few years. When average salinity levels are high, usually duringsummer, the percentage of fish infected isalso high. However, the percentage ofinfected fish during other seasons may farsurpass that percentage encountered duringa summer for which the average salinity waslow. More spotted seatrout have heavierinfections in the clean high saline water ofApalachee Bay, Florida, (an average of 4.4worms in 98% of fish over 25 cm and a maxi
mum of 16 worms in a fish) than in anyother region sampled. High saline waterprobably supports more potential intermediate hosts. An average of two worms infectsabout 40% of the eating-sized fish in Texas,Louisiana, and Mississippi. In pollutedTampa Bay, Florida, only 1.3 worms perfish infected 10% of the trout.
Figure 119. Close-up of tentacles of Poecllanclstrlum caryophyllum from "wormyseatrout." Note the different shapes andsizes of hooks, the muscle in the sheath thatretracts the tentacle, and the darkenedciliated pits at the anterior margin. Thesensory pits protruded and actively movedabout when the fixed worm was alive.
The above cestode plerocercoid belongs tothe trypanorhynch group and is diagnosedprimarily by the presence of four eversiblehooked tentacles (proboscises) (Figure 119).Contraction of muscles around the fluid-
filled proboscis bulb protrudes the tentacle(Figure 118). The hooks then allow thesetentacles to attach in the host gut. Anotherrelated trypanorhynch (Dasyrhynchus sp.)in the crevalle jack (Figure 120) can be observed commonly infecting the head and lessoften occurring elsewhere in the flesh. Theplerocercoid of many species infects crustaceans and an earlier example (Prochristianella hispida, Figures 70 and 72) represents an extremely abundant one.
Tentacle-hooks come in all sizes and
shapes, sometimes with considerable varietyon a single tentacle. Long spikes with smalldistal barbs (Figure 121) characterize aspecies (Pterobothrium heteracanthum)from the muscles and body cavity of Atlantic

croaker, and a variety of hooks (Figure 122),including a stout recurved one, occurs onanother species from the kidney of a striped
Figure 120. Individuals of a cestode larva(plerocercoid of Dasyrhynchus sp.) beingcut out from tissue of the crevalle jack. Theblastocyst has a long thin tail (caudalextension) and contains the plerocercoid.
Fishes 61
mullet. Hooks shaped like rose-thorns arethe most common type on trypanorhynchs.
Another cestode group (Tetraphyllidea)infecting local elasmobranchs has memberswith four similar muscular holdfast organswithout tentacles on the scolex. Each mayhave hooks, spines, suckers, or folds toattach to the gut. In order to appreciatethe variety of these holdfasts, some photographs are presented. Note the foldedmargins (Figure 123) on one from theAtlantic stingray (Phyllobothriumfoliatum). In lihinebothrium corymbum(Figure 124) from the southern stingraythere are 27 grooves (loculi) on each pedunculated bothridium. Rhinebothrium
maccallumi, also from the southern stingray, has fewer (Figure 125). Members of thegenus Acanthobothrium have triloculatebothridia, each with two forked hooksanteriorly. Compare the scolex (orAcanthobothrium paulum (Figure 126) from the
Figures 121 and 122. Variation intypesof hooks on the tentacles of trypanorhynch tapeworms. Leftshows small barbs on thin spikes (Pterobothrlum heteracanthum from flesh of Atlantic croaker).Right shows more than one type. Inaddition to numerous thin hooks, some robust ones shaped likea latch-hook provide the most conspicuous form (ICaltltetrarhynchus sp. from kidney of stripedmullet).

62 Marine Maladies?
Figures 123-125. Variety in scotices of nonhooked tetraphyllideans. Left, Phyllobothrlum sp. fromspiral valve of Atlantic stingray. Middle, Rhlnebothrium corymbum from a southern stingray hasmore "loculi" on each of the four bothridia than Rhlnebothrium maccalluml (right) from the samehost.
Figures 126-128. Two related species of the hooked tetraphyllidean tapeworm Acanthobothrlum.Left, scolex (attachment organ) of Acanthobothrlum paulum from spiral valve of southern stingray. Middle and right, mature proglottid (the reproductive segment) and scolex of Acanthobothrlum brevlsslme from the bluntnose stingray. Note the divided bothridia of both species and thebend (abnormal) in the hooks of the latter species occurring in both sets of hooks. The hooks alsobend on two unseen opposing bothridia.
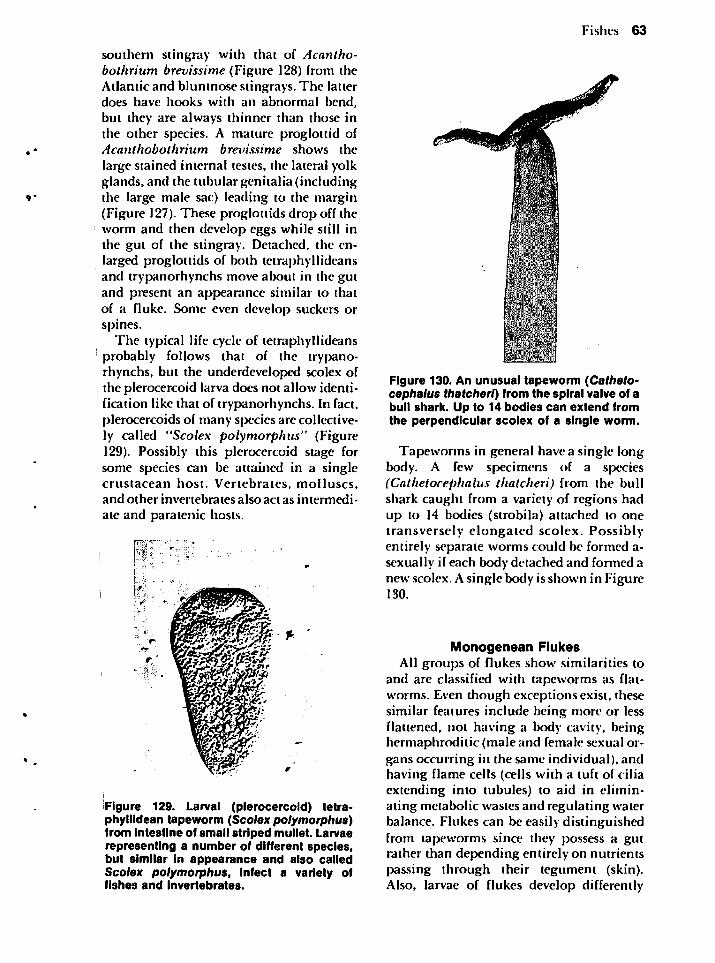
southern stingray with that of Acantho-bothrium brevissime (Figure 128) from theAtlantic and bluntnose stingrays. The latterdoes have hooks with an abnormal bend,but they are always thinner than those inthe other species. A mature proglottid ofAcanthobothrium brevissime shows the
large stained internal testes, the lateral yolkglands, and the tubular genitalia (includingthe large male sac) leading to the margin(Figure 127). These proglottids drop off theworm and then develop eggs while still inthe gut of the stingray. Detached, the enlarged proglottids of both tetraphyllideansand trypanorhynchs move about in the gutand present an appearance similar to thatof a fluke. Some even develop suckers orspines.
The typical life cycle of tetraphyllideansprobably follows that of the trypanorhynchs, but the underdeveloped scolex ofthe plerocercoid larva does not allow identification like that of trypanorhynchs. In fact,plerocercoids of many species are collectively called "Scolex polymorphus" (Figure129). Possibly this plerocercoid stage forsome species can be attained in a singlecrustacean host. Vertebrates, molluscs,and other invertebrates also act as intermedi
ate and paratenic hosts.
Figure 129. Larval (plerocercoid) tetraphyllidean tapeworm (Scolex polymorphus)from Intestine of small striped mullet. Larvaerepresenting a number of different species,but similar in appearance and also calledScolex polymorphus, infect a variety offishes and invertebrates.
Fishes 63
Figure 130. An unusual tapeworm (Catheto-cephalus thatcherl) from the spiral valve of abull shark. Up to 14 bodies can extend fromthe perpendicular scolex of a single worm.
Tapeworms in general have a single longbody. A few specimens of a species(Cathetocephalus thatcheri) from the bullshark caught from a variety of regions hadup to 14 bodies (strobila) attached to onetransversely elongated scolex. Possiblyentirely separate worms could be formed a-sexually if each body detached and formed anew scolex. A single body is shown in Figure130.
Monogenean FlukesAll groups of flukes show similarities to
and are classified with tapeworms as flat-worms. Even though exceptions exist, thesesimilar features include being more or lessflattened, not having a body cavity, beinghermaphroditic (male and female sexual organs occurring in the same individual), andhaving flame cells (cells with a tuft of ciliaextending into tubules) to aid in eliminating metabolic wastes and regulating waterbalance. Flukes can be easily distinguishedfrom tapeworms since they possess a gutrather than depending entirely on nutrientspassing through their tegument (skin).Also, larvae of flukes develop differently
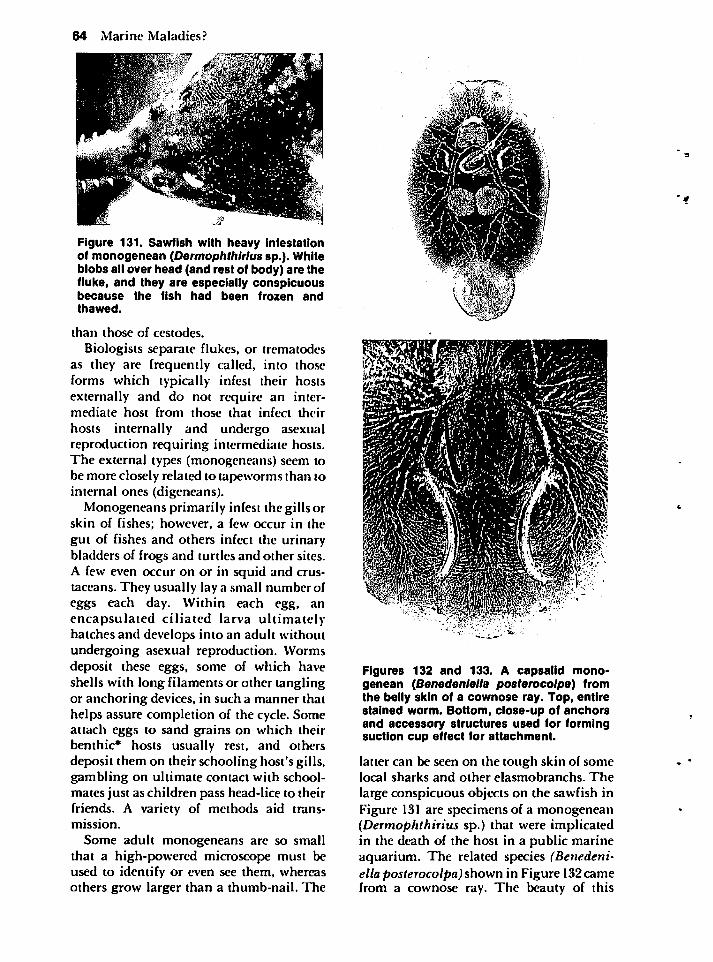
64 Marine Maladies?
Figure 131. Sawfish with heavy infestationof monogenean (Dermophthlrius sp.). Whiteblobs all over head (and rest of body) are thefluke, and they are especially conspicuousbecause the fish had been frozen andthawed.
than those of cestodes.
Biologists separate flukes, or trematodesas they are frequently called, into thoseforms which typically infest their hostsexternally and do not require an intermediate host from those that infect their
hosts internally and undergo asexualreproduction requiring intermediate hosts.The external types (monogeneans) seem tobe more closely related to tapeworms than tointernal ones (digeneans).
Monogeneans primarily infest the gills orskin of fishes; however, a few occur in thegut of fishes and others infect the urinarybladders of frogs and turtles and other sites.A few even occur on or in squid and crustaceans. They usually lay a small number ofeggs each day. Within each egg, anencapsulated ciliated larva ultimatelyhatches and develops into an adult withoutundergoing asexual reproduction. Wormsdeposit these eggs, some of which haveshells with long filaments or other tanglingor anchoring devices, in such a manner thathelps assure completion of the cycle. Someattach eggs to sand grains on which theirbenthic* hosts usually rest, and othersdeposit them on their schooling host's gills,gambling on ultimate contact with schoolmates just as children pass head-lice to theirfriends. A variety of methods aid transmission.
Some adult monogeneans are so smallthat a high-powered microscope must beused to identify or even see them, whereasothers grow larger than a thumb-nail. The
Figures 132 and 133. A capsalid monogenean (Benedenlella posterocolpa) fromthe belly skin of a cownose ray. Top, entirestained worm. Bottom, close-up of anchorsand accessory structures used for formingsuction cup effect for attachment.
latter can be seen on the tough skin of somelocal sharks and other elasmobranchs. The
large conspicuous objects on the sawfish inFigure 131 are specimens of a monogenean(Dermophthirius sp.) that were implicatedin the death of the host in a public marineaquarium. The related species (Benedeni-ella posterocolpa) shown in Figure 132camefrom a cownose ray. The beauty of this

common worm has been accentuated bystaining. In life, the transparent milky-colored worm on the belly of the ray can beeasily overlooked without critical examination. It attaches to its host using the largecentral anchors and small marginal hookson the large posterior organ (haptor, Figure133). It feeds with its anteriorly-locatedmouth.
Smaller monogeneans, but those stilllarge enough to observe with naked eyes,occur commonly on gills of fishes in theGulf. Jacks, drums, seatrouts, sharks, andmany other fishes often have them. Theseexhibit a reddish or blackish appearancecaused in part by ingested host blood or itsbreakdown products. Asa general rule, these(polyopisthocotylids) feed on blood insteadof tissue and mucus like the others (mono-pisthocotylids). Rather than having anchorsand hooks like those monopisthocotylidsdiscussed above from elasmobranchs, most
of these polyopisthocotylids have memberscharacterized by having clamps or suckers.Members of one family (Hexabothriidae)from sharks have large curved hooksassociated with suckers (Figure 134).
At the risk of leading the reader intothinking monogeneans are not primarilyparasites of fishes, an example of those withclamps also exemplifies a hyperparasitethat does not always infest fish. Largespecimens of this species with stalkedclamps (Choricotyle aspinachorda, Figure135) typically attach near the bases of thelegs of a symbiotic isopod (Cymothoaexcisa) lodged in the throat cavity (frequently on the tongue or gill rakers) of a
Figure 134. Holdfast (haptor) of a monogenean (Heteronchocotyle leucas) from thegills of a bull shark. Note the large curvedhooks (sclerites) associated with thesuckers.
Fishes 65
pigfish. Often when a pigfish is not infestedby the cymothoid isopod, it still hasimmature specimens of the monogenean onthe gills. Primarily large adult flukes favorthe isopod, and they deposit clumps of eggson it. The combined worms and eggs on theisopod present a flowery appearance. The
Figures 135 and 136. Monogeneans fromsymbiotic isopods on fish. Top, entire worm(Choricotyle aspinachorda) from isopod(Cymothoa excisa) In throat of pigfish. Noteposterior holdfast organ with eightpedunculated clamps. Bottom, close-up of aclamp from a related worm (Choricotyleloulslanensls) on isopod in southernkingfish.

66 Marine Maladies?
close-up of a clamp (Figure 136) actuallybelongs to a closely related worm(Choricotyle louisianensis) which has asimilar relationship with a cymothoidisopod on the southern kingfish (alsocommonly called ground mullet). Next timeone of these fiesty bottom feeders is caught,a reader should look for the isopod and itssymbionts.
Relatives of the isopod monogeneans thatinfest gills of fish occasionally causemortalities, but usually only when infestingconfined fish. Fishes such as the com
mercially valuable Florida pompanoacquire infestations heavy enough tosuccumb. Most relatives of the first men
tioned type (monopisthocotylids) on bonyfish, and there are many, never reach a sizevisible to the naked eye. These tiny tissue-feeding species comprise a much biggerthreat than polyopisthocotylids to confinedhosts, including freshwater fishes. If presentin an aquarium or small pond, however,infestations can usually be controlled with avariety of chemical treatments. Figure 137shows a couple of individuals in conjunction with a parasitic dinoflagellate stressingtheir ailing host.
In a manner similar to that of the
monogeneans on the isopod from the fishthroat, another species (Udonellacaligorum) often associates with copepods(usually species of Caligus) on which itattaches its eggs by filaments (Figure 138).Unusual because it lacks hooks or anchors,this monogenean can be commonly
Figure 137. Two small monogenean flukesmoving about near the attached parasiticdinoflagellate (Amyloodlnlum ocellatum) onthe gills of a sheepshead. Either of these twoparasites could kill a captive fish If enoughindividuals occur at one time.
Figure 138. Filamented eggs from amonogenean fluke (Udonella caligorum) ona copepod (Caligus praetextus) on the seacatfish. The segmented appearing structureat the lower right is the copepod's egg sac.
observed on caligids from the gills and skinof local sea catfish (hardhead catfish),seatrout, and striped mullet (popeyemullet). Apparently cosmopolitan, it infectsa wide range of hosts.
Monogeneans make good biological indicators (tags) because a single monogeneanspecies usually limits its infestations to asingle host species (see host-specificity inglossary). Many fish can be identified on thebasis of their parasites! Evolutionaryrelationships among fishes can also besuggested on the basis of these parasites.Even though not accepted by all ichthyologists, diskfishes (remoras and shark suckerswhich have their spinous dorsal finmodified into a laminated disk-mechanism
allowing attachment of the fish to turtles,sharks, rays, and large fish) seem to be closely related to thecobia (also called lemonfish,ling, and a variety of other names). Asparasitic evidence for this relationship,species of one family of monogeneans(Dionchidae) infest exclusively the cobiaand diskfishes. Since dionchids infest no

other noncaptive hosts and since membersof the two fish families host no other
monogeneans, a long period of isolation issuggested. Several other groups of monogeneans infest primarily members of certainfamilies, indicating strong evolutionarytrends.
Dlgenean FlukesDigenean flukes exhibit considerably
more variety in their adaptations thanmonogeneans and quite possibly more thanany other helminth group. Many differentgroups of animals serve as intermediatehosts for the larvae. In addition to fishes,adult digeneans infect amphibians, reptiles,birds, mammals, and occasionally crustaceans and molluscs. In fishes, most speciesinfect the alimentary tract, but a few encystin flesh, lodge in blood vessels, or occupyother sites. More species in birds andmammals than in fishes occupy spaces otherthan the alimentary tract. Some of these arcthe kidneys, air spaces, and the bile ducts.
Almost all digeneans utilize a snail orbivalve mollusc as a first intermediate host.
An earlier diagram (Figure 28) alreadyportrayed a complete cycle of a snail to crabto raccoon pattern, and more cycles will bediscussed in this section. In general, asexualreproduction takes place in the mollusc.This reproduction involves a few differentkinds of larvae. The larva from the egg infects the mollusc and produces a stage thateither yields additional individuals of theidentical type which then produce free-living* larvae (cercariae) or another stagewhich also produces the same kind offree-living larvae. The route of infection andkind of larvae depend on the species considered. Cercariae of some species crawl orencyst in the host mollusc without leavingit. Most, however, have long tails allowingthem to swim or to attract predators. Somepenetrate their hosts, some encyst externally, and some are ingested directly. Often, thetime of day, amount of light, temperature,tidal phase, or water pressure controlsrelease of these larvae. Once in or on their
proper host (or vegetation or some otherobject), the tail, if present, sheds and theminute larva (metacercaria) forms acapsular cyst, with or without more cyst
Fishes 67
Figures 139 and 140. The giant stomachworm (Hlrudlnella ventricosa) from "blue-water" fishes. Top, contracted, preservedspecimens from wahoo. Bottom, moreelongated preserved specimens from littletunny; living worms can extend further thanan opened hand. Note the two suckers. Thedark color represents breakdown productsof blood obtained from feeding on the host'sstomach.
layers added by the host. The metacercaria ofa few species develops without cysts(unencysted metacercaria), but examples ofthis type represent exceptions. Afterdeveloping for an adequate period, the larvacan infect its final host. Occasionally, it candebilitate or alter its intermediate host in
such a manner that the final host has easier
access to individuals that are infected: some
infected amphipods turn reddish or pale,making them more visible to final hosts,some snails do not burrow as deeply as uninfected ones, and some fish cannot swim asfast or as long as those without cysts. Thosespecies without special means to alter theintermediate host's behavior may have toproduce many more larvae in order to assurethat enough larvae will mature to completethe cycle.
One trematode that fishermen who fish
for billfish and other pelagic fish should beaware of is the giant stomach worm(Hirudinella ventricosa). Fishermen mayknow it because of its large size. Figure 139

68 Marine Maladies?
shows some preserved contracted ones("walnut worms") from the stomach of awahoo, whereas those in Figure 140 fromlittle tunny ("bonita") exhibit less contraction. When alive, the fluke can elongatemuch farther than shown. What appears tobe the same or at least similar species infectsa variety of "bluewater" fishes. When in thewahoo, usually only two individuals arepresent. This common occurrence of twoworms in fish shorter than 160 cm has
allowed many a wagering fisherman to wina six-pack of beer. One should use caution,however, because tunas and other fishes donot usually possess two per fish.
Most flukes have two suckers. A central
one primarily holds on, whereas an anteriorone operates in feeding, holding, andmoving. These suckers can be seen in thephotos and in most of the digenean illustrations in this guide. In addition to the
Figures 141 and 142. A sectioned digenean(Crasslcutls archosargl) expressing itsability to modify its tegument adjacent to thehost's (sheepshead) Intestinal tissue so thatit expands, fills all available space, andadheres firmly. Top, cross-section throughlevel of the fluke's testis. Bottom, especiallystained (periodic acid Schlff with diastase)worm-tegument showing positive reaction.Most digeneans attach to their host by theirventral sucker.
suckers, one species (Crassicutis archosargi)from a sheepshead can modify its tegumentinto an adhesive surface (Figures 141 and142).
The line drawings of three species(Figures 143 to 145) portray examples ofseveral features. The one on the left
(Lecithaster leiostomi) comes from theintestine of spot and a few other fishes. Unlike the related Hirudinella ventricosafrom billfish stomachs, it does not need athick skin (tegument) for protection fromstomach acid and from churning foodparticles. Also, Lecithaster leiostomi seldomreaches 1 mm in length. Most members of itsfamily (Hemiuridae) inhabit the stomach'slumen, but other exceptions exist. In fact,another small species(Satumius maurepasi)even dwells in the tissue under the lining ofthe striped mullet's "gizzard" (pyloricstomach). The middle worm (Glaucivermisspinosus) is a little shorter and has a thinnertegument than Lecithaster leiostomi, butalso infects the intestine (of the southernkingfish). The worm on the right(Neochasmus sogandaresi) is about twicethe size and infects the striped bass. It hasseveral relatives that infect a wide range offreshwater and marine fishes. One relative
without the spines about the mouth(Metadena spectanda) ranks among themost common, but seldom seen, animals inthe estuaries. For example, during Februaryof 1971 an average of 54.3 individuals infected each of 53% of the Atlantic croaker
examined near Ocean Springs that had theworm (90% of the fish larger than 7 cm wereinfected). Because croaker during thatperiod comprised the most common fishbesides the bay anchovy and many specimens of the fluke infected most of them,that fluke should be recognized as acommon constitutent of the animal
community. In fact, other fishes also hostthe worm! During the past couple of years,however, the population density of both thecroaker and digenean have decreased.
The three illustrated digeneans representdifferent families, but all have small eggs(about 30 u, or 0.030 mm, long). Severalother digenean groups have members witheggs several times larger. As a general rule,large eggs produce larvae (miracidia) that
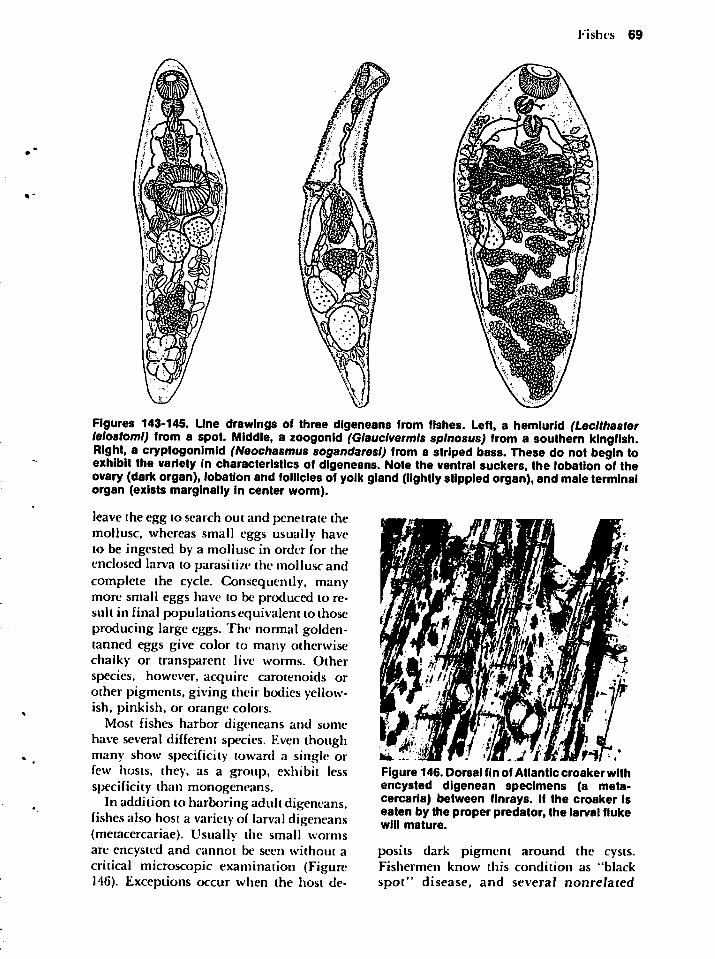
Fishes69
Figures143-145.Linedrawingsofthreedigeneansfromfishes.Left,ahemiurid(Lecithasterleiostomi)fromaspot.Middle,azoogonid(Glauclvermlssplnosus)fromasouthernkingfish.Right,acryptogonimid(Neochasmussogandaresl)fromastripedbass.Thesedonotbegintoexhibitthevarietyincharacteristicsofdigeneans.Notetheventralsuckers,thelobationoftheovary(darkorgan),lobationandfolliclesofyolkgland(lightlystippledorgan),andmaleterminalorgan(existsmarginallyincenterworm).
leavetheeggtosearchoutandpenetratethemollusc,whereassmalleggsusuallyhavetobeingestedbyamolluscinorderfortheenclosedlarvatoparasitizethemolluscandcompletethecycle.Consequently,many-moresmalleggshavetobeproducedtoresultinfinalpopulationsequivalenttothoseproducinglargeeggs.Thenormalgolden-tannedeggsgivecolortomanyotherwisechalkyortransparentliveworms.Otherspecies,however,acquirecarotenoidsorotherpigments,givingtheirbodiesyellowish,pinkish,ororangecolors.
Mostfishesharbordigeneansandsomehaveseveraldifferentspecies.Eventhoughmanyshowspecificitytowardasingleorfewhosts,they,asagroup,exhibitlessspecificitythanmonogeneans.
Inadditiontoharboringadultdigeneans,fishesalsohostavarietyoflarvaldigeneans(metacercariae).Usuallythesmallwormsareencystedandcannotbeseenwithoutacriticalmicroscopicexamination(Figure146).Exceptionsoccurwhenthehostde-
Figure146.DorsalfinofAtlanticcroakerwithencysteddigeneanspecimens(ametacercaria)betweenfinrays.IfthecroakerIseatenbytheproperpredator,thelarvalflukewillmature.
positsdarkpigmentaroundthecysts.Fishermenknowthisconditionas"black
spot"disease,andseveralnonrelated

70 Marine Maladies?
Figure 147. Black spot resulting frompigmentation (melanization) around larvalcysts of a strlgeoid fluke in the tidewatersilverside. A variety of freshwater and marineparasites can cause black spot.
digeneans cause it. In the southern U.S.,most cases of black spot result from strigeoidinfections. Most strigeoid species utilize afreshwater snail and a fish-eating bird in thecycle. The tidewater silverside from theestuaries in Mississippi, however, alsopossesses a strigeoid species. The one in thephoto (Figure 147) came from thePascagoula estuary. One large easilyvisible hemiurid (Stomachicola magnus)encysts in the flesh and elsewhere in a varietyof fish. The white (sand) seatrout often hasmany. If the curious finder pricks at thisblack object in a fresh fillet, out will popa pinkish-colored worm.
Normally, chromatophores (Figure 148)expand, spreading out their associatedmelanin to give a fish a dark color.Contraction produces a pale color. Otherchromatophores bear different pigmentsfor different colors. Most diseases in whichpigments are deposited, however, involvemelanin.
Less easy to see than the above hemiurid,a small heterophyid (Phagicola longus) infects the flesh and viscera of mullets alongthe Gulf and southeastern Atlantic states.
A diagram (Figure 149) shows the life cycle.In addition to herons, a large number ofother birds and mammals, including man,can probably act as final hosts. Although nohuman infections have been confirmed,
many people throughout the world whoeat raw fish harbor zoonotic* species relatedto this tiny worm. Most infections producesymptoms hardly more serious thandiarrhea and mild irritation; however,
serious complications occasionally develop.
Figure 148. Melanophores In the skin of afish. These are part of a complex system ofchromatophores which regulate colors andcolor changes in fish. This piece of sectionedtissue had an overabundance of pigmentsurrounding a lesion.
Heterophyids have many similaritieswith the microphallids already discussedearlier as digeneans infecting the blue craband shrimp. Small species infect birds andmammals, often in very large numbers. Asample of 14 brown pelicans from Louisiana averaged about 12,000 specimens ofPhagicola longus per bird. Both digeneanfamilies have a large number of species, and,in both groups, the second intermediate hostis usually more specific than the final one.Adult heterophyids appear very similar inappearance to microphallids, but minordifferences in their genitalia and otherfeatures set them apart. Differences in theirfree-living larvae (cercariae) accentuate thedifferences even more. From a more practical point of view, heterophyids use fishesas second intermediate hosts and micro
phallids use crustaceans.The key silverside shown in Figures 150
and 151 has damaged fin rays caused by anencysted metacercaria (of an acanthostome).One important fact concerning these infections is that the fish is rare and considered
endangered. Found in a pond in the FloridaKeys, the fish has to contend with a parasitein addition to man's encroaching developments. The final link in the cycle, either theAmerican crocodile or the American
alligator, also has status as an endangeredspecies.
Another group of digeneans or closelyrelated worms is the aspidogastrids. Thesehave a conspicuous ventral loculated holdfast organ and a life history lacking asexualreproduction. Although prevalent in some
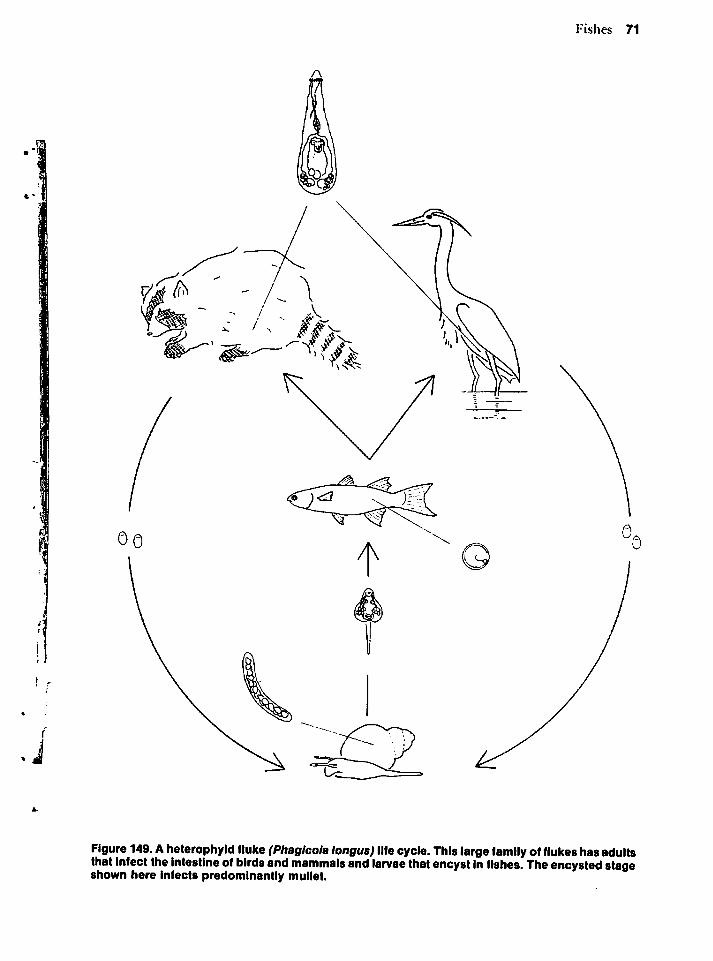
Fishes 71
Figure 149. A heterophyid fluke (Phagicola longus)lifecycle. This large family of flukes hasadultsthat Infect the intestine of birds andmammals andlarvae thatencystin fishes. Theencystedstageshown here infects predominantly mullet.

72 Marine Maladies?
Figures 150 and 151. Two specimens of thekey silverside infected with cysts of themetacercaria of an acanthostome digenean.Bottom, close-up view showing nature ofdamage to caudal fin rays. Cysts also occurelsewhere on these and other species of fish.A crocodilian completes the cycle for theparasite.
freshwater clams and turtles, aspidogastridsinclude species that also infect marinefishes. The most common Gulf species (Lo-batostoma ringens) occurs in the intestineof a variety of fishes, but if a person reallydesires to obtain a specimen, he (used in theantiquated sense to include also females)will have the best chance by examining aFlorida pompano. Fish apparently acquirethe worm by feeding on coquina clams inthe surf, but it may be harbored by othermolluscs.
The most attractive aspidogastrid(Multicalyx cristata) resides in the bileducts of large rays, sawfish, and otherelasmobranchs. Growing over 10 cm long,the narrow pale yellow body contrastssharply with a bright reddish holdfastorgan. The elasmobranchs may acquireinfections by feeding on bony fish withjuvenile worms (checkered puffer, southernkingfish, and others) which in turn pickedup the worms from a mollusc. A preadultspecimen (Cotylogaster basiri) from therectum of the sheepshead illustrates anaspidogastrid (Figure 152). An earlier study
Figure 152. A preadult aspidogastrid(Cofy/ogasfer basiri) from the rectum of thesheepshead. Presumably acquired from amollusc, this worm matures in a few fishesincluding the sheepshead.
showed completion of one aspidogastridcycle utilizing a freshwater clam as the onlynecessary host and the ability for turtlesfeeding on the clam to maintain an infection; many biologists assumed that speciesto be representative of the group. Actually,a vertebrate appears to be necessary formaturation of most species, includingCotylogaster basiri and Lobatostomaringens, rather than just an adjunct hosthelpful as a dispersing agent.
Roundworms
"Roundworm" is a common name for anematode. Not all nematodes parasitizeanimals. Some infect plants and others livefree without hosts in water or soil. Theyall have separate sexes, a digestive tract, alough cuticle (outer layer of worm), andinternal longitudinal muscles. The reasonfor mentioning this last feature is thatnematodes, unlike earthworms, and caterpillars, move in a whiplike fashion. Mostother animals that are called "worms" havea layer of circular muscles that allow acoordinated inchworm-type of locomotion.

Fishes 73
Figures 153-155. Some views of two adult roundworms from the intestine of fish. Left, anteriorendof camallanid (Splrocamallanus cricotus) showing spirally ribbed buccal capsule used for attachment to gut. The capsule is hard and golden-tan to orangish In life and followed by a long muscularesophagus. Middle, a male tail of the same worm showing the winglike flaps (caudal alae) withsensory papillae used in grasping the female during mating. Right, anterior end of ascaridoid(Thynnascaris Inqules) from a cobia. Attachment by this worm is achieved by the three large lips.The distinct ringed cuticle is not as dominant in related species.
The size and arrangement of the longitudinal body musculature in nematodesdictates the type of slashing movement itwill have. Before reading this guide, a readermay have never thought about parasiticnematodes, even though he may havepondered "vinegar eels" (Turbatrix aceti)thrashing about in plant juices or vinegar.Those small free-living worms had beenincorrectly credited in the past with theacetic properties of vinegar.
Parasitic nematodes occupy a widervariety of habitats than most worms in fish.The adult forms in fishes occur mostlyalong the alimentary tract, whereas mostlarval forms inhabit the mesentery. Theselarvae usually have thin cystsencapsulatingthem. One family of nematodes worthmentioning for its unusual locations is thePhilomeiridae. Members of this groupusually appear reddish. Some reside in thebody cavity, and others occur in the gonads,between fin rays, under the skin, and inmuscle tissue. Rather than releasing eggslike most nematodes, these worms releaseyoung larvae. Philometrids infect a varietyof local fishes including the Atlanticcroaker, redfish, tripletail, and southernflounder.
A few intestinal worms also deposit larvae rather than eggs. An especially notableone also has a reddish color. It was men
tioned earlier as a nematode larva in penaeidshrimp. This camallanid (Spirocamallanuscricotus, Figures 153 and 154) infects theintestine of at least 13 different estuarine
and nearshore fishes. When an intestine of a
fish is swollen, filled with liquid, or partially transparent, these relatively largeworms (up to 3.2 cm long) can be seenwiggling within the uncut gut. The reddishworm's activity, especially when severaloccur together, readily distinguishes it fromany food contents.
Why is Spirocamallanus cricotus red?Many worms, and other animals includingthe beautiful flamingo acquire their reddishcolor from components in their diets(mostly carotenoids). Such is not the case forSpirocamallanus cricotus and some otherreddish nematodes. Unlike most nematodes,these worms have hemoglobin* just likeman and fishes. Properties of the hemoglobins from Spirocamallanus cricotus,however, differ some from those of its
Atlantic croaker host. The worm attaches to
the gut wall with its golden colored,spiralled, scleritized* buccal capsule (Figure

74 Marine Maladies?
153) and feeds on host blood. The wormquickly digests the host blood, and a possibility exists that its components help toform the worm's own hemoglobin.Strangely though, properties of thishemoglobin in males differ from those infemale worms.
Fish can acquire infections of this reddishcolored nematode by feeding on copepodintermediate hosts. The penaeid shrimpsalso act as intermediate hosts and quitepossibly as paratenic (transfer) hosts.I have seen worms in the common small
squid (Lolliguncula brevis), and otheranimals may also serve as paratenic hosts. Aparatenic, or transfer, host is one in whichno development to the next stage occurs, andparatenic hosts have and will be discussedagain under other sections. This type hostshould not be confused with a reservoir
host* in which any stage could occur.Spirocamallanus cricotus takes several
months to mature and can live in a fish for
longer than a year. Since individuals growrelatively slowly, they probably do notcause excessive mortalities in juvenile fish.Several specimens crowded into one fishmight cause disease, and related species havekilled their hosts. The ripe female of one ofthese (Camallanus oxycephalus) sticks itsreddish body out the anus of a striped bass orother of its hosts. These can be commonlyseen in estuarine bayous and freshwaterhabitats of both Mississippi and Louisiana.When fully gravid, the protruded femaleruptures explosively and releases her larvae.Copepods serve as the intermediate host,and small fishes serve as paratenic ones.
Different groups of nematodes attach totheir host using different anterior structures.Philometrids do not develop a very elaborateanterior end because they embed in tissue.The camallanids have scleritized bivalve-
shaped or barrel-shaped buccal capsules,and adult ascaridoids possess three stronglips.
Common ascaridoids from local Gulf
fishes include members of the genusThynnascaris. Figure 155 shows the powerful lips of a species (Thynnascaris inquies)from the stomach and pyloric caeca of thecobia (lemonfish). Actually, the conspicuously-ringed cuticle constitutes an
atypical feature for members of that genus,but not for members of a related genus.Thynnascaris inquies, often present in thehundreds in a large cobia, occasionallycauses ulcerated lesions at sites where several
individuals attach clustered together.Other ascaridoids occur in the northern
Gulf, and one (Thynnascaris reliquens) isoccasionally common in inshore fishes.This worm, up to 13 cm long, may alsooccur in the hundreds in the intestine and
elsewhere along the alimentary tract ofsheepshead and a few other hosts. In severalother hosts, however, only few specimensoccur. When a host has been caught andlanded or otherwise placed under severestress, the worms vacate the host, leavingthrough the mouth, gill cavities, andanus, hence the Latin name reliquens referring to the abandonment. Additionalspecies in martin and other billfish alsoleave in mass. Such an activity, disgusting asit may appear, should have no bearing onwhether a person eats the fish. Even thoughsome species may be harmful to man, theescaping stage is easily visible, it is an adult,the worms are discarded when removing viscera, and their eggs need several days beforebecoming infective and then not to people.Nevertheless, cooking the fish alleviates anypotential threat. More information onpublic health aspects occurs below.
Something should be said about the sexlife of nematodes, since reproduction accounts for most of their expended energy.The figured copulating pair shows thecurled tail of the male wrapped about thefemale, the inserted right spicule, and a
Figure 156. A male nematode (Thynnascarisreliquens) mating with the female. A mucoidband girdles the female immediatelyposterior to her genital opening.
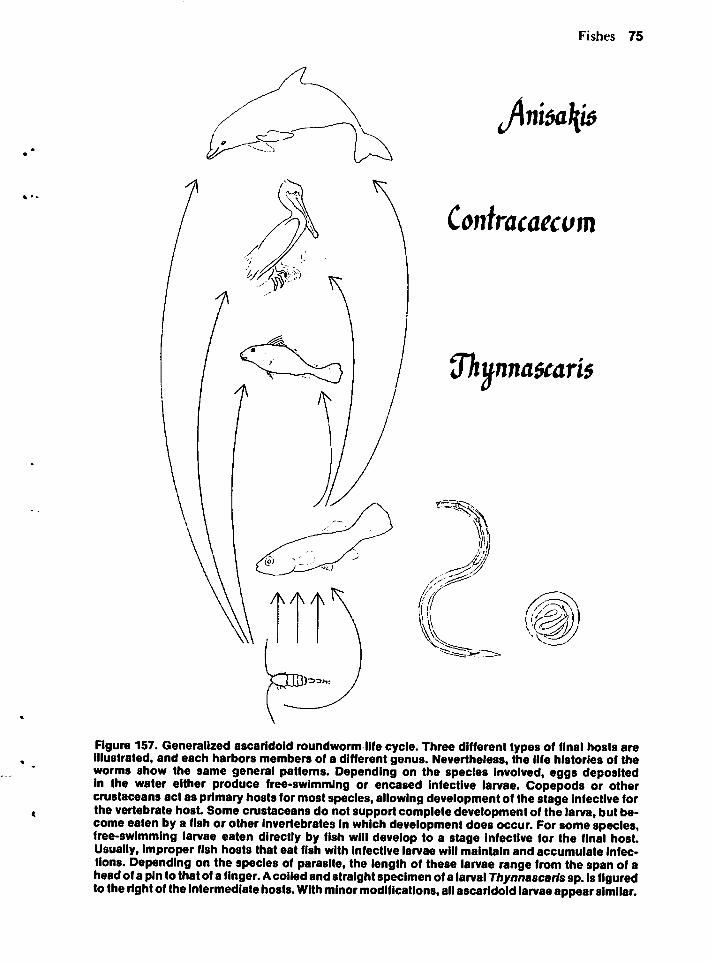
Fishes 75
J\ni$a\i$
Contracaecvm
7hunna$cari$
Figure 157. Generalized ascaridoid roundworm life cycle. Three different types of final hosts areillustrated, and each harbors members of a different genus. Nevertheless, the life histories of theworms show the same general patterns. Depending on the species Involved, eggs depositedin the water either produce free-swimming or encased infective larvae. Copepods or othercrustaceans act as primary hosts for most species, allowing development of the stage infective forthe vertebrate hosL Some crustaceans do not support complete development of the larva, but become eaten by a fish or other invertebrates in which development does occur. For some species,free-swimming larvae eaten directly by fish will develop to a stage infective for the final host.Usually, improper fish hosts that eat fish with infective larvae will maintain and accumulate infections. Depending on the species of parasite, the length of these larvae range from the span of aheadof a pinto thatof a finger. Acoiledandstraightspecimen of a larval Thynnascaris sp. is figuredto the right of the intermediate hosts.With minormodifications, all ascaridoid larvae appearsimilar.

76 Marine Maladies?
mucoid band about the female (Figure 156).The male's tail in this worm and all other
nematodes has some type of a modificationto cup or clasp the female. It is usuallylined ventrally with a specific pattern ofsensory papillae (see Figure 154 of the maletail of Spirocamallanus cricotus). Onceproperly aligned, the inserted spicule orspicules allow passage of sperm to fertilizethe ova. The position of the female openingand other aspects of both the female andmale reproductive systems vary considerablyamong different groups of worms and comprise important taxonomic characters.
While still concerned with ascaridoidsand their biology, I will discuss general lifecycles of species of three different genera.Mammals (porpoise), birds (pelican), andfishes (Atlantic croaker) act as hosts formembers of Anisakis, Contracaecum, andThynnascaris, respectively, as exemplifiedin Figure 157.Actually, several species existin each of these three and related genera ofparasites, and most probably have similarcycles. The major difference among thethree relates to the members' infecting amammal, a bird, or a fish, respectively, astheir definitive hosts. Eggs are released withfeces from the host. Usually an egg developsin a few days, and a small larva is released. Ifa proper crustacean feeds on the wigglinglittle larva, the worm develops into a stageinfective for the final host. Alternatively, ifan appropriate fish feeds on the free-swimming larva of some species, the wormcan develop without the copepod. On theother hand, some copepods do not promotedevelopment in the worm, but still allow afish to acquire the larva by feeding on them.Whether infective as a free larva or withina crustacean, some species can complete thecycle to maturity by utilizing a fishintermediate. Although usually not highlyhost-specific, an ascaridoid will not infectnor develop in all fishes. In order to assurecompletion of cycles, however, many fishesand invertebrates can act as paratenic hosts.As discussed earlier, they acquire infectivelarvae by feeding on other hosts with thesame stage larva. No development takesplace in the new host, but accumulation andtransference assure a greater supply of infective larvae in the entire habitat. Once an
intermediate or paratenic host iseaten by theproper definitive host, the worm matures indie alimentary tract and deposits eggs tostart another cycle.
Larval species of Anisakis and othergenera infective to marine mammals act aspotential threats to public health and theprimary cause of marine nematodezoonoses*. Even though species belongingin genera that normally infect non-mammalian hosts may infect man, they arenot as large and most are harmless. At leastone species of Thynnascaris from bothfishes and invertebrates can infect laboratory mice, and it was discussed earlier as ashrimp parasite. Fortunately, fewmammalian-type larvae occur in the inshorecoastal regions.
Less dangerous to man, but neverthelessdiscomforting to fishermen, are largespecies of larval Contracaecum. Thesecream-colored to yellowish to reddishworms coil up or occur straight, either freeor encysted in their hosts. The species indifferent mullets (Contracaecum "robust-um") most often infects their kidneys andliver. A related species in the redfish and afew other hosts infects the same sites, butalso commonly resides in the mesentery. Because an individual of both species canextend over 2 cm when uncoiled and the
worms heavily infect some fish, they constitute a logical cause for unnecessary worry.The tested worms have not infected experimental mammals, are usually discardednormally when cleaning fish, can be easilyavoided, and are killed with heat.
Past history, including the great depression of the 1930's did much to influence the
likes and dislikes of eating mullet on theGulf Coast. Since smoked mullet was a primary protein source for many years, main-people still prepare mullet in this and a variety of other ways. For this same reason,many people would be happy never to seeanother mullet or can of Spam. Actually, inthe 1800's many inland farmers made annual fall trecks to the Florida coast to catch
and salt large quantities of mullet to lastthrough the year. Before that, Indians madesimilar annual trips to catch, salt, andsmoke the fish. If fresh mullet is cleanedimmediately, I recommend it highly.
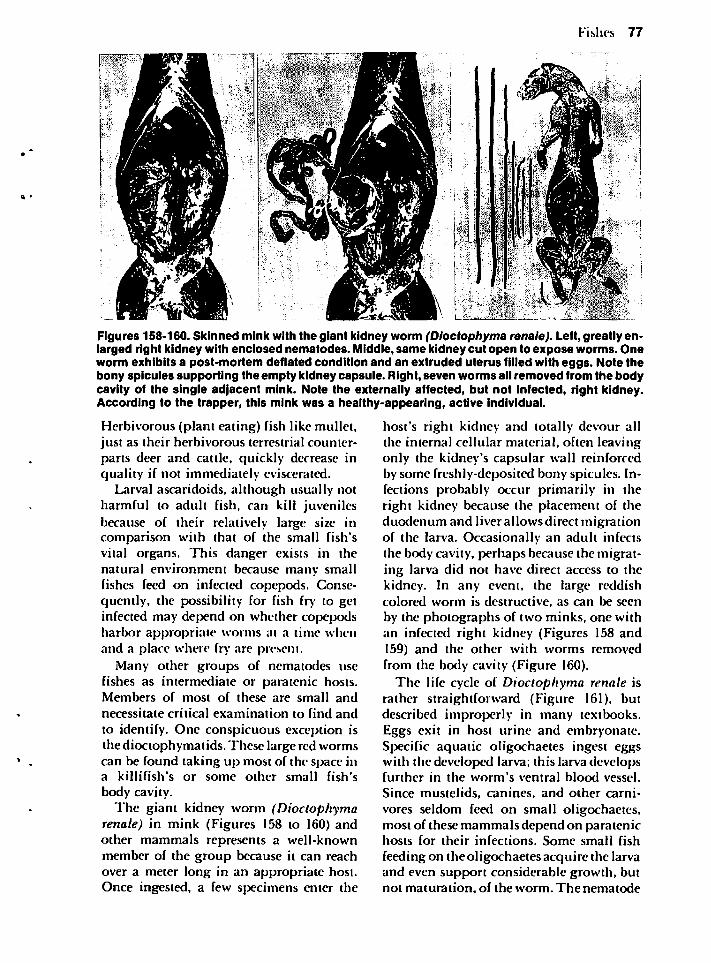
Fishes 77
Figures 158-160. Skinned mink with the giant kidney worm (Dloctophyma renale). Left, greatly enlarged right kidney with enclosed nematodes. Middle, same kidney cut open to expose worms. Oneworm exhibits a post-mortem deflated condition and an extruded uterus filled with eggs. Note thebony spicules supporting the empty kidney capsule. Right, seven worms all removed from the bodycavity of the single adjacent mink. Note the externally affected, but not infected, right kidney.According to the trapper, this mink was a healthy-appearing, active individual.
Herbivorous (plant eating) fish like mullet,just as their herbivorous terrestrial counterparts deer and cattle, quickly decrease inquality if not immediately eviscerated.
Larval ascaridoids, although usually notharmful to adult fish, can kill juvenilesbecause of their relatively large size incomparison with that of the small fish'svital organs. This danger exists in thenatural environment because many smallfishes feed on infected copepods. Consequently, the possibility for fish fry to getinfected may depend on whether copepodsharbor appropriate worms at a time whenand a place where fry are present.
Many other groups of nematodes usefishes as intermediate or paratenic hosts.Members of most of these are small and
necessitate critical examination to find and
to identify. One conspicuous exception isthe dioctophymatids. These large red wormscan be found taking up most of the space ina killifish's or some other small fish's
body cavity.The giant kidney worm (Dioctophyma
renale) in mink (Figures 158 to 160) andother mammals represents a well-knownmember of the group because it can reachover a meter long in an appropriate host.Once ingested, a few specimens enter the
host's right kidney and totally devour allthe interna] cellular material, often leavingonly the kidney's capsular wall reinforcedby some freshly-deposited bony spicules. Infections probably occur primarily in theright kidney because the placement of theduodenum and liver allows direct migrationof the larva. Occasionally an adult infectsthe body cavity, perhaps because the migrating larva did not have direct access to thekidney. In any event, the large reddishcolored worm is destructive, as can be seen
by the photographs of two minks, one withan infected right kidney (Figures 158 and159) and the other with worms removedfrom the body cavity (Figure 160).
The life cycle of Dioctophyma renale israther straightforward (Figure 161), butdescribed improperly in many textbooks.Eggs exit in host urine and embryonate.Specific aquatic oligochaetes ingest eggswith the developed larva; this larva developsfurther in the worm's ventral blood vessel.
Since mustelids, canines, and other carnivores seldom feed on small oligochaetes,most of these mammals depend on paratenichosts for their infections. Some small fish
feeding on the oligochaetes acquire the larvaand even support considerable growth, butnot maturation, of the worm. The nematode
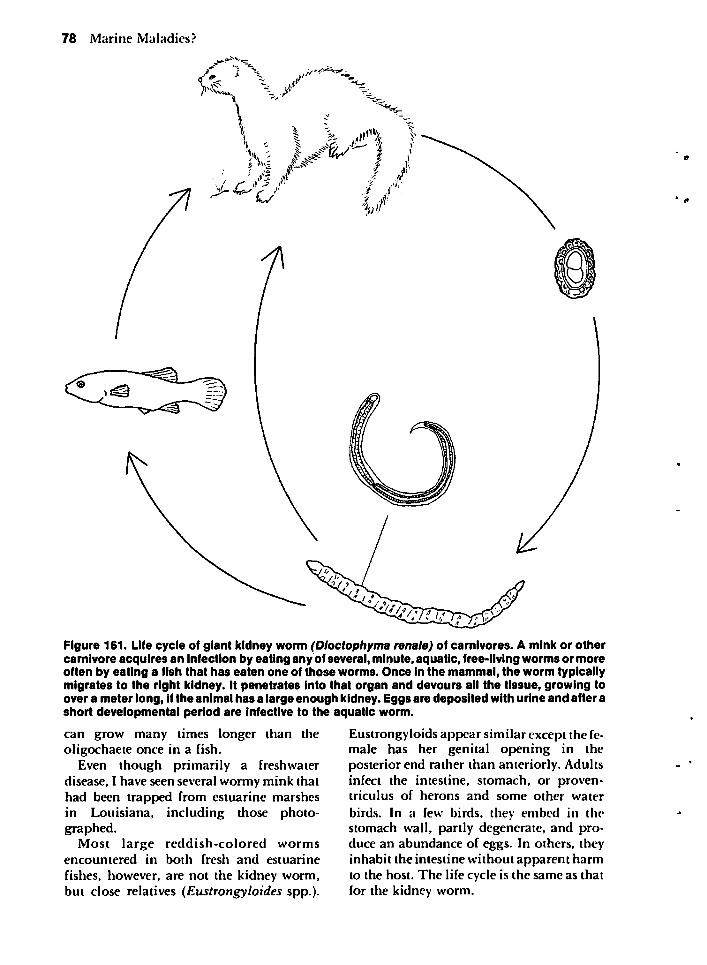
78 Marine Maladies?
—•**>=
'ii'i
,r
Figure 161. Life cycle of giant kidney worm (Dioctophyma renale) of carnivores. A mink or othercarnivore acquires an infection by eating any of several, minute, aquatic, free-living worms or moreoften by eating a fish that has eaten one of those worms. Once in the mammal, the worm typicallymigrates to the right kidney. It penetrates into that organ and devours all the tissue, growing toover a meter long, if the animal has a large enough kidney. Eggs are deposited with urine and after ashort developmental period are infective to the aquatic worm.
can grow many times longer than theoligochaete once in a fish.
Even though primarily a freshwaterdisease, I have seen several wormy mink thathad been trapped from estuarine marshesin Louisiana, including those photographed.
Most large reddish-colored wormsencountered in both fresh and estuarine
fishes, however, are not the kidney worm,but close relatives (Eustrongyhides spp.).
Eustrongyloids appear similar except the female has her genital opening in theposterior end rather than anteriorly. Adultsinfect the intestine, stomach, or proven-triculus of herons and some other water
birds. In a few birds, they embed in thestomach wall, partly degenerate, and produce an abundance of eggs. In others, theyinhabit the intestine without apparent harmto the host. The life cycle is the same as thatfor the kidney worm.

Eustrongyloids penetrate out of the fleshof dead infected paratenic hosts. Stripedbass maintained in a pond at GCRL as wellas mosquitofish, killifish, and tadpoles allhad worms escaping after a large scalemortality in the pond. Whether the wormshad any influence on the mortalities wasnot established, but investigating the relationship would be a good problem.Striped bass held in aquaria and fedgambusia acquired larval Eustrongyloidessp., whereas others not fed fish did not,suggesting the ability of the worm to bemaintained by a series of paratenic hosts.
Larval eustrongyloids have been implicated in deaths of herons and egrets. Actually,when a variety of birds feed heavily on infected fish, death can follow. Numerousred-breasted mergansers in Virginia died aday or so following engorgement on infected mosquitofish and silversides trapped beneath a thin sheet of ice on a brackish pond.Migrating worms reduced the birds' liver,proventriculus, and air sacs to a bloodypulp.
Thorny-headed WormsAcanthocephalans, or thorny-headed
worms, are all parasitic, even though livingworms presumably shed from fish do occurin benthic samples. They look superficiallylike roundworms. Actually, they have nogut, they have a protrusiblc proboscis withhooks, and their reproductive and mostother systems differ considerably from thoseof roundworms. They, like roundworms,have separate sexes.
Several different species occur in localfishes. The sheepshead minnow hosts one(Atactorhynchus verecundum) and thecobia (= lemonfish) hosts another(Serrasentis sagittifer). In order to see thefirst species, a microscope is necessary, butthe latter species stands out conspicuouslywhen one cuts into the intestine and pyloriccaeca of a cobia because it frequently growslonger (but substantially narrower) than afinger. Many other species occurcommonly,even in the fresh-brackish water interface
where one (Leptorhynchoides thecatum) infects most members of the sunfish family aswell as a variety of marine intruders. An
Fishes 79
Figure 162. Proboscises of juveniles of tworelated thorny-headed worms (acanthocephalans). Large one on left is Tegorhyn-chus turcatus that commonly infects thesouthern kingfish, and the smaller one isDollfusentls chandler! that commonlyinfects the Atlantic croaker. Even thoughboth can be transmitted by amphipods fromnear high salinity beaches, Dollfusentlschandler! usually infects estuarine hoststhat feed on amphipods from low salinityhabitats.
orange-colored species (Pomphorhynchusbulbocolli) may make a stronger impressionon an observer from this low salinity-freshwater habitat because its bulbous proboscispermanently embeds in the gut of thebowfin (= mudfish, Amia calva) and a fewother fishes.
As an extremely common estuarineexample, Dollfusentis chandleri portrays acharacteristic thorny-head (Figure 162).This species infects the rectum of theAtlantic croaker, spot, and a variety of otherfishes. It is quite similar to a larger species(Tegorhynchus furcatus, Figure 162) whichembeds in the intestine of the southern
kingfish (= ground mullet, Menticirrhus

80 Marine Maladies?
Figure 163. Typical example of a nearlymature acanthocephalan egg. The shell ofmost species looks like a diatom and thus asource of food for the arthropod intermediate host. The nearly infective acanthorcan be seen inside the shell.
americanus) and a few other fishes. In bothcases, many individuals can infect a singlehost. Indeed, over 450 specimens ofDollfusentis chandleri crowded into therectum of a relatively small (as long as ahand) Atlantic croaker without any apparent harm to the host.
Life cycles show one basic pattern,whether for species infecting fishes or othervertebrates. Developed eggs are sorted andshed by the female worm; they leave the hostwith its feces. Aquatic ones look similar toa diatom (Figure 163) and consequentlysimulate a food source for arthropodintermediate hosts. After being eaten by theproper arthropod, the larva (acanthor)forms another stage (acanthella) whichgradually develops into an infective juvenile. Finally, the arthropod with the wormis eaten by and the worm infects the propervertebrate host. In the case of acantho-
cephalans from fishes, most either use anostracod or an amphipod host, but few usecopepods, isopods, and other hosts. Sincethe cobia and many other host-fish do notfeed on such small crustaceans, the reader
may wonder how the cycle is completed.Once again, the paratenic host becomes a vital link in those cycles. Consequently, somebiologists call the host a transfer host.Fish usually fill this role and, in the case ofSerrasentis sagillifer which matures in thecobia, the Atlantic croaker, inshore lizardfish, lane snapper, and other fishes functionas the transfer hosts by harboring juveniles(cystacanths) in their mesenteries.
Dollfusentis chandleri (Figure 164) doesnot need a transfer host in its life cycle because its definitive hosts eat amphipods. Afew unrelated amphipods from habitatsranging from nearly-fresh to full-strengthseawater can host it (reported from Lepi-dactylus sp., Grandidierella bonnieroides,and Corophium lacustre). Up to threecystacanths, each folded over on itself andlonger than the amphipod, can inhabit thesame host (at least for the closely-relatedTegorhynchus furcatus in Lepidactylussp.). Tegorhynchus furcatus appears tocause an increase in the host's size; evidencesuggests infections may even make the affected amphipod a more accessible item ofprey.
Most croaker usually acquire Dollfusentischandleri in low salinity bayous about earlysummer. The worms can remain in this
principal final host for over a year, butfemales usually restrict egg-production to ashort period sometime in or near summer.Unlike many acanthocephalan species,males and females remain in equalnumbers. Many species of both acantho-cephalans and nematodes undergo a loss ofmales following fertilization, but thisacanthocephalan acts as an exception.
Crustaceans
Crustacean parasites from fishes evokemore interest than most parasites becausethey are readily visible and show such a widevariation in appearance. Isopods can beextremely large and fierce-looking, whereascopepods come in all sizes and shapes. Bothgroups have large numbers of nonparasiticrelatives. The parasitic ones have evolvedseveral features to assure their livelihood.
Some have evolved to the point where, without examining larvae and mouthparts. onecould hardly classify them as crustaceans.
Crustaceans have a protective, external,nonliving cuticle which sheds during maturation. This molting process wasdescribed earlier in conjunction with crabsymbionts. Most parasitic crustaceans alsopossess a variety of appendages the same asnonparasitic ones (see figure 48 of a brownshrimp).
Structures of the isopod (Lironeca ovalis)

Fishes 81
Figure 164. Life cycle of acanthocephalan (Dollfusentls chandleri). Male and female worms liveand mate in the rectum of the Atlantic croaker and other fishes. Specific amphipod crustaceansfeeding on deposited eggs acquire a stage that ultimately produces a juvenile worm. Croaker become Infected from eating these amphipods; however, some other acanthocephalans utilizedifferent arthropods as hosts, and some use additional transfer hosts to bridge the apparentgap between an arthropod and a predator that feeds on relatively large prey.
most commonly observed on the gills of avariety of fishes have been modified muchless than those on the bopyrid discussedearlier from under a grass shrimp's carapace (see Figures 82 and 83). Effective claws,however, do hold that asymmetricallyshaped parasite to the gills or adjacentregions of its fish-host. Oftentimes, most ofthe filaments from several gills erode awayand secondary infections ensue (see dis
cussion on bacteria and lymphocystis).When one isopod infests each side of a host,the debilitation must be considerable. Three
views (Figures 165 to 167) illustrate a sheepshead with two such unwanted "passengers."
Some other isopods display themselvesmore conspicuously when a host dies.Occasionally a large percentage of a stockof Gulf menhaden (pogy) will have a largespecies (Olencira praegustator) attached to

82 Marine Maladies?
Figures 165-167. Three views of a youngsheepshead with an isopod (LIroneca ovalls)in each gill chamber. Top, side view. Middle,top view showing bulge. Bottom, ventral(underside) view showing protrusion.Normally, the opercula do not protrude frombody.
the roof of the mouth. Actually, that is thefemale; smaller males and juveniles feed onthe gills, but can occur in the mouth. Allthese come crawling out on the deck or sorting tray of a fishing boat as the fish die.Some evidence suggests that many infestedor otherwise injured menhaden cannotfunction with their normal school and
therefore recuperate by swimming withslower younger schools or remain near shoreduring summer and fall.
The isopods situated in the oral cavitiesof pigfish and southern kingfish take moreeffort to find than simply opening themouth as in menhaden and some other
fishes.
Figure 168. An isopod (Nlroclla acuminata)attached to the dorsal fin of the sea catfish.The same isopod commonly infests the tailand also the fins of a few other fishes.
Externally-located species gain theattention of divers in clear tropical waters.An infested sergeant major in Panama or asquirrelfish in the Virgin Islands stand outobviously. Actually, a striped species(Nirocila acuminata), common on seacatfish and remoras, is just as conspicuous(Figure 168) in the northern Gulf. Apparently these may stay attached for longperiods because a small fish has a smallisopod and a large fish has a large one. Aspecies (Anilocra acuta) on the spotted gareither debilitates that host considerably orselectively infests an already weakened fish.Sick fish with extensive lesions associatedwith the isopod, especially on or nearthe bases of their fins, can be picked out ofthe water with bare hands or a dip net.
The life cycle for most of these isopods hasnot been investigated. Whereas a few speciesmay not utilize an intermediate host, mostdo, and different species of fishes apparentlyact as those hosts for several species. In theBahamas, nearly 100 tiny immature isopodscan be seen crawling about on the skin of amoderate-sized grouper. Intermediate hostsfor most species harbor fewer larvae. Presumably these larvae do not mature untilattaching to the next proper host. In theGulf, the immature form called Aegathoaoculata probably represents several differentisopod species. Usually just one or two ofthese large individuals infest a single host.
Copepods evoke curiosity because of theirrange of modifications. Indeed, two evolutionary lines of nonrelated copepods havecounterparts that superficially resembleeach other. These lines range from looking
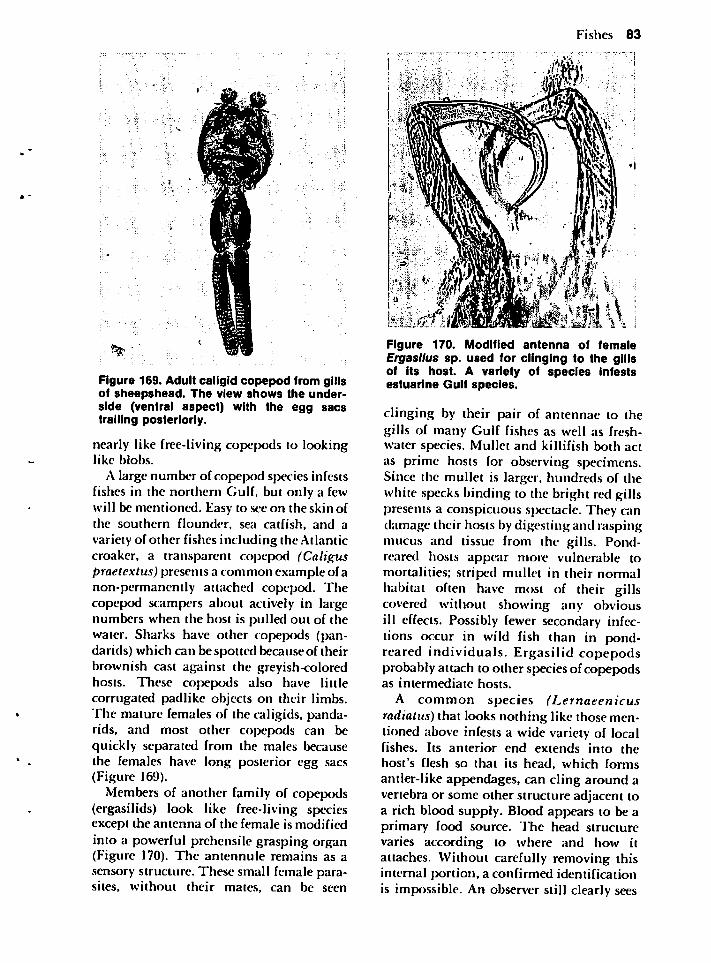
**
Figure 169. Adult caligid copepod from gillsof sheepshead. The view shows the underside (ventral aspect) with the egg sacstrailing posteriorly.
nearly like free-living copepods to lookinglike blobs.
A large number of copepod species infestsfishes in the northern Gulf, but only a fewwill be mentioned. Easy to see on the skin ofthe southern flounder, sea catfish, and avariety of other fishes including the Atlanticcroaker, a transparent copepod (Caliguspraetextus) presents a common example of anon-permanently attached copepod. Thecopepod scampers about actively in largenumbers when the host is pulled out of thewater. Sharks have other copepods (pan-darids) which can be spotted because of theirbrownish cast against the greyish-coloredhosts. These copepods also have littlecorrugated padlike objects on their limbs.The mature females of the caligids, panda-rids, and most other copepods can bequickly separated from the males becausethe females have long posterior egg sacs(Figure 169).
Members of another family of copepods(ergasilids) look like free-living speciesexcept the antenna of the female is modifiedinto a powerful prehensile grasping organ(Figure 170). The antennule remains as asensory structure. These small female parasites, without their mates, can be seen
Fishes 83
{•\.4f.t m \\ :Figure 170. Modified antenna of femaleErgasllus sp. used for clinging to the gillsof its host. A variety of species infestsestuarine Gulf species.
clinging by their pair of antennae to thegills of many Gulf fishes as well as freshwater species. Mullet and killifish both actas prime hosts for observing specimens.Since the mullet is larger, hundreds of thewhite specks binding to the bright red gillspresents a conspicuous spectacle. They randamage their hosts by digesting and raspingmucus and tissue from the gills. Pond-reared hosts appear more vulnerable tomortalities; striped mullet in their normalhabitat often have most of their gillscovered without showing any obviousill effects. Possibly fewer secondary infections occur in wild fish than in pond-reared individuals. Ergasilid copepodsprobably attach to other species of copepodsas intermediate hosts.
A common species (Lernaeenicusradiatus) that looks nothing like those mentioned above infests a wide variety of localfishes. Its anterior end extends into thehost's flesh so that its head, which formsantler-like appendages, can cling around avertebra or some other structure adjacent toa rich blood supply. Blood appears to be aprimary food source. The head structurevaries according to where and how itattaches. Without carefully removing thisinternal portion, a confirmed identificationis impossible. An observer still clearly sees
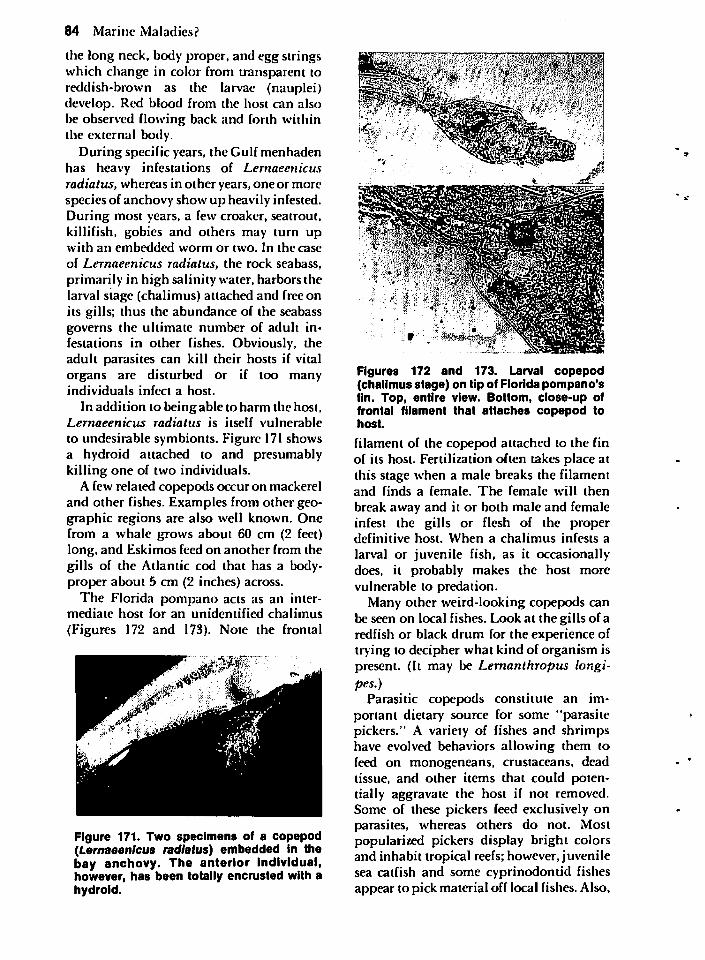
84 Marine Maladies?
the long neck, body proper, and egg stringswhich change in color from transparent toreddish-brown as the larvae (nauplei)develop. Red blood from the host can alsobe observed flowing back and forth withinthe external body.
During specific years, the Gulf menhadenhas heavy infestations of Lemaeenicusradiatus, whereas in other years, one or morespecies of anchovy show up heavily infested.During most years, a few croaker, seatrout,killifish, gobies and others may turn upwith an embedded worm or two. In the case
of Lemaeenicus radiatus, the rock seabass,primarily in high salinity water, harbors thelarval stage (chalimus) attached and free onits gills; thus the abundance of the seabassgoverns the ultimate number of adult infestations in other fishes. Obviously, theadult parasites can kill their hosts if vitalorgans are disturbed or if too manyindividuals infect a host.
In addition to being able to harm the host,Lemaeenicus radiatus is itself vulnerable
to undesirable symbionts. Figure 171 showsa hydroid attached to and presumablykilling one of two individuals.
A few related copepods occur on mackereland other fishes. Examples from other geographic regions are also well known. Onefrom a whale grows about 60 cm (2 feet)long, and Eskimos feed on another from thegills of the Atlantic cod that has a body-proper about 5 cm (2 inches) across.
The Florida pompano acts as an intermediate host for an unidentified chalimus
(Figures 172 and 173). Note the frontal
Figure 171. Two specimens of a copepod(Lemaeenicus radiatus) embedded in thebay anchovy. The anterior individual,however, has been totally encrusted with ahydroid.
T^mm^mimismssk
tfJ:-'-?$UX<*
••; ••:•••' i,:.fi!;;$p
Figures 172 and 173. Larval copepod(chalimus stage) on tip of Florida pompano'sfin. Top, entire view. Bottom, close-up offrontal filament that attaches copepod tohost.
filament of the copepod attached to the finof its host. Fertilization often takes place atthis stage when a male breaks the filamentand finds a female. The female will then
break away and it or both male and femaleinfest the gills or flesh of the properdefinitive host. When a chalimus infests a
larval or juvenile fish, as it occasionallydoes, it probably makes the host morevulnerable to predation.
Many other weird-looking copepods canbe seen on local fishes. Look at the gills of aredfish or black drum for the experience oftrying to decipher what kind of organism ispresent. (It may be Lemanthropus longi-pes.)
Parasitic copepods constitute an important dietary source for some "parasitepickers." A variety of fishes and shrimpshave evolved behaviors allowing them tofeed on monogeneans, crustaceans, deadtissue, and other items that could potentially aggravate the host if not removed.Some of these pickers feed exclusively onparasites, whereas others do not. Mostpopularized pickers display bright colorsand inhabit tropical reefs; however, juvenilesea catfish and some cyprinodontid fishesappear to pick material off local fishes. Also,

Figure 174. Argulids such as the one shownhere have acquired the common name "fishlice." They are not copepods. Note the twocompound eyes and cuplike suckersmodified from mouthparts. Eggs are laid instrings rather than carried like copepods,and argulids can damage their hosts.
some diskfishes cat parasitic copepods ofltheir primarily pelagic hosts.
Another external crustacean that appearssuperficially like a caligid copepod, butbelongs to the Branchiura group, often goesby the name argulid or "fish louse" (Figure174). In the typical argulid figured, note thecompound eyes and the modified mouth-parts forming two disks. This attractivelittle disk loses most of its intrigue when itreproduces in an aquarium. Females layeggs on a hard substratum and developmentof young ensues without intermediate hosts.Quickly, the piercing stylet, or "sting,"and proboscis of these active youngsymbionts can overpower aquarium fishes,occasionally producing an ulcerated depression where the argulid can reside.
When snorkeling among a school ofFlorida pompano, a diver might think hiseyes are playing tricks on him. Are smallbrownish specks moving about on the sidesof some of the fish? They are real! Actually,
Miscellaneous Hosts 85
the colored dots are an argulid (Argulusfuscus), occasionally numbering up to adozen per fish and involving about 10 to 30%of the pompano and Gulf kingfish amongthe breakers along the Mississippi barrierislands and northwestern Florida beaches.
Some species show more host specificitythan others. An exceptionally large species(Argulus funduli) infests the relatively smallkillifish, a smaller one (Argulus lepidostei)occurs in large numbers on the spotted gar,another small one (Argulus flavescens)commonly infests the southern flounder andseveral other fishes, and several other speciesalso infest local fishes.
MISCELLANEOUS HOSTS
The American Alligator
The establishment of an alligator seasonin Louisiana made several large parasitesmore accessible to hunters and other
interested people. If interested, one couldprobably see four ascaridoid nematodespecies in the stomach, a variety ofdigeneans from several families in theintestine, a pentastome (Sebekiaoxycephala) in the lungs and liver, and alarge leech (Placobdella multilineata) onthe skin and in the mouth. The leech acts as
an especially good indicator that theinfested gator was from or had recentlyresided in freshwater. Actually, most of thegator's parasites are acquired in freshwater,but some come from brackish water (e.g., theacanthostome digeneans). All the majorgroupings have been discussed previouslyexcept the pentastome, or tongue-worm,which belongs in a separate phylum.
Pentastomes have arthropod-1 ike nymphlarvae. Adults possess four powerfulanterior hooks (Figures 175 and 176) usedfor embedding their mouth region into hosttissues. They feed on tissue-fluids and bloodcells. The nymph of the alligator species(Sebekia oxycephala) infects a variety offishes. Even though not parasitic to man, itsrelative from rattlesnake lungs has a nymphthat can infect mice, and that nymph couldpotentially infect man.
Pentastomes typically infect the lungs of
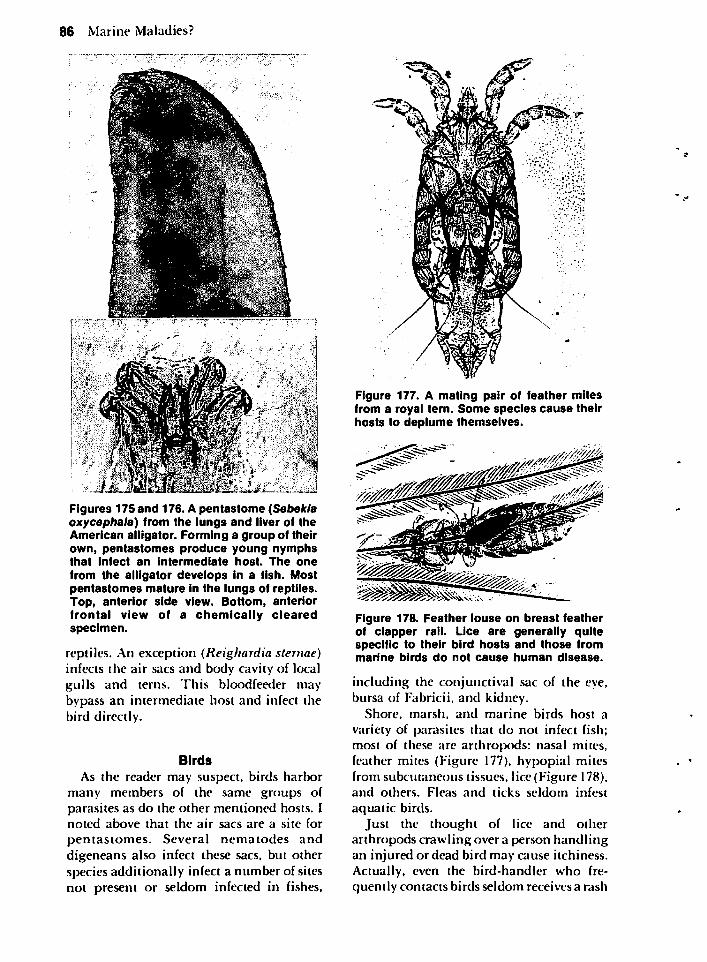
86 Marine Maladies?
Figures 175 and 176. A pentastome (Sebekiaoxycephala) from the lungs and liver of theAmerican alligator. Forming a group of theirown, pentastomes produce young nymphsthat infect an intermediate host. The one
from the alligator develops in a fish. Mostpentastomes mature in the lungs of reptiles.Top, anterior side view. Bottom, anteriorfrontal view of a chemically clearedspecimen.
reptiles. An exception (Reighardia sternae)infects the air sacs and body cavity of localgulls and terns. This bloodfeeder maybypass an intermediate host and infect thebird directly.
Birds
As the reader may suspect, birds harbormany members of the same groups ofparasites as do the other mentioned hosts. Inoted above that the air sacs are a site for
pentastomes. Several nematodes anddigeneans also infect these sacs, but otherspecies additionally infect a number of sitesnot present or seldom infected in fishes,
Figure 177. A mating pair of feather mitesfrom a royal tern. Some species cause theirhosts to deplume themselves.
Figure 178. Feather louse on breast featherof clapper rail. Lice are generally quitespecific to their bird hosts and those frommarine birds do not cause human disease.
including the conjunctival sac of the eye,bursa of Fabricii, and kidney.
Shore, marsh, and marine birds host a
variety of parasites that do not infect fish;most of these are arthropods: nasal mites,feather mites (Figure 177), hypopial mitesfrom subcutaneous tissues, lice (Figure 178),and others. Fleas and ticks seldom infest
aquatic birds.Just the thought of lice and other
arthropods crawling over a person handlingan injured or dead bird may cause itchiness.Actually, even the bird-handler who frequently contacts birds seldom receives a rash

from handling aquatic or marsh birds. A fewarthropods on non-aquatic birds do causerashes. The tropical fowl mite (Ornithonys-sus bursa) from the English sparrowoccasionally bites man, causing a rash(dermatitis). That mite, however, cannotlive longer than 10days without the sparrowor a few other domestic fowl which can host
it. Chiggers, or "red bugs," also cause irritation, but seldom infest aquatic hosts.
Searching for avian arthropods can befun. Try looking for eggs of feather liceattached to the feathers. The female of these
biting lice usually lays her eggs in rows onflight feathers safe from regions reachableby the bird's bill. After 3 or 4days of development, the nymph molts and undergoesgrowth for about a month at which time theadult emerges. Some species feed directly onfeathers whereas others puncture the quillto take blood and central pulp.
Lice reveal much about a host. If many-individuals infest a bird, it is probably soweak that it cannot keep them in check bypreening. The lice then can irritate andweaken the bird further. The number of
individuals on a healthy host seldom variesthroughout the year like it does for many-worms. Often several species of lice infest asingle bird. The color of these usually variesdirectly with the color of host; dark coloredlice infest dark colored hosts and so forth.
The reason for so many species of lice hasbeen thought to reflect evolution of thegroup spreading to a variety of unutilizedecological niches* during the period whenbirds first evolved. Apparently, ancestrallice lived in trees and later started eatingtissue-debris on reptiles. As feathers evolvedfrom the scales of reptiles, more habitatsbecame available for rapid speciation ofthese lice. Some later spread to mammals.
Lice also seem to indicate evolutionary-patterns of their hosts. Birds evolvedrapidly, so rapidly that few traces of theprimitive arrangements of bones andmuscles remain, and few fossils exist forbirds during this period when the fundamental changes occurred. The fragile natureof the bones of birds is not conducive tofossilization. Since so many lice speciesinfest birds and since they show consider-
Miscellaneous Hosts 87
able specificity toward a single species orgroup of birds, one can speculate with someassurance that both birds and their lice have
often evolved together. Since little aboutbird evolution can be deduced positivelyfrom modern evidence, that evidence
provided by parasites becomes especially-important. Evolution of lice should paralleland reflect that of birds. As an example,flamingos have been placed with storks andherons. Some species of feather lice onflamingos, however, have relatives thatoccur elsewhere only on swans, geese, andducks. Lice on storks and herons show norelationship to any of the lice on the otherindicated birds. Apparently the ancestors ofthe duck-lice were present before a commonancestor gave rise to flamingos and modernducks. Thus, maybe flamingos should beplaced more closely with ducks than withherons!
Some biting lice also seriously harm theirhosts. Juveniles of the while pelican anddouble-crested cormorants in their rookeries
may be killed by a species (Piagetiellaperalis) that heavily infests the oral cavity.Attached to the tissue of the pouch and roofof the cavity, over 500 of these 3 to 6 mmlong, transversely-striped lice can covermost of the mouth's available space.Additional immature forms occur elsewhereon the body surface. Young white ibis inrookeries also host a troublesome bitinglouse. Secondary bacterial infections(Staphylococcus sp.) often accompany thelouse infestation, and an infected youngbird looks as if it has mange. The host-parasite balance between young of nestingaquatic birds and their arthropod symbiontsneeds further investigation.
A final comment on bird parasitesconcerns white patches commonly found inthe breast of surface-feeding ducks, but alsoin birds from at least 21 other families.These cysts infect heart, leg, and neckmuscles, and they are all the same species(Sarcocystis rileyi). A definitive host for thiscoccidian protozoan has not beendetermined, but it is possibly a raptorial birdor carnivorous mammal. A related speciesin the muscles of a cow utilizes the intestineof man or dog as a final host.

88 Marine Maladies?
Marine Mammals
Marine and estuarine mammals harbor a
multitude of parasites, and the discussionfor most, just like for most birds, is beyondthe scope of this guide. Because of thepleasant and almost humanoid nature of theAtlantic bottlenose dolphin and the fact thatthis porpoise commonly makes its presenceknown in the Gulf, a few comments on someof its parasites seem warranted. The federalgovernment protects marine mammals,even when dead, reducing the chance ofseeing their internal parasites. The penetrating odors transmitted with oils in theblubber make cutting into a beachedmammal dead only a few hours a ratherunpleasant experience, anyway. If someinspired person in the northern Gulf doesget the opportunity to see a fresh stomachlining of a porpoise, he would probablysee two species of digeneans and possibly amass of nematodes. Large numbers of arather strange looking fluke (Brauninacordiformis) about the size of a marbleattach firmly to the stomach and can live forup to 3 years. The other digenean (Pholetergastrophilus) is smaller, but cannot be seenbecause a nodular cyst larger than theprevious fluke forms around the worm andhides it. The nematodes belong to one ormore species of Anisakis, the larvae of whichwhen present in raw seafood have beenimplicated in severe human gastric disease.Its life cycle was diagrammed earlier (Figure157).
A few other worms commonly infect theporpoise. One elongated fluke (Nasitremadelphini) inhabits the lateral air sinuses ofthe head near the internal ear. Less
commonly, a campulid fluke species infectsthe pancreatic ducts. The reason formentioning this infection is that worms inthis family have been implicated in somestrandings of marine mammals.
Several dolphins that had stranded singlyalong California beaches had livers badlydamaged by a campulid and had extensivelydamaged brains apparently resulting fromeasily diagnosed campulid eggs. Eggs andoccasionally adults pass through thepancreatic duct and ultimately invadevarious tissues including the brain, causingthem to degenerate. A few nematodes can
also invade the brain.
A lungworm (Halocercus lagenorhynchi)inhabits the air passages of the lungs. Thisnematode may cause severe bronchitis.Possibly because an infected porpoise hasa poorly developed cough reflex, it cannotdischarge resulting fluid. A healthyporpoise seems able to control the parasite,but a very young or stressed host apparentlyoften allows numerous individuals to
mature and thereby contribute to its deathor a serious disease. A heavy infection of arelated worm in the auditory capsule cancause deafness.
Conspicuous to someone admiring theantics of the bottlenose dolphin alongsidehis vessel are the skin disorders of some
individuals. Cuts and sores from fishingnets, shark attacks, or even self- or peer-induced trauma may progress intodebilitating infections. When a dominantporpoise bites its subordinate companion,the rake marks of its teeth orient parallel(Figure 179). Many animals also show avariety of healing wounds, often withsecondary infections (Figure 180). Othertypes of lesions occur on the skin. Some
Figures 179 and 180. Lesions on the Atlanticbottlenose dolphin. Top, flipper with deepwounds made by another dolphin. Bottom,secondary infection of wound in dorsal fin.

result from bacterial or fungal infectionsand the cause of some remains unknown.People who work with porpoises refer tosome common dark colored patchy lesionsas "tattoos." These do not appear to harmthe host, but their cause should be investi
gated.
Because marine mammals reach giganticsizes, a reader might expect a gigantic wormto infect large whales. One exists! Blue andfin whales have a nematode (Crassicaudasp.), the males of which occur in the penis orclitoris. The much larger female wormextends from this region down the ureter,through the kidneys, into blood vessels, andoccasionally to the liver. This is a longworm.
MISCELLANEOUS
SYMBIONTS
Nematode larvae and larval nematode-
like organisms infect insects and otherhosts in addition to fish and the mentionedshellfish. Because people can see theseworms, they deserve some comment.
Fairly common large nematode larvae ininsects may be mermithids. If so, farmersrefer to them as "cabbage snakes." Theinfected midge in Figure 181 from anestuarine bayou exemplifies a localinfection. Spiders, leeches, crustaceans, andother invertebrates harbor specificmermithid larvae, some of which reach50 cm in length. Because they have lifecyclesadapted to their specific hosts, produce
Figure 181. Mermithid roundworm abandoning a midge larva. Mermithid nematodeshave a parasitic larva which In some casesacts as a biological control agent to keeppopulations of annoying insects undercheck.
Miscellaneous Symbionts 89
large numbers of eggs, and are free-swimming or otherwise readily availableto infect the hosts, some act as good agentsfor biological control of pests.
In order to appreciate better thesepotential biological control agents, Ipresent a brief life history of one(Perutilimeris culicis) from the saltmarshmosquito. Free-living females lay thousands of eggs in the substratum, and thesmall preparasitic larva hatches andpenetrates an early instar* mosquito larva.It does not grow, but remains dormant untilthe mosquito pupates. At that time, thelarva migrates to the abdomen and enlargesrapidly soon after the mosquito matures andobtains a blood meal. The host becomessterile and then dies when the internalpostparasitic stage with its lance-like toothperforates an escape hole. Once in the water,the nematode molts, mates, and lays eggs.Some other species produce eggs that mustbe eaten and larvae that enlarge in the larvalinsect host.
Not to be confused with mermithids, butalso infecting insects and other arthropod
Figures 182 and 183. A horsehair worm(Gordlus robustus). Top, several malesthat had been found wiggling together In ashallow pond. Juveniles parasitize grasshoppers and a variety of other hosts. Bottom,posterior tip of a male worm. The female ofthis species has a rounded rather than alobed tail.
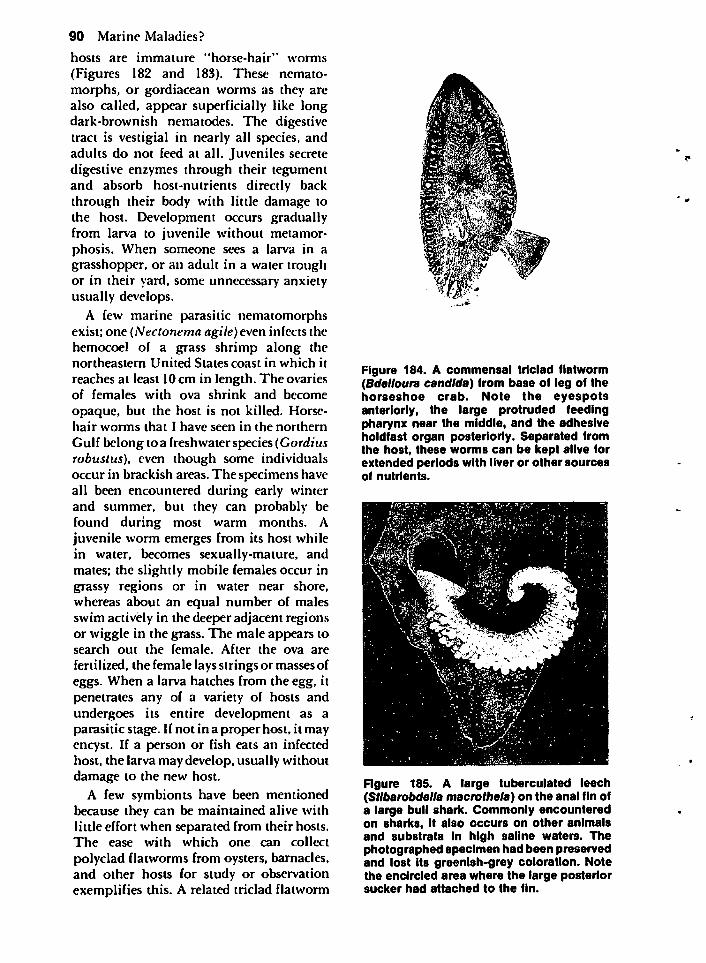
90 Marine Maladies?
hosts are immature "horse-hair" worms
(Figures 182 and 183). These nemato-morphs, or gordiacean worms as they arealso called, appear superficially like longdark-brownish nematodes. The digestivetract is vestigial in nearly all species, andadults do not feed at all. Juveniles secretedigestive enzymes through their tegumentand absorb host-nutrients directly backthrough their body with little damage tothe host. Development occurs graduallyfrom larva to juvenile without metamorphosis. When someone sees a larva in agrasshopper, or an adult in a water troughor in their yard, some unnecessary anxietyusually develops.
A few marine parasitic nematomorphsexist; one (Nectonema agile) even infects thehemocoel of a grass shrimp along thenortheastern United States coast in which it
reaches at least 10 cm in length. The ovariesof females with ova shrink and become
opaque, but the host is not killed. Horsehair worms that I have seen in the northern
Gulf belong to a freshwater species (Gordiusrobustus), even though some individualsoccur in brackish areas. The specimens haveall been encountered during early winterand summer, but they can probably befound during most warm months. Ajuvenile worm emerges from its host whilein water, becomes sexually-mature, andmates; the slightly mobile females occur ingrassy regions or in water near shore,whereas about an equal number of malesswim actively in the deeper adjacent regionsor wiggle in the grass. The male appears tosearch out the female. After the ova are
fertilized, the female lays strings or masses ofeggs. When a larva hatches from the egg, itpenetrates any of a variety of hosts andundergoes its entire development as aparasitic stage. If not in a proper host, it mayencyst. If a person or fish eats an infectedhost, the larva may develop, usually withoutdamage to the new host.
A few symbionts have been mentionedbecause they can be maintained alive withlittle effort when separated from their hosts.The ease with which one can collect
polyclad flatworms from oysters, barnacles,and other hosts for study or observationexemplifies this. A related triclad flatworm
Figure 184. A commensal triclad flatworm(Bdelloura Candida) from base of leg of thehorseshoe crab. Note the eyespotsanteriorly, the large protruded feedingpharynx near the middle, and the adhesiveholdfast organ posteriorly. Separated fromthe host, these worms can be kept alive forextended periods with liver or other sourcesof nutrients.
Figure 185. A large tuberculated leech(Stlbarobdella macrothola) on the anal fin ofa large bull shark. Commonly encounteredon sharks, it also occurs on other animalsand substrata in high saline waters. Thephotographed specimen had been preservedand lost its greenish-grey coloration. Notethe encircled area where the large posteriorsucker had attached to the fin.

(BdellouraCandida,Figure184)isalsoaninvitingorganismtostudy.Thislargewormcanbefoundonmostlargespecimensofthehorseshoecrab(notactuallyatruecrab)bylookingwherethelegsattachandatthebookgills(namedbecausetheleaf-likelamellaeonthebreathingappendagessuggestabook).
Leecheshavealreadybeenmentionedassymbiontsinfestingthebluecraband
Figure186.Medium-sizedduskysharkencircledbyacommercialplasticpackingband.Barnaclesattachtotheband.
MiscellaneousSymbionts91
shrimpsandasavectorofbloodparasites.EventhoughseveralotherleechesoccurinthenorthernGulf,one(Figure185)deservesattentionbecauseofitslargesizeanditselasmobranchhosts.Thetuberculatedleech
(Stibarobdellamacrothela)canreachalengthofabout15cm,butmostmeasureabout3cmlong.Ittypicallyinfeststhemouth,fins,claspers,andskinofanyofseveralsharks,aswellasoftheguitarfish,crabs,shells,andoccasionallybonyfishes.Itoccursworldwideinhighsalinehabitats.
Barnacleshavealreadybeendiscussedassymbiontsshowingvariousdegreesofassociationwithcrabs.Barnaclesalsoattach
andhavecommensalrelationshipstootherlivingorganisms,butoftentheyattachbecausetheorganismactsasasolidsubstratumratherthanasymbioticpartner.Threespeciesthatattachedtoaplasticbandaroundasharkdemonstratethis.TwoduskysharksoffPensacola,Florida,becameencircledwithincommercialplasticpackingbands(Figure186).Aclose-up
Figures187-190.Barnaclesattachedtoplasticpackingbandsencirclingtheduskyshark.Topleft,notethedamagecausedthesharkatitsgillslits,structuresthroughwhichwaterpassessothatthesharkcanobtainoxygen.Topright,acommongooseneckbarnacle(Lepasanatlfera)coveredbyafilamentousgreenalga.Bottomleft,therabbitearwhalebarnacle(Conchodermaaurltum)whichoftenattachestosessilebarnaclesonwhales.Bottomright,rareintheGulf,anotherrelatedbarnacle(Conchodermavlrgatum)oftenattachesdirectlytofishandtheirparasiticcopepods

92 Marine Maladies?
photograph (Figure 187) reveals the damagecaused to the body near the gill slits by theband. Similar harm caused to other fishes
will be discussed later, and I will focus myattention here on the barnacles. A common
goose-neck barnacle (Lepas analifera)frequently attaches to drifting wood andother objects. One individual on the plasticband (Figure 188) had become covered byattached filamentous green algae. Anotherstalked species (Conchoderma auritum,Figure 189) has the common name "rabbitear whale barnacle" because it has been
collected off whales. Actually, manyspecimens collected during modern timescome from hulls of ships that transittropical and subtropical Atlantic andMediterranean waters, but their commonpresence on whales seems more thanaccidental. Another species (Conchodermavirgatum, Figure 190), even though oftenattaching to ships and flotsam, does attachdirectly to small living organisms. Notcommon in the Gulf, it has been reportedthere from the dorsal fin of a halfbeak and
on a parasitic copepod (Lemaeolophussultanus) embedded in the flesh of a cobia.Considering such rare sightings of thisbarnacle, a rather unique combination ofconditions seem to be required for itsdevelopment. Perhaps one of theseconditions is a wound.
Whether one considers as parasites insectsthat must feed on blood to produce eggs isa matter of individual interpretation andinterest. Because biting gnats, even morethan mosquitoes, have such a dramaticinfluence on people's daily outdooractivities, I include a few comments onthem.
From a human point of view, gnats (atleast Culicoides furens and Culicoideshollensis), also called "no-see-ums,""punkies," and "sand flies," can vector trueparasites and often make collectingparasites or just being outside miserable.One species (Culicoides furens) acts as theintermediate host for Ozzard's filariasis
(Mansonella ozzardi) in South America andthe West Indies. Because people in NorthAmerica have not been found to harbor the
thin worm in their mesentery, the gnat inthe northern Gulf apparently has nothing
Figure 191. Underside of a biting gnat(Culicoides hollensls).Th\8 large gnat is oneof the two common gnats in the southeasternU.S. that breed in salt marshes and bite people. It is most prevalent in March-April andOctober-November and occurs throughoutthe day. The other species Is discussed in thetext.
to propagate.
The two indicated species commonly bitepeople along the Gulf Coast and Atlanticseaboard. Like mosquitoes, only the femalegnat sucks blood. Effects of the irritatingbite may persist for days in some people,whereas all symptoms disappear withina few hours for others. Along the northernGulf, adults of the larger (Culicoideshollensis, Figure 191) appear from mid-March through April and also duringOctober and November. They feed heavilythroughout the day on people, domesticmammals, and probably other animals. Onthe other hand, a smaller species (Culicoidesfurens) occurs predominantly from Maythrough September, biting near sunrise andagain from about an hour and a half beforesunset until after the sun sets. Both speciesbreed among the salt marsh vegetation.Larvae of some other species develop infreshwater habitats and those of one
(Culicoides melleus) develop in brackishintertidal sandy regions. Fortunately, thecoastal Gulf pests transmit no knownhuman diseases.
FISH-KILLS AND
MISCELLANEOUS
DISEASES
Fish-kills are not restricted to finfishes;
they also involve shellfishes. In general,shellfish mortalities present more difficulty

in assessing because the hosts are smallerand more easily overlooked, are more readilypreyed upon, and seldom swim to thesurface. Several examples of diseases thathave not been overlooked have already beendiscussed in the earlier invertebrate sections.
Fish-kills occur for a variety of reasons.Most cases have not been thoroughly-investigated, but residents usually knowwhen to take advantage of them. People fillwashtubs with fresh fish following a rapidtemperature drop or during certain otherconditions.
When water temperature gradually dropsduring the winter, fish usually have time toacclimate or move into deep or offshorewater. When the drop is rapid and when fishare additionally stressed such as by thepresence of low salinity water, some fishcome to the surface and die. Those that sink
to the bottom slowly bloat with gas and riseto the surface after about 2 to 3 weeks when
the water has warmed and algae has begunto cover them. When these fish surface,
people often worry unnecessarily aboutanother fish-kill not associated with low
temperatures. Dying fish or those in a comamake a safe product to eat if cooked and nototherwise contaminated.
Other naturally-occurring water conditions may also provide another source offood for the table. Residents in the northern
Gulf call the major condition a "jubilee."The eastern shore of Mobile Bay has gaineda reputation for its summer jubilees. Once,when the coastal region was less populated,observers would notify their neighborsabout these events; they usually occur justbefore sunrise. Now it is almost everybodyfor himself, and biologists seldom hearabout jubilees until all the fish arc collected.Rarely do animals die unless confined likecrabs in traps or sessile like oysters; mostbecome stunned for several hours and make
a dip net an easy means to gather them.Apparently a combination of specific windsand tides allows water with a low concentra
tion of oxygen to stratify and to move alongthe bottom of specific areas. Crabs andbottom fishes like flounders and eels move
shoreward and evade this encroaching watermass which soon overtakes them.
Summer kills also referred to as jubilees
Miscellaneous Kills and Diseases 93
occur along Bcllefontaine Beach andGulfport, Mississippi. Perhaps the cause ofthese differs from that for the jubilee inMobile Bay because the conditions relatedabove do not necessarily coincide with thoseencountered in Mississippi. Many fish die,but neither involved fishes nor the completestate of the water from jubilees has been investigated. Plankton* blooms have beenobserved during some of the mortalities.If algal toxins cause the mortalities, perhapsinvestigators should ascertain if affectedhosts can be safely eaten.
Devastating "red tides" result fromblooms of a few species of toxic planktonicdinoflagellates (primarily Gymnodiniumbrei'e). They occur periodically along thewest coast of Florida and less often along thewestern Gulf and perhaps elsewhere. Whena red tide occurs, large numbers of fish andinvertebrates die. an irritating aerosoloccurs throughout the nearby coastal areas,and less often, contact-rashes occur inhumans. The noxious tides alarm tourists
and residents alike.
The alga secretes a neurotoxin that killsany fish if exposed to a sufficient amount.Fish also die from oxygen depletion resulting from decomposing fish and algae.However, other mechanisms of death seem
to occur. Two of these are long term neuro-imoxication and long term blood disease.Different fishes respond differently, and,consequently, only specific fishes die from agroup of several species. As examples, lowlevels of toxin affect the ladyfish's neurological system and the mullet's blood. Themullet's blood clots slowly and the cellsbreak apart. Resulting deaths usually occura few weeks following a subsided red tide.Typically one or more fish of either speciesfrom a large school under continuous exposure to the toxin exhibits a frenziedbehavior and dies while others of the same
kind remain normal until they too areterminally affected. Some birds, primarilythe lesser scaup, also die following red tides.Perhaps this is because they feed onmolluscs which concentrate the algae, andthus the toxin. Man can also show symptoms of mild neuromuscular intoxication
following ingestion of contaminated shellfish.

94 Marine Maladies i
Probably the most common cause of fish-kills in the northern Gulf is oxygendepletion. This can occur naturally afterseveral days of overcast skies. Decreasedsunlight reduces photosynthesis* andthereby oxygen production, and this, inconjunction whith the loss of oxygen fromdecomposing organic material, results in anoxygen concentration too low to supportmost fish life. Domestic wastes and organicwastes from industrial plants intensify algalblooms and the amount of matter to be decomposed. Consequently, the number ofmass mortalities caused by oxygendepletion, especially in harbors and enclosed areas, increases with increasedpopulation growth.
Other kills result from fishing practices,toxic materials, and combinations ofnatural and man-controlled factors. Manyfish, occasionally entire catches, may bespilled from seines or "culled" from shrimpcatches. Pesticides, herbicides, and chemi
cals may be accidentally or carelesslyintroduced into a system and kill organisms.
Several other anomalous conditions
might be considered diseases. Unless fish areconfined in a rearing facility or in a stressedhabitat, most of these conditions are seldomobserved. Predators eat most affected fish before their genetic, nutritional, and injury-caused diseases become conspicuous tofishermen; however, a few affected fish canbe seen, and I will briefly mention a sample.
Anomalies seen by most shrimpersinclude albinism, ambicoloration, and reversal of flatfishes. Even though most arecongenital defects, some anomalies can becaused by trauma during early developmental stages of the fish. When flatfishdevelop normally, one eye migrates to theopposite side resulting in both eyes beingon the top or ocular side. This side possessespigmentation which can be controlled tovary in color according to the background sothat both predators and prey looking downhave difficulty detecting them. The blindunderside of the fish lacks pigment and provides a difficult target against the water'sreflecting surface for a predator from below.
When fish lack pigments in their normalregions, that is "albinism." Usually somepigments occur on the head or elsewhere
Figure 192. Reversal of a fringed flounder.The fish on the right is normal, whereas theone on the left has its eyes on the right side ofthe body.
and the lack of pigment is patchy. Replication of the eyed-side color pattern on theblind-side of flatfishes is termed "ambicoloration." The hogchoker (Trinectesmaculatus) in the northern Gulf often hasextensive shading or pigmentation on theblind-side, but ambicoloration is infrequentin these and other Gulf of Mexico flatfishes.
Reversal indicates the occurrence of eyesand pigmentation on the usual blind side. Anormal (right) and reversed (left) fringedflounder (Etropus crossotus) illustrate thiscondition (Figure 192).
By visiting a fish hatchery' where largequantities of fry are produced, one may beable to observe a variety of genetically deformed young fish. In the natural environment, many of these abnormal fish would beeaten. Still, increased heated effluents
and pollutants in the environment probablyproduce an increasing number ofabnormalities. In order to investigate thatlikelihood, eggs and embryos have beenexamined for chromosomal anomalies.
Those of the Atlantic mackerel from polluted New York Bight show a high numberof all sorts of chromosomal damage with atendency for more damage to occur in themore degraded regions.
The striped mullet illustrated in Figure193 might have been a genetic anomaly, anutritionally-deprived fish, a parasitizedindividual, or an individual confrontedwith excessive stress during early development. A variety of pollutants such assulfuric acid, herbicides, and pesticidescan produce such stress.
Figure 193. Anomalous striped mullet withsideways curvature of the spine (scoliosis).This condition, usually resulting from anutritional deficiency, can result from avariety of causes.
Figures 194 and 195. A stained section offatty liver tissue from the striped mullet. Thespecial stain (Sudan black B) stains the lipiddarkly, distinguishing the condition fromthat of excessive glycogen. Top, doublelayers of liver cells radiating normally fromcentral vein. Bottom, close-up of largestained fat globules in cells. The conditioncan be reversed in most instances; excess fatoccurs in the liver during starvation, certainvitamin deficiencies, spawning, and a fewother conditions. Young fish in aquaria fedexclusively on a diet of a single animal oftendevelop a fatty liver and die.
Fatty livers, livers with cells occupiedwith lipid material (Figures 194 and 195),commonly occur in naturally starved fishduring cold spells, in fish during theirspawning periods, and in fish maintainedon inadequate diets. Usually the process can
Miscellaneous Kills and Diseases 95
Figure 196. Fibrous tumors in a stripedmullet. This type of growth appears to beincreasing in prevalence in pollutedestuarine habitats during the past few years.
be reversed, and the liver will resume its
normal character after proper food is eaten.Fatty livers, however, can also result frompollutants.
Some contaminants also induce cancer in
fishes and other aquatic animals. So far, inthe northern Gulf, few tumors have been reported from fishes. Probably the mostprevalent one in estuarine fishes involvesfibrous outgrowths in the striped mullet(Figure 196). Usually less than \% of a catchof fish reveals these externally protrudingtumors. They, however, have been seen onlyin polluted regions of Galveston andCorpus Christi, Texas; Pensacola, CrystalRiver, and Miami, Florida; and OceanSprings, Mississippi. Long-time mulletfishermen say they have seen the tumorousfish only in recent years, further suggesting a relationship to increasing pollution.Composition of the growths apparentlydiffers among geographical regions. Thosein Mississippi fit most, but not all, of thecriteria of malignant cancers.
Most tumors on fishes are benign; that is,they do not grow fast, invade tissues, normetastasize. The most common of these, apapilloma somewhat similar to a wart,occurs occasionally on a few fish, the black

96 Marine Maladies?
Figure 197. Large black drum with benigntumorous growth on chin. This papilloma,similar to a wart, rarely occurs on fish fromthe northern Gulf. This fish from a publicaquarium developed the growth while in alarge display tank.
Figure 198. A rubber band girdling anAtlantic croaker. Tissue has grown aroundthe hole In the dorsum and separated someof the fish's tissue ventrally.
drum being an example (Figure 197). Thedrum acquired the growth of a long-timeresident displayed in a public aquarium.
Anomalies also result from injuries.Examples of wounds have already beenpresented. Chitinoclastic (chitin-attacking)bacteria essentially tattoo wounds on crustaceans. Fishes on occasion lose their tails or
other helpful but nonessential structures.Many of these fish survive, if sheltered frompredators. One local example of woundingconcerns an Atlantic croaker which
caught itself in a rubber band. After a periodof time the fish apparently grew around theband, leaving the band to perforate anddisfigure its body (Figure I98). Unfortunately, plastic beer can holders, rubbercondoms, and other items slow to degrademay cripple many fishes. A packing straparound a shark has already been illustrated
(Figures 186 and 187).A reader should also realize that plastic
bags, even though they may mess up a boat,do a lot of damage to the intestinal tract of asea turtle or other animal that normally-feeds on jellyfish and other such itemsresembling the bags. If someone is bent onthrowing garbage in the water, a can mightbe the easiest to rationalize. Most cans provide shelter for small gobies and blennieswhich in turn harbor some harmless parasites which most certainly should beperpetuated.
TECHNICAL ASPECTS
SYMBIOSIS
Textbooks on parasitology are beginningto treat many host-parasite associationsmore generally as symbiotic ones. Thevariability in the amount of dependencyof a particular symbiont on its host underdifferent conditions makes a more generalized symbiotic approach the most practicalone. Texts treating a variety of types ofsymbiotic relationships include those by-Henry (1966;l967), Read (1970), Noble andNoble (1971), Cheng (1973), and Schmidtand Roberts (1977). Few texts of any sortemphasize marine relationships. Those thatdo usually stress diseases (e.g., Sindermann,1970).
Dales (1957) presented a review of somecommensal relationships, listing numeroushydroids and anemones with their partners.Several additional uncited associations exist
in the Gulf.
The example of Clytia sp. on Donaxroemeri protracta and other species ofDonax noted above points out problemsin assigning a specific form of symbiosis,in this case phoresis, to any particularsymbiotic couple. A strict definition ofphoresis demands that neither party bephysiologically dependent on the other.The hydroid may depend on the clam. Itrarely infests any other substratum. RichardHeard has found it on old large specimens ofthe mole crab Emertia talpoida inGeorgetown, South Carolina, but why does

it not attach to other hard items along highenergy sandy beaches? Maybe it does. If so,maybe when it does, the item washes ashoreor becomes otherwise unable to support thehydroid or the hydroid's character changessuch that biologists consider it a differentspecies. Loesch (1957) reported an absenceof digenean parasites in clams with thehydroid or an attached alga. Tiffany (1968),however, did not obtain similar results. Hefound coquina over 25 mm long all haddigeneans, whether commensals were present or not. Actually, Loesch encounteredfew infected clams, and the suggested looseform of mutualism based on the hydroid'spreventing parasitic infections is doubtful.
In an unpublished study, Joyce (1961)suggested that the hydroid Podocorynecarnea (actually Podocoryne selena, seeMills, 1976) in Cedar Key, Florida, requireda living snail shell, whereas Hydractiniaechinata "preferred" a shell occupied by ahermit crab. Matthews and Wright (1970),Defenbaugh and Hopkins (1973), andMercando (1976) provided additionalobservations. AH three works reported atendency for hydroids to live on shells occupied by Pagurus longicarpus andPagurus pollicaris, but not other hermitcrabs. The first authors noted that colonies
of Hydractinia echinata were usually-moribund when on shells occupied by thecrab Clibanarius vittatus. When deprived ofa shell, 80%of the specimens of Clibanariusvittatus refused a shell covered with thathydroid, and those that accepted the shellremoved the polyps before entering it.Apparently Clibanarius vittatus is moresensitive to stinging than species ofPagurus, and those pagurids have an advantage over the more abundant Clibanariusvittatuswhen there isa fight over possessionof a shell. Laboratory experiments couldshed some light on this intriguing problem!
Anyone interested in the anemone-crabrelationship will find that several suchrelationships have been examined. Indeed,Ross (1974) gathered together most of theinformation known on the behavior, physiology, ecology, and zoogeography of thepartners and used that information toformulate ideas on the evolution of the
associations. He (e.g., 1967) has presented
Technical Aspects 97
other reviews on the behavioral aspects ofthe relationships.
The terms "zooxanthellae" and
"zoochlorellae" have no taxonomic significance when applied to invertebrate-algalrelationships. The names refer to the greenor brown colored algae just as "cyanellae"refers to a blue-green colored mutualist.Actually dinoflagellates, cryptomonads,diatoms, blue-green algae, red algae, andother algae have been reported as symbionts.Henry (1966) presented many examples.The inability to culture most algal speciesand to extract them from living hosts hasaffected the understanding of associations.Smith (1973) presented a brief review ofalgal associations, drawing mostly from theexamples of corals and their zooxanthellae,hydra and its chlorclla, molluscs and theirchloroplasts, and a small platyhelminth andits alga, a normal component of the adjacentphytoplankton. In the case of the flatworm,other related algal species can be establishedin it, but if the normal symbiont is thenexposed to it, the worm ejects the alien algaand acquires the usual one. The symbioticinteraction involves more than nutritional
processes! Taylor (1974) discussed thetaxonomy of all described symbiotic marinealgae and presented information on theirbiology. A very useful paper, it will helpanyone initiating a study on algalsymbionts.
In the two species of Zoanthus fromFlorida, I found the typical golden-brownvegetative stage of a dinoflagellateresembling those of the genusGymnodinium or Exuviaella (probablyGymnodinium microadriaticum) withinthe gastrodermis. Embedded in theepidermal tissue occurred bacteria, diatoms,and what may be another algal symbiontwhich did not fluoresce light. However, thepresence of chlorophyll a, as determinedwith a Beckman DK-2 spectrophotometer,was confirmed in tissue stripped of diatoms.Internally, a few species of colorlessflagellates resided. One large flagellateextended over 40 p. in length not includingthe flagellum, and another stored considerable starch. Even the vegetative stage of thedinoflagellate occurred in a flagellatedmotile carrier cell.

98 Marine Maladies?
A method of observing objects withchlorophyll is to use fluorescent microscopy. I found the dinoflagellate to fluorescered, and, under a squash preparation, a fewindividuals showed dividing stages.Fluorescent microscopy also revealedelongated homogeneous boat-shapedobjects that differed some between the twozoanthids. Instead of being red like that ofmost chlorophyll, the fluorescence was achartreuse green similar to that producedby members of Peridinium, dinoflagellatesthat possess armor in their motile stage.This material appeared to be packaged fromgranular material in mesogleal cells and wastransported throughout the organism. Itsfunction is unknown.
Considerable work has already beenconducted on zoanthid feeding. Trench(1974) provided an extensive study on thephotosynthetic contribution of theintracellular Gymnodinium species andhow it passes that material on to "Zoanthussociatus." Ultimately the algal cellsapparently undergo senesence, but still arenot digested by the host.
Food other than the photosynthate fromthe alga also is utilized. Digestion occursintracellularly after cells engulf smallparticles. Sebens (1977) assumed that thezoanthid studied by Trench was actuallyZoanthus solandri. The taxonomy remainsto be clarified. In any event, what Sebensconsidered Zoanthus sociatus feeds on
smaller items than Zoanthus solandri.
These include small crustaceans,
foraminifera, and apparently detritus.When not provided this external source offood, both mentioned species lost weight.Some other zoanthids tested by Sebens didnot lose weight, suggesting that they, unlikethe two species of Zoanthus, derived all theirenergetic needs from their algal symbionts.Reimer (1971) also studied different feedingbehaviors of zoanthids, but she used Pacificspecies.
Other than Trench (1974) who reportedaggregates of unidentified bacteria on thecuticle, no other authors to my knowledgehave reported more than the singledinoflagellate symbiont in a host. Theidentification, function, and interaction ofother symbionts needs investigation. Per
haps these organisms are vital to the life of azoanthid, and perhaps their presence orabundance varies according to locality orseason.
This knowledge would provide aremarkable understanding of a very-complex "simple animal." Even with thehelp of E.J.F. Wood and John F. Kimball,I was unable to culture any of the internalsymbionts using several modifications ofmethods used by McLaughlin and Zahl(1957) and Provasoli, McLaughlin, andDroop (1957). Perhaps someone using moremodern techniques and approaches cansucceed in this task.
THE AMERICAN OYSTER
The American oyster, Crassostreavirginica, acts as a host or substratum for alarge number of symbionts. The "bible" onall aspects of the oyster (Galtsoff, 1964)presents considerable information on mostof these. Cheng (1967), Sindermann andRosenfield (1967), and Sprague (1970b)provide more recent compilations onsymbionts from molluscs in general. Otherpertinent reports dealing with Crassostreavirginica include those by Sprague (1971)and Sindermann (1977). MacKenzie (1977)discussed some oyster symbionts inconjunction with an appraisal of the oysterfishery and an aquacultural program torehabilitate damaged reefs in Mississippi.Citations referring to work on specificorganisms occur in all the above articles andothers are cited below. Several infectious
and noninfectious diseases that involve
oysters from the Gulf have not been treatedhere.
Parasites of another oyster, the smallerGulf oyster (Ostrea equestris), have not beenreported as extensively. This oyster occurs inhigh salinity regions, has rows of small"teeth" bordering the inner margins of thevalves, and lacks pigmentation on theadductor muscle scar.
Overstreet and Howse (1977) provided abrief review of the public health aspects ofeating raw oysters and of coming intocontact with polluted waters in Mississippi.Unquestionably, the most dreaded disease

from eating raw oysters locally is hepatitis,but others, primarily bacterial and viral,also can be contracted. A few other types ofmicroorganisms may also have potentialpublic health significance (Harshbarger,Chang, and Otto, 1977).
Much of the ecological information aboutDermocystidium marinum came from astudy by Quick and Mackin (1971) inFlorida. Other extensive investigations havebeen made by Ray (1954), Andrews andHewatt (1957), and Mackin (1962). For thoseinterested in classification of the organism,the species has been transferred fromDermocystidium and was known asLabyrinthomyxa marina for several years.After some uncertainty, Perkins (1976a;1976b), using ultrastructural evidence,considered it a coccidian or relatedprotozoan, and Levine (1978) has nowestablished it as a protozoan namedPerkinsus marinus after Perkins.
Several other species exist in oysters andother molluscs; Quick (Dow ChemicalCompany, Frecport, Texas, personalcommunication) estimates at least four orfive Labyrinthomyxa-like organisms affectthe oyster alone. Most of these usually occurin low numbers and do not harm the oysterunless it experiences other stresses. Uponculturing with thioglycollate and othermedia, these organisms respond in a varietyof different ways and morphological types.Often more than one species will infect thesame individual oyster. Much more effortshould be given to understanding theseorganisms and their relationships with theirinvertebrate hosts and their environments.In fact, Labyrinthomyxa patuxent, firstdescribed in 1921, a potentially-lethalpathogen, and the focus of considerableresearch (Mackin and Schlicht, 1976), maybe related to what many authors considerneoplastic (tumor) diseases of molluscs.Whether that organism, with its incomplete response to thioglycollate medium,occurs in the Gulf has not been firmly-established. The method for the improvedculture medium has been documented anddiscussed by Quick (1972).
A disease not discussed above because itapparently does not affect oysters in the Gulfis "MSX" (for Multinucleate Sphere X).
Technical Aspects 99
Losses as high as 85%of planted oysters fromsome regions along the northeastern U.S.coast have been attributed to this disease,now known to be caused by the haplosporidan Minchinia nelsoni based onevidence by Couch, Farley, and Rosenfield(1966). Both it and Minchinia costalis causedisease and have attracted considerableattention concerning their morphology(Perkins, 1968; 1969), reduced reproductivecapacity of infected oysters (Couch andRosenfield, 1968), and mortalities caused bythe two haplosporidans (Andrews, Wood,and Hoese, 1962; Farley, 1975; Kern,1976). Even though most oysters in a reefmay be killed, survivors and their progeny-appear to develop a resistance. Consequently, either transplanting uninfectedoysters into an enzootic region orintroducing an infected oyster into a regionfree of the haplosporidan disease can haveshort-term devastating effects!
Apparently more than two species ofsporozoan gregarines infect the oyster.Spores of Nematopsis ostrearum lodgeprimarily in the mantle, and those ofNematopsis prytherchi and perhaps anotherundescribed species primarily involve thegills. Quick (1971) encountered unidentifiable spores near the digestive tissues ofoysters from central but not northernFlorida. Decapod hosts for Nematopsisostrearum include several mud crabs,whereas the host for spores in the gills is thestone crab. Mackin (1962: 203-205)questioned whether any of these speciescauses oyster-mortalities as suggested byseveral authors. Nematopsis spores,generally larger than those ofDermocystidium marinum, can be seen inhistological sections or in fresh tissuepartially digested for about a minute in afew drops of 10% sodium or potassiumhydroxide solution.
Nematopsis ostrearum appears to beinvolved in a dynamic equilibrium inoysters from Virginia (Feng, 1958). That is,when oysters heavily or lightly infected gettransplanted to other regions with oystershaving light and heavy infections,respectively, the transplants alter theintensity to assume average levels for theregion. Such ideas should be investigated

100 Marine Maladies?
further using oysters in the Gulf.At least two species of Stylochus plus
other polyclads occur in oyster beds ofMississippi (Hyman, 1940). Stylochusellipticus, a cream to brownish wormreaching about 25 mm long, is much moreprevalent than the grayish 50 mm longStylochus frontalis. The latter, much morecommon in Florida, was studied by Pearseand Wharton (1938), who presentedecological and biological data on it as thesynonym Stylochus inimicus as well as somedata on Stylochus ellipticus under the nameEustylochus meridianalis. The latterwithstands water of lower salinity andtemperature than Stylochus frontalis andis less specific toward oysters. Both specieslay single layers of egg-masses on oystershells or other hard surfaces.
The oyster crab rarely occurs in most Gulfregions, but because students often wonderabout problems they could undertake, Ithink it worthwhile to mention here that
several species of small pinnotherid andporcellanid crabs associate with a variety ofinvertebrates and provide an easily visibleanimal to study. If a person collectspcnshells or scallops, digs up burrows ofmud shrimps (callianassids). or investigatesthe tubes or burrows of polychaetes,holothurians, or sipunculids, he willprobably encounter several species of \kacrabs. They can probably be identified usingthe keys by Williams (1965) or Felder (1973).
Many questions concerning each differentpea crab could be asked. Are youngproduced over long periods? Does the firststage crab always invade the host? Are youngindividuals chemically attracted to hosts?What is the dependency and pathologicaleffect of the crab on the host? What is thelongevity of both sexes? Will additionalfemales enter an already occupied habitat?When several females associate with a host,
will some migrate to nonassociated hostswhen they change into a hard stage? Can thecrabs survive harsher conditions than theirhosts? Do attached hydroids aid the crabs? Aplace to initiate a literature search for anysuch investigation could be articles byChristensen and McDermott (1958), Haven(1959), Davenport, Camougis, and Hickok(1960), and Sastry and Menzel (1962).
The small pyramidellid gastropod alongthe edge of an oyster shell can be identifiedusing Abbott's (1974) book. Hopkins (1956c)and Allen (1958) both discussed theOdostomia impressa-oyster relationship.
Oyster growth shows a relationship withbucephalid infections (Menzel andHopkins, 1955), although more extensivestudies are needed.
The pathogenic effect by normallycommensal inhabitants to stressed oystershas not been well documented in most cases.
Stress from high temperatures apparentlycauses dermo to harm the host. Other
examples also exist. Ciliates occasionallyinfect a healthy oyster, but may become acomplicating factor in weakened hosts as asecondary invader (Mackin, 1962; Pauley,Chew, and Sparks, 1967).The latter workersobserved a ciliate to infect heavily the Pacificoyster after they injected turpentine, but nota nonirritant, in the adductor muscle or inconnective tissue.
Even though burrowing organisms affectoysters and other molluscs, few studies havetreated them in detail. Alfred Chestnut at
GCRL provided most of the information onthe oyster burrowing clam, or martcsia(Diplothyra smithii), used here. Hisdissertation will present details of thebiology of this intriguing clam. The mudworm Polydora websteri can be easily-identified using Blake's (1971) key. Anotherspecies, Polydora ligni, also excurs in oysterreefs, but forms fragile tubes of silt ratherthan burrowing in shells. It. however, canreproduce rapidly and smother the oysters.Hartman (1958) presented a taxonomictreatment on the several species of burrowing sponges, and Wells, Wells, andGray (1960) added sizesof perforations in theshell in addition to other characteristics.Hopkins (1956a; 1956b) reported ecologicaldata on the different species. He consideredthe presenceof Cliona celata to indicate thatsalinity remained below 10 ppt about onefourth of the time and that of Cliona truitti,a species favoring high salinity, to indicatethat the salinity seldom dropped below15 ppt. For those interested in animal-burrowing, an entire issue of AmericanZoologist (Carriker, Smith, and Wilce, 1969)provided many specific and general articles

treating the subject.The section on predation by Galtsoff
(1964) can be amplified by reading theinteresting findings of McDermott (1960)who studied predation of oysters by mudcrabs. Kay McGraw has been investigatingpredation by the blue crab in her doctoralwork on growth and survival of hatchery-reared oysters. McGraw and Gunier (1972)investigated predation by the drill inMississippi. Larval development of thismost important predator, Thaishaemastoma, was described by D'Asaro(1966).
THE BLUE CRAB
The blue crab fishery in Mississippi hasbeen discussed by Perry (1975). Others havetreated it in adjacent regions in the Gulf(e.g., Darnell, 1959; More. 1969; Jaworski,1972; and Adkins. 1972). Tagatz and Hall(1971) presented an annotated bibliographyon Callinectes sapidus, Pyle and Cronin(1950) described its anatomy, PhyllisJohnson is finishing a histological study,and Overstreet and Cook (1972) discussedsoft-shelled crabs and their potential forfurther exploitation.
So far I have not said anything about thelesser blue crab, Callinectes similis, or. as
some fishermen call it, the Gulf or scissorbill crab. It also occurs in the northern Gulf
and the eastern U.S. seaboard (Williams.1974) and helps make up a part of the bluecrab fishery. It, however, does not reach thesize of Callinectes sapidus and can be furtherseparated from it by possessing two largeand two small "teeth" projecting along thefront of the carapace between the eyes ratherthan just two large ones. This count doesnot include the projections where theeyestalks protrude. Also, small specimens ofCallinectes similis have a blue patch at the"elbow" and where the claw articulates,rather than being red like on Callinectessapidus.
The parasites infecting the two crabspecies also differ in some cases. A criticalstudy of all the parasites of the two has notbeen made, even though numerous paperscover individual species infectingCallinectes sapidus. As far as I know, neither
Technical Aspects 101
Ameson michaelis nor Loxothylacustexanus, both internal parasites, infectCallinectes similis. I tried to infect a few
individuals with the microsporidan withoutsuccess. On the other hand, I have seen a
nonencysted larval cestode plerocercoidextensively invading the skeletal muscles ofthe lesser blue crab (Figures 26 and 27). buthave never seen it in the regular blue crab.On the basis of examining muscle tissuefrom numerous specimens of Callinectessapidus. both Phyllis Johnson and RichardHeard (personal communication) have eachseen but a single specimen of a larval cestode. Heard's specimen was encountered inthe Florida Keys. Most external symbiontsinfest both crabs, and their presence dependson ecological variables. The remainingdiscussion relates entirely to infections inCallinectes sapidus.
The first of four viruses infecting easternblue crabs, according to Phyllis Johnson(1977b), is a reolike virus found inhemopoietic tissues, hemocytes, glia of thecentral nervous system, and other cells. Ifinfected hemolymph is injected into thebody- cavity of a crab, the crab may diewithin 3 days, probably from neurologicalinvolvement, compared to 12 to 32 days forcrabs fed injected tissue. The second is aherpeslike virus mainly in hemocytes(Figures 44 and 45) which takes about amonth to kill a confined crab and perhapsmuch longer to kill or severely affect anunstressed host in its natural environment.
Crabs do not appear distressed until neardeath, at which time their hemolymph turnschalky white. The third is a picornalikevirus infecting primarily neurosecretorycells, epidermis, and bladder epitheliumand secondarily invading blood cells andother tissues. After about a month, the crab,affected by an altered molting pattern andsometimes by blindness, may die. Thefourth is a baculovirus infecting the hepa-topancrcas that does not appear to causeovert disease. Most of these and other viruses
probably affect the blue crab in the Gulf.Infections to date have been confirmed
solely on the basis of ultrastructuralexamination of specimens from the Atlanticseaboard (P. Johnson, 1977b).
A variety of chitinoclastic bacteria have

102 Marine Maladies?
been isolated from crabs and shrimp inMississippi (Cook and Lofton, 1973). Onestrain, Beneckea type I. was isolated from allnecrotic lesions, making it suspect as aprimary causative agent that invadeswounded hosts. Shell disease in the blue
crab, caused by one or more opportunisticorganisms, has only been reported relativelyrecently (Rosen, 1967; Sandifer andEldridge, 1974), but has long been considered a problem to crabbers who maintaincrabs in floating cypress cages or who leavecrabs in traps for extended periods.Overstreet and Howse (1977) discussed theneed to investigate the relationship betweenstress and causative agents.
Crabs collected by (rapping, trawling, ordredging typically have systemic bacterialinfections which are undetectable other
than by culturing the bacteria. Many suchcrabs with bacteria in their hemolymphappear to be, and probably are. relativelyhealthy (Colwell. Wicks, and Tubiash, 1975;Sizemore, Colwell, Tubiash, and Lovelace,1975). Most of these bacteremias areprobably transient, and some may bemisdiagnoses based on cultures of contaminants from the cuticle. Disease typicallyensues only when the organism is especiallyvirulent or when the host is stressed. Heavyinfections can kill crabs (e.g., P. Johnson,1976). Overstreet and Howse (1977) reportmortalities of crabs in traps caused byVibrio parahaemolyticus.
Vibrio parahaemolyticus, a gramnegative, rod-shaped, motile bacterium hasalso been implicated in human disease.Many strains live and multiply rapidly inmarine waters, especially at the human body-temperature of 37°C. In contrast, moststrictly human pathogens cannot reproducein seawater. These organisms concentrate inmarine organisms near sewage outfalls andgive most raw seafood products anundeserved poor reputation. Just likeseveral other bacteria though, Vibrioparahaemolyticus usually producessymptoms of nausea, diarrhea, vomiting,and mild fever. It has also been implicated ininfections of the skin. Lee (1973) discussedinformation that seafood processors shouldknow about the organism.
Most of the microsporidans infecting the
blue crab as well as other hosts have been
treated by Sprague (Bulla and Cheng, 1977).I am presently working on another species.Sprague recently changed the name of themost prevalent species, Ameson michaelis,when he transferred it from the genusNosema. Some aspects of its developmenthave been described by Weidner (1970; 1972).Overstreet and Weidner (1974) suggestedthat the muscle disintegration induced byAmeson michaelis may be more severe andextensive than that caused by several othermicrosporidans because the spores of thisspecies lack a surrounding pansporoblast.A growth chamber bounded by one or moremembranes, the pansporoblast may trappotentially harmful metabolites.
If in the future the crab industry rears itsproduct for the soft-shelled crab market,means to prevent and control infections ofmicrosporidans will be necessary. Steps inthese directions have already been made(Overstreet. 1975; Overstreet and Whatley,1975; Sindermann, 1977).
A helpful listing of many protozoansfrom decapods was presented by Spragueand Couch (1971). Many other papers treatdifferent aspects of infections and theircausative agents in detail, and a few of thesewill be mentioned. P. Johnson (1977a)critically investigated paramoebiasis; theagent has been adequately described bySprague, Beckett, and Sawyer (1969) and by-Perkins and Castagna (1971). Newman andC. Johnson (1975) reported the dinoflagellate Hematodinium sp., and Couch(personal communication) has additionalstudies in progress. Newman, Johnson, andPauley (1976) described the Minchinia-likehaplosporidan. Three papers (Couch, 1966;1967; 1973) treated the ciliate Lagenophryscallinectes, and Overstreet and Whatley(1975) reported the internal "holotrich."
Bland and Amerson (1973a; 1973b)reported the fungus Lagenidiurn callinectesfrom crustacean ova, and continuing workby Bland's group has developed controlmeasures (Bland, Ruch, Salser, andLightner, 1976).
In a dissertation on microphaiiiddigeneans of fiddler crabs, Richard Heard(1976) additionally treated some speciesfrom the blue crab and from other

crustaceans. He corrected and added data to
the earlier reports on Microphallusbasodactylophallus (= Carneophallusbasodactylophallus) by Heard (1967) andBridgman (1969). Even considering thenomenclatural treatment of the species in amonograph by Deblock (1971), problemsstill exist, as pointed out by Heard. Heardalso elucidated additional aspects of the lifecycle of Levinseniella capitanea, a wormdescribed by Overstreet and Perry (1972).The cycle progresses much the same asMicrophallus basodactylophallus; however,fewer mammals and birds probably host theworm than for many microphallids, since Icould not establish specimens in mice, rats,or baby chicks.
Many microphaiiid metacercariae,including those discussed, produce eggswhen maintained in warm saline. This
ability reflects the lack of much biochemicaldependency on the host for egg productionin those microphallids. The importantaspect of the life cycles of those species seemsto be the need for adults to produce eggsquickly, a maneuver allowing infection ofintermediate hosts if the definitive host is
one that would soon migrate from theregion and thereby leave potential intermediate hosts behind.
Various histological studies treating thehyperparasite Urosporidium crescens fromthe Atlantic coast list digeneans of the bluecrab other than Microphallus basodactylophallus as probable hosts (De lurk,1940; Sprague, 1970a; Perkins, 1971; Couch,1974a); however, Heard and I haveexamined specimens of trematodes frominfected crabs from Virginia and foundthem to be Microphallus basodactylophallus, the same as in the Gulf. On theother hand, additional digeneans aresuitable hosts for this haplosporidanincluding Microphallus choanophallus andLevinseniella capitanea.
The acorn barnacle, Chelonibia patula,and the pedunculate barnacle, Octolasmismuelleri, have been reared in the laboratoryby Lang (1976a; 1976b). Those workssuggest that distribution of both speciesdepends on temperature. Breeding andlarval development ceases below aboutI5°C. Development from the first naupliar
Technical Aspects 103
to cyprid stage of Octolasmis muelleri takesabout 2 to 3 weeks at 21 to 27°C and less for
the other barnacle. Development to thatcyprid stage, the stage which attaches to thehost, prolongs considerably in cooler water.Diet also controls development, and thepedunculate barnacle necessitates a muchmore specific diet. Observations on thatspecies from North Carolina weredocumented long ago (Coker, 1902).However, whether or not specimens fromthe blue crab in America comprise ageographic population and a synonym ofthe cosmopolitan Octolasmis lowei has notbeen established decisively (e.g., Newman,1967).
Ecological studies of Loxothylacustexanus in the northern Gulf were presentedin some detail by Ragan and Matherne(1974) who additionally cited most of thereferences on that species in their bibliography. The only other reported host inaddition to Callinectes sapidus isCallinectes ornatus. Gordon Gunter has
seen that crab infected along the west coastof Florida (Christmas, 1969), and I have seenmany in the shallow hot regions of Bear Cutin Miami, Florida. Lawler and Shepard(1978) present a fairly complete bibliography on all rhizocephalans. FromMississippi, Christmas (1969) reportedinfected blue crabs and also mentioned the
presence of what he tentatively consideredLoxothylacus panopaei in mud crabs.
Loxothylacus panopaei is an importantexample of introduced species. After manyoysters in Chesapeake Bay died fromMinchinia nelsoni, oysters from the Gulf ofMexico were introduced in 1963 and 1964 tohelp reestablish the population. Incidentalto the oysters occurred mud crabs infectedwith the rhizocephalan. By 1965,native mudcrab populations started revealinginfections of Loxothylacus panopaei for thefirst time (see Van Engel. Dillon, Zwerner.and Eldrtdge, 1966; Daugherty, 1969).Couch (personal communication) has alsofound Eurypanopeus depressus infected inChincoteague Bay (Figure 36).
Soon after first being introduced, asuccessful species often produces excessivelylarge populations until such time that anequilibrium becomes established. Whether

104 Marine Maladies?
infections of Loxothylacus panopaei spreadexplosively has not been established, but theapparent abundance of mud crabs declined.Possibly the parasite did not influence thepopulation structure of the crabs, but,nevertheless, introductions of one
species may result ultimately in a decline inother species in the new habitat. Beforeintroducing any animal or plant, the needfor it and the possible side-effects from theintroduction should be carefully evaluated.
Humes (1942) presented an extensivetreatment on the nemertean Carcino-
nemertes carcinophila. which was laterfollowed byapapcron itsuscasan indicatorof host spawning (Hopkins, 1947) andanother on its larva (Davis, 1965).
The fact that the common leech
Myzobdella lugubris uses a fish to obtainblood and a crab on which to depositcocoons has been recognized only recently(Daniels and Sawyer, 1975; Sawyer, Lawler,and Overstreet, 1975). The latter referencegoes into considerable detail on salinity-tolerances of the adult and developing leech.Hutton and Sogandares-Bernal (1959)reported the association between the leechand crab mortality. Some data on thebranchiobdellid Cambrincola mesochoreus
were reported in an unpublished honor'sthesis by Blackford (1966), but moreextensive work could be done on that and
other branchiobdellids of both the crab
and crayfishes in the northern Gulf region.The authority on that group is Perry C. Holtat Virginia Polytechnic Institute and StateUniversity.
PENAEID SHRIMPS
Considerable data have been publishedon shrimp catches and shrimp biology.Perez Farfante (1969) provided a detailedtaxonomic and ecological treatment and areview of all members of the genus Penaeusin the Western Atlantic. Of the two common
species in the northern Gulf, the brownshrimp has a groove (lateral rostral sulcus)running along each side of the dorsal (top)midline ridge on the "head" (cephalo-thorax) as well as one on the last (sixth)
abdominal (tail) segment. The whiteshrimp lacks these, but not the pink shrimp,which may additionally have a distinguishing brownish lateral spot on thesecond and third segments. Cook andLindner (1970) and Lindner and Cook(1970) presented good summaries of data onthe brown and white shrimps to whichcould be added a paper on burrowing anddaily activity of shrimp by Wickham andMinkler(l975). Christmas and Etzold(1977)edited a report on a regional fisherymanagement plan for penaeid shrimp.
The parasites and diseases of commercialpenaeids, more than those of most any otherselect group of marine organisms, have beenstudied in some detail. A considerable
number of questions remain unanswered,but recent findings and summaries ofprevious works have been presented byOverstreet (1973), Feigenbaum (1975), S.Johnson (1971; 1975), Lightner (1975). andCouch (1978). Couch broadened his paperfurther by including the response of shrimpand their tissues to a variety of toxicchemicals and heavy metals. The observations on the tumorous growth, ahamartoma, discussed above were made byOverstreet and Van Devender (1978).
Couch (1974b) first observed the viralinclusion bodies in shrimp and followed hisreport with a series of studies assessing therelationship between stress and disease(Couch, 1974c; see citations by Couch andCourtney, 1977). The virus was characterized by Summers (1977). Flagellate andciliate protozoans additionally involvedsome of the dying larval shrimp, but nodoubt exists about the increased presenceof the inclusion bodies in shrimp experiencing certain stresses.
The normal bacterial flora of brown and
white shrimps was investigated byVanderzant, Mroz, and Nickelson (1970).Lightner (1975), who reported the epizooticof virulent chitinoclastic bacteria, reviewedbacterial diseases of shrimp in general.
Research on fungal diseases of crustaceans has just begun to receive seriousinvestigation. Unestam (1973), Bland(1975), and Lightner (1975) provided goodsummaries of the problems.

"Black gill disease" results from a varietyof both infectious and noninfectious agents.Massive accumulations of hemocytes settlewithin the gill filaments, ultimatelyassociated with dysfunction, destruction, orsecondary infections. Lightner and Redman(1977) showed histologically that theinflammatory responses typically involvemelanin deposition following exposure ofshrimps to cadmium, copper, and Fusariumsolani, as well as when shrimp have anascorbic acid deficiency, wounds, and avariety of undiagnosed conditions.Inflammatory response in penaeid shrimpinvolves fixed phagocytes lining bloodsinuses, phagocytes in loose connectivetissue, fibrocytes, and three forms ofhemocytes. Solangi and Lightner (1976)followed the response to the fungus,Fontaine and Lightner (1975) described theencapsulation, phagocytosis, and elimination of necrotic material as well as the
healing process, Couch (1977) presented anultrastructural study of lesions in gillsexposed to cadmium, and Sindermann(1971) reviewed the internal defenses ofcrustaceans in general.
The four common microsporidans differin several aspects (Overstreet, 1973) and willbe described because of the difficulty indistinquishing them and because of theirreceiving a recent resurgence of interest.Spores of Ameson (= Nosema) nelsonimeasure about 2.5 ;/ long by 1.5 fi wide(occasionally up to 3.5 ji by 2.3 ]i) with anarrow polar filament 23 /< long. The singlespore has no pansporoblast chamber, andmasses of them surround the abdominalmuscle bundles of white, brown, and pinkshrimps before eventually replacing them.Pleistophora sp. may be especially difficultto distinguish from the above species.Chromatophores along the dorsolateralsurfacesof shrimp infected with both speciesexpand and produce a bluish-blackappearance to the shrimp; however, theappearance usually is darker when theshrimp is infected by Pleistophora sp.Sporesof that species, 1.7 to3.0/< long by0.9to2.5 n wide withpolar filaments 42 to 125 ylong, develop in pansporoblasts containingas few as 14 spores or as many as severalhundred. The Pleistophora sp. infections
Technical Aspects 105
typically replace muscle tissue and sometimes invade anteriorly-located tissues inaddition to the tail; they involve primarilywhite and brown shrimps. The third speciesfrom the host tail, Thelohania duorara, lies
between muscle fibers, most commonly inpink shrimps. As with Pleistophora sp., itoccasionally invades heart and other tissues.Spores measure 4.7 to 6.8 }i long by 3.0 to4.2 u wide (averaging 6.0 by 3.7 /<) withuniformly wide polar filaments 97 to 142 //long; eight of them exist within eachpansporoblast.
The last species (Agmasoma /= Thelohania] penaei) invades tissue other thanthe abdominal muscle. Prevalent inblood vessels, foregut, and hindgut, it alsoinfects and destroys the germinal tissue ofthe gonads. Infections seen through thecarapace among the branchiostegalepithelium and in the appendages originatefrom tissue of the blood vessels. Only in anabnormal situation have I observed an
infection associated with abdominal
muscles (Overstreet, 1973). A pansporoblastharbors a group of eight pyriform sporeseither 2.5 to 4.7 /c long by 2.0 to 3.5 fi wideor 5.5 to 8.2 ]i by 3.5 to 4.2;^. Both the smalland large (megasporcs) spores occur in thesame shrimp. Also, the polar filamentsprotrude 65 to 87 }i and have a thin distalportion contrasting with the thick proximalpart. Other than some pond-reared pink andbrown shrimp, only white shrimp haverevealed infections. Iversen and Kelly (1976)apparently transmitted this or a relatedspecies to postlarval pink shrimp by feedingthem feces from spotted seatrout that hadeaten infected shrimp.
Descriptions of the different NorthAmerican gregarines of penaeids occur inworks by Sprague (1954), Kruse (1966a;1966b), Overstreet (1973), and Feigenbaum(1975). Kruse (1966a; 1966b) providedinformation on the biology of a few species,and Sprague (1970a) reviewed gregarines ofcrustaceans in general.
Descriptive data on most ciliates arelacking. Couch (1978) gave a recentsummary of most ciliate and flagellatespecies from shrimp, and papers byS. Johnson (1974) and by Lightner (1975)provided additional information. The

106 Marine Maladies?
relationship between Zoothamnium sp. andthe shrimp's gill has been studied by Foster,Sarphie, and Hawkins (in press), and thepossible detrimental effects caused by thatrelationship were discussed by Overstreet(1973).
Heard experimentally infected the shrimpshown in Figure 68 and plans to report theresults soon. Overstreet (1973) summarizedinformation on the other trematodes,including a description of Opecoeloidesfimbriatus by Kruse (1959).
Most of the common helminths have been
reported by Hutton, Sogandares-Bernal,Eldred, Ingle, and Woodburn (1959), Kruse(1959), Overstreet (1973), Feigenbaum(1975), and Couch (1978). Feigenbaum(1975) discovered two new trypanorhynchcestodes from the hepatopancreas and bodycavity of Penaeus brasiliensis near Miami,Florida, and Corkern (1978) found anencysted cyclophyllidean larva in shrimpsin Texas. No one has seriously tried todetermine the identity of the intestinallarval cestode. However, Tom Mattis hascompleted much of a dissertation problemat the Gulf Coast Research Laboratory onthe life history of Prochristianella hispida.Figure 70 illustrates his present understanding of its cycle. Prevalence of thatspecies, previously referred to asProchristianella penaei (see Campbell andCarvajal, 1975), has been treated by severalauthors (Aldrich, 1965; Corkern, 1970;Ragan and Aldrich, 1972; Overstreet, 1973),and Corkern (1970) also included data onParachristianella dimegacantha, a speciescommon in younger shrimp from GalvestonBay, Texas.
In white shrimp studied by Sparks andFontaine (1973), larval Prochristianellahispida developed a dense cyst in thehepatopancreas that was apparently soondestroyed and resorbed by the host. When inthe hemocoel, the larva had a thinner cystwith less inflammatory response,suggesting a more preferable site. Thoseauthors believed that worms are graduallylost in white shrimp. Because infections inbrown shrimp do not reveal a decrease inintensity with age as assumed for those inwhite shrimp from some areas, the authors
suggested brown shrimp may respond toinfections differently. When shrimp can bereadily infected, these hypotheses can betested. Overstreet (1973) did not detect adecreased intensity of infections over the 2 to3 month period shrimp were reared inLouisiana ponds.
Most investigators previous to or unawareof a study by Mudry and Dailey (1971)assumed shrimp acquired infections fromsmall infected crustacean intermediate
hosts. That study suggested that in cyclesusing a crustacean and elasmobranch, thecrustacean acquired its infection directly byfeeding on worm's eggs, whereas in cycleswith a plerocercoid in a fish, the fishacquired the infection from a crustacean.The first method may not be representativefor penaeid shrimp with Prochristianellahispida.
The larval ascaridoid from shrimpdescribed by Kruse (1959), Hutton,Sogandares-Bernal, Eldred. Ingle, andWoodburn (1959), and others asContracaecum sp. can presently beconsidered Thynnascaris type MA. Norrisand Overstreet (1976) briefly discussed itanddifferentiated it from type MB. Type MB is 1to 3 mm long with an esophagus 4 to 13%ofthat length, a ventricular appendage 14 to 68times the length of the short intestinalcaecum, and a spineless, bluntly-roundedtail. In contrast, type MA is 3 to 5 mm long,has an esophagus 14 to 15% of that length,has a ventricular appendage 2 to 3 times aslong as the intestinal caecum, and possessesa spiny projection on the tail. Norris andOverstreet recognized three or four formsfrom penaeids.
Adult Thynnascaris species infect avariety of fishes. Norris and Overstreet(1975) described two adult forms andThomas Deardorff of GCRL hopes to beable to correlate the adults of a few Gulf
species with their larval forms by experimentally completing portions of their lifecycles.
Spirocamallanus cricotus was firsttentatively reported from penaeid shrimp asSpirocamallanus pereirai (see Overstreet,1973). Fusco and Overstreet (1978), however,have now clearly established that the species

commonly referred to as Spirocamallanuspereirai from the Atlantic croaker and spotin the Gulf of Mexico constitutes a separate,closely-related species. Fusco (1978a) hasdescribed the larval stages of that worm.
Leptolaimus sp. (Leptolaimidae) from onthe gills and from both within the lumen ofand adjacent to the alimentary tract ofbrown and white shrimps was reported byOverstreet (1973).
Rigdon and Baxter (1970) first reportedspontaneous necrosis of abdominal musclesof penaeids, and that work has beenfollowed up by Lakshmi, Venkataramiah,and Howse (1978), who showed that thelesions, presumably caused by musclefatigue heal faster in oxygenated ratherthan aerated water. The cramped conditionwas first reported by de Sylva (1954) and thensubsequently by Johnson, Byron, Lara M.,and Ferreira de Souza (1975). Venkataramiah, Lakshmi, and Biesiot at GCRLhave found that cramping occurs most oftenin conditions of low salinity and temperature. They (personal communication) alsodetermined that it results from an internal
ionic imbalance, and they- are presently-trying to identify the specific ion or ionsinvolved. Both necrosis and crampingtypically affects shrimps in tanks for baitdealers and in culturing facilities. As culturing shrimp becomes more widespread,especially with heated or cooled water, theseand other diseases will appear. One of theseothers is gas-bubble disease (Lightner,Salser, and Wheeler, 1974), and it alsoaffects oysters and clams in water supersaturated with oxygen or nitrogen.
Overstreet (1973) discussed the goldenshrimp disease and also reported andreviewed some fouling organisms. Manyorganisms foul shrimp, even a barnacle thatsettles on the blue crab (Dawson, 1957).
Just like different tissues of all animals,those of shrimp degenerate at different ratesafter death. Lightner (1973) described thesepostmortem changes in the brown shrimp atdifferent temperatures.
GRASS SHRIMP
In addition to his illustrated handbook
Technical Aspects 107
(1975) on shrimp diseases for thoseculturing shrimp, S. Johnson (1977) alsoprovided a similar handbook on diseasesof crayfish and freshwater shrimp.Although not complete for diseases of grassshrimp, that pamphlet provides the onlysingle source of parasites of that host. Itcontains much useful information, sometaken from an earlier paper (S. Johnson,1974). Many of the protozoans symbiotic onthe grass shrimp were listed by Sprague andCouch (1971). The findings of more speciesplus completion of considerable detailedbiological studies on many protozoans ofgrass shrimp and other crustaceans haveoccurred since 1971. Clamp (1973) providedobservations on Lagenophrys lunatus, andPhyllis Bradbury has worked up severalother ciliate-grass shrimp relationshipsincluding that for Terebrospira chattoni(see Bradbury, Clamp, and Lyon, 1974;Bradbury and Goyal, 1976). Sprague (Bullaand Cheng, 1977) listed three microsporidans from Palaemonetes pugio andmore information on those have been
presented by Street and Sprague (1974) andOverstreet and Weidner (1974).
The bopyrid Probopyrus pandalicola hasbeen studied by Richard Heard (unpublished). Also, its respiration in both larvaland adult stages has been investigated inrelation to its two hosts (Anderson, 1975a;1975b). Both the infested copepod andshrimp respired less than their respectivecontrol counterparts. This decreased use ofoxygen may result from a lipid buildup ininfested hosts requiring less energy tometabolize. When fish have parasiticinfections, they generally respire more,which in turn increases their stressedcondition.
FISHES
A general treatment of the biology offishes can be obtained from a multitude of
books available in most any library.Identifications of most species from thenorthern Gulf of Mexico can be made usingkeys in a book by Hoese and Moore (1977).Walls (1975) also wrote a recent book onthese fishes. A standardized list of common

108 Marine Maladies?
and scientific names acceptable to theAmerican Fisheries Society was published(Bailey etal., 1970), and another edition willupdate it in the future. These are commonnames used in most scientific literature, andnot necessarily those used in specificgeographic localities by fishermen. Olderworks, such as an article by Earll (1887) usedfor comments on the early mullet fishery,often use scientific and common names no
longer in use today.Because of recent interest in elasmo
branchs and the potential for investigatingevolutionary aspects of some highly specificelasmobranch parasites, I mention a fewreferences. A book edited by Gilbert,Mathewson, and Rail (1967) covered mostaspects of elasmobranch biology except theparasites. More current evaluation of interrelationships among elasmobranch groupshas been presented by Zangerl (1973) andCompagno (1973; 1977). Means for criticalidentification of species have been given byBigelow and Schroeder (1948).
No single source covers parasites frommarine or estuarine fishes. Hoffman (1967).however, covered parasites in North American freshwater species, and that book mightbe helpful for identifying some parasites. Arather extensive compilation of mulletparasites and diseases (Paperna andOverstreet, in press) may also apply to ageneral investigation of conditions in anyfish. Control and treatment of freshwater
diseases have been covered extensively(Hoffman and Meyer, 1974). Methods fordiagnosing and controllingselected diseasesin salmon, pompano, and striped bass havebeen edited by Sindermann (1977), but thesame statements also apply to several otherhost-disease relationships.
Microbes
Plumb (1974) reviewed all known viraldiseases of fishes from the Gulf region.Lymphocystis from Atlantic croaker wasfirst described by Howse and Christmas(1970; 1971) and that from the silver perchby Lawler, Howse, and Cook (1974). A briefreview of infections including some newdata was presented by Overstreet and Howse(1977).
Bacterial flora including commensal anddisease-causing organisms have beeninvestigated more thoroughly than viruses.Horsley (1977) reviewed the bacterial flora offishes and concluded that the flora in a fish's
alimentary canal was about the same as thaton the gills and skin, which, in turn,depended on the state of the environment.In other words, monitoring bacteria andwater quality in ponds should help predictpotential disease outbreaks.
The natural bacterial epizootic in thenorthern Gulf was first reported by Plumb.Schachte, and Gaines (1974) and laterstudied in more detail by Cook and Lolton(1975). Overstreet and Howse (1977) discussed some bacterial infections in
Mississippi coastal areas.A book with parts by G. Bullock, Conroy,
and Snieszko (1971) and by G. Bullock(1971) deals specifically with the methods ofworking with bacteria from fishes. Two ofseveral papers concerning identification ofbacteria are by Glorioso, Amborski, Larkin,Amborski, and Culley (1974) and by Slums(1976). The primary reference is Bergey'sManual (Buchanan and Gibbons, 1974).
Protozoans
Protozoology and even the study ofparasitic protozoans represent fields untothemselves. The advances in chemistry,ultrastructure, and culturing techniqueshave allowed a much better understandingof the relationships among different groups.In 1964, Honigberg and a group of othersformulated the committee's revised
classification of the Protozoa. Since then
several major changes have been suggestedsuch as those by de Puytorac and others(1974)and Corliss (1974a; !974b)forciliates,and by Sprague (Bulla and Cheng, 1977) formicrosporans. Several books coverprotozoans in general such as those by Kudo(1966), Grell (1973), and Sleigh (1973).Levines' book (1973) specializes in parasiticforms primarily from domestic animals.Even though helpful, that book unfortunately did not treat species from fishes.The biochemistry of parasitic forms hasbeen summarized by Gutteridge andCoombs (1977). Several papers on specific

species or groups are cited below.Lawler (1977) has shown that all but 6 of
over 80 tested fishes (different species)succumbed to heavy infestations of thedinoflagellate Amyloodinium ocellatum.Those six fishes usually appeared refractoryto the invader. Occasionally an individualof one of those species would becomeinfested or even die, but in those instances,
another less susceptible fish was usuallypresent. Lawler suggested the resistanceof Cyprinodon variegatus, Fundulusgrandis, Opsanus beta, and others mightrelate to the ability of those fish to toleratelow oxygen concentrations or to produce anabundance of mucus, either with or withouta repelling substance. Lorn and Lawler(1973) described the attachment mechanismof the related Oodinium cyprinodontumwhich infests gills of killifish and othercyprinodontid fishes along the Atlantic andGulf coasts. Larger and more elongate thanAmyloodinium ocellatum, it harborschloroplasts. Rather than penetrating anddigesting host tissue like that comparedspecies, it attaches without harming the hostand produces its own nutrients like a typicalplant. That Oodinium cyprinodontum hassuch characteristics favor the host, since asmany as 2000 dinospores (about eight timesas many as for Amyloodinium ocellatum)can form from one trophont.
Several chemical treatments cited in
aquarium publications as curing Oodiniuminfestations have not been successful in
treating heavily-infested fish. Lawlei (1977)listed several of these and emphasized thevalue of a short freshwater bath.
A general treatment of piscine flagellates(Becker, 1977) covered all different groups.Becker and I are presently studying someblood parasites in fishes from Mississippi,and comments on leeches as vectors for these
occurred in the paper by Sawyer, Lawlei.and Overstreet (1975). Dealing specificallywith trypanosomes. Khan (1977) showedexperimentally that at least Trypanosomamurmanensis in Newfoundland, Canada,infected a variety of related and non-related hosts. After being transmitted bythe leech, the trypanosome continuously-increased in size for about 2 months. Thedifference in size of blood flagellates and
Technical Aspects 109
paucity of observations have influencedsome biologists to assume that several ratherspecific trypanosomes occurred in sonic-regions. Ultrastructure of stages in the leechhas been investigated by Desser (1976).
Amoebae have been encountered in
marine fishes. Orias and Noble (1971) listed the four reported species. All but oneinhabits the alimentary tract, and it occursin the branchial mucus and skin. None
appears pathogenic to the fish, but freshwater amoebae have been implicated inmortalities, possibly in conjunction withassociates (Rogers and Gaines, 1975;Sawyer, Hoffman, Hnatli, and Conrad.1975). Living individuals move aboutactively with their pseudopodia; in somecases they even feed on their own protectivecyst stage in addition to expected food particles (W. Bullock, 1966). Critical examination of intestinal tissue and contents mightreveal many amoebae in marine fishes.Based on other reports, individuals shouldbe more prevalent when intestinal flagellates also cx-cur. and they probably occur inclusters. Perhaps given appropriatestimulation, some of the commensalamoebae become virulent.
Several books treat coccidiosis in general(Kheysin, 1972; Levine, 1973; Hammondand Long, 1973). A brief treatment by Lorn(1970) summarized much of the informationabout piscine species, and many new specieshave been described since then. Lorn (1970)also treated hemogregarines and other protozoans of fish, whereas Becker (1970)confined his chapter in the same book toprotozoans in fish blood. Davies andJohnston (1976) studied Haemogregarinabigemina, a species that reproducesasexually in peripheral blood of a blennyand other hosts; they presented strongevidence that the isopod Gnathia maxillarisacted as the vector. For someone interestedin the fine structure and locomotion ofhemogregarine gameiocytes, the most observed stage in fish, Desser and Weller(1973)provided an informative paper. Olscn's(1974) book presented a diagram of a lifecycle for a species utilizing a turtle and aleech, and it also gave several references forobtaining information on all stages.Generally not considered pathogenic,

110 Marine Maladies?
hemogregarines include a species(Haemogregarina acipenseris) in a sturgeonfrom the Volga River that apparently causesanemia and wasting in its host (seeShulmanand Stein, 1962).
Microsporidans have been discussed indetail in the section on crustacean hosts.
The best single source of review articles andreferences on all aspects of the group is thetwo volume work edited by Bulla and Cheng(1976; 1977).
The extensive literature about myxosporidans primarily deals with spore sizeand shape and with hosts. Increased numbers of ultrastiuctural investigations on thespores and on developmental stages shouldenhance the knowledge of the groupconsiderably. Kudo (1920) and Shulman(1966) wrote the classic treatments on thegroup, and Paul Meglitsch of DrakeUniversity- has an ii|xlated monograph inprogress.
Lorn (1969), an active worker withmyxosporidans. compared the developmentof the polar filament and its external tubethai is drawn into the capsule with thesimilar process for nematoc yst developmentin cnidarians. He used this and other fea
tures, all of which contrast with those of a
microsporidan and its polar filament andwith a ciliate and its extrudiblc organelles,to relate myxosporidans more closely withcnidarians than with other protozoans. Bothgroups also have dissociated generativecells.
The possession of more than one cellneeds further explanation. When mostdisporous species develop (two sporeswithin a membrane), ten cells form the twospores and one forms the pansporoblastchamber. For a spore, one cell forms eachof the two valves and each of the two polarcapsules including their contents. Thenuclei of a binucleate cell often fuse to
produce the infective amoeboid stage.Pathological alterations in Cyprinodon
variegatus (sheepshead minnow) infectedwith Myxobolus lintoni have been treatedby Nigrelli and Smith (1938). Overstreet andHowse (1977), and others. Articles on troutwith whirling disease probably surpass innumber those on any other myxosporidan.Recently, Wolf and Markiw (1976) have
artificially- cultured spores from the vegetative stage. Hoffman (1974), who hasconducted much research on means to
control whirling disease, reported on the useof ultraviolet irradiation to control both it
and a variety of other fish parasites.Treating ponds in order to control the disease is not always entirely effective(Hoffman, 1976; Hoffman and O'Grodnick1977).
The myxosporidan cysts portrayedbetween the fin rays of the spot probablyare referable to the same species ofHenneguya that occurs between the soft raysof the dorsal and anal fins of the seatrouts
Cynoscion nebulosus and Cynoscionarenarius in Mississippi Sound and thatreported but never named from Cynoscionregalis in New York and Virginia byJakowska. Nigrelli, and Alperin (1954).
Other than three species, one from anannelid, one from an insect, and one from adigenean, all reported true myxosporidansinfect cold-blooded vertebrates. The pathogenic Fabespora vermicola from thedigenean Crassicutis archosargi and itsunusual ability to move by itself was reported by Overstreet (1976a). If the flukeharmed its sheepshead host, the myxosporidan could be considered for possibleuse as a biological control agent. The fluke,however, even though attaching to theintestine by an unusual adhesive mechanism (Overstreet, 1976b), apparently doesnot harm the fish.
References on ciliates in general were presented at the beginning of this section. Byculturing a few ciliates. applying theprotargol and silver impregnation techniques, maybe using an electronmicroscope, and exercising considerablepatience, students and other scientists canstill solve numerous economically andacademically important problems withciliates.
Trichodinids of marine fishes have been
studied most extensively by Lorn, whopresented a nicely illustrated study (1973)ontheir adhesive disk. They have been reportedto cause mortalities in marine hatcheries(e.g., Purdom and Howard, 1971; Pearse.1972), but dying fish often also harbormonogeneans, copepods, or dinoflagellates

in addition to the ciliates that may helppromote their multiplication. Severalspecies cause mortalities in freshwateraquaculture facilities. As examples (Meyer,1970), trichodinids kill channel catfish fry-within an hour after hatching and keepyoung golden shiners from feeding.
Red sore disease has been considered a
problem for some time to people withfreshwater fish-ponds (Rogers, 1972). Inaddition to low oxygen concentration andhigh organic content, other factors canpromote the disease. Esch, Hazen. Dimock,and Gibbons (1976) studied the Aeromonas-Epistylis complex in centrarchid fishesfrom a cooling pond for a nuclear production facility and found that more fishfrom the thermally-affected region hadred sores than did those from a region withwater of ambient temperature. Presumably,a variety of pollutants that stress the hostand encourage growth and reproduc lion ofthe ciliate and the bacterium will enhancered sore disease.
Overstreet and Howse (1977) studied theattachment of the stalk's terminal plate tohost tissue and suggested that the fibrillarattachment complex of Epistylis sp. slowlyinfiltrates and erodes the host tissue leavingscale or bone as the only remaining sites onwhich to attach. Fish heavily infested oftenhave exposed bones and extensive tissuedestruction and regeneration. Thoseauthors point out that hemorrhaginglesions seem to depend on the presence ofscales. Catfish, without any scales, harbordense growths of the same or similar speciesof Epistylis without much hemorrhagingeven though bones may be exposed.
The major group Holotrichia no longerhas acceptance in the recent classificationschemes of de Puytorac and others (1974)and Corliss (1974a; 1974b). The term,however, still has some useful value innontaxonomic works because of its longworkable and familiar nature. An outbreak
of Cryptocaryon irritans in fishes from bothexperimental and public aquaria wasdiscussed by Wilkie and Gordin (1969).Hoffman, Landolt, Camper, Coats,Stookey, and Burek (1975) provided a key to"holotrichs" of freshwater fishes anddiscussed pathological responses by hosts,
Technical Aspects 111
especially those resulting from Tetra-hymena corlissi. That facultative parasitekilled fish, but fish could not be experimentally infected with it.
Specimens of the sheepshead minnowfrom Horn Island infested with the
hypotric h were collected by Adrian Lawler.and the progress of the disease was followedby the two of us (unpublished data).
Cestodes
Yamaguti (1959) compiled a book on allcestodes known at that time, providingkeys to genera and higher groups. A moreup-to-date set of keys was written by-Schmidt (1970). but neither helps identifymost larval forms. Some of these larvae from
Gulf molluscs can be sorted out using a keyby Gike (1976). Wardle and McClouds(1952) book on tajM-wonns remains theclassic for a general treatment.
Aspects of the ecology and taxonomy ofthe pseudophyilidean Anantrum tortumwere presented by Overstreet (1968). Rees(1969) presented a more detailed account ofseveral of the characters of the same worm
using the junior synonym Acompsoceph-alum tortum. Before this present report,the worm had never been reported from theGulf of Mexico, even though the worm isextremely abundant. I have confirmed thatit has operculated eggs. Recently Jensen andHeckmann (1977) described a secondspecies, Anantrum histocephalum, from alizardfish in California. That worm,however, has a mushroom-shaped scolex toassure attachment within the gutepithelium.
Overstreet (1977b) reported the ecologicalaspects of infections of Poecilancistriumcaryophyllum in the spotted seatrout.Moreover, Tom Mattis has been investigating the life-history of that worm and ofseveral other elasmobranch cestodes. He is
the authority from the Gulf region onelasmobranch cestodes; others that work onelasmobranch cestodes are Ronald
Campbell of Southeastern MassachusettsUniversity and Murray Dailcy of CaliforniaState University at Long Beach. Dailey andOverstreet (1973) described the multi-strobilate forms of Cathetocephalusthatcheri from the bull shark.

112 Marine Maladies?
MonogeneansRather than being most closely related to
digeneans, monogeneans probably evolvedfrom an ancestral form that gave rise tocestode groups. Malmberg (I974) discussedhis ideas and reviewed much of the literature
on monogenean phylogeny.A book by Yamaguti (1963a) provides keys
to the families and genera of monogeneansand listings of species. A more updated keyfor North American genera by Schell (1970)may prove more economical and up to datefor a beginning student.
Many laxonomic problems still needattention, including numerous descriptions of pathogenic species. In fact, AdrianLawler at GCRL plans to describe the newspecies of Dermophthirius that harmed itssawfish host. Lawler and Cave (1978)already reported on Aspinatrium pogoniaewhich built up infestations to thousands ofindividuals and apparently killed a largecaptive black drum. Large worms measuredover 12 mm long, but most were smaller.Many species will multiply extensively incultured fish and kill or weaken them, but
few cases have been reported in theliterature.
Some monogeneans that occur inMississippi may be near the limit of theirgeographic range, being rare except underspecific conditions. The relatively rareAbsonifibula bychowskyi may be anexample of this (Lawler and Overstreet,1976). We found that the intensity ofinfestations related directly with watertemperature.
The best understood monogeneaninfesting skin does not occur in local waters.Still, a brief understanding of the biology ofEntobdella solea from the ventral surface of
the sole should inspire extensive investigations of local species (Kearn, 1971).This5 mm worm moves about on the bottom of
the flatfish, feeding on tissue overlyingscales. It lays eggs with long sticky filamentson sand grains during the day when the hostis buried. Fish feed during dark hours, andabout dawn they return to partially bury inthe sand. At that time, a ciliatedmonogenean larva (onchomiracidium)hatches and infests the exposed upper headof the sole. As it matures, the worm migrates
to the fish's belly. It embeds its anchors intothe host, but holds on primarily by suction.Accessory hook-like structures anterior tothe anchors actually prop up the central partof the holdfast making a cup. In some otherspecies, the anchors penetrate more deeplyand hold the host in a manner similar to an
ice long when the fish is active. When thefish slows down, some worms' marginalhooks suffice for attachment (Lester, 1972).
Studies by Paperna (1964) provided anunderstanding of infestations by more thanone monogenean species in pond-rearedfish. He found that heavy infestations ofDactylogyrus vastator on young carp causedincreased growth of host tissue (hyperplasia) and created an unfavorableenvironment for two other monogeneanspecies. The small worm can kill many carpfry, but fingerlings develop a resistance, sothat monogenean infestations in larger fishinclude other less pathogenic species only.If the effect of temperature, salinity, andother physical parameters on larval andadult worms and the time when fry firstappear in ponds are understood, fish can beproperly managed to avoid infestations(e.g., Paperna, 1963). A biologist, however,must realize that not all monogeneans' eggshave to hatch quickly; some develop-mentally inhibited larvae over-winter andhatch about the time young fish appear.
Another group of tiny monogeneans(gyrodactylids) produces embryos ratherthan eggs, and many species can coexist onthe same host (e.g., Williams and Rogers,1971, for species on the sheepshead minnow). Some species cause mortalities andothers inhabit atypical sites. One specieseven infests the brood pouch of the maleGulf pipefish (Holliman, 1963).
A large number of the marine species inthe northern Gulf have been reported ordescribed in papers by Koratha (1955a;1955b) and Hargis (1957, for summary).Koratha (1955a) also suggested a possiblephylogenetic relationship between cobiaand diskfishes indicated by theirmonogeneans. This relationship has beenquestioned by several ichthyologists.Several other groups of monogeneans arealso restricted to specific families or othergroups of fishes. On the basis of Hargis'

report on host-specificity, each of 67 monogeneans from the northern Gulf infested asingle host species, whereas only 8 parasitized two and 1 parasitized three. When morethan one host was infested, they belonged tothe same genus. Similar levels of specificityto single hosts occur in the White Sea,Barents Sea, Black Sea, and Great Barrier
Reef off Australia (Rohcle, 1977).
DigeneansYamaguti (1971) revised an earlier
compilation of all digeneans of vertebrates,giving keys to genera and families andlisting species with their hosts. He (1975)also compiled most known data on lifecycles. Both lengthy works are extensively-illustrated. Shell's (1970) key allows someidentifications and is much cheaper in pricethan Yamaguti's book, but does not includeas many local marine and estuarine species.
A book by Erasmus (1972) deals withexperimental investigations of trematodcbiology, emphasizing larval stages.Lumsden (1975) reviewed the function andstructure of helminth teguments, includingthose of trematodes, and Overstreet (1976b)described the unusual attachment to a fish
host by Crassicutis archosargi.For further discussion on some ol the
examples presented, the reader can refer tothe following works: Gibson and Bray(1977) considered the giant fish stomachworm from a variety of hosts as the singlespecies Hirudinella ventricosa until furtherevidence confirms a second species, Iversenand Yoshida (1957) reported the commonoccurrence of two specimens of that giantdigenean per wahoo, and Watertor (1973)discussed variations in infections of it fromtunas. A general discussion on melanin-containing cells in teleostswas presented byRoberts (1975), and Dubois' (1968; 1970)synopsis covered the taxonomy and lifehistories of most strigeoids. Overstreet(1970; 1971a; 1971b; 1971c; 1977a) treatedMetadena spectanda, Saturnius maurepasi,and the digeneans illustrated in Figures 143to 145.
Some discussed works are still in progress.Heard and I will describe the life history ofPhagicola longus, and I will report on
Technical Aspects 113
ecological aspects of many parasites fromthe Atlantic croaker including digeneans.Courtney and Forrester (1974) reported onthe worms from the brown pelican. Someheterophyids infect their intermediate hostsin large numbers. Paperna and Overstreet(in press) reported as many as 6,000heterophyid metacercariae per gram oftissue from some mullet in the Sinai.
Even though fishes do not act as paratenichosts for digeneans as frequently as forcestodes, nematodes, and acanthocephalans,some still function in that role. So can
cephalopods (squids and related molluscs)which parallel fishes in many respects inaddition to harboring the same digeneans(Overstreet and Hochberg, 1975).
For a fairly recent discussion onphylogenetic and taxonomic relationshipsof digeneans, readers should refer to a paperby Cable (1974). I hope Raymond Cable willshare more of his knowledge about cercariaewith us in the future.
Chuck Getter, a graduate student ofC. Richard Robins at the University ofMiami has been investigating the endangered key silverside, Menidia conchorum.He sent me preserved material infected withacanthostomes. Specimens of sheepsheadminnow and sailfin molly also hadinfections. To date, only threeacanthostomes have been reported from theUnited States, and the larva, based on thepresence of 24 oral spines and othercharacters, appeared most likeAcanthostomuw. coronarium (see Brooksand Overstreet, 1977); however, we havestudied no crocodilian worms from SouthFlorida and additional species with similarfeatures may occur there.
Anatomical aspects of the four species ofmarine aspidogastrids in the northern Gulfhave been treated by Hendrix and Overstreet(1977).
A review of American paragonimiasisincluding the pairing and encapsulation ofindividuals was presented by Sogandares-Bernal and Seed (1973). Literature on manymarine zoonoses (parasites, includingdigeneans, that could infect humans and,in some cases, pets) has been synthesized byWilliams and Jones (1978).

114 Marine Maladies?
Nematodes
As he has done for other helminths,Yamaguti (196I) has compiled a listing ofnematodes parasitic in vertebrates, and heincluded keys to genera and higher taxa.These keys, but not the listings, have beensuperseded by a series of keys edited by-Anderson, Chabaud, and Willmott (1974a,1974b. 1975a. 1975b). Number 1 in this
series also includes a glossary which shouldstraighten out some of the confusion amongterms. A series of books in Russian have
compiled much of the literature cm somegroups, and a few have been translated intoEnglish (e.g., Skrjabin, 1919; Ivashkin,Sobolev. and Khromova, 1971). A number ofother books has been written on various
aspec ts of nematodes, and, for the presentpur|K>ses, I mention those by Chitwood andChitwood (1950) and by Bird (1971) only.
Information on the striking reddishcamallanids in Gulf marine fishes has justbeen published (Fusco and Overstreet,1978). Spirocamallanus cricotus infectsshallow inshore fishes. Actually more thanthe 13 reported hosts can harbor infections.In water 27 meters and deeper, Spirocamallanus halitrophus infects at leastbothid flounders and a cusk-eel. Fusco
(1978b), using isoelectric focusing andspectrophotometric characterization, hasdifferentiated hemoglobins of the Atlanticcroaker, male Spirocamallanus cricotus,and female Spirocamallanus cricotus. He(1978a) also obtained experimentalinfections of this worm in a harpacticoidcopepod (Tigriopus californicus) and in thewhite shrimp.
Adults of the common Thynnascarisreliquens were described by Norris andOverstreet (1975). Those authors (1976) alsoreported larval members of that genus ininvertebrates and the ability of at least onemember to invade mice. Ebert (1976)examined this latter matter in more detail.
Thomas Deardorff of GCRL hopes to relatea few larval species of Thynnascaris withtheir adult forms. Much has been written
about the disease anisakiasis, especially inJapan. Oshima (1972) gave a good reviewwhich has been updated by several workers(e.g., Jackson, 1975; Margolis, 1977;and inprogress by John Smith of the Department
of Agriculture & Fisheries for Scotland.Aberdeen, Scotland).
A general treatment of nematodes inmarine fishes by Margolis (1970) gave afairly complete idea of the different majorgroups infecting fish. More groups thanthat, however, infect fish. A critical studyon the life cycle of Dioctophyma renale byMace and Anderson (1975) emended some ofthe interpretations by Karmanova (1962).who confirmed that earlier workers had
indeed mistaken a gordiacean larva from thebranchiobdellid from crayfish as Dioctophyma renale. The duck mortalities resulting from their eating fish withEustrongyloides sp. were described by-Locke, DeWitt, Menzie, and Kerwin (1964).
Rosenthal (1967) fed wild plankton tolarval herring and many of these young fishapparently started dying from larvalascaridoid larvae after about 11 days. Theworms grew rapidly and were quite active.After the fish reached 2 cm in length, theybecame less seriously affected by the worm.More studies of this nature should be con
ducted. Problems arise when usingpredatory fish, however, because theytypically cannibalize weakened individuals.Consequently, several predatory fish cannotoccupy the same aquarium and at the sametime provide meaningful results.
AcanthocephalansYamaguti (1963b) listed the known
acanthocephalans and presented keys to thehigher taxa. Golvan (1969) provided descriptions and keys to the species ofpaleoacanthocephalans, a major groupinfecting fishes. Authorities do not agreeabout the higher classification of acanthocephalans, even though about 1000 specieshave been described. W. Bullock (1969) discussed the anatomy of these worms anddiscussed many features in relation to theiruse in classification and systematics. A morerecent treatment on the phylum was presented by Schmidt and Roberts (1977).
One of several papers on acanthocephalans covers most of the marine fishspecies in the northern Gulf (W. Bullock,1957). Buckner, Overstreet, and Heard(1978) reported intermediate hosts for

Tegorhynchus furcatus and Dollfusentischandleri, and Overstreet is preparing aseveral-year ecological study on the latterspecies in the Atlantic croaker.
Crustaceans
Kabata (I970) and Schmidt and Roberts(1977) treated many aspects of crustaceanparasites. Kabata also has other books inprogress which should be classics. Schultz's(1969) key can be used to identify mostAmerican isopods, and Yamaguti's (1963c)book lists and illustrates most piscinecopepods.
The example discussing altered behaviorof infested menhaden was by Guthrie andKroger (1974), and that on infested and debilitated gar was by Overstreet and Howse(1977).
Life cycles of isopods on fishes needconsiderable attention, especially in light ofthe potential economic loss to cultured fishcaused by isopods. A group of isopods notmentioned in the first section because mem
bers do not appear to be a problem in thenorthern Gulf of Mexico is the Gnathiidae.They may be a threat in south Florida. Inthe Red Sea, one isopod that can bring fishculture to a halt has been investigated(Paperna and Por, in press; Paperna andOverstreet, in press). The larval gnathiid(praniza), unlike the adult, obtains a bloodmeal from almost any fish, leaving that fishwith integumentary wounds, anemic, andoften dead. The larva feeds on fluid from afish's gills or skin for about 2 to 4 hours atnight and may do this three times beforemolting into an adult. A variety of fishspecies maintained in cages near shore in1 to 2 meters of water attracted the isopod,but those in cages from deeper water of 6 to8 meters about 100 m offshore did not. Once
attacked, an individual fish rarely becomesinfested again. After obtaining the bloodmeals, the isopod molts to an adult in abouta week. Eggs develop in the female's broodpouch for about 3 weeks and then the larvae,numbering about 90 in a full-sized female,emerge. Larvae feed on fish after about 9 to10 days of development and live in the mudin shallow habitats.
Information on life cycles of cymothoid
Technical Aspects 115
species remains incomplete. Menzies,Bowman, and Alverson (1955) piecedtogether the cycle of Lironeca convexa onthe Pacific bumper. Presumably free-swimming juveniles find the appropriatehost, infest and feed on gill tissue, moltinto a male, transform into a female, mate,and produce young as a commensal in thehost's mouth. A similar progression doesnot occur for all species.
Two of the most troublesome copepodgroups to identify and understand arc theergasilids and caligids. Roberts (1970) andJohnson and Rogers (1973) provided keysand data on the tiny ergasilids, and Papernaand Zwcrner (1976) gave a good example ofecological aspects of an infestation ofErgasilus labracis on the striped bass.Margolis, Kabata, and Parker (1975) provided a catalogue and synopsis on the many-species of Caligus, and Cressey (1967)reviewed pandarids from elasmobranchs.
Myron Loman has investigatedLemaeenicus radiatus in my laboratory.Using the rock seabass as an intermediatehost, he obtained results similar to Shields(1970), who used the black seabass from theAtlantic coast. Copepodid larvae infest thefish, pass through a chalimus stage, mate(the male breaks its frontal filament andtransfers its spermatophores to the female),leave within 3 to 5 days, infest another fish,mature, and produce eggs within a week.The Gulf killifish acted as a good final host,but readily succumbed when more than acouple of individuals infected it. Since over100 infective larvae can infest a bass at onetime, a few killifish left with a bass in anaquarium will usually die unless removed atproper intervals.
Considerable data concerning biologicalaspects of copepods have been gathered.Cressey and Collette (1970) provided a goodexample of copcpod-needlefish relationships which could act as a guide for furtherworks on other copepod-host relationships.By examining the stomach contents of re-moras, sharksuckers, and other diskfish,Cressey and Lachner (1970) discovered thatcopepods appeared to play different rolesas the food source for different diskfish.Also, age of fish determined food preferences. Young Remora remora ate more

116 Marine Maladies?
parasites than older individuals, whereasolder Remora osteochir ate more than
younger fish. Bernett-Herkes (1974) foundthat the presence of some copepods fromdolphin (fish) appeared to be influenced bysize of host and by the presence of othercopepod species.
Many argulids prove difficult to identifyusing old literature. Most likely, such problems should be solved by preliminary andfuture keys by- Cressey (1972).
MISCELLANEOUS HOSTS
The American AlligatorThe alligator harbors parasites that, for
the most part, can be seen with the nakedeye. Large nematodes in the stomach may- beone or a combination of at least four adult
ascaridoids. A new species of Goezia is beingdescribed by Deardorff and Overstreet, andthe other three are being reviewed by JohnSprent of the University of Queensland. Hisworks will include most known crocodilian
nematodes.
The most conspicuous digeneans are theacanthostomes. Sometimes hundreds of
the nearly 1cm long species inhabit a singlealligator. Brooks and Overstreet (1977)described or presented supplemental data onthe three known species from the UnitedStates. Other digeneans occur in lessernumbers in hosts inhabiting fresher waters.Brooks, Overstreet, and Pence (1977) reported the proterodiplostomes, and Brooks andOverstreet (1978) described the onlyliolopid. The digenean Odhneriotremaincommodum from southern Florida deserves comment because of its unusuallocation even though it does not infectalligators in Mississippi and Louisiana.This large worm attaches firmly to thepharynx or buccal cavity of its host, and anobserver can see it by using extreme care. Itslife cycle presents a good example of aparasite that infects a specific sex ofhost. The Florida gar (Lepisosteusplatyrhincus), which does not occur inMississippi, harbors the encysted meta-cercariae primarily in the ovary, hence apreponderance of infections in the femalegar (Leigh, 1960).
By considering natural phylogeneticgroups of both crocodilians and crocodiliandigeneans, Brooks (dissertation in progress.University of Mississippi) presented evidence that both groups evolved together.About 200 million years ago all land massesthat harbor crocodilians today were joinedtogether as one body of land (NorthAmerica, South America, Africa, Asia,and Australia). By the end of the next 50million years, that land mass had completely broken up. Because several speciesof the worms belonging to the same generaoccur dispersed in most regions today andbecause intermediate hosts of most groupscannot tolerate saltwater, the actual orancestral digeneans appeared to already beinfecting crocodilians 200 million yearsago.
A stronger relationship between crocodilians and birds than between crocodiliansand other reptilians is suggested by thepresence of strigeoids, clinostomes, andechinostomes inhabiting both crocodiliansand birds, but not other reptiles. Brookswill provide details of the association in thenear future.
The presence of the turtle leechPlacobdella multilineata on the alligatorhas been reported only recently (Forresterand Sawyer, 1974) and then from Florida. InLouisiana, it can be seen commonly ongators in freshwater. Just as in Florida, thehosts have a species of Haemogregarinain the blood which could be transmitted by
that leech.
The pentastome Sebekia oxycephalaoccurs sometimes in large numbers in thelungs of alligator hosts. It also occurred inthe liver and mesentery of some hosts. Thealligator acquired its infections from avariety of fishes (Venard and Eangham,1941; Dukes, Shealey, and Rogers. 1971).Self (1969) presented a review and bibliography of pentastomes in general.
Birds
Symbionts and diseasesof birds have beentreated extensively in a book edited by-Davis, Anderson, Karstad, and Trainer(1971). Another general book byRothschildand Clay (1957) placed more emphasis on

the arthropods. In order to appreciate thewealth of information available on the
helminths of just the Anatidae. one can inspect the bibliography and catalogue by-McDonald (1969a; 1969b). Sources toidentify helminths are basically the sameas for helminths infecting fishes.
Marine and estuarine fish play an important role in transmitting helminths tobirds. As already shown earlier, they readilytransmit numerous digeneans and nematodes to birds. To a lesser extent, these
fishes transmit cestodes, acanihoccphalans.and other helminths. As an example,killifishes can be cited for hosting cestodesbelonging to the genus Parvitaeniawhich infect ibis, herons, pelicans, andother birds as determined by Richard Heard(personal communication) and the roughly5 cm long acanthocephalan Arhythmor-hynchus frassoni which infects the clapperrail (Nickol and Heard, 1970).
Much information known about parasitesof local birds has been reported by Danny-Pence of Texas Tech University who studiedthe numerous nasal mites from Louisiana
birds (Pence, 1973b; 1975). In addition tothose parasites, he reported on a variety ofworms that have- importance foi differentreasons. As examples, the digeneanAmphimerus elongatus occurred in thethousands in the- bile ducts of the- double-
crested cormorant, causing extensivecirrhosis and other pathological alterations(Pense and Childs. 1972), and the nematodeTetrameres aspinosa encysted in pairs in theproventriculus of the snowy egret (Pence,1973c). Nothing was particularly notableabout that tetramerid infection except thatanyone examining the proventriculus andstomach of any birds should be aware of thepresence of those encysting nematodes. Theembedded blood-red female, rather thanlooking nematode-like, is extended greatlyabout the midbody. Either the males occuiencysted with the females in fibrous cysts orfree in the stomach or proventriculus, depending on the species. The conspicuouscysts with red worms can be seen throughhost tissue, appearing at first glance likehemorrhagic lesions. An extensive reviewof the group is in preparation by TonyMollhagen (1977. abstract) working under
Technical Aspects 117
the direction of Pence.
Pence (1973a) reported the pe-ntastonu-Reighardia sternae from gulls and terns inLouisiana. It also infects the lungs and airsacs of at least the- laughing gull andcommon tern in Ocean Springs. Ratherunique in that it infects birds, it may alsobe unusual in not requiringan intermediate-host (Riley. 1972a). However, like thespecies in the alligator. Reighardia sternaemay also be transmitted by fishes (Bakke.1972). Those interested in the variety offeeding mechanisms that parasites use- toingest host blood should consult papers byRiley (1972b; 1973) who reported observations and described the method used by thispentastome.
In order to identify and study arthropods,a reader could initiate his literature search
using publications by Emerson (1972a:1972b; 1972c) for lice. Coolc-y and Kohls(1945) for ticks. Pence (1972) for subcu
taneous hypopial miles, and Pence (1975)for nasal mites. The reference to the bloody-ulcers produced by a louse in the- mouth ofthe white pelican was by Wobeser, Johnson,and Acompanado (197-1) and that to thepossible evolutionary history of flamingo-lice- was by Rothschild and Clay (1957).
This latter relationship among theflamingos and other gioups it-mains unsettled. Osteological evidence based on acombination of characters of fossil
flamingolike waders from the Lower Eocenesupports the- relationship of flamingoswith anseriforms (ducks) and also withcharadriiforms (shorebirds) (Feduccia.1976). In fact, similarities in cestodes occur
between flamingos and shorebirds (Baer,1957). However, the consensus of mostpresent-day ornithologists (Oscar T. Owrc,personal communication) supjxms Sibleyand Ahlquist (1972) who believe that flamingos, while related to ducks, align closerwith Ciconiiformes (herons), and that, assuggested by Mayer and von Keler (see review by Sibley and Ahlquist). the licetransferred recently from waterfowl toflamingos. As possible tools to help solvethe problem, the affinities of quill mites(syringophilids) and nasal mites (derman-yssids) from flamingos should be investigated.

118 Marine Maladies?
Some important protozoans, bacteria, andviruses of wild birds have been treated in
some detail by Davis, Anderson, Karstad,and Trainer (1971). Levine (1973) coveredmany more protozoans, includingSarcocystis rileyi.
Marine Mammals
Marine mammalian parasites have beendescribed in a book (Delyamure, 1955) andcompiled into a checklist (Dailey andBrownell, 1972) in another book that provides a wide range of diverse information onthese mammals (Ridgway. 1972). An annotated treatment of porpoise parasites(Zam, Caldwell, and Caldwell, 1971)supplements a review on this host's common diseases encountered by veterinarians(Sweeney and Ridgway, 1975). Lowery(1974) discussed marine mammals in thenorthern Gulf, Gunter and Overstreet (1974)reported parasites from a dwarf sperm whalein Mississippi Sound, and I have seen all thedolphin parasites discussed earlier in hostsfrom Mississippi. David Gibson and RodBray have designed a display of the largespecies of Crassicauda for the BritishMuseum (Natural History).
The possible role of campulids in singlestratidings was suggested by Ridgway andDailey (1972). Perhaps multiple strandingscould result from a dominant leader beingaffected by parasites and the subordinatesfollowing that leader to beach themselvesregardless of the consequences. Otherparasites also have the potential to interferewith the "sonar" system in these animals.
MISCELLANEOUS SYMBIONTS
A review of the mermithids was presentedby Nickle (1972). Petersen (1973) is one ofseveral who have reported on the use of thesenematodes in controlling mosquito populations. Presently, one species, Reesimermisnielseni, has been especially fruitful forthis purpose. It feeds on the hemolymph ofand kills over 20 species of mosquitoes. Thelarva can successfully infect a largepercentageof a mosciuitopopulation. Sincethe worm does not feed after it leaves its
host-mosquito, it can be handled easily, andthe preparasitic juvenile can be reared andcollected in large numbers. Preliminary-field applications of 1000 larvae per squaremeter killed most of the mosquitoes, and theworm became permanently established insome of the sites (Petersen and Willis, 1972).
The illustrated midge with the mermithidcame from Simmon's Bayou near OceanSprings when the salinity was 6 ppt (14April 1977)and the associated animals consisted primarily of estuarine species.
A classic work on nematomorphs by May(1919) described the life histories of twosjx»cies, and both Pennak (1953) and Cheng(1973) provided a general treatment of thegroup. Born (1967) reported Nectonemaagile from the grass shrimp.
The leech Stibarobdella macrothela was
recently treated by Sawyer, Lawler, andOverstreet (1975) and by Penner and Raj(1977). A few turbellarians were discussedas symbionts of oysters; Jennings (1974)discussed many more and showed progressive stages from totally free-livingto parasitic.
Barnacles from bands on the dusky sharkwere collected by Tom Mattis while insearch of elusive tapeworms. William A.Newman of Scripps Institution of Oceanography identified Conchoderma auritumand Cindy van Duyne of North CentralCollege in Naperville, Illinois, identifiedthe other two. According to Newman (personal communication), Conchodermaauritum usually attaches to sessilebarnacles on whales, but sometimes occurson baleen or on the teeth of toothedcetaceans when their lips are damaged.Newman got one off a submarine. Dawson(1969) previously reported the only recordofConchoderma virgatum from the Gulf ofMexico and listed cases in other parts ofthe world where the organism attacheddirectly to fishes and to parasitic copepods.Unexpectedly, all hosts are not those thatbask or remain in surface water where onemight consider the barnacle's larvae mostable to infest hosts.
Biting gnats (Culicoides spp.) that occurin the northern Gulf have been studied fromseveral localities, and their activities differsome from place to place (Khalaf, 1967.

*r
1969; Kline and Axtell. 1976). Because therewas a dense population of Culicoideshollensis last fall (1977) and because theweather has been so nice during the earlyweeks of this spring, the present populationof gnats has me satisfied being insidewriting this booklet rather than beingoutside.
According to Robert C. Lowrie, Jr.(personal communication. Delta PrimateCenter), our populations of gnats have notbeen implicated in transmitting viruses norhosting any local filarid nematodes. Lowriehas examined many individuals and neverseen natural filarial infections; however, hefound local gnats toexperimentally supportsome exotic filaria.
FISH-KILLS AND
MISCELLANEOUS DISEASES
Often a fish-kill results from a
combination of factors. Comments on the
physiology of fish related to a cold-kill weretreated by Overstreet (1974). Guntcr (1941)and Moore (1976) are two of several whohave reported on fishes killed by the cold inTexas where such mortalities occur more
frequently than in Mississippi. May (1973)reported on the occurrence of jubilees inMobile Bay and how the first account was in1867 even though he searched documentsdating back to 1821 which should havementioned early periodic fish-kills if theyhad been present. According to May, thereason for the increasing frequency ofjubilees appears to be the decreasing abilityfor the water of Mobile Bay to transport,deposit, dilute, and assimilate the amount oforganic matter in the bottom water. Most ofthe matter comes from natural sources of
wood and leaves, but is supplemented by theincreased amount of domestic wastes.
The jubilees in Mississippi, according toCharles Lyles (personal communication,Gulf States Marine Fisheries Commission),and possibly those in Alabama, too. resultfrom plankton blooms. Lyles has observedmany jubilees along Bellefontaine Beachsince the late 1930'sand found several items
common to all. They occur during neap
Technical Aspects 119
tides (tides with smallest differences betweenhigh and low tide occurring after the firstand last quarters of the moon) at nightbetween late June and early September. Usually rain precedes them, and the well-defined affected water has a tea color. Apparently the proper nutrients allow succession of a phytoplankter that mightproduce a similar toxin to that of dinoflagellate blooms of a red tide or a conditionthat otherwise creates an unhealthy watermass. These jubilees certainly demand critical investigation!
Numerous papers deal with red tide. Notuntil recently, however, has anyonecritically examined dead and dying animals.Quick and Henderson (1974, 1975) reportsuch findings, and Forrester and others(1977) go into more detail and speculationconcerning an epizootic affecting lesserscaup.
Sources of pollution are being examinedmore extensively in recent years Ixxause ofan increased concern for the environment.Overstreet and Howse (1977) briefly-reviewed pesticides, heavy metals, and othersources of pollution and possible stress-related diseases in Mississippi estuaries.Pollution-sources from other regions havealso been documented (e.g., Alabama byCrance, 1971). More extensive studies havedealt with potential and actual problems inChesapeake Bay (McErlean, Kerby, andWass, 1972). Sindermann (personalcommunication) is preparing a review ofvarious diseases associated with aquaticpollution.
Deaths and disease in the natural marineenvironment are hard to document and thecauses even harder to determine. Usually acombination of factors act together. One ofthose factors is often temperature. Somefish, especially pelagic ones, swim fasterwhen the temperature is either raised ordecreased under experimental conditions(Olla, Studholme, Bejda, Samel, andMartin, 1975). Increases in localized watertemperatures are rapidly becoming normalbiproducts of modern technology (e.g..Cairns, 1972). Primarily because of powerplants, critical research has been conductedon the effects of fish by increased temperature combined with chlorine (Meldrin,

120 Marine Maladies?
Gilt, and Petrosky. 1974) and with otherchemicals (Cairns, Health, and Parker,1975).
In addition to natural mortalities caused
by a rapid drop in temperature, mortalitiescan also result from sudden rainfall and
storms. Dawson (1965) documented deathsof the lancelet (Branchiostoma caribaeum)in Mississippi following heavy rains with asubsequent drop in salinity. Most likely-other animals also die under sue h adverse
conditions. Storms can kill fish, especiallytransient species not normally occupyingshallow inshore regions (Robins. 1957).Thefishes accumulate sediment in their gillchambers, and their gills fray. Regularinhabitants of shallow, wind-exposedregions appear capable of withstandingconsiderable turbulence.
Human activities have now stressed the
environment to a greater degree thancorresponding means have been designed toprotect that environment. Our wastes andother activities are taking their toll on theenvironment. Unfortunately, only whenfish die in large numbers do people worryabout the problems. Would many people beconcerned about dumping sewage sludge inthe New York Bight had not there- been anextensive kill (Bullock. 1976)? Dredging,bulkheading, and filling in natural marshesseldom cause extensive fish-kills, but thoseactivities nevertheless decrease numbers of
animals (e.g.. Trent, Pullen, Proctor. 1976).Even flushing detergents into our waters hasserious effects (Abel, 1974), but these effectsseldom meet public view. Man needssounder management programs, andperhaps these should be oriented towardprotecting important ecosystems ratherthan protecting specific species. Cairns(1975) commented on this strategy and onthe fact that our actions do not match our
public statements. In order for substantialchanges to occur, the life styles of peoplemust also change.
As pointed out several times, the mostimportant factor in transforming aninfection into a disease is stress. A readable
book by Wedemeyer, Meyer, and Smith(1976) on this problem provides a valuablebackground in regard to fishes.
A recent compilation of contributions on
genetics and mutagenesis of fishes goes intoconsiderable detail on many aspects ofgenetics (Schroder, 1973). Investigationsinto chromosomal anomalies in marine
fishes have been recent and very fruitful(Longwell, 1976). Apparently LongwelTsmethods of detecting anomalies allowevaluation of material from museum
collections and thereby a means ofcomparing collections from past years whenthe water was relatively clean with thosecollected recently from the same regionsafter water-quality had deteriorated. She(personal communication) has beencompiling much more extensive data thatcorroborate her earlier findings.
Nutrition of fish has been covered in
detail in a book edited by Halver (1972),andthat book gave a number of examples andreferences which should guide any studentinterested in different aspects of nutrition.Examples of scoliosis (spinal columntwisted in a sideways direction) such as thatshown for the striped mullet (Mugilcephalus) as well as cases of lordosis (spinalcurvature tending toward a verticaldirection) are presented. Diets deficient intryptophan or ascorbic acid produce bothconditions experimentally as does a lack ofcalcium phosphate or an infection with afew different bacteria and parasites. Anotherbook (Neuhaus and Halver, 1969) alsocovered nutrition, metabolism, genetics,and cancer. Representatives of most tumorsfrom fishes that have been studied scientifically have been deposited in the "Registry ofTumors in Lower Animals" at the National
Museum of Natural History. JohnHarshbarger, who directs that registry andidentifies most of the neoplasms, haspublished extensively on the availablematerial (e.g., 1977).That paper occurs withseveral others in a volume on "AquaticPollutants and Biological Effects withEmphasis on Neoplasia" (Kraybill, Dawe,Harshbarger, and Tardiff, 1977). Edwardsand Overstreet (1976) reported on theneoplasms in the striped mullet, andOverstreet and Edwards (1976) also reportedbenign fibromas in the southern flounderand sea catfish and reviewed the obser
vations of other tumors from the northern
Gulf.

That many tumors and other diseases arecaused by environmental contaminants isbased almost entirely on circumstantialevidence. Aquatic examples have beendocumented primarily from freshwaterhabitats because such habitats are oftenexcessively polluted and because specimensare easier to collect from these confinedregions than from open marine andestuarine ones. Edwards and Overstreet
(1976) suspected the tumor on mullet mightresult from pollutants. Mearns andSherwood (1976) also considered thispossibility for skin tumors in a southernCalifornia flatfish, but did not thinkimprovements in municipal waste treatments would reduce the prevalence oftumors unless the improvements increasedthe number of young fish. They (Sherwoodand Mearns. 1977), however, presentedstrong observational and experimentalevidence linking chlorinated hydrocarbonpollutants (e.g., DDT) with fin-erosion inthe same flatfish exposed to the dischargedmunicipal waste water.
Many examples of injuries that causeanomalies have been indexed in the
bibliographies by Dawson(l964; I966; I97I)and Dawson and Heal (1976). Bird (1978)reported straps encircling carcharhinidsharks, including a postscript on theaffected dusky sharks mentioned in thepresent treatment of barnacle symbionts.The Atlantic croaker with its rubber band
was reported by Oversteet and Lyles (1974).Even though a few fishermen maydeliberately place a band about a fish as ajoke, most fish ring themselves naturally.Lamont (1961) pointed out the abundanceof mackerel ringed with contraceptivesoccurring near sewage effluents.
Dawson's bibliographies cited referencesto all types of anomalies, including those offlatfish. According to the literature, reversalin Eutropus crossotus is rare. Taylor,Stickney, and Heard (1973) reported the firstcase from Georgia, and James Ray Warrenin the Fisheries Section at the Gulf Coast
Research Laboratory, who found thespecimen photographed above in Back Bayof Biloxi has found only one other exampleof a reversed fringed flounder, and itoccurred in a pass off Horn Island. Dawson
Acknowledgments 121
(1962) and Moore and Posey (1974) describedand reviewed many abnormalities in thehogchoker. Norman (1934) and Gudger(1934) laid the foundation for discussinganomalies in flatfishes, and that foundation
continues to be built upon today.
Est ut omne hoc vobis stercus sit,
sed mihi panis et butyrum est.
ACKNOWLEDGMENTS
I am grateful lo the many people who helped makethis guide possible, and the addresses of those not fromthe Gulf Coast Research Laboratory are indicated.Several people read different portions of the text andmade comments. C.E. Dawson. A.R. Lawler. andR.W. Heaid (Dauphin Island Sea I.all) read the entiretext; J.A. Quirk. Jr. (Dow Chemical Company.Freeporl. Texas). P.T. Johnson (National MarineFisheries Service. Oxford. Maryland), and J.A. Couch(Environmental Protection Agency. Gulf Bree/c,Florida) read the portions on invertebrates; and othersread specific- pans: F.M. Bayer (Smithsonian Institution) (zoanthids); H.M. Perry (blue crab): I'M. VanDevcnder (penaeid shrimps); W.J. Demoran. E.W.Cake. Jr.. J.T. Ogle. A.P. Chestnut, and K.A. McGraw(oyster); T.L. Deardorff and T.E. Mattis (fishes): andD.B. Pence (Texas Tech University, Lubbock. Texas)(birds). Other assistance was provided by R.G. Palmer.T.A. Miller. R.E. Buxton. T.L. Deardorff. T.E.
Mattis. D.R. Brooks, A.C. Fusco. M.A. Solangi.L.A. Toomey, S.L. Shepard. I..B. Ross, and V'.D. Jones(Mississippi-Alabama Sea Grant Consortium).
CA. Foster and R.E. Buxton pi (pared most of theblack and white prints. Negatives or other prints wereprovided by J.A. Couch (Figs. 36. 46. 47. 19. and 50).P.T. Johnson (Figs. 44 and 45). A.P. Chestnut (Figs. 15and 16). CA. Foster (Fig. 63). M.A. Solangi (Figs. 13.54 - 57. and 72). and T.E. Mattis (Figs. 186 - 190).B.C. Heard (Dauphin Island. Alabama) drew Figs. 23.28, 70. 78. 82. 86. 113, 149, 157. 161. and 164 andD.L. Ilolcomb drew Figs. 5 and 48.
Permission lo reproduce the same or similar figuresfrom previously published works was granted byAcademic Press — Figs. 56 and 57 from Solangi andLightner (1976) and Overstreet and Van Devender(1978); The American Microscopical Society — Fig.145 from Overstreet (1971a); American Society ofIchthyologists and Herpetologisls — Figs. 91 and 93from Lawler. Howse, and Cook (1974); AmericanSociety of Parasitologists — Fig. 109 from Overstreet(1976a). Fig. 117 from Overstreet (1977b). Figs. 141 and142 from Overstreet (1976b), Fig. 143 from Overstreet(1970). and Fig. 144 from Overstreet (1971 b);fluc/W«>io/Marine Science — Fig. 196 from Edwards andOverstreet (1976); Cambridge University Press — Figs.129 and 193 from Overstreet and Paperna (in press);Gulf Research Reports — Figs. 194 and 195 from

122 Marine Maladies?
Overstreet (I974) and Figs. I98 from Overstreet andLyles (I974); The Hclmitithological Society ofWashington — Fig. 162 from Buckner. Overstreet. andHeard (1978); The New York Academy of Sciences —Figs. 88. 89. 103. and 105 from Overstreet and Howse(1977); Springer-Verlag — Fig. 80 from Overstreet andWeidner (1974); and The Wildlife Disease Association— Figs. 25. 51. and 52 from Cook and Lofton (1973).
A number of people provided specimens for me tophotograph: A.P. Chestnut (Figs. 12 and 14), D.W.Cook (Figs. 25. 51. and 52). T.M. Van Devcnder (Figs.75-77). CD. Getter (Rosenstiel School of Marine and
Atmospheric Science. Miami, Florida) (Figs. 150 and131). R.W. Heard (Figs. 68 and 178), A.R. Lawler(Figs.90-93. 96-99. 137. and 165-168).M.J. Loman(Fig. 171).Marine Life. Inc. (Gulfport. Mississippi) (Figs. 130and197). T.E. Mattis (Figs. 71. 114-116. and 121-128). K.A.McGraw (Figs. 6-11. 17-21). K.R. Melvin (Figs. 33. 35,and 112). J.T. Ogle (Fig. I). 1I.M. Perry (Figs. 30, 31.and 39). J.R. Warren (Figs. 32. 34, 39. and 192). andsome others by summer students.
LITERATURE CITED
ABBOTT. R.T.. 1974. American Seashells. The Marine
Mollusca of the Atlantic and Pacific Coasts of NorthAmerica. 663 pp. New York: Van NostrandReinhold.
ABEL. P.D., 1974. Toxicity of synthetic detergents tofish and aquatic invertebrates, journal of fishBiology 6(3):279-298.
ADKINS. C 1972. A study of the blue crab fishery inLouisiana. Louisiana Wild Life and Fisheries Commission Technical Bulletin No. 3:1-57.
ALDRICH. D.V., 1965. Observations on the ecologyand life cycle of Prochristianella penaei Kruse(Cestoda: Trypanorhyncha). The Journal of Parasitology 51(3): 370-376.
ALLEN. J.F., 1958. Feeding habits of two species ofOdostomia. Na ulilus 72( I): 11-15.
ANDERSON. C, 1975a. Larval metabolism of the
epicaridian isopod parasite Probopyrus pandallcolaand metabolic effects of P. pandalicola on its copepod intermediate host Acartia tonsa. ComparativeBiochemistry and Physiology 50(4A):747-751.
ANDERSON. C... 1975b. Metabolic response of thecaridean shrimp Palaemonetes pugio to infection byche adult epibranchial isopod parasite Probopyruspandalicola. Comparative Biochemistry and P/»y-«ofogy52(lA):201-207.
ANDERSON. R.C, A.G. CHABAUD, AND S.WILLMO'IT (eds.), 1974a. CIH Keys to the Nema-lode Parasites of Vertebrates. No. 1. GeneralIntroduction, Glossary of Terms, by S. Willmolt,Keys to Subclasses, Orders and Superfamilies, byA.G. Chahaud. 17 pp. Farnham Royal: Commonwealth Agricultural Bureaux.
ANDERSON. R.C., A.G. CHABAUD. AND S.
WILLMOTT (eds.). 1974b. CIH Keys to the Nematode Parasites of Vertebrates. No. 2. Keys to Genera
of the Ascaridoidea, by G. Hartwich. 15 pp.Farnham Royal: Commonwealth AgricultiiialBureaux.
ANDERSON. R.C. A.G. CHABAUD. AND S.WILLMOTT (eels.). 1975a. CIH Keys to the Nematode Parasites of Vertebrates. No. 3. Keys to Generaof the Order Spirurida. Part I. Camallanoidra,liracunculoidea. Gnathostomatoidea, Physalopter-oidea, Rictularioidea and Thelazioidea, by A.G.Chabaud. 27 pp. Farnham Royal: CommonwealthAgricultural Bureaux.
ANDERSON, R.C, A.G. CHABAUD. AND S.
WILLMOTT (eds.). 1975b. CIH Keys to the Nematode Parasitesof Vertebrates. No. 1.Keys to Generaofthe Order Spirurida. Part 2. Spiruroidea, Habrone-matoidea and Acuartoidea, by A.G. Chabaud. 58pp. Farnham Royal: Commonwealth AgrkuliuialBureaux.
ANDREWS, J.D. AND W.G. HEWATT. 1957. Oystermortality studies in Virginia. II. The fungus diseasecaused by Dermocystidium marinum in oystcis ofChesapeake Bay. Ecological Monographs 27(1):1-25.
ANDREWS, J.D., J.L. WOOD. AND H.D. HOESE,1962. Oyster mortality studies in Virginia: 111.Epizootiology of a disease caused by Haplospor-idium costaleWood and Andrews. Journal of InsectPathology 4(3): 327-343.
BAER, J.G., 1957. Repartition el cndemieitc des cestodes chez les reptiles, oiseaux et mammifii'-s. InPremier Symposium sur la Specificite Parasitaue desParasites de Verlebres. International Union ofBiological Sciences Series B 32:270-292.
BAILEY, R.M. (ch.). J.E. FITCH, E.S. HERALD. E.A.LACHNER. CC LINDSEY. C.R. ROBINS. ANDW.B. SCOTT. 1970. A List of Common and
Scientific Names of Fishes from the United Statesand Canada. 3rd cd. American Fisheries SocietySpecial Publication No. 6:1-150.
BAKKE.T.A.. 1972.Reighardia.vlcmae(Diesing. 1864)Ward, 1899 [Pentastomida; Cephalobaenida] fromthe common gull (Lams canus I..) in a Norwegianlocality. .Vorteegion Journal of Zoology 20(4):273-277.
BECKER. CD.. 1970. Haemato/oa of fishes, withemphasis on North American records. In A Symposium on Diseases of Fishes and Shellfishes (cd. S.F.Snieszko). American Fisheries Society SpecialPublication No. 5:82-100.
BECKER. CD.. 1977. Flagellate parasites of fish. InParasitic Protozoa, Vol. 1, pp. 357-416. New York:Academic Press.
BIGELOW. H.B. AND W.C SCHROEDER. 1948.Sharks. In Fishes of the Western North Atlantic.Memoirs. Sears Foundation for Marine Research I(Pi. l):59-576.
BIRD, A.F., 1971.The Structure of Nematodes. 318 pp.New York: Academic: Press.
BIRD, P.M., 1978. Tissue regeneration in threecurcharhinid sharks encircled by embedded straps.Copeia !978(2):345-349.

BLACKFORD, S.K., 1966. A Study of Certain Aspectsof the Ecology and Morphology of lirancliiobdellidAnnelids Epizoic on Callinectes sapidus. HonorsThesis. Newcomb College. New Orleans. Louisiana.34 pp.
BLAKE, J.A., 1971. Revision of the genus Polydora(torn the east coast of North America (Polychaela:Spionidae). Smithsonian Contributions toZoology No. 75:1-32.
BLAND, C.E., 1975. Fungal diseases of marineCrustacea. Proceedings of the Third U.S.-JapanMeeting on Aquaculture at Tokyo. Japan,Oct. 15-16. 1974. pp. 41-48. Niigala: Fishery Agency.Japanese Government and Japan Sea RegionalFisheries Research laboratory.
BLAND. CE. AND H.V. AMERSON. 1973a. Electron
microscopy of zoosporogcnesis in Ilie marinephycomycete. Lagenidium callinectes Couch.Archiv fur Mikrobiologie 94:47-6-1.
BLAND. CE. AND H.V. AMERSON. 1973b. Obser
vations on Lagenidium callinectes: Isolation andsporangia! development. Mycologia e>5(2):310-320.
BLAND, C.E., D.C. RUCH, B.R. SALSER, ANDD.V. LIGHTNER, 1976. Chemical control of
Lagenidium, a fungal pathogen of marineCrustacea. 38 pp. UNC-SG-76-02. Raleigh: University of North Carolina Sea Gram Piogram. 1235
Burlington I.aboratories. North Cirolina SlateUniversity.
BORN, J.W., 1967. Palaemonetes vulgaris (Crustacea,Decapoda) as host for the juvenile stage ofNeclonema agile (Nematomorpha). The Journal ofParasitology 53(4):793-794.
BRADBURY. P.C. J.C. CLAMP, AND J.T. LYON.III. 1974. Terebrospira chattom sp. n.. a parasite olthe endocuticle of the shrimp Palaemonetes pugioHolthuis. The Journal of Protozoology 21(5):678-686.
BRADBURY. P.C. AND V. GOYAL. 1976. The fine
structure of a parasitic ciliate Terebrospira duringingestion of the exoskeleton of a shrimpPalaemonetes. Tissue and Cell 8(4):573-582.
BRIDGMAN. J.F., 1969. Life cycles of Carneophalluschoanophallus n. sp. and C. basodactylophallusn. sp. (Trematoda: Microphallidac). Tutane Studiesin Zoology and Botany 15(3):8I-I05.
BROOKS. D.R. AND R.M. OVERSTREET. 1977.
Acanthostome digeneans from the Americanalligator in the southeastern United Stales.Proceedings of the Biological Society of Washington90(4):1016-1029.
BROOKS, D.R. AND R.M. OVERSTREET, 1978. The
family Liolopidae (Digenea) including a new genusand ewo new species from crocodilians. InternationalJournal for Parasitology (in press).
BROOKS. DR., R.M. OVERSTREET. AND D.B.
PENCE, 1977. New records of proterodiplosiomedigeneans from Alligator mississippiensis andCaiman crocodilus fuscus. Proceedings of theHelminthological Society of Washington 44<2):237-238.
Literature Cited 123
BUCHANAN. R.E. AND N.E. GIBBONS (eels). 1974.Sergey's Manual of Determinative Bacteriology.8th ed. 1268 pp. Baltimore: Williams and Wilkin*.
BUCKNER. R.L.. R.M. OVERSTREET. AND R.W.
HEARD. 1978. Intermediate hosis for Tegorhynchusfurcatus and Dollfusentis chandleri (Acantho-cephala). Proceedings of the HelminthologicalSociety of Washington (in press).
BULLA, L.A., JR. AND T.C CHENG (ids.). 1976.Comparative Pathobiology. Vol. I. Biology of theMicrosporidia. 371 pp. New York: Plenum Press.
BULLA. L.A.. JR. AND T.C. CHENG (eds). 1977.Comparatn-e Pathobiology. Vol. 2. Systemalics ofthe Microsporidia. 510 pp. New York: Plenum Press.
BULLOCH. D.K.. 1976. Ocean kill in the New York
Bight - summer 1976. Underwater Naturalist10(1):!-12.
BULLOCK. G.I... 1971. Identification of FishPathogenic Bacteria, Book 2B. In Diseases of Fishes(eds. S.F. Snieszko and H.R. Axehoel), pp. Ml.Jersey City: T.F.H. Publications.
BULLOCK. G.I... DA. CONROY. AND S.F.
SNIESZKO. 1971. Bacterial Diseases of Fishes,Book 2A. In Diseases of Fishes (eds. S.F. Snic-s/ko andH.R. Axelrod). pp. 1-151. Jersey City: T.F.H. Publications.
BULLOCK. W.l... 1957. The acanihocephalanjKirasilesof the fishes of the Texas coast. Publicationsof the Institute of Marine Science l(2):278-283.
BULLOCK, W.L., 1966. Entamoeba gadi sp.n. fromthe rectum of lite pollock. Pollachius virens (I...1758). with some observations on its cytochemistry.77te* Journal of Parasitology 52<4):679-684.
BULLOCK. W.l... 1969. Morphological features astools and and as pitfalls in acanthocephalansystemalics. In Problems in Systemalics of Parasites(ed. CD. Schmidt), pp. 9-45. Baltimore: UniveisityPark Press.
BURNETT-HERKES. J.. 1974. Parasites of the gillsand buccal cavity of the dolphin. Coryphaenahippurus, from the Straits of Florida. Transactionsof the American Fisheries Society 103(1):101-106.
CABLE, R.M., 1974. Phylogeny and taxonomy ofnematodes with reference to marine- species. InSymbiosis in the Sea (ed. W.B. Vernbcrg).pp. 173-193.Columbia: University ol South CarolinaPress.
CAIRNS. J.. JR.. 1972. Coping with heated waste waterdischarges from steam-electtic power plants.flKj.«i>nce22(7):4l)-l20.
CAIRNS, J., JR.. 1975.Critical species, including man.within the biosphere. Die Naturwissenschaften62:193-199.
CAIRNS. J., JR.. A.G. HEATH. AND B.C. PARKER.1975. The effects ol temperature upon the toxicity ofchemicals lo aquatic organisms. Hydrobiologia47(1): 135-171.
CAKE. E.W.. JR.. 1976. A key to larval cestodes ofshallow-water, benthic mollusks of the northern

124 Marine Maladies?
Gulf of Mexico. Proceedings of the Helminthological Society of Washington 43(2): 160-171.
CAMPBELL. R.A. AND J. CARVAJAL. 1975. Arevision of some trypanorhynchs from the westernNorth Atlantic described by Edwin Linton. 77ie*Journal of Parasitology 61(6>:)016-1022.
CARRIKER. M.R.. E.H. SMITH. AND R.T. WILCE
(eds.). 1969. Penetralion of calcium carbonatesubstrates by lower plants and invc-itc-biates.American Zoologist 9(3. ed. 2):629-1020.
CHENG. T.C. 1967. Marine molluscs as hosts for
symbioses wilh a review of known iKirasitt-s ofcentimere-tally important species. Advances in MarineBiology 5:1-424.
CHENG.. T.C. 1973. General Parasitology. 965 pp.New Yolk: Academic Press.
CHITWOOD. B.G. AND MB. CHITWOOD. 1950.
Introduction lo Sematology. 334 pp. Baltimoie:University Patk Press.
CHRISTENSEN. A.M. AND J.J. Mc DERMO IT.1958. Life-bistcuy and biology of the oystc-i crab.Pinnotheres ostreum Say. Biological BulletinI14(2):I46-I79.
CHRISTMAS. J.Y.. 1969. Parasitic barnacles inMississippi estuaries wilh special iclcie-nee- loLoxolhylaius texanus Boschma in the blue crab(Callinectes sapidus). Proceedings of the Twenty-Second Annual Conference. Southeastern Association of Game and Fish Commissioners, pp. 272-275.
CHRISTMAS. J.Y. AND D.J. ETZOLD (eds.). 1977.The shrimp fishery of the Gull of Mexieo United
Stales: A regional management plan. Gulf CoastResearch Laboratory Technical Report SeriesNo. 2:1-128.
CLAMP. J.C, 1973. Observations on the hosi-symhi-ont relationships of Lagenophrys lunatus ItnamiiiaThe Journal of Protozoology 20(5):588-561.
COKER. R.E.. 1902. Neites on a species ol barnacleIDichelaspisI parasitic on the gills of edible crabs.Bulletin of The United Slates Fish Commision (for1901) 21:399-412.
COLWELL, R.R.. T.C WICKS. AND U.S.
TUBIASH. 1975. A comparative study of thebacterial flora ol the hemolymph of Callinectessapidus. Marine Fisheries Review 37(5-6):29-33.
COMPAGNO. L.J.V.. 1973. Interrelationships ofliving elasmobranchs. In Interrelationships of Fishes(eds. P.H. Greenwood. R.S. Miles, and C. Patterson).
Zoological Journal of the Ltnnean Society 53(Suppl. 1):I5-61.
COMPAGNO. L.J.V.. 1977. Phyletic relationships ofliving sharks and rays. American Zoologist 17(2):303-322.
COOK. ILL. AND M.J. LINDNER. 1970. Synopsis ofbiological data on the brown shrimp Penaeus azlecusaziecus Ives, 1891. FAO Fisheries Report 57(4):1471-1497.
COOK. D.W. AND S.R. LOFTON. 1973. Chiiinoclas-
cic bacteria associated with shell disease in Penaeus
shrimp and the blue crab (Callinectes sapidus).Journal of Wildlife Diseases 9(2):154-I59.
COOK. D.W. AND S.R. LOFTON. 1975. Patho
genicity studies with a Streptococcus sp. isolatedfrom fishes in an Alabama-Florida fish kill.
Transactions of the American Fisheries SocietyIO-l(2):286-288.
COOLEY. R.A. AND G.M. KOHLS. 1945. The genusIxodes in Noith America. National Institute of
Health Bulletin. U.S. Public Health Sen-lie 181:
1-216.
CORKERN. CC. JR.. 1970. Investigations onHelminths from the Hepatopancreas of the BrownShrimp, Penaeus aztccus Ives, from Galveston Hay,Texas. M.S. thesis. Texas A&M University, College-Station. 51 pp.
CORKERN, CC. II. 1978. A larval eye lophyllidean(Cestotla) parasiteol blown and whin- shiimp. 2 pp.[•"DDLS-10. College Station: Fish Disease Diagnosticlaboratory, Texas Agricultural Extension Service.Depart men Iof Wildlife and Fisheries Sc nines. TexasA&M University System.
CORLISS. JO.. 197 la. The changing world of ciliatesystematic s: historical analysis of past efforts anil anewly pioposed phylogenelic scheme- of classification for the pioiistan phylum Ciliophora.Systemalu Zoology 23(1):91-138.
CORLISS. J.O.. 1974b. Remarks on theconi|iositiou ofthe huge ciliate class Kinelofiagmophoia dePuyiorac et al.. 1974. and recognitor! of several newtaxa therein, wilh emphasis on the ptimiiivc orderI'riiiKHiliaticla n. ord. The Journal of Protozoology2l(2):207-220.
COUCI I, J.A.. 1966.'Two peritrichousciliates fiom thegills o( the blue crab. Chesapeake Science 7(3):171-173.
COUCH. J.A.. 1967. A new species of Lagenophrys(Ciliatea: Peritiiihida: Lagenophryidae) from amarinccrah, Callinectes sapidus. Transactions of theAmerican Microscopical Society 86(2):204-2l 1.
COUCH. J.A.. 1973. Uitrastructural and piotait;(>lstudies of Lagenophrys callinectes (Ciliophora:Peritrichida). The Journal of Protozoologx 20(51:638-647.
COUCH. J.A.. 1974a. Pathological effects ofUrosporidittm (Haplosporida) iuleclion inmicrophaiiid melacereariae. Journal of InvertebratePathology 23(3):389-396.
COUCI I. J.A.. 1974b. Free and occluded vims, similailo Baculovirus. in hepatopancreas of pink shiimp.Nature 247(54381:229-231.
COUCH. JA.. 1974c.An enzootic nuclear polyhedrosisvirus of pink shrimp: ulirastructure. prevalence, andenhancement. Journal of Invertebrate Pathology24(3 ):311-331.
COUCH. J.A.. 1977. Uitrastructural study of lesions ingills of a marine shiimp exposed to cadmium.Journal of Invertebrate Pathology 29(3):267-288.
COUCH. J.A.. 1978. Diseases, paiasites. and toxicresponses of commercial penaeid shrimps of the Gull

«•*•
of Mexico and south Atlantic coasts of Noith
America. Fishery Bulletin 76(l):l-44.
COUCH. J.A. AND I.. COURTNEY. 1977. Interactionof chemical pollutants and virus in a crustacean:a novel bioassay system. In Aquatic Pollutants andBiologic Effects with Emphasis on Neoplasia (eds.H.F. Kraybill. C.J. Dawe, J.C. Harshbarger. andR.C Tardiff). Annals of The New York Academy ofSciences 298:497-501.
COUCH. J.A., CA. FARLEY. AND A. ROSENFIELD. 1966. Speculation of Minchinia nelsoni(Haplosporida, Haplosporidiidae) in Crassostreavirginica (Gmelin). Science 153(3743):1529-I53I.
COUCH. J.A. AND A. ROSENFIELD. 1968. Epizex.ti-ology of Minchinia costalis and Minchinia nelsoniin oysters intrcxluced inio Cliiiicoleague Bay,Virginia. Proceedings of the National ShellfishenesAssociation 58:51 -59.
COURTNEY. C.H. AND D.J. FORRESTER. 1971.Helminth parasites of the brown pelican in Floridaand Louisiana. Proceedings of the HelminthologicalSociety of Washington 41(l):89-93.
CRANCE, J.H.. 1971. Description of Alabamaestuarine areas — coopcralive Gull of Mexicoestuarine inventory. Alabama Marine ResourcesBulletin 6:1-85.
CRESSEY. R.. 1967. Revision of the family Pandaridae(Copcpoda: Caligoida). Proceedings of the UnitedStates National Museum 12l(3570):l-133.
CRESSEY. R.F.. 1972. The genus Argulus (Crustacea:Branchiura) of the United Stales. Biota of Freshwater Ecosystems, Identification Manual. Environmental Protection Agency No. 2:1-14.
CRESSEY. R.F. AND B.B. COLLETTE. 1970.
Copepods and needlefishes: a study in host-parasiterelationships. Fishery Bulletin 68(3):317-432.
CRESSEY. R.F. AND E.A. LACHNER. 1970. The
parasitic copepod diet and life hisloiy of diskfishes(Echeneidac). Copeia 1970(2):310-318.
DAILEY. M.D. AND R.L. BROWNELL. JR.. 1972. Achecklist of marine mammal parasiu-s. In Mam ma Isof the Sea. Biology and Medicine(ed. S.H. Ridgway),pp. 528-589. Springfield: Charles C. Thomas.
DAILEY. M.D. AND R.M. OVERSTREET. 1973.
Cathetocepkalus thatcheri gen. ei sp. n. (Tcira-phyllidea: Cathctocephalidae fam. n.) from the bullshark: a s|>ecies demonstrating multistrobilizaiion.The Journal of Parasitology 59(3):469-473.
DALES. R.P., 1957. Commensalism. Interrelations of
organisms. In Treatise on Marine Ecology andPaleoecology (ed. J.W. Hcdg|x-th). The GeologicalSociety of America Memoir 67(1):391-412.
DANIELS. B.A. AND R.T. SAWYER. 1975. The
biology of the leech Myzobdella lugubris infestingblue crabs and catfish. Biological Bulletin 148(2):193-198.
DARNELL. R.M.. 1959. Studies of the life history ofthe blue crab (Callinectes sapidus Rathbun) inLouisiana waters. Transactions of the AmericanFisheries Society 88(0:294-304.
Literature Cited 125
D'ASARO. C.N.. 1966. The egg capsules, cmbiyo-genesis of a common oyster ptedaior. Thais haemas-loma florulana (Gastropoda: Prosobianc Ilia).Bulletin of Marine Science 16(0:884-9 II.
DAUG1IKRTY. S.J.. 1969. Aspet Isof the E, ologw LifeHistory, and Host-Parasite Relalion\hip olLoxothylacus panopaei tSaicul'nidaei m Chesapeake Bay. M.A. thesis. College of William and Man.Williamsburg. Virginia. 69 pp.
DAVENPORT. D.. G. CAMOUC1S. AND J.F.HICKOK. I960. Analyses ol the btliavimu olcommensals in host-lac tor. 1. A besioned polv-chaete and a piiinolheritl ctab. Animal Behaviour8(3-0:209-218.
DAVIES. A.J. AND M.R.L. JOHNSTON. 1976. Thebiology of Haemogregarina bigrmina Laverati feMesnil. a parasite of the marine fish Blenntus pholisLinnaeus. The Journal of Protozoology 23(2):
315-320.
DAVIS. CC. 1965. A study ol the hate hing process inaquatic invertebiales: XX. The blue erah. Callinei lessapidus. Ralhbun. XXL The nemeiieaii. Carttno-nemerles carcinophila (Kollikcr). ChesapeakeScience 6(0:201-208.
DAVIS. J.W., R.C ANDERSON. L. KARSTAD. ANDD.O. TRAINER. 1971. Infectious and ParasitiiDiseases of Wild Birds. 3-14 pp. Ames: Iowa StaleUniversity Press.
DAWSON. CE.. 1957. Balanus fouling of shrimp.Science 126(3282): 1068.
DAWSON. C.E.. 1962. Notes on anomalous Ameiic an
Helerosomata with descriptions of five new records.Copeia 1962(1):138-146.
DAWSON. C.E.. 1964. A bibliography of anomalies offishes. Gulf Research Reports 1(61:308-399.
DAWSON. CE., 1965. Rainstorm induced mortality oflancelets. Branchiostoma. in Mississippi Sound.Copeia I965(4):505-506.
DAWSON. C.E.. 1966. A bibliography ol anomalies offishes. Supplement 1. Gulf Research Reports 2(2):169-176.
DAWSON. C.E.. 1969. Records of the barnacle Concho-derma virgatum (rom twoGulfof Mexico fishes. TheProceedings of the Louisiana Academy of Sciences.32:58-62.
DAWSON. C.E.. 1971. A bibliography of anomalies offishes. Supplement 2. Gulf Research Reports 3(2):215-239.
DAWSON. C.E. AND E. HEAL. 1976. A bibliographyof anomalies of fishes. Supplement 3. Gulf ResearchReports o(2):35-il.
DEBLOCK. S.. 1971. Contributions a Telude des
MicrophallidaeTravassos. 1920 XXIV. Tentative dephylogciiic el de taxonomie. Bulletin du MuseumNational d'Histoire Naturelle 3e serie, no. 7,
Zoologie 7:353-469.
DEFENBAUGH, R.E. AND S.H. HOPKINS. 1973.
The Occurrence and Distribution of the Hydroids ofthe Galveston Bay. Texas, Area. TAMU-SG-73-210.

126 Marine Maladies?
202 pp. College Station: Department of Marine Resources Information. Center for Marine Resources.
Texas AfeM University.
DELYAMURE. S.L.. 1955. Helminthofauna of MarineMammals (Ecology and Phylogeny) (ed. K.I.Skrjabin. translated from Russian by M. Raveh,1968), 522 pp. Academy of Sciences of the U.S.S.R.,Helminthological Laboratory, Moscow. Jerusalem:Israel Program for Scientific 'Translations.
DESSER. S.S.. 1976. The ultrastructure of theepimasiigote stages of Trypanosoma rotalorium inihe leech Batracobdella picta. Canadian Journal ofZoology 54(I0):17I2-I723.
DESSER, S.S. AND I. WELLER. 1973. Structure,cytochemistry, and locomotion of Haemogregarinasp. from Rana berlandieri. The Journal of Protozoology 20(l):65-73.
De SYLVA, D.P., 1954. The live bait shrimp fisheryof the northeast coast of Florida. Florida State Boardof Consen'ation. Marine Laboratory. TechnicalSeries No. 11:1-35.
DtTURK. W.E., 1940. The Parasites and Commensalsof Some Crabs of Beaufort North Carolina. Ph.D.dissertation. Duke University. Durham. NorthCarolina. 105 pp.
DUBOIS, G.. 1968. Synopsis des Strigeidae et desDiplostomatidae (Trematoda). Memoires de laSocitle Neuchateloise des Sciences Naturelles 10
(Pt. 1):1-261.
DUBOIS. G.. 1970. Synopsis des Sirigeidae el desDiplostomatidae (Trematoda). Memoires de laSociete Neuchateloise des Sciences Naturelles 10
(Pt. 2): I-727.
DUKES. G.H.. JR.. R.M. SHEALEY. AND W.A.ROGERS. 1971. Sebekia oxycephala (Pentastomida)in large-mouth bass from Lake St. John, ConcordiaParish, Louisiana. The Journal of Parasitology57(5): 1028.
EARLL. R.E.. 1887. The mullet fishery. In TheFisheries and Fishery Industries of Ihe United Stales.Section V. History and Methods of the Fisheries. Vol.1. (ed. G.B. Goode). pp. 553-582. Washington:Government Printing Office.
EBERT, D.J.. 1976. The Behavior and PathologicalEffects of Larvae of Thynnascaris (Type MB)(Raphidascarinae) in the Mouse. M.S. thesis, University of Southern Mississippi. Hattiesburg. 47 pp.
EDWARDS, R.H. AND R.M. OVERSTREET, 1976.Mesenchymal tumors of some estuarine fishes of thenorthern Gulf of Mexico. I. Subcutaneous tumors,
probablyfibrosarcomas, in the stripedmullei, Mugilcephalus. Bulletin of MarineScience 26(l):S3-40.
EMERSON. K.C, 1972a. Checklist of the Mallophagaof North America (North of Mexico). Part I. SuborderIschnocera. 200 pp. Dugway, Utah: Deseret TestCenter, Dugway Proving Ground.
EMERSON. K.C. )972b. Checklist of the Mallophagaof North America (North of Mexico). Part II. Suborder Amblycera. 118 pp. Dugway, Utah: DeseretTest Center, Dugway Proving Ground.
EMERSON. K.C. 1972c. Checklist of the Mallophagaof North America (North of Mexico). Part IV. BirdHost List. 216 pp. Dugway, Utah: Deseret TestCenter. Dugway Proving Ground.
ERASMUS. D.A.. 1972. The Biology of Trematodes.312 pp. London: Edward Arnold.
ESCH. G.W..T.C HAZEN. R.V. D1MOCK.JR..ANDJ.W. GIBBONS. 1976. Thermal effluent and thecpizooiiology ol the ciliate Epistylis and thebacterium Aeromonas in assoe iation wilh central-
chiel fish. Transactions of the AmericanMicroscopical Society 95<4):687-ti93.
FARLEY. C.A., 1975. Epizootic and enzootic aspects olMinchinia nelsoni (Haplosporida) disease inMaryland oysters. VVic? Journal of Protozoology22(3): II8-427.
FEDUCCIA. A.. 1976. Osieological evidence lorshoti-birtl ailinilies ol ihe llamingos. The Auk 93(3):587-601.
FEIGENBAUM. D.l... 1975. Parasites of the commer
cial shrimp Penaeus vannamet Boone and Penaeusbrasiliensis Latreille. Bulletin of Marine Science25(0:491-514.
FEEDER. D.l... 1973. An Annotated Key to Crabs andLobsters IDecapocla. Reptantia)from Coastal Watersto the Northwestern Gulf of Mexico. LSI) -St; -73-02.103 pp. Baton Rouge: Center for Welland Resources.Louisiana Stale Univcisity.
FENG. S.Y.. 1958. Observations on distribution and
elimination ol spores of Nematopsis ostrearum inoysieis. Proceedings of the National ShellfishenesAssociation 48:162-173.
FONTAINE. C.T. AND D.V. LIGHTNER. 1975.
Cellular response- to injuiy in penaeid shrimp.Marine Fisheries Rei'iew 37(5-6):4-l0.
FORRESTER. D.J.. J.M. GASKIN. F.H. WHITE.N.P. THOMPSON. J.A. QUICK, JR.. G.E.HENDERSON. J.C WOODARD. AND W.D.ROBERTSON. 1977. An epizootic of waterfowlassociated with a red tide episode- in Florida. Journalof Wildlife Diseases I3(2):I60-I67.
FORRESTER. D.J. AND R.T. SAWYER. 1974. Pla-cobdella multilineata (Hiriidinea) from ihe
American alligator in Florida. The Journal ofParasitology 60(1):673.
FOSTER. CA.. E.G. SARPHIE. AND WE.
HAWKINS. In pi ess. Fine structure of the periiriclionsectocoiiimc'iisal Zoolhainniuin sp. with emphasis onits mode of altaihmenl to penaeid shiimp. Journalof Fish Diseases.
FUSCO. A.C. 1978a. The Life History and Development of Spirocamallanus cricotus. with Notes onSpirocamallanus and with an Annotated SpeciesListing. M.S. thesis. University ol .SouthernMississippi. Haitiesburg. 101 pp.
FUSCO. A.C. 1978b. Spirmamallanus critoim(Ni-iiialoda): Isoelectric' focusing and spc-ettophoici-meliie characterization of its hemoglobin and ihatolits piscine host. Micropogmiias undiilaliis. Experimental Parasitology 4-l(2):155-l60.

»<•
FUSCO. A.C. AND R.M. OVERSTREET. I978.
Spirocamallanus cricotus sp. n. and S. halitrophussp. n. (Nemaloda: t^amallanidea) from fishes in thenorthern Gulf of Mexico. The Journal of Parasitology 64(2):239-244.
GALTSOFF, P.S.. 1964. The American oyster
Crassostrea virginica Gmelin. Fishery Bulletin ofthe Fish and Wildiife Service 64:1-480.
GIBSON. D.I. AND R.A. BRAY. 1977. The Azygiidae.Hirudinellidae. Piythogonimidae, Sclerodislomidaeand Syncoeliidae (Digenea) of fishes from the northeast Atlantic. Bulletin of the British Museum(Natural History). Zoology Series 32(6): 167-245.
GILBERT. P.W.. R.F. MATHEWSON. AND D.P.
RAI.L (eds.), 1967. Sharks. Skates, and Rays. 624 pp.Baltimore: Johns Hopkins Press.
GLORIOSO. J.C. R.I.. AMBORSKI. J.M. LARKIN.G.F. AMBORSKI. AND DC CULI.EY. 1974.
Laboratory identification of bacterial pathogens ofaquatic animals. American Journal of VeterinaryResearch 35(3):447-150.
GOLVAN. Y.J.. 1969. Systcmatiquc des acantho-cephale's(/Ican(ftc«r/j/ia/a Rudolphi 1801). PremierePanic. L'ordre des Palaeacanlhocapliala Meyer 1931.Premier fascicule. I.a super-famille des Echinor-hynchoidea (Cobbold 1876) Cohan et Houin 1963.Memoires du Museum National d'Histoire Natur-
elle, Serie A 57(l'l. l):l-373.
GRELL. K.G.. 1973. Protozoology. 554 pp. Berlin:Springer-Verlag.
GUDGER. E.W.. 1934. Ambicoloraiion in the winter
flounder. Pseudopleuronectes americanus. American Museum Novitates 717:1-8.
GUNTER. C. 1941. Death of fishes due to cold on ihe
Texas coast, January. 1940. Ecology 22(2):203-208.
GUNTER. G. AND R. OVERSTREET, 1974. Ceta
cean notes. I. Sei and Rorqual whales on theMississippi coast, a correction. II. A dwarf spermwhale in Mississippi Sound and ils helminth parasites. Gulf Research Reports 4(3):479-481.
GUTHRIE. J.F. AND R.L. KROGER. 1974. Schooling habits of injured and parasitizeel menhaden.Ecology 55(l):208-210.
GUTTERIDGE, W.E. AND G.H. COOMBS. 1977.
Biochemistry of Parasitic Protozoa. 172 pp. Baltimore: University Park Press.
HALVER. J.E. (ed.). 1972. Fish Nutrition. 713 pp.New York: Academic Press.
HAMMOND, D.M. WITH P.I.. LONG (eels.). 1973.
The Coccidia. Eimeria, lsos|>ora. 'Toxoplasma, andRelated Genera. 482 pp. Baltimore: University ParkPress.
HARGIS. W.J.. JR.. 1957. The host specificity ofmonogenetic tremalodes. Experimental Parasitology6(6):6l()-625.
HARSHBARGER. J.C. 1977. Role of ihe registry oftumors in lower animals in the study of environmental carcinogenesis in aquatic animals. InAquatic Pollutants and Biologic Effects with
Literature Cited 127
Emphasis on Neoplasia (ed. I I.E. Kraybill. C.J.Dawe. J.C Harshbaiger. and R.G. Tardiff). Annalsof the New York Academy of Sciences 298:280-289.
HARSHBARGER. J.C., S.C CHANG. AND S.V.OTTO. 1977. Chlamydial- (with phages), mycoplasmas, and Rickettsiae in Chesapeake Bay bivalves..Science !96(4290):666-668.
HARTMAN. W.D.. 1958. Natural history ol the marinesponge-s in southern New England. PeabodyMuseum of Natural History, Yale University.Bulletin 12:1-555.
HAVEN. D.. 1959. Effects of pea crabs Pinnotheresostreum on oysters Crassostrea virginica. Proceedings of the National Sheltfisheries Association49:77-8(>.
HEARD. R.W.. 111. 1967. Life history stueiics on twoclosely related s|K-cies of Carneophallus (Micto-phallidae). 77ic* Journal of Parasitology 53(Suppl.):27.
HEARD, R.W.. JR.. 1976. Microphaiiid TrematodeMetacercariae in Fiddler Crabs of the Genus UcaLeach, 1814 from the Northern Gulf of Mexico.Ph.D. dissertation. University of Southern
Mississippi. Halliesburg. 179 pp.
HENDRIX. S.S. AND R.M. OVERSTREET. 1977.
Marine aspidogastrids (Trematoda) from fishes inthe northern Gull ol Mexico. The Journal of Parasitology 63(5):8I0-817.
HENRY. S.M. (ed.). 1966. Symbiosis. Associations ofMicroorganisms. Plants, and Marine Organisms.Vol. 1. 478 pp. New York: Academic Pre-ss.
HENRY. S.M. (ed). 1967. Symbiosis. Associations ofInvertebrates, Birds. Ruminants, and Other Biota.
Vol. II. 443 pp. New York: Academic Pre-ss.
HOESE, H.D. AND R.H. MOORE. 1977. Fishes of theGulf of Mexico. Texas, Louisiana, and AdjacentWaters. 327 pp. tiollege Station: Texas Affc.VI University Piess.
HOFFMAN. G.L.. 1967. Parasites of North AmericanFreshwater Fishes. -186 pp. Berkeley: University ofCalifornia Press.
HOFFMAN. G.L.. 1974. Disinfection of contaminated
water by ultraviolet irradiation, wilh emphasis onwhirling disease (Myxosoma cerebralis) and itseffect on fish. Transactions of the American FisheriesSociety IOS(3):54l-550.
HOFFMAN, G.I... 1976. Whirling disease of trout.U.S. Fish and Wildlife Sen'ice. Fish Disease Leaflet47:1-10.
HOFFMAN, G.L.. M. LANDOLT, J.E. CAMPER,D.W. COATS. J.L. STOOKEY, AND J.D. BUREK.1975. A disease of freshwater fishes caused byTetrahymena corltssi Thompson. 1955, and a key foridentification of holotrich ciliates ol freshwater
fishes. The Journal of Parasitology 6l(2):2l7-223.
HOFFMAN. G.L. AND F.P. MEYER, 1974. Parasites
of Freshwater Fishes: a Review of Their Control andTreatment. 224 pp. Neptune City: T.F.H. Publications.

128 Marine Maladies?
HOFFMAN. G.L. AND J.J. OGRODNICK. I977.(Join to) of whirling disease (Myxosoma cerebralis):effects ol drying, and disiiifet lion wilh hydrated lime01 chlorine. Journal of Fish Biology 10(2):175-179.
HOLLIMAN. R.B.. I963. Gyrodaclylus shorti. a newspec ies of monogeneiic irematode from the broodpoue h ol the southern pipefish. Syngnathusscovelli (Evermann and Kendall). Tulane Studies inZoology IO(2):83-86.
HON.CBERG. B.M., W. BALAMUTH. E.t:. BOVEE.
J.O. CORLISS. M. GOJDICS. R.P. HALL.R.R. KUDO. N.D. LEVINE. A.R. LOEBLICH. JR..J. WEISER. AND D.l I. WENRICH. I964. Arevisedclassifit alion of the phylum Protozoa. The Journalof Protozoology 11 (I): 17-20.
HOPKINS. S.H... I917. The nemertean Carcino-
nemertes as an indicator of ihe spawning history ofthe host. Callinectes sapidus. The Journal ofParasitology 33(2):146-150.
HOPKINS. S.H.. 1956a. Notes on the boring spongesin Gulf coast estuaries and their relation lo salinity.Bulletin of Marine Science of the Gulf and Caribbean6(1): 14-58.
HOPKINS. S.H.. 1956b. The boring s|Kinges which
aitac k South Carolina oysters, with notes on someassociated organisms. Contributions from BearsBluff Laboratories, Wadmalaw Island, SouthCarolina No. 23:1-30.
HOPKINS. S.H., 1956c. Odostomia impressa parasitizing southern oysters. Science 124(3223):628-629.
HORSLEY. R.W., 1977. A review of the bacterial flora
of icleosis and elasmobranchs. including methods lorits analysis. Journal of Fish Biology IO(6>:529-553.
HOWSE. H.D. AND J.Y. CHRISTMAS. 1970.Lymphocystis tumors: histochemical identificationof hyaline substances. Transactions of the AmericanMicroscopical Society 89(21:276-282.
HOWSE. H.D. AND J.Y. CHRISTMAS. 1971. Observations on the ultrastruciure of lymphocystis virus inthe Atlantic croaker. Micropogon undulalus(Linneaus). Virology 44(1):211-214.
HUMES, A.G.. 1942. The morphology, taxonomy, andbionomics of the nemertean genus Carcinonemertes.Illinois Biological Monographs 18(0:1-105.
HUTTON. R.F. AND F. SOGANDARES-BERNAL.
1959. Notes on the distribution of the leech,
Myzobdella lugubris l.eidy, and its association wilhmortality of the blue crab, Callinectes sapidusRaihbun. The Journal of Parasitology 45(4):38i.4(H.430.
HUTTON, R.F.. F. SOGANDARES-BERNAL.
B. EDLRED. R.M. INGLE, AND K.D.
WOODBURN. 1959. Investigations on the parasitesand diseases of saltwater shrimps (Penaeidae) ofsports and commercial importance to Florida.Florida State Board of Consewation. Marine Laboratory. Technical Series No. 26:1-38.
HYMAN. L.H., 1940. The polyclad flalworms of On-Atlantic coast of the United Stales and Canada.
Proceedings of the United States National Museum
89(310l):449-495.
IYASHKIN. V.M.. A.A. SOBOLEV. AND L.A.
KHROMOVA. 1971. Camallanata of Anina's andMan and Diseases Caused by Them. Essentials ofNematodology Vol. XXII. (ed. K.I. Skrjabin,translated by H. Hardin. 1977), 381 pp. Academy ofSciences of theUSSR. Helminthological laboratory,Moscow. Jentsalem: Israel Program for ScientificTranslations.
IVERSEN, E.S. AND J.F. KELLY. 1976. Micro-sporidiosis successfully transmitted experimentallyin pink shrimp. Journal of Ini'ertebrate Pathology27(3):407-l08.
IVERSEN. E.S. AND H.O. YOSHIDA. 1957. Notes on
ihe biology of the wahoo in the Line Islands. PacificScience 11(4):370-379.
JACKSON, G.J., 1975. The "new disease" status ofhuman anisakiasis and North American cases: a
review. Journal of Milk and Food Technology38(l2):769-773.
JAKOWSKA. S.. R.F. NIGRELLI. AND 1. ALPERIN.1954. A new Henneguya in the North Atlanticweakfish. Cynoscion regalis. The Journal of Protozoology l(Suppl.):13.
JAWORSKI. E.. 1972. The Blue Crab Fishery.Barataria Estuary. Louisiana. LSU-SG-72-01.
112 pp. Baton Rouge: O-nier for Wetland Resources.Louisiana State University.
JENNINGS. J.B., 1974. Symbioses in the Turbellariaand their implications in studies on the evolution ofparasitism. In Symbiosis in the Sea (ed. W.B.Vernberg). pp. 127-160. Columbia: University ofSouih Citolina Press.
JENSEN. LA. AND R.A. HECKMANN. 1977.Anantrum histocephalum sp. n. (Ccscoda: Boihrio-cephalidae) from Synodus lucioceps (Synodontidae)of southern California. The Journal of Parasitology63(3):47l-472.
JOHNSON. P.T.. 1976. Bacterial infection in the bluecrab, Callinectes sapidus: course of infection andhisiopalhology. Journal of Invertebrate Pathology28(l):25-36.
JOHNSON. P.T.. 1977a. Paramoebiasis in the blue-crab, Callinectes sapidus. Journal of InvertebratePathology 29(3):308-320.
JOHNSON, P.T.. 1977b. Virus diseases of ihe blue-crab, Callinectes sapidus. Program of the 2ndAnnual Eastern Fish Health Workshop. F.aston.Maryland, pp. 22-24. (also, Marine Fisheries Review.in press).
JOHNSON. S.K., 1974. Eclocommensals and Parasitesof Shrimp from Texas Rearing Ponds. TAMU-SG-74-207.20 pp. CollegeSialion: Department of MarineResoutces Information. Center (en Marine
Resources, Texas A&M University.
JOHNSON. S.K., 1977. Handbook of Crawfish andFreshwater Shrimp Diseases. TAMU-SG-77-605.19 pp. College Station: Department of MarineResources Information, Center for Marine
Resources. Texas AifcM University.

»«•
JOHNSON, S.K., 1978.Handbook of Shrimp Diseases.TAMU-SG-75-603 (revised ed.). 23 pp. CollegeStation: Department of Marine Resources Information, Center for Marine Resources, 'Texas A&M
University.
JOHNSON, S.K., D. BYRON. G. LARA M. AND T.P.FERREIRA DE SOUZA. I975. Cramped conditionin pond-reared shrimp. Penaeus brasilienscs.FDDL-S6. 2 pp. College Scation: Fish DiseaseDiagnostic Laboratory, Texas AgriculturalExtension Service, Department of Wildlife andFisheries Sciences, Texas A&M University System.
JOHNSON. S.K. AND W.A. ROGERS. 1973. Distribution of the genus Ergasilus in several Gulf ofMexico drainage basins. Agricultural F.xperimentStation, Auburn University, Bulletin -1-15:1-74.
JOYCE. E.A.. JR.. 1961. The Hydroida of the Seahorse, Key Area. M.S. thesis, University of Florida,
Gainesville. 116 pp.
KABATA. Z., 1970. Crustacea as Enemies of Fishes.Book 1. In Diseases of Fishes (eds. S.F. Snieszko andH.R. Axelrod). 171 pp. Jersey City: T.F.H. Publications.
KARMANOVA. E.M.. 1962. The life-cycle ofDioclophyme renale in its intermediate and definitive hosts. (In Russian) Trudy HelminthologicalLaboratory, USSR Academy of Science 12:27-36.
KEARN, G.C. 1971. Pie physiology and behaviour ofthe monogenean skin parasite Enlobdella soleae inrelation to its host (Solea solea). In Ecology andPhysiology of Parasites (ed. A.M. Fallis), pp. 161-187.Toronto: University of Toronto Press.
KERN. F.G.. 1976. Minchinia nW.Kini(MSX)diseaseofthe American oyster. Marine Fisheries Review 38( 10):22-24.
KHALAF. K.T., 1967. Seasonal incidence and population densities of Culicoides in ihe coastal areas of
Louisiana (Diptera: Ccralopogonidac). Journal ofthe Kansas Entomological Society 40<4):472-477.
KHALAF, K.T., 1969. Distribution and phenology ofCulicoides (Diptera: Ceralopogonidae) along theGulf of Mexico. Annab of the Entomological Societyof America 62(5):1153-1161.
KHAN, R.A.. 1977. Susceptibility of marine fish lotrypanosomes. Canadian Journal of Zoology 55(8):1235-1241.
KHEYSIN. Y.M. (ed. K.S. TODD. JR.). 1972. LifeCycles of Coccidia of Domestic Animals. ('Translatedby F.K. Pious, Jr.) 264 pp. Baltimore: UniversityPark Press.
KLINE. D.L. AND R.C. AXTELL. 1976. Salt marsh
Culicoides (Diptera: Ceralopogonidae): species,seasonal abundance and comparisons of trappingmelhods. Mosquito News 36(l):l-10.
KORATHA. K.J., 1955a. Studies on the monogcnelicnematodes of thcTexascoasc. I. Results of a survey ofmarine fishes al Port Aransas, wilh a review of
Monogenea reported from the Gulf of Mexico andnotes on euryhalinity, hosl-s)x-cificity, and relationship of ihe remora and the cobia. Publications of the
Literature Cited 129
Institute of Marine Science 4(l):233-250.
KORATHA, K.J., 1955b. Studies on the monogcnelictrematodes of the Texas coast. II, Descriptions ofspecies from marine fishes of Port Aransas.Publications of the Institute of Marine Science4(l):25l-278.
KRAYBILL, H.F., C.J. DAWE. J.C. HARSHBARGER. AND R.G. TARDIFF (eds.). 1977.
Aquatic Pollutants and Biologic Effects withEmphasis on Neoplasia. Annals of the New YorkAcademy of Sciences 298:1-604.
KRUSE, D.N.. 1959. Parasites of the commercial
shrimps, Penaeus aztecus Ives, P. duorarumBurkenroad and P. setiferus (Linnaeus). TulaneStudies in Zoology 7(4): 123-144.
KRUSE, D.N.. 1966a. Life cycle studies on Nematopsisduorari n. sp. (Gregarinia: Porosporidae), A parasiteof the pink shrimp (Penaeus duorarum) andpclecypod molluscs. Dissertation Abstracts27B:2919-B.
KRUSE. D.N., 1966b. Life Cycle Studies onNematopsis duorari n. sp. (Gregarinia: Porosporidae), A Parasite of the Pink Shrimp (Penaeusduorarum) and Pelecypod Molluscs. Ph.D. thesis,Florida State University, Tallahassee. 174 pp.
KUDO, R., 1920. Studies on Myxosporidia. Asynopsisof genera and species of Myxosporidia. IllinoisBiological Monographs 5(3-4):3-265.
KUDO, R.R., 1966. Protozoology. 5lh ed. 1,174 pp.Springfield: Charles C 'Thomas.
LAKSHMI, G.J.. A VENKATARAMIAH. AND H.D.HOWSE, 1978. Effect of salinity and temperaturechanges on spontaneous muscle necrosis in Penaeusaztecus Ives. Aquaculture l3(l):35-43.
LAMONT, J.M.. 1961. Fish with rubber bands.Scottish Fisheries Bulletin 15:17-18.
LANG. W.H., JR.. 1976a. The larval development ofthe barnacles Octolasmis mulleri and Chelonibia
patula. American Zoologist !G(2):219.
LANG, W.H., 1976b. The larval development andmetamorphosis of the pedunculate barnacleOctolasmis mulleri (Cokcr. 1902) reared in thelaboratory. Biological Bulletin l50(2):255-267.
LAWLER, A.R., 1977. The parasitic dinoflagellateAmyloodinium ocellatum in marine aquaria. Drumand Croaker 17(2):17-20.
LAWLER. A.R. AND R.N. CAVE. 1978. Deaths of
aquarium-held fishes caused by monogenetictrematodes. I. Aspinatrium pogoniae (Mae Odium.1913) on Pogonias cromis (Linnaeus). Drum andCroaker 18(l):3l-S3.
LAWLER, A.R.. H.D. HOWSE, AND D.W. COOK.
1974. Silver |>ereh,Bairdiella chrysura: new host forlymphocystis. Copeia 1974(1 ):266-269.
LAWLER, A.R. AND R.M. OVERSTREET, 1976.
Absonifibula bychowskyi gen. el sp. nov.(Monogenea: Absonifibulinae subfam. nov.) fromthe Atlantic croaker, Micropogon undulatus (L.).from Mississippi, U.S.A. In Studies on the

130 Marine Maladies?
Monogeneans. Proceedings of the Institute ofBiology and Pedology, Far-East Science Centre,Academy of Sciences of the USSR, New Series34(137):83-9I.
LAWLER. A.R. AND S.L. SHEPARD. I978. A
partially albino blue crab. Drum and Croaker )8( I):34-36.
LEE, J.S.. 1973. What seafood processors should knowabout Vibrio parahaemolyticus. Journal of Milk andFood Technology 36(8): 405-408.
LEIGH, W.H., I960. The Florida spotted gar. as theintermediate host of Odhneriotrema incommodum
(Leidy. I856) from Alligator mississippiensis.The Journal of Parasitology 46(5, Suppl.):!<>.
LESTER. R.J.G.. 1972. Attachment eif Cyrodeuty/imoGasterosteus and host response. 77ie Journal ofParasitology 58(4):7l7-722.
LEVINE, N.D., 1973. Protozoan Parasites of DomesticAnimals and of Man. 2nd ed. 406 pp. Minnea|x>lis:Burgess Publishing.
LEVINE. N.D.. 1978. Perkinsus gen. n. and other newlaxa in ihe protozoan phylum Apicomplexa. TheJournal of Parasitology 64(3):549.
LIGHTNER. D.V.. 1973. Normal postmortem changesin the brown shrimp. Penaeus aztecus. FisheryBulletin 72( 1):223-236.
LIGHTNER, D.V., 1975. Some potentially seriousdisease problems in the culture of penaeid shrimp inNorth America. In Proceedings of the Third U.S.Japan Meeting on Aquaculture at Tokyo, Japan,Oct. 15-16,1974. pp. 75-97. Niigala: Fishery Agency.Japanese Government and Japan Sea RegionalFisheries Ri*search Laboratory.
LIGHTNER. D.V. AND R. REDMAN. 1977.Histochemical demonstration of melanin in cellular
inflammatory processes of penaeid shrimp. Journalof Invertebrate Pathology 30(S):298-302.
LIGHTNER, D.V.. B.R. SALSER, AND R.S.
WHEELER. 1974. Gas-bubble disease in the brown
shrimp (Penaeus aztecus). Aquaculture 4(l):8l-84.
LINDNER, M.J. AND H.L. COOK, 1970. Synopsis ofbiological data on the white. shrimp Penaeussetiferus (Linnaeus) 1767. FAO Fisheries Report57(4): 1439-1469.
LOCKE, L.N., J.B. DeWITT, CM. MENZIE. ANDJ.A. KERWIN, 1964. A merganser die-off associatedwith larval Eustrongytides. Avian Diseases 8(3):420-427.
LOESCH, H.C., 1957. Studies of the ecology of twospecies of Donax on Mustang Island, Texas. Publications of the Institute of Marine Science4(2):201-227.
LOM. J., 1969. Notes on the ultrastructure and sporo-blast development in fish parasitizing myxo-sporidian of the genus Sphaeromyxa. Zeitschrift furZellforschung und Mikroskopische Anatomie 97(3):416-437.
LOM, J., 1970. Protozoa causing diseases in marinefishes. In A Symposium on Diseases of Fishes andShellfishes (ed. S.F. Snieszko). American Fisheries
Society Special Publication No. 5: 101-123.
LOM, J., 1973. The adhesive disc of Trichodinetlaepizootica — ultrastructure and injury to the hosttissue. Folia Parasitologica (Praha) 20(3): 193-202.
LOM. J. AND A.R. LAWLER. 1973. An uhrastruciuralstudy on the mode of attachment in dinoflagellatesinvading gills of Cyprinodontidae. Protistologica9(2):293-309.
LONGWEI.L, A.C. 1976. Chromosome mutagenesisin developing mackerel eggs sampled from ihe New-York Bight. NOAA Technical Memorandum ERLMESA-7, Marine Ecosystems Analysis ProgramOffice, Boulder, Colorado. 61 pp.
LOWERY. G.H.. JR.. 1974. The Mammals ofLouisiana and its Adjacent Waters. 565 pp. BatonRouge: Louisiana Slate University Press.
LUMSDEN. R.D.. 1975. Surface uhrasiructiire and
cytochemistry of parasitic helminths. ExperimentalParasitology 37(2):267-339.
MACKENZIE. CL.. JR.. 1977. Development of anaquacullural program for rehabilitation of damagedoyster reefs in Mississippi. Marine Fisheries Review39(8):I-I3.
McDERMOTT, J.J.. 1960. The predation of oystersand barnacles by crabs of the family Xanthidae.Proceedings of the Pennsylvania Academy of Science34:199-211.
MCDONALD. M.E.. 1969a. Annotated bibliography ofhelminths of waterfowl (Anatidae). Bureau of SportFisheries and Wildlife Special Scientific Report —Wildlife No. 125:1-333.
MCDONALD. M.E.. 1969b. Catalogue of helminths ofwaterfowl (Anatidae). Bureau of Sport Fisheries andWildlife Special Scientific Report — WildlifeNo. 126:1-692.
McERLEAN. A.J.. C. KERBY. AND M.L. WASS(eds.). 1972.Biota of the Chesapeake Bay. ChesapeakeScience l3(Suppl.):l-l97.
McGRAW. K.A. AND G. GUNTER, 1972. Obser
vations on killing of the Virginia oyster by the Gulfoyster borer, Thais haemastoma. with evidence fora paralytic secretion. Proceedings of the NationalShellfisheries Association 62:95-97.
Mclaughlin, j.j.a. and p.a. zahl. 1957.Studies in marine biology. II. In vitro culture ofzooxanthellae. Proceedings of the Society for Experimental Biology and Medicine 95:115-120.
MACE, T.F. AND R.C. ANDERSON. 1975. Development of the giant kidney worm, Dioctophyma renale(Goeze, 1782) (Ncmatoda: Dioctophymatoidea).Canadian Journal of Zoology 53(11):1552-1568.
MACKIN. J.C, 1962. Oyster disease caused byDermocystidium marinum and other microorganisms in Louisiana. Publications of theInstitute of Marine Science 7:132-229.
MACKIN, J.G. AND F.G. SCHLICHT. 1976. Aproteomyxan amoeba stage in the development ofLabyrinthomyxa patuxent (Hogue) Mackin andSchlicht, with remarks on the relation of ihe

proteomyxids to the neoplastic diseases of oysters andclams. Marine Fisheries Review 38(I0):16-I8.
MAI.MBERG. C. 1971. On the larval protonepliridialsystem ed Gyrocotyle and the evolution ofCercomeiomorphac (Plalyhelminthes). ZoologicaScripta 3:65-81.
MARGOLIS, L., 1970. Nemalexle- diseases ol marine
fishes. In A Symposium on Diseases of Fishes andShellfishes (ed. S.F. Snieszko). American FisheriesSociety Special Publication No. 5:190-208.
MARGOLIS. I... 1977. Public health aspects of"codworm" infection: a review. Journal of theFisheries Research Board of Canada 3!(7):887-898.
MARGOLIS. I... Z. KABATA. AND R.R. PARKER.
1975. dialogue and synopsis ol Caligus, a genus ofCopepoda (Crustacea) parasitic on fishes. Bulletinof the Fisheries Research Board of Canada 192:1-117.
MATTHEWS, G. AND H.O. WRIGHT. 1970. The
selection of hermit crab host by the commensalhydroid. Hydractinia sp. Abstracts of SouthwesternAssociation of Naturalists. 17th Annual Meeting,Texas AirM University. 23-26 April 1970:b.
MAY, E.B., 1973. Extensive oxygen depletion in MobileBay. Alabama. Limnology and Oceanography 18(3):353-366.
MAY. H.C.. 1919. Contributions to the life hisioiie-s of
Gordius robustus I.eicly and Paragordius varius(Leiely). Illinois Biological Monographs 5(2):1-118.
MEARNS. A.J. AND M.J. SHERWOOD. 1976. Oceanwastewater discharge and tumors in a southernCalifornia flatfish. Progress in Experimental TumorResearch 20:75-85.
MELDRIM. J.W.. J.J. GIFT. AND B.R. PETROSKY.1974. The effect ol lemperaluic and chemicalpollutants on the behavior ol several estuarineorganisms. Ichthyologual Associates Inc. BulletinNo. 11:1-129.
MENZEL. R.W. AND S.H. HOPKINS. 1955. The
growth of oysters parasitized by the fungusDermocystidium marinum and by the iremaiodeBucephalus cuculus. The Journal of Parasitology4l(4):333-342.
MENZIES. R.J.. T.E. BOWMAN. AND F.G.
ALVERSON. 1955. Studies ol the biology of ihe fishparasite l.ivoneca convexa Richardson (Crustacea.Isopcxla. Cymoihoidac). 77re Wasmann Journal ofBiology l3(2):277-295.
MERCANDO, N.A., 1976. A pattern of specificity inthe symbiotic assoe iation between Hydractiniaechinata and hermit crabs from coastal North
Carolina. 77»r Journal of the Elisha MitchellScientific Society 92(2):84-85.
MEYER. F.P.. 1970. Seasonal flue luations in the
incidence of disease on fish farms. In A Symposiumon Diseases of Fishes and Shellfishes (ed. S.F.Snieszko), American Fisheries Society SpecialPublication No. 5:21-29.
MILLS. C.E.. 1976. Podocoryne selena. a new speciesofhydroid from the Gulf of Mexico, and a comparisonwilh Hydractinia echinata. Biological Bulletin
Literature Cited 131
15l(l):2ll-224.
MOLLHAGEN. T.R.. 1977. A study of the sysiemaiicsand hosts of the parasitic nematode genusTetrameres (Habionemaioidea: Tetranicridae).Dissertation Abstracts International 38(2. Pt. B):
547-B.
MOORE. C.J. AND CR. POSEY. SR.. 1974. Pigmentation and morphological abnormalities in the hog-choker. Trinecles maculatus (Pisces, Solc-idac).
Copeia I97l(3):660670.
MOORE. R.H.. 1976. Observations on fishes killed bycold at Port Aransas, 'Texas. 11-12 January 1973.The Southwestern Naturalist 2()(l):461-16li.
MORE. W.R.. 1969. A contribution to the biology ofthe blue crab (Callinectes sapidus Rathbun) inTexas, wilh a description of the fishery. Texas Parksand Wildlife Department Technical SeriesNo. 1:1-31.
MUDRY. D.W. AND M.D. DAILEY. 1971. Postern-
bryonie development of certain tc-iraphyllidcan andirypaiiorhynchancc-slodcs wilba |x>ssil>lcalicmaii\clife cycle- for the order Trypanorhyncha. CanadianJournal of Zoology 19:1249-1253.
NEUHAUS, O.W. AND J.E. HALVER (eds.). 1969.Fish in Research. 311 pp. New York: AcademicPress.
NEWMAN. W.A.. 1967. Shallow-water versus deep-seaOctolasmis (Cirrirx-dia Thoiacica). Crustaceana12(1): 13-32.
NEWMAN. M.W. AND CA. JOHNSON. 1975. Adisease- ol blue crabs (Callinectes sapidus) causedby a parasitic dinoflagellate. Hemaiodimum sp.The Journal of Parasitology 6l(3):554-557.
NEWMAN. M.W.. CA. JOHNSON. HI. ANDG.B. PAULEY. 1976. A Afinc/iinia-like haplosporidan parasitizing bluecrabs. Callinectes sapidus.journal of Invertebrate Pathology 27(3):311-315.
NICKLE. W.R.. 1972. Acontribulion toour knowledgeof the Mermithidae (Nematoda). Journal ofNematology 4(2): 113-146.
NICKOL. B.B. AND R.W. HEARD. HI. 1970.
Arhythmorhynchus frassoni from the elappet rail.Rallus longirostns, in North America. The Journalof Parasitology 56(l):204-2O6.
NIGRELLI. R.F. AND G.M. SMITH. 1938. Tissue
responses of Cyprinodon variegatus to the myxo-sporidian parasite. Myxobolus lintoni Gurley.Zoologica 23(2): 195-202.
NOBLE. E.R. AND G.A. NOBLE. [971. Parasitology.The Biology of Animal Parasites. 3rd ed. 617 pp.Philadelphia: Lea & Febiger.
NORMAN, J.R., 1934. A Systematic Monograph of theFlatfishes (Heterosomata). Psettodidae, Bothidae,Pleuronectidae. Vol. I. 459 pp. I.ondon: OxfordUniversity Press.
NORRIS. D.E. AND R.M. OVERSTREET, 1975.
Thynnascaris reliquens sp. n. and T. habena(Linton. l900)(Nematcxla: -War ieloielea) from fishes
in the northern Gull of Mexico and eastern U.S.
seaboard. The Journal of Parasitology 61(2):330-336.

132 Marine Maladies?
NORRIS. D.E. AND R.M.OVERSTREET. 1976.The
public health implications of larval Thynnascarisnematodes from shellfish. Journal of Milk and FoodTechnology 39(l):47-54.
OLLA. B.L.. A.L. STUDHOLME. A.J. BEJDA. CSAMET. AND A.D. MARTIN. 1975. The effect of
temperature on the behaviour of marine fishes: Acomparison among Atlantic mackerel. Scomberscombrus. bluefish. Pomatomus saltatrix. and
tatifog. Tauloga onitis. In Combined Effects ofRadioactive, Chemical and Thermal Releases to the
Environment, pp. 299-308. Vienna: InternationalAtomic Energy Agency.
OLSEN. O.W.. 1974. Animal Parasites. Their LifeCycles and Ecology. 3rd ed. 562 pp. Baltimore:University Park Press.
OR1AS. J.D. AND E.R. NOBLE. 1971. Entamoebanezumia sp. u. and other parasites from a NorthAtlantic fish. 77ie Journal of Parasitology57(5):945-947.
OSHIMA, T., 1972. Anisakis and anisakiasis in Japanand adjacent area. Progress of Medical Parasitologyin Japan 4:301-393.
OVERSTREET. R.M., 1968. Parasites of the inshore
lizardfish, Synodus foetens, from south Florida,including a description of a new genus of Cesloda.Bulletin of Marine Science 18(2):l44-470.
OVERSTREET. R.M.. 1970. Two new species ofDigenea from the spot, Leiostomus xanthurusLaccpede. from the Gulf of Mexico. 77re Journal ofParasitology 56(6):1055-I057.
OVERSTREET. R.M.. 1971a. A'eoc/ia.imic.t
sogandaresi n. sp. (Trematoda: Crypiogonimidae)from the slri|x-d bass in Mississippi. Transactionsof the American Microscopical Society 90( 1):87-89.
OVERSTREET. R.M.. 1971b. Glaucivermis spinosusgen. ei sp. n. (Digenea: Zoogoniclac) from thesouthern kingfish, Menticirrhus americanus(Linnaeus), in the coastal waters of Mississippi.The Journal of Parasitology 57(3):536-538.
OVERSTREET. R.M., 1971c. Metadata spectandaTravassos, Freilas, and Biihrnheim, 1967 (Digenea:Cryptogonimidae) in estuarine fishes from theGulf of Mexico. Proceedings of the Helminthological Society of Washington 38(2):156-I58.
OVERSTREET, R.M., 1973. Parasites of some penaeidshrimps with emphasis on reared hosts. Aquaculture2(2):105-140.
OVERSTREET. R.M.. 1974. An estuarine low-
le*mperalure fish-kill in Mississippi, wilh remarkson restricted necropsies. Gulf Research Reports4(3):328-350.
OVERSTREET. R.M.. 1975. Buquinolale as a preventive drug to control microsporidosis in the bluecrab. Journal of Invertebrate Pathology 26(2):213-216.
OVERSTREET. R.M.. 1976a. Fabespora vermicolasp. n.. the first myxosporidan from a platyhclmimh.The Journal of Parasitology 62(5):680-684.
OVERSTREET. R.M., 1976b. A redescriplion of
Crassicutis archosargi. a digenean exhibiting anunusual tegumental attachment. 77ie Journal ofParasitology 62(5):702-708.
OVERSTREET. R.M.. 1977a. A revision of Saturnius
Mauler. 1969 (Hemiuridae: Butiocoiylinae) wilhdescriptions of two new species from the stripedmullet. UniversidadNacionalBMexico,Inslilutode
Biologia Publicaciones Especiales 4:273-284.
OVERSTREET. R.M.. 1977b. Poecilancistrium
caryophyllum and other trypanorhynch cestodeplerocercoids from the musculature of Cynoscionnebulosus and oilier sciaenid fishes in the Gulf ol
Mexico. The Journal of Parasitology 63(5):780-789.
OVERSTREET. R.M. AND D.W. COOK. 1972. An
underexploiied Gulf coast fishery: soft shelledcrabbing. The American Fish Farmer 3(9):12-17.
OVERSTREET. R.M. AND R.H. EDWARDS. 1976.
Mesenchymal tumors of some estuarine fishes of thenoil hern Gulf of Mexico. II. Subc utaneous fibromas
in the southern flounder, I'aratichthys lethostigma.and ihe sea catfish, Arius felis. Bulletin of MarineScience 26(I ):41-48.
OVERSTREET. R.M. AND F.G. HOCHBERG. JR..1975. Digenetic nematodes in ccphalopods. journalof the Marine Biological Association of the UnitedKingdom 55(0:893-910.
OVERSTREET. R.M. AND H.D. HOWSE. 1977.
Some parasites and diseases of estuarine fishes inpolluted habitats of Mississippi. In AquaticPollutants and Biologic Effects with Emphasis onNeoplasia (eds. C.J. Dawe. J.C. Harshbarger. andR.G. Tardiff). Annals of the New York Academy ofSciences 298:427-162.
OVERSTREET. R.M. AND C.H. LYLES. 1974. A
rubber band around an Atlantic croaker. GulfResearch Reports 4(3): 176-178.
OVERSTREET, R.M. AND l-I.M. PERRY. 1972. A
new microphaiiid trematode from the blue crab inthe northern Gulf of Mexico. Transactions of theAmerican Microscopical Society 9l(S):436-440.
OVERSTREET. R.M. AND T. VAN DEVENDER.
1978. Implication of an environmentally inducedhamartoma in commercial shrimps. Journal ofInvertebrate Pathology 3l(2):234-238.
OVERSTREET. R.M. AND E. WEIDNER. 1971.
Differentiation of microsixiridian spore-tails inInodosporus spraguei gen. el sp. n. Zeilschrift fiirParasitenkunde 44(3):169-186.
OVERSTREET. R.M. AND E.G. WHATLEY. JR..1975. Prevention of microsporidosis in the blue crab,with notes on natural infections. Proceedings ofSixth Annual Workshop World Mariculture Society:335-345.
PAPERNA., I., 1963. Some observationson the biologyand ecology of Dactylogyrus vastalor in Israel.Bamidgeh 15(l):8-29.
PAPERNA, I.. 1964. Competitive exclusion ofDactylogyrus extensus by Dactylogyrus vastalor(Trematoda, Monogenea) on ihe gills of reared carp.The Journal of Parasitology 50(l):94-98.

PAPERNA. I. AND R.M. OVERSTREET. In press.Parasites and diseases of mullets (Mugilidae). InInternational Biological Programme, The Mullets(ed. OH. Orel)I. Loudon: Cimbiidgc- UniccisilvPress.
PAPERNA. I. AND F.D. POR. In press. Preliminarydata on theGnailiiidac(I.sopoda)of ihe northern RedSea. the Bitter Lakes and the eastern Mediterraneanand the biology of Gnathia piscivora n. sp. AWTCongress of I.C.S.E.M.. October l'>76. Split.Yugoslavia.
PAPERNA, I. AND D.E. /.WERNER. 1976. Studies onErgasilus labracis Krjtyer (Cyclopidea: Ergasilidae)parasitic on striped bass. Morone saxaltlis. from thelower Chesapeake Bay. I. Distribution, life cycle,and seasonal abundance. Canadian Journal ofZoology 54(0:149-462.
PAULEY. G.B.. K.K. CHEW. AND A.K. SPARKS.1967. Exix-rimenlal infection of oystets (Crassostreagigas) with ihigmotiichid ciliates. Journal ofInvertebrate Pathology 9(2):230-231.
PEARSE. L.. 1972. A note on a marine iiithodiiiid
ciliate parasitic on the skin of captive flatfish.Aquaculture l(3):26l-266.
PEARSE. A.S. AND C.W. WHARTON. 1938. The
oyster "leech." Stylochus immiciis Pa lorn hi.associated with oysters on ihe coasts of Florida.Ecological Monographs 8(0:605-655.
PENCE. D.B.. 1972. The hypopi (Acarina:Sarcoptiformes: Hypoderidae) from the subcutaneous tissues of birds in Louisiana. Journal ofMedical E.ntomology 9(5): 135-438.
PENCE. D.B.. 1973a. Reighardia sternae (Cephalo-bacnida: Reighardiidae). a pentaslome from gullsand terns in Louisiana. Proceeding: of theHelminthological Society of Washington 40(1):161-165.
PENCE. D.B.. 1973b. The nasal mites ol birds from
Louisiana. fX. Synopsis. The Journal of Parasilo-focj>-59(5):88l-892.
PENCE. D.B.. 1973c. Tetrameres aspinosa n. sp. fromthe snowy egree and a new host record for T. robusta(Nematoda: Telramericlae). Transactions of theAmerican Microscopical Society 92(3):522-525.
PENCE. D.B.. 1975. Keys, species and host list, andbibliography for nasal mites of North Americanbirds (Acarina: Rhinonyssinae. Turbinoptinae.Speleognathinae, and Cytoditidae). 77ie Museum,Texas Tech University, Special PublicationsNo. 8:1-148.
PENNAK. R.W.. 19S3. Fresh-Water Invertebrates of theUnited States. 769 pp. New York: Ronald Press.
PENNER. L.R. AND P.J.S. RAJ. 1977. Concerningthe marine leech, Pontobdella macrothela Schmarda,1861 (Piscicolidae: Himdinea). Excerta Parasi-tologica en Memoria del Doctor Eduardo Caballero yCaballero, Universidad Nacional Autonoma de
Mexico, Institute) de Biologia PubltcacionesEspeciales 4:519-530.
PENSE [sic], D.B. AND G.E. GUILDS. 1972.
Literature Cited 133
Pathology of Amphtmerus etongatus (Digcuca:Opislhoichiidar) in the- liver of ihe double-c lesicclcoimotmu. Journal of WildlifeDi\en.\r\Htfr.?>\-?>\.
PEREZ l-ARFANTE. I.. 1969. Western Atlantic
sill imps ol the genus Penaeus. Fishery Bulletin67(3 ):-!<> I-591.
PERKINS. F.O.. I968. Fine- structure ol ihe- oisici
pathogen Minchinia nelsoni <Ilaplospcn ida.Haplospoiidiidac). Journal of lit.erlehiiitcPathology Ul(2):287-.t05.
PERKINS. F.O., 1969. Election microseopc- studies olspeculation in the- ocsiei palhogeu. Minchiniacostalis (Sporozoa: Haplospoiida). 77ic Journal ofParasitology 55(5):897-920.
PERKINS. F.O.. 197I. Spoiutalion in ihe- lli'iu.ilodc-lnpi'lpaiasilc e" impioiiliuiii <)c\trn\ l)i I ink. Ml III(Haplospoiida: Haplospoiidiidac) — an electronmicroscope study. The journal of Paiasilology57(l):9-23.
PERKINS. F.O.. 1976a. Dermocystidium marinuminfection in oystets. Marine Fisheries /eYi-iet<-38(l(l|:19-21.
PERKINS. F.O.. 197tib. Zoospores ol the oysterpathogen. Dermocystidium marinum. I. l-inestiiie-ture of the conoid and other spoiozoau-likcorganelles. 77i<- journal <d Paia\itiilog\- l>2elii:959-974.
PERKINS, F.O. AND M. CASTAGNA. I97l. Ullra-
slruclureof ihe Nebenkorper ti\-"secondary line lens"of the parasitic amoeba Paramoeba prrnitiosa(Amocbida. Paramoebidae). Journal of InvertebratePathology 17(2): 186-193.
PERRY. H.M.. 1975. The- blue crab fishery inMississippi. Gulf Research Reports 5(l):39-57.
PETERSEN. J.J.. 1973. Role of mermithid nematodesin biological control of mosquitoes. ExperimentalParasitology 33(2):239-217.
PETERSEN. J.J. AND O.R. WILLIS. 1972. Resultsof preliminary field applications of Reesimermisnielseni (Mermithidae: Nemaioda) lo controlmosciuito larvae. Mosquito News 32(3):3I2-316.
PLUMB. J.A.. 1974. Viral diseases of fishes ol the Gulfof Mexico legion. In 1974 Proceedings. Gulf CoastRegional Symposium cm Diseases of AquaticAnimals (eels. R.L. Amborski. M.A. Hood, and R.R.Miller), pp. 55-75. LSU-SG-74-05. Baton Rouge:Center for Wetland Resources. Louisiana Slate
University.
PLUMB, J.A., J.H. SCHACHTE. AND J.L. GAINES.
1974. Streptococcus sp. from marine fishes alongthe Alabama and northwest Florida coast of ihe Gulf
of Mexico. Transactions of the American FisheriesSociety I03(2):358-361.
PROVASOI.I. L.. J.J.A. Mc:LAUCHLIN. AND M.R.DROOP. 1957. The development of artificial mediafor marine algae. Archiv fur Mihrobiologie25:392-428.
PURDOM. CE. AND A.E. HOWARD. 1971. Ciliate
infestations: a problem in marine fish farming.Journal du Conseil. Cornell Permanent Inter
national pour {'Exploration de la At>r33(3):5l I-511.

134 Marine Maladies?
DE PUYTORAC P.. A BATISSE. J. BOHATIER.J.O. CORLISS. G. DEROUX. P. DIDIER. J.DRACESCO. G. FRYD-VERSAVEL. J. GRAIN.C GROLIERE. R. HOVASSE. F. IFTODE.
M. LAVAL. M. ROQUE. A SAVOIE. AND M.TUFFRAU. I974. Pio|>osiiion d'uut-c lassifitaliondu phylum Ciliophora Dollein. I90I. CompteRendu de I'Academic des Sciences. Paris 278:
2799-2802.
l'YI.E. R. AND E. CRONIN. I950. The generalanatomy ol the- blue crab Callinectes sapidusRalhbun. Chesapeake Biological Laboratory.Solomons Island. Manland. Publication No. 87:
I-10.
QUICK. J.A., JR.. 1971. Pathological and para-silological effects of elevated lemperaiutes on theoyster Crassostrea virginica wilh emphasis on thepathogen Labyrinthomyxa marina. In A Preliminary Investigation: The Effect of ElevatedTemperature on the American Oyster Crassostreavirginica (Gtnelin) (ed. J.A. Quick. Jr.). FloridaDepartment of Natural Resources, Marine ResearchLaboratory. Professional Papers Series No. 15:105-176.
QUICK. J.A.. JR.. 1972. Fluid thioglycollate mediumassay of/.afcyriiil/ioHiyvo parasites in oysters. FloridaDepartment of Natural Resources. Marine ResearchLaboratory, Leaflet Series 6(Pt. 4. No. 3):l-12.
QUICK. J.A.. JR. AND G.E. HENDERSON. 1974.Effects of Gymnodinium breve reel tide on fishesanil birds: A preliminary return on behavior,anatomy, hematology, and histopalhology. In 1974Proceedings. Gulf Coast Regional Symposium onDiseases of Aquatic Animals (eds. R.I.. Amborski.M.A. Hood, and R.R. Miller), pp. 85-113. USU-SG-74-05. Baton Rouge: Center for Wetland Resources,Louisiana Stale University.
QUICK. J.A., JR. AND G.E. HENDERSON. 1975.Evidences of new ichthyoimoxicative phenomena inGyninodinium breve red Iides. In Proceedingsof TheFirst International Conference on Toxic Dinoflagellate Blooms. Nov. 1974, Boston,Mass. (cd. V.R.LoCiccro). pp. 413-422.
QUICK. J.A.. JR. AND J.G. MACKIN. 1971. Oysterparasitism by Labyrinthomyxa marina in Florida.Florida Department of Natural Resources. MarineResearch Laboratory Professional Papers SeriesNo. 13:1-55.
RAGAN. J.G. AND D.V. ALDRICH. 1972. Infection ofbrown shrimp, Penaeus aztecus Ives by Prochristianella penaei Kruse (Cestoda:Trypanorhyntha) in southeastern Louisiana bays.Transactions of the American Fisheries Society10l(2):226-238.
RAGAN. J.G. AND B.A. MATHERNE. 1974. Studiesof Loxothylacus texanus. In 1974 Proceedings, GulfCoast Regional Symposium on Diseases of AquaticAnimals (eds. R.L. Amborski, M.A. Hood, and R.R.Miller), pp. 185-203. LSU-SG-74-05. Baton Rouge:Center for Wetland Resources, Louisiana State
University.
RAY. S.M.. 1954. Biological studies of Dermocystidium marinum. The Rice Institute Pamphlet,Special Issue Monograph in Biology. 114 pp.
READ. C.P.. 1970. Parasitism and Symbiology. AnIntroductory Text. 316 pp. New York: Ronald Press.
REES. C. 1969. ti-slexles Irom Bermuda fishes and anaccount of Acompsocephalum tortum (Linton, 1905)gen. nov. from the lizard fish Synodus intermedins(Agassiz). Parasitology 59(3):519-548.
REIMER. A.A.. 1971. Feeding behavior in iheHawaiian zoanthids Palythoa and Zoanthus.Pacific Science 25(4):5l2-520.
RIDGWAY, S.H. (ed.). 1972. Mammals of the Sea.Biology and Medicine. 812 pp. Springfield: CharlesC Thomas.
RIDGWAY. S.H. AND M.D. DAILEY. 1972. Cerebral
and cerebellar involvement of trematexle parasitesin dolphins and theii possible role in stranding.Journal of Wildlife Diseases 8( 11:33-13.
RICDON. R.H. AND K.N. BAXTER, 1970. Spontaneous necrosis in muse les of brown shrimp. Penaeusaztecus Ives. Transactions of the American FisheriesSociety 99(3):583-587.
RILEY. J.. 1972a. Some observations on the life-cycleol Reighardia sternae Diesing 186-1 (Pentaslomida).Zeitschrift ftir Parasilenkunde 40( I ):49-59.
RILEY. J., 1972b. Histochcmical and uitrastructuralobservations on feeding and digestion in Reighardiasternae (Pentaslomida: Cephalobaenida). Journalof/oology. London 167:307-318.
RILEY. J.. 1973.The structure of the buccal cavity andpharynx in relation to ihe method of feeding ofReighardia sternae Diesing 186-1 (Penlastomida).International Journal for Parasitology 3(2):I49-156.
ROBERTS. L.S.. 1970. Ergasilus (Copepoda: Cyclo-poida): Revision and key to species in NorlhAmerica. Transactions of the American Microscopical Society 89(1): 134-161.
ROBERTS. R.J.. 1975. Melanin-containing cells ofleleost fish and their relation lo disease. In 77ie
Pathology of Fishes (eds. W.E. Ribclin and G.Migaki). pp. 399-428. Madison: University ofWisconsin Press.
ROBINS. C.R.. 1957. Effects of storms on the shallow-
water fish fauna off Southern Florida with new re
cords of fishes from Florida. Bulletin of MarineScience of the Gulf and Caribbean 7(3):266-275.
ROGERS. W.A., 1972. Disease in fish due lo the
protozoan Epistylis (Ciliata: Peril lie ha) in thesoutheastern U.S. Proceedings of the 25th AnnualConference of the Southeastern Association of Gameand Fish Commmioners:493-496.
ROGERS, W.A. AND J.J. GAINES. JR.. 1975. Lesionsof protozoan diseases in fish. In The Pathology ofFishes (eds. W.E. Ribclin and G. Migaki), pp.117-141. Madison: University of Wisconsin Press.
ROHDE. K.. 1977. Si>ccies diversity of monogeneangill parasites ol fish on the Great Barrier Reef. InProceedings of the Third International Coral Reef

Symposium. Vol. 1, Biology, pp. 586-591. Miami.Florida: Rosensiiel School ol Marine and Atmospheric Science.
ROSEN. B.. 1967. Shell disease of ihe blue nab.Callinectes sapidus. Journal of InvertebratePathology 9(3):348-353.
ROSENTHAL. II., 1967. Parasiies in larvae of thehening (Clupea harengus I..) led with wild plankton. Marine Biology 1(1 ):I()-I5.
ROSS. DM.. 1967. Behavioural and ecological tela-•ionships between sea anemones and ciibeiinvertebrates. In Oceanography andMarine BiologyAnnual Review (ed. H. Barnes), 5:291-316. London:George Allen and Unwin.
ROSS. D.M.. 1974.Evolutionary aspects of associationsbetweencrabsand seaanemones. In Symbiosis in theSea (ed. W.B. Vernbcrg). pp. 111-125. Oilumbia:.University of South Carolina Press.
ROTHSCHILD, M. AND T. CLAY. 1957. Fleas,Flukes and Cuckoos. A Study of Bird Parasites.305 pp. New York: MacMillan.
SANDIFER. PA. AND P.J. ELDRIDGE. 1974. Observations on the incidence of shell disease in SouthCarolina blue crabs, Callinectes sapidus. In 1974Proceedings, Gulf Coast Regional Symposium onDiseases of Aquatic Animals (eds. R.I.. Amborski.M.A. Hood, and R.R. Miller), pp. 161-181. I.SU-SG-74-05. Baton Rouge: Center for Wetland Resources, Louisiana State University.
SASTRY. A.N. AND R.W. MENZEL. 1962. Influence
of hosts on the behavior of the commensal crab
Pinnotheres maculatus Say. Biological Bulletin123(2):388-395.
SAWYER. T.K.. G.L. HOFFMAN. J.G. HNATH.AND J.F. CONRAD. 1975. Infection of salmonidfish gills by aquatic ami has (Amoebida: Thecamoe-bidae). In 77ie Pathology of Fishes (eds. W.E.Ribelin and G. Migaki). pp. 143-150. Madison:University of Wisconsin Press.
SAWYER. R.T., A.R. LAWLER, AND R.M.
OVERSTREET, 1975. Marine leeches of the eastern
United States and the Gulf of Mexico wilh a key tothe species. Journal of Natural History 9(6):633-667.
SCHELL. S.C. 1970. How to Know the Trematodes.
355 pp. Dubuque: William C. Brown.
SCHMIDT. CD.. 1970. How toKnowthe Tapeworms.266 pp. Dubuque: William C Brown.
SCHMIDT. CD. AND L.S. ROBERTS, W7.Foundations of Parasitology. 60-1 pp. St. Louis: CV.Mosby.
SCHRODER. J.H. (ed.). 1973. Genetics andMutagenesis of Fish. 356 pp. New York: Springer-Verlag.
SCHULTZ. G.A.. 1969. How to Know the MarineIsopod Crustaceans. 359 pp. Dubuque: William C.Brown.
SEBENS. K.P.. 1977. Autotrophic and heterotrophicnutrition of coral reef zoanthids. In Proceedings,Third International CoralReefSymposium. Vol. 1,Biology, pp. 397-404. Miami, Florida: Rosenstiel
Literature' Cited 135
School of Marine and Atmospheric- Science.
SELF, J.T., I969. Biological relationships of thePi-uiasiomida: a bibliography on the- Pt-niastomida.Experimental Parasitology 24(l):63-l 19.
SHERWOOD, M.J. AND A.J. MEARNS. 1977.Environmental significance of fin erosion insouthern California demersal fishes. In AquaticPollutants and Biologic Effects with Emphasis onNeoplasia (eds. H.F. Kraybill. C.J. Dawe, J.C.Harshbarger. and R.C Tardiff). Annals of The NewYork Academy of Sciences 298:177-189.
SHIELDS, R.J., 1970. Rate of development of themarine parasitic copepod, Lemaeenicus radiatus.The Journal of Parasitology 56(4, Sect. 2, Pt. 1):316.
SHOTTS, E.B.. JR., 1976. Rapid diagnosticapproaches in the identification of gram-negativebacterial diseases of fish. Fish Pathology 10(2):187-190.
SHULMAN, S.S., 1966. Myxosporidia in the Fauna ofU.S.S.R. 507 pp. Moscow: U.S.S.R. Academy ofScience (In Russian).
SHULMAN. S.S. AND G.A. SHTEIN. 1962. Protozoa.
In Key to Parasites of Freshwater Fish of the U.S.S.R.pp. 5-236. (Translated from Russian, 1964).Jerusalem: Israel Program for Scientific Translations.
SIBLEY. CC AND J.E. AHLQUIST. 1972. Acomparative study of the egg white proteins of non-passerine birds. Peabody Museum of NaturalHistory, Yale University, Bulletin 39:1-276.
SINDERMANN, C.J.. 1970. Principal Diseases ofMarine Fish and Shellfish. 369 pp. New York:Academic Press.
SINDERMANN. C.J.. 1971. Internal defenses ofCrustacea: a review. Fishery Bulletin 69(3):455-489.
SINDERMANN. CJ. (ed.), 1977. Disease Diagnosisand Control in North American Marine Aquaculture. Developments in Aquaculture and FisheriesScience, 6. 329 pp. Amsterdam: Elsevier Scientific.
SINDERMANN, C.J. AND A ROSENFIELD, 1967.Principal diseases of commercially importantmarine bivalve Mollusca and Crustacea. FisheryBulletin 66(2):335-385.
SIZEMORE, R.K.. R.R. COLWELL. U.S. TUBIASH.
AND T.E. LOVELACE. 1975. Bacterial flora of the
hemolymph of the blue crab, Callinectes sapidus:numerical taxonomy. Applied Microbiology 29(3):393-399.
SKRJABIN, K.I. (ed.). 1949. AVy to ParasiticNematodes. Vol. I Spirurata and Filariata. 497 pp.Leningrad: Academy of Sciences of ihe USSR,Helminthological Laboratory ('Translated and
edited by M. Raveh, Israel Program for Scientific-Translations. 1968).
SLEIGH. M.A.. 1973. The Biology of Protozoa. 315 pp.New York: American Elsevier.
SMITH, D.C., 1973. Symbiosis of algae with invertebrates. In Oxford Biology Readers No. 43 (eds. J.J.Head and O.E. Lowenstein), 16 pp. London: OxfordUniversity Press.

136 Marine Maladies?
SOGANDARES-BERNAL. F. AND JR. SEED. 1973.Aineilean paragonimiasis. In Current Topics mComparative Pathobiology. Vol. 2 (ed. T.C. Cheng),pp. 1-56. New York: Academic Pre-ss.
SOLANGI. M.A. AND D.V. LIGHTNER. 1976.
C-llulat inflammatory icsixinsc ol Penaeus aztecusanil /'. setiferus to ihe pallioginii liuigus. Fu.sariumsp.. isolated from the California blown shrimp./'. laliforniensis. Journal of Invertebrate Pathology•lit I ):77-86.
SPARKS. A.K. AND C.T. FOX TAINE. 1973. Host
H's|h>iisi- in the white shiimp. Penaeus setiferus. tomice lion by ilie laival mpaiioihyic hid ee-slode.Prtu hrisliciiiella penaei. Journal of InvertebratePathology 22(21:213-219.
.SPRAGUE. V.. 1954. Pioto/oa. In Gulf of Mexico. ItsOrigin. Welters, and Marine Life. Fishery Bulletin ofol the Fish and Wildlife Service 55:213-256.
SPRAGUE. V.. 1970a. Some piotozoan paiasiie-s andhvpe-ipaiasiii-s in marine decapod Crustacea. In ASymposium on Diseases <>/ Fishes and Shellfishes(ed. S.F. Snies/ko). American Fisheries SocietySpecial Publication No. 5:116-130.
SPRAGUE. V . 1970b. Some piotozoan paiasites andhxpeipaiasiles ill marine bivalve molluscs. In ASymposium on Diseases of Fishes and Shellfishes(e-d. S.F. Suii-s/ko). American Fisheries SocietySpecial Publication No. 5:511-526.
SPRAGUE. V.. 1971. Diseases of oysieis. AnnualReview of Mic robiology 25:211-230.
SPRAGUE. V.. R.L. BECKETT. ANDT.K. SAWYER.
1969. A new s|x-cics ol Paramoeba (Anux-bida.Paiaiiioc'biclac) paiasilic in die crab Callinectessalndu.s. Journal of Inveitebrate Pathology 14(2):167-171.
SPRAGUE. V. AND J. COUCH. 1971. An aimoiaicdlist of piolo/oan parasites, hyperparasites. andcommensals of decapod Ciustacca. 77ie Journal ofProtozoology 18(3 ):526-537.
STREET. D.A. AND V. SPRAGUE. 1974. A new
species of Pleistophora (Microsporida: Pleisto-pheiiidae) patasilit in the shrimp Palaemonetespugio. Journal of Invertebrate Pathology 23(2):153-156.
SUMMERS. M.D.. 1977. Chaiacii-iization of shrimpbaculovirus. EPA-600 3-77-130. Ft ologicalResearch Series. Environmental Research Labora
tory. Gulf Breeie. Florida. 36 pp.
SWEENEY. J.t;. AND S.H. RIDGWAY. 1975.Common diseases of small cetaceans. Journal of theAmerican Veterinary Medical Association 167(7):533-540.
TACATZ. M.E. AND A.B. HALL. 1971. Annotated
bibliography on the fishing industry and biology ofihe blue etab. Callinectes sapidus. NO AA. TechnicalReport NMFS SSRF 640:1-94.
TAYLOR. D.l... 1974. Symbiotic marine algae:taxonomy and biological fitness. In Symbiosis inthe Sea (ed. W.B. Vernberg). pp. 245-262. Columbia:University of South Carolina Press.
TAYLOR. C.R.R..STICKNEY.AXDR.HEARD.nl.
1973. Two anomalous flounders (Boihidae. Elropuscrossotus) from Georgia esiuarine waters.Chesapeake Science I4(2):I47.
TIFFANY. W.J.. III. 1968. The Life Cycle and Ecologyof the Beach Clam Donax variabilis Say (Mollusta:Pelecypoda: Donacidae). M.S. thesis. Florida StaleUniversity. Tallahassee. 79 pp.
TRENCH. R.K.. 1974. Nutritional potentials inZoanthus sotiatus (Coc-lc-iiu-iaia. Anthozoa).
Helgotander Wissenschaflliche Meeresuntersuchun-gen 26(2):174-216.
TRENT. 1... E.J. PUI.I.EN, AND R. PROCTOR.1976. Abundance of macrocrustaccans in a natural
marsh and a marsh altered by dredging, bulk-heading, and filling. Fishery Bulletin 74(1): 195-200.
UNESTAM. T.. 1973. Fungal diseases of Crustacea.Review of Medical & Veterinary Mycology 8(1):I-20.
YANDERZANT. C. E. MROZ. AND R.
MCKEI.SON. 1970. Microbial flora of Gull ol
Mexico and pond shrimp. Journal of Milk and FoodTechnology SSWMli-VM.
VAN ENGEL. W.A.. W.A. DILLON. D.ZW'ERNER.
ANDD. ELDRIDGE. 1966. Loxothylacus panopaei(Cirri|M-dia. Sacciilinidae) an introduced parasite ona xaiiibid crab in Chesapeake Bay. U.S.A. Crusta-tecum I(i(l):lll)-I12.
VENARD. CE. AND R.V. BANGHAM. l941..S>eVAicj
oxycephala (Peniastomida) from Florida fishes andsome notes on die morphology of the larvae. OhioJournal of Science 41(l):23-27.
WALLS. J.t;.. 1975. Fishes of the Northern Gulf ofMexico. 432 pp. Xepiune Cily. New Jersey: T.F.H.Publications.
WARDl.E. R.A. AND J.A. McLEOD. 1952. TheZoology of Tapeworms. 780 pp. Minneapolis:University of Minnesota Press.
WATERTOR. J.L.. 1973. Incidence- of Hirudmellamarina Gaicin. 1730 ( Tii-inaicxla: Hinuliiu llidac)
in tunas hom die- Atlantic Ocean. 77ie Journal ofParasitology 59( I ):207-208.
WEDEMEYER. G.A.. F.P. MEYER. AND L. SMI TIL
1976. Environmental Stress and Fish Diseases.
Book V In Diseases of Fishes (eds. S.F. Snieszko andH.R. Axehotl). 192 pp. Neptune Cily. New Jersey:T.F.H. Publications.
WEIDNER. E.. 1970. Uhrastructtiral study of mieio-sixuidian development. [.Nosema sp. Sprague. 1965in Callinectes sapidus Rathbun. Zeilschrifl furZellforschung undMikroshopische Anatomic 105(1):33-54.
WEIDNER. E.. 1972. Uitrastructural study of ruiero-sporidian invasion into cells. Zeilschrifl furParasitenkwide 40(3):227-242.
WELLS. 11.W.. M.J. WELLS. AND I.E. GRAY. I960.Marine sponges of North Carolina. Journal of IheElisha Mitchell Scientific Society 76(2):200-245.
WICKHAM. D.A. AND F.C. M1NKI.ER. HI. 1975.
Laboratory observations on daily patients ol

burrowing and locomotor activity of pink shrimp.Penaeus duorarum. brown shrimp. Penaeus aztecus,and while shrimp, Penaeus seliferus. Contributionsin Marine Science 19:21-35.
WILKIE, D.W. AND H. GORDIN. 1969. Outbreak of
cryplocaryoniasis in marine aquaria at Scripps In-siiiuiion of Oceanography. California Fish andGame 55(3):227-236.
WILLIAMS. A.B., 1965. Marine decapod crusiaccansofthe Carolinas. Fishery Bulletin 65(l):l-298.
WILLIAMS, A.B., 1974. The swimming crabs ol ihegenus Callinectes (Dccapcxla: Poriunidae). FisheryBulletin 72(3):685-798.
WILLIAMS, E.H.. JR. AND W.A. ROGERS. 1971.Two new species of Gyrodactylus ('Trematoda:Monogenea) and a redescriplion and new host iccoidlor G. prolongis Hargis. 1955. The Journal ofParasitology 57(4):845-847.
WILLIAMS. H. AND A. JONES, 1978. MarineHelminths and Human Health. Commonwealth
Institute of Helminthology, CommonwealthAgricultural Bureau. CIH Miscellaneous Publication No. 3:1-17.
WOBESER. C. G.R. JOHNSON. AND G.ACOMPANADO. 1974. Stomatitis in a juvenile-white pelican due lo Piagetiella peraiis (Mallophaga:Meno|)onidae). Journal of Wildlife Diseases 10(2):135-138.
WOLF. K. AND M.E. MARKIW. 1976. Myxosomacerebralis: In vitro spotulalion ol the myxosporidanof salmonid whirling disease. 77m* Journal of Protozoology 23(31:425-427.
Glossary 137
YAMAGUTI. S.. 1959. Syslema llelmiiitliiini. Vol.II. The Cestodes of Vertebrates. 860 pp. New York:Interscience.
YAMAGUTI. S.. \W\\. Syslema Helminlhum. Vol. 111.The Nematodes of Vertebrates. 2 Pans. 1261 pp.New York: Interscience.
YAMAGUTI. S.. 1963a. Syslema Helminlhum.Vol. IV. Monogenea and Aspidocolylea. 699 pp.New York: Inleiseienee.
YAMAGUTI. S.. 1963b. .Syslema Helminlhum.Vol. V. Acctiilhocephitla. 423 pp. New York:Inleiseienee-.
YAMAGUTI. S.. 1963c. Parasitic Copepoda andBranchiura of Fishes. 1104 pp. New York: Interscience.
YAMAGUTI. S.. 1971. Synopsis of Digenetic Trematodes of Vertebrates. Vols. ML 1074 pp.. 349 pis.Tokyo: Keigaku.
YAMAGUTI. S., 1975. A Synoptical Review of LifeHistories of Digenetic Trematodes of Vertebrateswilh Special Reference to the Morphology of theirLarval Forms. 590 pp.. 219 pis. Tokyo: Keigaku.
ZAM. S.G.. D.K. CALDWELL. AND M.C
CALDWELL. 1971. Some endoiiarasitcs horn small
odontocele cetaceans collected in Florida and
Georgia. Cetology 2:1-11.
ZANGERL, R., 1973. Interrelationships eif earlychoiidiie luliyans. In Interrelationships of Fishes(eds. P.II. Gree-nwood, R.S. Miles.and CPatierson),Zoological Journal of the Linnean Society 53(Suppl. I):l-I4.
GLOSSARY
The following definitions are included toaid those not familiar with technical terms.
One desiring more information on specificterms dealing with parasites should referto one of the cited works or some other
treatment of the group in question. Toolarge a number of vertebrate and invertebrate host-groups are mentioned in the textto discuss or classify here; therefore, a readerwanting more information on a host shouldexamine an encyclo|X"dia. a book on ihegeneral group in c,uestion, or specificpapers, several of which have been cited inthe text.
apron of crab - The thin abdomen which isfolded forward on the crab's underside;it is sometimes called the tail.
benthic - Living in. living on. or referring toihe bottom.
carapace - The hard outer protective covering of some animals. In crustaceans, itcovers the head and gill region.
chitin - A skeletal material of arthropodsand other invertebrates that decomjDosesto specific chemicals (glucosamine andacetic acid), if boiled with concentratedhydrochloric acid. When impregnatedwith calcium salts, it may help form afirm exoskeleton.
chlorophyll - Any of several pigmentscapable of jjhotosynthesis. They providethe green and yellow colors of manyplants.
chloroplast - The structure containingchlorophyll embedded in a cell.
chromatophore - A pigment-containing cellthat by exjjansion or contraction canchange the overall color of an organism.

138 Marine Maladies?
community (biotic) - The group of speciesof plants and animals living in and characteristic of a given habitat.
cuticle - The noncellular (therefore nonliving), usually horny, protective outercovering of many invertebrates producedby the animal.
disease - z\n abnormal condition of an
organism or part that impairs normalphysiological functioning.
estuary - The brackish water regions of andnear river mouths influenced by tides.
fibroblast - A connective-tissue cell that canproduce collagen, the primary component of scar tissue.
fish)fishes - "Fish" may be singular for aspecies or an individual or plural whenreferring to more than one individual ofone or more species. "Fishes" is usedwhen referring to two or more species,disregarding the number of individuals.The term for a group of individuals comprising a variety of species used herein is"fish." but some ichthyologists prefer theterm "fishes."
free-living - An organism or stage notsymbiotic with a host; the organism maybe parasitic during one stage in its lifehistory and free-living in another.
gamete -The mature sperm or ovum capableof participating in fertilization.
hemoglobin - One of several pigmentsused to transport oxygen, consisting ofaniron-complex coupled to a protein. Itoccurs in the red blood cells of most
vertebrates and in body fluids or corpuscles of a variety of nonrelatedinvertebrates.
hemolymph!hemocytes - Hemolymph isthe circulatory fluid that bathes tissues ofsome invertebrates that do not have the
closed circulatory system found in vertebrates. It is functionally comparable toblood and lymph of vertebrates. Theblood cells are called hemocytes.
host-specificity - The degree to which aparasite can mature in more than onehost species. If a parasite infects only asingle species, it is highly host-specific.
humoral-cellular mechanisms - When re
ferring to immunity, responses are bothblood-mediated (humoral) and cell-mediated. Antigenic responses involveboth systems.
hyaline - Refers to being translucent ortransparent without the presence of fibersor granules.
hyperparasite - An organism which parasitizes another parasite.
hypha/mycelium - The mycelium is thevegetative filamentous stage of a fungus,and an individual branch is called a
hypha.
inclusion body - A particulate body foundin cells of tissue infected with a virus.
infection!infestation - An infection is theinvasion or state resulting from an invasion by a parasite or a pathogen into ahost. An infestation is an external rather
than internal invasion. If both internal
and external organisms occur on one host,all the associations are collectively referred to as infections. Some biologists, however, restrict the term "infestation" to
either internal or external associations
with metazoans. Neither an infection nor
an infestation necessarily implies disease.
instar - A larval stage of an insect characterized by the animal's size and form betweenmolts.
intermediate host - A host in which a larval
parasite develops, but not to sexualmaturity.
long-line - A fishing line usually maintained by an anchor and buoy withnumerous, spaced, hooked lines.
lorica - A secreted noncellular (thereforenon-living) protective case. The examplesin the text all involve ciliate protozoans.
measurements - For those confused about
metric units, the following aids mighthelp.
multiplied bythe following
number is
centimeters
inches
feet
0.3937 inches
2.54 centimeters
30.48 centimeters

liters 0.2642 gallonsgallons 3.785 liters
1 meter = 100 centimeters (cm) =39.37 inches
1 cm =10 millimeters (mm) =0.394 inches
1 mm = 1000 microns (u)0.039 inches
The diameter of a human headhair varies
among individuals; those of a few friendsrange from 60 to 80 // wide.
A common wooden pencil is about 7 mm(5/32 inch) wide.
A penny is 19mm wide by about 1mm thick.
A stick of gum is 75 mm (7.5 cm or about3 inches) long.
A 10 ounce (about 0.3 liter) Coca-Cola bottleis 24.5 cm (9 21/32 inches) high.
metabolic - Pertaining to the complex ofphysical and chemical processes involvedin maintaining life.
necrotic - Refers to the pathologic death ofliving tissue while still on a living body asopposed to that which continually diesand is sloughed.
niche - The position or status of anorganism within its community resultingfrom structural adaptations, physiological responses, and innate or learnedbehavior.
photosynthesis - The synthesis of simpleorganic matter from carbon dioxide andwater, using light as the energy source.
phylogenetic- Referring to theevolutionarydevelopment of a plant or animal.
phylum (pi. -la) - A primary taxonomicdivision of the animal (or plant) kingdom. As an example, snails, clams,chitons, and octopus all belong todifferent classes in the phylum Mollusca.
plankton- The floating and drifting aquaticorganisms whose primary movementsresult from water movement rather than
their swimming efforts.
prepotent period - That period betweeninvasion by a disease-agent and demonstration of disease or a parasite in the body(such as by eggs or cysts in feces).
Glossary 139
prevalence! incidence - Prevalence is thedegree to which a disease or infectiveagent occurs. It is often measured as thepercentage of a population affected with aparticular organism at a given time. Manybiologists define incidence identically.More precisely, the term "incidence"indicates the rate of occurrence of new-
cases of a particular infection in apopulation.
reservoir host - An organism serving as asource of infection for a specific organismby harboring the infectious agent.
resistance - The ability of an organism towithstand the effects of physical, chemical, and biological agents which mightotherwise debilitate it.
salinity - The sum of the various saltcomponents in seawater usually represented as parts per thousand (ppt). Openocean salinities remain about 35 ppt (3.5%salt), and in Mississippi Sound, salinityvalues usually range between 6 and 15ppt depending primarily on the directionand force of the wind.
sclerotized - Hardened by substances otherthan chitin.
serum!plasma - Plasma is the watery fluidcontaining salts, proteins, and otherorganic compounds in which blood cellsare suspended. After the blood clots andloses many constituents, the clearremaining fluid is serum.
sign'symptom - A sign is any objectiveevidence of a disease or disorder, asopposed to a symptom, which is thesubjective complaint of a patient.
sp. - Abbreviation for species, and whenused in conjunction with a generic name,it indicates the identification of the
species is unknown or uncertain. If morethan one species of a genus is beingreferred to, spp. is used.
spent - Referred to in above context asdepleted of most reproductive products.
spore - A term defining a different structurefor different organisms. Typically asingle - or few-celled resting or resistantstage produced asexually.

140 Marine Maladies?
substrate/substratum (pis. -tes, -ta) - Asubstrate is the material or substance
upon which a chemical enzyme acts,whereas a substratum is the underlyingmaterial on which an organism grows orattaches.
systemic - Pertaining to the entire body,such as an infection with a bacterium
dispersed through the body by the bloodstream.
tumor'cancer - "Tumor" refers to a
swelling serving no physiological function; it may be a neoplasm, a parasiticoutgrowth, or any other growth. Aneoplasm may be benign or malignant,the latter being characterized by its abilityto invade tissue and metastasize to newsites. Typically, only a malignantneoplasm is referred to as a cancer.
vector - An invertebrate or in some cases
. wind, water, or other agents whichtransmit a parasite or disease organism,usually to a vertebrate recipient. Ifdevelopment of a parasite occurs, the hostis called an intermediate host unless
development includes sexual reproduction and then it is called an
invertebrate host. Man is actually theintermediate host for malaria organismswhich have their sexual stage inmosquitoes.
zooid - An individual forming part of acolony in the Protozoa, Cnidaria, andother groups. A zooid can specialize inreproduction, feeding, food capture,protection, or locomotion.
zoonosis - A disease of invertebrates or
vertebrates other than man which may betransmitted to man.





























