Embed Size (px)
Citation preview

J. evol. Biol. 2: 317-338 (1989) 101&061X/89/05317-22 $ 1.50 +0.20/O 0 1989 Birkhtbser Verlag, Base1
What controls haploid-diploid ratio in the red alga, Gracilaria verrucosa?
Christophe Destombe, Myriam Valero ‘, Philippe Vernet’ and Denis Couvet*
Laboratoire d’Algologie et de Biologie Vegetale Marine, Equipe de Biologie des Populations, Universite des Sciences et Techniques de Lille Flandres-Artois, 59655 Villeneuve d’dscq cedex, France. ‘Laboratoire de Genetique Ecologique et de Biologie des Populations Vegetales, UA-CNRS 1185. Universite des Sciences et Techniques de Lille, 59665 Villeneuve d’Ascq cedex, France. *B2P-CNRS Route de Mende, B.P. 5051, 34033 Montpellier cedex, France
Key words: Haploiddiploid ratio; alternation of generations; life cycle variations; isomorphic red algae; population demography.
Abstract
The conditions for maintenance of a haploiddiploid life cycle in the species Gracilaria verrucosa were studied. This species is a red alga, where haploid plants have separate sexes. In the two natural populations studied, male and female haploid individuals were in equal proportions, and the frequency of diploid individuals reached 0.5.
A two-fold advantage in viability for diploid relative to haploid juveniles was observed in the field. This advantage can account for a frequency of 0.5 of diploid individuals in natural populations. Different types of anomalies in the reproduction of diploid individuals were observed, all of which lead to a reduction of the haploid stage.
Introduction
The reproductive cycle of eukaryotic individuals is generally characterized by an alternation of generations between a haploid state and diploid state. These two phases vary in structure and in duration among different kinds of organisms. The diploid and haploid phases are usually very different morphologically and differ greatly in their exposure to selective factors (Wright, 1968). In most organisms, one of the two phases, haploid or diploid, is predominant during the life cycle. Higher plants and animals are diploids with only brief haploid interludes. To attempt to answer the question why, in most species, haploid individuals or diploid individuals
317

318 Destombe et al.
are predominant, it may be especially relevant to look at haploiddiploid species such as exist in many red algae and some fungi, where the haploid and diploid stages are independent plants. Indeed, population studies of such species can furnish data about the range of variations of the haploiddiploid ratio (in terms of number and time) and permit one to investigate the evolutionary patterns from haploidy to diploidy.
The life history of the red alga is diplohaplontic (Fig. 1) with haploid (gameto- phyte) and diploid (tetrasporophyte) plants. In the genus Gracilaria gametophytes are dioecious; they are either male or female. Fertilization occurs on female individuals. After fertilization, one brief generation (carposporophyte) develops on the female haploid plant. The zygote undergoes successive mitoses to produce a gonimoblast from which many thousands of identical diploid spores are formed. They are liberated in a mucilage and give rise to independent diploid plants. Meiosis occurs in specialized cells (tetrasporocysts) produced by these plants: many haploid spores are liberated and germinate to produce independent haploid individuals (either male or female gametophytes). Van der Meer and Todd (1977) have shown that the sex is controlled by one mendelian locus (heterozygotic in diploid tetrasporophyte).
The species studied Graciluria verrucosu (Hudson) Papenfuss is found off the coast of France and shows this typical life history: the “Polysiphoniu type” of Yamanouchi ( 1906) (Fig. 1). Both haploid individuals (gametophyte) and diploid individuals (tetrasporophyte) have the same morphology and are perennial. In such
Fig. 1. Life cycle of Gracilariu verrucosa. m = mitosis, M = meiosis, F = fertilization, 2 = zygote. Thalli and reproductive organs of each type of individual are described Fig. 2. Erect thalli of haploid
gametophytes (male and female) and diploid tetrasporophytes are isomorph. After fertilization (F) the carposporophyte (cystocarp) develops on the female haploid gametophyte. The zygote (Z) undergoes successive mitoses (m) to produce many thousand of diploid spores. Each diploid spore gives a tetrasporophytic plant. Meiosis (M) occurs on this plant. Many thousand of haploid spores are liberated
and germinate to produce male or female independent haploid gametophytes.

What controls haploid4iploid ratio? 319
cases a comparison of haploidy and diploidy will not be confused by other unrelated differences of morphology or physiology.
The evolutionary significance of this three-phase life cycle has received very little attention. Searles (1980) considered that the function of the “small generation” (carposporophyte) which gives many thousands of spores from only one zygote is to compensate for the difficulties of fertilization (due to the lack of motile male gametes).
In this study of Gracilaria verrucosa we attempt to answer the following questions: 1. What are the relative frequencies of haploid and diploid plants in natural
populations? 2. What is the relative survivorship of haploid and diploid individuals at the
juvenile and adult stages? 3. Are the observed frequencies of haploid and diploid plants in agreement with
their expected frequencies within populations, calculated on the basis of their estimated fertility (Destombe, 1987) and survivorship?
Material
The individual plant of G. verrucosa consists of a perennial holdfast fixed to a rocky substrate that produces annually numerous erect thalli. When the individual is not reproducing, i. e. has no erect thalli with reproductive organs, it is called vegetative. The different types of individuals (haploid male and female gametophytes and diploid tetrasporophytes) can be recognized by cytological observation of the thallus during the reproductive period (Fig. 2). The female gametophyte is often identified only after fertilization, when the “fruit” (cystocarp) is formed. The chromosomes of G. gracilaria are very tiny and difficult to observe (Magne, 1964). On the one hand, for two tetrasporophytes sampled in our studied sites, Maggs (unpublished data) observed 32 chromosomes in mitosis of the diploid vegetative cells, and 16 chromosomes in tetraspores (haploid cells resulting from meiosis). On the other hand, Godin et al. (in prep.) have measured the ploidy level, using two different methods, on individuals from our studied populations. First, the nuclear DNA content of 10 individuals (5 haploids and 5 diploids) was measured using a cytofluorometric method described by Hull et al. (1982). They found two times more nuclear DNA content in diploids (tetrasporophytes) than in haploids (male and female gametophytes). Secondly, the number of chromosomes were counted in mitosis preparations for microscopic observation. In three female gametophytes, 18 chromosomes were counted in the gametophytic tissues and 36 chromosomes in the diploid gonimoblast resulting from fertilization. In two tetrasporophytes, they counted 36 chromosomes in the tetrasporophytic tissues and 18 chromosomes in the haploid tetraspores resulting from meiosis.
Study sites
Populations of G. verrucosa are found in rock pools at the lower end of the intertidal zone. The individuals occur aggregated in pools that are exposed only at
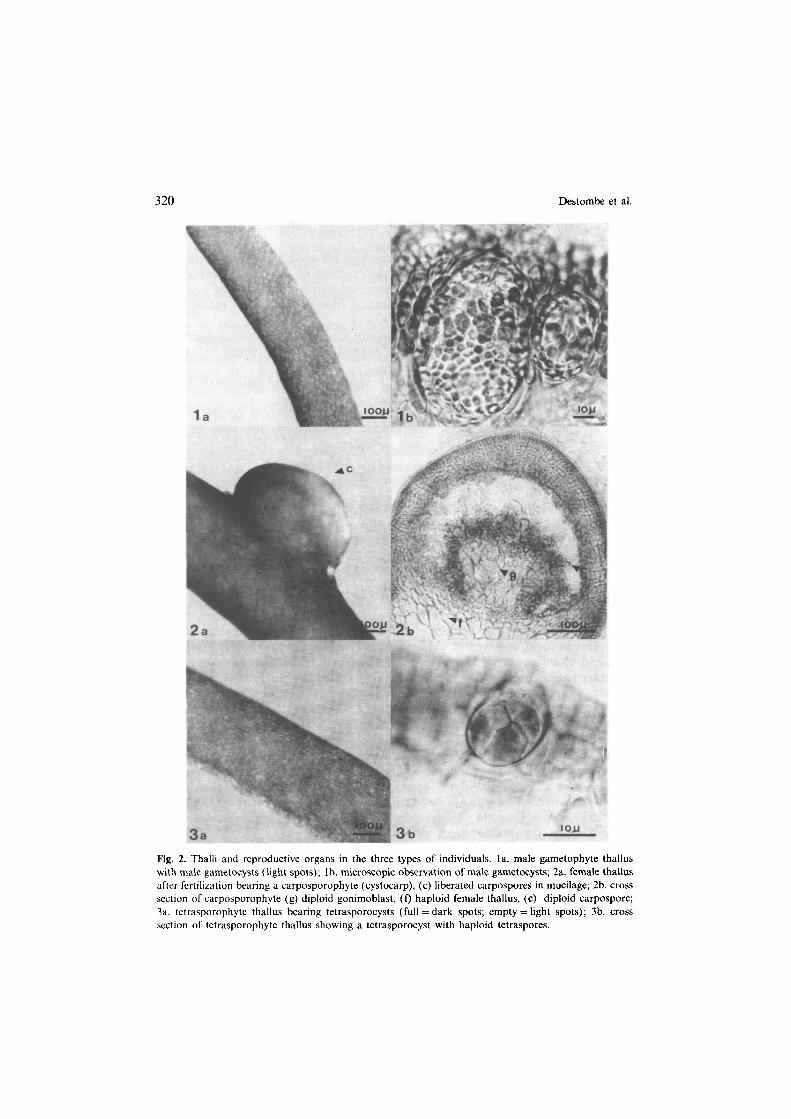
320 Destombe et al.
Fig. 2. Thalli and reproductive organs in the three types of individuals. la. male gametophyte thallus with male gametocysts (light spots); lb. microscopic observation of male gametocysts; 2a. female thallus
after fertilization bearing a carposporophyte (cystocarp), (c) liberated carpospores in mucilage; 2b. cross section of carposporophyte (g) diploid gonimoblast, (f) haploid female thallus, (c) diploid carpospore; 3a. tetrasporophyte thallus bearing tetrasporocysts (full = dark spots; empty = light spots); 3b. cross section of tetrasporophyte thallus showing a tetrasporocyst with haploid tetraspores.
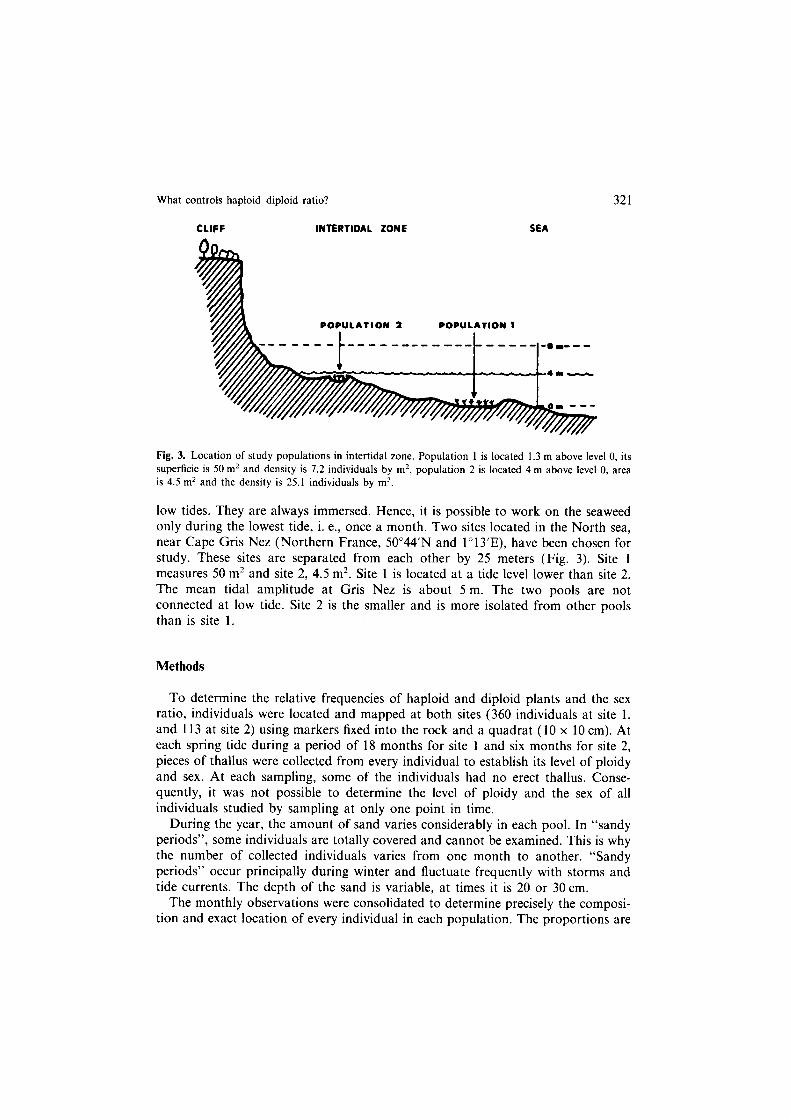
What controls haploid+liploid ratio? 321
Fig. 3. Location of study populations in intertidal zone. Population 1 is located 1.3 m above level 0, its
superficie is 50 mz and density is 7.2 individuals by m’, population 2 is located 4 m above level 0, area is 4.5 mz and the density is 25.1 individuals by m2.
low tides. They are always immersed. Hence, it is possible to work on the seaweed only during the lowest tide, i. e., once a month. Two sites located in the North sea, near Cape Gris Nez (Northern France, 50”44’N and 1 o 13’E), have been chosen for study. These sites are separated from each other by 25 meters (Fig. 3). Site 1 measures 50 m2 and site 2, 4.5 m2. Site 1 is located at a tide level lower than site 2. The mean tidal amplitude at Gris Nez is about 5 m. The two pools are not connected at low tide. Site 2 is the smaller and is more isolated from other pools than is site 1.
Methods
To determine the relative frequencies of haploid and diploid plants and the sex ratio, individuals were located and mapped at both sites (360 individuals at site 1, and 113 at site 2) using markers fixed into the rock and a quadrat ( 10 x 10 cm). At each spring tide during a period of 18 months for site 1 and six months for site 2, pieces of thallus were collected from every individual to establish its level of ploidy and sex. At each sampling, some of the individuals had no erect thallus. Conse- quently, it was not possible to determine the level of ploidy and the sex of all individuals studied by sampling at only one point in time.
During the year, the amount of sand varies considerably in each pool. In “sandy periods”, some individuals are totally covered and cannot be examined. This is why the number of collected individuals varies from one month to another. “Sandy periods” occur principally during winter and fluctuate frequently with storms and tide currents. The depth of the sand is variable, at times it is 20 or 30 cm.
The monthly observations were consolidated to determine precisely the composi- tion and exact location of every individual in each population. The proportions are

322 Destombe et al.
therefore calculated with regard to the total number of individuals present in populations.
To estimate the relative survivorship of haploids and diploids the viability was assessed in vitro, on juvenile individuals, and in the field, on juvenile and adult individuals.
To collect spores, plants of G. verrucosa were collected at the end of July at Cap Gris Nez. Five tetrasporophytic plants and five female plants with car- posporophytes were washed repeatedly in sterile sea water and then placed in plastic petri dishes filled with filtered sea water (0.4 pm) in a culture chamber at 19°C and 40~E.rn~*.s-’ under cool white fluorescent lights with a photoperiod of 12 : 12 h LD. The water in the petri dish was changed once a day during the first week, then weekly for the next month. Spores were shed in abundance after a few days.
The viability of haploid and diploid juveniles was compared in three successive experiments; the juveniles go through two successive stages: (i) as floating spores (that is, spores after liberation and before fixation on the substrate) and then (ii) as fixed spores (that is, after fixation on the substrate, when they start dividing to give juvenile individuals). To estimate the relative survivorship of haploid and diploid spores, the number of spores which remained in the petri dish was counted (dead spores disintegrated) and the number of living spores in the same dish was counted 24 h afterwards, and again 10 days later.
To estimate the relative survivorship of juvenile haploid and diploid individuals developed from fixed spores in vitro, the filtered sea water in the petri dishes was replaced by an artificial sea water (ASP6F modified by Bodard 1973). The number of juveniles still alive was counted periodically at the bottom of the dish and the artificial sea water was changed twice a week.
To estimate the relative survivorship of haploid and diploid young individuals in the field, spores obtained in vitro and fixed on artificial substrates (glass slides) were transplanted, 24 h after the spores had been emitted, from the laboratory to the field. The number of fixed haploid spores was extrapolated from the count on one square centimeter of a microscope slide. The number of fixed diploid spores was counted. Ten slides of tetraspores and thirteen slides of carpospores were studied. The culture slides (25 mm x 76 mm) were fixed, at random, on a PVC slide-holder attached to a breeze-block (Dion and Delepine, 1983) located near site 1. The number of young plants able to survive after one year in situ was counted.
Results
Relative frequencies of haploid and diploid individuals
Because sexual phenotypes may change over the season, because sexual maturity may occur at different times and because there is regular silting-over of individu- als, frequencies of haploid and diploid individuals vary each month.

What controls haploiddiploid ratio?
Proportion of vegetative individuals
323
In both populations the proportion of vegetative individuals was not constant over time (Fig. 4). The maximum number of vegetative individuals was obtained in May and the minimum in September. Nevertheless, we have never observed a total absence nor an exclusive presence of vegetative individuals in either population. Some individuals remained vegetative during all the observations; the frequency of such vegetative individuals is significantly greater in population 1 than in popula- tion 2 (Table 1: x2 = 6.71, 1 d.J, P -c 0.01).
Proportion of fertile individuals; haploidjdiploid ratio and sex ratio The proportion of fertile haploid and diploid individuals observed in both
populations over time is given in Fig. 4. For haploid and diploid plants, the reproductive period was primarily from July to December 1985 in both populations. In 1984, when we studied only population 1, the reproductive period occurred at about the same period (August-January). The ratio haploid/diploid is not signifi- cantly different from 1 : 1 (P > 0.05) in either population, except in April 1985 for population 1 (x2 = 3.94, 1 d.f., P < 0.05) and December for population 2 (x2 = 6.09, 1 d.f., P < 0.02), where haploids were significantly more numerous than diploids.
In both populations, the sex ratio is roughly 1 : 1 (Fig. 4). Only one sample, in November 1985, showed in site 1 a significant deviation from a 1 : 1 sex ratio with a deficiency of male individuals (x2 = 27.50, 1 d.f., P < 0.001).
Fig. 4. Monthly number of collected and vegetative individuals, haploid and diploid individuals and male and female individuals. Time scale: monthly records from August 1984 to February 1986 (noted: 2.86). (C) collected individuals, (V) vegetative individuals, (H) haploid individuals, (D) diploid individ- uals, (F) female individuals, (M) male individuals, (0) significantly different (P < 0.05).

Tabl
e 1.
Ha
ploi
ds,
dipl
oids
an
d se
x ra
tio
in
both
po
pula
tions
hapl
oid
dipl
oid
vege
tativ
e N
dipl
oid
frequ
ency
+
num
ber
%
POP.
l
fem
ale
79
21.9
”
mal
e
15
20.8
”
tetra
.
122 33
.9’
R.S.
P.
23 6.4b
61
17
”
num
ber
%
154
145
61
360
0.48
5’
(0.0
57)1
42.8
’ 40
.2’
17a
num
ber
%
POP.
2
27
32
41
5 8
23.9
a 28
.3”r
36.3
” 4.
4b
7.1’
num
ber
%
59
46
8 11
3 0.
438’
(0.0
96)*
52.2
’ 40
.7’
7.1’
For
each
lin
e,
valu
es
indi
cate
d by
a
diffe
rent
le
tter
are
signi
fican
tly
diffe
rent
. Di
ffere
nces
(t-
test
, P
< 0.
05).
pop.
=
popu
latio
n te
tra.
= di
ploi
d pl
ants
(te
trasp
orop
hyte
). R.
S.P.
=
dipl
oid
plan
ts
with
ra
re
sexu
al
phen
otyp
es
(abe
rratio
ns
of
life
cycle
) N
= po
pula
tion
size
* =
conf
iden
ce
inte
rval
+
= it
is c
ompu
ted
as:
dipl
oid/
(hap
loid
+
dipl
oid)
What controls haploiddiploid ratio? 325
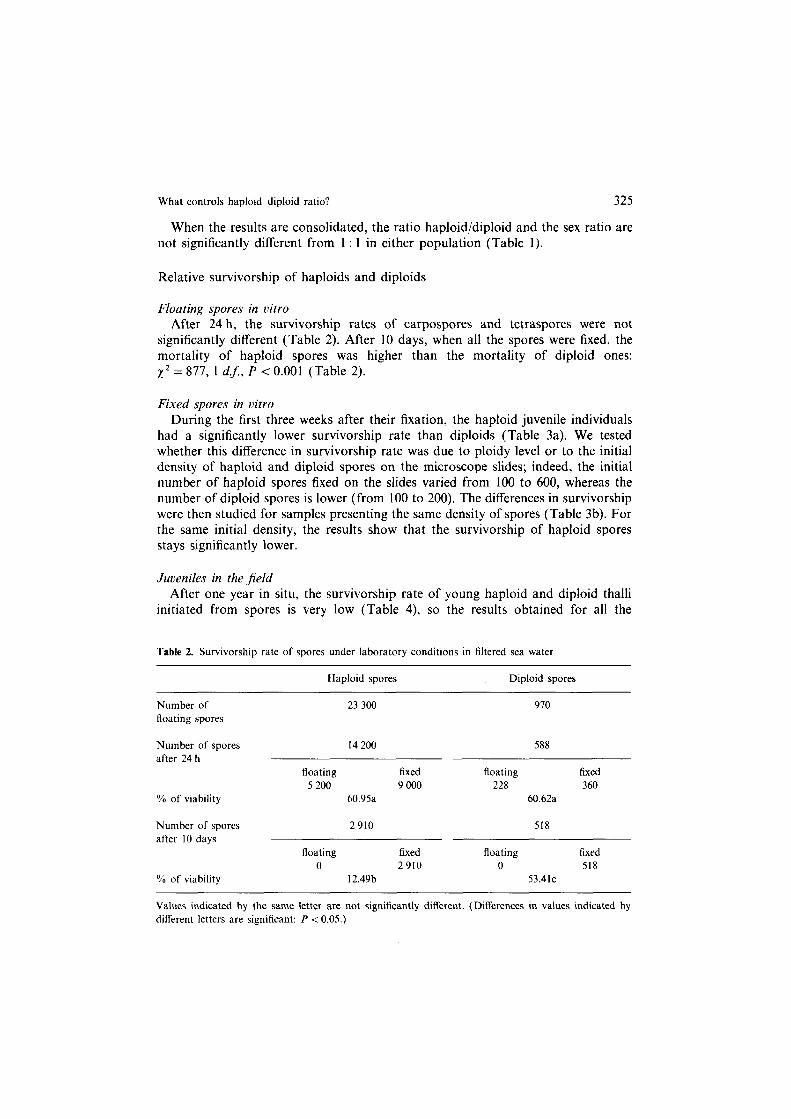
When the results are consolidated, the ratio haploid/diploid and the sex ratio are not significantly different from 1 : 1 in either population (Table 1).
Relative survivorship of haploids and diploids
Floating spores in vitro After 24 h, the survivorship rates of carpospores and tetraspores were not
significantly different (Table 2). After 10 days, when all the spores were fixed, the mortality of haploid spores was higher than the mortality of diploid ones: x2 = 877, 1 d.$, P < 0.001 (Table 2).
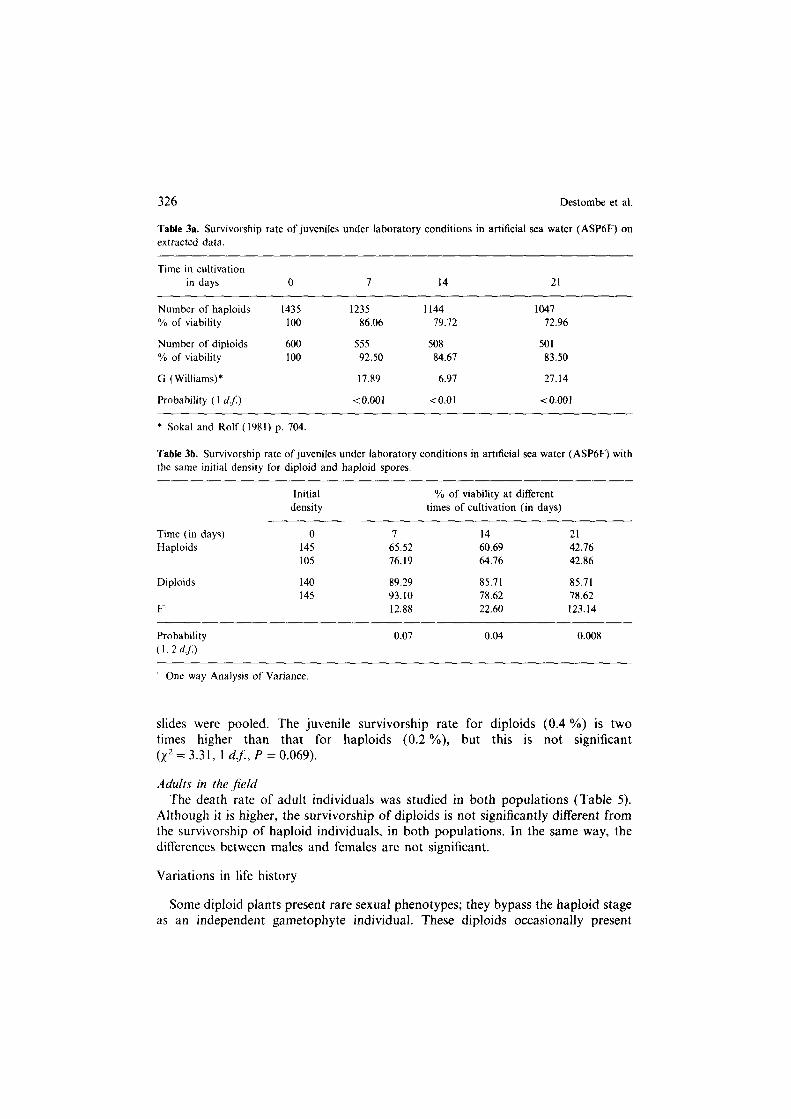
Fixed spores in vitro During the first three weeks after their fixation, the haploid juvenile individuals
had a significantly lower survivorship rate than diploids (Table 3a). We tested whether this difference in survivorship rate was due to ploidy level or to the initial density of haploid and diploid spores on the microscope slides; indeed, the initial number of haploid spores fixed on the slides varied from 100 to 600, whereas the number of diploid spores is lower (from 100 to 200). The differences in survivorship were then studied for samples presenting the same density of spores (Table 3b). For the same initial density, the results show that the survivorship of haploid spores stays significantly lower.
Juveniles in the jield After one year in situ, the survivorship rate of young haploid and diploid thalli
initiated from spores is very low (Table 4), so the results obtained for all the
Table 2. Survivorship rate of spores under laboratory conditions in filtered sea water
Number of
floating spores
Haploid spores Diploid spores
23 300 970
Number of spores after 24 h
% of viability
14200 588
floating fixed floating fixed 5200 9000 228 360
60.95a 60.62a
Number of spores after 10 days
% of viability
2910 518
floating fixed floating fixed
0 2910 0 518 12.49b 53.4lc
Values indicated by the same letter are not significantly different. (Differences in values indicated by different letters are significant: P < 0.05.)
326 Destombe et al.
Table 3a. Survivorship rate of juveniles under laboratory conditions in artificial sea water (ASP6F) on extracted data.
Time in cultivation in days 0 I 14 21
Number of haploids 1435 1235 1144 1047 % of viability 100 86.06 19.12 12.96
Number of diploids 600 555 508 501
% of viability 100 92.50 84.67 83.50
G (Williams)* 17.89 6.91 27.14
Probability (1 d.J) <0.001 <O.Ol t0.001
* Sokal and Rolf (1981) p. 704.
Table 3b. Survivorship rate of juveniles under laboratory conditions in artificial sea water (ASP6F) with the same initial density for diploid and haploid spores.
Initial % of viability at different density times of cultivation (in days)
Time (in days) 0 7 14 21 Haploids 145 65.52 60.69 42.16
105 76.19 64.76 42.86
Diploids 140 89.29 85.71 85.71 145 93.10 78.62 78.62
F 12.88 22.60 123.14
Probability
(1, 2 4f.l
0.07 0.04 0.008
One way Analysis of Variance
slides were pooled. The juvenile survivorship rate for diploids (0.4 %) is two times higher than that for haploids (0.2 %), but this is not significant (x2 = 3.31, 1 d$, P = 0.069).
Adults in the field The death rate of adult individuals was studied in both populations (Table 5).
Although it is higher, the survivorship of diploids is not significantly different from the survivorship of haploid individuals, in both populations. In the same way, the differences between males and females are not significant.
Variations in life history
Some diploid plants present rare sexual phenotypes; they bypass the haploid stage as an independent gametophyte individual. These diploids occasionally present
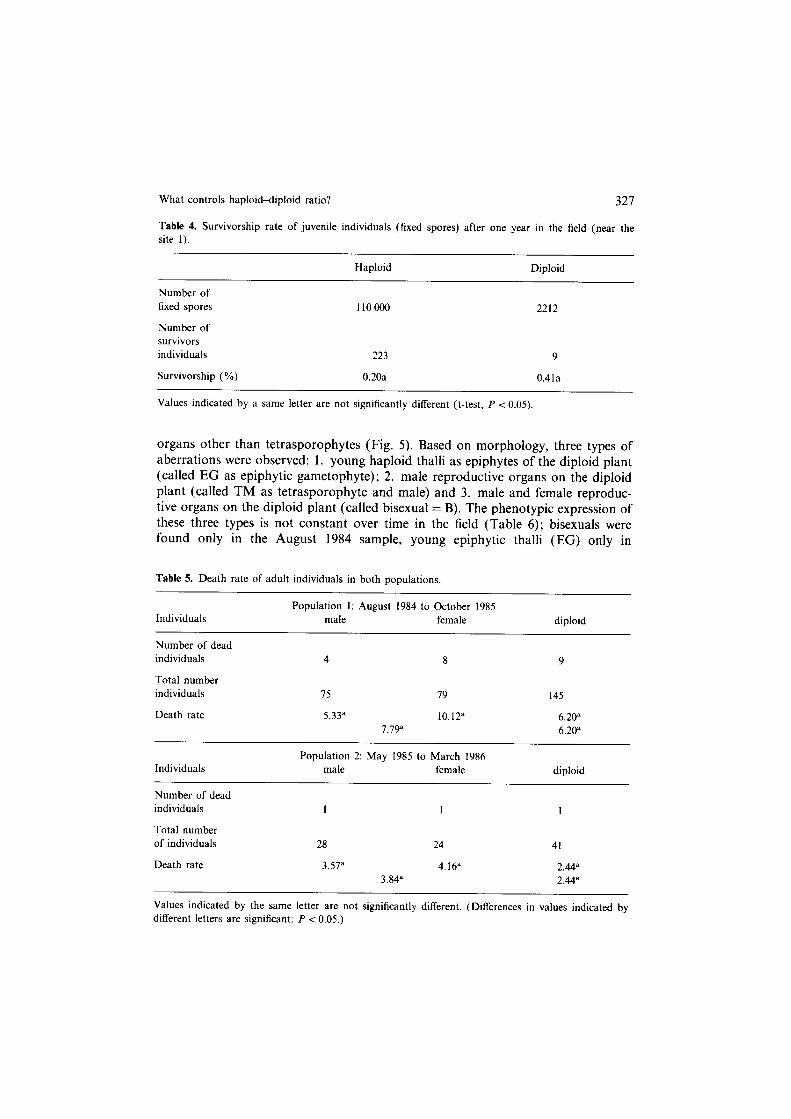
What controls haploiddiploid ratio? 327
Table 4. Survivorship rate of juvenile individuals (fixed spores) after one year in the field (near the site 1).
Number of fixed spores
Number of survivors
individuals
Haploid Diploid
110000 2212
223 9
Survivorship (X) 0.20a 0.41a
Values indicated by a same letter are not significantly different (t-test, P < 0.05).
organs other than tetrasporophytes (Fig. 5). Based on morphology, three types of aberrations were observed: 1. young haploid thalli as epiphytes of the diploid plant (called EG as epiphytic gametophyte); 2. male reproductive organs on the diploid plant (called TM as tetrasporophyte and male) and 3. male and female reproduc- tive organs on the diploid plant (called bisexual = B). The phenotypic expression of these three types is not constant over time in the field (Table 6); bisexuals were found only in the August 1984 sample, young epiphytic thalli (EG) only in
Table 5. Death rate of adult individuals in both populations.
Population 1: August 1984 to October 1985 Individuals male female diploid
Number of dead individuals 4 8 9
Total number
individuals 15 19 145
Death rate 5.33” 10.12” 6.20” 7.79” 6.20”
Population 2: May 1985 to March 1986 Individuals male female diploid
Number of dead
individuals 1 1 I
Total number of individuals 28 24 41
Death rate 3.51” 4.16” 2.44” 3.84a 2.44”
Values indicated by the same letter are not significantly different. (Differences in values indicated by
different letters are significant: P < 0.05.)
328 Destombe et al.
Fig. 5. The 4 types of abnormal individuals. (EC?) tetrasporophyte with epiphytic male (emg) and female (efg) gametophytes; (TM) male tetrasporophyte; (B) bisexual tetrasporophyte; (EGT) tetrasporophyte with epiphytic gametophytes (emg and efg) and epiphytic tetrasporophytes (et). (0) diploid thallus;
(~3) haploid thallus; (0) tetrasporocyst, where tetraspores are formed during meiosis; (3) tetrasporo- cyst, where male gametes ( = spermaties) are formed; (0) carpogone, containing one female gamete after fertilization, numerous mitosis of the zygote give the gonimoblast from which carpospores are
differentiated within a cystocarp.
February 1986 and male organs on diploid plants (TM) throughout the year. The proportions of the various types of aberration is given in Table 7. The proportion of these rare sexual phenotypes is 6.1 % in population 1 and 4.4 % in population 2.
In culture, the five diploid plants collected in the populations showed no spore liberation: spores germinated and increased in size as epiphytes on the diploid plant (this aberration is called EGT as epiphytic gametophyte and tetrasporophyte). This type of aberration is often observed on diploid individuals at the end of sporulation when they are grown in vitro (Destombe, in prep.). After two weeks of culture, numerous small thalli become visible on diploid fronds. Six weeks later, the young fronds measure 0.5 to 2 mm. After ten weeks, the size of the fronds is 6 to 10 mm and the first reproductive organs appear. On the diploid thallus each tetrad produced a yound frond and cross sections of this diploid thallus, at this level, showed traces of dead spores at the bottom of the young fronds. The number of
What controls haploiddiploid ratio?
Table 6. Stability of individual phenotypic expression for the “rare sexual phenotypes”.
329
Population 1 Month
number: 08 10 II 01 03 04 05 08 09 10 11 02
Year 84 85 86
No. of plant
1 -
2 3
4 V 5 B
6 7
8 9 B
10 T 11 TM 12 B
13 T 14 15 V
16 B 17
18 TM 19 V
20 21 TM 22
T
T T
T T
TM V
- - T T
T
T ~
T ~
v v T
T TM v
T - - -
T
T
- V - -
T V
T T
- T - -
T
T T
T T
TM TM
- V
V T V T
T
T T T
T T
V
V
V
T T
V T V
T V
TM
TM T T V T T T - EC
TM TM - V
TM T T - EC T T T T V
T V T EC T T T EC
TM T T TM T TM V
T T TM ~ TM T T T T
T TM T T TM
TM + 1
T T V T V
TM T T - T TM T T TM V TM - T T
TM T T T T T T TM T T - T T TM
Month
number
Year
04
85
05
Population 2
06 07 08 09 10 11
1 T - TM T 2 T - - T TM T V 3 V - - T T T V TM 4 T T TM 5 T T TM
T = (tetrasporophyte) normal diploid plant with tetrasporocyts. TM = (male tetrasporophyte) diploid plant bearing male organs and tetrasporocysts.
B = (bisexual) diploid plant bearing male organs and carposporophytes (female organs fertilizated) without tetrasporocysts.
EC = diploid plant bearing young epiphytic male and female gametophytes. V = vegetative diploid plant.
= “Sandy period” thalli covered by sand. = no erected thallus.
330 Destombe et al.
Table 7. Proportion of various types of rare sexual phenotype in the field.
TM B EG B+TM TM+EG Total
number
% in total number of rare sexual
number
% in total
number of rare sexual
14
61
5
100
4
17.4
Population 1 3 1 1 23
13 4.3 4.3
Population 2
5
TM = diploid individuals (tetrasporophyte) bearing male organs and tetrasporocysts.
B = diploid individuals (tetrasporophyte) with male organs and carposporophytes with tetrasporocysts called bisexual. EG = diploid individuals (tetrasporophyte) bearing young thallus (male and female) as epiphyte.
small thalli is, on average, 25 per linear centimeter of the parental diploid thallus. The offspring grew directly on the diploid plant and we counted the number of various types of young plants obtained. Three types of plants were identified: male, female and tetrasporophyte on the same individual (Table 8). On average, the proportion is respectively 36 % of males, 31 % of females and 32 % of tetrasporo- phytes. Thus, under laboratory conditions, the five diploid individuals give rise directly to $ of haploid individuals and j of diploid individuals.
Table 8. Number of young male, female and tetrasporophyte epiphyte thalli (EGT) per cm of parental
diploid thallus six months after their germination.
Parental thalli Sex and generations n Phenotype Male Female Tetra. Veg.
06 T 2.0 1.0 1.0 21 16 T 2.6 3.3 4.0 44 24 T 1.0 0 0.5 64 02 T 2.4 2.9 2.0 35 05 TM 0.3 0 0 16
Tetra = young diploid plants: tetrasporophytes. Veg. = vegetative Phenotypes of parental thallus:
T = normal diploid plant (tetrasporophyte). TM = diploid plant (tetrasporophyte) bearing male organs and tetrasporocysts.
Culture conditions:
strength of illumination 40 PE. m-* SK’, photoperiod: 12 h dark/l2 h light, temperature 19 k 1” C.

What controls haploid&ploid ratio?
Discussion
331
Relative frequencies of male and female haploid individuals and diploid individuals: sex ratio and haploidjdiploid ratio
Whereas selective factors might induce variation in the male/female and hap- loid/diploid ratios, fluctuations could also result from incorrect or inadequate sampling. For example, biased ratios observed by unique sampling could be due to non-concordant “reproducing” periods for the different types of individuals, or to flowering differences according to the sex (as for example in Asparagus, Vernet, 1971). In Gracilaria sp. Bird ( 1976) pointed out a deficiency of male individuals collected from an American population in July. Similar observations have been reported in many other studies of G. verrucosa (Isaac, 1956; Jones, 1959; Trono and Azanza-Corrales, 1981); however, when Bird (1976) cultured in the laboratory all the vegetative individuals he collected in July from the American population, he observed a sex-ratio of 1 : 1. Other studies of tetraspores in culture (Ogata et al., 1972 and Bird et al., 1977) also show equal numbers of males and females. Consequently it is necessary to examine marked individuals at regular time inter- vals to know exactly the composition of populations. Our study of marked individuals showed that: (i) haploids and diploids mainly reproduce at the same time from July to December (Fig. 4) although mature individuals can be found at any time of year; (ii) there are large differences from one sampling time to the other. For example in November 1985 there was a significant difference between male and female gametophyte frequencies in site 1; although the sex-ratio over the year (given by consolidated data) of this population is 1 : 1, as expected in panmictic populations (Fisher, 1930).
In both of the populations studied, the percentage of haploids and diploids is not significantly different. This conclusion would not be altered if vegetative individuals were taken into account because the frequency of vegetative individu- als does not seem to vary with ploidy. Bird ( 1976) has shown that “infertile individuals” are haploid and diploid individuals in equal proportions.
The results reported in the literature depend on the species studied and on the geographical location. In Gracilaria spp., Trono and Azanza-Corrales (1981) found an excess of haploid individuals in Manila bay populations, whereas Whyte et al. ( 1981) noted an excess of diploid individuals in British Columbia. In G. cuneata Hay and Norris (1984) have described haploids and diploids in the same proportions in one population in Panama. In most studies of red algae, however, populations are described as having an excess of diploids: G. domingensis (Hay and Norris, 1984), Hypnea cervicornis (Mshigeni, 1976), Iridea cordata (Hansen and Doyle, 1976) and Callithamion byssoides (Kapraun, 1978). Excess of diploid individuals has often been related to the species range, i. e., diploids are more numerous at the limits of the species’ geographical distribution (Dixon, 1965) or at the limit of tide levels (Mathieson and Prince, 1973).

332 Destombe et al.
How are haploid and diploid individuals maintained?
The coexistence of haploid and diploid individuals in the populations allows the question of the relative advantages of the haploid and the diploid stages to be posed. How is the frequency of haploid and diploid individuals determined in this life cycle? To answer this question, we built a model examining the relative effect of annual fertility (number of spores produced), survivorship of juvenile individuals, and annual survival rate of adult individuals on haploid/diploid ratios (appendix). If survivorship rates of haploid and diploid adult individuals are equal, the frequency of diploids depends only on the relative fertility of female and diploid plants, and on the survivorship rate of juveniles. Because the aim of the model is to look at the relative proportion of haploid and diploid individuals, the female fertility is here considered as the number of diploid spores produced by the female and not as the number of zygotes it produces. The relative fertility (K) of female gameto- phytes over diploid individuals was measured in populations of G. uerrucosa (Destombe 1987); this study shows that tetrasporophytes have a higher fertility than gametophytes (0.5 < K < 1).
If we assume that haploids and diploids have an equal probability of establish- ment and equal survival rates as adults, then for the relative fertility of K = 1 (i. e. equal female and diploid fertilities), the model gives the frequency of diploid individuals P = 0.41; and for K = 0.5, P = 0.33. The expected frequency of diploids is significantly below what is observed (Table 1).
Differences in viability between haploids and diploids were observed on juveniles in the laboratory (Tables 2 and 3) but were not significant in the field (Tables 4 and 5); this could be due to the low number of spores at the end of the experiment. One argument advanced in the literature is that diploid individuals may be protected against the deleterious effects of somatic mutations and cover up, by complementa- tion, the effect of deleterious genes (Bernstein et al., 1985; Maynard Smith, 1986) in contrast to haploids, which possess only a singly copy of each gene. Actually, diploids of G. verrucosa have been shown to be less susceptible to mutagenesis due to ultraviolet irradiation than haploids (Destombe, in prep.). A significant differ- ence of mortality of 10 % was observed between haploid and diploid juveniles in vitro (Table 3a) which is consistent with a difference in mortality two times higher in haploids after a year in the field. We introduced into the model the relative survivorship rate of juvenile and adult individuals observed in the field studies (Tables 4 and 5).
If we assume demographic equilibrium of the populations, the model yields for K = 0.5, P = 0.48 and for K = 1, P = 0.55. The model now describes accurately what is observed. Thus the advantage observed in this study to the benefit of diploids seems to be a good explanation for the higher-than-expected frequency of diploids observed in the field. A two-fold advantage in viability of diploids is necessary to maintain diploids at a frequency around 0.50.
A result of this model is that the more fertile female haploid individuals are, the more frequent diploid individuals will be. In general, the more fertile one of the stages is, the more frequent will be the other stage. As a result, diploid plants will

What controls haploiddiploid ratio? 333
be more frequent if their death rate is lower, if their probability of establishment is higher, or if their fecundity is lower. This means that the frequency of diploid individuals is not directly related to their fitness if both viability and fertility of diploid individuals are affected; their frequency can vary downwards or upwards. An uncertainty in the observations is that we do not know if vegetative individuals are haploids or diploids. If haploids were slower to develop, i. e. to reach the adult stage, a majority of the vegetative individuals could be haploids, lowering the actual frequency of diploid individuals closer to what is observed, between 43 and 48 % (Table 1).
Variations of life cycle
In both populations, anomalies of life cycle were noticed at appreciable frequencies (5 %). All these rare phenotypes appear at the diploid stage. They all skip a part of the haploid stage.
These anomalies are discussed below, according to the way they skip the haploid stage.
1. The haploid stage is no longer an independent stage: Small gametophyte thalli (EG) develop as epiphytes on parental diploid fronds in
the field. This phenomenon is quite common in red algal species (Svedelius, 1937; Tokida and Yamamoto, 1965; Bodard, 1966). It can be explained by non-liberation of tetraspores and by germination within the thallus (Fritsch, 1948). Thus male or female gametophytes develop on the sporophyte. The new thalli have small vegetative fronds and no real holdfast. They mature rapidly and are able to produce spores in less than six months. The life cycle is thus considerably shortened. The haploid stage is no longer independent of the diploid plant.
2. The haploid stage is totally reduced to the reproductive organs as in higher plants:
This is the case for TM (male reproductive organs and tetrasporocysts are observed on the tetrasporophyte) and for bisexuals (male and female reproductive organs on the diploid individual). Van der Meer (pers. corn.) shown that 100 % of tetrasporophytes of G. tikvahiae, G. foliiera and of G. sjoestidtii produced spermaties (male gametes). The same hypothesis of tetraspore germination in the thallus has been suggested (Cabioch, 1972) to explain the presence of male organs on the diploid thallus. Thus, the bisexual plants (B) observed by us, by Kling (1978) also at Cape Gris Nez and by Oliveria (1968) in a population in Brittany, can be interpreted in the same way, with male and female spores giving directly male and female organs. The occurrence of male, female and tetrasporophyte organs on the same thallus was observed in our populations. This situation has been described in G. verrucosa by Church ( 1919) and in Gracifaria sp. by Van der Meer and Todd ( 1977). Van der Meer and Todd (1977) have shown that these aberrant fronds may arise by mitotic recombination of diploids; the diploid thallus is heterozygous for a sexual locus which gives, without meiosis, by mitotic recombination only, male and female tissues appearing as homozygous patches. On these patches, diploid gametes are able to differentiate and to give, after fertilization, tetraploid or triploid new plants.

334 Destombe et al.
3. The haploid stage may no longer be present: The rare sexual phenotype EGT observed in the laboratory only (Table 8) is
characterized by the production not only of young tetrasporophytes but also of male and female gametrophytes directly on the parental diploid tetrasporophyte. The EG observed in the field may be the same as EGT, because in the field epiphytic tetrasporophytes on tetrasporophytes are difficult to observe and the maturity of tetrasporocysts is later than that of male and female reproductive organs. All diploid plants are of this type under laboratory conditions, which suggests that this type of rare sexual phenotype might be more common under special circumstances than was observed in the two populations studied. Moreover, the field study shows that all these aberrations are not stable in time and that one type of aberration is able to change to another type (Table 6). For example, in culture in vitro the diploid thallus number 05 (Table 8) had, before culture, male and tetraspore organs. After a few months of culture this diploid thallus (TM) still bore male and tetraspore organs but also small male gametopytes. If, within populations, all diploids behaved as in the laboratory, giving $ of tetrasporophytes and f of gametophytes, the frequency of diploids (P) can be recalculated (see appendix). The model yields with K ranging from 0.5 to 1:
(i) 0.43 < P < 0.50 with equal viabilities of haploid and diploid individuals (S=Sl and d=dl) and;
(ii) 0.60 < P < 0.65 with the two-fold differences in viabilities like those observed in the field (Tables 4 and 5).
Thus, the existence of such behavior of diploid individuals can explain, not only the two-fold viability advantage of diploids, but also why they reach a frequency of 50 % in natural populations. When these two features are combined, the frequency of diploids reaches more than 0.60, which is higher than what is observed. For example, only a small proportion of diploid individuals may present such aberra- tions. That would yield at equilibrium a frequency of diploid individuals closer to 50 %. Thus, under laboratory conditions, attached offspring of diploids skip at least the juvenile part of the haploid stage. That could be a reaction to certain environmental conditions that have been selected. Laboratory conditions were less disturbed in terms of water currents, temperature and salinity compared to field conditions. In such “stable” conditions, reproductive success may be higher for spores that germinate on an already-existing thallus, whereas in more disturbed environments, there may be a higher success rate for spores which germinate on a new site rather than on a thallus, because thalli are regularly broken under such conditions (Destombe, 1987) and are not able to regenerate because they have no disks.
The phenotypic plasticity observed suggests that there could be no developmental constraints preventing the appearance of variants and leading from a haploid- diploid life cycle towards a diploid life cycle. Although in our study no variant gametophyte was observed to bypass the diploid stage, one might ask whether evolution towards a haploid life cycle is possible. It sometimes happens in ferns (Farrar, 1967) another major haplo-diploid group of organisms. In G. edufis and in G. corticata, cystocarpic plants (female) have been directly obtained from

t
carpospores, i. e. the tetrasporophyte phase was eliminated (Krishnamurthy et al., 1969). In this case, the stability of such a haploid-diploid life cycle can be questioned.
A direct way to address the question of evolution of the life cycle of red algae would be to compare the relative success of one variant that leads to suppression of the individualized haploid stage, with that of another variant that leads to suppres- sion of the individualized diploid stage within the haploid-diploid life cycle, in order to investigate the selective factors maintaining such a haploid-diploid life cycle.
A population genetics model can be designed to look at the outcome of selection between three different alleles; one that determines that the haploid stage will produce haploid spores only (after fecundation and meiosis), one that determines that the diploid stage will produce diploid spores only, and the “wild type” allele (which determines the haploid-diploid life cycle).
In a closed population, one of the two mutant alleles will spread, depending on the respective life-time fecundity of the haploid and diploid stages; i. e. it is profitable to produce the spores that present the higher life-time fecundity. In every case, the wild type allele will be eliminated whatever the dominance relationship between the three alleles are. The wild type allele can be maintained if haploids and diploids present the same life-time fecundity, but such an equilibrium is unstable (Couvet et al., in prep.). A reason for maintenance of a haploid-diploid life cycle could be that haploids and diploids present different selective advantages. Haploid spores have higher dispersal abilities (Destombe, 1987) whereas diploid spores have higher survivorships. Thus a metapopulation effect can be invoked (Slatkin and Wade, 1978; Couvet et al., 1985) where diploids expand in established populations and haploids colonise new sites.
Acknowledgements
We would like to thank Dr. Janis Antonovics, Dr. Graham Bell, Dr. Judith Bronstein, Dr. Christopher Grant, Dr. Martine Hossaert, Dr. David A. Jones, Dr. Doyle McKey and Dr. John P. Van
der Meer for discussions on the first drafts of this paper and for correcting our English. We are grateful to Dr. Marcel Bodard, Dr. Jose Godin, Dr. Claude Lefebvre. Marc Nochet, and Juliette Bidiet for their help in many aspects of this study. Finally, special thanks to Jean Delivinquier, Michele Delecourt and
Godin’s family for their technical assistance. We would like to thank the two anonymous reviewers for the suggestions they have made about the paper. This research has been supported by the CNRS (ATP Biologie des Populations and RCP n”720) and by an IFREMER-Region Nord Pas-de-Calais grant.
References
Bernstein, H., H. C. Byerly, F. A. Hopf, and R. E. Michod. 1985. Genetic damage, mutation and the
evolution of sex. Science 229: 1277~1281. Bird, N. 1976. Studies on Gradaria: ecology of an attached population of Graciluriu sp. at Barrachois
Harbour, Colchester Co., N.S. Proc. N. S. Inst. Sci. 27: 144158. Bird, N., J. McLachlan, and D. Grund. 1977. Studies on Gruciluriu. 5. In vitro history of Gracihria sp.
from the maritime provinces. Can. J. Bot. 55: 128221290.

336 Destombe et al.
Bodard, M. 1966. Sur le development des tttrasporocystes d’Anatheca monfugnei (Schmitz) Solieriacees Gigartinales. Bull. I.F.A.N. 27 ser. A, n”3: 8677894.
Bodard, M. 1973. Reflexions sur les cultures d’Algues rouges a partir de boutures. Bull. Sot. Phycol. Fr.
18: 2&30. Cabioch, J. 1972. Un nouveau cas d’anomalie du cycle des Gigartinales. C.R. Acad. SC. Paris
275: 197991981.
Church, A. H. 1919. Historical review of the Floridem. J. Bot. 57: I-332.
Couvet, D., P. Gouyon, F. Kjellberg, I. Olivieri, D. Pomente, and G. Valdeyron. 1985. De la metapopulation au voisinage: la genbtique des populations en destquilibre. Gen. Select. Evol. 17: 407417.
Destombe, C. 1987. Biologie des populations de I’algue rouge Graciluria verrucosa (Hudson) Papenfuss,
interit et implication en aquaculture. These de doctorat, Universite de Lille Flandre-Artois. Dion, P., and R. Delepine. 1983. Experimental ecology of Gigurtina stellata (Rhodophyta) at Roscoff,
France, using an in situ culture method. Bot. Mar. 26: 201-211. Dixon, P. S. 1965. Perennation, vegetative propagation and algal life histories, with special reference to
Asparagopsis and other Rhodophyra. Botanica Gothobury 3: 67-74.
Farrar, D. R. 1967. Gametophyte of four tropical fern genera reproducing independently of their sporophytes in the southern Appalachians. Science 155: 1266 1267.
Fisher, R. A. 1930. The genetical theory of natural selection. Oxford University Press, Oxford. Fritsch, F. E. 1948. The structure and reproduction of the Algae. Vols. 1 & 2. Cambridge University
Press, Cambridge. Hansen, J. E., and W. Doyle. 1976. Ecology and natural history of Iridaea corduta (Rhodophyta;
Gigartinaceae): population structure. J. Phycol. 12: 2733278. Hay, M. E., and J. N. Norris. 1984. Seasonal reproduction and abundance of six sympatric species of
Grucilaria Grev. (Gracilariaceae; Rhodophyta) on a Caribbean subtidal sand plain. Hydrobiolog-
ica 116/l 17: 63 94. Hull, H. M., R. W. Hosmaw, and J. C. Wang. 1982. Cytofluorometric determination of nuclear DNA
in living and preserved algae. Stain Technology 57: 2733282. Isaac, W. E. 1956. The ecology of Graciluria conferooides (L.) Grev. in South Africa with special
reference to its ecology in Saldanha-Langebaan Lagoon. Proc. Int. Seaweed Symp. 2: 1733185. Jones, W. E. 1959. The growth and fruiting of Gracilaria uerrucosa (Hudson) Papenfuss. J. Mar. Biol.
Assoc. U.K. 38: 47756.
Kapraun, D. F. 1978. Field and cultures studies on selected North Carolina Polysiphoniu species. Bot. Mar. 21: 1433153.
Kling, R. 1978. Observation d’un cas de monoecie chez Gruciluria uerrucosa (Huds) Papenfuss (Gigarti-
nales). Bull. Sot. Bot. N. Fr. 31: 59960. Krishnamurthy, V., P. Venkataraju, and R. Venugopaz. 1969. An aberrant life-history in Graciluriu
edulis (Gurel) Silva and Gracilaria corticuta. J. Ag. Curr. Sci. 38: 343-344.
Magne, F. 1964. Recherches caryologiques chez les Floridees (Rhodophycees). Cah. Biol. Mar. 5: 461- 671.
Mathieson, A. C., and J. S. Prince. 1973. Ecology of Chondrus crispus Stackhouse pp. 53381. In Harvey M. J., and McLachlan J. (eds) Chondrus crispus. Nova Scotian Institute of Sciences Halifax.
Maynard Smith, J. 1986. The Problems of Biology. Oxford University Press, New York. Mshigeni, K. E. 1976. Studies on the reproduction of selected species of Hypnea (Rhodophyta,
Gigartinales) from Hawaii. Bot. Mar. 19(6): 341-346. Ogata. E., E. Matusi, and T. Nakamura. 1972. The life cycle of Gruciluriu verrucosa (Rhodophyceae,
Gigartinales) in vitro. Phycologia 1 I( 1): 75580. Oliveira, J. C. 1968. Recherches sur le dtveloppement et les organes reproducteurs de Gracilariu de la
Manche. These 32me Cycle de Biologie V&g&tale, Universite de Paris VI. Searles, R. B. 1980. The strategy of the red algal life history. Am. Nat. 115: 113~120. Slatkin, M, and M. J. Wade, 1978. Group selection on a quantitative character. Proc. natn. Acad. Sci.
USA 75: 3531-3534.

What controls haploiddiploid ratio? 337
Sokal, R. R., and F. J. Rolf. 1981. Biometry. W. H. Freeman and Company, New York. Svedelius, N. 1937. The apomeiotic tetrad division in Lomenfaria rosea. Symp. Bot. Ups]. 2(2): l-51. Tokida, J., and H. Yamamoto. 1965. Syntagmatic germination of tetraspores in Puchymeniopsis yendoi.
Phycologia 5(I): 15-20. Trono, G. C., and R. Azanza-Corrales, Jr. 1981. The seasonal variation in the biomass and reproductive
states of Gracilariu in Manilia Bay. Xth International Seaweed Symposium: 743-748. Van Der Meer, J. P., and E. R. Todd. 1977. Genetics of Graciluria sp: IV. Mitotic recombination and
its relationship to mixed phases in the life history. Can. J. Bot. 55: 281&2817.
Vernet, P. 1971. La proportion des sexes chez Asparagus ucuf[folius L. Bull. Sot. Bot. Fr. 118: 3455358.
Whyte, J.N.C., J.R. Englar, R.G. Sanders, and J.C. Lindsay. 1981. Seasonal variations in the biomass quantity of agar, from the reproductive and vegetative stages. Bot. Mar. 24: 493-501.
Wright, S. 1968. Evolution and genetics of populations. Vol. I: Genetics and Biometric Foundations, The University of Chicago Press, Chicago.
Yamanouchi, S. 1906. The life-history of Polysiphoniu uiolacea (preliminary note). Bot. Gaz. 41:
425 433.
Received I1 March 1988; accepted 28 December 1988. Corresponding Editor: P.-H. Gouyon
Appendix
Model examining the relative effects of annual fertility survivorship of juvenile individuals and annual survival rate of adult individuals on haploid/diploid ratio.
Type of individual
Frequency
(1)
Annual
fertility
(2)
Survivorship
rate of young individuals
(3)
Annual
survival rate of adults
(4)
diploid tetrasporophyte
haploid male
haploid female
P
(1--p)P
(1 - o/2
N
Nl=KN
s
SI
Sl
l-d
1 -dl
I -dl
NI: number of diploid spores produced by carposporophytes growing on the haploid female thallus. N: number of haploid spores produced by diploid individual after meiosis, K: relative fertility of haploid individuals K = NI/N. d: mortality of adult diploid individuals.
(/I: mortality of adult haploid individuals. S: probability for a diploid spore to germinate and to reach the adult stage (one year). 51: probability for a haploid spore to germinate and to reach the adult stage (one year).
P’, the frequency of tetrasporophyte the following is:
P(l-d) +((l-P)/2). KNS
P’=P(l-d)+(l-P)(l-dl)+((1-P)/2).KNS+PNSl

338 Destombe et al.
To compute P’ one has to notice that column (1) x column (4) yields the individuals surviving the following year, and that column (2) x column (3) yields the new individuals reaching the adult stage the following year. At equilibrium P = P’. It is possible to calculate P, which depends on d, dl, S, Sl, K and N.
If we consider that all diploids behave as in the laboratory, giving i of diploid spores (tetrasporophytes) and f of haploid spores (gametophytes); the equation gives:
p’ = P(l-d)+((l-P)/2).KNS+~.PNS
p(l-d)+(l-P)(l-dl)+((l-P)/2).KNS+$.PNSl+f.PNS





























