Embed Size (px)
Citation preview

Molecular Cell, Vol. 6, 1243–1252, November, 2000, Copyright 2000 by Cell Press
Vesicular Stomatitis Virus Matrix ProteinInhibits Host Cell Gene Expressionby Targeting the Nucleoporin Nup98
rRNA, and mRNA without affecting the export of tRNAmolecules (Her et al., 1997).
Nuclear transport occurs through nuclear pore com-plexes (NPCs) and is mediated by saturable transportreceptors that shuttle between the nucleus and cyto-
Cayetano von Kobbe,*‖ Jan M. A. van Deursen,†Joao P. Rodrigues,‡ Delphine Sitterlin,*Angela Bachi,* Xiaosheng Wu,†Matthias Wilm,* Maria Carmo-Fonseca,‡and Elisa Izaurralde*§
plasm. Most of the transport receptors identified to date*European Molecular Biology Laboratoryare members of a large family of RanGTP binding pro-Meyerhofstrasse 1teins exhibiting limited sequence similarity with the RanD-69117binding domain of importin-b, and have been termedHeidelbergimportins/exportins or karyopherins. The interaction ofGermanythese receptors with their cargoes or with nucleoporins†Mayo Clinicis regulated by the small GTPase, Ran (reviewed byRochester, Minnesota 55905Nakielny and Dreyfuss, 1999).‡ Institute of Histology and Embryology
In vertebrates, export of U snRNA, tRNA, and 5S rRNAFaculty of Medicineis mediated by members of the importin-b-like family ofUniversity of Lisbonreceptors. tRNA export is mediated by exportin-t, which1699 Lisboa Codexdirectly binds to the tRNA molecules in the presence ofPortugalRanGTP (Arts et al., 1998; Kutay et al., 1998). CRM1 isthe export receptor for polymerase II-transcribed spliceo-somal U snRNA (Fornerod et al., 1997) and has alsoSummarybeen implicated in the export of 5S rRNA (Fischer et al.,1995). Export of cellular mRNAs might not be mediatedVesicular stomatitis virus matrix protein (VSV M) hasby any member of the importin-b-related receptor familybeen shown to inhibit both transcription and nucleocy-(reviewed by Nakielny and Dreyfuss, 1999). The NPC-toplasmic transport. We have isolated a mutant formassociated proteins TAP/Mex67p and RAE1/Gle2p haveof M, termed M(D), lacking both inhibitory activities.been proposed to fulfill the role of export receptors forHeLa cells expressing M, but not M(D), accumulatemRNA (Bachi et al., 2000).polyadenylated RNAs within the nucleus. Concomi-
The different RNP export pathways converge at thetantly, a fraction of M, but not of the M(D) mutant,NPC. The overall three-dimensional architecture of thelocalizes at the nuclear rim. Additionally, the nucleo-NPC is evolutionarily conserved and consists of threeporin Nup98 specifically interacts with M but not withstructural units: a ring-like central framework embracingM(D). In Nup982/2 cells, both the levels of M at thethe central channel of the pore is sandwiched betweennuclear envelope and its inhibitory effects on host cell–the cytoplasmic ring, from which eight cytoplasmic fi-
directed expression of reporter genes were signifi-brils emanate, and the nuclear ring, which anchors the
cantly reduced. Together, our data demonstrate thatnuclear basket (Yang et al., 1998; Stoffler et al., 1999).
VSV M inhibits host cell gene expression by targeting The structural units of the NPC are composed of multiplea nucleoporin and primarily blocking nuclear export. copies of up to about 50 polypeptides called nucleopor-
ins. Nucleoporins often contain multiple phenylalanine-Introduction glycine dipeptide repeats (FG) clustered in domains,
which in vertebrates are glycosylated by addition ofThe vesicular stomatitis virus (VSV) belongs to the Rhab- O-linked N-acetylglucosamine (GlcNAc) (reviewed bydoviridiae family and causes acute infections in a wide Rout and Wente, 1994).range of mammalian hosts (reviewed by Wagner and Evidence of a role for vertebrate nucleoporins in RNARose, 1995). Its matrix protein (VSV M) has been shown export came from the use of anti-nucleoporin antibodiesto play a key role in virus assembly and budding and in and of wheat germ agglutinin (WGA), a lectin that bindsthe inhibition of host cell gene expression during infec- to O-linked GlcNAc residues. WGA or antibodies thattion. When expressed in vertebrate cells, in the absence recognize the FG repeat region of various nucleoporinsof any other viral protein, VSV M produces dramatic block several export processes including those of Ualterations in cellular RNA metabolism and mRNA ex- snRNA, 5S rRNA, and tRNA (Featherstone et al., 1988;pression (Black and Lyles, 1992; Ahmed and Lyles, 1998; Neuman de Vegvar and Dahlberg, 1990; Terns andand references therein). Among its pleiotropic effects, Dahlberg, 1994). These reagents bind to a common sub-VSV M has been shown to shut off transcription by host set of glycosylated, FG repeat–containing nucleoporins
such as p62, Nup98, and CAN. The role of specificRNA polymerases I and II (Ahmed and Lyles, 1998). Fur-nucleoporins in export has been analyzed by using spe-thermore, when expressed in Xenopus laevis oocytes,cific antibodies and by overexpression of nucleoporinVSV M inhibited nuclear export of U snRNA, rRNA, 5Sdomains in cultured cells. Microinjection of antibodiesraised against the nucleoporin Nup98 in Xenopus oo-§ To whom correspondence should be addressed (e-mail: izaurralde@cytes inhibited U snRNA and mRNA export without af-embl-heidelberg.de).fecting tRNA export (Powers et al., 1997), whereas over-‖ Present address: Laboratory of Molecular Genetics, NIH, Balti-
more, Maryland 21224. expression of Nup153 in mammalian cells resulted in
Molecular Cell1244
nuclear accumulation of polyadenylated RNA (Bastoset al., 1997).
To investigate the mechanism of nuclear export inhibi-tion by VSV M, we first mapped the inhibitory domainof M protein to residues found between positions 44 and77. Within this domain, substitution of residues 52–54 byalanines resulted in a mutant form of the protein M(D),lacking the export inhibitory activity. HeLa cells express-ing M protein, but not the M(D) mutant, accumulatepolyadenylated RNA within the nucleus. At the sametime, a fraction of wild-type protein, but not of the M(D)mutant, localizes at the nuclear rim. We identify thenucleoporin Nup98 as a cellular protein able to discrimi-nate between wild-type M and the M(D) mutant. Experi-ments performed in Nup98-deficient embryonic mousecells confirmed that Nup98 represents the major bindingsite for VSV M at the NPC. Furthermore, in these cellsVSV M–mediated inhibition of the host cell transcriptionwas dramatically impaired. Together, our data indicatethat VSV M acts to deregulate nucleocytoplasmic trans-port, and thereby cellular gene expression, by targetingNup98.
Results
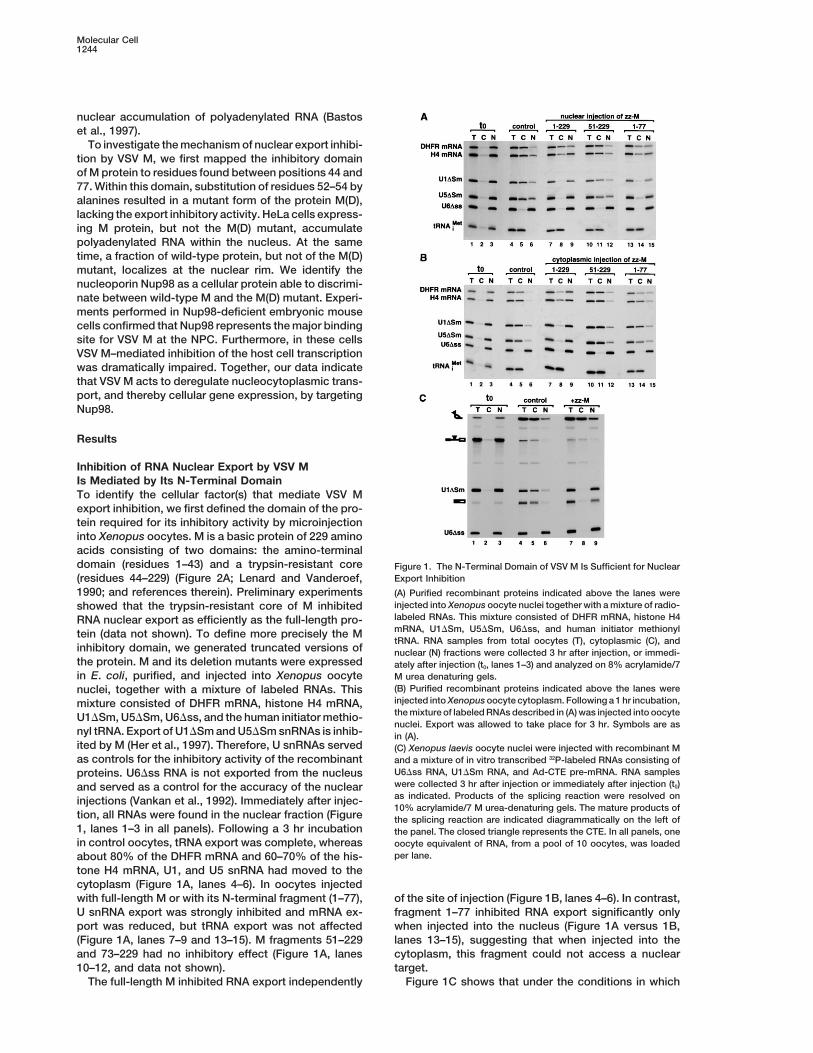
Inhibition of RNA Nuclear Export by VSV MIs Mediated by Its N-Terminal DomainTo identify the cellular factor(s) that mediate VSV Mexport inhibition, we first defined the domain of the pro-tein required for its inhibitory activity by microinjectioninto Xenopus oocytes. M is a basic protein of 229 aminoacids consisting of two domains: the amino-terminaldomain (residues 1–43) and a trypsin-resistant core Figure 1. The N-Terminal Domain of VSV M Is Sufficient for Nuclear
Export Inhibition(residues 44–229) (Figure 2A; Lenard and Vanderoef,1990; and references therein). Preliminary experiments (A) Purified recombinant proteins indicated above the lanes were
injected into Xenopus oocyte nuclei together with a mixture of radio-showed that the trypsin-resistant core of M inhibitedlabeled RNAs. This mixture consisted of DHFR mRNA, histone H4RNA nuclear export as efficiently as the full-length pro-mRNA, U1DSm, U5DSm, U6Dss, and human initiator methionyltein (data not shown). To define more precisely the MtRNA. RNA samples from total oocytes (T), cytoplasmic (C), and
inhibitory domain, we generated truncated versions of nuclear (N) fractions were collected 3 hr after injection, or immedi-the protein. M and its deletion mutants were expressed ately after injection (t0, lanes 1–3) and analyzed on 8% acrylamide/7in E. coli, purified, and injected into Xenopus oocyte M urea denaturing gels.
(B) Purified recombinant proteins indicated above the lanes werenuclei, together with a mixture of labeled RNAs. Thisinjected into Xenopus oocyte cytoplasm. Following a 1 hr incubation,mixture consisted of DHFR mRNA, histone H4 mRNA,the mixture of labeled RNAs described in (A) was injected into oocyteU1DSm, U5DSm, U6Dss, and the human initiator methio-nuclei. Export was allowed to take place for 3 hr. Symbols are as
nyl tRNA. Export of U1DSm and U5DSm snRNAs is inhib- in (A).ited by M (Her et al., 1997). Therefore, U snRNAs served (C) Xenopus laevis oocyte nuclei were injected with recombinant Mas controls for the inhibitory activity of the recombinant and a mixture of in vitro transcribed 32P-labeled RNAs consisting of
U6Dss RNA, U1DSm RNA, and Ad-CTE pre-mRNA. RNA samplesproteins. U6Dss RNA is not exported from the nucleuswere collected 3 hr after injection or immediately after injection (t0)and served as a control for the accuracy of the nuclearas indicated. Products of the splicing reaction were resolved oninjections (Vankan et al., 1992). Immediately after injec-10% acrylamide/7 M urea-denaturing gels. The mature products of
tion, all RNAs were found in the nuclear fraction (Figure the splicing reaction are indicated diagrammatically on the left of1, lanes 1–3 in all panels). Following a 3 hr incubation the panel. The closed triangle represents the CTE. In all panels, onein control oocytes, tRNA export was complete, whereas oocyte equivalent of RNA, from a pool of 10 oocytes, was loaded
per lane.about 80% of the DHFR mRNA and 60–70% of the his-tone H4 mRNA, U1, and U5 snRNA had moved to thecytoplasm (Figure 1A, lanes 4–6). In oocytes injectedwith full-length M or with its N-terminal fragment (1–77), of the site of injection (Figure 1B, lanes 4–6). In contrast,
fragment 1–77 inhibited RNA export significantly onlyU snRNA export was strongly inhibited and mRNA ex-port was reduced, but tRNA export was not affected when injected into the nucleus (Figure 1A versus 1B,
lanes 13–15), suggesting that when injected into the(Figure 1A, lanes 7–9 and 13–15). M fragments 51–229and 73–229 had no inhibitory effect (Figure 1A, lanes cytoplasm, this fragment could not access a nuclear
target.10–12, and data not shown).The full-length M inhibited RNA export independently Figure 1C shows that under the conditions in which

VSV M Inhibits Gene Expression by Targeting Nup981245
Figure 2. Substitution of M Residues 52–54by Alanines Abolishes Its RNA Export Inhibi-tory Activity
(A) Positions of the alanine scan mutants areshown on the primary amino acid sequenceof VSV M. Residues important for the inhibi-tory effect of M in RNA export are locatedaround position 52. The position of the M51Rmutation is indicated by a circle while theAPPPY motif implicated in budding is under-lined.(B) Purified recombinant M mutants indicatedabove the lanes were coinjected into Xeno-pus oocyte nuclei with the mixture of radiola-beled RNAs described in Figure 1A. RNAsamples were collected 3 hr after injection,or immediately after injection (t0, lanes 1–3),and analyzed as indicated in Figure 1A.(C) Xenopus oocyte nuclei were injected withzz-M or zz-M(D). Protein samples from totaloocytes (T), cytoplasmic (C), and nuclear (N)fractions were collected 4.5 hr after injection,or immediately after injection (t0), and ana-lyzed by Western blot.
(D) Human 293 cells were transfected with a mixture of plasmids encoding b-gal, CAT, and either GFP alone or C-terminally fused to M orM(D). Cells were collected 36 hr after transfection and b-gal or CAT activity was determined. Data from three separate experiments wereexpressed as the percentage of the activities measured when GFP alone was coexpressed. The data are means 6 standard deviations. Similarresults were obtained in experiments performed with different preparations of plasmid DNA. Protein expression levels were determined byWestern blot using anti-GFP antibodies.
M strongly inhibited export of U1DSm RNA and of a plasmid, either without insert or encoding M and M(D)proteins C-terminally fused to green fluorescence pro-spliced mRNA, the export of an excised intron lariat
bearing the constitutive transport element (CTE) of sim- tein (GFP). Western blot analysis revealed that the ex-pression levels of M were reproducibly lower than thoseian retrovirus-1 was not affected (Figure 1C, lanes 7–9
versus 4–6). Similarly, as reported by Her et al. (1997), of M(D), as M inhibits its own expression (Black andLyles, 1992). Furthermore, M protein inhibited expres-M did not interfere with the export of multiple tRNA
species (i.e., tRNASer, tRNAiMet, tRNALeu, and tRNATyr; Fig- sion of both cat and lacZ reporter genes, as previouslyreported (Black and Lyles, 1992), while coexpression ofure 1 and data not shown). These results indicate that
the inhibitory effect of VSV M in export is mediated mutant M(D) had no effect (Figure 2D). Since this mutantalso lacks export inhibitory activity, we conclude thatby its N-terminal domain and is not exerted simply by
blocking the central channel of the NPC. mutation of the same residues of M relieves the inhibitionof both transcription and export.
Mutation of M Residues 52–54 Abolishes Its InhibitorEffect in Transcription and Nuclear Export
Wild-Type M, but Not the M(D) Mutant, LocalizesTo define M residues important for RNA export inhibi-to the Nuclear Envelopetion, we constructed alanine substitutions of residuesNext, we compared the subcellular localization of M withwithin positions 1–77 (Figure 2A). The resulting mutantsthat of the M(D) mutant fused to GFP in transfectedwere tested by injection into Xenopus oocyte nuclei asHeLa cells. Both the wild-type protein and the M(D)described above. Figure 2B shows that substitution ofmutant distributed evenly between the nuclear and cyto-residues 52–54 by alanines (mutant D) abolished theplasmic compartments (Figures 3A and 3E). A fractioninhibitory effect of M in RNA export, whereas alanineof M, but not of M(D), was also detected in a rim aroundscan mutants A, B, C, and E retained the inhibitory activ-the nuclear periphery. When transfected HeLa cells wereity. The nucleocytoplasmic distribution of mutant M(D)permeabilized with digitonin prior to fixation, the nucleo-was similar to that of wild-type M following a 4.5 hrplasmic and cytoplasmic pools of M were solubilized,incubation (Figure 2C), suggesting that mutant M(D) isbut the fraction associated with the nuclear rim wasproperly folded. Thus, residues important for VSV Mresistant to detergent extraction (Figure 3B). The nuclearexport inhibition are located around position 52. Pre-rim–associated fraction of M colocalized with the label-viously, a methionine-to-arginine substitution at positioning produced by the monoclonal antibody 414 directed51 (M51R) was shown to render M protein defectiveagainst nucleoporins (compare Figure 3B with 3D), indi-in shutting off host RNA synthesis (Ferran and Lucas-cating that M associates with the NPC. In contrast, inLenard, 1997; Ahmed and Lyles, 1998). Prompted bycells transfected with the M(D) mutant, the GFP signalthese observations, we tested the effect of mutant M(D)disappeared on digitonin extraction (Figure 3F). Thus,on the expression of various reporter genes in a cotrans-wild-type M localizes to the nuclear envelope while mu-fection assay. Human 293 cells were cotransfected withtant M(D), which is unable to inhibit transcription andthe reporter genes cat (chloramphenicol acetyltransfer-
ase) and lacZ (b-galactosidase) together with pEGFPN3 export, was not detected at the nuclear rim.
Molecular Cell1246
Figure 3. A Fraction of VSV M Localizes to the Nuclear Rim
(A–H) HeLa cells were transfected with pEGFPN3 plasmid deriva-tives expressing GFP fusions of M or the M(D) mutant. In (B), (D),(F), and (H), HeLa cells were extracted with digitonin prior to fixation.In (C), (D), (G), and (H), HeLa cells expressing M-GFP or M(D)-GFPwere double labeled with the monoclonal antibody 414 directedagainst nucleoporins. Bar, 10 micrometers.
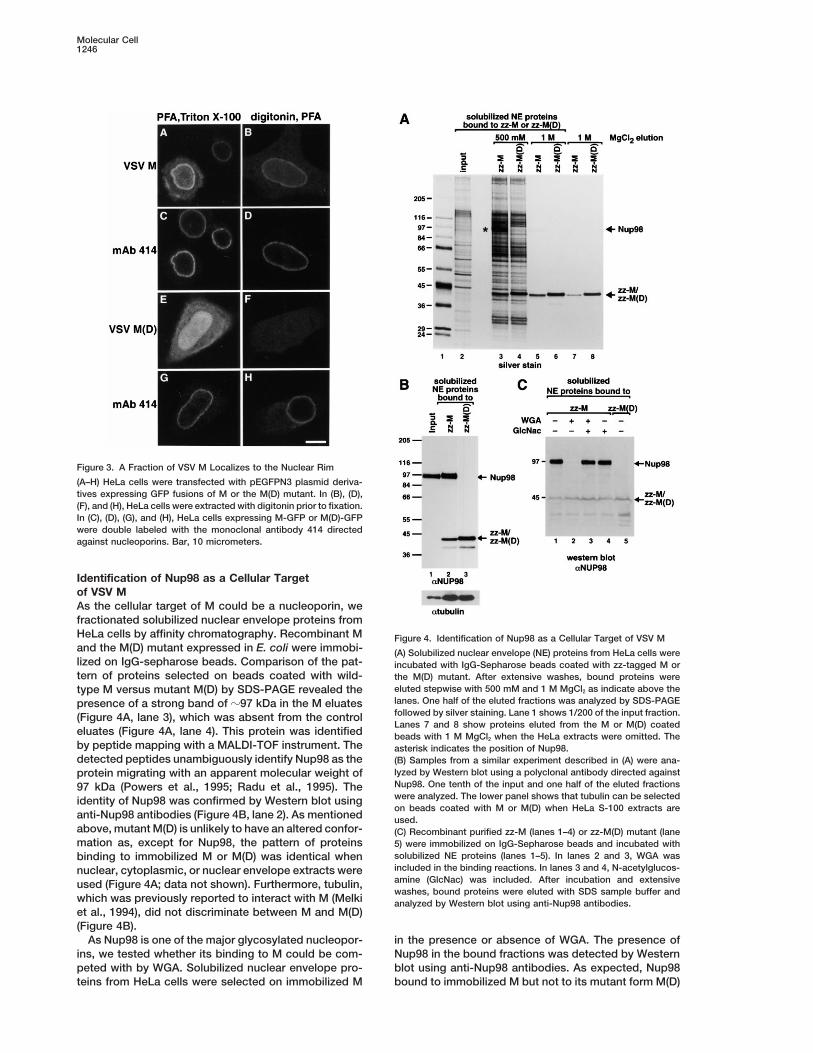
Identification of Nup98 as a Cellular Targetof VSV MAs the cellular target of M could be a nucleoporin, wefractionated solubilized nuclear envelope proteins fromHeLa cells by affinity chromatography. Recombinant M Figure 4. Identification of Nup98 as a Cellular Target of VSV Mand the M(D) mutant expressed in E. coli were immobi- (A) Solubilized nuclear envelope (NE) proteins from HeLa cells werelized on IgG-sepharose beads. Comparison of the pat- incubated with IgG-Sepharose beads coated with zz-tagged M ortern of proteins selected on beads coated with wild- the M(D) mutant. After extensive washes, bound proteins were
eluted stepwise with 500 mM and 1 M MgCl2 as indicate above thetype M versus mutant M(D) by SDS-PAGE revealed thelanes. One half of the eluted fractions was analyzed by SDS-PAGEpresence of a strong band of z97 kDa in the M eluatesfollowed by silver staining. Lane 1 shows 1/200 of the input fraction.(Figure 4A, lane 3), which was absent from the controlLanes 7 and 8 show proteins eluted from the M or M(D) coated
eluates (Figure 4A, lane 4). This protein was identified beads with 1 M MgCl2 when the HeLa extracts were omitted. Theby peptide mapping with a MALDI-TOF instrument. The asterisk indicates the position of Nup98.detected peptides unambiguously identify Nup98 as the (B) Samples from a similar experiment described in (A) were ana-
lyzed by Western blot using a polyclonal antibody directed againstprotein migrating with an apparent molecular weight ofNup98. One tenth of the input and one half of the eluted fractions97 kDa (Powers et al., 1995; Radu et al., 1995). Thewere analyzed. The lower panel shows that tubulin can be selectedidentity of Nup98 was confirmed by Western blot usingon beads coated with M or M(D) when HeLa S-100 extracts are
anti-Nup98 antibodies (Figure 4B, lane 2). As mentioned used.above, mutant M(D) is unlikely to have an altered confor- (C) Recombinant purified zz-M (lanes 1–4) or zz-M(D) mutant (lanemation as, except for Nup98, the pattern of proteins 5) were immobilized on IgG-Sepharose beads and incubated with
solubilized NE proteins (lanes 1–5). In lanes 2 and 3, WGA wasbinding to immobilized M or M(D) was identical whenincluded in the binding reactions. In lanes 3 and 4, N-acetylglucos-nuclear, cytoplasmic, or nuclear envelope extracts wereamine (GlcNac) was included. After incubation and extensiveused (Figure 4A; data not shown). Furthermore, tubulin,washes, bound proteins were eluted with SDS sample buffer and
which was previously reported to interact with M (Melki analyzed by Western blot using anti-Nup98 antibodies.et al., 1994), did not discriminate between M and M(D)(Figure 4B).
As Nup98 is one of the major glycosylated nucleopor- in the presence or absence of WGA. The presence ofNup98 in the bound fractions was detected by Westernins, we tested whether its binding to M could be com-
peted with by WGA. Solubilized nuclear envelope pro- blot using anti-Nup98 antibodies. As expected, Nup98bound to immobilized M but not to its mutant form M(D)teins from HeLa cells were selected on immobilized M

VSV M Inhibits Gene Expression by Targeting Nup981247
(Figure 4C, lane 1 versus 5). Binding of Nup98 to M wasprevented by WGA (Figure 4C, lane 2). This effect wasspecific as it was reversed by addition of GlcNAc (Figure4C, lane 3); GlcNAc alone had no effect (Figure 4C, lane4). In summary, Nup98 binds to M but not to the M(D)mutant. This binding is prevented by WGA, suggestingthat the O-linked sugars may contribute to the inter-action.
VSV M Interacts with the FG RepeatDomain of Nup98To determine the domain of Nup98 implicated in theinteraction with VSV M, we performed in vitro bindingassays. Full-length Nup98, an N-terminal fragment con-taining the FG repeats (residues 66–515), and a fragmentcomprising part of the repeats and the completeC-terminal domain (residues 222–920) were expressedin 293 cells as fusions with glutathione S-transferase(GST) (Figure 5A). The recombinant proteins were solubi-lized with 1% Triton X-100 and tested for binding toimmobilized M. Figure 5B shows that both the full-lengthNup98 and its N-terminal domain (66–515) bound to im-mobilized M (lanes 2 and 8). These interactions musthave been specific as they were competed with by WGA(Figure 5B, lanes 3 and 9). In contrast, fragment 222–920and also fragment 66–281 were not selected on immobi-lized M (Figure 5B, lane 5 and not shown). Thus, the Mbinding domain of Nup98 is contained within residues66–515. This fragment encompasses most of the FGdipeptide repeats (Powers et al., 1995; Radu et al., 1995),the hRAE1/Gle2 binding site, or GLEBS-like motif(Pritchard et al., 1999), and most of the predicted glyco-sylation sites of the nucleoporin (Figure 5A).
Figure 5. VSV M Interacts with the FG Repeat Domain of Nup98To determine whether the interaction between M andNup98 was direct, Nup98 fragment 66–515 was also (A) Full-length Nup98 and fragments 222–920 or 66–515 were ex-
pressed in 293 cells as GST fusions. Cells were lysed with 1% Tritonexpressed in E. coli as a GST fusion and selected onX-100, and the presence of each protein in the lysates was detectedbeads coated with M or the M(D) mutant. Figure 5Cby Western blot using a polyclonal anti-GST antibody. Schematicshows that the bacterially expressed fragment did notrepresentation of Nup98: numbers indicate the amino acid position.
interact specifically with immobilized M (lanes 5–8), even The vertical bars show the position of the FG repeats. The blackthough binding reactions were supplemented with 293 rectangle represents the GLEBS-like motif, and the branches thecell lysates. Also, in vitro translated full-length Nup98 position of putative glycosylation sites.
(B) Lysates from human 293 cells expressing full-length Nup98 oror fragment 66–515 failed to interact with M (data notfragments 222–920 or 66–515 were incubated with IgG-Sepharoseshown). In contrast, both full-length Nup98 and fragmentbeads coated with purified zz-M. Binding reactions were performed66–515, translated in vitro or expressed in E. coli, werein the presence (1) or absence (2) of WGA, as indicated above the
able to interact with three putative Nup98 partners: lanes. After extensive washes, bound proteins were eluted with SDSRAE1 (Pritchard et al., 1999), CRM1 (Zolotukhin and sample buffer. One tenth of the inputs (lanes 1, 4, and 7) and oneFelber, 1999), and TAP (Bachi et al., 2000) (data not quarter of the bound fractions (lanes 2, 3, 5, 6, 8, and 9) were
analyzed by Western blot using anti-GST antibodies.shown). Thus, their failure to interact with M was unlikely(C) Nup98 fragment 66–515 fused to GST was expressed in eitherto be due to a misfolding of the proteins. Since these293 cells or E. coli. The corresponding cell lysates were incubatedproteins interact with M only when expressed in 293with IgG-Sepharose beads coated with zz-M or the M(D) mutant
cells, we hypothesized that M recognizes the glycosy- as indicated above the lanes. Binding to zz-M coated beads waslated form of Nup98. The observation that Nup98 frag- performed in the presence (1) or absence (2) of WGA. Note that E.ments expressed in E. coli could not interact with M coli lysates were supplemented with lysates prepared from non-
transfected 293 cells.even in the presence of 293 cell extracts supports themodel that M binds only to glycosylated Nup98.
Nup98 Mediates the Inhibitory Effect of VSV M treating transfected cells with Triton X-100 prior to fixa-tion. As in HeLa cells, in wild-type mouse cells M-GFPin Cellular Gene Expression
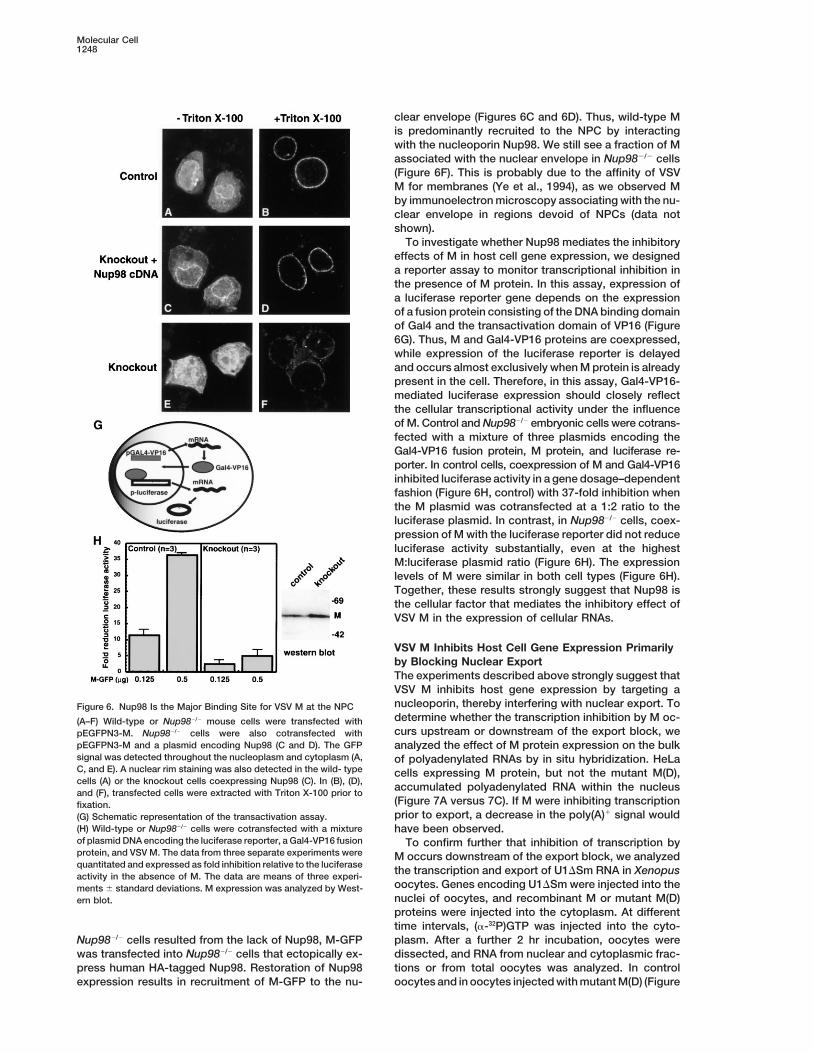
To determine whether binding to Nup98 accounts for localizes evenly within the cytoplasm and nucleoplasm,and concentrates at the nuclear rim (Figures 6A and 6B).the localization of VSV M at the nuclear envelope, we
compared the subcellular localization of M-GFP in wild- In contrast, in Nup982/2cells M-GFP was hardly detectedat the nuclear envelope (Figures 6E and 6F). To confirmtype to Nup982/2 mouse embryonic cells. The nuclear
rim–associated fraction of M-GFP was visualized by that the failure to detect M at the nuclear envelope of
Molecular Cell1248
clear envelope (Figures 6C and 6D). Thus, wild-type Mis predominantly recruited to the NPC by interactingwith the nucleoporin Nup98. We still see a fraction of Massociated with the nuclear envelope in Nup982/2 cells(Figure 6F). This is probably due to the affinity of VSVM for membranes (Ye et al., 1994), as we observed Mby immunoelectron microscopy associating with the nu-clear envelope in regions devoid of NPCs (data notshown).
To investigate whether Nup98 mediates the inhibitoryeffects of M in host cell gene expression, we designeda reporter assay to monitor transcriptional inhibition inthe presence of M protein. In this assay, expression ofa luciferase reporter gene depends on the expressionof a fusion protein consisting of the DNA binding domainof Gal4 and the transactivation domain of VP16 (Figure6G). Thus, M and Gal4-VP16 proteins are coexpressed,while expression of the luciferase reporter is delayedand occurs almost exclusively when M protein is alreadypresent in the cell. Therefore, in this assay, Gal4-VP16-mediated luciferase expression should closely reflectthe cellular transcriptional activity under the influenceof M. Control and Nup982/2 embryonic cells were cotrans-fected with a mixture of three plasmids encoding theGal4-VP16 fusion protein, M protein, and luciferase re-porter. In control cells, coexpression of M and Gal4-VP16inhibited luciferase activity in a gene dosage–dependentfashion (Figure 6H, control) with 37-fold inhibition whenthe M plasmid was cotransfected at a 1:2 ratio to theluciferase plasmid. In contrast, in Nup982/2 cells, coex-pression of M with the luciferase reporter did not reduceluciferase activity substantially, even at the highestM:luciferase plasmid ratio (Figure 6H). The expressionlevels of M were similar in both cell types (Figure 6H).Together, these results strongly suggest that Nup98 isthe cellular factor that mediates the inhibitory effect ofVSV M in the expression of cellular RNAs.
VSV M Inhibits Host Cell Gene Expression Primarilyby Blocking Nuclear ExportThe experiments described above strongly suggest thatVSV M inhibits host gene expression by targeting anucleoporin, thereby interfering with nuclear export. ToFigure 6. Nup98 Is the Major Binding Site for VSV M at the NPCdetermine whether the transcription inhibition by M oc-(A–F) Wild-type or Nup982/2 mouse cells were transfected withcurs upstream or downstream of the export block, wepEGFPN3-M. Nup982/2 cells were also cotransfected with
pEGFPN3-M and a plasmid encoding Nup98 (C and D). The GFP analyzed the effect of M protein expression on the bulksignal was detected throughout the nucleoplasm and cytoplasm (A, of polyadenylated RNAs by in situ hybridization. HeLaC, and E). A nuclear rim staining was also detected in the wild- type cells expressing M protein, but not the mutant M(D),cells (A) or the knockout cells coexpressing Nup98 (C). In (B), (D), accumulated polyadenylated RNA within the nucleusand (F), transfected cells were extracted with Triton X-100 prior to
(Figure 7A versus 7C). If M were inhibiting transcriptionfixation.prior to export, a decrease in the poly(A)1 signal would(G) Schematic representation of the transactivation assay.
(H) Wild-type or Nup982/2 cells were cotransfected with a mixture have been observed.of plasmid DNA encoding the luciferase reporter, a Gal4-VP16 fusion To confirm further that inhibition of transcription byprotein, and VSV M. The data from three separate experiments were M occurs downstream of the export block, we analyzedquantitated and expressed as fold inhibition relative to the luciferase the transcription and export of U1DSm RNA in Xenopusactivity in the absence of M. The data are means of three experi-
oocytes. Genes encoding U1DSm were injected into thements 6 standard deviations. M expression was analyzed by West-nuclei of oocytes, and recombinant M or mutant M(D)ern blot.proteins were injected into the cytoplasm. At differenttime intervals, (a-32P)GTP was injected into the cyto-plasm. After a further 2 hr incubation, oocytes wereNup982/2 cells resulted from the lack of Nup98, M-GFP
was transfected into Nup982/2 cells that ectopically ex- dissected, and RNA from nuclear and cytoplasmic frac-tions or from total oocytes was analyzed. In controlpress human HA-tagged Nup98. Restoration of Nup98
expression results in recruitment of M-GFP to the nu- oocytes and in oocytes injected with mutant M(D) (Figure

VSV M Inhibits Gene Expression by Targeting Nup981249
interaction with Nup98. Interfering with nuclear exportby targeting a nucleoporin points to a novel strategy bywhich cytoplasmic RNA viruses can inhibit host cell geneexpression, and represents a novel mechanism for cyto-toxicity.
A Fraction of VSV M Localizes to the Nuclear RimM was previously shown to be evenly distributed withinthe cell or specifically localized at the plasma membrane(Lyles et al., 1988; Blondel et al., 1990; Ye et al., 1994;Harty et al., 1999). Its subcellular localization appearedto be subject to variation depending on the fixation pro-cedure utilized and the cell type in which the protein wasexpressed (Peeples, 1988). We analyzed the subcellularlocalization of M fused to GFP in various cultured celllines. The GFP tag allowed us to detect the fusion proteindirectly in living cells, avoiding possible artifacts fromthe fixation procedure. In fixed or unfixed cells, M distrib-utes evenly within the nucleoplasm and cytoplasm.Moreover, a fraction of the protein localized at theplasma membrane and at the nuclear envelope. We areconfident that the M-GFP fusion reflects the localizationof wild-type M in human cells for the following reasons.First, similar results were obtained using an N-terminalzz-tagged version of M in two human cell lines: HeLacells and U-2 OS cells (data not shown). Second, M-GFPinduced cytopathic cell rounding in transfected HeLa,U-2 OS, and 293 cells. Third, M-GFP inhibited the ex-pression of reporter genes in 293 cells and mouse cells.Thus, the GFP tag did not interfere with the cytotoxic
Figure 7. VSV M Inhibits Host Cell Gene Expression Primarily byeffects or with the inhibitory activity of the proteinBlocking Nuclear Export(Blondel et al., 1990; Black and Lyles, 1992).(A–D) HeLa cells expressing wild-type M or mutant M(D), C-termi-
nally fused to GFP, were double labeled for poly(A) RNA.(E) Oocytes were injected with U1DSm gene followed by recombi- VSV M Interferes with Nuclear Export by a Mechanismnant M or M(D) proteins and a-32P-GTP as indicated diagrammati-
Distinct from Inhibiting Nup98 Functioncally on the top panel. RNA was analyzed as in Figure 1. BandsIt is striking that microinjection of anti-Nup98 antibodiesindicated by asterisks on the edge of the figure correspond to en-
dogenous transcripts. or of recombinant M in Xenopus oocytes leads to thesame pattern of export inhibition. Both inhibit U snRNAexport strongly, partially reduce mRNA export, and have
7E, lanes 1–6), U1DSm transcripts were present almost no effect on the export of tRNA (Her et al., 1997; Powersexclusively in the cytoplasmic fractions (lanes 2 and 5). et al., 1997; Figure 1). However, while the anti-Nup98In contrast, in oocytes injected with wild-type M (Figure antibody did not inhibit protein import, M was reported7E, lanes 7–9), the U1 transcripts were retained in the to inhibit protein import when expressed in Xenopusnucleus, indicating that M protein was blocking nuclear oocytes by microinjection of its mRNA (Her et al., 1997).export efficiently. Unexpectedly, M failed to inhibit tran- We have not been able to detect protein import inhibitionscription of U1, various endogenous transcripts (indi- by microinjecting recombinant M under conditions incated by asterisks in Figure 7E), and a tRNA gene (data which U snRNA export was strongly inhibited. In ournot shown). Since the basic transcriptional machinery in experimental conditions, M was injected in the cyto-Xenopus is very unlikely to differ from that of mammalian plasm 1 hr prior to the injection of the reporter proteins.cells, we conclude that VSV M does not directly target In the experiments reported by Her et al. (1997), oocytesthe transcription apparatus. expressing M were incubated for 22 hr prior to the injec-
tion of the karyophilic proteins. These oocytes accumu-late importin-a within the nucleus. Thus, it is possibleDiscussionthat protein import inhibition by M was a secondaryconsequence of the block of importin-a nuclear export.The matrix protein of vesicular stomatitis virus (VSV M)
shuts off host cell gene expression by inhibiting both Consistent with this, we observed that coinjection of Mwith labeled importin-a within oocyte nuclei partiallytranscription and nucleocytoplasmic transport. In this
study, we show that the nucleoporin Nup98 is critical for inhibits export of importin-a to the cytoplasm (data notshown). Thus, VSV M inhibits export mediated by CRM1,the inhibitory effect of M on host cell gene expression.
Furthermore, we demonstrate that the primary effect of CAS, and the mRNA export receptor(s), but does notdirectly interfere with protein import or with export medi-VSV M is to inhibit nuclear export; its effect on transcrip-
tion is thus seen to be an indirect consequence of its ated by exportin-t. Moreover, we have shown that M did

Molecular Cell1250
not inhibit TAP-mediated export of CTE-bearing intron Thus, the primary effect of M is the inhibition of nuclearexport. Taking into account the growing list of cellularlariats.
M binding to Nup98 may competitively inhibit the in- proteins whose activity is regulated at the level ofnucleocytoplasmic transport (reviewed by Kaffman andteraction with some of its putative partners such as
CRM1 (Zolotukin and Felber, 1999), TAP (Bachi et al., O’Shea, 1999), it is reasonable that a block in nuclearexport might lead to the pleiotropic effects associated2000), and hRAE1/Gle2p (Pritchard et al., 1999), thereby
interfering with its function. However, the possibility that with VSV M expression.M acts solely by disrupting Nup98 function is incompati-
Experimental Proceduresble with the observation that Nup982/2 cells do not dis-play an overt export inhibition phenotype (Kasper et al.,
Plasmids and Protein Expression in E. coli2000). Although we cannot rule out that in the Nup982/2cDNAs encoding the M protein of VSV (Indiana serotype, San Juan
cells a compensatory mechanism bypasses Nup98 strain) or the deletion mutants were amplified by PCR and clonedfunction, our data support a model in which M interferes within the NcoI-BamHI sites of vector pQE60zz. This vector is a
derivative of pQE60 (Qiagen) and has a zz tag inserted within thewith nuclear export by a mechanism distinct from inhibi-EcoRI and BglII restriction sites. Thus, proteins expressed usingtion of Nup98 function. It is likely that anti-Nup98 anti-pQE60zz vector have a zz tag at their N termini and a hexahistidinebodies act in a similar way. Nup98 has been localizedtag at their C termini. M cDNA was also cloned into the NcoI-BamHIat the baskets of the NPC (Radu et al., 1995; Fontoura sites of pBSpAlter, a derivative of pALTER-Ex1 (Promega). For ex-
et al., 1999). Studies on the export of the Balbiani ring pression in human cells, VSV M cDNA was excised from the pQE60zzmRNP from Chironomus tentans indicate that the nu- or the pBSpAlter vectors as an EcoRI-BamHI fragment and inserted
into the EcoRI-BamHI sites of the pEGFPN3 vector (Clontech).clear baskets with their terminal rings are dynamic struc-Expression of the resulting constructs in human cells generates antures that can open when large RNPs are translocatedM-GFP fusion with or without a zz tag at the N terminus, respectively.(Kiseleva et al., 1996). Thus, M and anti-Nup98 antibod-Alanine substitutions were introduced using an oligonucleotide-ies bound to the nuclear baskets via Nup98 may interfere directed in vitro mutagenesis system from Stratagene (Quick-
with the dynamic behavior of these structures. change site-directed mutagenesis) following the instructions of themanufacturer. Mutants were generated in the pQE60zz-M vector.Mutations were confirmed by restriction mapping and by sequenc-The Link between Transcription and Transporting. For expression in human cells, full-length Nup98 cDNA wasInhibition by VSV Minserted within the BamHI-NotI sites of pEBG vector. The pEBG-The Xenopus oocyte system allowed us to investigate Nup98-222-920 was generated by digestion and religation of plas-
the effects of M in RNA export independently of its mid pEBG-Nup98 with BamHI-KpnI. Nup98 fragment 66–515 clonedinhibitory effects on transcription. This is in contrast into pGEX-4T vector was kindly provided by Dr. Francoise Stutz.
This fragment was subcloned into the pEBG vector for expressionwith studies using transient transfections in culturedin 293 cells.cells, in which the ability of M to inhibit the expression
E. coli BL21(DE3) pLysS (Stratagene) strain was used for expres-of a reporter gene is measured. In this case, the effectssion of proteins in the pGEX-4T vector and M15 pRP4 strain (Qiagen)of M on mRNA transcription and export cannot be disso- for expression of proteins in the pQE60zz vector. Proteins were
ciated. Using the oocyte system, we showed that inhibi- purified on glutathione agarose beads (Sigma) or Ni1-NTA sepha-tion of RNA export by M is rapid and direct, as it can rose fast flow (Qiagen) as described by Gruter et al. (1998).be observed by coinjecting the recombinant protein to-
Xenopus laevis Oocyte Microinjections and RNA Analysisgether with the labeled RNAs within the nucleus. In anDNA templates for in vitro RNA synthesis of Ad-CTE pre-mRNA,early study, Fresco et al. (1987) showed that the effectsDHFR mRNA, histone H4 mRNA, U5DSm, U1DSm, and U6Dss RNA,of M on U snRNP processing and assembly, which areand human methionyl tRNA have been previously described (Saave-
likely to reflect an export block, were extremely rapid dra et al., 1997). Oocyte injections and analysis of microinjectedand observed prior to the onset of transcription inhibi- RNA by denaturing gel electrophoresis and autoradiography analy-tion. This raised the possibility that inhibition of tran- sis were performed as previously described (Hamm and Mattaj,
1990). The concentration of recombinant proteins in the injectedscription by M was a secondary consequence of ansamples was 1 mg/ml. Isolation of protein from oocytes and SDS-export block. Three sets of experiments demonstratePAGE was carried out as described (Kambach and Mattaj, 1992).that the effect of M on transcription is downstream of
the export inhibition. First, in Nup98-deficient cells MDNA Transfection, CAT, b-Galactosidase,
fails to inhibit the expression of a luciferase reporter. and Luciferase AssaysSecond, expression of M in HeLa cells results in accu- Human 293 cells were transfected using the calcium phosphatemulation of polyadenylated RNA within the nucleus. method. To test the effect of M on lacZ and cat gene expression,
cells were cotransfected with a mixture of plasmid DNA. This mixtureThird, although M strongly inhibits export in Xenopusconsisted of 2 mg of pCH110 (Pharmacia), 0.5 mg of pcDNA3-CAToocytes, no inhibition of transcription by polymerase II(Invitrogen), and 2 mg of pEGFP-N3 without insert or encoding C-ter-and III was observed. Since the basic transcriptionalminal GFP-tagged versions of M or M(D). Cells were harvested 36
machinery in Xenopus is very unlikely to differ from that hr posttransfection. CAT activity was measured as described byof mammalian cells, we interpret these results as an Morency et al. (1987). Gal4-VP16 was tested for transcriptional acti-indication that M does not target the transcription appa- vation properties on the Gal4-responsive reporter construct G5B/
pGL2 essentially as described in Kasper et al. (1999).ratus directly. The effects of M on transcription in mam-malian cells may be explained by a higher turnover of
Expression and Purification of GST-Nup98 Fusiontranscription factors compared to the oocyte system,Proteins in Human 293 Cellsand thus a requirement in mammalian cells to exportHuman 293 cells were transfected with either pEBG-Nup98(1–920),
mRNA-encoding labile transcription factors. It is also pEBG-Nup98(222–920), or pEBG-Nup98(66–515) using the calciumpossible that by blocking export, M interferes with some phosphate method. Approximately 40 hr after transfection, cells
were collected, washed once with ice-cold PBS containing 1 mMregulatory mechanism that does not exist in the oocytes.

VSV M Inhibits Gene Expression by Targeting Nup981251
PMSF, and lysed in lysis buffer (150 mM NaCl, 50 mM Tris-HCl [pH gift of VSV M cDNA. We are grateful to Maarten Fornerod, Iain W.Mattaj, and Christel Schmitt for critical reading of the manuscript8.0], 0.5 mM EDTA, 1% Triton X-100, 1 mM PMSF, 1 mg/ml leupeptin,
and 1 mg/ml aprotinin) for 15 min in ice. The lysate was then cleared and to Nicolas Roggli for graphic work. This study was supportedby the German Ministry of Research and Technology (BMBF), theby centrifugation at 13,000 RPM for 15 min at 48C. Protein expression
levels were determined by Western blot using anti-GST antibodies. European Molecular Biology Organization (EMBO), the Human Fron-tier Science Program Organization, and the Junta Nacional de In-vestigacao Cientifica e Tecnologica (Program PRAXIS XXI), Por-Antibodiestugal.A polyclonal anti-GST serum was obtained by immunizing rabbits
with bacterially expressed GST. A polyclonal antiserum to humanNup98 was kindly provided by Dr. Aurelian Radu. Monoclonal anti- Received May 11, 2000; revised September 18, 2000.body mAb414 to nucleoporins was purchased from BAbCO.
ReferencesImmunofluorescence in HeLa Cells and Nup982/2 Mouse CellsHeLa cells were transfected with either pEGFPN3-M or M(D) using Ahmed, M., and Lyles, D.S. (1998). Effect of vesicular stomatitis virusFuGene. Mouse cells were transfected with pEGFPN3-M using Su- matrix protein on transcription directed by host RNA polymerases I,perfect. Approximately 20 hr after transfection, cells were either II, and III. J. Virol. 72, 8413–8419.fixed with 3.7% formaldehyde for 10 min and subsequently perme- Arts, G.-J., Fornerod, M., and Mattaj, I.W. (1998). Identification of aabilized with 0.5% Triton X-100, or extracted first with 0.1% digitonin nuclear export receptor for tRNA. Curr. Biol. 6, 305–314.or Triton X-100 for 1 min and then fixed in formaldehyde. Double-
Bachi, A., Braun, I.C., Rodrigues, J.P., Pante, N., Ribbeck, K., vonlabeling experiments and in situ hybridizations with oligo U wereKobbe, C., Kutay, U., Wilm, M., Gorlich, D., Carmo-Fonseca, M., andperformed essentially as described previously (Calado et al., 2000).Izaurralde, E. (2000). The C-terminal domain of TAP interacts withthe nuclear pore complex and promotes export of specific CTE-Solubilization of Nuclear Envelope Proteinsbearing RNA substrates. RNA 6, 136–158.About 2.5 3 109 HeLa cells were lysed according to Dignam et al.Bastos, R., Lin, A., Enarson, M., and Burke, B. (1997). Targeting and(1983) to obtain nuclear and cytoplasmic extracts. After nuclearfunction in mRNA nuclear export of nuclear pore complex proteinlysis, the insoluble pellet containing chromatin and nuclear envelopeNup153. J. Cell Biol. 134, 1141–1156.proteins was resuspended in 2 ml of ice-cold buffer containing 0.1
mM MgCl2, 2 mM CaCl2, 5 mg/ml microccocal nuclease (Calbio- Black, B.L., and Lyles, D.S. (1992). Vesicular stomatitis virus matrixchem), and protease inhibitors. Subsequent steps were performed protein inhibits host cell-directed transcription of target genes inas described by Fontoura et al. (1999). vivo. J. Virol. 66, 4058–4064.
Blondel, D., Harmison, G.G., and Schubert, M. (1990). Role of matrixBinding Assays protein in cytopathogenesis of vesicular stomatitis virus. J. Virol.IgG-Sepharose beads (Pharmacia) were washed extensively with 1 64, 1716–1725.M NaCl, 25 mM HEPES (pH 7.9). Approximately 50 ml of packed
Calado A., Kutay, U., Kuhn, U., Wahle, E., and Carmo-Fonseca, M.beads was incubated with 50 mg of recombinant purified zz-tagged(2000). Deciphering the cellular pathway for transport of poly(A)-M or M(D) in 1 M NaCl, 25 mM HEPES (pH 7.9) for 1 hr at 48C. Beadsbinding protein II. RNA 6, 245–256.were then washed extensively with NP-40 buffer (1% NP-40, 50 mMDignam, J., Lebowitz, R., and Roeder, R. (1983). Accurate transcrip-Tris-HCl [pH 8], 5 mM EDTA, 5 mM EGTA, 15 mM MgCl2, 2 mM DTT,tion initiation by RNA polymerase II in a soluble extract from mam-150 mM NaCl, and protease inhibitors), and incubated with 1 ml ofmalian nuclei. Nucleic Acids Res. 11, 1475–1589.solubilized nuclear envelope proteins previously precleared with 50
ml of IgG-Sepharose beads. Beads were rotated for 1 hr at 48C, Featherstone, C., Darby, M.K., and Gerace, L. (1988). A monoclonalrecovered by gentle centrifugation, and washed six times with NP- antibody against the nuclear pore complex inhibits nucleocytoplas-40 buffer containing 200 mM KCl instead of 150 mM NaCl. Bound mic transport of protein and RNA in vivo. J. Cell Biol. 107, 1289–1297.proteins were eluted stepwise with 500 mM and 1 M MgCl2, 25 mM Ferran, M.C., and Lucas-Lenard, J.M. (1997). The vesicular stomati-HEPES (pH 7.9). The eluted proteins were precipitated with 20% tis virus matrix protein inhibits transcription from the human betatrichloroacetic acid (final concentration), resuspended in SDS sam- interferon promoter. J. Virol. 71, 371–377.ple buffer, and analyzed by SDS-PAGE followed by silver staining
Fischer, U., Huber, J., Boelens, W.C., Mattaj, I.W., and Luhrmann,or Western blot. In the experiments shown in Figure 5, beads wereR. (1995). The HIV-1 Rev activation domain is a nuclear export signalincubated with lysates prepared from human 293 cells transfectedthat accesses an export pathway used by specific cellular RNAs.with pEGB vectors expressing Nup98 full-length or fragments 222–Cell 82, 475–483.920 or 66–515. After washing, beads were eluted with SDS sampleFontoura, B.M.A., Blobel, G., and Matunis, M.J. (1999). A conservedbuffer and analyzed by SDS-PAGE followed by Western blot usingbiogenesis pathway for nucleoporins: proteolytic processing of aanti-GST antibodies. When indicated, WGA (Sigma) and GlcNAc186 kilodalton precursor generates Nup98 and the novel nucleoporin(Sigma) were added to the binding reactions at a final concentrationNup96. J. Cell Biol. 144, 1097–1112.of 4.5 mM and 50 mM, respectively.
Fornerod, M., Ohno, M., Yoshida, M., and Mattaj, I.W. (1997). CRM1Protein Identification is an export receptor for leucine-rich nuclear export signals. CellProteins eluted from the affinity columns were analyzed on SDS- 90, 1051–1060.PAGE followed by silver staining. Bands of interest were excised Fresco, L.D., Kurilla, M.G., and Keene, J.D. (1987). Rapid inhibitionand in-gel digested with trypsin (Shevchenko et al., 1996). Of the of processing and assembly of small nuclear ribonucleoproteinstotal digest solution, 0.3 ml was analyzed by peptide mass mapping after infection with vesicular stomatitis virus. Mol. Cell. Biol. 7, 1148–on a Bruker REFLEX MALDI time-of-flight mass spectrometer 1155.(Bruker-Franzen, Bremen, Germany) using the fast evaporation tech-
Gruter, P., Tabernero, C., von Kobbe, C., Schmitt, C., Saavedra, C.,nique for matrix preparation (Vorm et al., 1994).Bachi, A., Wilm, M., Felber, B.K., and Izaurralde, E. (1998). TAP,the human homologue of Mex67p, mediates CTE-dependent RNA
Acknowledgmentsexport from the nucleus. Mol. Cell 1, 649–659.
Hamm, J., and Mattaj, I.W. (1990). Monomethylated cap structuresThe technical assistance of Michaela Rode is gratefully acknowl-facilitate RNA export from the nucleus. Cell 63, 109–118.edged. We are indebted to Drs. Lionel Arnaud, Ralph Bischoff, and
Nelly Pante for their help. We wish to thank Dr. Aurelian Radu for Harty, R.N., Paragas, J., Sudol, M., and Palese, P. (1999). A proline-rich motif within the matrix protein of vesicular stomatitis virus andthe kind gift of anti-Nup98 antibodies, Dr. Barbara Felber for the
Nup98 cDNA, Dr. Douglass Lyles for providing M51R-mutant cDNA, rabies virus interacts with WW domains of cellular proteins: implica-tions for viral budding. J. Virol. 73, 2921–2929.and Drs. John K. Rose, Yves Gaudin, and Danielle Blondel for the

Molecular Cell1252
Her, L.S., Lund, E., and Dahlberg, J.E. (1997). Inhibition of Ran gua- complex: from molecular architecture to functional dynamics. Curr.Opin. Cell Biol. 11, 391–401.nosine triphosphatase-dependent nuclear transport by the matrix
protein of vesicular stomatitis virus. Science 276, 1845–1848. Terns, M.P., and Dahlberg, J.E. (1994). Retention and 59 cap trimeth-ylation of U3 snRNA in the nucleus. Science 264, 959–961.Kaffman, A., and O’Shea, E.K. (1999). Regulation of nuclear localiza-
tion: a key to a door. Annu. Rev. Cell Dev. Biol. 15, 291–339. Vankan, P., McGuigan, C., and Mattaj, I.W. (1992). Domains of U4and U6 snRNAs required for snRNP assembly and splicing comple-Kambach, C., and Mattaj, I.W. (1992). Intracellular distribution of thementation in Xenopus oocytes. EMBO J. 11, 335–342.U1A protein depends on active transport and nuclear binding to U1
snRNA. J.Cell Biol. 118, 11–21. Vorm, O., Roepstorff, P., and Mann, M. (1994). Improved resolutionand very high sensitivity in MALDI TOF of matrix surfaces made byKasper, L.H., Brindle, P.K., Schnabel, C.A., Pritchard, C.E.J., Cleary,fast evaporation. Anal. Chem. 66, 3281–3287.M.L., and van Deursen, J.M.A. (1999). CREB binding protein interacts
with the nucleoporin-specific FG repeats that activate transcription Wagner, R.R., and Rose, J.K. (1995). Rhabdoviridiae. In Virology,and mediate Nup98-HOXA oncogenicity. Mol. Cell. Biol. 19, 764–776. B.N. Fields, D.M. Knipe, and P.M. Howley, eds. (New York: Lippin-
cott, Williams, and Wilkins) pp. 1121–1135.Kasper, L.H., Wu, X., Mantcheva, R., Fontoura, B.M.A., Mantchev,G., and van Deursen, J.M.A. (2000). NUP98 controls stochiometry Yang, Q., Rout, M.P., and Akey, C.W. (1998). Three-dimensionalof multiple FG repeat–containing nuclear pore complex proteins. J. architecture of the isolated yeast nuclear pore complex: functionalCell Biol., in press. and evolutionary implications. Mol. Cell 1, 223–234.
Ye, Z., Sun, W., Suryanarayana, K., Justice, P., Robinson, D., andKiseleva, E., Goldberg, M.W., Daneholt, B., and Allen, T.D. (1996).RNP export is mediated by structural reorganization of the nuclear Wagner, R.R. (1994). Membrane-binding domains and cytopatho-
genesis of the matrix protein of vesicular stomatitis virus. J. Virol.pore basket. J. Mol. Biol. 260, 304–311.68, 7386–7396.Kutay, U., Lipowsky, G., Izaurralde, E., Bischoff, F.R., Schwarzmaier,Zolotukhin, A.S., and Felber, B. (1999). Nucleoporins Nup98 andP., Hartmann, E., and Gorlich, D. (1998). Identification of a tRNA-Nup214 participate in nuclear export of human immunodeficiencyspecific nuclear export receptor. Mol. Cell 90, 1061–1071.virus type 1 Rev. J. Virol. 73, 120–127.Lenard, J., and Vanderoef, R. (1990). Localisation of the membrane-
associated region of vesicular stomatitis virus M protein at the Nterminus, using the hydrophobic, photoreactive probe 125I-TID. J.Virol. 64, 3486–3491.
Lyles, D.S., Puddington, L., and McCreedy, B.J., Jr. (1988). Vesicularstomatitis virus M protein in the nuclei of infected cells. J. Virol. 62,4387–4392.
Melki, R., Gaudin, Y., and Blondel, D. (1994). Interaction betweentubulin and the viral matrix protein of vesicular stomatitis virus:possible implications in the viral cytopathic effect. Virology 202,339–347.
Morency, C.A., Neumann, J.R., and Russian, K.O. (1987). A novelrapid assay for chloramphenicol-acetyltransferase gene expression.Biotechnology 5, 444–448.
Nakielny, S., and Dreyfuss, G. (1999). Transport of proteins andRNAs in and out of the nucleus. Cell 99, 677–690.
Neuman de Vegvar, H.E., and Dahlberg, J.E. (1990). Nucleocytoplas-mic transport and processing of small nuclear RNA precursors. Mol.Cell. Biol.. 10, 3365–3375.
Peeples, M.E. (1988). Differential detergent treatment allows immu-nofluorescence localization of the Newcastle disease virus matrixprotein within the nucleus of infected cells. Virology 162, 255–259.
Powers, M.A., Macaulay, C., Masiarz, F.R., and Forbes, D.J. (1995).Reconstituted nuclei depleted of a vertebrate GLFG nuclear poreprotein, p97, import but are defective in nuclear growth and replica-tion. J. Cell Biol. 128, 721–736.
Powers, M.A., Forbes, D.J., Dahlberg, J.E., and Lund, E. (1997). Thevertebrate GLFG nucleoporin, Nup98, is an essential component ofmultiple RNA export pathways. J. Cell Biol. 136, 241–250.
Pritchard, C.E.J., Fornerod, M., Kasper, L.H., and van Deursen,J.M.A. (1999). RAE1 is a shuttling mRNA export factor that binds toa GLEBS-like Nup98 motif at the nuclear pore complex throughmultiple domains. J. Cell Biol. 145, 237–253.
Radu, A., Moore, M.S., and Blobel, G. (1995). The peptide repeatdomain of nucleoporin Nup98 functions as a docking site in transportacross the nuclear pore complex. Cell 81, 215–222.
Rout, M.P., and Wente, S.R. (1994). Pores for thought: nuclear porecomplex proteins. Trends Cell Biol. 4, 357–365.
Saavedra, C., Felber, B.K., and Izaurralde, E. (1997). The simianretrovirus-1 constitutive transport element CTE, unlike HIV-1 RRE,utilizes factors required for cellular RNA export. Curr. Biol. 7,619–628.
Shevchenko, A., Wilm, M., Vorm, O., and Mann, M. (1996). Massspectrometric sequencing of proteins from silver-stained polyacryl-amide gels. Anal. Chem. 68, 850–858.
Stoffler, D., Fahrenkrog, B., and Aebi, U. (1999). The nuclear pore





























