Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Amdras Semjen á Hans-Henning Schulze á Dirk Vorberg
Timing precision in continuation and synchronization tapping
Received: 16 November 1998 /Accepted: 21 April 1999
Abstract Wing and Kristo�erson (1973) have shownthat temporal precision in self-paced tapping is limitedby variability in a central timekeeper and by variabilityarising in the peripheral motor system. Here we test anextension of the Wing±Kristo�erson model to synchro-nization with periodic external events that was proposedby Vorberg and Wing (1994). In addition to the time-keeper and motor components, a linear phase correctionmechanism is assumed which is triggered by the last orthe last two synchronization errors. The model is testedin an experiment that contrasts synchronized and self-paced trapping, with response periods ranging from200±640 ms. The variances of timekeeper and motordelays and the error correction parameters were esti-mated from the auto-covariance functions of the inter-response intervals in continuation and the asynchroniesin synchronization. Plausible estimates for all parame-ters were obtained when equal motor variance was as-sumed for synchronization and continuation.Timekeeper variance increased with metronome period,but more steeply during continuation than duringsynchronization, suggesting that internal timekeepingprocesses are stabilized by periodic external signals.First-order error correction became more important asthe metronome period increased, whereas the contribu-tion of second-order error correction decreased. It isconcluded that the extended two-level model accountswell for both synchronization and continuation perfor-mance.
Introduction
The temporal precision of rhythmic motor behavior hasbecome an important topic in the behavioral and brainsciences (see, for instance, Tallal, Galaburda, LlinaÁ s, &von Euler, 1993). In psychological approaches to humanmotor behavior, a distinction has been made betweentasks which involve only timing constraints internal tothe person and tasks which involve some form of co-operative timing. Walking or running alone and playinga solo piece of music are examples of the former type oftask, whereas walking or running with other people,playing in an orchestra, or simply tapping in synchronywith a metronome are examples of the second type oftask.
The timing of periodic motor actions is often mod-eled in terms of an internal timekeeper which providesthe motor system with temporal information, which thenimplements the neuromuscular events that support theexecution of the intended action. One model of this kind,developed for self-paced periodic tapping, is the two-level model proposed by Wing and Kristo�erson (1973).Until recently (Mates, 1994a, b; Vorberg & Wing, 1994,1996; Pressing, 1998), however, theoretical work onsynchronization has ignored the two-level timingframework and the evidence put forward for it, althoughthere is no a priori reason why self-paced tapping shouldinvolve timing mechanisms fundamentally di�erent fromthose in synchronization with an external periodic sig-nal. Here we investigate the assumption that synchro-nization di�ers from self-paced tapping mainly by thecontribution of corrective mechanisms which keep thesubject's responses locked to the metronome events,counteracting timekeeper variability.
The aims of this paper are (1) to examine the em-pirical validity of such an extension of the two-levelmodel from self-paced to synchronic tapping and (2) todetermine whether the properties of the postulatedcentral timekeeper remain invariant over the two typesof task. In the ®rst part of the paper we summarize the
Psychological Research (2000) 63: 137±147 Ó Springer-Verlag 2000
A. Semjen (&)Centre National de la Recherche Scienti®que,Centre de Recherche en Neurosciences Cognitives,31, Chemin Joseph-Aiguier,13402 Marseille Cedex 20, Francee-mail: [email protected]
H.-H. SchulzePhilipps UniversitaÈ t Marburg, Fachbereich Psychologie
D. VorbergTechnische UniversitaÈ t Braunschweig,Institut fuÈ r Psychologie

original two-level timing model for self-paced tappingand its extension to synchronous tapping. Next wepresent an experiment that contrasts continuation andsynchronization performance at a range of tempi. Thenwe apply the model to the data and compare estimates oftimekeeper variability in synchronization and continu-ation. Finally, we discuss the merits and limitations ofthe two-level model for synchronization.
The two-level timing model for self-pacedand synchronized tapping
The timing model for continuation
Wing and Kristo�erson (1973) assumed that, in self-paced tapping, a succession of command pulses is gen-erated by an internal timekeeper. Each pulse initiates amotor implementation process which leads, after somedelay (``motor delay''), to an observable response. Theintervals marked o� by the timekeeper as well as themotor delays are assumed to be subject to chance ¯uc-tuations. If these assumptions (plus some technical as-sumptions, see Vorberg & Wing, 1994, 1996) hold, thenthe intervals between responses (``inter-response inter-vals'') are decomposable into contributions of thetimekeeper and of the motor system. Each inter-re-sponse interval In consists of the current timer intervalTn, plus the motor delay Mn+1 that leads to the responsewhich terminates In, minus the delay Mn that leads tothe response which initiates In:
In � Tn �Mn�1 ÿMn �1�Wing and Kristo�erson (1973) assumed that timekeeperintervals and motor delays are independent randomvariables with variance r2
T and r2M, respectively. They
also assumed mutual independence between thetimekeeper intervals and the motor delays. These as-sumptions imply that the variance of the observableinter-response intervals equals the timekeeper varianceplus twice the motor delay variance:
var�In� � r2T � 2r2
M �2�Moreover, the model predicts that adjacent inter-re-sponse intervals covary negatively with each other be-cause they have a motor delay in common, as is re¯ectedby the covariance
cov�In; In�1� � ÿr2M �3�
Note that in the model negative dependence betweensuccessive inter-response intervals follows from thetwo-level architecture and not from some feedbackmechanism. The degree of dependence that the modelcan accommodate is limited, however; the serial corre-lation between adjacent intervals is bounded by ÿ1=2and 0, and non-adjacent intervals must be uncorrelated.
Equations 2 and 3 make it possible to assess howmuch of the temporal variability in tapping is contrib-uted by the assumed timekeeper and how much by the
motor system. It is straightforward to estimate thetimekeeper variance, r2
T, and the motor delay variance,r2M, from the variance and the lag 1 auto-covariance of
the inter-response intervals. Empirical tests of the modelhave been reviewed by Vorberg and Wing (1994, 1996).
Modelling synchronization
As suggested above, synchronization might di�er fromself-paced tapping by the contribution of a mechanismthat locally corrects for synchronization errors. Such amechanism could take each successive asynchrony (i.e.,the temporal di�erence between a metronome signal andits corresponding tap) as input and correct the up-coming timekeeper interval appropriately. Voillaume(1971) has pointed out that, in general, the ``cost'' oferror correction is increased variability and dependencebetween inter-response intervals (IRIs). Therefore, theWing±Kristo�erson approach described above for sep-arating timekeeper from motor delay variability is validfor continuation data only. Applying it directly to IRIsequences from a synchronization experiment is likely toattribute part or all of the variance arising from errorcorrection to the peripheral motor processes and tounderestimate the variance contribution of the centraltimekeeper.
Extensions of the two-level model to synchronizationhave been examined by Vorberg and Wing (1994, 1996),Pressing (1998), and Vorberg and Schulze (1999). Herewe sketch the special case of isochronous tapping insynchrony with a perfect metronome. The two-level ar-chitecture is supplemented with a linear feedbackmechanism that corrects for phase errors. As in theoriginal model, observable inter-response intervals arelinear combinations of central (timekeeper) intervals andmotor delay components:
In � T�n �Mn�1 ÿMn �4�The central component T�n consists of an interval gen-erated by the internal timekeeper and a correction term.A crucial assumption is that the timekeeper mean isapproximately set to the metronome period and neednot be adjusted; in fact, only phase di�erences are evercorrected, whereas the timekeeper period is assumed toremain una�ected. The correction is done by subtractinga ®xed proportion, a, of the last synchronization error,An, from the timekeeper interval Tn:
T�n � Tn ÿ aAn �5�This ®rst-order phase correction scheme may be gener-alized such that the next-to-the-last synchronization er-rors are also taken into account:
T�n � Tn ÿ aAn ÿ bAnÿ1 �6�Obviously, the ®rst-order model can be derived from thesecond-order model as a special case by setting b � 0.
Like the model for self-paced tapping, the phase-correction model permits decomposition of the variance
138
in synchronization performance into separate sourcesdue to timekeeper and motor delays. The procedure ismore involved, however, and requires estimating thecorrection factors a and b as well as the variance pa-rameters r2
T and r2M from the empirical auto-covariance
function (ACVF) of the asynchronies.
Variability and dependence predictions
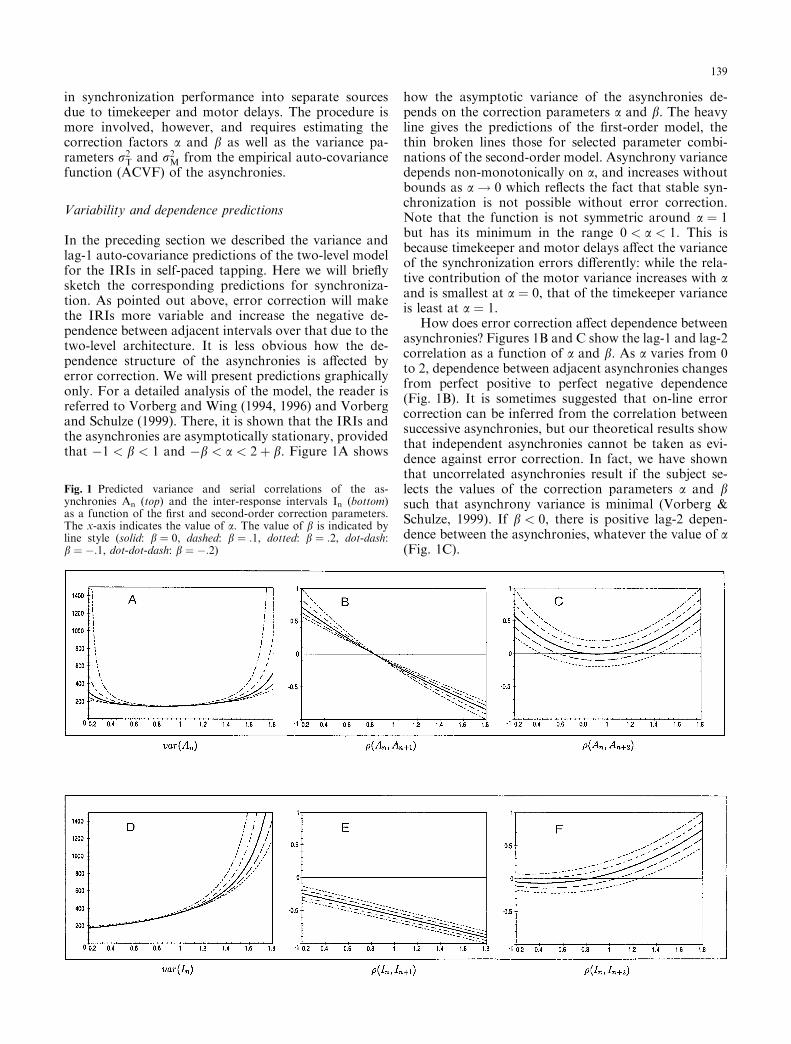
In the preceding section we described the variance andlag-1 auto-covariance predictions of the two-level modelfor the IRIs in self-paced tapping. Here we will brie¯ysketch the corresponding predictions for synchroniza-tion. As pointed out above, error correction will makethe IRIs more variable and increase the negative de-pendence between adjacent intervals over that due to thetwo-level architecture. It is less obvious how the de-pendence structure of the asynchronies is a�ected byerror correction. We will present predictions graphicallyonly. For a detailed analysis of the model, the reader isreferred to Vorberg and Wing (1994, 1996) and Vorbergand Schulze (1999). There, it is shown that the IRIs andthe asynchronies are asymptotically stationary, providedthat ÿ1 < b < 1 and ÿb < a < 2� b. Figure 1A shows
how the asymptotic variance of the asynchronies de-pends on the correction parameters a and b. The heavyline gives the predictions of the ®rst-order model, thethin broken lines those for selected parameter combi-nations of the second-order model. Asynchrony variancedepends non-monotonically on a, and increases withoutbounds as a! 0 which re¯ects the fact that stable syn-chronization is not possible without error correction.Note that the function is not symmetric around a � 1but has its minimum in the range 0 < a < 1. This isbecause timekeeper and motor delays a�ect the varianceof the synchronization errors di�erently: while the rela-tive contribution of the motor variance increases with aand is smallest at a � 0, that of the timekeeper varianceis least at a � 1.
How does error correction a�ect dependence betweenasynchronies? Figures 1B and C show the lag-1 and lag-2correlation as a function of a and b. As a varies from 0to 2, dependence between adjacent asynchronies changesfrom perfect positive to perfect negative dependence(Fig. 1B). It is sometimes suggested that on-line errorcorrection can be inferred from the correlation betweensuccessive asynchronies, but our theoretical results showthat independent asynchronies cannot be taken as evi-dence against error correction. In fact, we have shownthat uncorrelated asynchronies result if the subject se-lects the values of the correction parameters a and bsuch that asynchrony variance is minimal (Vorberg &Schulze, 1999). If b < 0, there is positive lag-2 depen-dence between the asynchronies, whatever the value of a(Fig. 1C).
Fig. 1 Predicted variance and serial correlations of the as-ynchronies An (top) and the inter-response intervals In (bottom)as a function of the ®rst and second-order correction parameters.The x-axis indicates the value of a. The value of b is indicated byline style (solid: b � 0, dashed: b � :1, dotted: b � :2, dot-dash:b � ÿ:1, dot-dot-dash: b � ÿ:2)
139

Figure 1D illustrates the variance predictions for theIRIs. In contrast to the asynchronies, asymptotic IRIvariance increases monotonically with a. The least IRIvariance is achieved without error correction, but this isincompatible with keeping the asynchrony mean andvariance within bounds. This shows that there are per-formance goals in synchronization experiments whichcon¯ict with each other. Figures 1E and F illustrate thedependence predictions. Increasing the gain of ®rst-ordercorrection magni®es the negative correlation betweenadjacent IRIs, and the e�ect is reinforced or attenuatedby changes in the gain of second-order correction. As forthe asynchronies, b < 0 implies positive lag-2 depen-dence between IRIs, whatever the value of a (Fig. 1F).
Parameter estimation
In this section, we will sketch several problems thatpertain to the estimation of the parameters of the modeland their interpretation. The reader not interested intechnical details can omit this section without loss ofcontinuity.
From here on, we will simplify our notation bywriting m and t for the motor and timekeeper varianceparameters, that is, m � r2
M and t � r2T. A particular
model is thus characterized by the parameter vector(a; b;m; t).
Estimation problems
Calculating quantitative predictions from a model for agiven set of parameters is straightforward, but the re-verse problem, that is, inferring the parameters fromdata, is more complex. There are three kinds of prob-lems from which di�culties can arise: identi®ability,parameter tradeo�s, and sampling variability.
If the parameters are speci®ed, the theoretical ACVFis uniquely determined by the model. The parameters ofa model are called identi®able if the opposite is also true,that is, if it is possible to uniquely determine their valuesfrom the theoretical ACVF. It can be shown that theparameters of both the ®rst-order and the second-ordererror correction model are identi®able almost every-where in the parameter space (Vorberg & Schulze, 1999).
In practice, identi®ability is not enough, because itdoes not safeguard against numerical problems. Thesemay arise when di�erent parameter vectors lead tomathematically distinct yet numerically close predic-tions. For a given goodness-of-®t measure, compare theACVF predictions of the model for di�erent parametervectors. Let (a0; b0;m0; t0) be the ``true'' parameters ofthe model. How much will the predictions change if wereplace them by (a; b;m; t)? Due to limited numericalresolution, in general it will not be possible to distin-guish between model versions if their predictions di�er(in terms of the ®t measure) by less than some smallvalue. For a given numerical precision e we de®ne the
tradeo� region by the set of all vectors (a; b;m; t) whichdi�er by less than e in their predictions from(a0; b0;m0; t0). Obviously, all models in the tradeo� re-gion will ®t an empirical ACVF about equally as wellas the true model, implying that changes in a parametermay be compensated for by changes in the other pa-rameters without appreciable loss of ®t. Numericalanalyses have indeed revealed huge tradeo� regions inthe space of the second-order model (Vorberg &Schulze, 1999), which make interpretation of the valuesof parameter estimates risky. It is possible to overcomethese problems by restricting the parameter space or byimposing invariance constraints across conditions, aswe will show below.
In contrast to identi®ability and tradeo� problemswhich depend on how predictions and parameters arerelated, sampling variability problems arise in stochasticmodels because of the inherent noise in the data. Basi-cally, they relate to the question as to whether we canrecover the parameter values from the data if we knowthe true model. We provide some examples from MonteCarlo studies of the Wing±Kristo�erson continuationmodel and of the synchronization model (for details, seeVorberg & Schulze, 1999).
Simulating continuation
We simulated self-paced and synchronized tappingfrom sequences of independent timekeeper intervalsand motor delays, each variable generated from agamma distribution with k � 4 and scaled to givevariances of m � 52 and t � 72 or 152. A simulatedcontinuation experiment involved sequences of 35 re-sponses each, yielding N � 34 IRIs. For a given pa-rameter combination, Ns � 800 such sequences weregenerated. From each IRI series, we calculated unbi-ased estimates of m and t (see Vorberg & Wing, 1996).We determined the frequency with which the varianceestimators assumed negative values. This can occureven when the underlying model is known to be freefrom defects and the estimators provide perfect long-run estimates of the variances, simply because esti-mates calculated on sample data must scatter aroundtheir expected values. The question is how likely suchapparently defective estimates are, i.e., what is the riskof erroneously rejecting the right model on such evi-dence? It turns out that the risk is unexpectedly high,even for reasonable parameter values and sample sizes:Variance estimates were negative in 8.25% of the ex-periments with t � 72 and m � 52, but the percentagerose to 30.5% when the timekeeper variance was in-creased to t � 152. Note that in the long run theseestimates are quite accurate: the mean of all 800 esti-mates came close to the underlying parameters t (7.02
and 14.92 ms2, respectively) and m (4.972 and5.152 ms2), which strongly argues against the commonpractice of discarding or replacing negative estimateswith 0.
140

Simulating synchronization
Timekeeper intervals and motor delays were generatedas in continuation, except that as a simulated seriesunfolded, the metronome-tap asynchrony was calculatedfor each tap and used to correct the up-coming time-keeper interval. A simulated experiment consisted ofNs � 100 sequences of 31 responses each (yielding N �30 asynchronies). For each series, we calculated thesample ACVF of the asynchronies and averaged themacross all Ns series. Parameters were estimated by ®ttingthe model to the mean ACVF of the simulated data atlags 0 through 3; details are described in the Model ®tand parameter estimates section. Sampling distributionsof the estimators were computed by replicating eachexperiment 1000 times. Note that the sample size used(100 sequences of 31 responses) is much larger than inmost psychological experiments, which implies that evenmore drastic sampling problems have to be expected inrealistic settings than those reported here.
Severe estimation problems were noticed for themodelwith second-order error correction, in particular when thetrue correction parameters were small, such as a < :2 andjbj < :2; themain problem is the extreme dependence. Fora model with some ®xed set of parameters (a; b;m; t), theestimates (a; b; m; t) are strongly correlated with eachother across di�erent realizations. Consequently, esti-mation error in one of the parameters is likely to a�ect theother estimates aswell. For example, overestimating awilltend to produce in¯ated timekeeper variance and lowmotor variance estimates. Dependence is less extreme forlarger values of the correction parameters, but still ap-
preciable, as Fig. 2 shows for simulations based on a � :4and b � :3. This dependence between the estimators ismainly due to the parameter-tradeo� problems men-tioned above, as was evidenced by the fact that it becomesstronger (rather than weaker) with sample size. Note howmuch uncertainty in the remaining parameters is reducedif one of them is known. As an example, consider thespread in the estimates of a; b, and t that remains ifwe inferfrom continuation data, say, that m is close to 25.
Deciding from the parameter estimates whether errorcorrection is ®rst-order or second-order is likely to fail,as the following example demonstrates. We ®tted thesecond-order model (b free) to data generated by the®rst-order model (b � 0). The results are devastating,with estimates ¯uctuating wildly around the true pa-rameter values (see Fig. 3). Note that the estimatespartition into two disjoint sets: If b happens to be esti-mated correctly, the other estimators are well behaved.However, if by chance the estimate of b is far from thetrue parameter value 0, there is little hope of recoveringthe values of the other parameters. Fitting the ®rst-ordermodel as well and comparing the resulting goodness-of-®t measures seems a more promising approach for de-ciding whether b � 0. This is the approach we use below.
We tested the empirical validity of the synchroniza-tion model and examined how the estimated model pa-rameters depend on tempo in synchronized tapping in anexperiment that uses the synchronization-continuationparadigm, with tapping rate as the main independentvariable. We were particularly interested in studyingparameter invariance across task (synchronization vs.continuation) and tempo conditions.
Fig. 2 Estimator variability inthe second-order model. Histo-grams and scatter plots of pa-rameter estimates in N = 1000simulated experiments witht � 100, m � 25, a � :4 andb � :3
141

Method
Subjects. Twelve right-handed subjects (3 women and 9 men, agedbetween 16 and 56) participated in the experiment. The data of 2male subjects were discarded from the analyses because of theirerratic performance in the synchronization task. All remainingsubjects (N = 10) had at least three years of experience in playinga musical instrument (piano, guitar, or percussion); all but one(the ®rst author) were naive with respect to the aim of theexperiment.
Apparatus and measurements. The response key was a circulartouch plate mounted on a wooden support ®xed to a table. Thesubject was seated with his or her forearm lying on the table andthe index ®nger placed above the response key. The response keywas interfaced to an Olivetti PC (M 290). Any onset and o�set ofcontact between the ®nger and the response key was detected. In-ter-response intervals were measured to the nearest millisecondbetween successive contact onsets. Via a sound card, the computeralso controlled the periodic auditory clicks (1000-Hz square waves,3-ms duration) which served as metronome signals. The clicks werepresented through loudspeakers above a video monitor, about60 cm in front of the subject. Subjects' responses did not producetones, but the sound of the ®nger touching the response plate wasclearly audible.
Task and design. The task was to tap periodically on the responsekey, ®rst in synchrony with the clicks of the metronome (``syn-chronization''), and then without the metronome (``continuation'').At the start of a trial, the metronome rate in e�ect (period = 200,240, 280, 320, 400, 480, 560, or 640 ms) was shown on the screen.After 10 s, the screen was cleared and the metronome clicks began.Subjects could listen to the metronome until they felt ready to enter
at the required tempo. They were urged to maintain synchroniza-tion between clicks and taps as precisely as possible throughout theseries. After 35 taps the metronome was turned o�, and the subjectcontinued for another 35 cycles at the same tempo. Subjects were toavoid any interruption, acceleration, or deceleration of the tapswhen passing from synchronization to continuation. A continuoustone of 500 ms duration signaled the end of the trial.
The IRIs were checked on-line. Trials were interrupted if anIRI deviated by more than 50% from the target value. In suchcases, a message indicated the defective value, and the trial wasimmediately repeated. At the end of each trial, performance feed-back was given. The coe�cient of variation of the IRIs (i.e., 100*standard deviation/mean) was shown, with a message whether theseries was accepted. The aim of this feedback procedure was tomotivate the subjects to maintain high-level performancethroughout the experimental session. Series with a coe�cient largerthan 7% occurred only in a few instances; they were immediatelyrepeated.
The experiment consisted of two sessions of roughly one houreach, separated by one to seven days. The sessions di�ered in themetronome period used. The shorter periods (200, 240, 280, and320 ms) were presented in one session, the longer ones (400, 480,560, and 640 ms) in the other. Half of the subjects started with theshorter periods, and the other half with the longer periods. Withina session, each subject performed a total of 64 correct trials, 16 foreach period. Within a block of 4 successive trials, the periods werepresented once each, in random order.
Data analysis. The experimental data consisted of the IRIs plus theasynchronies during synchronization. Asynchronies were de®ned asthe delay between tap onset and metronome onset, with negativevalues indicating that the tap preceded the metronome. The ®rst®ve values in each sequence were discarded, leaving 30 inter-re-sponse times or asynchronies per series. For each single series,means and lag 0 to 3 serial auto-covariances of the IRIs and theasynchronies were calculated, using the estimators given by Vor-
Fig. 3 Results of ®tting thesecond-order model to datagenerated from the ®rst-ordermodel. Histograms and scatterplots of estimates in N = 1000simulated experiments witht � 100, m � 25, a � :2 andb � 0
142
berg and Wing (1996). These statistics were then averaged over the16 trials for a given tempo ´ task ´ subject combination.
Results
IRI and asynchrony means
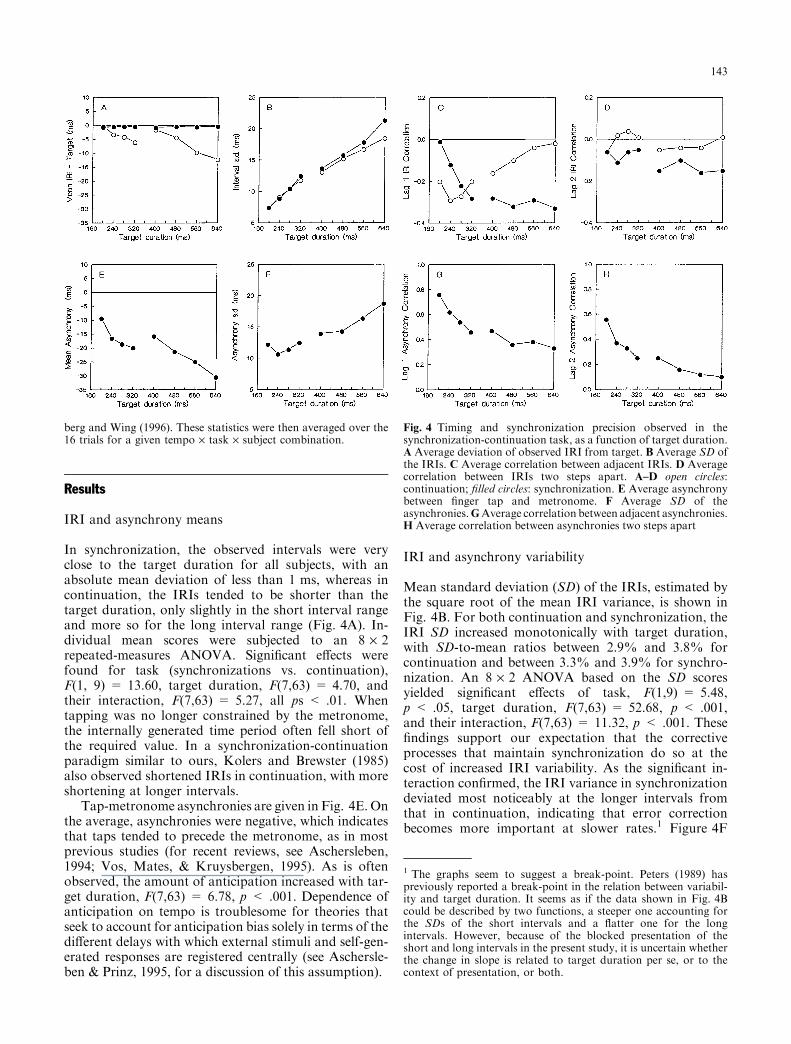
In synchronization, the observed intervals were veryclose to the target duration for all subjects, with anabsolute mean deviation of less than 1 ms, whereas incontinuation, the IRIs tended to be shorter than thetarget duration, only slightly in the short interval rangeand more so for the long interval range (Fig. 4A). In-dividual mean scores were subjected to an 8 ´ 2repeated-measures ANOVA. Signi®cant e�ects werefound for task (synchronizations vs. continuation),F(1, 9) = 13.60, target duration, F(7,63) = 4.70, andtheir interaction, F(7,63) = 5.27, all ps < .01. Whentapping was no longer constrained by the metronome,the internally generated time period often fell short ofthe required value. In a synchronization-continuationparadigm similar to ours, Kolers and Brewster (1985)also observed shortened IRIs in continuation, with moreshortening at longer intervals.
Tap-metronome asynchronies are given in Fig. 4E. Onthe average, asynchronies were negative, which indicatesthat taps tended to precede the metronome, as in mostprevious studies (for recent reviews, see Aschersleben,1994; Vos, Mates, & Kruysbergen, 1995). As is oftenobserved, the amount of anticipation increased with tar-get duration, F(7,63) = 6.78, p < .001. Dependence ofanticipation on tempo is troublesome for theories thatseek to account for anticipation bias solely in terms of thedi�erent delays with which external stimuli and self-gen-erated responses are registered centrally (see Aschersle-ben & Prinz, 1995, for a discussion of this assumption).
IRI and asynchrony variability
Mean standard deviation (SD) of the IRIs, estimated bythe square root of the mean IRI variance, is shown inFig. 4B. For both continuation and synchronization, theIRI SD increased monotonically with target duration,with SD-to-mean ratios between 2.9% and 3.8% forcontinuation and between 3.3% and 3.9% for synchro-nization. An 8 ´ 2 ANOVA based on the SD scoresyielded signi®cant e�ects of task, F(1,9) = 5.48,p < .05, target duration, F(7,63) = 52.68, p < .001,and their interaction, F(7,63) = 11.32, p < .001. These®ndings support our expectation that the correctiveprocesses that maintain synchronization do so at thecost of increased IRI variability. As the signi®cant in-teraction con®rmed, the IRI variance in synchronizationdeviated most noticeably at the longer intervals fromthat in continuation, indicating that error correctionbecomes more important at slower rates.1 Figure 4F
Fig. 4 Timing and synchronization precision observed in thesynchronization-continuation task, as a function of target duration.A Average deviation of observed IRI from target. B Average SD ofthe IRIs. C Average correlation between adjacent IRIs. D Averagecorrelation between IRIs two steps apart. A±D open circles:continuation; ®lled circles: synchronization. E Average asynchronybetween ®nger tap and metronome. F Average SD of theasynchronies.GAverage correlation between adjacent asynchronies.H Average correlation between asynchronies two steps apart
1 The graphs seem to suggest a break-point. Peters (1989) haspreviously reported a break-point in the relation between variabil-ity and target duration. It seems as if the data shown in Fig. 4Bcould be described by two functions, a steeper one accounting forthe SDs of the short intervals and a ¯atter one for the longintervals. However, because of the blocked presentation of theshort and long intervals in the present study, it is uncertain whetherthe change in slope is related to target duration per se, or to thecontext of presentation, or both.
143

shows that the SD of the asynchronies also followed anincreasing trend in the 240 to 640-ms target intervalrange. An ANOVA indicated that the variation of theSD with the target interval was highly signi®cant,F(7,63) = 30.71, p < 0.001.
IRI and asynchrony dependence
For each subject, the correlation between adjacent IRIswas calculated from the average ACVF at lags 1 and 0,that is r = ACVF(1)/ACVF(0). These correlation co-e�cients, averaged over subjects, are presented inFig. 4C. As expected, adjacent IRIs were negativelycorrelated in both tasks. In synchronization, negativedependence became stronger with target duration, but incontinuation it diminished. (For some subjects, evenpositive correlations were observed at the slowest rates.)An ANOVA of the z-transformed correlations backsthese observations, yielding signi®cant e�ects for task,F(1,9) = 5.94, p < 0.5, target duration, F(7,63) = 3.35,p < .01, and their interaction, F(7,63) = 31.25, p <.001. Figure 4D shows the corresponding correlationsfor IRIs separated by one intervening interval, calcu-lated in an analogous fashion. For continuation, thesecorrelations came close to zero, as expected, whereas forsynchronization, they were still negative. An ANOVAcon®rmed this di�erence by yielding a signi®cant e�ectfor task, F(1,9) = 13.99, p < .01; the e�ect of targetduration did not reach statistical signi®cance (p > .05).
Figures 4G and 4H show the dependence betweenadjacent asynchronies (lag-1 correlation) and betweenasynchronies two steps apart (lag-2 correlation). Thelag-1 correlation was generally positive and was inv-ersely related to target interval. Adjacent asynchronieswere more strongly correlated than asynchronies twosteps apart. ANOVAs con®rmed that the e�ects of tar-get interval was statistically signi®cant for both lag-1,F(7,63) = 29.06, and lag-2, F(7,63) = 17.22, correla-tion, ps < .001.
Model ®t and parameter estimates
As sketched above, parameters were estimated by ®ttingthe predicted to the empirical IRI and asynchronyACVFs in continuation and synchronization, respec-tively. Estimator bias was taken into account byfollowing the procedure recommended by Vorberg andWing (1996): (a) for a given parameter vector, the the-oretical ACVF is computed; (b) from this, the expected
sample ACVF (which includes bias) is derived; (c)goodness-of-®t is evaluated by comparing the expectedand observed ACVFs at lags 0 through 3, with thesummed squared deviations as ®t measure. Numericaloptimization (Nelder-Mead simplex search, procedure,fmins, MATLAB, 1995) was used for determining theparameter vector that minimizes the ®t measure.
Above, we discussed problems in recovering themodel parameters from data which may arise fromsampling variability and parameter tradeo�s. To controltheir impact, we ®tted the second-order model with un-constrained parameters and compared its ®t and para-meter estimates to those under constraints: (1) withmotor variance invariant across tasks, that is, equalmotor variance in synchronization and continuation, and(2) with ®rst-order error correction only, that is, b � 0.At each target duration, we thus estimated six para-meters (mc;ms; tc; ts; a; b) for the general model, ®veparameters (m; tc; ts; a; b) under constraint 1, and fourparameters (m; tc; ts; a) under the joint constraint 1 & 2;the indices c and s stand for continuation and synchro-nization. The variances mc and tc (which correspond tothe parameters in the Wing±Kristo�erson model) wereestimated directly from the lag-0 and lag-1 IRIauto-covariances. Parameters were estimated separatelyfor each subject.
Table 1 reports the ®t of the three versions of themodel averaged across subjects. As is to be expected, themodel without constraints gave by far the best overall ®tto the data, but the parameter patterns that resulted forsynchronization are hard to understand and seem to bea�ected by the parameter tradeo�s noted in our MonteCarlo studies: 27 out of 80 estimates of the motor vari-ance were zero or negative; moreover, motor variancetended to increase with target duration almost as muchas timekeeper variance (cf. Fig. 5A and B). This wasunexpected, because in continuation the parameter es-timates showed the usual dissociation pattern, that is,timekeeper variance increasing with target duration, andmotor variance approximately constant.
These problems were greatly reduced when con-straints were imposed on the motor variance. Assumingmc � ms � m seems reasonable, because the samemovements are required in synchronization as in con-tinuation, but estimating a single parameter m for syn-chronization and continuation at a given tempo still ledto many zero or negative estimates. Therefore, we esti-mated mc from the continuation data and kept ms ®xedat this value in estimating the remaining parameters ts, a,and b from the synchronization data.
Table 1 Root mean squaresConstraint Target duration (ms)
200 240 280 320 400 480 560 640
None 7.2 8.6 8.9 10.1 6.6 18.0 10.9 21.2ms � mc 20.0 19.9 23.4 31.8 50.5 41.5 40.9 99.9ms � mc, b � 0 29.0 49.5 46.8 46.3 58.9 45.0 58.5 115.7
144
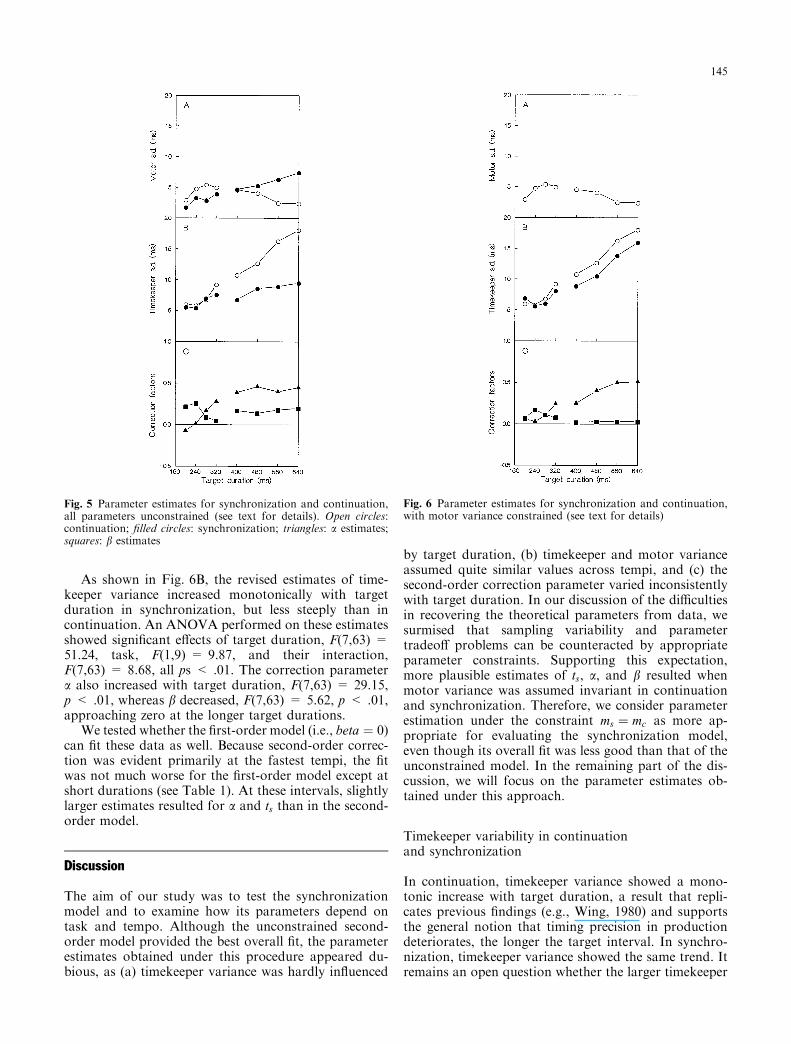
As shown in Fig. 6B, the revised estimates of time-keeper variance increased monotonically with targetduration in synchronization, but less steeply than incontinuation. An ANOVA performed on these estimatesshowed signi®cant e�ects of target duration, F(7,63) =51.24, task, F(1,9) = 9.87, and their interaction,F(7,63) = 8.68, all ps < .01. The correction parametera also increased with target duration, F(7,63) = 29.15,p < .01, whereas b decreased, F(7,63) = 5.62, p < .01,approaching zero at the longer target durations.
We tested whether the ®rst-order model (i.e., beta � 0)can ®t these data as well. Because second-order correc-tion was evident primarily at the fastest tempi, the ®twas not much worse for the ®rst-order model except atshort durations (see Table 1). At these intervals, slightlylarger estimates resulted for a and ts than in the second-order model.
Discussion
The aim of our study was to test the synchronizationmodel and to examine how its parameters depend ontask and tempo. Although the unconstrained second-order model provided the best overall ®t, the parameterestimates obtained under this procedure appeared du-bious, as (a) timekeeper variance was hardly in¯uenced
by target duration, (b) timekeeper and motor varianceassumed quite similar values across tempi, and (c) thesecond-order correction parameter varied inconsistentlywith target duration. In our discussion of the di�cultiesin recovering the theoretical parameters from data, wesurmised that sampling variability and parametertradeo� problems can be counteracted by appropriateparameter constraints. Supporting this expectation,more plausible estimates of ts, a, and b resulted whenmotor variance was assumed invariant in continuationand synchronization. Therefore, we consider parameterestimation under the constraint ms � mc as more ap-propriate for evaluating the synchronization model,even though its overall ®t was less good than that of theunconstrained model. In the remaining part of the dis-cussion, we will focus on the parameter estimates ob-tained under this approach.
Timekeeper variability in continuationand synchronization
In continuation, timekeeper variance showed a mono-tonic increase with target duration, a result that repli-cates previous ®ndings (e.g., Wing, 1980) and supportsthe general notion that timing precision in productiondeteriorates, the longer the target interval. In synchro-nization, timekeeper variance showed the same trend. Itremains an open question whether the larger timekeeper
Fig. 5 Parameter estimates for synchronization and continuation,all parameters unconstrained (see text for details). Open circles:continuation; ®lled circles: synchronization; triangles: a estimates;squares: b estimates
Fig. 6 Parameter estimates for synchronization and continuation,with motor variance constrained (see text for details)
145

variability in continuation than in synchronization(tc > ts) is an artifact of parameter tradeo�s or indicatesa genuine increase in timekeeper stability due to themetronome. If parameter tradeo� was the main cause ofthe lower timekeeper variance estimates in synchroni-zation, constraints on the parameter space shouldgreatly in¯uence the estimates. However, we did notobserve any systematic change in the value ts in thecritical range of long intervals, whether second-order or®rst-order error correction was assumed. We thereforeconclude that the lower timekeeper estimates in syn-chronization indicate improved timing precision whenthere is external temporal information.2 A possible ex-planation of this ®nding is that the target duration canbe repeatedly encoded when the metronome is present,allowing for more accurate representation or more pre-cise reproduction of that duration. This conjecture issupported by empirical results which show that multiplepresentation of the standard leads to less variable per-ception and production of time intervals (Ivry & Ha-zeltine, 1995).
Error correction at slow and fast tempi
Under the constraint ms � mc the correction factor a in-creased from 0.06 at the shortest target duration to 0.51 atthe longest duration. In contrast, b showed the reversetrend, tending to zero at long durations. The latter ®ndingsuggests that at fast tempi the e�ects of detecting andcorrecting tap-sound asynchronies are delayed by oneresponse cycle, perhaps due to the time constants of theerror correction loop.A related e�ect has beenobserved insynchronization with a randomly ¯uctuatingmetronome,where the maximum cross-correlation between met-ronome interval and IRI was found to shift from lag 1 tolag 2 at fast metronome rates (Schulze, 1992).
First-order error correction increasing with targetduration would be predicted if subjects tried to optimizesynchronization performance. For example, the optimalstrategy for minimizing asynchrony variance is to selectb � 0 and a � a�, where a� < 1 depends on the ratio oftimekeeper and motor variance (Vorberg & Schulze,1999). In the present study, this ratio increased withtarget duration, as m was approximately constant and tincreased. The increase of a with target duration may
thus indicate that subjects tried to keep asynchronyvariance at a low level, even though they undershot theoptimal a� values considerably: At the four longest du-rations, where b was close to 0, the average estimates ofa were 0.25, 0.40, 0.50, and 0.51 (see Fig. 6B). From thevariance estimates of t and m, the optimal a� is computedas 0.78, 0.84, 0.94, and 0.95.
The two-level timing model and alternative accounts
The two-level timing model has proved successful inseparating the timekeeping and error correction pro-cesses that underlie synchronization with a metronome.The model provides plausible estimates for the time-keeper variance t and the correction parameters a and bwhen reasonable constraints are imposed on the pa-rameters. Whereas, in the present study, these parame-ters were estimated from the auto-covariance pattern ofthe stationary time series of asynchronies, in a previousstudy we estimated them from the pattern of mean asyn-chronies during the initial (transient) phase of synchro-nization. In that study, the metronome was activatedafter a series of self-paced ®nger taps performed at thesubject's preferred tempo. The metronome period wasequal to the mean of the self-paced intervals and rangedfrom 428 to 725 ms in a group of six subjects(mean = 565 ms). The initial phase di�erence of themetronome from the taps was varied across trials. Theestimates of the correction terms, averaged over sub-jects, were a � 0:29 and b � 0:13 (Semjen, Vorberg, &Schulze, 1998). Experiments are in progress to clarifywhether a and b are invariant during the transient andasymptotic phases of synchronization.
From the increase in timekeeper precision in syn-chronization as compared to continuation, we concludethat internal timekeeping processes are stabilized byperiodic external signals, an intuitively appealing con-clusion that is corroborated by previous ®ndings (Ivry &Hazeltine, 1995), but goes beyond the basic assumptionsof the two-level framework of timing. In this framework,the metronome is not supposed to exert any in¯uence onthe internal timekeeper beyond that of initially settingthe target interval. Rather, its function is to provide theperceptual cues against which synchronization is as-sessed. The internal timekeeper is self-triggering in thesense that the end of a central period Tn initiates the nextperiod Tn+1, implying that the timekeeper is a type ofinterval generator.3 It is remarkable how well such aninterval generator, augmented by a ®rst-order feedbackmechanism for local phase adjustment, accounts for thestochastic aspects of synchronization performance.Other models have considered the internal timekeeper to
2 In a simulation study, we generated synchronization-continuationtapping sequences using parameter values (t � 122, m � 52, a � :5,b � 0) close to those obtained in the present experiment for thelongest interval. We generated 35 taps per sequence, 16 sequencesper block, and a total of N = 80 blocks. For each block, m, wasrecovered from the continuation ACVF and used as a ®xedparameter when estimating t, a, and b from the synchronizationACVF. In ®tting the second-order model, ts was slightly lower thantc and b was estimated slightly larger (0.03) than the true value. In®tting the ®rst-order model, ts was slightly higher than tc. Since®tting the ®rst-order or second-order model to our experimentaldata did not lead to similar changes in the ts estimates, we considerit highly unlikely that the observed relationship tc > ts was due toan estimation artifact.
3 This makes unnecessary to incorporate any other ``reset event'' inthe model. In ``tap reset'' models, the beginning of the timekeeperintervals is marked o� by response events, whereas in ``metronomereset'' models, they are marked o� by registered external events(see, e.g., Hary & Moore, 1987; Schulze, 1992).
146

be a limit-cycle oscillator coupled by resonance to ex-ternal timing signals. One such approach has provedsuccessful in a connectionist model of the perception ofmusical meter (Large & Kolen, 1994), but it remains tobe seen whether such a model can also account for thevariability and dependence patterns in IRIs and as-ynchronies as well.
Acknowledgements Preparation of this article was partially sup-ported by a CNES (Centre National d'Etudes Spatiales) researchgrant (No. 95/CNES/0244) to A. Semjen.
References
Aschersleben, G. (1994). A�erente Informationen und die Syn-chronisation von Ereignissen [A�erent information and thesynchronization of events]. Frankfurt am Main: Peter Lang.
Aschersleben, G., & Prinz, W. (1995). Synchronizing actions withevents. The role of sensory information. Perception & Psycho-physics, 57, 305±317.
Hary, D., & Moore, P. (1987). Synchronizing human movements toan external clock source. Biological Cybernetics, 56, 305±311.
Ivry,R. B.,&Hazeltine,R. E. (1995). The perception andproductionof temporal intervals across a range of duration: Evidence for acommon timing mechanism. Journal of Experimental Psycholo-gy: Human Perception and Performance, 21, 3±18.
Kolers, P. A., & Brewster, J. M. (1985). Rhythms and responses.Journal of Experimental Psychology: Human Perception andPerformance, 11, 150±167.
Large, E. W., & Kolen, J. F. (1994). Resonance and the perceptionof musical meter. Connection Science, 16, 177±208.
Mates, J. (1994a). A model of synchronization of motor acts tostimulus sequence: I. Timing and error corrections. BiologicalCybernetics, 70, 463±473.
Mates, J. (1994b). A model of synchronization of motor acts tostimulus sequence: II. Stability analysis, error estimation andsimulations. Biological Cybernetics, 70, 475±484.
Peters, M. (1989). The relationship between variability of interlapintervals and interval duration. Psychological Research, 51,38±42.
Pressing, J. P. (1998). Error correction processes in temporal pat-tern production. Journal of Mathematical Psychology, 42,63±101.
Schulze, H. H. (1992). The error correction model for the trackingof a random metronome: Statistical properties and empiricaltest. In: F. Macar, V. Pouthas, & W. J. Friedman (Eds.), Time,action, and cognition: Towards bridging the gap (pp. 275±286).Dordrecht: Kluwer Academic Publisher.
Semjen, A., Vorberg, D., & Schulze, H. H. (1998). Getting syn-chronized with the metronome: Comparisons between phaseand period correction. Psychological Research, 61, 44±55.
Tallal, P., Galaburda, A. M., LlinaÁ s, R., & von Euler, C. (Eds.).Temporal information processing in the nervous system. Annalsof the New York Academy of Sciences, 1993, Vol. 682.
Voillaume, C. (1971). ModeÁ les pour l'e tude de la re gulation demouvements cadence s. L'AnneÂe Psychologique, 71, 347±358.
Vorberg, D., & Schulze, H. H. (1999). The two-level timing modelfor synchronization: Stochastic properties and parameter esti-mation. Manuscript submitted for publication.
Vorberg, D., & Wing, A. (1994). Modelle fuÈ r VariabilitaÈ t undAbhaÈ ngigkeit bei der zeitlichen Steuerung. In H. Heuer & S. W.Keele (Eds.), Psychomotorik: EnzyklopaÈ die der PsychologieVol. 3 (pp. 223±320), GoÈ ttingen: Hogrefe.
Vorberg, D., & Wing, A. (1996). Modeling variability and depen-dence in timing. In H. Heuer & S. W. Keele (Eds), Handbook ofperception and action: Vol. 3. Motor Skills (pp. 181±262).London: Academic Press.
Vos, P. G., Mates, J., & van Kruysbergen, N. W. (1995). Theperceptual centre of a stimulus as the cue for synchronization toa metronome: Evidence from asynchronies. The QuarterlyJournal of Experimental Psychology, 48A, 1024±1040.
Wing, A. M. (1980). The long and short of timing in responsesequences. In G. E. Stelmach & J. Requin (Eds), Tutorials inmotor behaviour (pp. 469±486). Amsterdam: North Holland.
Wing, A. M., & Kristo�erson, A. B. (1973). Response delays in thetiming of discrete motor responses. Perception & Psychophysics,14, 5±12.
147





























