Embed Size (px)
Citation preview

The Departments of Urology' and Clinical Chemistry2, University of Lund, Malmo General Hospital, Malmo/Sweden
Three predominant prostatic proteins
Drei vorherrschende Prostataproteine
P.-A. Abrahamsson' and H. Lilja2
Key words. Prostatic acid phosphatase, kallikreins, prostate-specific antigen, serine proteases, y-seminopro- tein, P-microseminoprotein, PSP,,, P-inhibin.
Summary. Prostatic acid phosphatase (PAP), prostate-specific antigen (PSA; or y- seminoprotein), and 0-microseminoprotein (P-MSP; PSP,, or P-inhibin) are the three predominant proteins secreted by the normal human prostate gland. In the epithelium of normal and hyperplastic prostatic acini and ducts PAP, PSA and P-MSP have an identical immunohistochemical localization. Highly differentiated (grade I) carcinomas contain an almost equal number of PAP-, PSA- and 0-MSP-immunoreactive cells; the incidence of these cells is lower and they display a greater staining variability in the moderately and poorly (grade 11-111) differentiated tumours. Especially in poorly differentiated tumours PSA seems to be a more sensitive immunohistochemical marker than PAP or prostatic carcinomas. Moreover, the use of PAP as a marker for prostatic carcinomas is complicated by the reported structural similarities between the prostatic secreted acid phosphatase and lysosomal acid phosphatase occurring in all tissues. The use of P-MSP as a marker for prostatic carcinomas may be limited by indications of non-prostatic production of this protein.
Zusammenfassung. Saure Prostata-Phosphatase (PAP), Prostata-spezifisches Antigen (PSA; oder y-Seminoprotein) und P-Mikroseminoprotein (P-MSP; PSP,, oder P-Inhibin) sind die drei vorherrschenden Proteine, die von der normalen Prostata sezerniert werden. Im Epithel normaler und hyperplastischer Prostata-Azini und Drusengange haben PAP, PSA und 0-MSP eine identische immunhistochemische Lokalisation. Hoch differenzierte (Grad I) Karzinome enthalten annahernd gleiche Anzahlen von PAP-, PSA- und /?-MSP- immunreaktiven Zellen; das Auftreten dieser Zellen in maBig und wenig differenzierten (Grad 11, 111) Tumoren ist geringer und sic zeigen eine erhohte Variabilitat in der Anfarbung. Vor allem in wenig differenzierten Prostatakarzinomen scheint PSA ein sensitiverer immunhistochemischer Marker als PAP zu sein. Uberdies ist die Anwendung von PAP als Marker fur Prostatakarzinome durch die beschriebene strukturelle Ahnlich- keit zwischen der sekretorischen Prostataphosphatase und der lysosomalen sauren Phos- phatase aus allen ubrigen Geweben kompliziert. Die Anwendung von P-MSP als Marker fur Prostatakarzinome konnte durch die extraprostatische Produktion dieses Proteins belastet sein.

Three predominant prostatic proteins 123
Introduction
Results obtained from the electrophore- tic analyses of split ejaculates (Dub6 et al., 1987a) and of azoospermic ejaculates ob- tained from two subjects with functional deficiency of aplasia of the seminal vesi- cles (Lilja & Abrahamsson, 1988) have demonstrated three proteins to be the predominant protein components in the human prostatic secretion. These pro- teins are the 100 kDa prostatic acid phos- phatase (PAP) (Choe & Rose, 1982), the 33 kDa prostate-specific antigen (PSA) (Wang et al., 1979) and the 94-amino acid P-microseminoprotein (Tsuda et al., 1982). The latter protein is also known as PSP,, (Dub6 et al., 1987a) or P-inhibin (Thakur et al., 1978) but it will, from now on, be referred to as P-microseminopro- tein (P-MSP). Final evidence to consider the prostatic epithelium to be the site of synthesis for these three proteins has also been provided by the immunohisto- chemical demonstration of PAP-, PSA-, and P-MSP-specific immunoreactivity in the secretory epithelial (SE) cells in the normal prostate gland (Jobsis et al., 1981; Papsidero et al., 198 1; Doctor et al., 1986; Dub6 et al., 1987b; Lilja & Abrahamsson, 1988), and the isolation of PAP-, PSA-, and P-MSP-coding DNAs constructed from human prostatic mRNA (Vihko et al., 1988; Sharief et al., 1989; Lundwall and Lilja, 1987; Mbikay et al., 1987). As a result of transudation from the intercellu- lar compartment the prostatic secretion also contains albumin at concentrations corresponding to 1/100 to 1/50 of that in blood plasma. Recently, human Zn-cw,- glycoprotein, a protein that has been pro- posed to be a truncated secretory major histocompatibility complex-related mole- cule (Araki et al., 1988), has been shown to be an abundant prostatic-secreted protein although it occurs in semen at concentra- tions considerable below those of PAP, PSA, and 0-MSP, respectively (Frenette et al., 1987; Burg1 et al., 1989).
Prostatic acid phosphatase. Acid phos- phatases (EC 3.1.3.2) are a group of isoenzymes that hydrolyze phosphate es- ters under acidic conditions. They may be differentiated according to structural, ca- talytic, and immunological properties, as well as according to tissue distribution and subcellular localization. Since 1935, the human prostate has been recognized to secrete high concentrations of acid phosphatases (Kutscher & Wolbergs, 1935), and the PAP is present in the semi- nal plasma at concentrations approxi- mating 1 g/l (Ronnberg et al., 1981). It belongs to the group of acid phospha- tases that are sensitive to inhibition to L- tartrate. The physiological function of PAP is not well understood but recent reports have suggested it to function as a phosphotyrosyl-protein phosphatase (Lin & Clinton, 1986; Li et al., 1984; Chevalier et al., 1988). PAP is a glycoprotein exhib- iting heterogeneity in charge mainly due to structural variations of the polypep- tide chain (Morris et al., 1989). It con- tains two subunits of identical (48 to 52 kDa) size (Luchter-Wasylewska & Os- trowski, 1974). Most recently, the prim- ary structure of PAP has been established (Vihko et al., 1988; Sharief et al., 1989). The mature enzyme is a 354-amino acid protein that has structural resemblance (about 50% identity) to the lysosomal acid phosphatase occurring in all tissues (Sharief et al., 1989; Peters et al., 1989).
Prostate-specific antigen. Prostate-specif- ic antigen (PSA; or y-seminoprotein), is a single chain glycoprotein, first purified from normal human prostatic tissue by Wang et al. in 1979 (Wang et al., 1979). PSA is a serine protease (Lilja, 1985) and it is present in the seminal plasma at concentrations of some 0.6 to 0.7 g/l (Wang et al., 1982; Lilja et al., 1987). The physiologic substrates to PSA in semen are secreted by the seminal vesicles (Lilja, 1985; Lilja et al., 1987; McGee & Herr, 1988). These seminal vesicle-secreted

124 P.-A. Abrahamsson and H. Lilja
proteins participate in the formation of a weak gel-structure at ejaculation and the proteolytic fragmentation of the gel-forming proteins, that accompanies the dissolu- tion of this gel, is due to the action of PSA on these substrates (Lilja, 1985; Lilja et al., 1987; McGee & Herr, 1988).
By the comparison of the NH,-termi- nal sequence of PSA with that of several trypsin-like proteases, PSA was suggest- ed to belong to the family of glandular kallikreins (Lilja, 1985), a subgroup of the trypsin-like serine proteases. This suggestion was confirmed by the estab- lishment of the primary structure of PSA (Lundwall & Lilja, 1987; Watt et al., 1986; Schaller et al., 1987). Most recently, evidences to suggest that another kalli- krein-like protease, hGK-1 (Schedlich et al., 1987), may be produced by the human prostate have come from the identifica- tion in prostatic tissue of a transcript coding for this protease (Chapdelaine et al., 1988) that has a very close structural similarity (about 80% identity) to the primary structure of PSA (Schedlich et al., 1987). The hGK-1 has not yet been identified and characterized by protein chemical methods. However, it must be emphasized that the structural similari- ties of PSA and the hGK-1 may result in extensive immunological cross-reactivi- ty, thus complicating separation of these two most similar proteases.
P-Microseminoprotein. P-MSP is a non- glycosylated 94-amino acid protein mi- grating as a 14- to 16-kDa protein on sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE). It is an abundant protein constituent of the hu- man seminal plasma where it is present at concentrations of about 1 g/l (Dub6 et al., 1987a). The primary structure of P- MSP, which is unrelated to any other known protein, is known from sequence determinations on the purified seminal plasma protein (Seidah et al., 1984; Jo- hansson et al., 1984; Akiyama et al.,
1985) as well as by complementary DNA sequencing (Mbikay et al., 1987). This protein was originally isolated from se- men as a factor inhibiting the pituitary release of follicle-stimulating hormone (Thakur et al., 1978). Later, this has been questioned as highly purified prepara- tions of the protein were devoid of this activity (Kohan et al., 1986; Gordon et al., 1987) and the biological function of P-MSP is therefore at present unknown.
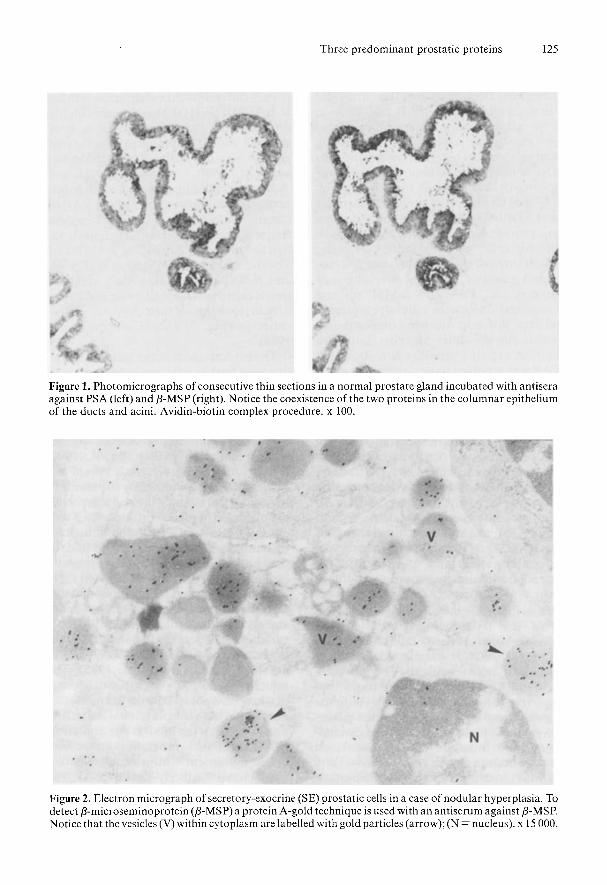
Immunohistochemical distribution of the predominant secretory proteins in the parenchyma of hyperplastic andneoplastic prostate glands. Immunohistochemical localization of PAP, PSA, and P-MSP in nodular hyperplasia. Our finding that each PAP-immunoreactive cell was inva- riably immunoreactive both to PSA- and P-MSP-specific antisera in the epithelium of the acini and of the ducts in the nodules of prostatic hyperplasia prostatic SE cells (Fig. 1) (Abrahamsson et al., 1988) is in agreement with a number of reports (Job- sis et al., 1981; Nadji et al., 1981; Stein et al., 1982; Wang et al., 1982; Tsuda et al., 1983; Okabe & Eto, 1983; Sheth et al., 1985). On the whole, the immunohisto- chemical localization of PAP, PSA, and P-MSP to the SE cells in nodular hyper- plasia is in agreement with the distribu- tion of the three proteins in the normal prostate gland (Fig. 1).
Ultrastructural identification of P-MSP To detect P-MSP on the ultrastructural level we used a protein A-gold technique in a case of prostatic hyperplasia. There- by, P-MSP was found to be distributed throughout the endoplasmic reticulum and cytoplasmic vesicles of the SE cells (Fig. 2), and within the lumina of prostat- ic acini. Control grids revealed only weak, non-specific background labelling.
These observations imply that the P-MSP, synthetised by the rough endo- plasmic reticulum, is stored in vesicles and released into the glandular lumina by exocytosis. A recent report (Warhol &
Three predominant prostatic proteins 125
Figure 1. Photomicrographs of consecutive thin sections in a normal prostate gland incubated with antisera against PSA (left) and p-MSP (right). Notice the coexistence of the two proteins in the columnar epithelium of the ducts and acini. Avidin-biotin complex procedure. x 100.
Figure 2. Electron micrograph of secretory-exocrine (SE) prostatic cells in a case of nodular hyperplasia. To detect P-microseminoprotein (p-MSP) a protein A-gold technique is used with an antiserum against p-MSP. Notice that the vesicles (V) within cytoplasm are labelled with gold particles (arrow); (N = nucleus). x 15 000.

126 P.-A. Abrahamsson and H. Lilja
Longtine, 1985) described the ultrastruc- tural localization of PAP and PSA in hyperplastic and neoplastic human pros- tate glands, the pattern of labelling being almost identical both in hyperplastic cells and in neoplastic tumour cells. In agree- ment with our findings for P-MSP (Fig. 2), the PSA-specific immunoreactivity was distributed throughout the endoplas- mic reticulum and cytoplasmic vesicles of the SE cells, whereas the PAP-specific immunoreactivity was confined to lyso- soma1 granules. Thus these results may suggest that PSA and P-MSP on one hand, and PAP on the other, are segregat- ed into different storage compartments within the glandular SE cells; though in another report (Aumiiller & Seitz, 1985) PAP was reported to be confined to cyto- plasmic vesicles. The lysosomal localiza- tion of PAP reported by Warhol & Long- tine (1985) might be explained by the fact that there is a considerable structural re- semblence between the prostatic-secreted acid phosphatase (PAP) and acid phos- phatase of lysosomal origin; the antisera used in their study may crossreact with the lysosomal acid phosphatase. Al- though some controversy exists regard- ing PAP, it might be concluded that PSA and 0-MSP are localised in the same cyto- plasmic vesicles and released into the glandular lumina by exocytosis.
Immunohistochemical localization of PAP, PSA, and P-MSP in neoplastic pvostate glands. Highly differentiated carcinomas (grade I, WHO) manifested an abun- dance of cells specifically immunoreac- tive to PAP, PSA, and 0-MSP respective- ly (Abrahamsson et al., 1988). As in the normal and hyperplastic prostate paren- chyma, the highly differentiated carcino- mas were found to contain an almost equal number of PAP-, PSA-, and P-MSP-immunoreactive cells. Semiquan- titatively assessed, these PAP-, PSA-, and P-MSP-immunoreactive cells were found to be less abundant in the moderately and
poorly differentiated carcinomas (grades I1 and 111, WHO) than in the highly differentiated ones; the tumour cells of grade I1 and I11 carcinomas also showed greater staining variability (Abrahams- son et al., 1988).
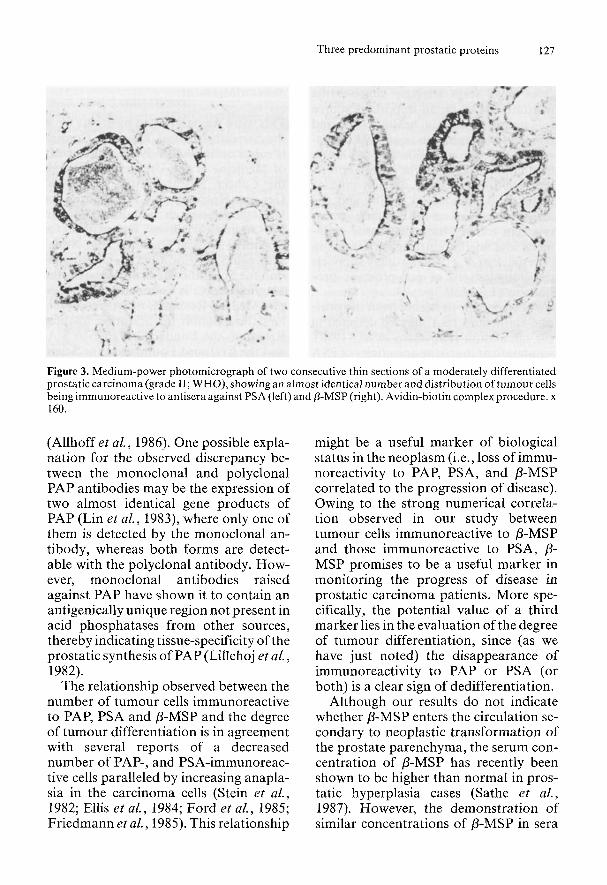
Tumour cells immunoreactive to a mo- noclonal antibody raised against PAP in carcinomas of grade I1 and 111 were less frequent than tumour cells immunoreac- tive to antisera against PSA, P-MSP and a polyclonal antiserum against PAP. PSA and 0-MSP showed an almost identical tissue distribution in all prostatic carci- nomas regardless of their degree of differ- entiation (Fig. 3) (Abrahamsson et al., 1988).
The heterogeneity of the staining inten- sity, seen both in adjacent individual cells and in different areas of the same tumour, is consistent with several reports of con- siderable variation in staining intensity of PAP-, and PSA-immunoreactive cells in prostatic carcinomas of various degrees of differentiation (Allhoff et al., 1983; Ellis et al., 1984; Purnell et al., 1984; Ford et al., 1985). The variability in immuno- histochemical staining intensity, especial- ly noticeable in pourly differentiated car- cinomas, indicates a variation in the rate of synthesis of the three proteins and/or the rate of cell turnover by prostate tumour cells (Papsidero et al., 198 1).
There are several reports indicating PSA to be a more sensitive immunohisto- chemical marker than PAP of prostatic carcinomas, especially in poorly differen- tiated tumors (Allhoff et al., 1983; Ellis et al., 1984; Friedmann et al., 1985). The limitations of PAP as a histochemical marker, here observed with the monoclo- nal but not with the polyclonal antiserum against PAP in the moderately and poor- ly differentiated prostatic carcinomas, are in agreement with a report where a monoclonal antibody against PAP was said to be inferior to a polyclonal anti- body in detecting the antigenic profile altered by malignant transformation
Three predominant prostatic proteins 127
Figure 3. Medium-power photomicrograph of two consecutive thin sections of a moderately differentiated prostatic carcinoma (grade 11; WHO), showing an almost identical number and distribution of tumour cells being immunoreactive to antisera against PSA (left) and 8-MSP (right). Avidin-biotin complex procedure. x 160.
(Allhoff et al., 1986). One possible expla- nation for the observed discrepancy be- tween the monoclonal and polyclonal PAP antibodies may be the expression of two almost identical gene products of PAP (Lin et al., 1983), where only one of them is detected by the monoclonal an- tibody, whereas both forms are detect- able with the polyclonal antibody. How- ever, monoclonal antibodies raised against PAP have shown it to contain an antigenically unique region not present in acid phosphatases from other sources, thereby indicating tissue-specificity of the prostatic synthesis of PAP (Lillehoj et al., 1982).
The relationship observed between the number of tumour cells immunoreactive to PAP, PSA and P-MSP and the degree of tumour differentiation is in agreement with several reports of a decreased number of PAP-, and PSA-immunoreac- tive cells paralleled by increasing anapla- sia in the carcinoma cells (Stein et al., 1982; Ellis et al., 1984; Ford et al., 1985; Friedmann et al., 1985). This relationship
might be a useful marker of biological status in the neoplasm (i.e., loss of immu- noreactivity to PAP, PSA, and P-MSP correlated to the progression of disease). Owing to the strong numerical correla- tion observed in our study between tumour cells immunoreactive to P-MSP and those immunoreactive to PSA, P- MSP promises to be a useful marker in monitoring the progress of disease in prostatic carcinoma patients. More spe- cifically, the potential value of a third marker lies in the evaluation of the degree of tumour differentiation, since (as we have just noted) the disappearance of immunoreactivity to PAP or PSA (or both) is a clear sign of dedifferentiation.
Although our results do not indicate whether P-MSP enters the circulation se- condary to neoplastic transformation of the prostate parenchyma, the serum con- centration of P-MSP has recently been shown to be higher than normal in pros- tatic hyperplasia cases (Sathe et al., 1987). However, the demonstration of similar concentrations of P-MSP in sera

128 P.-A. Abrahamsson and H. Lilja
from both sexes strongly suggests the prostate not to be an important source for serum p-MSP in healthy men and women (Abrahamsson et al., 1989). Mo- reover, the demonstration of p-MSP-spe- cific immunoreactivity in certain epithe- lial cells in the gastric antrum mucosa (Lilja & Abrahamsson, 1988) and the identification of p-MSP-coding trans- cripts in tracheo-bronchial tissue speci- mens and in the antral gastric mucosa (Ulvsback et al., 1989) strongly supports these suggestions of non-prostatic pro- duction of p-MSP.
As secondary to release from the pros- tate epithelium PSA may be detected in the circulation (Papsidero et al., 1980), validation of the tissue-specificity of PSA is obtained by the reportedly undetect- able levels of PSA in the circulation of prostatic cancer patients after successful radical prostatectomy (Oesterling et al., 1988). The results of recent longitudinal prospective studies have shown patients clinical status to correlate more strongly to their serum PSA concentration than to their serum PAP concentration (Siddall et al., 1986; Guinan et al., 1987).
Moreover, in a comparison of the se- rum concentrations of PAP, PSA, and alkaline phosphatase as markers of the risk of progression, a significant relation- ship was found for the serum PSA con- centration only (Killian et al., 1986). In several recent reports PSA is claimed to be more sensitive than PAP in the detec- tion of prostatic carcinoma, and that PSA will probably be the more useful of the two in monitoring response to treat- ment and signalling any recurrence after treatment (Ahmann & Schifman, 1987; Stamey et al., 1987).
Prospective studies are required to compare the usefulness of PAP, PSA, and p-MSP as tumour markers for staging, determining prognosis and monitoring prognosis in cases of prostatic carcino- ma.
Acknowledgements - *
This work was supported by grants from the Swedish Medical Research Council (Project no B-89-13X-7903), the Faculty of Medicine at the University of Lund, the Cancer Research fund at Malmo General Hospital, the Foundation for Urological Research in Malmo, the John och Augusta Perssons Foundation for Scientific Medical Research, the Anna- Lisa och Sven-Eric Lundgrens Founda- tion for Scientific Medical Research, the Magnus Bergvalls Foundation for Scien- tific Medical Research, the Alfred Oster- lund Foundation for Scientific Medical Research, the Fundaci6n Federico S.A. and the Crafoord Foundation for Scien- tific Medical Research.
References
Abrahamsson, P.-A,, H. Lilja, S. Falkmer & L. B. Wadstrom (1988): Immunohistochemical distri- bution of the three predominant secretory pro- teins in the parenchyma of hyperplastic and neo- plastic prostate glands. The Prostate 12: 39-46.
Abrahamsson, P.-A., C. Anderson, T. Bjork, P. Fernlund, H. Lilja,A. Murne & H. Weiber (1989): Radioimmunoassay of P-microseminoprotein, a prostatic-secreted protein present in sera of both men and women. Clin. Chem. 35: 1497-1503.
Ahmann, F.R. & R. B. Schifman (1987): Prospec- tive comparison betweeen serum monoclonal prostate specific antigen and acid phosphatase measurements in metastatic prostatic cancer. J. Urol. 137: 431-434.
Akiyama, K., Y. Yoshioka, K. Schmid, G. D. Offner, R. F. Troxler, R. Tsuda & M. Hara (1985): The amino acid sequence of human p-microsemi- noprotein. Biochem. Biophys. Acta 829: 288-294.
Allhoff, E.P., K. H. Proppe, C. M. Chapman, C. W. Lin & G. R. Prout, JR (1983): Evaluation of prostate specific acid phosphatase and prostate specific antigen in identification of prostatic cancer. J. Urol. 129: 315-318.
Allhoff, E., R. Fischer & R. Engelking (1986): Die Bedeutung immunhistochemischer Techniken fur Diagnose und Therapie des Prostatakarzinoms.
Araki, T., F. Gejyo, K. Takagaki, H. Haupt, H. G. Schwick, W. Burgi, T. Marti, J. Schaller, E. Rickli, R. Brossmer, P. H. Atkinsson, F. W. Putnam & K. Schmid (1988): Complete amino acid sequence of human plasma Zn-a,-glycoprotein and its homo-
Akt. Urol. 17: 113-118.

Three predominant prostatic proteins 129
logy to histocompatibility antigens. Proc. Natl. Acad. Sci (USA) 85: 679-683.
Aumuller, G. & J. Seitz (1985): Cytochemistry and biochemistry of acid phosphatases VI: Immuno- electron microscopic studies on human prostatic and leukocytic acid phosphatases. The Prostate 7:
Biirgl, W., S. Simonen, S. Baudner & K. Schmid (1989): Unusually high concentrations of Zn-a,- glycoprotein and the lack of a,HS-glycoprotein in human ejaculates. Clin. Chem. 35: 1649-1650.
Chapdelaine, P., G. Paradis, R. R. Tremblay & J. Y. Dub& (1988): High level of expression in the prostate of a human glandular kallikrein mRNA related to prostate-specific antigen. FEBS Lett. 236: 205-20S
Chevalier, S., D. Landry & A. Chapdelaine (1988): Phosphotyrosine phosphatase activity of human and canine acid phosphatases of prostatic origin. The Prostate 12: 209-219.
Choe, B.-K. & N. R. Rose (1982): Prostatic acid phosphatase: A marker for human prostatic ade- nocarcinoma. Meth. Cancer Res. 19: 199-207.
Doctor, V. M., A. R. Sheth, M. M. Simha, N. J. Arbatti, J . P. Aaveri &N. A. Sheth (1986): Studies on the immunocytochemical localization of in- hibin-like material in the human prostatic tissue: Comparison of its distribution in normal, benign, and malignant prostates. Br. J. Cancer 53: 547- 554.
Dub&, J.Y., G. Frenette, R. Paquin, P. Chapde- laine, J . Tremblay, R. R. Tremblay, C. Lazure, N. Seidah & M. ChrCtien (1987a): Isolation from human seminal plasma of an abundant 16 kDa protein originating from the prostate, its identifi- cation with a 94-residue peptide originally de- scribed as b-inhibin. J. Androl8: 182-189.
Dub&, J.Y., G. Pelletier, P. Gagnon & R. R. Tremb- lay (1987b): Immunohistochemical localization of a prostatic secretory protein of 94-amino acids in normal prostatic tissue, in primary prostatic tumors and in their metastases. J. Urol. 138: 883- 887.
Ellis, D.W., S. Leffers, J. S. Davies & A. B. P. Ng (1984): Multiple immunoperoxidase markers in benign hyperplasia and adenocarcinoma of the prostate. Am. J. Clin. Pathol. 81: 279-284.
Ford, T.F., D. N. Butcher, J. R. W. Masters & M. C. Parkinson (1985): Immunocytochemical localiza- tion of prostate-specific antigen: Specificity and application to clinical practice. Br. J. Urol. 57: 50- 55.
Frenette, G., J . Y. Dubt, C. Lazure, G. Paradis, M. Chrktien & R. R. Tremblay (1987): The major 40 kDa glycoprotein in human prostatic fluid is identical to Zn-a,-glycoprotein. The Prostate 11:
Friedman, W., J. Steffens, H. Lobeck, S. Bliimcke & R. Nagel (1985): Immunohistochemical dem-
16 1-169.
257-270.
onstration of tumor-associated antigens in pros- tatic carcinomas of various histological differen- tiations. Eur. Urol. 11: 52-56.
Gordon, W.L., W. K. Liu & K. Akiyama (1987): b- Microseminoprotein (P-MSP) is not an inhibin. Biol. Reprod. 36: 829-835.
Guinan, P., Bhatti, R. & P. Ray (1987): An evalua- tion of prostate specific antigen in prostatic cancer. J. Urol. 137: 686-689.
Jobsis, A.C., G. P. de Vries, A. E. F. H. Meijer & J. S. Ploem (1981): The immunohistochemical de- tection of prostatic acid phosphatase: its possibil- ities and limitations in tumour histochemistry. Histochem. J. 13: 961-973.
Johansson, J., A. Sheth, E. Cederlund & H. Jorn- vall (1984): Analysis of an inhibin preparation reveals apparent identity between a peptide with inhibin-like activity and a sperm-coating antigen. FEBS Lett. 176: 21-26.
Killian, C.S., L. J . Emrich, F. P. Vargas et al. (1986): Relative reliability of five serially measured markers for prognosis of progression in prostate cancer. JNCI 76: 179- 185.
Kohan, S., B. Froysa, E. Cederlund, T. Fairwell, R. Lerner, J . Johansson, S. Khan, M. Ritzen, H. Jornvall, S. Cekan & E. Diszfalusy (1986): Pep- tides of postulated inhibin activity: Lack of in vitro inhibin activity of a 94-residue peptide iso- lated from human seminal plasma, and of a syn- thetic replicate of its C-terminal 28-residue seg- ment. FEBS Lett. 199: 242-248.
Kutscher, W. & H. Wolbergs: Prostataphosphatase (1935): 2. Physiol. Chem. 236: 237-240.
Li, H.-C., J. Chernoff, L. B. Chen, A. Kirschon- baum (1984): A phosphotyrosyl-protein phos- phatase activity associated with acid phosphatase from human prostate gland. Eur. J. Biochem. 138:
Lilja, H. (1985): A kallikrein-like serine protease in prostatic fluid cleaves the predominant seminal vesicle protein. J. Clin. Invest. 76: 1899-1903.
Lilja, H., J . Oldbring, G. Rannevik & C. B. Laurel1 (1987): Seminal vesicle-secreted proteins and their reactions during gelation and liquefaction of human semen. J. Clin. Invest. 80: 281-285.
Lilja, H. & P.-A. Abrahamsson (1988): Three pre- dominant proteins secreted by the human pros- tate gland. The Prostate 12: 29-38.
Lillehoj, H.S., B. K. Choe & N. R. Rose (1982): Monoclonal antibodies to human prostatic acid phosphatase: probes for antigenic study. Proc. Natl. Acad. Sci. (USA) 79: 5061-5065.
Lin, M.F., C. Lee, S. S. L. Li & T. M. Chu (1983): Purification and characterization of a new human prostatic acid phosphatase isoenzyme. Biochem- istry 22: 1055-1062.
Lin, M.-F. & G. M. Clinton (1986): Human prostat- ic acid phosphatase has phosphotyrosyl protein phosphatase activity. Biochem. J. 235: 351-357.
45-5 1.

130 €‘.-A. Abrahamsson and H. Lilja
Luchter-Wasylewska, E. & W. Ostrowski (1974): Subunit structure of human prostatic acid phos- phatase. Biochim. Biophys. Acta 265: 349-359.
Lundwall, A& H. Lilja (1987): Molecular cloning of human prostate-specific antigen cDNA. FEBS Lett. 214: 317-322.
Mbikay, M., S. Nolet, S. Fournier, S. Benjannet, P. Chapdelaine, G. Paradis, J. Y. Dubt, R. Tremb- lay, C. Lazure, N. G. Seidah & M. Chrttien (1987): Molecular cloning and sequence of the cDNA for a 94-amino-acid seminal plasma pro- tein secreted by the human prostate. DNA 6: 23- 29.
McGee, R.S., J. C. Herr (1988): Human seminal vesicle-specific antigen is a substrate for prostate- specific antigen (or P-30). Biol. Reprod. 39: 499- 510.
Morris, M.F., A. Waheed, J. M. Risley & R. L. Van Etten (1989): Carbohydrate removal fails to elim- inate the heterogeneity of human prostatic acid phosphatase. Clin. Chim. Acta 182: 9-20.
Nadji, M., S. Z. Tabei, A. Castro, T. M. Chu, G. P. Murphy, M. C. Wang & A. R. Morales (1981): Prostatic-specific antigen: An immunohistologic marker for prostatic neoplasms. Cancer 48: 1229- 1232.
Okabe, T. & K. Eto (1983): Clinical studies of prostatic antigens (y-seminoprotein, p-microse- minoprotein). Examination of immunohistologi- cal study. Jpn. J. Urol. 74: 1313-1319.
Oesterling, J.E., D. W. Chan, J . I. Epstein, A. W. Kimball, Jr., D. J. Bruzek, R. C. Rock, C. B. Brendler & P. C. Walsh (1988): Prostate-specific antigen in the preoperative and postoperative evaluation of localized prostatic cancer treated with radical prostatectomy. J. Urol. 139: 766-772.
Papsidero, L.D., M. C. Wang, L. A. Valenzuela, G. P. Murphy & T. M. Chu (1980): A prostate anti- gen in sera of prostatic cancer patients. Cancer Res. 40: 2428-2432.
Papsidero, L.D., M. Kuriyama, M. C. Wang, J. Horoszewicz, S. S. Leong, L. Valenzuela, G. P. Murphy & T. M. Chu (1981): Prostate antigen: A marker for human prostate epithelial cells. J. Nut. Cancer Inst. 66: 37-41.
Peters, C., C. Geier, R. Pohlmann, A. Waheed, K. von Figura, K. Roiko, P. Virkkunen, P. Henttu & P. Vihko (1989): High degree of homology be- tween primary structure of human lysosomal acid phosphatase and human prostatic acid phospha- tase. Biol. Chem. Hoppe-Seyler 370: 177-181.
Purnell, D.M., B. M. Heatfield & B. F. Trump (1984): Immunocytochemical evaluation of hu- man prostatic carcinomas for carcinoembryonic antigen, nonspecific cross-reacting antigen, P- chorionic gonadotrophin, and prostatc-specific antigen. Cancer Res. 44: 285-292.
Ronnberg, L., P. Vihko, E. Sajanti & R. Vihko (1981): Clomiphene citrate administration to nor-
mogonadotropic subfertile men: blood hormone changes and activation of acid phosphatase in seminal fluid. Int. J. Androl. 4: 372-378.
Sathe,V.S.,N. A. Sheth,M. A. Phadke, A. R. Sheth & Y. P. Zaveri (1987): Biosynthesis and localiza- tion of inhibin in human prostate. The Prostate 10: 33-43.
Schaller, J., K. Akiyama, R. Tsuda, M. Hara, T. Marti & E. Rickli (1987): Isolation, characteriza- tion and amino-acid sequence of y-seminopro- tein, a glycoprotein from human seminal plasma. EUK J. Biochem. 170: 111-120.
Schedlich, L.J., B. H. Bennets & B. J . Morris (1987): Primary structure of a human glandular kallikrein gene. DNA 6: 429-437.
Seidah, N.G., N. J. Arbatti, J. Rochemont, A. R. Sheth & M. Chrttien (1984): Complete amino acid sequence of human seminal plasma P-inhib- in. FEBS Lett. 175: 349-355.
Sharief, ES., H. Lee, M. M. Leuderman, A Lund- wall, L. L. Deaven, C. Lee & S. S. L. Li (1989): Human prostatic acid phosphatase: cDNA clon- ing, gene mapping and protein sequence homol- ogy with lysosomal acid phosphatase. Biochim. Biophys. Res. Cornm. 160: 79-86.
Sheth, N.A., V. M. Doctor & A. R. Sheth (1985): Cellular immunolocalization of inhibinlike pep- tide in human benign prostatic hyperplasia. Arch. Androl. 14: 155-159.
Siddall, J.K., E. H. Cooper, D. W. W. Newling, M. R. G. Robinson & P. Whelan (1986): A evaluation of the immunochemical measurement of prostat- ic acid phosphatase and prostatic specific antigen in carcinoma of the prostate. Eur. Urol. 12: 123- 130.
Stamey, T.A., N. Yang, A. R. Hay, J . E. McNeal, F. S. Freiha & E. Redwine (1987): Prostate-specific antigen as a serum marker for adenocarcinoma of the prostate. N. Engl. J. Med. 317: 909-916.
Stein, B.S., R. 0. Petersen, S. Vangore & A. R. Kendall (1982): Immunoperoxidase localization of prostate-specific antigen. Am. J. Surg. Pathol.
Thakur, A.N., A. Y. Vaze, B. Dattatreyamurthy, N. H. Arbatti & A. R. Sheth (1978): Isolation and characterization of inhibin from human seminal plasma. Indian J. Exp. Biol. 16: 854-856.
Tsuda, R., T. Inoue & M. Hara (1982): A seminal plasma specific antigen of prostate gland. Jpn. J. Leg. Med. 36: 703-709.
Tsuda, R., M. Hara, T. Inoue & T. Okabe (1983): Immunochemical localization of y-seminopro- tein and P-microseminoprotein in prostatic glands. - Forensic immunological studies of body fluids and secretion. Jpn. J. Leg. Med. 31: 16-19.
Ulvsback, M., C. Lindstrorn. ) I . Weiber, P. A. Abrahamsson, H. Lilja & A. Lundwall (1989): Molecular cloning of a small prostate protein, known as P-microseminoprotein, PSP,, or p-in-
6: 553-557.

Three predominant prostatic proteins 131
hibin, and demonstration of transcripts in non- genital tissues. Biochem. Biophys. Res. Commun. (in press).
Vihko, P., P. Virkkunen, P. Henttu, K. Roiko, T. Solin & M. L. Huhtala (1988): Molecular cloning and sequence analysis of cDNA encoding human prostatic acid phosphatase. FEBS Lett. 236: 275- 281.
Wang, M.C., L. A. Valenzuela, G. P. Murphy & T. M. Chu (1979): Purification of a human prostate specific antigen. Invest. Urol. 17: 159-163.
Wang, M.C., M. Kuriyama, L. D. Papsidero, R. M. Loor, L. A. Valenzuela, G. P. Murphy & T. M. Chu (1982). Prostate antigen of human cancer patients. Meth. in Cancer Res. 19: 179-197.
Warhol, M.J. & J. A. Longtine (1985): The ultra- structural localization of prostatic specific anti- gen and prostatic acid phosphatase in hyperplas- tic and neoplastic human prostates. J. Urol. 134:
Watt, K.W.K., P.-J. L. Lee, T. M. M’Timkulu, W.- P. Chan & R. Loor (1986): Human prostate- specific antigen: Structural and functional sim- ilarity with serine proteases. Proc. Natl. Acad. Sci
607-616.
(USA) 83: 3166-3170.
Address: Professor Dr P.-A. Abrahamsson, De- partment of Urology, Malmo General Hospital, S- Forstadsgatan 101, S-21 401 Malmo, Sweden.





























