Embed Size (px)
Citation preview

Thermal evolution of Tethyan surface waters during the Middle-Late
Jurassic: Evidence from D18O values of marine fish teeth
Christophe Lecuyer,1,2 Stephanie Picard,1 Jean-Pierre Garcia,3 Simon M. F. Sheppard,4
Patricia Grandjean,1 and Gilles Dromart1
Received 9 November 2002; revised 27 March 2003; accepted 16 May 2003; published 25 September 2003.
[1] Oxygen isotope compositions of phosphate from vertebrate tooth enamel were measured to determine theevolution of tropical sea surface (<�200 m depth) temperatures in the western Tethys during the Middle-LateJurassic. On the basis of a high-resolution stratigraphic framework with a 1 Myr time resolution, vertebrate teethwere sampled on Aalenian to Portlandian isochrons over the Anglo-Paris Basin. Asteracanthus sharks andPycnodontidae teleosteans, identified as sea surface dwellers, have enamel with d18O values that range from 18.5to 22.3%. Thermal variations of tropical surface waters, with amplitudes of a few degrees per few million years,suggest that Middle to Late Jurassic climates were quite variable. Assuming a seawater d18O value of 0% forsurface tropical waters in the absence of polar ice caps, temperatures increased from 25 to 29�C from the mid-Bajocian to mid-Bathonian. During the middle to late Bathonian, a strong geographic zonation in isotopiccompositions is observed between the eastern and western parts of the basin. High d18O values of fish toothenamel (up to 22.3%) could reflect the arrival of a cold water current from the Arctic during the opening of theNorth Sea rift. An apparent large drop of temperatures from 28 to 21�C is identified at the Callovian-Oxfordianboundary over no more than �2–3 Myr. This cooling is compatible with previous paleobotanical andgeochemical studies and can be precisely correlated with the migration of boreal ammonites into the Tethyandomain. Because isotopic sea surface temperatures are probably too low to be compatible with tropical climaticconditions, the d18O value of seawater could have been >0% owing to limited growth of continental ice duringthe early middle Oxfordian. The resulting sea level fall is estimated to be at least 50 m and is compatible with aglobal regression stage. The middle Oxfordian thermal minimum is followed by a new warming stage of 3–4�Cfrom the middle to the late Oxfordian. INDEX TERMS: 1040 Geochemistry: Isotopic composition/chemistry; 1045
Geochemistry: Low-temperature geochemistry; 1620 Global Change: Climate dynamics (3309); KEYWORDS: oxygen isotope, phosphate,
shark, Jurassic, seawater, climate
Citation: Lecuyer, C., S. Picard, J.-P. Garcia, S. M. F. Sheppard, P. Grandjean, and G. Dromart, Thermal evolution of Tethyan surface
waters during the Middle-Late Jurassic: Evidence from d18O values of marine fish teeth, Paleoceanography, 18(3), 1076,
doi:10.1029/2002PA000863, 2003.
1. Introduction
[2] The Jurassic has long been considered to have been awarm and equable period of Earth history, based on floral andfaunal distribution patterns [e.g., Frakes and Francis, 1990;Frakes et al., 1992; Hallam, 1998]. However, recent paleo-botanical [e.g., Philippe and Thevenard, 1996; Hubbard andBoulter, 1997; Morgans et al., 1999; Riding and Hubbard,1999; Van Aarssen et al., 2000] and oxygen isotope studieson belemnites and brachiopods [e.g.,Ditchfield, 1997; Veizeret al., 1997; Podlaha et al., 1998; Riboulleau et al., 1998]
have provided evidence for fluctuations of both paleotemper-atures and humidity during the Jurassic. Continuous paleo-climatic curves, however, cannot be established from thelimited, and fragmentary database of samples from variablewater depths.[3] Fossil biogenic apatite, especially enamel, shows
more resistance than carbonate shells to oxygen isotopeexchange during diagenesis [Kolodny et al., 1983; Kolodnyand Raab, 1988; Lecuyer et al., 1993]. Biogenic apatites,that crystallize in apparent equilibrium with seawater, haved18O values that are fixed by the temperature and theisotopic composition of water [Longinelli and Nuti, 1973;Kolodny et al., 1983]. Empirical fractionation equationshave been determined with present-day invertebrate andvertebrate ectotherms, leading to a single temperature-de-pendent curve [Longinelli and Nuti, 1973; Kolodny et al.,1983; Lecuyer et al., 1996a]. This intrinsic property of thephosphate oxygen-water system can be applied to extinctspecies to derive past temperatures, assuming a d18O valuefor the environmental water. The oxygen isotope composi-tion of well-preserved biogenic apatites has already beensuccessfully used as a marine paleothermometer [Kolodny
PALEOCEANOGRAPHY, VOL. 18, NO. 3, 1076, doi:10.1029/2002PA000863, 2003
1Laboratoire Paleoenvironnements et Paleobiosphere UMR CNRS5125, Centre Nationale de Recherche Scientifique, Campus de la Doua,Universite Claude Bernard Lyon 1, Villeurbanne, France.
2Also at Institut Universitaire de France, Paris, France.3Centre des Sciences de la Terre, Centre Nationale de Recherche
Scientifique, Universite de Bourgogne, Dijon, France.4Laboratoire de Sciences de la Terre, Centre Nationale de Recherche
Scientifique, Ecole Normale Superieure de Lyon, Lyon, France.
Copyright 2003 by the American Geophysical Union.0883-8305/03/2002PA000863$12.00
21 - 1

and Raab, 1988; Kolodny and Luz, 1991; Lecuyer et al.,1993, 1996b; Picard et al., 1998; Vennemann and Hegner,1998].[4] Before the Latest Jurassic-Early Cretaceous radiation
of carbonate plankton [Roth, 1989], ectotherm vertebratesliving in surface waters, defined here as about the top 100 mof the water column, can be considered as potential sub-stitutes of paleorecorders of sea surface temperatures [Picardet al., 1998]. To characterize the living depth of Jurassic fish,their d18O values were compared with a previously obtaineddata set of coexisting brachiopods [Picard et al., 1998]. Theoxygen isotope compositions of brachiopods record bottom-water temperatures from the shallowest to the deepest depo-sitional environments of the Anglo-Paris Basin. The oxygenisotope compositions of brachiopods appeared to be in goodagreement with independent hydrodynamic and sedimentarycriteria [Guillocheau, 1991].[5] Well documented sedimentological and paleogeo-
graphic studies of the 600 km � 800 km Anglo-Paris Basin[Ziegler, 1988; Guillocheau, 1991; Robin et al., 2000] andthe high-resolution stratigraphic framework, set up byGarcia et al. [1996], Gaumet et al. [1996] and Garciaand Dromart [1997], may help us to understand changes inthe hydrologic budget and oceanic circulation patterns thataffected its water masses. This Basin was located at tropicallatitudes (20�–30�N) during the Middle and Late Jurassic[Dercourt et al., 1985; Ziegler, 1988] with water depthsdown to about 200 m [Guillocheau, 1991; Picard et al.,1998]. This epicontinental sea, representing the occidentalpart of the equatorial Tethyan ocean, was connected to theopening central Atlantic ocean. Since the middle Bathonian,the development of seaways to the north probably enabledefficient communication of surface waters between theTethyan Basin and boreal realms.[6] The Anglo-Paris Basin, with its high-resolution strati-
graphic framework, thus constitutes an especially interest-ing case study to apply oxygen isotope compositions ofMiddle to Late Jurassic fish tooth enamel to (1) reconstructtropical surface seawater temperature variations with time,(2) document the existence of paleocurrents and theirinfluence on faunal migration at the few million yeartimescale, and (3) explore the consequences of tropicaltemperature variations in terms of sea level variations andthe possible presence of a polar ice cap.
2. Depositional Environments
[7] Depositional environments were dominated by threemain carbonate platform domains (Burgundy, Ardennes andNormandy), Normandy being separated from the two othersby a marly furrow and bordered by land masses during theBathonian and the middle and late Oxfordian (Figure 1).The Callovian is characterized by the drowning of relativelyhigh areas by clayey and sandy wedges with a maximumflooding surface in the middle Callovian [Gaumet et al.,1996]. A regressive-transgressive second-order sequencewas recorded in the late Bajocian to middle Calloviancarbonate series [Guillocheau, 1991; Garcia et al., 1996].This sequence includes (1) a regressive hemicycle from theupper Bajocian to the lower-middle Bathonian boundarycorresponding to an overall infill of the basin by prograda-
tion of carbonate platforms until subaerial exposure, and(2) a transgressive hemicycle from the middle Bathonianuntil the transgressive peak of middle Callovian age (Jasonzone). Siliciclastic sands and silts were deposited at thesame time in southern England and over the Channel(between England and France). The next regressive-trans-gressive cycle corresponds to the progradation of siliciclas-tic (middle Callovian to middle Oxfordian) then carbonatewedges from northwest to southeast.[8] The water depth of the fair weather and storm wave
limits may fluctuate depending on both coastal morphologyand occurrence of submarine topographic breaks. Nonethe-less, these hydrodynamic parameters can be used to definefour main depositional environments in the Anglo-Parisbasin. (1) The protected environment where wave actionis limited by topographic barriers. These environments cansometimes correspond to relatively deep locations comparedto emersive lagoonal environments. Shallow environmentsshow evidence of limited subaerial exposure such as mudcracks or birds’ eyes, but without evaporites (excludingdolomite) or significant siliciclastics. (2) The shorefaceabove the fair weather wave base forms barriers that wereoolitic and skeletal in the Bathonian and Callovian, andwere often made up of reefal mounds in the Oxfordian.(3) The upper offshore located between the fair weather andthe storm wave bases, and (4) the lower offshore occursbelow the storm wave base.
3. Sample Collection
[9] Eighty nine teeth (including seventy eight enamelsand eleven whole teeth) and two bones of Chondrichtyanand Osteichtyan Jurassic fish were selected from shallow todeep marine environments of the Anglo-Paris Basin(Table 1; Figure 1). In addition, eleven bulk teeth and onetooth enamel of reptiles, mainly marine crocodilians, wereselected when found associated with fish teeth. Vertebrateremains were sampled along sedimentary isochron profiles[Garcia et al., 1996; Gaumet et al., 1996] with a resolutionof about 1 Myr from the Bajocian to the Tithonian, with oneAalenian tooth. Isochrons consist of sedimentary bedsdefined by monospecific or plurispecific brachiopod asso-ciations that are laterally continuous throughout the basinand coincide with maximum platform floodings [Garciaand Dromart, 1997]. These brachiopod shell beds wereaccurately identified on the basis of one hundred and fortyexploratory oil wells that were used to construct highfrequency cycle correlations [Garcia and Dromart, 1997].They were also calibrated with ammonite standard zonationand Jurassic timescale [Gradstein et al., 1994].[10] Chondrichtyan fish are sharks represented by four
main genera that are Asteracanthus (crushing fish),Notidanus, Sphenodus and Odontaspis (tearing fish).Asteracanthus feeds on shells and is characterized by platyand massive teeth with an enamel structure [Cuny, 1998].Jurassic Osteichtyan fish are represented by durophageousfish, Pycnodontidae with tabular teeth, except one genus,Sphaerodus, having hemispherical teeth, and Gyrodontidae(Gyrodus). Ecology and, especially, living depths of Juras-sic fish, cannot simply be determined from paleontologicalor paleoenvironmental criteria. Depositional environments,
21 - 2 LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER

Figure 1. Sampling locations of Jurassic fish and reptile remains. The size of open circles isproportional to the number of sites. Biogenic phosphates were selected from the western (Normandy,Boulonnais, south England) and the eastern (Burgundy, Jura, and southeast basin) parts of the Anglo-Paris Basin. The paleogeographic map (modified after Dercourt et al. [1985] and Ziegler [1988]) showsthat Western Europe occupied a tropical position during the Middle-Late Jurassic; a latitude is shown andlabelled 20�–30� to give an estimate of uncertainties. Since the Bathonian, the Anglo-Paris basin, locatedat the occidental extremity of the Tethys, was connected northward to boreal realms. North-southseaways, formed during the initial rifting phases of the North Sea, were generally filled by shallowmarine (early-middle Bathonian), then by deeper marine deposits (late Bathonian). Abbreviations are asfollows: B, Boulonnais; N, Normandy; P, Poitou.
LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER 21 - 3

Table
1.Oxygen
IsotopeCompositionsofAalenianto
PortlandianVertebrate
Phosphates
AlongWithTaxonDetermination,Sam
ple
Locations,Ages,andEnvironmentsa
Vertebrate
Taxa
Code
Sam
ple
Stratigraphic
Age
AmmoniteZones
Isochron
Age,
Myr
Location
Depositional
Environments
d18O,
%P205,
%
Odontapsis
93399
T-e
Aalenian
Concavum
Concavum
177.1
Cu
Hte
Saone-Coulevon
O19.3
43
Asteracanthus
D66b
T-e
E.Bajocian
Sauzei
Sauzei
174.3
St
Var-Sollies-Toucas
UO
18.7
41
Pycnodus
L9b
T-e
M.Bajocian
Humphriesianum
Humphriesianum
173.2
Se
Saoneet
Loire-Sennecey
UOPZ
20.5
44
Oxyrhina
L12
T-b
M.to
L.Bajocian
Humphriesianum
Humphriesianum
172
Li
Saoneet
Loire-StSorlin
UOPZ
19.5
37
Pycnodus
93401b
T-e
M.Bajocian
Humphriesianum
Humphriesianum
172
Li
Saoneet
Loire-Laives
UOPZ
19.9
44
Fish
D28
BL.Bajocian
Acris
Acris
170.2
Po
Cote
d’O
r-Pouillenay
UOpPZ
14
28
Asteracanthus
L10b
T-e
L.Bajocian
Acris
Acris
170.2
HSaoneet
Loire-Hurigny
UOBPZ
20.1
34
Marinereptile
L11
T-b
L.Bajocian
Acris
Acris
170.2
HSaoneet
Loire-Hurigny
UOBPZ
17.5
30
Hybodus
O1
T-b
L.Bajocian
Parkinsoni
Parkinsoni
169.3
OCalvados-Omahabeach
UO
21.2
29
Pycnodont
D111-p
bT-e
E.Bathonian
Convergens
Convergens
169.1
Bn
Var-Bandol
UO
19.7
38
Asteracanthus
93357b
T-e
E.-M.Bathonian
Progracilis
Globata
167.9
SStonesfield(U
K)
P20.5
33
Crocodylian
93369
T-b
E.-M.Bathonian
Progracilis
Globata
167.9
SStonesfield(U
K)
P20.8
28
Asteracanthus
D109-A
bT-e
E.-M.Bathonian
--
-Vy
Cher-Vallenay
S19.4
42
Asteracanthus
AST53b
T-e
E.-M.Bathonian
Progracilis-Subcontractus
Globata-Multiplicata
167.9
Me
Jura-M
erey
s/Montrond
P18.7
35
Asteracanthusc
D6b
T-e
M.Bathonian
Progracilis-Subcontractus
Globata-Multiplicata
167.6
VCote
d’O
r-Vanvey
S19.7
35
Asteracanthus
D103-p
bT-e
M.Bathonian
--
-Gl
Gloucester
(UK)
S19.8
35
Asteracanthus
AST54b
T-e
M.-L.Bathonian
Hodsoni-Subcontractus
Concinna-Multiplicata
166.8
Na
Ain-N
antua
LO
19.6
33
Notidanus
93365Nb
T-e
M.-L.Bathonian
Hodsoni
Concinna
166.4
Tr
Isere-Trept
UOp
19.4
32
Pycnodont
93365Pb
T-e
M.-L.Bathonian
Hodsoni
Concinna
166.4
Tr
Isere-Trept
UOp
19.4
39
Asteracanthus
P6-e
bT-e
M.-L.Bathonian
Hodsoni-Retrocostatum
Concinna-Digona
166.4-165.3
Au
Aisne-Aubenton
SPE
20.4
35
Mesodon
P1b
T-e
M.-L.Bathonian
Hodsoni-Retrocostatum
Concinna-Digona
166.4–165.3
Ra
Calvados-Ranville
UOPZ
21
45
Asteracanthus
P3-e
bT-e
M.-L.Bathonian
Hodsoni-Retrocostatum
Concinna-Digona
166.4–165.3
Ma
Sarthe-Mam
ers
S/UOPZ
19.9
42
Asteracanthus
P13b
T-e
M.-L.Bathonian
Hodsoni-Retrocostatum
Concinna-Digona
166.4–165.3
Mr
Pas-de-Calais-Marquise
S20.6
37
Machimosaurusb.
92210
T-b
L.Bathonian
post-H
odsoni
postConcinna
<166.4
Ga
Indre-StGaultier
P19.4
40
Asteracanthus
93362b
T-e
L.Bathonian
post-H
odsoni
postConcinna
<166.4
Ga
Indre-StGaultier
P20.3
37
Asteracanthus
SERA-e
bT-e
L.Bathonian
Retrocostatum
Digona
165.3
Nu
Cote
d’O
r-NuitsSt.Georges
protected
20.2
36
Asteracanthus
93360b
T-e
L.Bathonian
post-Retrocostatum
post-D
igona
<165.3
WWiltshire(U
K)
P(bay)
20.5
30
Asteracanthus
D71b
T-e
L.Bathonian
justeante
Discus
ante
Eudesia
<164.6
Lq
Pas-de-Calais-Locquinghen
UOPZ
21.7
42
Asteracanthus
P12b
T-e
L.Bathonian
ante
Discus
ante
Eudesia
164.6
Mr
Pas-de-Calais-Marquise
UOPZ
22.3
40
Asteracanthus
Ja1b
T-e
L.Bathonian
Discus
Eudesia
164.6
JaYonne-Jaulges
LO
20.3
40
Asteracanthusc
D21Ab
T-e
L.Bathonian
Discus
Eudesia
164.6
LP
Cote
d’O
r-Les
Perrieres
LO
19.1
34
Pycnodontc
D19b
T-e
L.Bathonian
Discus
Eudesia
164.6
LP
Cote
d’O
r-Les
Perrieres
LO
18.8
35
Pycnodontc
D21Pb
T-e
L.Bathonian
Discus
Eudesia
164.6
LP
Cote
d’O
r-Les
Perrieres
LO
18.7
34
Asteracanthus
D35b
T-e
L.Bathonian
Discus
Eudesia
164.6
Ri
Boulonnais-Rinxent
LO
21.3
37
Asteracanthus
Be-1Ab
T-e
L.Bathonian
Discus-Bullatus
Eudesia-D
ivio
164.6–163.9
Me
Hte
Saone-Merey
S19.1
38
Asteracanthus
Bib
T-e
E.Callovian
Bullatus
Divio
163.9
Bi
Yonne-Brion
S20.2
35
Asteracanthusc
D25b
T-e
E.Callovian
Bullatus
Divio
163.9
DCote
d’O
r-Dijon
UOPZ
19.6
34
Asteracanthus
AST55b
T-e
E.Callovian
Bullatus
Divio
163.9
La
Cote
d’O
r-Ladoix
UOPZ
19.0
36
Fish
D22
T-b
E.Callovian
Koenigi
Kalli
163.3
Ta
Cote
d’O
r-Talant
P17.5
33
Asteracanthus
Be-2Ab
T-e
E.Callovian
Koenigi
Kalli
163.3
Or
Doubs-Ornans
UOPZ
19.6
39
Crocodylian
D8
T-b
E.Callovian
Calloviense
Torqui
162.2
ECote
d’O
r-Etrochey
UOPZ
16.6
34
Pycnodusc
D11ab
T-e
E.Callovian
Calloviense
Torqui
162.2
ECote
d’O
r-Etrochey
UOPZ
20.0
37
Asteracanthusc
D10b
T-e
E.Callovian
Calloviense
Torqui
162.2
ECote
d’O
r-Etrochey
UOPZ
18.8
35
Metriorhynchus
N2
T-b
E.Callovian
Calloviense
Torqui
162.2
Ba
Calvados-Bavent
OBPZ
19.3
39
21 - 4 LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER

Table
1.(continued)
Vertebrate
Taxa
Code
Sam
ple
Stratigraphic
Age
AmmoniteZones
Isochron
Age,
Myr
Location
Depositional
Environments
d18O,
%P205,
%
Asteracanthus
L6-e
bT-e
M.Callovian
Jason
Orbignyana
161.5
At
Hte
Saone-Authoison
UOBPZ
19.1
37
Asteracanthus
M2b
T-e
M.Callovian
Coronatum
Oxoniensis
161.1
Pi
Vienne-Carrieredes
Lourdines
?19.8
43
Notidanus
A1-L
T-b
M.Callovian
Coronatum
Oxoniensis
161.1
-Wallucke(D
)LOBPZ
17.7
18
Asteracanthus
D37b
T-e
M.Callovian
Coronatum
Oxoniensis
161.1
ECote
d’O
r-Etrochey
UOBPZ
19.2
34
Asteracanthus
P2b
T-e
L.Callovian
Athleta
Athleta
160.3
Mi
Vendee-St.Michel
enl’Herm
OBPZ
20.8
34
Asteracanthus
D36b
T-e
L.Callovian
Athleta
Athleta
160.3
Pr
Cote
d’O
r-Pruslys/ource
UOBPZ
18.9
39
Asteracanthus
D27b
T-e
L.Callovian
Athleta
Athleta
160.3
Ta
Cote
d’O
r-Talant
UOBPZ
19.5
30
Gyrodus
P5
T-b
L.Callovian
Athleta
orLam
berti
Athleta
orLam
berti
160.3–153.6
Mb
Sarthe-Marolles
lesBraults
UOBPZ
20.3
41
Sphenodus
93355
T-e
L.Callovian
Athleta
orLam
berti
Athleta
orLam
berti
160.3–153.6
Vo
Ardeche-LaVoulte
UOdBZP
19.5
11Marinereptile
93405
T-b
L.Callovian
Athleta
orLam
berti
Athleta
orLam
berti
160.3–153.6
Vi
Calvados-Vaches
noires
UOBZP
21.7
38
Fish
N3
BL.Callovian
Athleta
orLam
berti
Athleta
orLam
berti
160.3–153.6
Vi
Calvados-Villers-sur-mer
LOBPZ
20.3
27
Marinereptile
93404R
T-b
L.Callovian
Lam
berti
Lam
berti
159.6
At
Hte
Saone-Authoison
LOBPZ
18.7
28
Odontaspisl.
L41-e
bT-e
L.Callovian
Lam
berti
Lam
berti
159.6
At
Hte
Saone-Authoison
LOBPZ
20.7
44
Notidanusm.
L126-b
T-b
L.Callovian
Lam
berti
Lam
berti
159.6
At
Hte
Saone-Authoison
LOBPZ
19.4
42
Asteracanthus
W46b
T-e
L.Call-E.Oxf.
--
159.4
Vd
Vaud-Villeneuve
?20.5
36
Asteracanthus
D39b
T-e
E.Oxfordian
Mariae
Mariae
159.1
-Woodham
(UK)
?19.1
43
Notidanusm.
L5-b
T-b
E.Oxfordian
Cordatum-M
ariae
Cordatum-M
ariae
158.8
Ty
Jura-LaBillode
O18.4
16
Asteracanthus
Be-3Ab
T-e
E.Oxfordian
Cordatum-M
ariae
Cordatum-M
ariae
158.8
Or
Doubs-Ornans
O20.9
42
Asteracanthus
P11
bT-e
E.Oxfordian
Cordatum
Cordatum
158.5
To
Calvados-Trouville
s/mer
UOBPZ
20.6
22
Asteracanthus
L3b
T-e
E.Oxfordian
Cordatum
Cordatum
158.5
Ty
Jura-Pontdudiable
O20.4
34
Lam
nidae
B5L-e
bT-e
E.Oxfordian
Cordatum
Cordatum
158.5
Ue
Jura
suisse-U
ken
(CH)
LOBPZ
21.5
33
Notidanusm.
93402-N
-eb
T-e
E.Oxfordian
Cordatum
Cordatum
158.5
Ty
Doubs-Tarcenay
O20.7
44
Odontaspisl.
93402-O
bT-e
E.Oxfordian
Cordatum
Cordatum
158.5
Ty
Doubs-Tarcenay
O19.4
28
Reptile
D32
T-b
E.Oxfordian
Cordatum
Cordatum
158.5
Ar
Jura-A
rcs/Montenot
O18.7
34
Notidanuss.
W22-b
T-b
E.Oxfordian
Cordatum
Cordatum
158.5
Mf
Jura-M
ontfaucon(CH)
O20.3
20
Pycnodus
P15pb
T-e
M.Oxfordian
--
-Gn
Cote
d’O
r-Gigny
OBPZ
20.9
37
Reptile
P15R-e
T-e
M.Oxfordian
--
-Gn
Cote
d’O
r-Gigny
OBPZ
19.3
40
Marinereptile
L14
T-b
M.Oxfordian
Plicatilis
Plicatilis
157.8
FHte
Marne-Foug
UO(PZ)
20.8
32
Sphenodussp.
R1b
T-e
M.Oxfordian
Plicatilis-Antecedens
Plicatilis-Transversarium
157.8–157.6
Vo
Ardeche-Ravin
duCheynier
UOdBZP
20.8
45
Pycnodont
D100-p
bT-e
M.Oxfordian
Plicatilis-Transversarium
Plicatilis-Transversarium
157.8–157.6
-Yorkshire-(U
K)
S20.5
40
Pycnodontidae
B3pb
T-e
M.Oxfordian
Transversarium
Transversarium
157.6
UJura-StUrsanne(CH)
UOPZ
21.4
37
Sphenodusl.
93398-e
bT-e
M.Oxfordian
Transversarium
Transversarium
157.2
Cr
Ardeche-Crussol
UOdBZP
21
36
Sphaerodussp.
D70b
T-e
M.Oxfordian
Transversarium
Transversarium
156.6
Ta
Cote
d’O
r-Talant
LOBPZ
21.5
45
Lam
niform
B4L-e
bT-e
M.Oxfordian
Transversarium
Transversarium
156.9
Si
Jura-Siblingen
(CH)
UOBPZ
21.1
27
Pycnodont
D106-p
bT-e
M.-L.Oxfordian
Transversarium-Bifurcatus
Transversarium-Bifurcatus
156.9–155.9
Vn
Meuse-Verdun
P21.2
42
Pycnodus
P16b
T-e
L.Oxfordian
Bifurcatus
Bifurcatus
155.9
Ml
Yonne-Maillyle
Chateau
UOPZ/S
20.6
40
Sphenodusl.
L1-d
T-b
L.Oxfordian
Bifurcatus
Bifurcatus
155.9
Ch
Jura-Chatelneuf
OBPZ
18
34
Sphenodusl.
L1-e
bT-e
L.Oxfordian
Bifurcatus
Bifurcatus
155.9
Ch
Jura-Chatelneuf
OBPZ
20.8
37
Sphenodusl.
L8-d
T-b
L.Oxfordian
Bifurcatus
Bifurcatus
155.9
Sa
Jura-Savigna
OBPZ
18.9
33
Sphenodusl.
L8-e
bT-e
L.Oxfordian
Bifurcatus
Bifurcatus
155.9
Sa
Jura-Savigna
OBPZ
20.6
37
Pycnodontc
L13b
T-e
L.Oxfordian
Bifurcatus
Bifurcatus
155.9
PMeurtheet
Moselle-Pagny
P20.6
34
Asteracanthus
93363b
T-e
L.Oxfordian
Bifurcatus
Bifurcatus
155.9
He
Calvados-Hennequeville
UO
18.5
31
LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER 21 - 5

constituting a condensed image of fauna and flora activityoccurring from surface to bottom waters, also cannotprovide information on living depths in the water column.Previous studies, however, indicate that pycnodont fishhave been mostly marine, and only in some cases estuarineto freshwater dwellers [Martill, 1990; Nursall, 1996;Poyato-Ariza et al., 1998]. Pycnodonts must have been animportant component of tropical and subtropical coral reefcommunities, analogous to balistid fish in modern oceans[Nursall, 1996]. Asteracanthus sharks are considered asbeing essentially marine, but their ecology is poorly known[Cappetta, 1987]. Other sharks, with lamniform teeth, wereprobably piscivorous [Cappetta, 1987; Martill, 1990].[11] Enamel is more resistant than dentine or bone to post-
depositional oxygen isotope exchange [e.g., Sharp et al.,2000] because of its high degree of mineralization, thereforethis phosphatic tissue was preferentially selected and sepa-rated mechanically with a microdrill under a binocular(Table 1). In the case of the smallest teeth, a few bulkisotopic analyses were performed because it was not possi-ble to separate enough enamel.[12] Similarly, carbonate ions that are present in the apatite
structure are supposed to be less resistant than phosphateions to oxygen isotope exchange during diagenetic alteration[Iacumin et al., 1996;McArthur and Herczeg, 1990]. To testthe state of preservation of Jurassic teeth, we have measuredthe oxygen isotope compositions of both phosphate andcarbonate in fifteen fish and six reptile teeth.
4. Analytical Methods
[13] Carbonate apatite samples were first treated with a3% sodium hypochlorite solution for 4 hours to removeorganic matter and rinsed three times with distilled water.Then they were treated with a 1M acetic acid-acetate bufferfor two days to remove exogenous carbonates according tothe method presented by Bocherens et al. [1996]. Oxygenand carbon isotope ratios from the carbonate in substitutionin the apatite were determined using the phosphoric acidmethod [McCrea, 1950] with a reaction time of 48 hours at30�C. Reproducibility for carbon and oxygen isotopemeasurements in carbonates were ±0.05% and ±0.1%,respectively.[14] Phosphate radicals were isolated from biogenic apa-
tites as Ag3PO4 crystals using the method of Crowson et al.[1991] modified by Lecuyer et al. [1993]. Silver phosphatewas reacted with graphite at 1100�C to produce pure CO2
[Lecuyer et al., 1998] (modified after O’Neil et al. [1994]).The quality of the reaction, marked by the absence of COproduction, was monitored by the carbon isotope composi-tion of CO2 derived from the graphite (Figure 2a). Carbondioxide was analyzed with a VG Prism2 mass spectrometer.All data are quoted in the d notation relatively to SMOW,via calibration on NBS18 and NBS19 international stand-ards. Several tooth enamels, including all those with ex-treme d values were duplicated. Repeated analyses of thephosphorite NBS120c gave a mean d18O value of 21.68 ±0.19% (Figure 2b). The CO2 gave a mean d13C value of�23.51 ± 0.11% (Figure 2c), similar to that of graphiteanalyzed by the CuO method (d13C = �23.5 ± 0.05%). TheP2O5 contents of samples were calculated from their chem-T
able
1.(continued)
Vertebrate
Taxa
Code
Sam
ple
Stratigraphic
Age
AmmoniteZones
Isochron
Age,
Myr
Location
Depositional
Environments
d18O,
%P205,
%
Pycnodont
B1p-e
bT-e
L.Oxfordian
Bim
ammatum
Bim
ammatum
155.2
Ga
Jura-G
achlingen
(CH)
UOBPZ
20.5
38
Asteracanthusc
93364b
T-e
L.Oxfordian
Planula
Planula
154.4
TYonne-Tonnerre
UOZP
19.7
36
Teleosaurides
92154
T-b
L.Oxfordian
Planula
Planula
154.4
TYonne-Tonnerre
UOZP
16.5
33
Microdon
D108-p
bT-e
L.Oxfordian
Planula
Planula
154.4
TYonne-Tonnerre
UOpPZ
20
47
Asteracanthus
P19b
T-e
L.Oxf.to
E.Kim
.Planula
toCymodoce
Planula
toCymodoce
155.2–152.3
Mc
Saone&
Loire-Flace
Macon
P/UOPZ
19.7
42
Peloneustes
W19
T-b
E.Kim
meridgian
--
154.1–152.6
Ha
Seinemaritim
e-LeHavre
O20.7
34
Sphaerodussp.
W29
T-e
E.to
L.Kim
meridgian
--
-Ne
Jura-LaNeuveville
(CH)
P/UOPZ
22
41
Pycnodont
D69
T-e
Kim
meridgian
--
-Gr
Hte
Saone-Arc-les
Gray
?21.3
44
Pycnodont
D33b
T-e
L.Kim
meridgian
MutabilisorEudoxus
MutabilisorEudoxus
152.3–151.4
Do
Aube-Dolancourt
UOpPZ
20
39
Sphaerodussp.
W26b
T-e
L.Kim
meridgian
--
152.6–150.7
Vd
Vaud-Ste
Croix
(CH)
UOpPZ
21.4
43
Pycnodont
D107-p
bT-e
E.Tithonian
Gigas
Gigas
150.2
Bo
Aube-Bourguignons
UOpPZ
20.8
41
Sphaerodussp.
D45b
T-e
E.Tithonian
Gigas
Gigas
150.2
Mt
Haute
Saone-Mantoche
UOpPZ
20.7
37
Mesodongigas
D46b
T-e
E.Tithonian
Gigas
Gigas
150.2
Mt
Haute
Saone-Mantoche
UOpPZ
20.1
42
Sphaerodussp.
D68b
T-e
E.Tithonian
Gravesiana
Gravesiana
150.2
No
Haute
Saone-Noiron
UOpPZ
20.4
43
Pycnodus
93396b
T-e
Portlandian
--
144.2–145.6
Sl
Soleure
(CH)
?20.0
44
Pycnodusgigas
Be5-p
bT-e
Portlandian
--
144.2–145.6
Cx
LaChauxdefonds(CH)
?21.1
42
Mesodongigas
93471
T-e
Portlandian
--
144.2–145.6
Vl
Doubs-Villers
-le-lac
?20.6
24
aNotethatMicrodon,Mesodon,PycnodusgeneraareallPycnodontidae.Stratigraphicages
forisochronswerereported
ontheam
monitebiostratigraphicscaleandontheabsolutescaleproposedbyGradsteinet
al.[1994].P2O5wt%
ofbiogenicapatites
werecalculatedconsideringameanvalueof80±3%
forthesilverphosphatechem
icalyieldobtained
withNISTNBS120c.Depositionalenvironments:P,protected;S,=
shoreface;O,offshore;U,upper;L,lower;PZ,photiczone;
BPZ,below
photiczone;
p,proxim
al;d,distal.Sam
ples:B,bone;
T,tooth;b,bulk;d,dentine;
e,enam
elloıd.
bValues
usedin
Figure
5.
cDatathat
werepreviouslypublished
inthework
ofPicard
etal.[1998].
21 - 6 LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER

ical phosphate yields using the average yield of 80 ± 3%obtained for the NBS120c phosphorite that contains 33.33%of P2O5 (Table 1).[15] Diagenetic alteration of fossil apatite was detected in
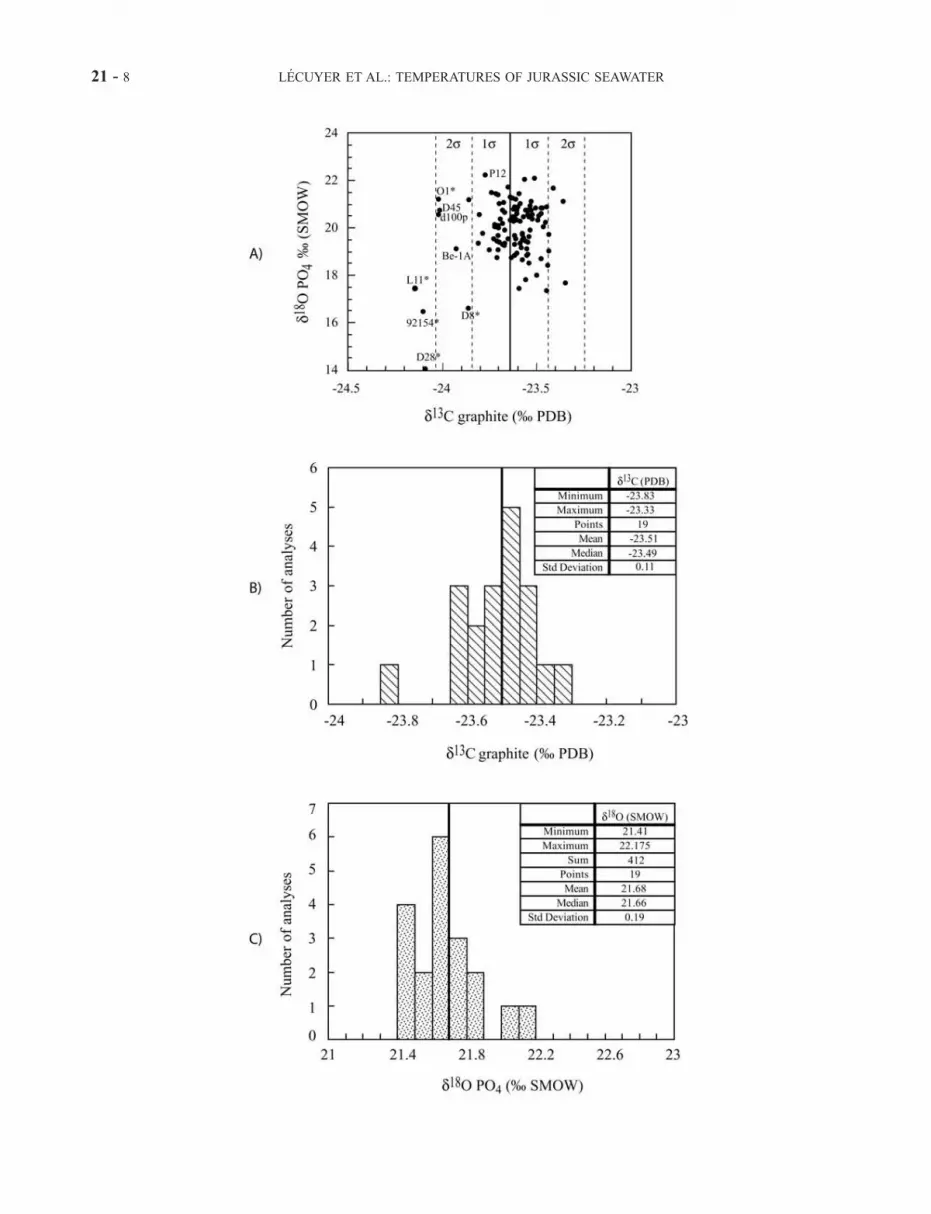
eleven samples during the chemical procedures using thefollowing criteria: (1) recovery of silver phosphate down to60% (Table 1), because of the replacement of phosphates bycarbonates, silica or metal oxides (samples A1-L, 93355,L5b, 93471), (2) solutions that turned from red to yellowduring neutralization with KOH (O1, N3, D22, 93355,D28), revealing the presence of metal oxides (Fe, Mn,Al), and (3) both low d18O values of PO43� and d13Cvalues of graphite (Figure 2a) for samples L11, 92154, D8and D28, indicating the presence of exotic organic mole-cules that were not totally eliminated during the wetchemical procedures [Lecuyer, 2003].
5. Results
[16] Oxygen isotope compositions of phosphate fromJurassic fish and reptiles are reported in Table 1. Analysesof associated dentine and enamel from two teeth of Chon-drichtyan lamnidae (samples L8 and L1) gave chemicalphosphate yields that are about 10% lower for the dentinethan for the enamel. The dentine has d18O values from 1.7 to2.8% lower than the enamel (Table 1), which indicatesstrong isotopic disequilibria between the two coexistingphosphatic tissues. Similar observations were made bySharp et al. [2000] and Puceat et al. [2003]. Massiveenamel of Asteracanthus and pycnodonts have high meanP2O5 percentages with relatively low variance (36.7 ± 3.6%and 39.3 ± 3.4%, respectively), bracketing those obtainedon enamels from tearing teeth (38.2 ± 6.8%; n = 13) thathave a microstructure close to that of modern lamnidae. Bycontrast, bulk fish teeth and bones have variable and lowP2O5 contents (31.6 ± 6.9%) and constitute a fossil materialthat commonly suffered chemical and isotopic alterations.[17] The range and mean d18O values of all well-preserved
vertebrate taxa (where no indicators of diagenesis wereobserved) are reported in Table 2. Similar ranges andmean d18O values were obtained with Asteracanthus andPycnodontidae enamels from the Bathonian-Callovian(19.5 ± 0.5 and 19.6 ± 0.7%) and Oxfordian-Tithonian(20.1 ± 0.6 and 20.7 ± 0.5%) periods. Mean d18O valuesfor other vertebrates like Notidanus (20 ± 0.7%) andOdontaspis (20 ± 0.7%) sharks, though based on a verylimited number of samples, have similar values to thosedocumented for Asteracanthus and Pycnodontidae. Meand18O values for Chondrichtyan Sphenodus (20.8 ± 0.2%)and Osteichtyan Sphaerodus (21.2 ± 0.6%) are higherthan those for Asteracanthus and Pycnodontidae sensustricto, but they are mainly restricted to the Late Jurassic,a period characterized by higher d18O values than thosedocumented throughout the Middle Jurassic. The morepositive d18O values for Sphenodus and Sphaerodus mayindicate that they lived in deeper waters and do not recordsurface temperatures unlike Asteracanthus seems to do(Figure 3). However, we note that Pycnodontidae haved18O values indistinguishable from those of Asteracanthusduring the Bathonian-Callovian and they constitute the
most abundant fossil fauna analyzed during the Oxfor-dian-Tithonian period (Table 2). Therefore the high d18Ovalues of Oxfordian fish teeth can be considered as alsorecording the temperature of the upper part of the watercolumn.[18] Mean d18O values of middle Bathonian to late
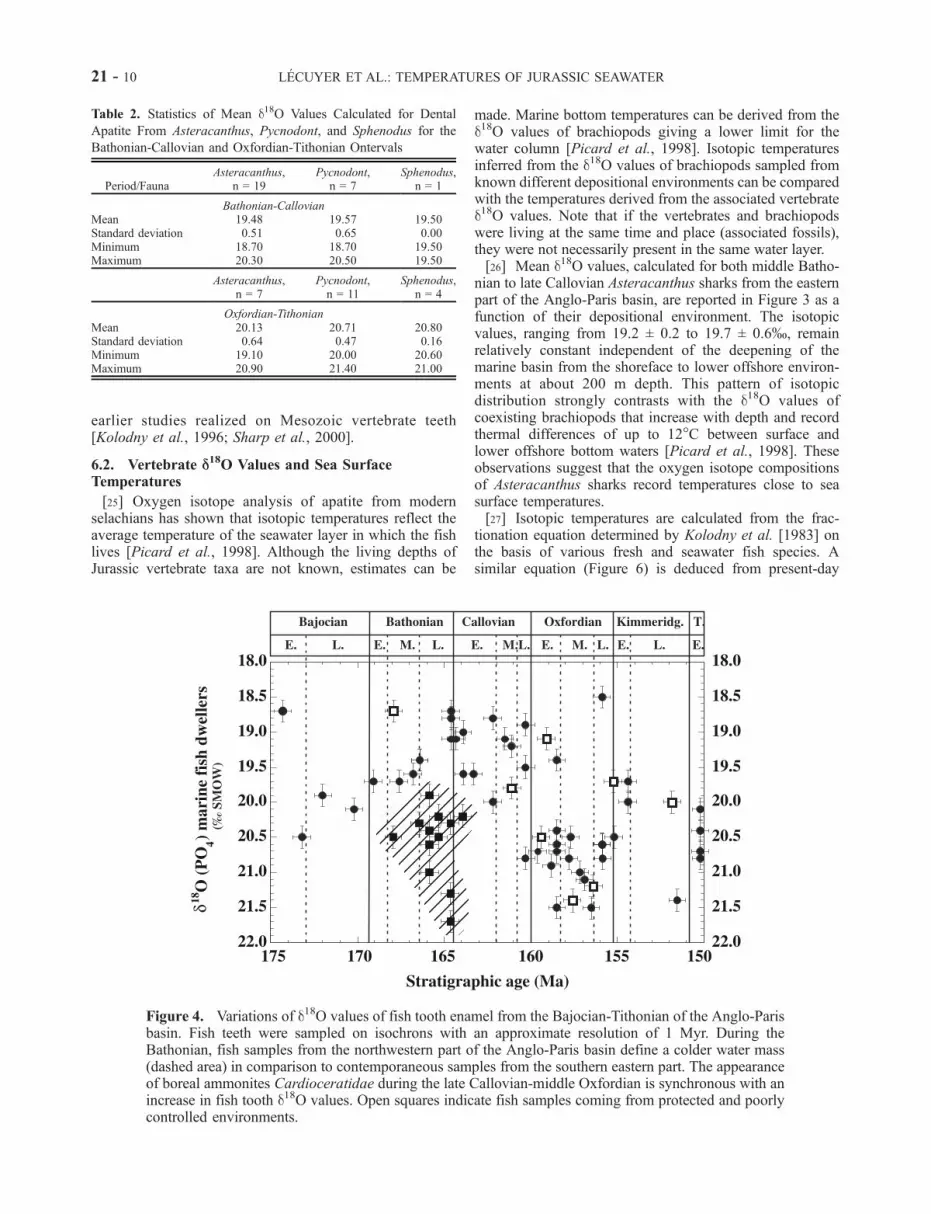
Callovian (168–160 Myr) Asteracanthus have been cal-culated for the various depositional paleoenvironmentsdocumented in the eastern part of the Basin (Figure 3).Note the uniformity of the d18O values of Asteracanthusteeth (from 19.2 ± 0.2 to 19.7 ± 0.6%) that are preservedin inner to outer depositional environments. Middle to lateBathonian samples from the western and northwestern partof the basin (Boulonnais, Normandy, and Poitou regions)have d18O values that are on average 1.5% higher than theirsoutheastern counterparts (Table 1). Samples from thecentral region (Ja1, Bi, SERA, Table 1) have intermediateoxygen isotope ratios, confirming the existence of anisotopic gradient across the whole basin.[19] The Bajocian to the Tithonian fish tooth samples
from the eastern part of the basin show d18O variations over20 Myr (Figure 4). These isotopic variations define anenvelope from 0.5 to 1.5% wide for the various isochrons.This isotopic range is comparable to the one measured byVennemann et al. [2001] on recent fish teeth, which istypically between 0.6 and 1.1% for different teeth from asingle shark. This naturally occurring variation in oxygenisotope compositions recorded in fish teeth may reflect thediversity of the living environments which includes:(1) thermal gradients within the water column; (2) seasonalthermal variations since the growth of a fish tooth representsless than a year (several weeks to several months dependingon species), but they should remain limited under tropicallatitudes; (3) coastline proximity and the influence of fresh-water inputs; and (4) the possible occurrence of surficialoceanic currents.[20] Three major stages may be distinguished: (1) a
period of decreasing d18O values from 20 to 19% for theBajocian to Bathonian interval, (2) a rapid increase of d18Ovalues up to 21% initiated during the late Callovian andreaching a maximum during the early-middle Oxfordian,and (3) another stage of decreasing d18O values, similar inamplitude to stage 1, starting in the late Oxfordian to reachvalues of 20% around the Oxfordian-Kimmeridgianboundary.[21] Oxygen isotope compositions of carbonates from
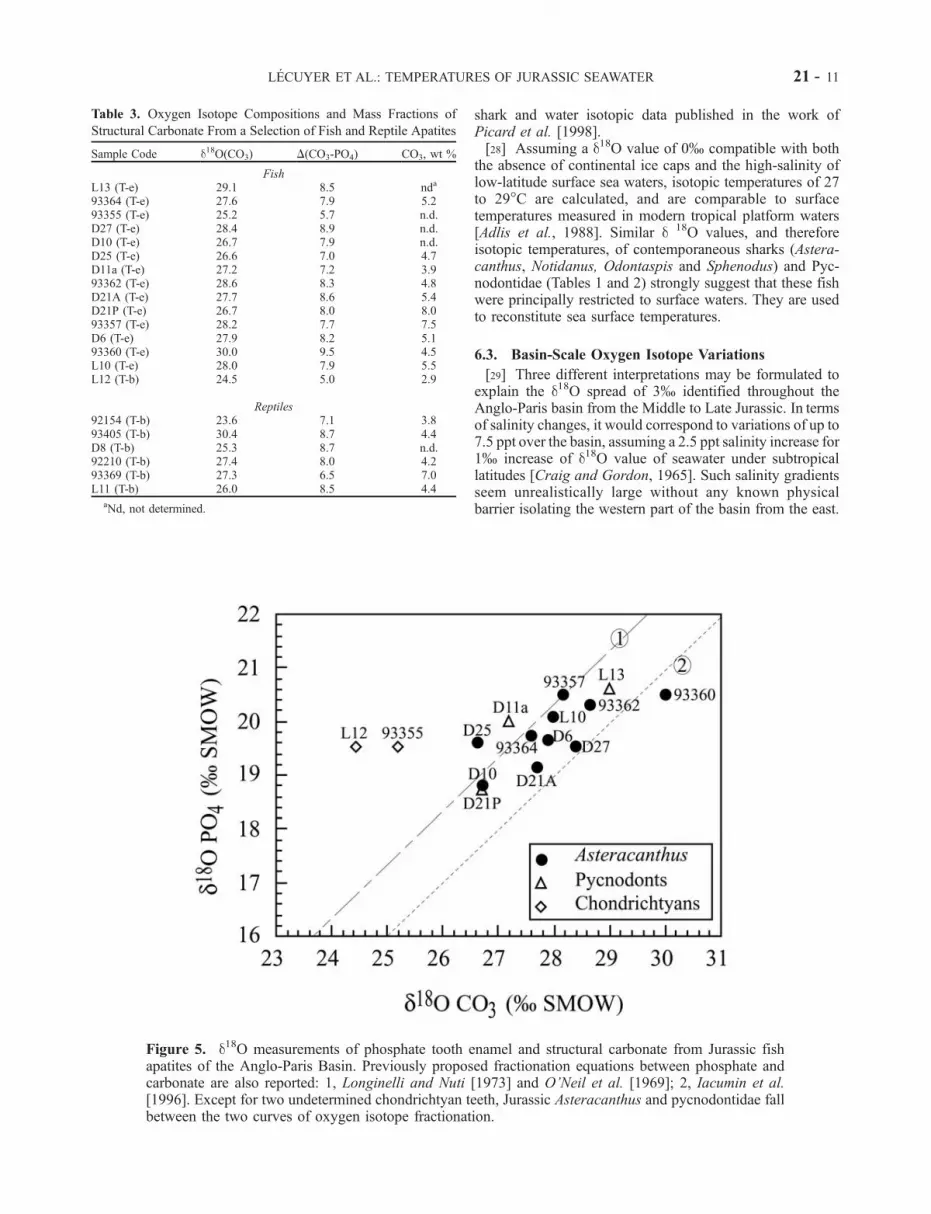
Jurassic fish teeth range from 24.5 to 30.0% whilst thoseof reptiles range from 23.6 to 30.4% (Table 3). For fishteeth, differences between the d18O values measured inapatite carbonate and its associated phosphate oxygen(Dc-p) range from 7.0 to 9.5%, except for two samples; awhole tooth (L12; Dc-p = 5.0%) and one altered sample(93355; Dc-p = 5.7%). These values are within or close tothe known fractionations (Figure 5) between apatite carbon-ate and phosphate oxygen [Longinelli and Nuti, 1973;O’Neil et al., 1969; Iacumin et al., 1996].[22] The amount of carbonate substituted in the apatite
structure of Jurassic fish tooth enamel ranges from 3.9 to5%, except for samples D21P and 93357 (Table 3). Thesevalues match those of 4–5% that were measured in present-
LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER 21 - 7
21 - 8 LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER

day vertebrate teeth [LeGeros and LeGeros, 1984; Michel etal., 1995]. The bulk tooth L12 has a low CO3
2� content of2.9% and a low Dc-p of +5 both diagnostics of a diageneticalteration of the carbonate component of the apatite. Thereptile bulk teeth have CO3
2� contents that range from 3.8to 7%.
6. Discussion
6.1. Diagenetic Perturbations
[23] We conclude from our observations during the chem-ical procedures that seven vertebrate whole teeth (i.e., <8%of the enamel samples) and the two fish bone fragmentshave been chemically and isotopically modified by diage-netic processes. Only one enamel from a Sphenodus tooth(93355) and one enamel from a massive tooth (93471) aresuspect on the basis of the selected criteria. Some of thesephosphate samples are depleted in 18O relative to coexisting
or associated enamel (D28; D22; A1-L; Table 1). Moreover,only two fish teeth (enamel 93355 and whole tooth L12)plot outside the domain of apparent oxygen isotopic equi-librium between phosphate and carbonate, limited by pre-viously proposed fractionation equations (Figure 5). Thissuggests that most enamels from Jurassic teeth have con-served their primary isotopic signatures.[24] On the basis of oxygen isotope analyses of modern
teeth, dentines are depleted in 18O by up to 1.7% relative toassociated enamel [Vennemann et al., 2001]. For compari-son, Jurassic dentines from two lamniform teeth show d18Ovalues up to 2.8% lower than coexisting enamel, suggestingthat these dentines have been significantly altered. Conse-quently, we do not consider bulk samples for paleoenvir-onmental studies even if some of them have d18O values inthe range of contemporaneous fish enamel (W22-b, L126-b,L12; Table 1). The greater resistance of Jurassic enamels todiagenetic processes than either dentine or bone supports
Figure 2. (opposite) Stable isotope compositions of CO2 from Ag3PO4 of Jurassic apatites and phosphorite standardNBS120c. (a) d13C and d18O values of CO2 from Ag3PO4 of Jurassic apatites after reaction with graphite at 1100�C. Notethat most samples having d13C values close to the lower limit of the envelope (defined with 2s a 95% of confidence level)were diagenetically altered (*), except D45, d100p, and Be-1A (see text for more explanations). (b and c) Frequencyhistograms of carbon and oxygen isotope compositions of CO2 from repeated analyses of the phosphorite NBS120c afterreaction of silver phosphate with graphite at 1100�C. The small standard deviations for O and C, of 0.19% and 0.11%respectively, indicate that oxygen from silver phosphate was converted into CO2 without significant production of CO[Lecuyer, 2003].
Figure 3. Mean d18O values calculated for the studied collection of Asteracanthus sharks through theBathonian-Callovian interval. Oxygen isotope compositions are reported along a general depositionalenvironment profile (modified from Gaumet et al. [1996]). Environments were defined by hydrodynamiclimits that are the fair weather base (FWB), the storm wave base (SWB), and by the occurrence ofphotosynthetic organisms (PZB = base of the photic zone). Note the relative constancy withinuncertainties of sharks’ d18O values with deepening of the basin, thus indicating that they record seasurface temperatures.
LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER 21 - 9
earlier studies realized on Mesozoic vertebrate teeth[Kolodny et al., 1996; Sharp et al., 2000].
6.2. Vertebrate D18O Values and Sea Surface
Temperatures
[25] Oxygen isotope analysis of apatite from modernselachians has shown that isotopic temperatures reflect theaverage temperature of the seawater layer in which the fishlives [Picard et al., 1998]. Although the living depths ofJurassic vertebrate taxa are not known, estimates can be
made. Marine bottom temperatures can be derived from thed18O values of brachiopods giving a lower limit for thewater column [Picard et al., 1998]. Isotopic temperaturesinferred from the d18O values of brachiopods sampled fromknown different depositional environments can be comparedwith the temperatures derived from the associated vertebrated18O values. Note that if the vertebrates and brachiopodswere living at the same time and place (associated fossils),they were not necessarily present in the same water layer.[26] Mean d18O values, calculated for both middle Batho-
nian to late Callovian Asteracanthus sharks from the easternpart of the Anglo-Paris basin, are reported in Figure 3 as afunction of their depositional environment. The isotopicvalues, ranging from 19.2 ± 0.2 to 19.7 ± 0.6%, remainrelatively constant independent of the deepening of themarine basin from the shoreface to lower offshore environ-ments at about 200 m depth. This pattern of isotopicdistribution strongly contrasts with the d18O values ofcoexisting brachiopods that increase with depth and recordthermal differences of up to 12�C between surface andlower offshore bottom waters [Picard et al., 1998]. Theseobservations suggest that the oxygen isotope compositionsof Asteracanthus sharks record temperatures close to seasurface temperatures.[27] Isotopic temperatures are calculated from the frac-
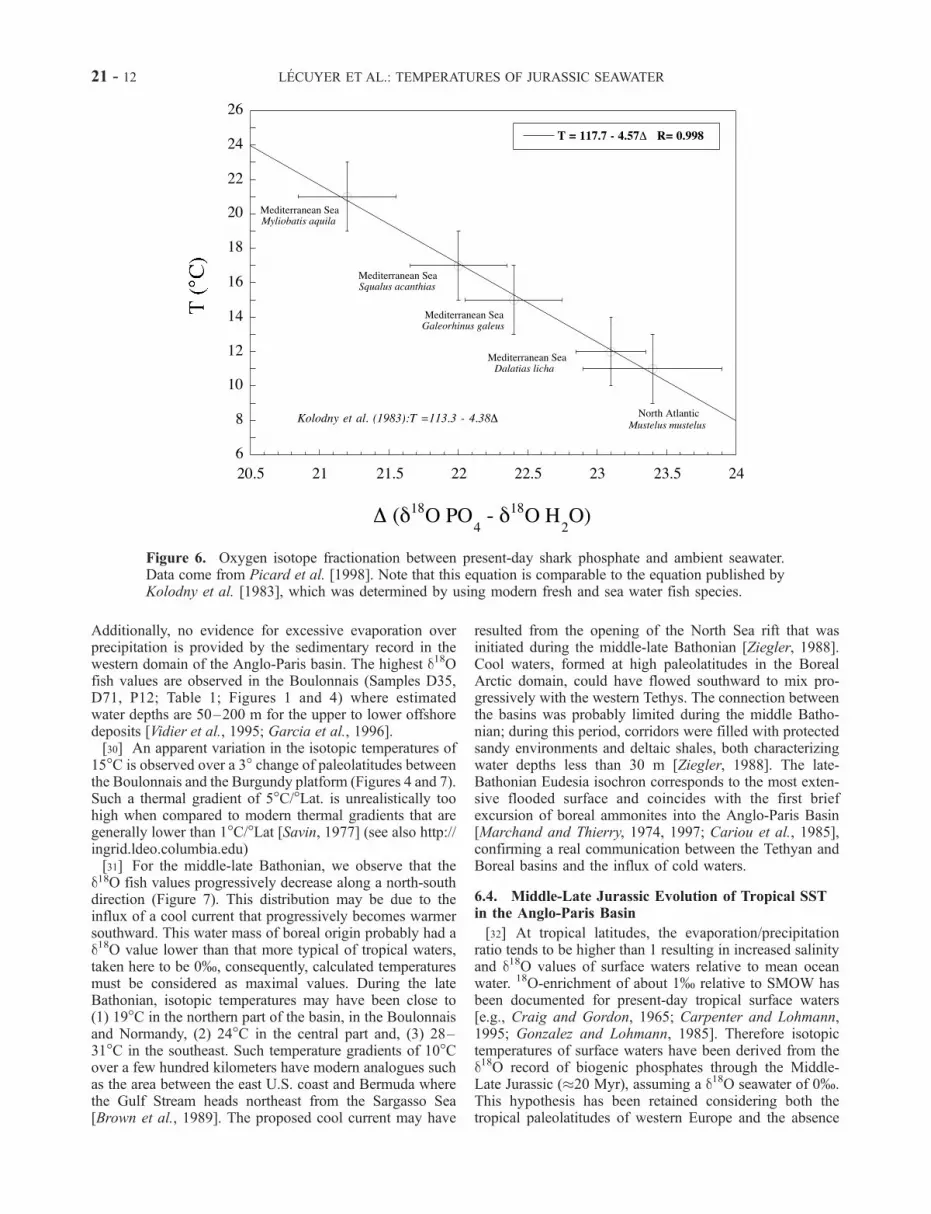
tionation equation determined by Kolodny et al. [1983] onthe basis of various fresh and seawater fish species. Asimilar equation (Figure 6) is deduced from present-day
Table 2. Statistics of Mean d18O Values Calculated for Dental
Apatite From Asteracanthus, Pycnodont, and Sphenodus for the
Bathonian-Callovian and Oxfordian-Tithonian Ontervals
Period/FaunaAsteracanthus,
n = 19Pycnodont,
n = 7Sphenodus,
n = 1
Bathonian-CallovianMean 19.48 19.57 19.50Standard deviation 0.51 0.65 0.00Minimum 18.70 18.70 19.50Maximum 20.30 20.50 19.50
Asteracanthus,n = 7
Pycnodont,n = 11
Sphenodus,n = 4
Oxfordian-TithonianMean 20.13 20.71 20.80Standard deviation 0.64 0.47 0.16Minimum 19.10 20.00 20.60Maximum 20.90 21.40 21.00
Figure 4. Variations of d18O values of fish tooth enamel from the Bajocian-Tithonian of the Anglo-Parisbasin. Fish teeth were sampled on isochrons with an approximate resolution of 1 Myr. During theBathonian, fish samples from the northwestern part of the Anglo-Paris basin define a colder water mass(dashed area) in comparison to contemporaneous samples from the southern eastern part. The appearanceof boreal ammonites Cardioceratidae during the late Callovian-middle Oxfordian is synchronous with anincrease in fish tooth d18O values. Open squares indicate fish samples coming from protected and poorlycontrolled environments.
21 - 10 LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER
shark and water isotopic data published in the work ofPicard et al. [1998].[28] Assuming a d18O value of 0% compatible with both
the absence of continental ice caps and the high-salinity oflow-latitude surface sea waters, isotopic temperatures of 27to 29�C are calculated, and are comparable to surfacetemperatures measured in modern tropical platform waters[Adlis et al., 1988]. Similar d 18O values, and thereforeisotopic temperatures, of contemporaneous sharks (Astera-canthus, Notidanus, Odontaspis and Sphenodus) and Pyc-nodontidae (Tables 1 and 2) strongly suggest that these fishwere principally restricted to surface waters. They are usedto reconstitute sea surface temperatures.
6.3. Basin-Scale Oxygen Isotope Variations
[29] Three different interpretations may be formulated toexplain the d18O spread of 3% identified throughout theAnglo-Paris basin from the Middle to Late Jurassic. In termsof salinity changes, it would correspond to variations of up to7.5 ppt over the basin, assuming a 2.5 ppt salinity increase for1% increase of d18O value of seawater under subtropicallatitudes [Craig and Gordon, 1965]. Such salinity gradientsseem unrealistically large without any known physicalbarrier isolating the western part of the basin from the east.
Figure 5. d18O measurements of phosphate tooth enamel and structural carbonate from Jurassic fishapatites of the Anglo-Paris Basin. Previously proposed fractionation equations between phosphate andcarbonate are also reported: 1, Longinelli and Nuti [1973] and O’Neil et al. [1969]; 2, Iacumin et al.[1996]. Except for two undetermined chondrichtyan teeth, Jurassic Asteracanthus and pycnodontidae fallbetween the two curves of oxygen isotope fractionation.
Table 3. Oxygen Isotope Compositions and Mass Fractions of
Structural Carbonate From a Selection of Fish and Reptile Apatites
Sample Code d18O(CO3) D(CO3-PO4) CO3, wt %
FishL13 (T-e) 29.1 8.5 nda
93364 (T-e) 27.6 7.9 5.293355 (T-e) 25.2 5.7 n.d.D27 (T-e) 28.4 8.9 n.d.D10 (T-e) 26.7 7.9 n.d.D25 (T-e) 26.6 7.0 4.7D11a (T-e) 27.2 7.2 3.993362 (T-e) 28.6 8.3 4.8D21A (T-e) 27.7 8.6 5.4D21P (T-e) 26.7 8.0 8.093357 (T-e) 28.2 7.7 7.5D6 (T-e) 27.9 8.2 5.193360 (T-e) 30.0 9.5 4.5L10 (T-e) 28.0 7.9 5.5L12 (T-b) 24.5 5.0 2.9
Reptiles92154 (T-b) 23.6 7.1 3.893405 (T-b) 30.4 8.7 4.4D8 (T-b) 25.3 8.7 n.d.92210 (T-b) 27.4 8.0 4.293369 (T-b) 27.3 6.5 7.0L11 (T-b) 26.0 8.5 4.4
aNd, not determined.
LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER 21 - 11
Additionally, no evidence for excessive evaporation overprecipitation is provided by the sedimentary record in thewestern domain of the Anglo-Paris basin. The highest d18Ofish values are observed in the Boulonnais (Samples D35,D71, P12; Table 1; Figures 1 and 4) where estimatedwater depths are 50–200 m for the upper to lower offshoredeposits [Vidier et al., 1995; Garcia et al., 1996].[30] An apparent variation in the isotopic temperatures of
15�C is observed over a 3� change of paleolatitudes betweenthe Boulonnais and the Burgundy platform (Figures 4 and 7).Such a thermal gradient of 5�C/�Lat. is unrealistically toohigh when compared to modern thermal gradients that aregenerally lower than 1�C/�Lat [Savin, 1977] (see also http://ingrid.ldeo.columbia.edu)[31] For the middle-late Bathonian, we observe that the
d18O fish values progressively decrease along a north-southdirection (Figure 7). This distribution may be due to theinflux of a cool current that progressively becomes warmersouthward. This water mass of boreal origin probably had ad18O value lower than that more typical of tropical waters,taken here to be 0%, consequently, calculated temperaturesmust be considered as maximal values. During the lateBathonian, isotopic temperatures may have been close to(1) 19�C in the northern part of the basin, in the Boulonnaisand Normandy, (2) 24�C in the central part and, (3) 28–31�C in the southeast. Such temperature gradients of 10�Cover a few hundred kilometers have modern analogues suchas the area between the east U.S. coast and Bermuda wherethe Gulf Stream heads northeast from the Sargasso Sea[Brown et al., 1989]. The proposed cool current may have
resulted from the opening of the North Sea rift that wasinitiated during the middle-late Bathonian [Ziegler, 1988].Cool waters, formed at high paleolatitudes in the BorealArctic domain, could have flowed southward to mix pro-gressively with the western Tethys. The connection betweenthe basins was probably limited during the middle Batho-nian; during this period, corridors were filled with protectedsandy environments and deltaic shales, both characterizingwater depths less than 30 m [Ziegler, 1988]. The late-Bathonian Eudesia isochron corresponds to the most exten-sive flooded surface and coincides with the first briefexcursion of boreal ammonites into the Anglo-Paris Basin[Marchand and Thierry, 1974, 1997; Cariou et al., 1985],confirming a real communication between the Tethyan andBoreal basins and the influx of cold waters.
6.4. Middle-Late Jurassic Evolution of Tropical SSTin the Anglo-Paris Basin
[32] At tropical latitudes, the evaporation/precipitationratio tends to be higher than 1 resulting in increased salinityand d18O values of surface waters relative to mean oceanwater. 18O-enrichment of about 1% relative to SMOW hasbeen documented for present-day tropical surface waters[e.g., Craig and Gordon, 1965; Carpenter and Lohmann,1995; Gonzalez and Lohmann, 1985]. Therefore isotopictemperatures of surface waters have been derived from thed18O record of biogenic phosphates through the Middle-Late Jurassic (�20 Myr), assuming a d18O seawater of 0%.This hypothesis has been retained considering both thetropical paleolatitudes of western Europe and the absence
Figure 6. Oxygen isotope fractionation between present-day shark phosphate and ambient seawater.Data come from Picard et al. [1998]. Note that this equation is comparable to the equation published byKolodny et al. [1983], which was determined by using modern fresh and sea water fish species.
21 - 12 LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER

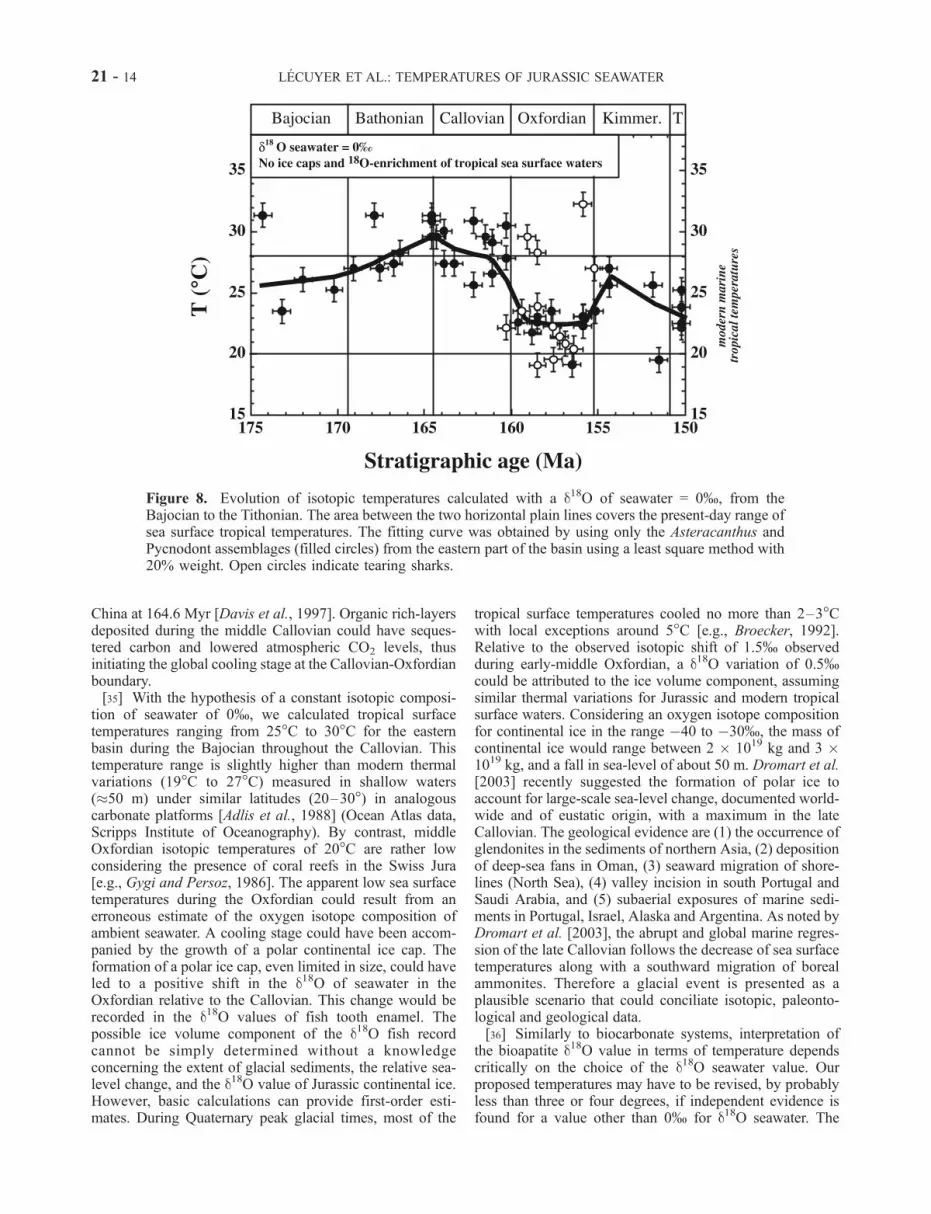
of polar ice caps as a first approximation. Sea surfacetemperatures (SST) inferred from surface dwellers (Aster-acanthus and Pycnodontidae) increased from 25�C to 30�Con average over �7 Myr from late Bajocian to late Batho-nian then decreased down to 20�C in the middle Oxfordianboundary in �8 Myr (Figure 8). This thermal minimum isfollowed by a warming of 5�C of surface waters during thelate Oxfordian and early Kimmeridgian.[33] The apparent cooling of surface waters that was
initiated during the late Callovian Lamberti zone and thatattained a maximum during the early-middle Oxfordianrelates with the colonization of the western Tethys by theboreal ammonites of the Cardioceratidae family [Marchandand Thierry, 1974, 1997; Cariou et al., 1985]. Such acooling episode may have corresponded to the southwardpropagation of a boreal oceanic current and/or to a global
climate change. There is no available evidence for theexistence of a cold oceanic current throughout the majorpart of the Oxfordian. Studies of the distribution of thewood genus Xenoxylon and palynomorphs argue for atemperate and wet climate during the early Oxfordian inwestern Europe [Philippe and Thevenard, 1996; Abbink etal., 2001]. All these data support the existence of a coolingepisode that affected both marine and terrestrial environ-ments of the western Tethyan domain.[34] Perturbations of the carbon cycle due to volcanism or
changes in the rates of burying organic matter could haveinduced temperature oscillations at the timescale of a fewmillion years. According to Dromart et al. [1996], the warmBathonian-Callovian phase could be related to large CO2
fluxes released by contemporaneous arc-related volcanismin Patagonia at 164.1 Myr [Feraud et al., 1999] and South
Figure 7. Geographical map of the Anglo-Paris Basin with the regional distribution of fish tooth d18Ovalues for the middle-late Bathonian. The gray area encloses isotopic compositions higher than 20%.This regional distribution of d18O values could have resulted from the presence of a cold oceanic currentoriginating from boreal realms.
LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER 21 - 13
China at 164.6 Myr [Davis et al., 1997]. Organic rich-layersdeposited during the middle Callovian could have seques-tered carbon and lowered atmospheric CO2 levels, thusinitiating the global cooling stage at the Callovian-Oxfordianboundary.[35] With the hypothesis of a constant isotopic composi-
tion of seawater of 0%, we calculated tropical surfacetemperatures ranging from 25�C to 30�C for the easternbasin during the Bajocian throughout the Callovian. Thistemperature range is slightly higher than modern thermalvariations (19�C to 27�C) measured in shallow waters(�50 m) under similar latitudes (20–30�) in analogouscarbonate platforms [Adlis et al., 1988] (Ocean Atlas data,Scripps Institute of Oceanography). By contrast, middleOxfordian isotopic temperatures of 20�C are rather lowconsidering the presence of coral reefs in the Swiss Jura[e.g., Gygi and Persoz, 1986]. The apparent low sea surfacetemperatures during the Oxfordian could result from anerroneous estimate of the oxygen isotope composition ofambient seawater. A cooling stage could have been accom-panied by the growth of a polar continental ice cap. Theformation of a polar ice cap, even limited in size, could haveled to a positive shift in the d18O of seawater in theOxfordian relative to the Callovian. This change would berecorded in the d18O values of fish tooth enamel. Thepossible ice volume component of the d18O fish recordcannot be simply determined without a knowledgeconcerning the extent of glacial sediments, the relative sea-level change, and the d18O value of Jurassic continental ice.However, basic calculations can provide first-order esti-mates. During Quaternary peak glacial times, most of the
tropical surface temperatures cooled no more than 2–3�Cwith local exceptions around 5�C [e.g., Broecker, 1992].Relative to the observed isotopic shift of 1.5% observedduring early-middle Oxfordian, a d18O variation of 0.5%could be attributed to the ice volume component, assumingsimilar thermal variations for Jurassic and modern tropicalsurface waters. Considering an oxygen isotope compositionfor continental ice in the range �40 to �30%, the mass ofcontinental ice would range between 2 � 1019 kg and 3 �1019 kg, and a fall in sea-level of about 50 m. Dromart et al.[2003] recently suggested the formation of polar ice toaccount for large-scale sea-level change, documented world-wide and of eustatic origin, with a maximum in the lateCallovian. The geological evidence are (1) the occurrence ofglendonites in the sediments of northern Asia, (2) depositionof deep-sea fans in Oman, (3) seaward migration of shore-lines (North Sea), (4) valley incision in south Portugal andSaudi Arabia, and (5) subaerial exposures of marine sedi-ments in Portugal, Israel, Alaska and Argentina. As noted byDromart et al. [2003], the abrupt and global marine regres-sion of the late Callovian follows the decrease of sea surfacetemperatures along with a southward migration of borealammonites. Therefore a glacial event is presented as aplausible scenario that could conciliate isotopic, paleonto-logical and geological data.[36] Similarly to biocarbonate systems, interpretation of
the bioapatite d18O value in terms of temperature dependscritically on the choice of the d18O seawater value. Ourproposed temperatures may have to be revised, by probablyless than three or four degrees, if independent evidence isfound for a value other than 0% for d18O seawater. The
Figure 8. Evolution of isotopic temperatures calculated with a d18O of seawater = 0%, from theBajocian to the Tithonian. The area between the two horizontal plain lines covers the present-day range ofsea surface tropical temperatures. The fitting curve was obtained by using only the Asteracanthus andPycnodont assemblages (filled circles) from the eastern part of the basin using a least square method with20% weight. Open circles indicate tearing sharks.
21 - 14 LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER

presence or absence of continental polar ice caps is a crucialelement in this discussion. From a purely isotopic point ofview for the period from the Bajocian through to theThithonian (Figure 8), a tentative case can only be madefor the presence of a significant continental ice reservoirduring the Oxfordian, the Thithonian and perhaps the Bajo-cian-Bathonian, the three stages with relatively high d18Oapatite values. These propositions are supported, in part, bythe documented periods of major regression [Guillocheau,1991; Garcia et al., 1996] and occurrence of possible glacialphenomena in other part of the world [Price, 2000].
7. Conclusion
[37] Oxygen isotope compositions of marine vertebratetooth enamel, that were not diagenetically altered, vary from18.5 to 22.5% for the Middle-Late Jurassic of the westernTethyan basin. These data are interpreted as evidence forthermal variations of sea surface (�100 m) temperaturesover a few Myr scale. A drop of isotopic temperatures(about 3�C to 5�C) occurred around the Callovian-Oxfor-dian boundary. This cooling of surface waters is associatedwith the southward migration of boreal ammonites and thedevelopment of temperate flora in the Anglo-Paris Basin.This cooler episode precedes a thermal increase that beganduring the late Oxfordian until the Kimmeridgian. Apparentisotopic temperatures not exceeding 20�C are low fortropical latitudes even when assuming an isotopic compo-sition of seawater of 0% compatible with 18O-enriched low-
latitude sea surface waters and the absence of permanentwell-developed polar ice caps. Therefore we cannot excludethe existence of an Oxfordian glaciation event that couldexplain the contemporaneous southward migration of borealammonites and the global sea-level fall.[38] Strong oxygen isotope gradients (3%) over 500 km
from the northwest to the southeast of the basin suggest thedevelopment of a cool oceanic current during the middle-late Bathonian that could be related to the opening of theNorth Sea rift. According to paleontological and paleobo-tanical studies, we suggest that the late Callovian-earlyOxfordian thermal decrease could be the expression of aglobal event rather than a more local geographic effect ofthese cool oceanic currents. In the absence of isotopicrecords provided by foraminifera, those available from fishphosphatic remains reveal that Jurassic climates were not soconsistently warm and equable as previously thought.
[39] Acknowledgments. Sampling would not have been possiblewithout the help of A. Prieur (Universite Claude Bernard de Lyon),B. Laurin, J.-H. Delance (Universite de Bourgogne), M. Weidmann(Universite de Lausanne), A. Boullier (Universite de Besancon), A. Lena,G. Gallio (Museum de la Citadelle, Besancon), J. P. Goujon (MNHN,Paris), Museum de Bale, J. M. Mazin (Universite de Poitiers), and G. Breton(Museum du Havre), who provided some of the vertebrate samples. We alsothank M. Emery for her help in sample analyses. The authors thank thereviewers David Dettman and Christina Hartman, who helped us toimprove the scientific content of this manuscript. This work was supportedby the CNRS through the programs ‘‘Interieur de la Terre’’ (Comportementdynamique des plates-formes carbonatees) and ‘‘ECLIPSE.’’
ReferencesAbbink, O., J. Targarona, H. Brinkhuis, andH. Visscher, Late Jurassic to earliest cretaceouspalaeoclimatic evolution of the southern NorthSea, Global Planet. Change, 30, 231–256,2001.
Adlis, D. S., E. L. Grossman, T. E. Ancey, andR. D. McLerran, Isotope stratigraphy andpaleodepth changes of Pennsylvanian cyclicalsedimentary deposits, Palaios, 3, 487–506,1988.
Bocherens, H., P. L. Koch, A. Mariotti,D. Geraads, and J. J. Jaeger, Isotopic biogeo-chemistry (C-13, O-18) of mammalian enamelfrom African Pleistocene hominid sites, Pa-laios, 11, 306–318, 1996.
Broecker, W. S., The Glacial World According toWally, 318 pp., Lahmont-Doherty Earth Obser-vatory, Palisades, N. Y., 1992.
Brown, J., A. Colling, D. Park, J. Phillips,D. Rothery, and J. Wright, Ocean Circulation,238 pp., Open Univ. Press, Milton Keynes,U. K., 1989.
Cappetta, H., Mesozoic and Cenozoic Elasmo-branchii, in Handbook of Paleoichtyology,Chondrichthyes II, edited by H.-P. Schultze,pp. 193, Springer-Verlag, New York, 1987.
Cariou, E., D. Contini, J.-L. Dommergues,R. Enay, J. R. Geyssant, C. Mangold, andJ. Thierry, Biogeographie des Ammonites etevolution structurale de la Tethys au cours duJurassique, Bull. Soc. Geol. Fr., 8, 679–697,1985.
Carpenter, S. J., and K. C. Lohmann, d18Oand d13C values of modern brachiopod shells,Geochim. Cosmochim. Acta, 59, 3749–3764,1995.
Craig, H., and L. I. Gordon, Deuterium and oxy-gen-18 variations in the ocean and the marine
atmosphere, in Stable Isotopes in Oceano-graphic Studies and Paleotemperatures, editedby E. Tongiorgi, pp. 9–130, Spoleto, Cons.Naz. delle Ric., Lab. di Geol. Nucl., Pisa,1965.
Crowson, R. A., W. J. Showers, E. K. Wright,and T. C. Hoering, A method for preparation ofphosphate samples for oxygen isotope analy-sis, Anal. Chem., 63, 2397–2400, 1991.
Cuny, G., Primitive neoselachian sharks: A sur-vey, Oryctos, 1, 3–21, 1998.
Davis, D.W., R. J. Sewell, and S. D. G. Campbell,U-Pb dating of Mesozoic igneous rocks fromHong Kong, J. Geol. Soc. London, 154,1067–1076, 1997.
Dercourt, J., L. P. Zonenshaim, L. P. Ricou, V. G.Kazmin, X. Le Pichon, and A. L. Knipper,Presentation de neuf cartes paleogeographi-ques au 1/20000000 s’etendant de l’Atlantiqueau Pamir pour la periode du Lias a l’actuel,Bull. Soc. Geol. Fr., 8, 637–652, 1985.
Ditchfield, P. W., High northern palaeolatitudeJurassic-Cretaceous palaeotemperature varia-tion: new data fromKongKarls Land, Svalbard,Palaeogeogr.Palaeoclimatol.Palaeoecol.,130,163–175, 1997.
Dromart, G., P. Allemand, J.-P. Garcia, andC. Robin, Variation cyclique de la productioncarbonatee au Jurassique le long d’un transectBourgogne-Ardeche, Est-France, Bull. Soc.Geol. Fr., 167, 423–433, 1996.
Dromart, G., J.-P. Garcia, S. Picard, F. Atrops,C. Lecuyer, and S. M. F. Sheppard, Ice age atthe Middle-Late Jurassic transition?, EarthPlanet. Sci. Lett, in press, 2003.
Feraud, G., V. Alric, M. Fornair, H. Bertrand,and M. Haller, 40Ar/39Ar dating of the Jurassicvolcanic province of Patagonia: Migrating
magmatism related to Gondwana break-upand subduction, Earth Planet. Sci. Lett., 172,83–96, 1999.
Frakes, L. A., and J. E. Francis, Cretaceouspalaeoclimates, in Cretaceous Resources,Events and Rhythms, edited by R. N. Ginsburgand B. Beaudoin, pp. 373–387, Kluwer Acad.,New York, 1990.
Frakes, L. A., J. E. Francis, and J. I. Syktus,Climate Modes of the Phanerozoic, 274 pp.,Cambridge Univ. Press, New York, 1992.
Garcia, J.-P., and G. Dromart, The validity of twobiostratigraphic approaches in sequence strati-graphic correlations: Brachiopod zones andmarker-beds in the Jurassic, Sediment. Geol.,114, 55–79, 1997.
Garcia, J.-P., G. Dromart, F. Guillocheau,P. Allemand, F. Gaumet, C. Robin, andG. Sambet, Bathonian-Callovian Paris BasinSubalpine Basin intercorrelations along anArdennes-Ardeche cross-section, C. R. Acad.Sci., Ser. II, 323, 697–703, 1996.
Gaumet, F., J.-P. Garcia, G. Dromart, andG. Sambet, Controle stratigraphique des fa-cies, geometries et profils de depot de laplate-forme carbonatee bourguignonne auBathonien-Callovien, Bull. Soc. Geol. Fr.,167, 409–421, 1996.
Gonzalez, L. A., and K. C. Lohmann, Carbonand oxygen isotopic composition of Holocenereefal carbonates, Geology, 13, 811 – 814,1985.
Gradstein, F. M., F. P. Agterberg, J. G. Ogg,J. Hardenbol, P. Van Veen, J. Thierry, andZ. Huang, A Mesozoic time scale, J. Geophys.Res., 99, 24,051–24,074, 1994.
Guillocheau, F., Mise en evidence des grandscycles transgression-regression d’origine tecto-
LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER 21 - 15

nique dans les sediments mesozoıques du Bas-sin de Paris, C. R. Acad. Sci., Ser. II, 312,1587–1593, 1991.
Gygi, R. A., and F. Persoz, Mineralostratigra-phiy, litho- and biostratigraphiy combined incorrelation of the Oxfordian (Late Jurassic)formations of the Swiss Jura range, EclogaeGeol. Helv., 79, 385–454, 1986.
Hallam, A., The determination of Jurassic envi-ronments using palaeoecological methods,Bull. Soc. Geol. Fr., 169, 681–687, 1998.
Hubbard, N. L. B., and M. C. Boulter, MidMesozoic floras and climates, Palaeontology,40, 43–70, 1997.
Iacumin, P., H. Bocherens, A. Mariotti, andA. Longinelli, Oxygen isotope analyses of co-existing carbonate and phosphate in biogenicapatite: A way to monitor diagenetic alterationof bone phosphate?, Earth Planet. Sci. Lett.,142, 1–6, 1996.
Kolodny, Y., and B. Luz, Oxygen isotopes inphosphates of fossil fish—Devonian to recent,in Stable Isotope Geochemistry: A Tribute toSamuel Epstein, edited by H. P. Taylor, J. R.O’Neil, and I. R. Kaplan, pp. 105–119, Geo-chem. Soc., San Antonio, Tex., 1991.
Kolodny, Y., and M. Raab, Oxygen isotopes inphosphatic fish remains from Israel: Paleother-mometry of tropical Cretaceous and Tertiaryshelf waters, Palaeogeogr. Palaeoclimatol.Palaeoecol., 64, 59–67, 1988.
Kolodny, Y., B. Luz, and O. Navon, Oxygenisotope variations in phosphate of biogenicapatites, I. Fish bone apatite-rechecking therules of the game, Earth Planet. Sci. Lett.,64, 398–404, 1983.
Kolodny, Y., B. Luz, M. Sander, and W. A.Clemens, Dinosaur bones: Fossils or pseudo-morphs?: The pitfalls of physiology recon-struction from apatitic fossils, Palaeogeogr.Palaeoclimatol. Palaeoecol., 126, 161–171,1996.
Lecuyer, C., Oxygen isotope analysis of phos-phates, in Handbook of Stable Isotope Analy-tical Techniques, Elsevier Sci., New York, inpress, 2003.
Lecuyer, C., P. Grandjean, J. R. O’Neil, H. Cap-petta, and F. Martineau, Thermal excursions inthe ocean at the Cretaceous-Tertiary boundary(northern Morocco): The d18O record of phos-phatic fish debris, Palaeogeogr. Palaeoclima-tol. Palaeoecol., 105, 235–243, 1993.
Lecuyer, C., P. Grandjean, and C. C. Emig,Determination of oxygen isotope fractionationbetween water and phosphate from living lin-gulids: Potential application to palaeoenviron-mental studies, Palaeogeogr. Palaeoclimatol.Palaeoecol., 126, 101–108, 1996a.
Lecuyer, C., P. Grandjean, F. Paris, M. Robardet,and F. Robineau, Deciphering ‘‘temperature’’and ‘‘salinity’’ from biogenic phosphates: Thed18O of coexisting fishes and mammals of themiddle Miocene sea of western France,Palaeogeogr. Palaeoclimatol. Palaeoecol.,126, 61–74, 1996b.
Lecuyer, C., P. Grandjean, J. A. Barrat, C. C.Emig, J. Nolvak, F. Paris, and M. Robardet,d18O and REE contents of phosphatic brachio-pods: A comparison betweenmodern and lowerPaleozoic populations, Geochim. Cosmochim.Acta, 62, 2429–2436, 1998.
LeGeros, R. Z., and J. P. LeGeros, Phosphateminerals in human tissues, in Phosphate Miner-als, edited by J. O. Nriagu and P. B. Moore, pp.351–385, Springer-Verlag, New York, 1984.
Longinelli, A., and S. Nuti, Revised phosphate-water isotopic temperature scale, Earth Planet.Sci. Lett., 19, 373–376, 1973.
Marchand, D., and J. Thierry, Les influencesmesogeennes et boreales dans le Callovien deBourgogne, Bull. Soc. Geol. Fr., 16, 476–484,1974.
Marchand, D., and J. Thierry, Enregistrement desvariations morphologiques et de la composi-tion des peuplements d’ammonites durant lecycle regressif/transgressif de 2eme ordreBathonien inferieur-Oxfordien inferieur enEurope occidentale, Bull. Soc. Geol. Fr., 168,121–132, 1997.
Martill, D. M., Predation on Kosmoceras bySemionotid fish in the middle Jurassic lowerOxford clay of England, Paleontology, 33,739–742, 1990.
McArthur, J. M., and A. Herczeg, Diagenetic sta-bility of the isotopic composition of phosphate-oxygen: palaeoenvironmental implications, inPhosphorite Research and Development, editedby A. J. G. Notholt and I. Jarvis, pp. 119–124,Geol. Soc. Am., Boulder, Colo., 1990.
McCrea, J. M., On the isotopic chemistry of car-bonates and a paleotemperature scale, J. Chem.Phys., 18, 849–857, 1950.
Michel, V., P. Ildefonse, and G. Morin, Chemicaland structural changes in Cervus elaphus toothenamels during fossilization (Lazaret cave): Acombined IR and XRD Rietveld analysis,Appl. Geochem., 10, 145–159, 1995.
Morgans, H., S. P. Hesselbo, and R. A. Spicer,The seasonal climate of the Early-Middle Jur-assic Cleveland Basin, England, Palaios, 14,261–272, 1999.
Nursall, J. R., Distribution and ecology of pyc-nodont fishes, in Mesozoic Fishes 2- Systema-tics and Fossil Record, edited by G. Arratiaand H.-P. Schultze, pp. 115 –124, F. Pfeil,Munich, 1996.
O’Neil, J. R., R. N. Clayton, and T. K. Mayeda,Oxygen isotope fractionation in divalent metalcarbonates, J. Chem. Phys., 51, 5547–5558,1969.
O’Neil, J. R., L. J. Roe, E. Reinhard, and R. E.Blake, A rapid and precise method of oxygenisotope analysis of biogenic phosphate, Isr.J. Earth Sci., 43, 203–212, 1994.
Philippe, M., and F. Thevenard, Distribution andpaleoecology of the Mesozoic wood genus Xe-noxylon: Palaeoclimatological implications forthe Jurassic of western Europe, Rev. Palaeo-bot. Palyno., 91, 353–370, 1996.
Picard, S., J. P. Garcia, C. Lecuyer, S. M. F.Sheppard, H. Cappetta, and C. Emig, d18Ovalues of coexisting brachiopods and fish: Tem-perature differences and estimates of paleo-water depths, Geology, 26, 975–978, 1998.
Podlaha, O. G., J. Mutterlose, and J. Veizer, Pre-servation of d18O and d13C in belemnite rostrafrom the Jurassic/Early Cretaceous succes-sions, Am. J. Sci., 298, 324–347, 1998.
Poyato-Ariza, F. J., M. R. Talbot, M. AFregenal-Martınez, N. Melendez, and S. Wenz,First isotopic and multidisciplinary evidencefor nonmarine coelacanths and pycnodonti-form fishes: Palaeoenvironmental implications,Palaeogeogr. Palaeoclimatol. Palaeoecol.,144, 65–84, 1998.
Price, G., The evidence and implications of polarice during the Mesozoic, Earth Planet. Sci.Lett., 48, 183–210, 2000.
Puceat, E., C. Lecuyer, S. M. F. Sheppard, G.Dromart, S. Reboulet, and P. Grandjean, Ther-mal evolution of Cretaceous Tethyan marinewaters inferred from oxygen isotope composi-tion of fish tooth enamels, Paleoceanography,18(2),1029,doi:10.1029/2002PA000823,2003.
Riboulleau, A., F. Baudin, V. Daux, P. Hantzper-gue, M. Renard, and V. Zakharov, Evolution de
la paleotemperature des eaux de la plateformerusse au cours du Jurassique superieur, C. R.Acad. Sci., Ser. II, 326, 239–246, 1998.
Riding, J. B., and N. L. B. Hubbard, Jurassic(Toarcian to Kimmeridgian) dinoflagellatecysts and paleoclimates, Palynology, 23, 15–30, 1999.
Robin, C., F. Guillocheau, P. Allemand, S. Bour-quin, G. Dromart, J.-M. Gaulier, and C. Prijac,Echelles de temps et d’espace du controle tec-tonique d’un bassin flexural intracratonique: Lebassin de Paris, Bull. Soc. Geol. Fr., 171, 181–196, 2000.
Roth, P. H., Ocean circulation and calcareousnannoplankton evolution during the Jurassicand Cretaceous, Palaeogeogr. Palaeoclimatol.Palaeoecol., 74, 111–126, 1989.
Saelen, G., P. Doyle, and M. R. Talbot, Stable-isotope analyses of Belemnite rostra from theWhitby Mudstone Fm, England: Surface waterconditions during deposition of a marine blackshale, Palaios, 11, 97–117, 1996.
Savin, S. M., The history of the Earth’s surfacetemperature during the past 100 million years,Ann. Rev. Earth Planet. Sci., 5, 319 –355,1977.
Sharp, Z. D., V. Aturodei, and H. Furrer, Theeffect of diagenesis on oxygen isotope ratiosof biogenic phosphates, Am. J. Sci., 300, 222–237, 2000.
Van Aarssen, B. G. K., R. Alexander, and R. I.Kagi, Higher plant biomarkers reflect palaeo-vegetation changes during Jurassic times, Geo-chim. Cosmochim. Acta, 64, 1417 – 1424,2000.
Veizer, J., et al., Oxygen isotope evolution ofPhanerozoic seawater, Palaeogeogr. Palaeocli-matol. Palaeoecol., 132, 159–172, 1997.
Vennemann, T. W., and E. Hegner, Oxygen,strontium, and neodymium isotope composi-tion of fossil shark teeth as a proxy for thepalaeoceanography and palaeoclimatology ofthe Miocene northern Alpine Paratethys,Palaeogeogr. Palaeoclimatol. Palaeoecol.,142, 107–121, 1998.
Vennemann, T. W., E. Hegner, G. Cliff, and G. W.Benz, Isotopic composition of recent sharkteeth as a proxy for environmental conditions,Geochim. Cosmochim. Acta, 65, 1583–1599,2001.
Vidier, J.-P., J.-P. Garcia, J. Thierry, andD. Fauconnier, Le Dogger du Boulonnais:Nouveaux decoupages chronologiques et se-quentiel des formations carbonatees jurassi-ques en bordure du massif Londres-Brabant,C. R. Acad. Sci., Ser. II, 320, 219–226, 1995.
Ziegler, P. A., Evolution of the Artic-NorthAtlantic and the western Tethys, Am. Assoc.Petrol. Geol. Mem., 43, 198, 1988.
�������������������������G. Dromart, P. Grandjean, C. Lecuyer, and
S. Picard, Laboratoire Paleoenvironnements etPaleobiosphere, CNRS UMR 5125, Campus de laDoua, Universite Claude Bernard Lyon 1, F-69622 Villeurbanne, France.([email protected]; [email protected];[email protected]; [email protected])J.-P. Garcia, Centre des Sciences de la Terre,
CNRS UMR 5561, Universite de Bourgogne,6 Boulevard Gabriel, F-21000 Dijon, France.([email protected])S. M. F. Sheppard, Laboratoire de Sciences de
la Terre, CNRS UMR 5570, Ecole NormaleSuperieure de Lyon, 46 allee d’Italie, F-69364Lyon, France. ([email protected])
21 - 16 LECUYER ET AL.: TEMPERATURES OF JURASSIC SEAWATER





























