Embed Size (px)
Citation preview

The Transcription Factor VvMYB5b Contributes to theRegulation of Anthocyanin and ProanthocyanidinBiosynthesis in Developing Grape Berries1,2[W]
Laurent Deluc3,4, Jochen Bogs3,5, Amanda R. Walker, Thilia Ferrier, Alain Decendit, Jean-Michel Merillon,Simon P. Robinson, and Francxois Barrieu*
Institut des Sciences de la Vigne et du Vin, UMR 1287 Ecophysiologie et Genomique Fonctionnelle de laVigne INRA, Universite de Bordeaux 1, Universite Victor Segalen Bordeaux 2, Centre INRA de Bordeaux,33883 Villenave d’Ornon, France (L.D., T.F., F.B.); CSIRO Plant Industry, Glen Osmond, South Australia 5064,Australia (J.B., A.R.W., S.P.R.); and Groupe d’Etude des Substances Vegetales a Activite Biologique, EA 3675,Institut des Sciences de la Vigne et du Vin, Universite Victor Segalen Bordeaux 2, UFR SciencesPharmaceutiques, 33076 Bordeaux cedex, France (A.D., J.-M.M.)
Among the dramatic changes occurring during grape berry (Vitis vinifera) development, those affecting the flavonoid pathwayhave provoked a number of investigations in the last 10 years. In addition to producing several compounds involved in theprotection of the berry and the dissemination of the seeds, final products of this pathway also play a critical role in berry andwine quality. In this article, we describe the cloning and functional characterization of VvMYB5b, a cDNA isolated from a grapeberry (V. vinifera ‘Cabernet Sauvignon’) library. VvMYB5b encodes a protein belonging to the R2R3-MYB family of transcriptionfactors and displays significant similarity with VvMYB5a, another MYB factor recently shown to regulate flavonoid synthesisin grapevine. The ability of VvMYB5a and VvMYB5b to activate the grapevine promoters of several structural genes of theflavonoid pathway was confirmed by transient expression of the corresponding cDNAs in grape cells. Overexpression ofVvMYB5b in tobacco (Nicotiana tabacum) leads to an up-regulation of genes encoding enzymes of the flavonoid pathway andresults in the accumulation of anthocyanin- and proanthocyanidin-derived compounds. The ability of VvMYB5b to regulateparticularly the anthocyanin and the proanthocyanidin pathways is discussed in relation to other recently characterized MYBtranscription factors in grapevine. Taken together, data presented in this article give insight into the transcriptionalmechanisms associated with the regulation of the flavonoid pathway throughout grape berry development.
The phenylpropanoid biosynthetic pathway leads tothe synthesis of numerous compounds that play a keyrole in many physiological and biochemical processesin plants (Kutchan, 2005). Some of these compounds,belonging to the flavonoid family, contribute to thepigmentation of flowers, fruit, seeds, and leaves, butare also involved in other plant processes, including
the signaling between plants and microbes (Hahlbrockand Scheel, 1989; Koes et al., 2005), the male fertility ofsome species, and plant defense in the form of anti-microbial agents and feeding deterrents (Dixon et al.,1996). In grapevine (Vitis vinifera), flavonoids such asflavonols and catechins protect the plant against UVradiation, whereas others, such as anthocyanins, helpattract seed-dispersal agents. Flavonoid compoundshave a critical role in the quality of wine by contrib-uting to the bitterness and astringency of wine (proan-thocyanidins [PAs]) or color (anthocyanins) and arealso known to have nutrient and health benefits forhumans. For example, grape seed PA extract has abeneficial effect on physical health, especially affectingthe bones (Kamitani et al., 2004), the heart (Bagchiet al., 2000), and the immune system (Lin et al., 2002).As a consequence, moderate consumption of red winemay reduce risk of cardiovascular disease and cancer(Klatsky, 2002).
In plants, the regulation of the phenylpropanoidbiosynthetic pathway has been extensively studied,and particularly the control of the anthocyanin pathwayis well characterized (Broun, 2005; Koes et al., 2005;Ramsay and Glover, 2005). Three regulatory gene fam-ilies encoding basic helix-loop-helix (bHLH), WDR(WD40 repeats), and MYB-like proteins appear to be
1 This work was supported by grants from the ‘‘Conseil Inter-professionnel du Vin de Bordeaux,’’ The Grape and Wine Researchand Development Corporation and the Cooperative Research Centrefor Viticulture, and the GABI-Future program of the German Min-istry of Education and Research.
2 This article is dedicated to the memory of our friend andcolleague, Professor Saıd Hamdi.
3 These authors contributed equally to the article.4 Present address: Department of Biochemistry, University of
Nevada, 1664 North Virginia St., Reno, NV 89557.5 Present address: Heidelberger Institut fur Pflanzenwissenschaften,
69120 Heidelberg, Germany.* Corresponding author; e-mail [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Francxois Barrieu ([email protected]).
[W] The online version of this article contains Web-only data.www.plantphysiol.org/cgi/doi/10.1104/pp.108.118919
Plant Physiology, August 2008, Vol. 147, pp. 2041–2053, www.plantphysiol.org � 2008 American Society of Plant Biologists 2041 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved. www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved. www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

directly involved and have been extensively charac-terized in various species such as maize (Zea mays;Hernandez et al., 2004), snapdragon (Antirrhinum majus;Schwinn et al., 2006), petunia (Petunia hybrida; Speltet al., 2000), and Ipomoea (Morita et al., 2006). In fruitcrops, characterization studies of MYB factors revealedtheir role in the regulation of the anthocyanin pathwayin strawberry (Fragaria 3 ananassa; Aharoni et al., 2001)and tomato (Solanun lycopersicum; Mathews et al., 2003).Likewise, in apple (Malus domestica), a MYB transcrip-tion factor was recently characterized and its functionsprovided new evidence of the conserved mechanismrelated to the regulation of the flavonoid pathwaywithin the plant kingdom (Takos et al., 2006; Banet al., 2007; Espley et al., 2007). In Arabidopsis (Arabi-dopsis thaliana), it was shown that a combinatorialcontrol through the action of three different families ofregulatory factors (MYB, bHLH, and WDR proteins)modulates the expression of the structural Banyuls(BAN) gene encoding anthocyanidin reductase, whichis directly involved in proanthocyanidin (PA) biosyn-
thesis (Baudry et al., 2004, 2006; Lepiniec et al., 2006). Ingrape berries, even though the identification of struc-tural genes associated with the flavonoid pathway hasbeen undertaken (Boss et al., 1996a; Kennedy et al.,2000; Downey et al., 2003a, 2003b; Bogs et al., 2005), littleis known about the transcriptional regulation of thestructural genes involved in flavonoid biosynthesisthroughout berry development. Recent studies in grape-vine have pointed out the key role of the VvMYBA1and VvMYBA2 transcription factors that regulate spe-cifically the expression of the UDP-Glc:flavonoid3-O-glucosyltransferase (UFGT) gene, which encodes anenzyme responsible for conversion of anthocyanidinsto anthocyanins (Kobayashi et al., 2002, 2004; Walkeret al., 2007). Another R2R3-MYB protein has also beenrecently identified as a key regulator of PA synthesis inberry skin and seeds (Bogs et al., 2007). However, theremaining regulatory genes, controlling the expressionof genes that encode enzymes located upstream ofUFGT, still remain to be identified. The coordinateexpression of some structural genes in berry suggests
Figure 1. Protein sequence alignmentof VvMYB5b and other R2R3-MYBtranscription factors from variousplant species. Identical residues areshown in black, conserved residuesin dark gray, and similar residues inlight gray. The lines above the align-ment locate the different domainsand motifs within the proteins. TheDNA binding domain correspondsto the R2R3-MYB repeats. The IDdomain, involved in the interactionwith bHLH proteins, and the C1 andC3 motifs are in bold and italics.GenBank accession numbers are asfollows (in parentheses): VvMYB5b(AAX51291), VvMYB5a (AAS68190),BNLGHi233 (AAK19611), PH4(AAY51377), AtMYB5 (AAC49311),OsMYB4 (BAA23340), VvMYBPA1(CAJ90831), and VvMYBA1(BAD18977).
Deluc et al.
2042 Plant Physiol. Vol. 147, 2008 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

the contribution of at least two distinct regulatorycomplexes involved in the early and late steps of berrydevelopment, respectively (Boss et al., 1996a). In ourgroup, we have recently identified a MYB gene namedVvMYB5a associated with the regulation of the fla-vonoid pathway during the early phase of berry de-velopment (Deluc et al., 2006). Identification oftranscriptional regulators involved in the later stepsof berry development (i.e. berry ripening) is requiredto understand the coordinate regulatory mechanismsof this biosynthetic pathway throughout berry devel-opment.
In this article, we describe the functional character-ization of a MYB transcription factor named VvMYB5bisolated from a cDNA library of Cabernet Sauvignongrape berries harvested at the onset of ripening,termed the veraison stage. Transcript analysis through-out berry development indicated that the VvMYB5bgene is expressed preferentially during berry ripening.Ectopic expression of VvMYB5b in tobacco (Nicotianatabacum) plants enhanced the expression levels of mostflavonoid structural genes and results in accumulationof anthocyanidin- and PA-derived compounds in re-productive tissues. In grape cells, VvMYB5b is able toactivate several promoters of structural genes in-volved not only in the common steps of the flavonoidpathway but also in some specific branches such as PAsynthesis. Experiments performed with the VvMYB5aprotein (Deluc et al., 2006) lead to similar resultsindicating a possible functional relationship betweenthose two MYB transcription factors. The putativebiological functions of VvMYB5a and VvMYB5b arediscussed according to their expression patterns dur-ing berry development and to the functional charac-terization data in homologous and heterologoussystems. Taken together, data presented in this articlerepresent an important step toward understanding therole of R2R3-MYB transcription factors in the regula-tory mechanisms of the flavonoid biosynthetic path-way in developing grape berries.
RESULTS
VvMYB5b Sequence Analysis
A cDNA clone named VvMYB5b, encoding a puta-tive R2R3-MYB protein, was isolated by PCR from agrape berry cDNA library. The open reading frame(ORF) encodes a protein of 311 amino acid residueswith a predicted mass of 34 kD. The deduced aminoacid sequence contains near its amino-terminal ex-tremity the R2R3 imperfect repeats (DNA bindingdomain) involved in binding to target DNA sequencesand highly conserved within MYB proteins (Fig. 1). Aphylogenetic analysis (Fig. 2) of 28 plant MYB proteinsassociated with different functions indicated thatVvMYB5b belongs to the same cluster as VvMYB5a,another R2R3-MYB protein recently characterized ingrapevine (Deluc et al., 2006). This small cluster con-tains six MYB proteins involved in the control ofvarious physiological and developmental processeslike the phenylpropanoid pathway in grapevine forVvMYB5a and the chilling tolerance in rice (Oryzasativa) for OSMYB4 (Vannini et al., 2004). AtMYB5 maybe involved in cell fate determination and trichomedevelopment in Arabidopsis (Li et al., 1996), whereasPH4 activates vacuolar acidification in petunia(Quattrocchio et al., 2006). The last member of thiscluster, the BNLGHi233 protein from cotton (Gossypiumhirsutum), has not yet been characterized. The otherMYB transcription factors from grapevine, like VvMYBA1and VvMYBA2 (Kobayashi et al., 2002, 2004; Walker
Figure 2. Phylogenetic relationships between VvMYB5b and R2R3-MYBtranscription factors from grape and other plant species. Phylogeneticand evolutionary analyses were performed using the neighbor-joiningmethod by the MEGA version 4 program (Kumar et al., 2004). The scalebar represents the number of substitutions per site and the numbersnext to the nodes are bootstrap values from 1,000 replicates. Theputative regulatory functions of the different R2R3-MYB proteins in thecontrol of secondary metabolite biosynthesis or other biological pro-cesses are indicated. GenBank accession numbers are as follows(in parentheses): VvMYBA1 (BAD18977), VvMYBA2 (BAD18978),VvMYBA3 (BAD18979), LeANT1 (AAQ55181), TT2 (CAC40021),AtMYB75 (AAG42001), PH4 (AAY51377), AmRosea1 (ABB83826),AmVenosa (ABB83828), MdMYB10 (ABB84753), ZmMYBPL(AAB67720), ZmMYBC1 (AAA33482), VvMYBPA1 (CAJ90831),PmMBF1 (AAA82943), OSMYB4 (BAA23340), AtMYB5 (AAC49311),BNLGHi233 (AAK19611), VvMYB5b (AAX51291), VvMYB5a(AAS68190), FaMYB1 (AAK84064), AtMYB7 (AAA98762), HOS10(AAF20989), HvMYB5 (CAA50221), TFMYB6 (AAS19480), GhMYB1(AAN28270), LeTHM27 (CAA64614), TaMYB1 (AAT37167), andZmMYB38 (BAD34380).
Grapevine MYBs Regulate the Flavonoid Pathway
Plant Physiol. Vol. 147, 2008 2043 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.
et al., 2007) or VvMYBPA1 (Bogs et al., 2007), clearlybelong to other clusters, indicating a possible func-tional divergence between these proteins and VvMYB5aand VvMYB5b. Regarding the sequence similarity, theVvMYB5b protein sequence displays 65% and 56%overall similarity to its nearest neighbors, the VvMYB5aand BNLGHi233 proteins, respectively. These overalllevels of similarity appear relevant if we consider thatR2R3-MYB proteins usually exhibit significant simi-larities only within the amino-terminal region wherethe DNA binding domain is located (Fig. 1; Kranzet al., 1998; Stracke et al., 2001).
The alignment of the VvMYB5b protein sequencewith other known R2R3-MYB regulators from variousplant species reveals the appearance of conservedpeptidic motifs in the C-terminal regions of the pro-teins (Fig. 1). Apart from the very well-conservedDNA binding domain, referred to as the R2R3 domain,three distinct motifs were found within the analyzedsequences. The first one, [D/E]Lx2[R/K]x3Lx6Lx3R, isinvolved in the interaction with bHLH proteins andis present in the majority of the R2R3-MYB proteins(ID domain; Fig. 1; Grotewold et al., 2000; Strackeet al., 2001) The second conserved motif, called C1(Lx3GIDPxTHKPL) and initially described by Kranzet al. (1998) in the proteins of the MYB subgroup 4, isfound in the six proteins belonging to the VvMYB5bcluster identified by phylogenetic analysis (Fig. 2).Another conserved motif, tentatively named C3(DDxF[S/P]SFL[N/D]SLIN[E/D]), appears only inthe sequences of the six members of the VvMYB5bcluster and is not found in other MYB proteins char-acterized so far. In conclusion, sequence analysis dataindicate that VvMYB5b belongs to the R2R3-MYBfamily of transcription factors and, more precisely, toa small group of MYB proteins characterized by thepresence of two specific conserved motifs, but involvedin various physiological or developmental processes.
The Expression of VvMYB5b Is Highest in Berries
after Veraison
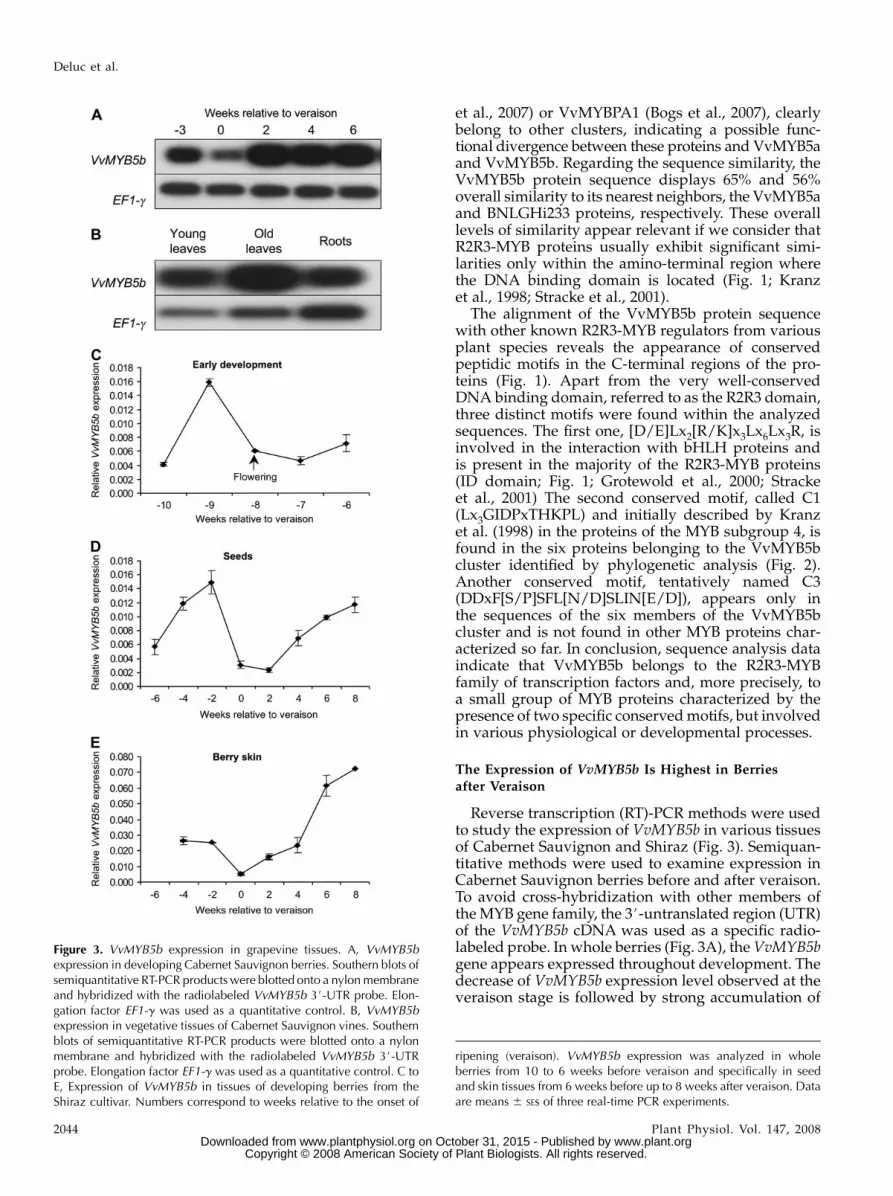
Reverse transcription (RT)-PCR methods were usedto study the expression of VvMYB5b in various tissuesof Cabernet Sauvignon and Shiraz (Fig. 3). Semiquan-titative methods were used to examine expression inCabernet Sauvignon berries before and after veraison.To avoid cross-hybridization with other members ofthe MYB gene family, the 3#-untranslated region (UTR)of the VvMYB5b cDNA was used as a specific radio-labeled probe. In whole berries (Fig. 3A), the VvMYB5bgene appears expressed throughout development. Thedecrease of VvMYB5b expression level observed at theveraison stage is followed by strong accumulation of
Figure 3. VvMYB5b expression in grapevine tissues. A, VvMYB5bexpression in developing Cabernet Sauvignon berries. Southern blots ofsemiquantitative RT-PCR products were blotted onto a nylon membraneand hybridized with the radiolabeled VvMYB5b 3#-UTR probe. Elon-gation factor EF1-g was used as a quantitative control. B, VvMYB5bexpression in vegetative tissues of Cabernet Sauvignon vines. Southernblots of semiquantitative RT-PCR products were blotted onto a nylonmembrane and hybridized with the radiolabeled VvMYB5b 3#-UTRprobe. Elongation factor EF1-g was used as a quantitative control. C toE, Expression of VvMYB5b in tissues of developing berries from theShiraz cultivar. Numbers correspond to weeks relative to the onset of
ripening (veraison). VvMYB5b expression was analyzed in wholeberries from 10 to 6 weeks before veraison and specifically in seedand skin tissues from 6 weeks before up to 8 weeks after veraison. Dataare means 6 SEs of three real-time PCR experiments.
Deluc et al.
2044 Plant Physiol. Vol. 147, 2008 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.
the transcripts 2 weeks after veraison. After this largeincrease, VvMYB5b remained expressed at a high levelup to 6 weeks after veraison.
VvMYB5b expression was also investigated in skinand seeds of berries from the Shiraz cultivar usingreal-time quantitative PCR. During the early stages ofberry development (10–6 weeks before veraison),VvMYB5b expression level remained low in the berrywith a slight transitory increase 9 weeks before theonset of ripening (Fig. 3B). Between 6 and 4 weeksbefore veraison, it was possible to separate skin andseed from the developing berries and thus to analyzeVvMYB5b transcript abundance in both tissues (Fig. 3,C and D). Generally, VvMYB5b expression was higherin skin than in seeds throughout berry development,except at the veraison stage where the expression wasvery low in both tissues. In seeds, VvMYB5b wasalways expressed at lower levels than in skins with amaximal level 2 weeks before veraison (Fig. 3C). In theskin (Fig. 3D), the VvMYB5b expression pattern re-sembles the one observed in Cabernet Sauvignonberries with a decrease of gene expression at theveraison stage followed by a continuous increase oftranscript abundance up to the mature stages (6–8weeks after veraison).
In summary, the VvMYB5b gene is expressed ingrape berries throughout development and also invegetative tissues of grapevine (Fig. 3E), where expres-sion was detected in young and old leaves and roots. Inberries, expression is high in the skin tissues with a firstphase of gene activation observed before veraison anda second one, more pronounced, beginning just afterveraison. It is interesting to note that the VvMYB5bexpression pattern during berry development was very
similar in Cabernet Sauvignon and Shiraz berries grownin France and Australia, respectively. This finding indi-cates that the regulatory mechanisms of gene expressionmay be conserved in two Vitis cultivars exposed todifferent environmental conditions.
VvMYB5b and VvMYB5a Activate Promoters ofGrapevine Flavonoid Genes
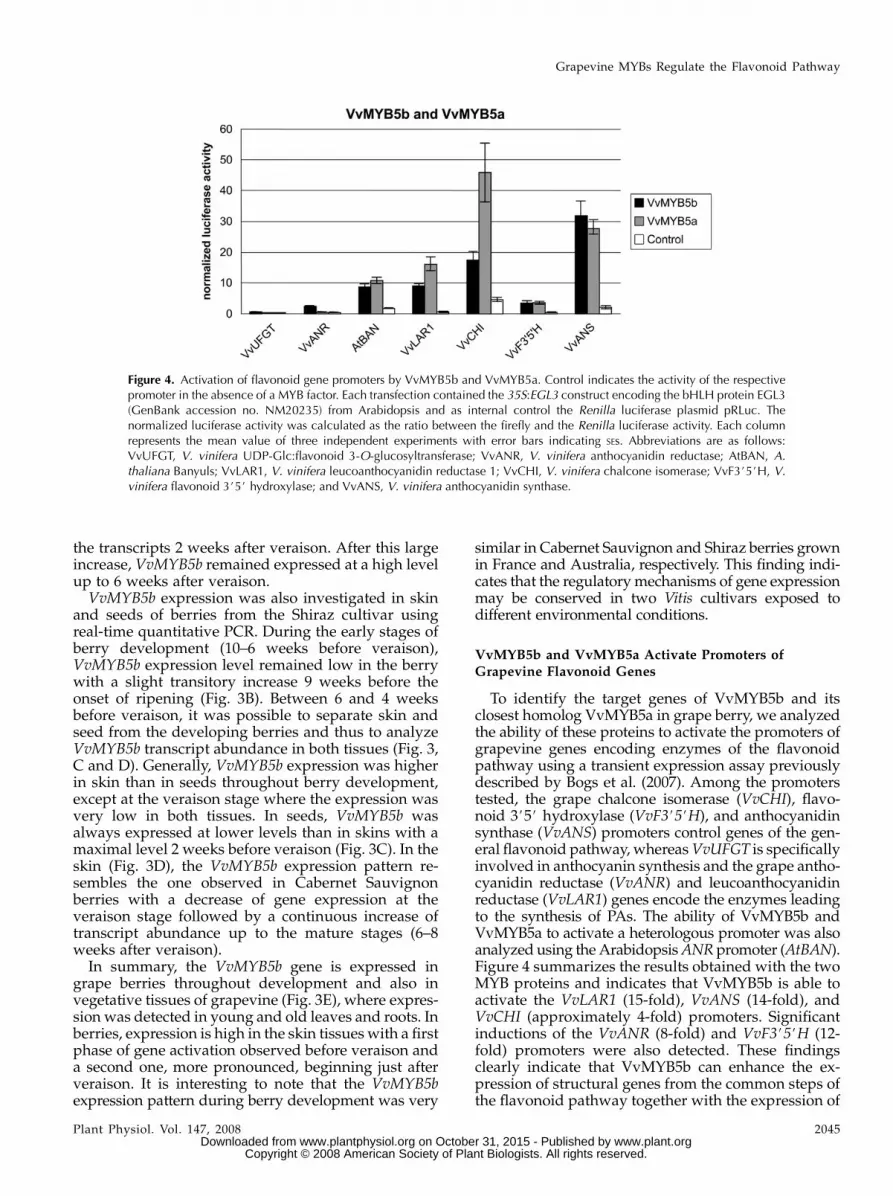
To identify the target genes of VvMYB5b and itsclosest homolog VvMYB5a in grape berry, we analyzedthe ability of these proteins to activate the promoters ofgrapevine genes encoding enzymes of the flavonoidpathway using a transient expression assay previouslydescribed by Bogs et al. (2007). Among the promoterstested, the grape chalcone isomerase (VvCHI), flavo-noid 3#5# hydroxylase (VvF3#5#H), and anthocyanidinsynthase (VvANS) promoters control genes of the gen-eral flavonoid pathway, whereas VvUFGT is specificallyinvolved in anthocyanin synthesis and the grape antho-cyanidin reductase (VvANR) and leucoanthocyanidinreductase (VvLAR1) genes encode the enzymes leadingto the synthesis of PAs. The ability of VvMYB5b andVvMYB5a to activate a heterologous promoter was alsoanalyzed using the Arabidopsis ANR promoter (AtBAN).Figure 4 summarizes the results obtained with the twoMYB proteins and indicates that VvMYB5b is able toactivate the VvLAR1 (15-fold), VvANS (14-fold), andVvCHI (approximately 4-fold) promoters. Significantinductions of the VvANR (8-fold) and VvF3#5#H (12-fold) promoters were also detected. These findingsclearly indicate that VvMYB5b can enhance the ex-pression of structural genes from the common steps ofthe flavonoid pathway together with the expression of
Figure 4. Activation of flavonoid gene promoters by VvMYB5b and VvMYB5a. Control indicates the activity of the respectivepromoter in the absence of a MYB factor. Each transfection contained the 35S:EGL3 construct encoding the bHLH protein EGL3(GenBank accession no. NM20235) from Arabidopsis and as internal control the Renilla luciferase plasmid pRLuc. Thenormalized luciferase activity was calculated as the ratio between the firefly and the Renilla luciferase activity. Each columnrepresents the mean value of three independent experiments with error bars indicating SEs. Abbreviations are as follows:VvUFGT, V. vinifera UDP-Glc:flavonoid 3-O-glucosyltransferase; VvANR, V. vinifera anthocyanidin reductase; AtBAN, A.thaliana Banyuls; VvLAR1, V. vinifera leucoanthocyanidin reductase 1; VvCHI, V. vinifera chalcone isomerase; VvF3#5#H, V.vinifera flavonoid 3#5# hydroxylase; and VvANS, V. vinifera anthocyanidin synthase.
Grapevine MYBs Regulate the Flavonoid Pathway
Plant Physiol. Vol. 147, 2008 2045 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

specific genes associated with the PA biosyntheticpathway such as VvLAR1 and VvANR.
Our results also indicate that the VvMYB5a proteinis able to activate the VvLAR1 (27-fold), VvANS (12.5-fold), VvF3#5#H (12-fold), and VvCHI (7-fold) pro-moters (Fig. 4). However, neither VvMYB5a norVvMYB5b appears to play a key role in the control ofthe VvUFGT promoter where the level of activation wasvery low. In contrast, both VvMYB5a and VvMYB5bappeared able to activate the Arabidopsis AtBAN pro-moter with almost the same efficiency (between 4- and5-fold). All promoter assays included a construct ex-pressing AtEGL3 (ENHANCER OF GLABRA3) in thetransient expression system, which encodes a bHLHprotein involved in flavonoid pathway regulation in
Arabidopsis (Ramsay et al., 2003). Similar to otherVvMYB regulators, VvMYB5a and VvMYB5b werenot able to induce promoter activities significantlywithout expression of AtEGL3 in our transient assay(data not shown; Bogs et al., 2007; Walker et al., 2007).
Taken together, promoter activation data indicatethat VvMYB5a and VvMYB5b are both able to activatethe expression of genes encoding enzymes of thegeneral flavonoid pathway involved in synthesis ofanthocyanins, PAs, and flavonols (e.g. VvCHI, VvF3#5#H,and VvANS). In addition, VvMYB5b can activate the ex-pression of the two genes specifically involved in PA bio-synthesis (e.g. VvLAR1 and VvANR), whereas VvMYB5aappears only implicated in the control of VvLAR1 ex-pression.
Figure 5. Phenotypic analysis of trans-genic tobacco flowers overexpressingVvMYB5b. Flowers of transgenic plants (Band C) showed an increased pigmentationin petal and stamen epidermal cells com-pared to control flowers (A). DMACA stain-ing of petals (E) and stamen (G) cells fromtransgenic flowers shows condensed tan-nin accumulation when compared to con-trol lines (D and F). H and I, Delay in pollenrelease observed in anthers at developmen-tal stages 10, 11, and 12 (from left to right,according to Koltunow et al. [1990]) fromVvMYB5b sense lines (I) compared to con-trol plant anthers (H). J and K, Cross sec-tions of anthers at stage 10 from control(J) and VvMYB5b-overexpressing tobaccoplants (K). Boxed areas are magnified in Land M, respectively. Arrows indicate en-dothecial cells developing complete (L) orincomplete (M) lignified fibers. Bars 5 500mm (J and K) and 75 mm (L and M).
Deluc et al.
2046 Plant Physiol. Vol. 147, 2008 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Accumulation of Anthocyanin- and PA-DerivedCompounds in Transgenic Tobacco FlowersExpressing VvMYB5b
Significant changes in the pigmentation of flowerswere observed in tobacco plants constitutively ex-pressing VvMYB5b. The strongest modifications werefound in stamens where the amount of pigmentationwas much greater in transgenic stamens at variousdevelopmental stages (Fig. 5, B and C) when com-pared to control plants (Fig. 5A). Transgenic petals alsoappeared deeper pink than for control flowers. Thisenhanced reddish pigmentation of petals resultedfrom an unusual distribution of reddish and uncoloredcells in the epidermal layer (Fig. 5, B and C). Quanti-fication of anthocyanin content was achieved by spec-trophotometry (535 nm) in stamens and petals (TableI). An accumulation of total anthocyanins was de-tected in transgenic stamens (2.52 6 0.08 mg/g of dryweight), whereas no significant level of anthocyaninswas detected in control stamens. In petals, the totalanthocyanin content in transgenic lines was nearlytwice (2.96 6 0.09 mg/g of dry weight) the amountmeasured in petals of control plants (1.48 6 0.26 mg/gof dry weight). Like for the VvMYB5a transcriptionfactor (Deluc et al., 2006), HPLC analysis indicates thatthe increase of anthocyanins in transgenic petals andstamens is linked to the accumulation of keracyanin(cyanidin-3-rhamnoglucoside), which is the main an-thocyanin compound in tobacco (data not shown).
Additional analyses of the transgenic plants alsorevealed modifications of the PA metabolic pathway.Accumulation of condensed tannins in petals andstamens was detected using dimethylaminocinnamal-dehyde (DMACA) staining (Xie et al., 2003). A bluecoloration, linked to the presence of PA, was observedin the epidermal cell layers of petals and anther tips oftransgenic flowers (Fig. 5, E and G), but not in controlline flowers (Fig. 5, D and F). HPLC analysis andquantification confirmed a strong accumulation of freemonomer units of epicatechin-derived compound intransgenic petals (10.51 6 0.09 mg/g of fresh weight)compared to control petals (1.48 6 0.26 mg/g of freshweight; Table II). Taken together, these results indi-
cated a strong accumulation of anthocyanins and PAsin flowers of plants overexpressing VvMYB5b andappeared similar to those obtained with the VvMYB5acDNA (Deluc et al., 2006), suggesting similar regula-tory functions in tobacco for both MYB factors. Aspreviously observed with VvMYB5a (Deluc et al.,2006), a delay in anther dehiscence was observed.Whereas the pollen was completely released 12 weeksafter bloom in control plants (Fig. 5H), the antherswere still closed in VvMYB5b transgenic sense lines(Fig. 5I). Cross section analyses of the anthers fromVvMYB5b sense lines revealed changes in the lignifi-cation network of the endothecial cell wall, responsibleof the stomium breaking and leading to the pollenrelease. In transformed plants, fewer endothecial cellsdeveloped intact lignified fibers. Most of the time, theywere incomplete as observed in Figure 5M comparedto the control that exhibited complete fibers coveringthe entire radial walls (Fig. 5L).
VvMYB5b Affects the Expression of Flavonoid
Biosynthetic Genes in Transgenic Tobacco Flowers
Expression analysis performed on RNA extractedfrom petals and stamens of three independent trans-genic lines of tobacco expressing the VvMYB5b cDNAindicated that VvMYB5b activated the expression ofseveral structural genes associated with the flavonoidpathway (Fig. 6, A and B). In petals and stamens oftransgenic plants, expression of the chalcone synthase(CHS), chalcone isomerase (CHI), flavanone 3 hydrox-ylase (F3H), and anthocyanidin synthase (ANS) geneswas higher than that in control flowers (Fig. 6, A andB). In addition, dihydroflavonol reductase (DFR) geneexpression appeared differentially regulated in trans-genic petals and stamens indicating the appearance ofadditional regulatory mechanisms. These results indi-cate the involvement of VvMYB5b in the regulation offlavonoid genes expression in tobacco flowers. How-ever, this regulatory action may vary depending on thetissue studied as suggested by the contrasting expres-sion of the DFR gene in transgenic flowers.
DISCUSSION
VvMYB5b Encodes a New Member of the R2R3-MYB
Family in Grapevine
During their development, grape berries accumu-late many different products of the flavonoid pathway.Although there is a separation in timing during whichPAs, flavonols, or anthocyanins are synthesized, tis-sues such as the skin contain relatively large quantitiesof these compounds at ripeness. The regulatory mech-anisms involved in the control of the complex branch-ing pathway that leads to the synthesis of thesevarious flavonoid compounds remain unclear. In thisarticle, we present the identification and the charac-terization of the grape VvMYB5b transcriptional reg-ulator and describe one of the missing parts of this
Table I. Total anthocyanin content (mg/g DW) of stamens and petalextracts from control and transgenic plants
Quantitative determination of total anthocyanin content in tobaccoflowers has been performed by spectrophotometry at 535 nm. Apurified keracyanin standard of was used as a control. Data are themean of three replicates 6 SD. DW, Dry weight tissue; N.D., notdetected.
Sample Stamens Petals
VvMYB5b line A 2.74 6 0.08 2.87 6 0.05a
VvMYB5b line B 2.48 6 0.07 2.98 6 0.09a
VvMYB5b line C 2.35 6 0.09 3.03 6 0.13a
Control N.D. 1.48 6 0.26
aSignificantly different from wild type (P , 0.001; Student’s t test).
Grapevine MYBs Regulate the Flavonoid Pathway
Plant Physiol. Vol. 147, 2008 2047 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

regulatory puzzle. Phylogenetic analyses indicate thatthe VvMYB5b protein is clearly distinct from theVvMYBA factors and belongs to a small cluster thatincludes the recently characterized VvMYB5a gene(Deluc et al., 2006). The presence, in this small cluster,of the Arabidopsis AtMYB5 protein may be interestingif we consider that AtMYB5 appears as a singleisolated MYB gene in Arabidopsis and does not belongto any identified cluster of R2R3-MYB proteins (Kranzet al., 1998; Stracke et al., 2001). According to the recentrelease of the grape genome (http://www.genoscope.cns.fr/externe/English/Projets/Projet_ML/index.html),VvMYB5a and b localize to chromosomes 8 and 6,respectively, and appear to be under the control ofdifferent cis-regulatory elements as indicated by theiralmost opposite expression patterns in developingberries. Thus, VvMYB5a and VvMYB5b may representparalogous genes that have originated from a dupli-cation event followed by genomic dispersion. Takentogether, data presented in this article indicate thatthe spatiotemporal expression of VvMYB5b, combinedwith the action of specific regulators like VvMYBA1and VvMYBPA1, controls the biosynthesis of both
anthocyanins and PAs throughout grape berry devel-opment (Fig. 7). Moreover, taking into account thefindings of Deluc et al. (2006), our results suggest thatVvMYB5b and VvMYB5a encode functionally relatedproteins and that their unique roles in the control ofthe flavonoid pathway in grape berry may be entirelydue to differences in their cis-regulatory sequences, asalready demonstrated in Arabidopsis for the WER andGL1 paralogous MYB genes (Lee and Schiefelbein,2001).
Implication of VvMYB5b in the Regulatory
Mechanisms of Anthocyanin Biosynthesis
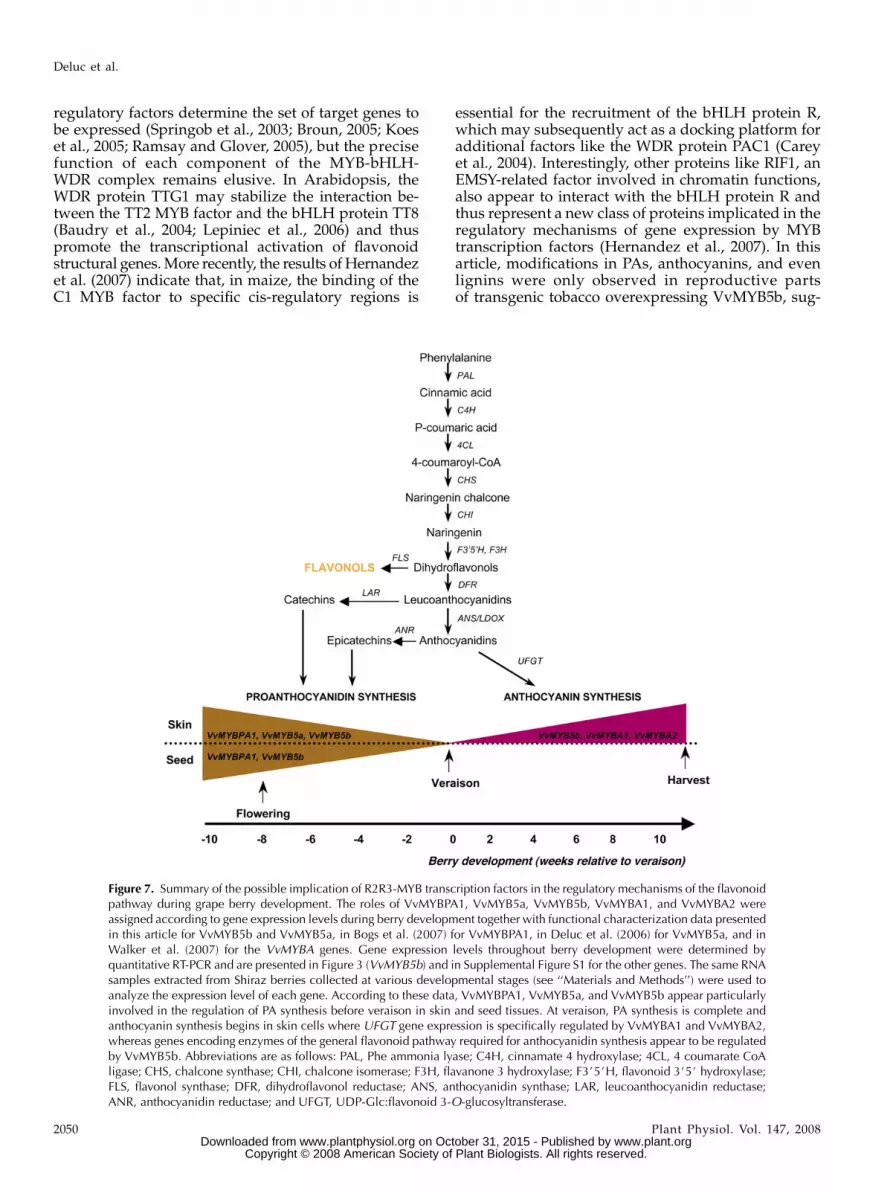
Promoter activation experiments in grape cells in-dicated that VvMYB5b is able to activate several genesof the general pathway, but not UFGT, the last enzymeof the anthocyanin biosynthetic pathway. However,the recent findings of Walker et al. (2007), combinedwith previous work from Kobayashi et al. (2002, 2004),clearly indicate the role of the two MYB genes VvMYBA1and VvMYBA2 in the specific control of UFGT geneexpression in ripening grape berries. Interestingly, bothMYBA factors are not functional in white grapes andUFGT is not expressed, whereas the genes encodingenzymes of the general flavonoid pathway (e.g. CHS,DFR, F3H, ANS), which produce anthocyanin precur-sors (i.e. anthocyanidin) are still expressed in whitegrapes after veraison (Boss et al., 1996b; Walker et al.,2007). These observations suggest the presence of anadditional regulator controlling the expression of genesencoding enzymes of the general flavonoid pathwaywhen anthocyanins are synthesized. Therefore, accord-ing to its high expression in skin after veraison andits ability to activate the promoters of general flavo-noid genes in grape cells, we postulate that VvMYB5bacts together with VvMYBA1 and VvMYBA2 to regu-late anthocyanin biosynthesis in ripening grape berries(Fig. 7).
Table II. Epicatechin and catechin content (mg/g FW) of petalextracts from control and transgenic plants
Quantitative determination of catechin and epicatechin content intobacco flowers has been performed by HPLC. The eluate was mon-itored at 280 nm. Purified standards of catechin and epicatechin wereused as controls. Data are the mean of two replicates 6 SD. FW, Freshweight tissue; N.D., not detected.
Sample Catechins Epicatechins
VvMYB5b line A N.D. 9.65 6 0.05a
VvMYB5b line B N.D. 10.84 6 0.09a
VvMYB5b line C N.D. 11.05 6 0.13a
Control N.D. 1.48 6 0.26
aSignificantly different from the control (P , 0.005; Student’s t test).
Figure 6. Analysis of flavonoid gene expression in flow-ers of transgenic tobacco overexpressing VvMYB5b. A,Transcripts for four flavonoid biosynthetic genes weredetected by semiquantitative RT-PCR in petals and sta-mens from three VvMYB5b independent lines (1–3) andcompared to control lines (Ct). VvMYB5b indicates thetransgene expression level. UBI was used as a quantita-tive control. Abbreviations are as follows: CHS, chalconesynthase; CHI, chalcone isomerase; F3H, flavanone 3hydroxylase; and F3#5#H, flavonoid 3#5# hydroxylase. B,Northern-blot analysis of anthocyanidin synthase (ANS)gene expression in petals and stamens from control (Ct)and VvMYB5b lines. Total RNA was extracted fromtissues harvested at the opening of flowers. Experimentswere repeated with three VvMYB5b and control lineswith similar results.
Deluc et al.
2048 Plant Physiol. Vol. 147, 2008 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

VvMYB5b Participates in the Regulation of PA
Biosynthesis in Developing Grape Berries
In grape, PA synthesis is complete by veraison, afterwhich expression of genes encoding ANR and LARdeclines and then remains very low during berryripening (Davies and Robinson, 2000; Downey et al.,2003a, 2003b; Bogs et al., 2005). Catechins, synthesizedby LAR, are the predominant flavan-3-ols found inskin, whereas epicatechin, synthesized by ANR, are themain flavan-3-ols present in grape seeds (Bogs et al.,2005). To date, the R2R3-MYB protein VvMYBPA1 isthe only specific regulator of PA synthesis identifiedin grapevine (Bogs et al., 2007). However, data pre-sented in this article indicate that VvMYB5b may alsobe involved in the regulatory mechanisms of PAbiosynthesis. Expression of VvMYB5b was observedin flowers and in the early steps of berry developmentin both seed and skin tissues. Combined with itsability to activate the VvLAR1 and, to a lesser extent,the VvANR promoters, this finding suggests thatVvMYB5b could play a part in the regulation of bothcatechin and epicatechin biosynthesis occurring dur-ing the early stages of berry development. However,because VvMYB5b appeared expressed at a higherlevel in young berry skin compared to seed and acti-vated preferentially the VvLAR1 promoter, its mainfunction in PA biosynthesis regulation seems to be thecontrol of catechin synthesis in skin cells.
In this article, we also provide additional data re-garding the biological function of VvMYB5a, an R2R2-MYB transcription factor expressed only during thefirst phase of berry development (i.e. before veraison;Deluc et al., 2006). The similarities observed betweenVvMYB5a and VvMYB5b include the appearance ofthe same phenotypic changes in transgenic tobaccoplants. In both cases, activation of flavonoid structuralgene expression accompanied by the accumulation ofanthocyanin and PA-derived compounds was ob-served in transgenic flowers. The analysis of promoteractivation by transient expression of VvMYB5a ingrape cells showed that this transcription factor isable to induce promoters of the general flavonoidpathway genes (e.g. VvANS, VvF3#5#H, or VvCHI), ofthe catechin-specific gene VvLAR1, but not of theVvANR gene. Thus, these new functional characteriza-tion data indicate that VvMYB5a appears also involvedin the regulation of PA biosynthesis and especially inthe control of catechin synthesis in skin cells.
In summary, VvMYBPA1, VvMYB5a, and VvMYB5bappear able to regulate PA biosynthesis in developing
grape berries and their expression patterns in differentberry tissues combined with their target gene speci-ficities could contribute to the different levels andcatechin/epicatechin composition of PAs detected inskins and seeds (Fig. 7; Downey et al., 2003a; Bogset al., 2005).
Evidence for Additional Regulators of the FlavonoidBiosynthetic Pathway in Grape Berry
In some cases, gene expression patterns do notalways correlate exactly with physiological events ormetabolite biosynthetic activities unravel by func-tional characterization approaches. As an example, inthis article, promoter activation assays indicated theability for VvMYB5b to activate VvLAR and VvANRpromoters. However, VvMYB5b is expressed in berriesafter veraison when PA biosynthesis appears to becomplete and both VvANR and VvLAR expressionremains very low (Bogs et al., 2005). A similar situationwas observed by Bogs et al. (2007) with the VvMYBPA1gene. The discrepancy of the ability for VvMYB5b orVvMYBPA1 to activate VvLAR and VvANR promoterswhile they are expressed after veraison when antho-cyanins begin accumulating and PA biosynthesis iscomplete is unclear. Differences between transcrip-tional regulators and target gene expression imply theeffect of additional regulatory mechanisms. In the caseof PA synthesis, one possibility is the expression inberries after veraison of a strong negative regulatorthat could act by direct repression or by competitionwith activators on binding motifs of the VvLAR andVvANR promoters and thus prevent transcription.Repression of phenylpropanoid biosynthesis by MYBtranscription factors has already been demonstrated(Jin et al., 2000; Aharoni et al., 2001). In strawberry, theFaMYB1 protein represses flavonoid gene transcrip-tion and anthocyanin synthesis at the latter stages offruit maturation (Aharoni et al., 2001). FaMYB1-relatedgenes are likely to be present in grape and may havesimilar effects on PA biosynthesis and thus may refinethe control of the timing of PA accumulation duringberry ripening.
An alternative hypothesis related to the decline ofPA gene expression after veraison may be the absenceof an interacting protein partner for the MYB proteins.It is now well established that the transcriptionalregulators for flavonoid biosynthesis include membersof the R2R3-MYB family, the bHLH family, and theWDR proteins. Combinations between these three
Table III. Primers for RT-PCR analysis
Genes Accession No. Sense Primers Antisense Primers
NtCHS AF311783 5#-CCTTTGGGAATTTCTGATTGG 5#-TCCCACAATATAAGCCCAAGCNtF3H AF036169 5#-AGCTAGAGACTACTCCAGGTG 5#-AACCGTGATCCAAGTTTTGCCANtUBI U66264 5#-TCCAGGACAAGGAGGGTAT 5#-GAGACCTCAAGTAGACAAAGCVvMYB5b3#-UTR AY899404 5#-GATGACAGCCGGTGTTCTTT 5#-AACAACACAACCACATACAVvEF1-g AF176496 5#-GCGGGCAAGAGATACCTCAA 5#-TCAATCTGTCTAGGAAAGGAAG
Grapevine MYBs Regulate the Flavonoid Pathway
Plant Physiol. Vol. 147, 2008 2049 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.
regulatory factors determine the set of target genes tobe expressed (Springob et al., 2003; Broun, 2005; Koeset al., 2005; Ramsay and Glover, 2005), but the precisefunction of each component of the MYB-bHLH-WDR complex remains elusive. In Arabidopsis, theWDR protein TTG1 may stabilize the interaction be-tween the TT2 MYB factor and the bHLH protein TT8(Baudry et al., 2004; Lepiniec et al., 2006) and thuspromote the transcriptional activation of flavonoidstructural genes. More recently, the results of Hernandezet al. (2007) indicate that, in maize, the binding of theC1 MYB factor to specific cis-regulatory regions is
essential for the recruitment of the bHLH protein R,which may subsequently act as a docking platform foradditional factors like the WDR protein PAC1 (Careyet al., 2004). Interestingly, other proteins like RIF1, anEMSY-related factor involved in chromatin functions,also appear to interact with the bHLH protein R andthus represent a new class of proteins implicated in theregulatory mechanisms of gene expression by MYBtranscription factors (Hernandez et al., 2007). In thisarticle, modifications in PAs, anthocyanins, and evenlignins were only observed in reproductive partsof transgenic tobacco overexpressing VvMYB5b, sug-
Figure 7. Summary of the possible implication of R2R3-MYB transcription factors in the regulatory mechanisms of the flavonoidpathway during grape berry development. The roles of VvMYBPA1, VvMYB5a, VvMYB5b, VvMYBA1, and VvMYBA2 wereassigned according to gene expression levels during berry development together with functional characterization data presentedin this article for VvMYB5b and VvMYB5a, in Bogs et al. (2007) for VvMYBPA1, in Deluc et al. (2006) for VvMYB5a, and inWalker et al. (2007) for the VvMYBA genes. Gene expression levels throughout berry development were determined byquantitative RT-PCR and are presented in Figure 3 (VvMYB5b) and in Supplemental Figure S1 for the other genes. The same RNAsamples extracted from Shiraz berries collected at various developmental stages (see ‘‘Materials and Methods’’) were used toanalyze the expression level of each gene. According to these data, VvMYBPA1, VvMYB5a, and VvMYB5b appear particularlyinvolved in the regulation of PA synthesis before veraison in skin and seed tissues. At veraison, PA synthesis is complete andanthocyanin synthesis begins in skin cells where UFGT gene expression is specifically regulated by VvMYBA1 and VvMYBA2,whereas genes encoding enzymes of the general flavonoid pathway required for anthocyanidin synthesis appear to be regulatedby VvMYB5b. Abbreviations are as follows: PAL, Phe ammonia lyase; C4H, cinnamate 4 hydroxylase; 4CL, 4 coumarate CoAligase; CHS, chalcone synthase; CHI, chalcone isomerase; F3H, flavanone 3 hydroxylase; F3#5#H, flavonoid 3#5# hydroxylase;FLS, flavonol synthase; DFR, dihydroflavonol reductase; ANS, anthocyanidin synthase; LAR, leucoanthocyanidin reductase;ANR, anthocyanidin reductase; and UFGT, UDP-Glc:flavonoid 3-O-glucosyltransferase.
Deluc et al.
2050 Plant Physiol. Vol. 147, 2008 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

gesting interaction with tissue-specific partners. Forthe grapevine MYB proteins VvMYB5a, VvMYB5b,and VvMYBPA1, the presence of a bHLH protein wasessential to activate gene expression in our promoterexperiments, indicating the necessity of an interactingpartner for both proteins. Thus, the lack of a specificinteracting partner after veraison in skin cells mightexplain the absence of VvANR and VvLAR expressioneven in the presence of MYB regulators. This hypoth-esis implies that MYB factors and their interactingproteins not only cooperate in directing tissue-specificproduction of flavonoid compounds, but also that aparticular interaction may control a specific branch ofthe pathway like PA biosynthesis (Hartmann et al.,2005; Park et al., 2007). In any case, future work willalso have to integrate the important role of bHLHs,WDRs, and EMSY-related proteins in the control andthe specificity of the mechanisms of gene expressioninvolving MYB transcription factors.
MATERIALS AND METHODS
Plant Material and Growth Conditions
Berries from grape (Vitis vinifera ‘Cabernet Sauvignon’) plants from
Domaine du Grand Parc (INRA) were sampled at 2-week intervals during
the 2002 and 2003 growing seasons. Grapevine tissues of Shiraz grape were
collected from a commercial vineyard (Adelaide, Australia) during the 2000 to
2001 season. Approximately 100 berries from at least 20 bunches were
collected at weekly intervals throughout berry development from floral
initiation until harvest, as described by Downey et al. (2003a). Berry devel-
opment stages were chosen according to criteria including size, titers of
soluble sugars, and softening and color of the berries (Boss et al., 1996a;
Downey et al., 2003a). Berries for RNA extraction were sampled and imme-
diately frozen in liquid nitrogen before storage at 280�C pending further
analysis. Separate skin and flesh samples were obtained by peeling fresh
berries just before freezing in liquid nitrogen.
Control and transgenic tobacco (Nicotiana tabacum ‘Xanthii’) seeds were
sterilized in 70% ethanol for 2 min, incubated in 2.5% potassium hypochlorite
solution for 10 min, and finally washed three times in sterile water. After cold
treatment at 4�C for 48 h, seeds were germinated in tissue culture conditions at
23�C under a 16-h-light/8-h-dark regime on Murashige and Skoog medium
(Murashige and Skoog, 1962) containing 3% (w/w) Suc and 100 mg/mL
kanamycin for transgenic plants. Tobacco plants were finally transferred to
soil and grown in individual pots in a greenhouse.
Cloning of VvMYB5b cDNA and Plant Transformation
A cDNA library from grape Cabernet Sauvignon berries (veraison stage)
was constructed using the Smart cDNA library construction kit (CLONTECH)
and used as template for PCR cloning. Two degenerate oligonucleotides from
plant MYB cDNA conserved sequences, 5#-GANRTMAARAAYTAYTGGA-
ACWCN-3# in forward and 5#-NGTGTTCCARTARTTYTTKAYNTC-3# in
reverse orientation, were used in combination with the T7 and 5# primers
located in the pTriplex vector (CLONTECH). PCR products were subse-
quently ligated to pGemTeasy (Promega) and the insert DNA was sequenced
on both strands (Genome Express). Specific oligonucleotides were then
defined within the 5# and 3# noncoding regions of sequenced cDNA and
used to amplify the complete full-length cDNA. For plant transformation, the
complete coding sequence of VvMYB5b cDNA was amplified with a specific
forward primer designed to introduce an XbaI restriction site (5#-GGGGT-
CTAGAGAGAAAGAAGAA-3#) and a reverse primer designed to introduce a
SacI restriction site (5#-TTAACTATAGAGCTCATTGCA-3#). The XbaI/SacI
fragment was ligated to the binary vector pGiBin19 (provided by Dr. D. Inze,
Ghent, Belgium) between the 35S promoter of cauliflower mosaic virus and
the nopaline synthase (nos) poly(A) addition site, creating the pGiBin19-
VvMYB5b plasmid. The construct was introduced into the Agrobacterium
tumefaciens LB4401 strain. Leaf disc transformation and regeneration of
transgenic plants were performed as previously described by Horsch et al.
(1986). Transformed tobacco plants were selected using kanamycin (100 mg/
mL) as a selective marker. Fifteen independent lines from the T2 progeny of
transgenic plants showing no phenotypic variations in their broad aspect were
used for further analysis.
RNA Extraction and Analysis
Different methods of RNA extraction were used depending on the plant
material. Total RNA was isolated from the various Shiraz berry tissues as
described in Downey et al. (2003b). For Cabernet Sauvignon berry tissues,
RNA extraction was carried out according to Asif et al. (2000). RNAs from
control and transformed tobacco leaves were extracted using RNeasy plant
mini kits (Qiagen) according to the manufacturer’s instructions. Finally, total
RNAs from tobacco flowers were extracted as described by Verwoerd et al.
(1989). All RNA samples were treated with Rnase-free Dnase I (Promega),
followed by phenol/chloroform extraction and ethanol (95%) precipitation.
No DNA contamination was detected by PCR amplification. For semiquan-
titative RT-PCR assays, gene-specific primers were preferentially chosen in the
3#-UTR regions of the target mRNAs as described by Deluc et al. (2006; Table
III; see Supplemental Fig. S1). To estimate the transcript amounts for the
VvMYB5b transgene in tobacco, we used a forward specific primer of
VvMYB5b (5#-GATAAGCGGTTCTGACAGC-3#) in combination with a re-
verse primer designed from the 3# transcribed region of the nos terminator
(5#-TCATCGCAAGACCGGCAACA-3#). Specific primers for tobacco CHI and
DFR were designed from cDNA clones generously provided by Dr. Cathie
Martin (John Innes Centre), to whom requests may be addressed. In all cases,
cloning and sequencing of the RT-PCR products confirmed the amplification
specificity. The ANS cDNA probe from petunia (Petunia hybrida) was provided
by Dr. Asaph Aharoni (Plant Research International).
For RT-PCR analysis, 1 mg of total RNA was reverse transcribed with
oligo(dT)12-18 in a 20-mL reaction mixture using the Moloney murine leukemia
virus reverse transcriptase (Promega) according to the manufacturer’s in-
structions. After heat inactivation of the reaction mixture, PCR was performed
using 1 mL of the first-strand cDNA sample with 25 pmol of the primers
shown in Table I in a 50-mL reaction. For all experiments presented in this
article, 15 cycles of PCR amplification were performed, except for the
detection of VvMYB5b transgene in tobacco (Fig. 5, A, B, and D), where 20
cycles were performed. RT-PCR products were separated on agarose gels and
analyzed by DNA gel-blot hybridization using random-primed 32P-gene-
specific probes. RNA gel blots were prepared and hybridized according to
standard protocols (Sambrook et al., 1989). Equal loading of RNA samples and
uniform transfer onto membranes were confirmed by ethidium bromide
staining of the RNA gels and hybridization with a 32P-labeled ubiquitin probe.
After hybridization at 42�C, membranes were washed at room temperature
three times for 20 min each in 13 SSC and 0.1% SDS, followed by one wash in
0.23 SSC and 0.1% SDS at 65�C for 15 min before exposure to a phosphor
imager screen (Molecular Imager FX; Bio-Rad).
Expression analysis by real-time PCR was performed according to Bogs
et al. (2007). The primers VvMYB5BF (5#-GGTGTTCTTTAATTTGGCTTCA-3#)
and VvMYB5BR (5#-CACAACAACACAACCACATACA-3#) were used to
amplify a 143-bp PCR product from the 3#-UTR of the VvMYB5b cDNA
(AY899404). The primers MYB5AF (5#-CATGTCTCCCTGAAAATGATGA-3#)
and MYB5AR (5#-TGCAAGGATCCATTTCACATAC-3#) were used to am-
plify a 179-bp PCR product from the 3#-UTR of the VvMYB5a cDNA
(AY555190). Data were normalized according to the VvUbiquitin1 (TC53702;
The Institute for Genomic Research database, VvGi5) gene expression level,
which is relatively stable throughout berry development (Downey et al.,
2003b; Bogs et al., 2005). All samples were measured in triplicate.
HPLC Analyses and Quantification ofAnthocyanin- and PA-Derived Compounds
HPLC analyses and quantification of total anthocyanins and PA-derived
compounds from petal limbs and stamens harvested from transgenic and
control tobacco were performed as described in Deluc et al. (2006). The
histochemical staining of condensed tannins in epidermal cell layers in
tobacco flowers was conducted according to Porter (1989) by staining tissues
in a solution of ethanol: 6 M HCl (1:1) containing 0.1% (w/v) DMACA (Sigma-
Aldrich) for 3 to 6 min, then by washing three times with water.
Grapevine MYBs Regulate the Flavonoid Pathway
Plant Physiol. Vol. 147, 2008 2051 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Transient Transfection Experiments and
Dual-Luciferase Assay
As described in Bogs et al. (2007), a transient assay was applied using a cell
suspension of a Chardonnay petiole callus culture and the dual-luciferase
system. Gold particles were coated with a mixture of DNA constructs (150 ng
of the respective plasmid, giving a total plasmid concentration of 750 ng/shot)
by the method described in Ramsay et al. (2003) and used to bombard
Chardonnay cells at a helium pressure of 350 kPa within a vacuum of 75 kPa
and a distance of 14 cm (Torregrossa et al., 2002). For the dual-luciferase assay,
each bombardment contained a positive control of 3 ng of the Renilla luciferase
plasmid pRluc (Horstmann et al., 2004). Cells were harvested 48 h after
transfection and lysed by grinding on ice in 150 mL of passive lysis buffer
(Promega). After centrifugation of the lysates for 2 min at 500g, measurement
of the luciferase activities was performed with a dual-luciferase reporter assay
system (Promega) by sequential addition of 25 mL of LARII and Stop & Glo to
10 mL of the lysate supernatant. Light emission was measured with a TD-20/
20 luminometer (Turner Design) and the relative luciferase activity was
calculated as the ratio between the firefly and the Renilla (control) luciferase
activity. All transfection experiments were performed in triplicate and each set
of promoter experiment was repeated with similar relative ratios to the
respective control. Cloning of all promoter luciferase and bHLH constructs
used in the transfection experiments of this study is described by Bogs et al.
(2007). For transient expression of VvMYB5b, the ORF was amplified by PCR
from Shiraz grape cDNA using PfuTurbo polymerase (Stratagene) and the
primers Myb5bartF (5#-ATGCTCGAGGGGACGAGAGAGAAAGAAGAAA-3#)
and Myb5bartR (5#-AGCTCTAGAATATCTCATTGCAGGGTGTTGA-3#). The
VvMYB5a ORF was amplified using the primers MYB5aartF (5#-TAGCTC-
GAGAAGCCAGAGGGATGAGAAATC-3#) and MYB5aartR (5#-TAGTCTA-
GATGATTCATCATTTTCAGGGAGA-3#). The generated PCR fragments
were purified, digested with XhoI and XbaI, and cloned in the vector pART7
(Gleave, 1992).
Sequence Analyses
Comparison and analysis of the VvMYB5b sequence were conducted with
the advanced BLAST program (Altschul et al., 1990) at the National Center for
Biotechnological Information (http//www.ncbi.nlm.nih.gov). Multiple se-
quence alignments were performed using the ClustalW (version 1.83.1)
program (Ramu et al., 2003) and the edition of aligned sequence was
performed using GeneDoc software (version 1.6). Phylogenic analyses were
achieved using MEGA package software (Kumar et al., 2004; version 3.2.1).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. Quantitative RT-PCR analysis of the relative
transcript levels of the VvMYB5a, VvMYBPA1, and VvMYBA genes in
Shiraz berries over complete berry development.
ACKNOWLEDGMENTS
We are grateful to Dr. Cathie Martin and Dr. Asaph Aharoni for providing
some of the cDNA probes used in this work and to Dr. Laurence Geny for PA
analysis.
Received March 14, 2008; accepted May 28, 2008; published June 6, 2008.
LITERATURE CITED
Aharoni A, De Vos CH, Wein M, Sun Z, Greco R, Kroon A, Mol JN,
O’Connell AP (2001) The strawberry FaMYB1 transcription factor
suppresses anthocyanin and flavonol accumulation in transgenic to-
bacco. Plant J 28: 319–332
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local
alignment search tool. J Mol Biol 215: 403–410
Asif M, Dhawan P, Nath P (2000) A simple procedure for the isolation of
high quality RNA from ripening banana fruit. Plant Mol Biol Rep 18:
105–119
Bagchi D, Bagchi M, Stohs SJ, Das DK, Ray SD, Kuszynski CA, Joshi SS,
Pruess HG (2000) Free radicals and grape seed proanthocyanidin
extract: importance in human health and disease prevention. Toxicology
148: 187–197
Ban Y, Honda C, Hatsuyama Y, Igarashi M, Bessho H, Moriguchi T (2007)
Isolation and functional analysis of a MYB transcription factor gene that
is a key regulator for the development of red coloration in apple skin.
Plant Cell Physiol 48: 958–970
Baudry A, Caboche M, Lepiniec L (2006) TT8 controls its own expression
in a feedback regulation involving TTG1 and homologous MYB and
bHLH factors, allowing a strong and cell-specific accumulation of
flavonoids in Arabidopsis thaliana. Plant J 46: 768–779
Baudry A, Heim MA, Dubreucq B, Caboche M, Weisshaar B, Lepiniec L
(2004) TT2, TT8, and TTG1 synergistically specify the expression of
BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana.
Plant J 39: 366–380
Bogs J, Downey MO, Harvey JS, Ashton AR, Tanner GJ, Robinson SP
(2005) Proanthocyanidin synthesis and expression of genes encoding
leucoanthocyanidin reductase and anthocyanidin reductase in devel-
oping grape berries and grapevine leaves. Plant Physiol 139: 652–663
Bogs J, Jaffe FW, Takos AM, Walker AR, Robinson SP (2007) The
grapevine transcription factor VvMYBPA1 regulates proanthocyanidin
synthesis during fruit development. Plant Physiol 143: 1347–1361
Boss PK, Davies C, Robinson SP (1996a) Analysis of the expression of
anthocyanin pathway genes in developing Vitis vinifera L. cv Shiraz
grape berries and the implications for pathway regulation. Plant Physiol
111: 1059–1066
Boss PK, Davies C, Robinson SP (1996b) Expression of anthocyanin
biosynthesis pathway genes in red and white grapes. Plant Mol Biol
32: 565–569
Broun P (2005) Transcriptional control of flavonoid biosynthesis: a complex
network of conserved regulators involved in multiple aspects of differ-
entiation in Arabidopsis. Curr Opin Plant Biol 8: 272–279
Carey CC, Strahle JT, Selinger DA, Chandler VL (2004) Mutations in the
pale aleurone color1 regulatory gene of the Zea mays anthocyanin
pathway have distinct phenotypes relative to the functionally similar
TRANSPARENT TESTA GLABRA1 gene in Arabidopsis thaliana. Plant
Cell 16: 450–464
Davies C, Robinson SP (2000) Differential screening indicates a dramatic
change in mRNA profiles during grape berry ripening. Cloning and
characterization of cDNAs encoding putative cell wall and stress re-
sponse proteins. Plant Physiol 122: 803–812
Deluc L, Barrieu F, Marchive C, Lauvergeat V, Decendit A, Richard T,
Carde JP, Merillon JM, Hamdi S (2006) Characterization of a grapevine
R2R3-MYB transcription factor that regulates the phenylpropanoid
pathway. Plant Physiol 140: 499–511
Dixon RA, Lamb CJ, Masoud S, Sewalt VJ, Paiva NL (1996) Metabolic
engineering: prospects for crop improvement through the genetic ma-
nipulation of phenylpropanoid biosynthesis and defense responses—a
review. Gene 179: 61–71
Downey M, Harvey J, Robinson S (2003a) Analysis of tannins in seeds and
skins of Shiraz grapes throughout berry development. Aust J Grape
Wine Res 9: 15–27
Downey M, Harvey J, Robinson S (2003b) Synthesis of flavonols and
expression of flavonol synthase genes in developing grape berries
of Shiraz and Chardonnay (Vitis vinifera L.). Aust J Grape Wine Res 9:
110–121
Espley RV, Hellens RP, Putterill J, Stevenson DE, Kutty-Amma S, Allan
AC (2007) Red colouration in apple fruit is due to the activity of the MYB
transcription factor, MdMYB10. Plant J 49: 414–427
Gleave A (1992) A versatile binary vector system with a T-DNA organisa-
tional structure conducive to efficient integration of cloned DNA into
the plant genome. Plant Mol Biol 20: 1203–1207
Grotewold E, Sainz MB, Tagliani L, Hernandez JM, Bowen B, Chandler
VL (2000) Identification of the residues in the Myb domain of maize C1
that specify the interaction with the bHLH cofactor R. Proc Natl Acad
Sci USA 97: 13579–13584
Hahlbrock K, Scheel D (1989) Physiology and molecular biology of
phenylpropanoid metabolism. Annu Rev Plant Physiol Plant Mol Biol
40: 347–369
Hartmann U, Sagasser M, Mehrtens F, Stracke R, Weisshaar B (2005)
Differential combinatorial interactions of cis-acting elements recognized
by R2R3-MYB, BZIP, and BHLH factors control light-responsive and
Deluc et al.
2052 Plant Physiol. Vol. 147, 2008 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.

tissue-specific activation of phenylpropanoid biosynthesis genes. Plant
Mol Biol 57: 155–171
Hernandez JM, Feller A, Morohashi K, Frame K, Grotewold E (2007) The
basic helix loop helix domain of maize R links transcriptional regulation
and histone modifications by recruitment of an EMSY-related factor.
Proc Natl Acad Sci USA 104: 17222–17227
Hernandez JM, Heine GF, Irani NG, Feller A, Kim MG, Matulnik T,
Chandler VL, Grotewold E (2004) Different mechanisms participate in
the R-dependent activity of the R2R3 MYB transcription factor C1. J Biol
Chem 279: 48205–48213
Horsch RB, Klee HJ, Stachel S, Winans SC, Nester EW, Rogers SG, Fraley
RT (1986) Analysis of Agrobacterium tumefaciens virulence mutants in
leaf discs. Proc Natl Acad Sci USA 83: 2571–2575
Horstmann V, Huether C, Jost W, Reski R, Decker E (2004) Quantitative
promoter analysis in Physcomitrella patens: a set of plant vectors activating
gene expression within three orders of magnitude. BMC Biotechnol 4: 13
Jin H, Cominelli E, Bailey P, Parr A, Mehrtens F, Jones J, Tonelli C,
Weisshaar B, Martin C (2000) Transcriptional repression by AtMYB4
controls production of UV-protecting sunscreens in Arabidopsis. EMBO
J 19: 6150–6161
Kamitani Y, Maki K, Tofani I, Nishikawa Y, Tsukamoto K, Kimura M
(2004) Effects of grape seed proanthocyanidins extract on mandibles in
developing rats. Oral Dis 10: 27–31
Kennedy J, Troup G, Pilbrow J, Hutton D, Hewitt D, Hunter C, Ristic R,
Iland P, Jones G (2000) Development of seed polyphenols in berries
from Vitis Vinifera L. cv Shiraz. Aust J Grape Wine Res 6: 244–254
Klatsky AL (2002) Alcohol and cardiovascular diseases: a historical over-
view. Ann N Y Acad Sci 957: 7–15
Kobayashi S, Goto-Yamamoto N, Hirochika H (2004) Retrotransposon-
induced mutations in grape skin color. Science 304: 982
Kobayashi S, Ishimaru M, Hiraoka K, Honda C (2002) Myb-related genes
of the Kyoho grape (Vitis labruscana) regulate anthocyanin biosynthesis.
Planta 215: 924–933
Koes R, Verweij W, Quattrocchio F (2005) Flavonoids: a colorful model for
the regulation and evolution of biochemical pathways. Trends Plant Sci
10: 236–242
Koltunow AM, Truettner J, Cox KH, Wallroth M, Golberg RB (1990)
Different temporal and spatial gene expression patterns occur during
anther development. Plant Cell 2: 1201–1224
Kranz HD, Denekamp M, Greco R, Jin H, Leyva A, Meissner RC, Petroni
K, Urzainqui A, Bevan M, Martin C, et al (1998) Towards functional
characterisation of the members of the R2R3-MYB gene family from
Arabidopsis thaliana. Plant J 16: 263–276
Kumar S, Tamura K, Nei M (2004) MEGA3: integrated software for
molecular evolutionary genetics analysis and sequence alignment. Brief
Bioinform 5: 150–163
Kutchan TM (2005) A role for intra- and intercellular translocation in
natural product biosynthesis. Curr Opin Plant Biol 8: 292–300
Lee MM, Schiefelbein J (2001) Developmentally distinct MYB genes
encode functionally equivalent proteins in Arabidopsis. Development
128: 1539–1546
Lepiniec L, Debeaujon I, Routaboul JM, Baudry A, Pourcel L, Nesi N,
Caboche M (2006) Genetics and biochemistry of seed flavonoids. Annu
Rev Plant Biol 57: 405–430
Li SF, Santini JM, Nicolaou O, Parish RW (1996) A novel myb-related gene
from Arabidopsis thaliana. FEBS Lett 379: 117–121
Lin LC, Kuo YC, Chou CJ (2002) Immunomodulatory proanthocyanidins
from Ecdysanthera utilis. J Nat Prod 65: 505–508
Mathews H, Clendennen SK, Caldwell CG, Liu XL, Connors K, Matheis
N, Schuster DK, Menasco DJ, Wagoner W, Lightner J, et al (2003)
Activation tagging in tomato identifies a transcriptional regulator of
anthocyanin biosynthesis, modification, and transport. Plant Cell 15:
1689–1703
Morita Y, Saitoh M, Hoshino A, Nitasaka E, Iida S (2006) Isolation of
cDNAs for R2R3-MYB, bHLH and WDR transcriptional regulators and
identification of c and ca mutations conferring white flowers in the
Japanese morning glory. Plant Cell Physiol 47: 457–470
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassays with tobacco tissue culture. Physiol Plant 15: 473–497
Park K, Ishikawa N, Morita Y, Choi J, Hoshino A, Iida S (2007) A bHLH
regulatory gene in the common morning glory, Ipomoea purpurea, con-
trols anthocyanin biosynthesis in flowers, proanthocyanidin and phy-
tomelanin pigmentation in seeds and seed trichome formation. Plant J
49: 641–654
Porter LJ (1989) Tannins. In JB Harbourne, ed, Methods in Plant Biochem-
istry. Plant Phenolics, Vol 1. Academic Press, London, pp 389–419
Quattrocchio F, Verweij W, Kroon A, Spelt C, Mol J, Koes R (2006) PH4 of
petunia is an R2R3 MYB protein that activates vacuolar acidification
through interactions with basic-helix-loop-helix transcription factors of
the anthocyanin pathway. Plant Cell 18: 1274–1291
Ramsay NA, Glover BJ (2005) MYB-bHLH-WD40 protein complex and the
evolution of cellular diversity. Trends Plant Sci 10: 63–70
Ramsay NA, Walker AR, Mooney M, Gray JC (2003) Two basic-helix-loop-
helix genes (MYC-146 and GL3) from Arabidopsis can activate anthocy-
anin biosynthesis in a white-flowered Matthiola incana mutant. Plant
Mol Biol 52: 679–688
Ramu C, Sugawara H, Koike T, Lopez R, Gibson T, Higgins D, Thompson
J (2003) Multiple sequence alignment with the Clustal series of program.
Nucleic Acids Res 31: 3497–3500
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular Cloning: A Labora-
tory Manual, Ed 2. Cold Spring Harbor Laboratory Press, Cold Spring
Harbor, NY
Schwinn K, Venail J, Shang Y, Mackay S, Alm V, Butelli E, Oyama R,
Bailey P, Davies K, Martin C (2006) A small family of MYB-regulatory
genes controls floral pigmentation intensity and patterning in the genus
Antirrhinum. Plant Cell 18: 831–851
Spelt C, Quattrocchio F, Mol JN, Koes R (2000) anthocyanin1 of petunia
encodes a basic helix-loop-helix protein that directly activates tran-
scription of structural anthocyanin genes. Plant Cell 12: 1619–1632
Springob K, Nakajima J, Yamazaki M, Saito K (2003) Recent advances in
the biosynthesis and accumulation of anthocyanins. Nat Prod Rep 20:
288–303
Stracke R, Werber M, Weisshaar B (2001) The R2R3-MYB gene family in
Arabidopsis thaliana. Curr Opin Plant Biol 4: 447–456
Takos AM, Jaffe FW, Jacob SR, Bogs J, Robinson SP, Walker AR (2006)
Light-induced expression of a MYB gene regulates anthocyanin biosyn-
thesis in red apples. Plant Physiol 142: 1216–1232
Torregrossa L, Verries C, Tesniere C (2002) Grapevine (Vitis vinifera L.)
promoter analysis by biolistic-mediated transient transformation of cell
suspensions. Vitis 41: 27–32
Vannini C, Locatelli F, Bracale M, Magnani E, Marsoni M, Osnato M,
Mattana M, Baldoni E, Coraggio I (2004) Overexpression of the rice
Osmyb4 gene increases chilling and freezing tolerance of Arabidopsis
thaliana plants. Plant J 37: 115–127
Verwoerd T, Dekker B, Hoekema A (1989) A small scale procedure for the
rapid isolation of plant RNAs. Nucleic Acids Res 17: 2362
Walker A, Lee E, Bogs J, McDavid D, Thomas M, Robinson S (2007) White
grapes arose through the mutation of two similar and adjacent regula-
tory genes. Plant J 49: 772–785
Xie DY, Sharma SB, Paiva NL, Ferreira D, Dixon RA (2003) Role of
anthocyanidin reductase, encoded by BANYULS in plant flavonoid
biosynthesis. Science 299: 396–399
Grapevine MYBs Regulate the Flavonoid Pathway
Plant Physiol. Vol. 147, 2008 2053 www.plant.org on October 31, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2008 American Society of Plant Biologists. All rights reserved.






























