Embed Size (px)
Citation preview

Page 1
This article is protected by copyright. All rights reserved. 1
The Top 10 oomycete pathogens in molecular plant pathology 1 1
Sophien Kamoun1*, Oliver Furzer1, Jonathan D.G. Jones1, Howard S. Judelson2, Gul Shad Ali3, Ronaldo J. 2 D. Dalio4, Sanjoy Guha Roy5, Leonardo Schena6, Antonios Zambounis7, Franck Panabières8, David Cahill9, 3 Michelina Ruocco10, Andreia Figueiredo11, Xiao-Ren Chen12, Jon Hulvey13, Remco Stam14, Kurt Lamour15, 4 Mark Gijzen16 Brett M. Tyler17, Niklaus J. Grünwald18, M. Shahid Mukhtar19, 20, Daniel F. A. Tomé21, Mahmut 5 Tör22, Guido Van den Ackerveken23, John McDowell24, Fouad Daayf25, William E. Fry26, Hannele Lindqvist-6 Kreuze27, Harold J.G. Meijer28, Benjamin Petre1,29, Jean Ristaino30, Kentaro Yoshida1, Paul R.J. Birch14, and 7 Francine Govers28 8
1The Sainsbury Laboratory, Norwich Research Park, Norwich, NR4 7UH, UK. 9 2Department of Plant Pathology and Microbiology, University of California, Riverside, CA 92521 USA 10 3Department of Plant Pathology and MREC, IFAS, University of Florida, Apopka, FL 32703, USA 11 4Biotechnology lab, Centro de Citricultura Sylvio Moreira / Instituto Agronomico, Cordeirópolis-Sao Paulo, 12 Brazil 13 5Department of Botany, West Bengal State University, Barasat, Kolkata-700126, India 14 6Dipartimento di Gestione dei Sistemi Agrari e Forestali, Università degli Studi Mediterranea, 89122 Reggio 15 Calabria, Italy 16 7UMR1290 BIOGER-CPP, INRA-AgroParisTech, 78850 Thiverval-Grignon, France 17 8INRA, UMR1355, Univ. Nice Sophia Antipolis, CNRS, UMR 7254, ISA, F-06903 Sophia Antipolis, France 18 9Deakin University, Geelong, Victoria, 3217, Australia 19 10Portici Division of The Italian National Research Council (CNR) Institute for Sustainable Plant Protection 20 (IPSP) Via Università 133, 80055 Portici (NA) Italy 21 11Centre for Biodiversity, Functional and Integrative Genomics, Faculty of Sciences, University of Lisboa, 22 1749-016 Lisboa, Portugal. 23 12College of Horticulture and Plant Protection, Yangzhou University, China 24 13Stockbridge School of Agriculture, University of Massachusetts Amherst, USA 25 14Division of Plant Sciences, College of Life Sciences, University of Dundee (at James Hutton Institute), 26 Errol Road, Invergowrie DD2 5DA, UK 27 15Department of Entomology and Plant Pathology, University of Tennessee, USA 28 16Agriculture and Agri-Food Canada, 1391 Sandford Street, London, Ontario N5V 4T3, Canada 29 17Center for Genome Research and Biocomputing, and Department of Botany and Plant Pathology, Oregon 30 State University, Corvallis, OR 97331, USA. 31 18USDA ARS, Plant Pathology Horticultural Crops Research Lab. 3420 NW Orchard Ave., Corvallis, Oregon 32 97330, United States 33 19Department of Biology, University of Alabama at Birmingham, Birmingham, AL 35294-1170, USA 34 20Nutrition Obesity Research Center, University of Alabama at Birmingham, Birmingham, 35294, AL, USA 35 21School of Life Sciences, University of Warwick, Coventry, CV4 7AL, UK 36 22National Pollen and Aerobiology Research Unit, The University of Worcester, Henwick Grove, Worcester 37 WR2 6 AJ, UK 38 23Plant- Microbe Interactions, Department of Biology, Utrecht University, Padualaan 8, 3584 CH Utrecht, 39 Netherlands 40 24Department of Plant Pathology, Physiology and Weed Science, Virginia Tech, Blacksburg, VA 24061, USA 41 25Department of Plant Science, University of Manitoba, Winnipeg, MB, Canada 42 26Department of Plant Pathology and Plant-Microbe Biology, Cornell University, Ithaca, New York 14853, 43 USA 44
This article has been accepted for publication and undergone full peer review but has not
been through the copyediting, typesetting, pagination and proofreading process, which
may lead to differences between this version and the Version of Record. Please cite this
article as doi: 10.1111/mpp.12190 Acc
epte
d A
rticl
e

Page 2
This article is protected by copyright. All rights reserved. 2
27International Potato Center, Apartado 1558, Lima 12, Peru 45 28Laboratory of Phytopathology, Wageningen University, NL-1-6708 PB Wageningen, Netherlands 46 29INRA, UMR 1136 Interactions Arbres/Microorganismes, Centre INRA Nancy Lorraine, 54280 47 Champenoux, France 48 30Department of Plant Pathology, North Carolina State University, Raleigh, North Carolina 27695, USA 49 50 * Corresponding author E-mail: [email protected] 51
Running header: Top 10 oomycete plant pathogens 52 53 SUMMARY 54 55 Oomycetes form a deep lineage of eukaryotic organisms that includes a large number of plant pathogens 56 that threaten natural and managed ecosystems. We undertook a survey to query the community for their 57 ranking of plant pathogenic oomycete species based on scientific and economic importance. In total, we 58 received 263 votes from 62 scientists in 15 countries for a total of 33 species. The Top 10 species and their 59 ranking are: (1) Phytophthora infestans; (2, tied) Hyaloperonospora arabidopsidis; (2, tied) Phytophthora 60 ramorum; (4) Phytophthora sojae; (5) Phytophthora capsici; (6) Plasmopara viticola; (7) Phytophthora 61 cinnamomi; (8, tied) Phytophthora parasitica; (8, tied) Pythium ultimum; and (10) Albugo candida. The article 62 provides an introduction to these 10 taxa and a snapshot of current research. We hope that the list will serve 63 as a benchmark for future trends in oomycete research. 64 65 66 INTRODUCTION 67 68 Oomycetes are eukaryotic organisms that superficially resemble filamentous fungi but are phylogenetically 69 related to diatoms and brown algae in the stramenopiles (Gunderson et al., 1987; Jiang and Tyler, 2012; 70 Lamour and Kamoun, 2009; Thines, 2014; Thines and Kamoun, 2010). Fossil evidence indicates that a 71 number of oomycetes emerged as endophytes of land plants at least by the Carboniferous period ~300-350 72 million years ago (Krings et al., 2011). One species, Combresomyces williamsonii, described from ~320 73 million year-old petrified stem cortex and rootlets of a seed fern may have even been parasitic (Strullu-74 Derrien et al., 2011). Phylogenetic analyses of modern taxa revealed that plant parasitism has evolved 75 independently in three lineages of oomycetes (Thines and Kamoun, 2010). Well known plant pathogens, 76 namely downy mildews, Phytophthora and Pythium, appear to have radiated from a common plant parasitic 77 ancestor (Thines and Kamoun, 2010). The impact of oomycetes on mankind is well documented both as 78 persistent threats to subsistence and commercial farming, and as destructive pathogens of native plants 79 (Agrios, 2005; Erwin and Ribeiro, 1996; Lamour and Kamoun, 2009). As a result, news related to plant 80 diseases caused by oomycetes tends to capture the interest of the general public and is frequently featured 81 in the media. 82
In the last two decades, increased awareness of the distinctive phylogeny and biology of 83 oomycetes drove the emergence of a specialist research community that is currently organized under the 84 umbrella of the “Oomycete Molecular Genetics Network”. This community has moved the field beyond the 85 gloomy view of the 1980s that oomycetes are a ‘fungal geneticist’s nightmare’ (Shaw, 1983; discussed in 86 Schornack et al, 2009). It produced novel paradigms in understanding host-microbe interactions, effector 87 biology and genome evolution (Bozkurt et al., 2012; Govers and Gijzen, 2006; Jiang and Tyler, 2012; 88 Schornack et al., 2009; Vleeshouwers et al., 2011). The oomycete community was one of the first in plant 89 pathology to initiate coordinated transcriptome and genome sequencing projects, and subsequently to 90 exploit the resulting resources to drive conceptual advances (Schornack et al., 2009; Govers and Gijzen, 91 2006; Pais et al., 2013). These days, with the genomes serving as unique resources for basic and applied 92 research, oomycetes are best portrayed as a ‘genomicist’s dream’ (Jiang and Tyler, 2012; Schornack et al., 93 2009; Govers and Gijzen, 2006; Pais et al., 2013). 94 A
ccep
ted
Arti
cle

Page 3
This article is protected by copyright. All rights reserved. 3
It is, therefore, particularly fitting that the oomycetes are at last covered by the Top 10 review 95 series of Molecular Plant Pathology. The process to generate the list and the aim of this article are similar to 96 previous contributions on plant pathogenic viruses (Scholthof et al., 2011), fungi (Dean et al., 2012), 97 bacteria (Mansfield et al., 2012) and nematodes (Jones et al., 2013). We undertook a survey to query the 98 community for their ranking of plant pathogenic oomycete taxa based on scientific and economic 99 importance. In total, we received 263 votes from 62 scientists in 15 countries that yielded the Top 10 100 species (Table 1). To some degree the results reflect the sizes of the sub-communities. Six of the ten 101 species belong to the genus Phytophthora, which is more commonly studied than any other oomycete 102 genus. Obligate parasites are also well represented with three species (Hyaloperonospora arabidopsidis, 103 Plasmopara viticola, and Albugo candida, Table 1). Another 23 species received votes but ranked outside 104 the Top 10 (Table 2). The fish parasite Saprolegnia parasitica received enough votes to place it in the Top 105 10 but was removed given that the list focuses on plant pathogens (Table 2). 106 The article is based on contributions from the voters to provide an introduction to the 10 taxa and a 107 snapshot of current research. Each section starts with a brief overview of the selected species, e.g. its 108 importance, pathology, host range, and life cycle. This is followed by a review of current research themes, 109 particularly the unique findings that emerged from studying the particular species, and an outlook on future 110 research. We hope that this article will serve as a reference and resource for novice and experienced 111 readers alike, as well as provide a benchmark for future trends in oomycete research. 112 113 114 1. PHYTOPHTHORA INFESTANS 115 116 Phytophthora infestans causes potato late blight (Fig. 1), a disease with major historical impact. Providing 117 twice the calories of rye and wheat per hectare, potato was central to European agriculture between 1750 118 and 1850. In 1844 the arrival of P. infestans changed this situation. In addition to the well-documented Irish 119 famine (Fig. 2), crop failures in 1845 and 1846 contributed to an estimated 750,000 hunger-associated 120 deaths in Continental Europe (Zadoks, 2008). The discovery that late blight was caused by a microbial 121 pathogen, fifteen years before Pasteur’s formal confirmation of Germ Theory, places the 19th century 122 migration of P. infestans as a significant milestone in the foundation of plant pathology as a scientific 123 discipline. Today, late blight remains a major constraint to the production of potato, the world’s third largest 124 staple crop, and is thus a constant threat to food security (Fisher et al., 2012; Haverkort et al., 2008). 125 In the 1840s, trade in potatoes likely facilitated long-distance migration of the pathogen. Whole-126 genome sequencing of European herbarium samples has revealed populations belonging to the HERB-1 127 genotype that were different from US-1, the genotype dominating the population globally in the mid-to-late 128 20th century (Goodwin et al., 1994; Martin et al., 2013; Martin et al., 2014; Yoshida et al., 2013; Yoshida et 129 al., 2014). Waves of migration and genotype displacements in the 20th century have been well documented. 130 The introduction of the A2 mating type to Europe in the 1970s (Drenth et al., 1994), and more recently, the 131 emergence of new, more aggressive lineages such as 13_A2 (Blue 13) in Europe (Cooke et al., 2012), US-132 22 in Eastern United States (Fry et al., 2013; Hu et al., 2012), and US-23/US-24 in Canada (Peters et al., 133 2014) impose constant challenges to disease resistance breeding. 134
In the 1990s, P. infestans research led the development of molecular approaches to study 135 oomycete pathology and biology. The first DNA transformation system (Judelson et al., 1991) and the first 136 transcriptional profiling during infection (Pieterse et al., 1991) paved the way to detailed molecular 137 investigations of sexual and asexual development and pathogenicity (reviewed in Judelson et al., 1997). 138 DNA fingerprinting revolutionised population studies (Goodwin et al., 1992) and was a prelude to the first 139 genome-wide genetic map of a Phytophthora species (van der Lee et al., 1997). Observations that gene 140 silencing occurred in P. infestans transformants (e.g. Judelson and Whittacker, 1995) sparked important 141 studies of the mechanisms underlying transcriptional silencing (van West et al., 1999). The first large-scale 142 transcript sequencing studies (Kamoun et al., 1999) ushered in the genomics era of Phytophthora research. 143
In the last 15 years, advanced genomic and functional approaches put P. infestans at the forefront 144 of research to understand oomycete development (Judelson and Blanco, 2005) and pathogenicity (Kamoun, 145 A
ccep
ted
Arti
cle

Page 4
This article is protected by copyright. All rights reserved. 4
2006). An elegant combination of bioinformatics, proteomics and functional genomics revealed secreted 146 proteins as potential factors influencing host-P. infestans interactions (Torto et al., 2003). Apoplastic 147 effectors that inhibit host secreted proteases were first described in P. infestans (Tian et al., 2004) and the 148 apoplastic phytotoxin-like SCR74 (Liu et al., 2005) provided support for the use of diversifying selection as a 149 criterion for identifying candidate effectors. The apoplastic effectors EPIC1 and EPIC2B were found to 150 converge on the host protease RCR3 that is also targeted by an independently evolved effector from a 151 fungal plant pathogen (Song et al., 2009). Recently, diversifying selection of the EPIC1 protease inhibitor 152 was implicated in functional adaptation to the equivalent protease target after ‘jumps’ to a new host (Dong et 153 al., 2014). 154
The first reported P. infestans avirulence effector was AVR3a, the counterpart of resistance protein 155 R3a (Armstrong et al., 2005). Comparison with avirulence effectors from Phytophthora sojae and H. 156 arabidopsidis identified conserved amino acid motifs RXLR and EER (Rehmany et al., 2005), which act as 157 signals for translocation into host cells (Whisson et al., 2007). The genome sequence of P. infestans 158 revealed hundreds of genes predicted to encode RXLR effectors plus a second class of candidate effectors 159 called Crinklers (CRNs) (Haas et al., 2009), which may also be delivered inside plant cells (Schornack et al., 160 2010). These effector genes are rapidly evolving and are typically variable between different P. infestans 161 isolates (Cooke et al., 2012). RXLR and CRN reside in gene-sparse, repeat-rich regions, potentially subject 162 to rearrangement and rapid mutation, raising the concept of a ‘two-speed’ genome in evolutionary terms 163 (Haas et al., 2009; Raffaele et al., 2010). Recent years have witnessed intense efforts to determine the 164 biochemical function of RXLR effectors, notably by determining their protein targets in the host. The first 165 target identified was ubiquitin E3 ligase CMPG1, which AVR3a stabilizes to prevent programmed cell death 166 in response to elicitors such as INF1 (Bos et al., 2010). 167
Breeding for late blight resistance was initiated by James Torbitt of Belfast in the 1870s, inspired by 168 the theories of Charles Darwin, with whom he shared considerable correspondence (DeArce, 2008). More 169 than 130 years later, breeding efforts have yet to provide a durable solution. Occasionally, local successes 170 are reported; for example variety C88, widely cultivated in China for over a decade, retains its resistance (Li 171 et al., 2011). Many resistance (R) genes effective against races of P. infestans have been identified 172 (Rodewald and Trognitz, 2013). Only with the recent identification of avirulence genes, all of which encode 173 RXLR effectors, can breeders incorporate knowledge of how P. infestans evades detection into their 174 breeding strategies (Vleeshouwers et al., 2011). Such information is critical to predicting the durability of R 175 genes. Using effectors as tools to rapidly identify R genes is a powerful step towards rational selection and 176 combination of lasting resistance (Vleeshouwers et al., 2008). 177
We still know relatively little about how P. infestans effectors manipulate host plants to establish 178 disease. How do they translocate into host cells? Do they define host range? How do they work in concert to 179 modulate complex networks of regulatory processes occurring in the host? Only recently have researchers 180 started to adopt structural biology to fully investigate functional relationships between interacting pathogen 181 and plant proteins (Wirthmueller et al., 2013). Could such research catalyse the modification of molecular 182 interactions in favor of the plant immune system through synthetic biology approaches? 183
The basic knowledge gained for P. infestans in the last decade is starting to impact management of 184 the late blight disease. For controlling late blight, farmers largely rely on agrochemicals, many with unknown 185 modes-of-action. Emergence of insensitive strains is possible due to high mutation rates in the pathogen 186 (Randall et al., 2014). The phenylamide Metalaxyl was one of the first chemicals that exhibited specificity to 187 oomycetes. Soon after its introduction and widespread use, fully insensitive strains emerged, but only 35 188 years later was the molecular basis of this insensitivity uncovered (Randall et al., 2014). Mining the P. 189 infestans genome has revealed many novel proteins with domain combinations that are unique for 190 oomycetes (Seidl et al., 2011). Several likely function in signalling might be promising fungicide targets. 191 Mode-of-action studies are more accessible thanks to the available genomics resources, transformation 192 tools and marker strains with tags to visualize the cytoskeleton and various subcellular compartments (Ah-193 Fong and Judelson, 2011; Meijer et al., 2014). In resistance breeding the efficacy of stacking R genes into 194 favored cultivars, either by introgression or trans- or cisgenesis, is being investigated (Tan et al., 2010). 195 However, for success, the R genes to be combined should be carefully selected based on our knowledge of 196 A
ccep
ted
Arti
cle

Page 5
This article is protected by copyright. All rights reserved. 5
variation in the pathogen population, and the attendant vulnerability to resistance being overcome 197 (Vleeshouwers et al., 2011). Future efforts should consider introducing multiple barriers, perhaps combining 198 R proteins with, for instance, membrane-localized pattern recognition receptors or components governing 199 nonhost resistance to P. infestans. Even then it is unlikely that plant resistance can fully control late blight. 200 Agrochemicals should not be abandoned. On the contrary, for more rational fungicide design, novel targets 201 should be identified, an endeavour that will be enhanced by an even more profound insight into the biology 202 of P. infestans. 203 204 2. HYALOPERONOSPORA ARABIDOPSIDIS 205 206 H. arabidopsidis (formerly H. parasitica and Peronospora parasitica), is one of 700 downy mildew species 207 within the Peronosporaceae (Thines, 2014). Downy mildew pathogens cause harmful diseases on many 208 important crops, notably Peronospora and Hyaloperonospora spp. on brassica crops, P. viticola on grape, 209 Peronosclerospora spp on maize and sorghum, Pseudoperonospora cubensis on cucurbits, and Bremia 210 lactucae on lettuce (Lucas et al., 1995). 211
Most downy mildews have narrow host ranges and are completely dependent on their host for 212 growth and reproduction. They can survive in the soil as quiescent oospores that initiate infection through 213 roots. Spread of the pathogen mostly occurs through airborne sporangiospores that are formed on the lower 214 leaf surface, giving downy patches (Fig. 3). These spores germinate on the plant surface and penetrate by 215 forming appressoria (Koch and Slusarenko, 1990). Once past the epidermis, hyphae grow intercellularly 216 and, similar to Phytophthora spp. and other downy mildews, develop haustoria, specialized structures that 217 may function in feeding and suppression of host defense by targeted secretion of effectors (Fig. 4, Whisson 218 et al., 2007) 219
H. arabidopsidis is a prominent pathogen in natural populations of Arabidopsis thaliana (Holub, 220 2008; Coates and Beynon, 2010). As such, it was adopted in the 1980’s as one of two pathogens of 221 Arabidopsis, along with the bacterium Pseudomonas syringae (Koch and Slusarenko, 1990). The top 10 222 ranking of H. arabidopsidis reflects the subsequent success of the Arabidopsis- H. arabidopsidis 223 pathosystem. H. arabidopsidis was initially utilized as a “physiological probe” of the Arabidopsis immune 224 system (Holub et al., 1994). This research led to the cloning of the first plant disease resistance (R) genes 225 against an oomycete, better understanding of the evolutionary dynamics of R genes, definition of broadly 226 important immune system regulators, identification of downy mildew-resistant mutants, and genetic 227 definition of the complexity of the plant immune signalling network (reviewed in Slusarenko and Schlaich, 228 2003, Coates and Beynon, 2010, Lapin & Van den Ackerveken, 2013). On the pathogen side, research is 229 hampered by the lack of protocols for culturing and genetic transformation, established techniques with 230 other oomycetes like P. infestans. However, work in the early 2000s led to the development of genetic maps 231 and DNA libraries that enabled discovery of the first avirulence effector (Allen et al., 2004), and later to the 232 RXLR effector family (Rehmany et al 2005). 233
Genome sequencing of H. arabidopsidis isolate Emoy2, completed in 2010, unveiled 134 predicted 234 RXLR effectors and other components of the H. arabidopsidis secretome (Baxter et al., 2010). Notably, this 235 report also revealed important genomic signatures of obligate biotrophy that have evolved convergently in 236 other obligate oomycete and fungal lineages (reviewed in McDowell, 2011). Protein interaction assays 237 showed that H. arabidopsidis effectors target a highly interconnected host machinery helping to define a 238 representative plant-pathogen interaction network (Mukhtar et al., 2011). In addition, several high 239 throughput functional studies have investigated effectors’ subcellular localizations, suppression of immune 240 responses, molecular targets, and cognate immune receptors (Cabral et al., 2011, Cabral et al., 2012, 241 Caillaud et al., 2011, Fabro et al., 2011). 242
Future studies with the H. arabidopsidis experimental system will include a) direct or 243 Agrobacterium-mediated transformation for genetic manipulation required for the molecular analysis of 244 downy mildew pathogenicity; b) establishment of the temporal hierarchy of effectors during penetration, 245 colonization and sporulation which may serve as a blueprint for a better understanding of the molecular 246 basis of biotrophy; c) revealing the role of genetic recombination and epigenetics on the emergence of new 247 A
ccep
ted
Arti
cle

Page 6
This article is protected by copyright. All rights reserved. 6
effectors; d) development of tools to understand how plant-originated molecules regulate pathogen 248 response; and e) relevance of interspecies transfer of small RNAs. These investigations on H. arabidopsidis 249 will continue to provide new insights into the molecular mechanisms of downy mildew pathogenicity, and 250 contribute to comparative and functional analysis of (obligate) biotrophic oomycete and fungal pathogens. 251 252 3. PHYTOPHTHORA RAMORUM 253
Phytophthora ramorum is the most destructive disease of oaks worldwide and the cause of sudden oak 254 death, sudden larch death and ramorum blight (Werres et al., 2001; Grünwald et al., 2008; Rizzo et al., 255 2005; Brasier and Webber, 2010) (Fig. 5). The host range of P. ramorum is one of the widest of any 256 Phytophthora spp., and includes many species of hardwood trees and ornamentals. To date, four distinct 257 clonal lineages are recognized by various molecular markers and two mating types are observed (Van 258 Poucke et al., 2012; Grünwald et al., 2009). These lineages appear anciently diverged, yet have emerged 259 repeatedly in both North America and Europe (Grünwald et al., 2012; Goss et al., 2009a) (Fig. 6). Lineages 260 NA1, NA2 and EU1 are currently found in Canada and the US, while EU1 and EU2 exist in Europe (Goss et 261 al., 2009b, 2011; Van Poucke et al., 2012; Grünwald et al., 2012). These lineages differ in aggressiveness 262 in controlled assays, while field experiments have not been conducted given quarantine restrictions 263 (Grünwald et al., 2008; Elliott et al., 2011; Kasuga et al., 2012; Van Poucke et al., 2012; Hüberli et al., 264 2012). Unique ecological attributes such as a wide host range, survival of dry, hot Mediterranean summers, 265 combined with the ability to reproduce from chlamydospores, are thought to provide the basis for sudden 266 oak death in California (Garbelotto and Hayden, 2012). 267
The availability of the genome sequence just a few years after the identification of the pathogen 268 provided rapid and novel insights into the biology of this pathogen (Tyler et al., 2006). Like other 269 Phytophthora spp., P. ramorum has a large number of candidate effectors interacting with the plant host 270 including RXLR, Crinkler and the necrosis and ethylene-inducing peptide 1 (Nep1)-like protein (NLP) gene 271 families (Tyler et al., 2006; Goss et al., 2013). RXLR effectors diversify rapidly despite the clonality of this 272 organism, using mechanisms such as loss or gain of repeated domains, recombination or gene conversion 273 among paralogs, and selection on point mutations (Goss et al., 2013). Recent work focused on the 274 discovery of endogenous small RNAs and description of the silencing machinery (Fahlgren et al., 2013). P. 275 ramorum and two other Phytophthora spp. that were examined produce two primary, distinct 21- and 25-276 nucleotide small RNA classes including a novel microRNA family. Two argonaute classes and two dicer-like 277 proteins appear to be involved in each pathway (Fig. 7), but this remains to be formally tested. Epigenetic 278 mechanisms were recently implicated in phenotypic diversification, namely colony morphology, colony 279 senescence, and virulence on coast live oak and California bay laurel (Kasuga et al., 2012). 280
P. ramorum provides a unique opportunity for studying the evolution of a genome that has two 281 mating types yet appears to lack sexual reproduction in the known field populations. The epigenome, 282 transcriptome and proteome of P. ramorum remain a mystery. The apparent documented phenotypic 283 variation driven by host association might be epigenetic in nature and invites further study. Several basic 284 questions remain unanswered. Why does P. ramorum have such a high number of RXLR effectors given 285 that the genome is adapted to a wide host range? What are the genes conferring a wide host range 286 compared to P. infestans and P. sojae? Is there a center of origin? This center might be located in Asia 287 where the closest relative, P. lateralis has been found in an old growth Chamaecyparis forest (Brasier et al., 288 2010). Discovery of a center of origin might also reveal a host with which P. ramorum might have coevolved 289 providing for a mechanism of R gene discovery, a critical tool needed for managing sudden oak death. 290 Comparative genomics of related species like P. lateralis, P. syringae and P. hibernalis, amongst others will 291 provide novel insights into core effectors and genes under purifying selection in the clade, but diverged 292 among clades. 293 A
ccep
ted
Arti
cle

Page 7
This article is protected by copyright. All rights reserved. 7
4. PHYTOPHTHORA SOJAE 294 295 Root rot disease of soybean caused by Phytophthora sojae was first recognized in North America in the 296 1950s (Kaufmann & Gerdemann, 1958, Hildebrand, 1959). The alternative names P. megasperma var. 297 sojae and P. megasperma f. sp. glycinea were commonly used for this species in the past (Erwin & Ribeiro, 298 1996). Infection of soybean by P. sojae, which is a hemibiotroph, typically initiates below ground and 299 eventually produces spreading cankers that destroy root tissues and travel up the stem (Fig. 8). The 300 pathogen thrives in wet conditions and in compacted or heavy clay soils. Motile, water-borne zoospores are 301 released from sporangia and are attracted to soybean root exudates (Morris & Ward, 1992). P. sojae is 302 homothallic and creates abundant, thick-walled sexual oospores that are long-lived and provide a soil-borne 303 inoculum. P. sojae has a narrow host range and its economic damage is limited to soybean. It has been 304 suggested that P. sojae originated in North America as a pathogen of lupins as some 26 species of the 305 genus Lupinus are susceptible to infection (Erwin & Ribeiro, 1996). In laboratory tests, P. sojae can also be 306 made to infect lima bean (Phaseolus lunatus), string bean (Phaseolus vulgaris), and cranesbill (Geranium 307 carolinianum) (Hildebrand, 1959, Erwin & Ribeiro, 1996). P. sojae is among the most damaging disease 308 problems that confront soybean growers (Wrather & Koenning, 2006). Management of P. sojae in soybean 309 has primarily relied on breeding for resistance. 310
Research on P. sojae has focused on mechanisms of pathogen virulence, host resistance, and the 311 molecular basis of recognition between P. sojae and soybean (Tyler, 2007, Gijzen & Qutob, 2009, Dorrance 312 & Grünwald, 2009). P. sojae, along with P. ramorum, was the first oomycete to have its genome sequenced 313 (Tyler et al., 2006). Genomic information, including gene expression data and multiple genome sequences, 314 have driven rapid progress (Qutob et al., 2000, Torto-Alalibo et al., 2007, Wang et al., 2011). The genome of 315 P. sojae, and later other oomycetes (Haas et al., 2009), was found to be partitioned into stable regions rich 316 in housekeeping genes displaying extensive synteny with other oomycete genomes, interspersed with 317 dynamic, transposon-rich regions that contain rapidly evolving genes implicated in virulence (Tyler et al., 318 2006, Jiang & Tyler, 2012) (Fig. 9). Multiple large, rapidly diversifying families of genes with functions in 319 virulence were discovered (Tyler et al., 2006, Jiang & Tyler, 2012) including hydrolases, hydrolase inhibitor 320 proteins, toxin-like proteins such as the NLP family (Qutob et al., 2006), and two huge, diverse classes of 321 effector proteins (RXLR and Crinkler effectors) (Tyler et al., 2006, Jiang et al., 2008, Haas et al., 2009) that 322 can cross into the cytoplasm of host plant cells (Dou et al., 2008, Kale et al., 2010) (Fig. 9). All 11 avirulence 323 genes cloned from P. sojae proved to encode RXLR effectors detected by host R proteins (Jiang & Tyler, 324 2012) (Fig. 9). Genetic inheritance studies in P. sojae using molecular markers led to discoveries of high-325 frequency gene conversion (also referred to as loss of heterozygosity) (Chamnanpunt et al., 2001) and 326 epigenetic silencing (Qutob et al., 2013) as important mechanisms underlying pathogen variation. 327
Many areas of future research into P. sojae will be common to other oomycete and fungal 328 pathogens. One will be defining in detail how apoplastic and host cell-entering effectors of P. sojae 329 manipulate soybean physiology to enable infection, including the soybean molecules targeted by the 330 effectors (Fig. 9), and the mechanisms by which RXLR and Crinkler effectors gain entry (Fig. 9). Identifying 331 the P. sojae effectors most important for infection (e.g. using new tools for genetic manipulation such as 332 tailored nucleases) should lead to new targets for disease control. The cell biology of P. sojae infection 333 (Enkerli et al., 1997) must be defined in much more detail, incorporating the spatial distributions of pathogen 334 and host transcripts, proteins and metabolites. Understanding fully the role of epigenetic mechanisms in 335 generating variation in pathogen populations will also be important for designing effective strategies for 336 disease control. Sequencing the genomes and transcriptomes of numerous isolates of P. sojae and its 337 nearest sister species will shed light on the genetic adaptability of the pathogen. In practical terms, a large 338 translational research effort will be required to convert the rapidly accumulating knowledge about the basic 339 biology and pathology of P. sojae and other oomycetes into effective disease control solutions. 340 341 5. PHYTOPHTHORA CAPSICI 342 343 A
ccep
ted
Arti
cle

Page 8
This article is protected by copyright. All rights reserved. 8
Phytophthora capsici is a highly destructive invasive pathogen that attacks solanaceous (pepper, 344 tomato), legume (lima and snap beans) and most cucurbit hosts (Hausbeck & Lamour, 2004, Leonian, 345 1922). Disease is favored by warm (25 to 28 °C) and wet conditions and the asexual epidemiology is often 346 explosive (Granke et al., 2009). Initial infection is biotrophic followed by transitions within 24 to 48 hours to 347 necrotrophy and the production of deciduous sporangia on the surface of infected tissues (Fig. 10). The 348 importance of the sexual stage differs based on region. In Peru and Argentina and across much of China, 349 long-lived and widely dispersed clonal lineages dominate (Gobena et al., 2012, Hurtado-Gonzales et al., 350 2008, Sun et al., 2008, Hu et al., 2013). This is in contrast to the United States, South Africa and the 351 northern provinces of China where populations have high levels of genotypic diversity and outcrossing is 352 frequent (Gobena et al., 2012, Meitz et al., 2010, Dunn et al., 2010). Once introduced to a field site, P. 353 capsici is difficult to control and often impossible to eradicate. P. capsici grows fast and sporulates heavily 354 on simple media and isolates from sexual field populations are often fecund (Gobena et al., 2012, Lamour & 355 Hausbeck, 2000). P. capsici is one of the most genetically diverse eukaryotic organisms yet described and 356 there is significant genetic variation in the form of single nucleotide polymorphisms within individual 357 genomes and across world populations (Lamour et al., 2012). 358
Current research includes investigation of resistance (host and nonhost) in commercial vegetable 359 and experimental plants, discovery and characterization of genes and proteins driving pathogenicity and 360 virulence, and studies to measure the evolution of natural and laboratory populations. Under controlled 361 conditions, P. capsici infects at least 26 plant families (Granke et al., 2012). Natural resistance to P. capsici 362 in pepper and cucurbits appears to be rare (Mallard et al., 2013) but in tomato, it may be extensive 363 (Quesada-Ocampo & Hausbeck, 2010). Recent molecular studies of the CRN class of effectors indicate 364 they are often localized to the host nucleus and play an important role in the infection process (Chen et al., 365 2013, Stam et al., 2013a, Stam et al., 2013b). Transcriptome studies using RNAseq and microarrays 366 indicate dramatic changes from pre-infective spores through the early biotrophic and later necrotrophic 367 stages of infection (Chen et al., 2013, Jupe et al., 2013). Studies of individual effectors suggest some 368 manipulate the host to allow infection while others trigger plant cell death (Chen et al., 2013, Feng et al., 369 2010, Feng & Li, 2013, Sun et al., 2009). 370
A high quality reference genome and a high density SNP-based genetic linkage map were recently 371 completed (Lamour et al., 2012). These important resources illuminated an important source of asexual 372 genetic variation known as loss of heterozygosity (LOH) (Lamour et al., 2012). LOH occurs when variable 373 length tracts (300bp to >1Mbp) of the diploid genome switch to one of the two possible haplotypes. LOH has 374 been described in laboratory-produced sexual progeny, field isolates maintained on agar media, and field 375 isolates genotyped directly from naturally infected tissue (Hu et al., 2013, Gobena et al., 2010, Hulvey et al., 376 2010). LOH is associated with spontaneous switches from the A2 to the A1 mating type, loss of 377 pathogenicity and reduced virulence (Hu et al., 2013, Lamour et al., 2012). 378
The diversity and plasticity of P. capsici present challenges and unique opportunities for research 379 (Fig. 10). Future research questions include: What is the significance of a highly diverse effector arsenal? 380 Are these effectors important for host-specific interactions and the broad host range? How important is LOH 381 in laboratory and field populations? The genomic plasticity of P. capsici and the ease of laboratory 382 manipulation provide a unique opportunity to measure genome stability and adaptive evolution at a fine 383 scale and may provide useful insights into this and other oomycete pathogens (Lamour & Hu, 2013). 384 385 6. PLASMOPARA VITICOLA 386 387 The obligate biotrophic oomycete P. viticola, causal agent of grape downy mildew, is native to North 388 America and was inadvertently introduced into Europe at the end of the 19th century (Viennot-Bourgin, 1949, 389 Millardet, 1881) causing severe damage to Vitis vinifera, which had evolved in the absence of this pathogen 390 (Galet, 1977, Gessler et al., 2011). Plasmopara viticola belongs to the family Peronosporaceae; its life cycle 391 includes an asexual multiplication phase occurring during the plant vegetative period and a sexual phase 392 that ensures pathogen overwinter survival (Wong et al., 2001). The pathogen overwinters as oospores in 393 dead leaf lesions or as mycelium in infected twigs. In spring, and in particular during rainy periods, oospores 394 A
ccep
ted
Arti
cle

Page 9
This article is protected by copyright. All rights reserved. 9
germinate to produce sporangia, which are transported by wind or water to the wet leaves near the ground, 395 which they infect through stomata on the lower surface. The mycelium then spreads into the intercellular 396 spaces of the leaf from which sporangiophores arise and emerge through the stoma, ready to start new 397 infection cycles (Gobbin et al., 2005). Downy mildew affects leaves, fruit, and shoots, with young tissues 398 being particularly susceptible to infection (Fig. 11) (Kennelly et al., 2007). A disease cycle may take from 5 399 to 18 days, depending on the temperature, humidity, and varietal susceptibility (Agrios, 2005, Gessler et al., 400 2011). 401
Downy mildew is still most destructive in Europe and in the eastern half of the United States, where 402 it may cause severe epidemics year after year (Agrios, 2005; Madden et al.,1995). When the weather is 403 favorable and no protection against the disease is provided, downy mildew can easily destroy up to 75% of 404 production in a single season (Madden et al., 2000, Rossi & Caffi, 2012). Occasionally, P. viticola can be 405 destructive in other humid parts of the world where V. vinifera is cultivated. 406 In the last fifty years, research on grapevine downy mildew has focused on the pathogen life cycle 407 and epidemiology (reviewed in Gessler et al., 2011), and on the genetic identification of host resistance loci 408 for the establishment of molecular markers (revised in Töpfer et al., 2011). In the last decade, studies on P. 409 viticola have highlighted its enormous potential in developing fungicide resistance (Chen et al., 2007, Blum 410 et al., 2010) and breaking down plant resistance of interspecific hybrids such as ‘Bianca’ (Peressotti et al., 411 2010, Casagrande et al., 2011) and ‘Regent’ (Delmotte et al., 2013). Population genetic studies have been 412 carried out (Fontaine et al., 2013 and references therein) to assess pathogen dynamics and diversity in 413 Europe. Moreover, it was proposed that grapevine downy mildew is not caused by a single species, but 414 instead by a complex of cryptic species that have diverged on Vitaceae (Rouxel et al., 2013). Trade-offs 415 between the size and the number of sporangia produced were reported, leading to ecological advantages 416 for this pathogen (Delmotte et al., 2013). For these reasons the Vitis - P. viticola pathosystem was elected 417 as a prime candidate for studying host specialization of biotrophic plant pathogens (Rouxel et al., 2013) and 418 pathogen adaptation to partial host resistance (Delmotte et al., 2013). 419
Control methods against P. viticola are essentially based on the use of chemical fungicides and 420 application of disease models that serve as decision support systems (Rossi et al., 2013, Vercesi et al., 421 2010, Rossi et al., 2009, Lalancette et al., 1987). Due to the growing public concern on the use of chemical 422 pesticides, and the necessity to adhere to the European Directive 2009/128/EC encompassing a framework 423 “to achieve a sustainable use of pesticides” by promoting Integrated Pest Management (IPM), finding 424 alternative methods for the control of P. viticola on grapevine appears urgent. Several alternative 425 approaches have been proposed in the last 20 years, but none has been transferred to practical use 426 (Gessler et al., 2011). Moreover, the lack of precise characterization of P. viticola isolates is dramatically 427 limiting reliable control of the pathogen. Although molecular studies have previously confirmed a high 428 diversity in the pathogen population, there is a surprising lack of phenotypic characterization of pathotype 429 strains or races which could be used to study the mechanisms of interaction with host genotypes with 430 different levels of resistance (Gómez-Zeledón et al., 2013). In conclusion, interdisciplinary studies and 431 consistent resources should be invested for the study of the P. viticola-V. vinifera pathosystem in order to 432 find new, effective alternatives for the IPM of this highly adaptive and destructive pathogen. 433 434 7. PHYTOPHTHORA CINNAMOMI 435 436 Some call it the ‘biological bulldozer’ for its capacity to destroy natural plant communities across the globe, 437 and disease caused by P. cinnamomi has broad and economically important impacts in forestry and 438 horticulture and in the nursery industry. Like other Phytophthora spp., P. cinnamomi has a number of 439 strategies for survival, propagation and dissemination. The motile zoospore is recognised as the main 440 infective propagule following encystment and attachment to roots and stems (Fig. 12). Recent work 441 suggests that in addition to chlamydospores, P. cinnamomi is also associated with lignituber formation that 442 enables survival under harsh conditions (Jung et al. 2013). Both A1 and A2 mating types are pathogenic but 443 wide differences in distribution and frequency of occurrence suggests the A2 mating type to be more 444 invasive and it is generally recognised as being the more aggressive of the two mating types. Where 445 A
ccep
ted
Arti
cle

Page 10
This article is protected by copyright. All rights reserved. 10
introduced, this pathogen has had enormous impacts on natural systems including those of Australia, 446 southern Europe and the United States. In Australia, for example, it has been estimated that >3000 largely 447 endemic plant species from numerous plant families are under threat and a new National Threat Abatement 448 Plan has been urgently implemented (Australian Government Department of Environment, 2014) (Fig. 13). 449 Control of disease especially across the large areas of native vegetation affected is still a great problem but 450 some success has been achieved by the use of phosphite in both natural systems and in agriculture 451 (Akinsanmi and Drenth 2013; Crane and Shearer 2013), by the use of calcium amendments to soil (Serrano 452 et al. 2012), and by containment and eradication of spot infections (Dunstan et al. 2010). Climate change is 453 predicted to have a significant influence on the intensity and distribution of disease (Thompson et al. 2014). 454
P. cinnamomi is a challenging organism to work with. Sequencing of its genome has recently been 455 completed (JGI; http://genome.jgi-psf.org/) and has opened up a variety of opportunities to investigate the 456 critical features that enable this pathogen to be so wide-ranging in its hosts. A real change in emphasis in 457 research on P. cinnamomi has occurred in recent times with an increasing number of groups using genomic, 458 transcriptomic and proteomic approaches to understand the pathogen and/or its interaction with the host (for 459 example Reeksting et al. 2014). Experimental plants, including Arabidopsis, Zea mays and Eucalyptus, for 460 which comprehensive genomic information is available, are increasingly used as host-pathogen platforms 461 (for example Allardyce et al. 2013; Dempsey 2012; Rookes et al. 2008). Fundamental studies on P. 462 cinnamomi plant cell wall degrading enzymes and the genes that encode them are being undertaken (Hee 463 et al. 2013; Adrienne Hardham, Australian National University, pers. comm.) as well as examination, at the 464 molecular level, of defense pathways and their control (Eshraghi et al. 2014; Gunning et al. 2013). 465
High throughput, next generation genome sequencing is emerging as an exciting way to examine 466 the pathogen, its hosts and the soil environment and is now being used to examine pathogen ecology and 467 diversity within soils and for analysis of the response of host plants (Treena Burgess, Murdoch University, 468 Australia pers. comm.). Much of our progress in understanding the host-pathogen interactions will come 469 from such approaches. We still don’t know enough about why plants are resistant to this pathogen or what 470 the basis for induced resistance is, for example, following phosphite treatment. The search for host 471 resistance to P. cinnamomi is an active area of research in which signalling pathways and their control are 472 being elucidated. Mapping the disease over the large invaded areas is still not straight forward although 473 advances in remote sensing, high resolution digital photography, hyperspectral imaging (David Guest, 474 Sydney University, Australia pers. comm.) and ‘drone’ technology will likely see our ability in this area to be 475 greatly enhanced in the near future. The era of nanotechnology offers new opportunities for the delivery of 476 molecules that can influence disease outcome yet this area has not been explored to date although 477 preliminary research with various nanoparticle systems (Nadiminti et al. 2013; Hussain et al. 2014) indicates 478 that these may be useful in delivering molecules, including pathogen effectors, DNA and miRNA that may 479 modify host processes and their response to pathogen infection. 480
Future research will focus on the following questions: 1) What are the virulence factors that enable 481 P. cinnamomi to infect and colonise susceptible species and therefore be targeted for control? 2) What 482 constitutes host resistance and how can it be manipulated? Can we look to nanotechnology for some 483 answers? 3) Is phosphite the only chemical that we can use to control P. cinnamomi and how does 484 phosphite alter the host response? 485 486 8. PHYTOPHTHORA PARASITICA 487 488 Phytophthora parasitica (= P. nicotianae) is a worldwide distributed pathogen (Erwin & Ribeiro, 1996). 489 Primarily known to cause tobacco black shank and citrus root rot and gummosis (Fig. 14), it is also 490 responsible for severe foliar and fruit diseases as well as root and crown rots on herbaceous and perennial 491 plant species in more than 250 genera, including solanaceous crops (Fig. 15), and horticultural and fruit 492 trees (Cline et al., 2008). P. parasitica produces both asexual zoospores and thick-walled sexual oospores. 493 Oospores constitute a potential source of genetic variation, and with resting chlamydospores contribute to 494 survival in unfavorable conditions in soil or within infected plant tissues. 495 A
ccep
ted
Arti
cle

Page 11
This article is protected by copyright. All rights reserved. 11
Contrasting the broad host range observed at the species level, most individual P. parasitica 496 isolates display host preference (Erwin & Ribeiro, 1996). Frequent cases of differential virulence on a range 497 of hosts have been revealed (Colas et al., 1998; Matheron & Matejka, 1990), pointing out the need to 498 decipher the genetic structure of P. parasitica on a global scale. In agreement with pathogenicity tests, 499 recent single nucleotide polymorphism (SNPs) analyses conducted with mitochondrial and nuclear genes 500 revealed a specific association between host of origin and genetic grouping that was particularly evident for 501 tobacco and citrus isolates (Mammella et al., 2011; Mammella et al., 2013). In contrast, no clear genetic 502 structure was revealed for isolates from other hosts, especially potted ornamentals in nurseries. A significant 503 geographic structuring was revealed for tobacco but not for citrus isolates (Bonnet et al., 1994; Colas et al., 504 2001; Mammella et al., 2013). Further studies relying on whole-genome sequencing programs (see below) 505 are necessary to determine if these molecular groups represent evidence of physiological races, pathotypes 506 or even subspecies within P. parasitica. 507
Since some isolates may also infect many hosts, including Arabidopsis thaliana (Attard et al., 2008; 508 Attard et al., 2010), P. parasitica emerges as an ideal pathogen to develop specific studies aiming to 509 advance our knowledge on mechanisms underlying general pathogenicity and those governing host 510 specificity. Furthermore, P. parasitica is phylogenetically related to P. infestans in Phytophthora clade 1, and 511 overlaps its host range, which includes potato (Taylor et al., 2008). Comparative analyses on these two 512 species varying in genome size (83 and 240 Mb for P. parasitica and P. infestans, respectively) will facilitate 513 the understanding of the evolution behind pathogenicity and host range among Phytophthora spp. Through 514 the sequencing of the genome of isolates of diverse host range and geographical origins, the international 515 "Phytophthora parasitica genome initiative" project will enable the characterization of genes that determine 516 host range. In-depth genomic and transcriptomic analyses of 14 sequenced genomes are currently being 517 performed. Special efforts are devoted to characterize the repertoire of effector proteins. As a broad host 518 range pathogen, P. parasitica provides a unique opportunity for intra- and interspecific comparative 519 analyses, looking for the presence of various effector families, their organization, their role in plant 520 recognition and infection, and their evolution among strains and species that display broad or restricted host 521 ranges. The identification of conserved effector groups, as well as of other pathogenicity genes, will allow 522 evaluation of the evolutionary pressures of exposure to different host-defense responses to the 523 diversification of effectors and their role in adaptation to host plants. 524
P. parasitica has not been as heavily studied as its economic importance would warrant. However, 525 it is likely to gain importance in the foreseeable future due the following reasons. First, it is prominent in 526 nurseries of potted ornamentals and fruit tree species, the trade of which seems to represent one of the 527 most efficient dissemination pathways of Phytophthora (Moralejo et al., 2009, Olson & Benson, 2011). This 528 makes P. parasitica an ideal species to study diffusion pathways of Phytophthora and other soilborne 529 pathogens on a global scale. For aesthetic reasons, the ornamental industry requires extensive use of anti-530 oomycete chemicals (Olson et al., 2013). The global nursery trade thus increases the risk of the rapid 531 spread of resistant P. parasitica strains to new areas following an inappropriate use of fungicides and 532 constitutes a growing threat to local agriculture and natural ecosystems (Brasier, 2008). In addition, P. 533 parasitica will likely benefit from the warming climate. Its host range generally includes those of other 534 species of prime economic importance (P. infestans, P. capsici, P. citrophthora) but generally requires 535 warmer conditions than these potential competitors (Erwin & Ribeiro, 1996). Consequently, the foreseen 536 global warming is likely to provide a catalyst for the geographical expansion of this species, as has already 537 been proposed under Mediterranean climates (Andres et al., 2003; Saadoun & Allagui, 2008) and Eastern 538 India (Guha Roy et al., 2009). 539 540 9. PYTHIUM ULTIMUM 541 542 Pythium resides in the peronosporalean lineage of oomycetes, with Phytophthora and downy mildews (Dick, 543 2001). One of its most significant members is Py. ultimum, which causes damping-off and root rot on >300 544 diverse hosts including corn, soybean, wheat, Douglas fir, and ornamentals (Farr and Rossman, 2014). Py. 545 ultimum is a common inhabitant of fields, ponds and streams, and decomposing vegetation worldwide. 546 A
ccep
ted
Arti
cle

Page 12
This article is protected by copyright. All rights reserved. 12
Contributing to the species' ubiquity is its ability to grow saprotrophically in soil and plant residue, a trait 547 shared by most Pythium spp. but not other peronosporaleans, which must colonize living hosts. Py. ultimum 548 is also homothallic, producing oospores capable of long-term survival (Martin and Loper, 1999). Mycelia 549 and oospores in soil thus can initiate infections of seeds or roots leading to wilting, reduced yield, and 550 mortality (Fig. 16a). Disease management is difficult but focuses on sanitation, fungicides, and biological 551 control, particularly in greenhouses. Quantitative resistance is reported in several hosts but has a limited 552 effect (Lucas and Griffiths, 2004, Wang and Davis, 1997). 553 The role of asexual spores depends on the strain. Py. ultimum is a species complex that includes Py. 554 ultimum var. ultimum and var. sporangiiferum, which have indistinguishable ITS sequences (Schroeder et 555 al., 2013). Sporangia and zoospores are produced rarely by the former, but abundantly by the latter. Py. 556 ultimum var. ultimum makes sporangia-like hyphal swellings which germinate to form infective hyphae (Fig. 557 16b; Stanghellini and Hancock, 1971). Similar to other oomycetes it also produces sexual oospores (Fig. 558 16c). 559 Genome sequencing of several Pythium spp. including Py. ultimum var. ultimum and var. 560 sporangiiferum has provided insight into their biology (Levesque et al., 2010, Adhikari et al., 2013). 561 Annotated in the ca. 43 Mb genomes of two subspecies are 15,290 and 14,086 genes, respectively, which is 562 less than the smallest Phytophthora but more than downy mildews. Whether this difference in gene number 563 is significant requires more analysis, since the two assemblies vary in quality and only single isolates were 564 sequenced. Large size variation between isolates was reported based on electrophoretic karyotyping 565 (Martin, 1995). 566 Evident nevertheless from genome analysis are many differences between Pythium and Phytophthora, 567 including a 25% reduction in Pythium of the fraction of proteins that are secreted. In part, this reflects the 568 absence of RXLR effectors, which may be unneeded for necrotrophic lifestyles. Pythium also lacks 569 haustoria, which biotrophic and hemibiotrophic oomycetes appear to use to deliver RXLRs to host cells 570 (Whisson et al., 2007). Another likely sign of the necrotrophy of Pythium is its greater abundance of lipases 571 and proteases. Pythium however lacks cutinases and pectin esterases, but this is probably because it 572 infects primarily nonsuberized tissue while Phytophthora (which encodes those enzymes) can penetrate 573 plant cuticles. Pythium encodes sufficient enzymes for macerating host cell walls, but cannot degrade some 574 ubiquitous wall components such as xylans, which suggests that soluble sugars may be a more important 575 carbon source (Zerillo et al., 2013). 576 Markers derived from Py. ultimum genomes should help reveal what distinguishes the subspecies and 577 provide more resolution to population studies, which can help identify the sources of outbreaks. Studies of 578 Py. ultimum var. ultimum indicated that populations are not clonal and some outcrossing may occur (Francis 579 and St. Clair, 1997). A bigger evolutionary question concerns the organization of the Pythium genus as a 580 whole. Division of the >120 Pythium species over five new genera was proposed based on sequence 581 analysis of two loci (Uzuhashi et al., 2010). If accepted by the community, Py. ultimum would be renamed 582 Globisporangium ultimum. 583 Of much interest is the interaction between Py. ultimum and biocontrol agents such as Trichoderma, 584 Streptomyces, Pythium oligandrum (a mycoparasitic member of the genus), and others (Gracia-Garza et al., 585 2003, Martin and Loper, 1999). The extent to which these organisms function by altering rhizosphere 586 microflora or attacking Py. ultimum directly is an open question (Naseby et al., 2000, Vallance et al., 2012); 587 Py. ultimum competes poorly with organisms having higher saprotrophic capabilities. 588 Py. ultimum is just one of many destructive members of the genus, with Py. aphanidermatum and Py. 589 irregulare also topping lists of important pathogens (Martin and Loper, 1999). Opportunities for 590 understanding their biology, ecology, and evolution have been enhanced by genome sequencing. DNA-591 mediated transformation was achieved for several species, but few studies of gene function are reported 592 (Weiland, 2003, Grenville-Briggs et al., 2013). Since core promoter structure appears conserved within the 593 Peronosporales, vectors and technologies applied to species such as Phytophthora should be transferable 594 to Pythium (Roy et al., 2013). Pythium spp. are easily grown and have the potential to develop into valuable 595 experimental systems for necrotrophic and saprotrophic oomycete lifestyles. Their growth on byproducts of 596 the agro-food industry has also attracted interest as sources of polyunsaturated fatty acids for human 597 A
ccep
ted
Arti
cle

Page 13
This article is protected by copyright. All rights reserved. 13
consumption (Stredansky et al., 2000). 598 599 10. ALBUGO CANDIDA 600 601 White blister rust is a disease caused in many dicotyledonous plant species by obligate biotrophic parasites. 602 For example, A. candida infection of Brassica juncea (Indian mustard) can result in significant crop losses in 603 India (Awasthi et al., 2012), Canada (Rimmer et al., 2000) and Australia (Kaur et al., 2008). The white rusts, 604 order Albuginales, are phylogenetically distant from the Peronosporales and probably represent an 605 independent acquisition of biotrophy (Thines and Spring, 2005; Thines and Kamoun 2010). All Albugo 606 species infecting the Brassicaceae were thought to be races of A. candida, but molecular studies of isolates 607 from various hosts and locations led to the description of specialists, for example Albugo laibachii on 608 Arabidopsis thaliana (Thines et al., 2009; Thines 2014). Specific A. candida races can grow on diverse plant 609 hosts, including Brassicaceae, Cleomaceae and Capparaceae (Thines, 2014). Albugo spp. provide an 610 interesting experimental system for the study of plant immune suppression, disease resistance and host-611 pathogen co-evolution. 612
Albugo spp. reproduce asexually via zoosporangia, which release flagellated motile zoospores 613 upon incubation in water. On the surface of a plant leaf, zoospores settle in stomata, and each extends a 614 germ tube into the sub-stomatal chamber (Holub et al., 1995). Coenocytic hyphae then grow intercellularly 615 through the plant. Small globose haustoria penetrate into plant cells (Soylu et al., 2003). When an Albugo 616 infection is mature, zoosporangia rupture the plant epidermis with force and enzymatic digestion (Heller and 617 Thines, 2009). This results in characteristic “white blister” pustules. Albugo also has a sexual cycle, 618 producing tough oospores that can survive difficult environmental conditions (Petrie, 1975). During systemic 619 infection of Brassicacea hosts, the inflorescences become misshapen, forming so-called ‘stagheads’ (Fig. 620 17A). 621
Albugo infection has long been associated with “green islands” where infected tissue appears 622 healthy and senescence is delayed. Infection by Albugo also greatly enhances susceptibility to co-infections 623 with downy mildews (Bains and Jhooty, 1985; Crute et al., 1994). Cooper et al (2008) investigated the ability 624 of A. laibachii and A. candida to suppress host immunity. They showed that A. laibachii can suppress the 625 “runaway cell death” of Arabidopsis lsd1 mutants after inoculation with avirulent H. arabidopsidis. 626 Furthermore, when pre-infected with virulent A. laibachii, resistant Arabidopsis accessions were no longer 627 resistant to avirulent H. arabidopsidis isolates (Fig. 17B), lettuce downy mildew or powdery mildew. 628 Suppression was also observed on B. juncea with A. candida and Brassica downy mildew (Cooper et al., 629 2008). These results suggest that Albugo is effective at broad suppression of plant immunity, including 630 effector-triggered-immunity. 631
The first step to understanding how Albugo spp. impose such susceptibility is to examine their 632 genomes. Links et al (2011) and Kemen et al (2011) sequenced A. candida and A. laibachii genomes, 633 respectively. The genomes are around 40 Mb and compact; about 50% of the assemblies consist of coding 634 sequences (see Fig. 18). Both genomes show adaptions to obligate biotrophy; they are missing sulfite 635 oxidases, nitrate and nitrite reductases and in the case of A. laibachii the whole molybdopterin biosynthesis 636 pathway. The A. candida secretome consists of 929 proteins (without transmembrane domains) compared 637 to 672 in A. laibachii, perhaps reflecting its wider host range. Within the secretomes there is no enrichment 638 of putative RXLR effectors. Kemen et al (2011) discovered the CHXC (cysteine, histidine, any amino acid, 639 cysteine) motif at the N-terminus of a class of candidate effectors. The CHXC-containing N-terminus is 640 sufficient to translocate the C-terminus of P. infestans AVR3a (an RXLR effector) into host cells (Kemen et 641 al., 2011). 642
Several A. candida races can infect some but not all A. thaliana accessions and from crosses 643 between resistant and susceptible accessions, an R gene against four A. candida races, WRR4 (encoding a 644 TIR-NB-LRR R-protein), was identified (Borhan et al., 2008). WRR4 can also provide resistance to A. 645 candida when transformed into susceptible cultivars of B. napus and B. juncea (Borhan et al., 2010). In A. 646 thaliana, RAC1 (also encoding a TIR-NB-LRR R-protein) confers resistance to A. laibachii (Borhan et al., 647 2004). The inheritance of avirulence of a B. juncea isolate (Ac2V) was studied through a cross between two 648 A
ccep
ted
Arti
cle

Page 14
This article is protected by copyright. All rights reserved. 14
A. candida isolates; this work predicted a single avirulence gene for the incompatibility between Ac2V and 649 B. rapa (Adhikari et al., 2003). 650
There are open questions about Albugo from both fundamental and translational perspectives. 651 Thines (2014) speculated that the broad host range of the A. candida meta-population is maintained through 652 frequent genetic exchange where the host range of individual isolates overlap. Comparing the genomes of 653 multiple isolates from different hosts would test this hypothesis and build up a clear picture of population 654 variation. This would also aid the discovery of new effector candidates through the identification of secreted 655 proteins under strong selective pressure. A more extensive analysis of Albugo effectors should be made. 656 The presence of the CHXC effectors inside host cells needs to be confirmed and the translocation 657 mechanism elucidated. It is unclear which Albugo effector proteins are recognised by the few known R-658 proteins. A. thaliana cannot be colonised by most A. candida isolates. The molecular basis for this 659 resistance could be exploited to introduce durable resistance to Brassica crops. 660
Lastly and perhaps most interestingly: how does the remarkable defense suppression by Albugo 661 work? We need to define the extent of this suppression- what other pests or pathogens can grow on plants 662 infected with Albugo? What changes occur in the microbiome of Albugo-infected leaves in the field? Recent 663 data suggested that Albugo could be widespread as an asymptomatic endophyte (Ploch and Thines, 2011). 664 The implications of these infections as a reservoir for i) Albugo and ii) further Albugo suppression-enabled 665 infections by other species remain to be discovered. 666 667 CONCLUSION 668 669 Although the study of oomycete plant pathogens has always been an important topic in plant pathology, it 670 has recently taken an even more central role with the advent of genomics and the discovery of large effector 671 repertoires in oomycete genomes. This article provides a benchmark for future trends. Many topics remain 672 to be investigated in more depth. For example, the large number of pathogenic species and the wide 673 diversity of host species and host ranges should provide a rich basis for investigating the genetic and 674 physiological basis for host adaptation and specialization. It will be also be interesting to track how the Top 675 10 list will develop in the coming years. Many pathogenic oomycetes among the 33 listed (Table 2) have 676 evolved unique adaptations in their parasitic lifestyle that will undoubtedly reveal fascinating processes and 677 mechanisms. Thus future research efforts should take into account a diverse spectrum of taxa beyond the 678 widely studied species. 679 680 ACKNOWLEDGEMENTS 681
The authors would like to thank Dr. Diane Hird for assistance with several aspects of the project. 682 Sophien Kamoun, Oliver J. Furzer and Jonathan D.G. Jones received funding from the European Research 683 Council (ERC), the UK Biotechnology and Biological Sciences Research Council (BBSRC), and the Gatsby 684 Charitable Foundation. Ronaldo Dalio thanks the National Council for Scientific and Technological 685 Development (CNPq/CsF Brazil - 313139/2013-0) for financial support. Michelina Ruocco receives support 686 from Conoscenze Integrate per Sostenibilità ed Innovazione del Made in Italy Agroalimentare (CISIA –687 MIUR). Leonardo Schena was supported by grant FIRB 2010 - RBFR10PZ4N from the Italian Ministry of 688 Education, University and Research (MIUR). Andreia Figueiredo receives support from the Portuguese 689 Foundation for Science and Technology (Grant nº SFRH / BPD / 63641 / 2009). Xiao-Ren Chen was 690 financed by National Natural Science Foundation of China (Grant no. 31101395) and Jiangsu Province 691 Basic Research Program (Natural Science Foundation) of China (Grant no. BK2011443). Howard S. 692 Judelson and Brett M. Tyler were supported by grants 2011-68004-30154 and 2011-68004-30104, 693 respectively, from the Agriculture and Food Research Institute of the National Institute of Food and 694 Agriculture of the USDA. Mark Gijzen was supported by Agriculture and Agri-Food Canada GRDI program. 695 Niklaus J. Grünwald was supported by grant 2011-68004-30154 from the USDA National Institute of Food 696 and Agriculture, the National Research Initiative Competitive Grants Program grant 2008-35600-18780, the 697 USDA Agricultural Research Service CRIS 5358-22000-039-00D, the Northwest Center for Nursery Crop 698 A
ccep
ted
Arti
cle

Page 15
This article is protected by copyright. All rights reserved. 15
Research, the US Forest Service, the USDA ARS Floriculture Nursery Initiative and Oregon Department of 699 Agriculture/ Oregon Association of Nurseries. Daniel F.A.Tomé is funded by the UK Biotechnology and 700 Biological Sciences Research Council grant BB/G015066/1. John McDowell is supported by the U.S. 701 Department of Agriculture–Agriculture and Food Research Initiative (2009-03008 and 2011-68004), the 702 National Science Foundation (ABI-1146819), and the Virginia Tech Institute for Critical Technology and 703 Applied Sciences. Fouad Daayf received funding from NSERC, MB-ARDI, McCain Foods, KPPA, and Peak 704 of the Market. Harold J.G. Meijer is funded by The Dutch Technology Foundation STW-NWO (VIDI grant 705 10281). Benjamin Petre is supported by an INRA Contrat Jeune Scientifique and has received the 706 support of the European Union, in the framework of the Marie-Curie FP7 COFUND People 707 Programme, through the award of an AgreenSkills’ fellowship (under grant agreement n° 708 267196). Francine Govers receives support from the Food for Thought program, Wageningen University 709 Fund. 710 711 712 AUTHOR CONTRIBUTIONS 713 714 All authors voted and contributed to writing the 10 sections. SK oversaw the voting and writing. Other 715 authors are listed in reverse order based on the species ranking. The species coordinator(s) are listed last in 716 their section and are preceded by other contributors listed in alphabetical order. The species coordinators 717 are: A. candida (OF and JDGJ), Py. ultimum (HJ), P. parasitica (FP), P. cinnamomi (DC), Pl. viticola (MR 718 and AF), P. capsici (KL), P. sojae (MG and BMT), P. ramorum (NJG), H. arabidopsidis (GvdA and JMcD), 719 and P. infestans (PRJB and FG). 720 721 REFERENCES 722
723 Adhikari, B. N., Hamilton, J. P., Zerillo, M. M., Tisserat, N., Levesque, C. A. and Buell, C. R. (2013) 724
Comparative genomics reveals insight into virulence strategies of plant pathogenic oomycetes. PloS 725 One 8, e75072 726
Adhikari, T. B., J. Q. Liu, S. Mathur, C. R. X. Wu, and S. R. Rimmer. 2003. Genetic and molecular 727 analyses in crosses of race 2 and race 7 of Albugo candida. Phytopathology no. 93 (8):959-965. 728
Agrios, G. N. (2005) Plant Pathology. United States of America: Elsevier Academic Press Publications. 729 Ah Fong, A.M.V. and Judelson, H.S. (2003) Cell cycle regulator Cdc14 is expressed during sporulation but 730
not hyphal growth in the fungus-like oomycete Phytophthora infestans. Mol. Microbiol. 50, 487-494. 731 Akinsanmi, O.A. and Drenth, A. (2013) Phosphite and metalaxyl rejuvenate macadamia trees in decline 732
caused by Phytophthora cinnamomi. Crop Protection 53, 29-36 733 Allardyce, J.A., Rookes J.E., Hussain, H.I. and Cahill, D.M. (2013) Transcriptional profiling of Zea mays 734
roots reveals roles for jasmonic acid and terpenoids in resistance against Phytophthora cinnamomi. 735 Functional and Integrative Genomics 13, 217-228 736
Allen, R. L., Bittner-Eddy, P. D., Grenvitte-Briggs, L. J., Meitz, J. C., Rehmany, A. P., Rose, L. E., et al. 737 (2004) Host-parasite coevolutionary conflict between Arabidopsis and downy mildew. Science, 306, 738 1957-1960. 739
Andres, J. L., Rivera, A. and Fernandez, J. A. (2003) Phytophthora nicotianae pathogenic to pepper in 740 northwest spain. J Plant Pathol, 85, 91-98. 741
Attard, A., Gourgues, M., Callemeyn-Torre, N. and Keller, H. (2010) The immediate activation of defense 742 responses in Arabidopsis roots is not sufficient to prevent Phytophthora parasitica infection. New 743 Phytol, 187, 449-460. 744
Attard, A., Gourgues, M., Galiana, E., Panabieres, F., Ponchet, M. and Keller, H. (2008) Strategies of 745 attack and defense in plant-oomycete interactions, accentuated for Phytophthora parasitica Dastur (syn 746 P. nicotianae Breda de Haan). J. Plant Physiol., 165, 83-94. 747
Australian Government Department of Environment (2014) Threat abatement plan for disease in natural 748 ecosystems caused by Phytophthora cinnamomi. Commonwealth of Australia, Canberra, 33 pp. 749 Acc
epte
d A
rticl
e

Page 16
This article is protected by copyright. All rights reserved. 16
Armstrong, M.R., Whisson, S.C., Pritchard, L., Bos, J.I., Venter, E., Avrova, A.O., Rehmany, A.P., 750 Bohme, U., Brooks, K., Cherevach, I., Hamlin, N., White, B., Fraser, A., Lord, A., Quail, M.A., 751 Churcher, C., Hall, N., Berriman, M., Huang, S., Kamoun, S., Beynon, J.L. and Birch, P.R. (2005) 752 An ancestral oomycete locus contains late blight avirulence gene Avr3a, encoding a protein that is 753 recognized in the host cytoplasm. Proc. Natl. Acad. Sci. USA, 102, 7766-7771. 754
Awasthi, R. P., N. I. Nashaat, S. J. Kolte, A. K. Tewari, P. D. Meena, and Bhatt Renu. 2012. Screening 755 of putative resistant sources against Indian and exotic isolates of Albugo candida inciting white rust in 756 rapeseed-mustard. Journal of Oilseed Brassica no. 3 (1):27-37. 757
Bains, S. S., and J. S. Jhooty. 1985. Association of peronospora-parasitica with albugo-candida on 758 brassica-juncea leaves. Phytopathologische Zeitschrift-Journal of Phytopathology no. 112 (1):28-31. 759
Baxter, L., Tripathy, S., Ishaque, N., Boot, N., Cabral, A., Kemen, E., et al. (2010) Signatures of 760 adaptation to obligate biotrophy in the Hyaloperonospora arabidopsidis genome. Science, 330, 1549-761 1551. 762
Blum, M., Waldner, M. and Gisi, U. (2010) A single point mutation in the novel PvCesA3 gene confers 763 resistance to the carboxylic acid amide fungicide mandipropamid in Plasmopara viticola. Fungal 764 Genetics and Biology, 47, 499-510. 765
Bonnet, P., Lacourt, I., Venard, P. and Ricci, P. (1994) Diversity in pathogenicity to tobacco and in elicitin 766 production among isolates of Phytophthora parasitica. J. Phytopathol., 141, 25-37. 767
Borhan, M. H., N. Gunn, A. Cooper, S. Gulden, M. Tör, S. R. Rimmer, and E. B. Holub. 2008. WRR4 768 encodes a TIR-NB-LRR protein that confers broad-spectrum white rust resistance in Arabidopsis 769 thaliana to four physiological races of Albugo candida. Mol Plant Microbe Interact no. 21 (6):757-68. 770 doi: 10.1094/MPMI-21-6-0757. 771
Borhan, M. H., E. B. Holub, J. L. Beynon, K. Rozwadowski, and S. R. Rimmer. 2004. The Arabidopsis 772 TIR-NB-LRR gene RAC1 confers resistance to Albugo candida(white Rust) and is dependant on EDS1 773 but not PAD4. Molecular Plant-Microbe Interactions no. 17 (7):711-719. 774
Borhan, M. H., E. B. Holub, C. Kindrachuk, M. Omidi, G. Bozorgmanesh-Frad, and S. R. Rimmer. 775 2010. WRR4, a broad-spectrum TIR-NB-LRR gene from Arabidopsis thaliana that confers white rust 776 resistance in transgenic oilseed brassica crops. Molecular Plant Pathology no. 11 (2):283-291. doi: 777 10.1111/j.1364-3703.2009.00599.x. 778
Bos, J.I., Armstrong, M.R., Gilroy, E.M., Boevink, P.C., Hein, I., Taylor, R.M., Zhendong, T., 779 Engelhardt, S., Vetukuri, R.R., Harrower, B., Dixelius, C., Bryan, G., Sadanandom, A., Whisson, 780 S.C., Kamoun, S. and Birch, P.R. (2010) Phytophthora infestans effector AVR3a is essential for 781 virulence and manipulates plant immunity by stabilizing host E3 ligase CMPG1. Proc. Natl. Acad. Sci. 782 USA, 107, 9909-9914. 783
Bozkurt, T.O., Schornack, S., Banfield, M.J. and Kamoun, S. (2012) Oomycetes, effectors, and all that 784 jazz. Curr Opin Plant Biol, 15:483-492. 785
Brasier, C., and Webber, J. 2010. Plant pathology: Sudden larch death. Nature. 466:824–825 786 Brasier, C. M. (2008) The biosecurity threat to the UK and global environment from international trade in 787
plants. Plant Pathol., 57, 792-808. 788 Brasier, C. M., Vettraino, A. M., Chang, T. T., and Vannini, A. 2010. Phytophthora lateralis discovered in 789
an old growth Chamaecyparis forest in Taiwan. Plant Pathol. 59:595–603 790 Cabral, A., Oome, S., Sander, N., Kufner, I., Nurnberger, T. and Van den Ackerveken, G. (2012) 791
Nontoxic Nep1-like proteins of the downy mildew pathogen Hyaloperonospora arabidopsidis: repression 792 of necrosis-inducing activity by a surface-exposed region. Mol Plant Microbe Interact, 25, 697-708. 793
Cabral, A., Stassen, J. H., Seidl, M. F., Bautor, J., Parker, J. E. and Van den Ackerveken, G. (2011) 794 Identification of Hyaloperonospora arabidopsidis transcript sequences expressed during infection 795 reveals isolate-specific effectors. Plos One, 6, e19328. 796
Caillaud, M.-C., Piquerez, S. J. M., Fabro, G., Steinbrenner, J., Ishaque, N., Beynon, J., et al. (2011) 797 Subcellular Localization of the H. arabidopsidis RxLR Effector Repertoire Identifies the Extrahaustorial 798 Membrane-Localized HaRxL17 that Confers Enhanced Plant Susceptibility. The Plant Journal 69, 52. 799 Acc
epte
d A
rticl
e

Page 17
This article is protected by copyright. All rights reserved. 17
Casagrande, K., Falginella, L., Castellarin, S., Testolin, R. and Di Gaspero, G. (2011) Defence 800 responses in Rpv3-dependent resistance to grapevine downy mildew. Planta, 234, 1097-1109. 801
Chamnanpunt, J., Shan, W. X. and Tyler, B. M. (2001) High frequency mitotic gene conversion in genetic 802 hybrids of the oomycete Phytophthora sojae. Proc Natl Acad Sci U S A, 98, 14530-14535. 803
Chen, W., Delmotte, F., Richard-Cervera, S., Douence, L., Greif, C. and Corio-Costet, M. (2007) At 804 least two origins of fungicide resistance in grapevine downy mildew Populations. Applied and 805 Environmental Microbiology, 73, 5162-5172. 806
Chen, X. R., Xing, Y. P., Li, Y. P., Tong, Y. H. and Xu, J. Y. (2013) RNA-Seq reveals infection-related 807 gene expression changes in Phytophthora capsici. PloS one, 8, e74588. 808
Cline, E. T., Farr, D. F. and Rossman, A. Y. (2008) Synopsis of Phytophthora with accurate scientific 809 names, host range, and geographic distribution. Plant Health Progress, doi:10.1094/PHP-2008 0318-810 01-RS. 811
Colas, V., Conrod, S., Venard, P., Keller, H., Ricci, P. and Panabieres, F. (2001) Elicitin genes 812 expressed in vitro by certain tobacco isolates of Phytophthora parasitica are down regulated during 813 compatible interactions. Mol. Plant Microbe Interact., 14, 326-335. 814
Colas, V., Lacourt, I., Ricci, P., Vanlerberghe-Masutti, F., Poupet, A. and Panabieres, F. (1998) 815 Diversity of virulence in Phytophthora parasitica on tobacco, as reflected by nuclear RFLPs. 816 Phytopathology, 88, 205-212. 817
Cooke, D.E.L., Cano, L.M., Raffaele, S., Bain, R.A., Cooke, L.R., Etherington, G.J., Deahl, K.L., Farrer, 818 R.A., Gilroy, E.M., Goss, E.M., Grünwald, N.J., Hein, I., Maclean, D.J., McNicol, J.W., Randall, E., 819 Oliva, R.F., Pel, M.A., Shaw, D.S., Squires, J.N., Taylor, M.C., Vleeshouwers, V.G.A.A., Birch, 820 P.R.J., Lees, A.K. and Kamoun, S. (2012) Genome analyses of an aggressive and invasive lineage of 821 the Irish potato famine pathogen. PLoS Pathog. 8, e1002940. 822
Cooper, AJ, AO Latunde-Dada, A Woods-Tör, J Lynn, JA Lucas, IR Crute, and EB Holub. 2008. Basic 823 compatibility of Albugo candida in Arabidopsis thaliana and Brassica juncea causes broad-spectrum 824 suppression of innate immunity. Mol Plant Microbe Interact 21 (6):745-56. 825
Crane, C.E. and Shearer, B.L. (2014) Comparison of phosphite application methods for control of 826 Phytophthora cinnamomi in threatened communities. Australasian Plant Pathology 43, 143-149 827
Crute, I, J Beynon, J Dangl, E Holub, B Mauch-Mani, A Slusarenko, B Staskawicz, and F Ausubel. 828 1994. Microbial pathogenesis of Arabidopsis. In Arabidopsis: Cold Spring Harbor Laboratory Press. 829
DeArce, M. (2008) Correspondence of Charles Darwin on James Torbitt's project to breed blight-resistant 830 potatoes. Arch. Nat. Hist. 35, 208-222. 831
Dean, R., Van Kan, J.A.L., Pretorius, Z.A., Hammond-Kosack, K.E., Di Pietro, A., Spanu, P.D., Rudd, 832 J.J., Dickman, M., Kahmann, R., Ellis, J. and Foster, G.D. (2012) The top 10 fungal pathogens in 833 molecular plant pathology. Mol. Plant Pathol. 13, 414–430. 834
Delmotte, F., Mestre, P., Schneider, C., Kassemeyer, H.-H., Kozma, P., Richart-Cervera, S., et al. 835 (2013) Rapid and multiregional adaptation to host partial resistance in a plant pathogenic oomycete: 836 Evidence from European populations of Plasmopara viticola, the causal agent of grapevine downy 837 mildew. Infection, Genetics and Evolution: http://dx.doi.org/10.1016/j.meegid.2013.10.017 838
Dempsey, R.W., Merchant, A. and Tausz, M. (2012) Differences in ascorbate and glutathione levels as 839 indicators of resistance and susceptibility in Eucalyptus trees infected with Phytophthora cinnamomi. 840 Tree Physiology 32, 1148-1160 841
Dick, M. W. (2001) Straminipilous Fungi. Dordrecht: Kluwer Academic Publishers. 842 Dong, S., Stam, R., Cano, L.M., Song, J., Sklenar, J., Yoshida, K., Bozkurt, T.O., Oliva, R., Liu, Z., 843
Tian, M., Win, J., Banfield, M.J., Jones, A.M.E., van der Hoorn, R.A.L. and Kamoun, S. (2014) 844 Effector specialization in a lineage of the Irish Potato Famine Pathogen. Science 343, 552-555. 845
Dorrance, A. and Grünwald, N. J. (2009) Phytophthora sojae: Diversity among and within Populations. In: 846 Oomycete Genetics and Genomics: Diversity, Interactions, and Research Tools. (Lamour, K. and 847 Kamoun, S., eds.). John Wiley & Sons, Inc., pp. 197-212. 848 Acc
epte
d A
rticl
e

Page 18
This article is protected by copyright. All rights reserved. 18
Dou, D., Kale, S. D., Wang, X., Jiang, R. H. Y., Bruce, N. A., Arredondo, F. D., et al. (2008) RXLR-849 mediated entry of Phytophthora sojae effector Avr1b into soybean cells does not require pathogen-850 encoded machinery. Plant Cell, 20, 1930–1947. 851
Drenth, A., Tas, I.C.Q. and Govers, F. (1994) DNA fingerprinting uncovers a new sexually reproducing 852 population of Phytophthora infestans in the Netherlands. Eur. J. Plant Pathol. 100, 97-107. 853
Dunn, A., Milgroom, M., Meitz, J., McLeod, A., Fry, W., McGrath, M., Dillard, H. and Smart, C. (2010) 854 Population structure and resistance to mefenoxam of Phytophthora capsici in New York State. Plant 855 Disease, 94, 1461-1468. 856
Dunstan, W. A., Rudman, T., Shearer, B. L., Moore, N. A., Paap, T., Calver, M. C., Dell, B., Hardy, G. E. 857 St J. (2010) Containment and spot eradication of a highly destructive, invasive plant pathogen (P. 858 cinnamomi) in natural ecosystems. Biological Invasions 12, 913–925. 859
Elliott, M., Sumampong, G., Varga, A., Shamoun, S. F., James, D., Masri, S., and Gruenwald, N. J. 860 2011. Phenotypic differences among three clonal lineages of Phytophthora ramorum. For. Pathol. 41:7–861 14 862
Enkerli, K., Hahn, M. G. and Mims, C. W. (1997) Ultrastructure of compatible and incompatible interactions 863 of soybean roots infected with the plant pathogenic oomycete Phytophthora sojae. Can. J. Bot., 75, 864 1494-1508. 865
Erwin, D. C. and Ribeiro, O. K. (1996) Phytophthora diseases worldwide. St. Paul, MN. American 866 Phytopathological Society. 867
Eshraghi, L., Anderson, J.P., Aryamanesh, N., McComb, J.A., Shearer B.S. and Hardy, G.StJ.E. (2014) 868 Suppression of the auxin response pathway enhances susceptibility to Phytophthora cinnamomi while 869 phosphite-mediated resistance stimulates the auxin signalling pathway. BMC Plant Biology 14:68 870
Fabro, G., Steinbrenner, J., Coates, M., Ishaque, N., Baxter, L., Studholme, D. J., et al. (2011) Multiple 871 Candidate Effectors from the Oomycete Pathogen Hyaloperonospora arabidopsidis Suppress Host 872 Plant Immunity. PLoS Pathogens, 7, e1002348. 873
Fahlgren, N., Bollmann, S. R., Kasschau, K. D., Cuperus, J. T., Press, C. M., Sullivan, C. M., 874 Chapman, E. J., Hoyer, J. S., Gilbert, K. B., Grünwald, N. J., and Carrington, J. C. 2013. 875 Phytophthora have distinct endogenous small RNA populations that include short interfering and 876 microRNAs. PLoS One. 8:e77181 877
Farr, D. F. and Rossman, A. Y. (2014) Fungal Databases, Systematic Mycology and Microbiology 878 Laboratory, ARS, USDA. http://nt.ars-grin.gov/fungaldatabases/. 879
Feng, B. Z. and Li, P. Q. (2013) Molecular characterization and functional analysis of the Nep1-like protein-880 encoding gene from Phytophthora capsici. Genetics and Molecular Research, 12, 1468-1478. 881
Feng, B. Z., Li, P. Q., Wang, H. M. and Zhang, X. G. (2010) Functional analysis of Pcpme6 from oomycete 882 plant pathogen Phytophthora capsici. Microb Pathogenesis, 49, 23-31. 883
Fisher, M.C., Henk, D.A., Briggs, C.J., Brownstein, J.S., Madoff, L.C., McCraw, S.L., et al. (2012) 884 Emerging fungal threats to animal, plant and ecosystem health. Nature 484,186–94. 885
Fontaine, M., Austerlitz, F., Giraud, T., Labbe, F., Papura, D., Richard-Cervera, S., et al. (2013) Genetic 886 signature of a range expansion and leap-frog event after the recent invasion of Europe by the grapevine 887 downy mildew pathogen Plasmopara viticola. Molecular Ecology, 22, 2771-2786. 888
Francis, D. M. and St. Clair, D. A. (1997) Population genetics of Pythium ultimum. Phytopathology 87, 454-889 461. 890
Fry, W.E., McGrath, M.T., Seaman, A., Zitter, T.A., McLeod, A., Danies, G., Small, I.M., Myers, K., 891 Everts, K., Gevens, A.J., Gugino, B.K., Johnson, S.B., Judelson, H., Ristaino, J., Roberts, P., 892 Secor, G., Seebold, K., Snover-Clift, K., Wyenandt, A., Grünwald, N.J. and Smart, C.D. (2013) The 893 2009 Late Blight Pandemic in the Eastern United States – Causes and Results. Plant Dis. 97, 296-306. 894
Galet, P. (1977) Mildiou. Paris: Lavoisier. 895 Garbelotto, M., and Hayden, K. J. 2012. Sudden oak death: interactions of the exotic oomycete 896
Phytophthora ramorum with naïve North American hosts. Eukaryot. Cell. 11:1313–23 897 Gessler, C., Pertot, I. and Perazzolli, M. (2011) Plasmopara viticola: a review of knowledge on downy 898
mildew of grapevine and effective disease management. Phytopathol Mediterr, 50, 3-44. 899 Acc
epte
d A
rticl
e

Page 19
This article is protected by copyright. All rights reserved. 19
Gijzen, M. and Qutob, D. (2009) Phytophthora sojae and Soybean. In: Oomycete Genetics and Genomics: 900 Diversity, Interactions, and Research Tools. (Lamour, K. and Kamoun, S., eds.). John Wiley & Sons, 901 Inc., pp. 303-329. 902
Gobbin, D., Jermini, M., Loskill, B., Pertot, I., Raynal, M. and Gessler, C. (2005) Importance of 903 secondary inoculum of Plasmopara viticola to epidemics of grapevine downy mildew. Plant Pathology, 904 54, 522-534. 905
Gobena, D., McGrath, M. T. and Lamour, K. H. (2012) Survival and spread of Phytophthora capsici on 906 Long Island, New York. Mycol. Prog., 11, 761-768. 907
Gobena, D. J., Roig, J., Hulvey, J. and Lamour, K. (2010) Genetic diversity of the vegetable pathogen 908 Phytophthora capsici in Argentina. Phytopathology, 100, S41-S41. 909
Goodwin, S.B., Cohen, B.A. and Fry, W.E. (1994) Panglobal distribution of a single clonal lineage of the 910 Irish potato famine fungus. Proc. Natl. Acad. Sci. USA, 91, 11591-11595. 911
Goodwin, S.B., Drenth, A. and Fry, W.E. (1992) Cloning and genetic analyses of two highly polymorphic, 912 moderately repetitive nuclear DNAs from Phytophthora infestans. Curr. Genet. 22, 107-115. 913
Goss, E. M., Carbone, I., and Grünwald, N. J. 2009a. Ancient isolation and independent evolution of the 914 three clonal lineages of the exotic sudden oak death pathogen Phytophthora ramorum. Mol. Ecol. 915 18:1161–1174 916
Goss, E. M., Larsen, M., Chastagner, G. A., Givens, D. R., and Grünwald, N. J. 2009b. Population 917 genetic analysis infers migration pathways of Phytophthora ramorum in US nurseries. PLoS Pathog. 918 5:e1000583 919
Goss, E. M., Larsen, M., Vercauteren, A., Werres, S., Heungens, K., and Grünwald, N. J. 2011. 920 Phytophthora ramorum in Canada: evidence for migration within North America and from Europe. 921 Phytopathology. 101:166–71 922
Goss, E. M., Press, C. M., and Grünwald, N. J. 2013. Evolution of RXLR-class effectors in the oomycete 923 plant pathogen Phytophthora ramorum. PLoS One. 8:e79347 924
Govers, F. and Gijzen, M. (2006) Phytophthora genomics: the plant destroyers’ genome decoded. Mol. 925 Plant–Microbe Interact. 19, 1295– 1301. 926
Gracia-Garza, J. A., Little, M., Brown, W., Blom, T. J., Schneider, K., Allen, W., et al. (2003) Efficacy of 927 various biological control agents and biorationals against Pythium root rot in poinsettia. Horttechnology 928 13, 149-153. 929
Granke, L., Quesada-Ocampo, L., Lamour, K. and Hausbeck, M. K. (2012) Advances in research on 930 Phytophthora capsici on vegetable crops in the United States. Plant Disease, 95, 1588-1600. 931
Granke, L., Windstam, S., Hoch, H., Smart, C. and Hausbeck, M. (2009) Dispersal and movement 932 mechanisms of Phytophthora capsici sporangia. Phytopathology, 99, 1258-1264. 933
Grenville-Briggs, L. J., Horner, N. R., Phillips, A. J., Beakes, G. W. and Van West, P. (2013) A family of 934 small tyrosine rich proteins is essential for oogonial and oospore cell wall development of the 935 mycoparasitic oomycete Pythium oligandrum. Fungal Biol. 117, 163-172. 936
Grünwald, N. J., Garbelotto, M., Goss, E. M., Heungens, K., and Prospero, S. 2012. Emergence of the 937 sudden oak death pathogen Phytophthora ramorum. Trends Microbiol. 20:131–8 938
Grünwald, N. J., Goss, E. M., Ivors, K., Garbelotto, M., Martin, F. N., Prospero, S., Hansen, E. M., 939 Bonants, P. J. M., Hamelin, R. C., Chastagner, G., Werres, S., Rizzo, D. M., Abad, G., Beales, P., 940 Bilodeau, G. J., Blomquist, C. L., Brasier, C., Brière, S. C., Chandelier, A., Davidson, J. M., 941 Denman, S., Elliott, M., Frankel, S. J., Goheen, E. M., de Gruyter, H., Heungens, K., James, D., 942 Kanaskie, A., McWilliams, M. G., Veld, W. M. in ‘t, Moralejo, E., Osterbauer, N. K., Palm, M. E., 943 Parke, J. L., Sierra, A. M. P., Shamoun, S. F., Shishkoff, N., Tooley, P. W., Vettraino, A. M., 944 Webber, J., and Widmer, T. L. 2009. Standardizing the nomenclature for clonal lineages of the sudden 945 oak death pathogen, Phytophthora ramorum. Phytopathology. 99:792–795 946
Grünwald, N. J., Goss, E. M., and Press, C. M. 2008. Phytophthora ramorum: a pathogen with a 947 remarkably wide host-range causing sudden oak death on oaks and ramorum blight on woody 948 ornamentals. Mol. Plant Pathol. 9:729–740 949 Acc
epte
d A
rticl
e

Page 20
This article is protected by copyright. All rights reserved. 20
Guha Roy, S., Mukherjee, S. K., Bhattacharyya, S. and Khatua, D. C. (2009) Molecular identification of 950 Phytophthora sp. affecting some economicaly important crops in Eastern India through ITS-RFLP and 951 sequencing of the ITS region. J. Phytopathol., 157, 666-674. 952
Gunderson, J. H., Elwood, H., Ingold, H., Kindle, A., and Sogin, M. L. (1987) Phylogenetic relationships 953 between chlororphytes, chrysophytes, and oomycetes. Proc. Natl. Acad. Sci. U.S.A. 84:5823-5827. 954
Gunning, K., Conlan, X.A., Parker, R.M., Dyson, G.A., Adams, M.J., Barnett, N.W. and Cahill, D.M. 955 (2013) Profiling of secondary metabolites in blue lupin inoculated with Phytophthora cinnamomi 956 following phosphite treatment. Functional Plant Biology, 40, 1089-1097 957
Haas, B.J., Kamoun, S., Zody, M.C., Jiang, R.H.Y., Handsaker, R.E., Cano, L.M., Grabherr, M., Kodira, 958 C.D., Raffaele, S., Torto-Alalibo, T., Bozkurt, T.O., Ah-Fong, A.M.V., Alvarado, L., Anderson, V.L., 959 Armstrong, M.R., Avrova, A.O., Baxter, L., Beynon, J.L., Boevink, P.C., Bollmann, S.R., Bos, 960 J.I.B., Bulone, V., Cai, G., Cakir, C., Carrington, J.C., Chawner, M., Conti, L., Costanzo, S., Ewan, 961 R., Fahlgren, N., Fischbach, M.l.A., Fugelstad, J., Gilroy, E.M., Gnerre, S., Green, P.J., Grenville-962 Briggs, L.J., Griffith, J.M., Grünwald, N.J., Horn, K., Horner, N.R., Hu, C.H., Huitema, E., Jeong, 963 D.H., Jones, A.M.E., Jones, J.D.G., Jones, R.W., Karlsson, E.K., Kunjeti, S.G., Lamour, K., Liu, Z., 964 Ma, L.J., Maclean, D.J., Chibucos, M.C., McDonald, H., McWalters, J., Meijer, H.J.G., Morgan, W., 965 Morris, P.F., Munro, C.A., O'Neill, K., Ospina-Giraldo, M.D., Pinzon, A., Pritchard, L., Ramsahoye, 966 B., Ren, Q., Restrepo, S., Roy, S., Sadanandom, A., Savidor, A., Schornack, S., Schwartz, D.C., 967 Schumann, U.D., Schwessinger, B., Seyer, L., Sharpe, T., Silvar, C., Song, J., Studholme, D.J., 968 Sykes, S., Thines, M., van de Vondervoort, P.J.I., Phuntumart, V., Wawra, S., Weide, R., Win, J., 969 Young, C., Zhou, S., Fry, W.E., Meyers, B.C., van West, P., Ristaino, J.B., Govers, F., Birch, 970 P.R.J., Whisson, S.C., Judelson, H.S. and Nusbaum, C. (2009) Genome sequence and analysis of 971 the Irish potato famine pathogen Phytophthora infestans. Nature, 461, 393-398. 972
Haverkort, A., Boonekamp, P., Hutten, R., Jacobsen, E., Lotz, L., Kessel, G., Visser, R. and van der 973 Vossen, E. (2008) Societal costs of late blight in potato and prospects of durable resistance through 974 cisgenic modification. Potato Res. 51, 47-57. 975
Hausbeck, M. K. and Lamour, K. H. (2004) Phytophthora capsici on vegetable crops: research progress 976 and management challenges. Plant Disease, 88, 1292-1303. 977
Hee, W. Y., Torreña, P. S., Blackman, L. M. and Hardham, A. R. (2013) Phytophthora cinnamomi in 978 Australia. In: Lamour, K. (ed) Phytophthora: a global perspective, CABI Plant Protection Series, No.2, 979 pp. 124-134 980
Heller, A., and M. Thines. 2009. Evidence for the importance of enzymatic digestion of epidermal walls 981 during subepidermal sporulation and pustule opening in white blister rusts (Albuginaceae). Mycol Res 982 no. 113 (Pt 6-7):657-67. doi: S0953-7562(09)00031-8 [pii] 983
10.1016/j.mycres.2009.01.009. 984 Hildebrand, A. A. (1959) A root and stalk rot of soybeans caused by Phytophthora megasperma Drechsler 985
var. sojae var. nov. Canadian Journal of Botany, 37, 927-957. 986 Holub, E., Brose, E., Tör, M., Clay, C., Crute, I. R. and Beynon, J. L. (1994) Phenotypic and genotypic 987
variation in the interaction between Arabidopsis thaliana and Albugo candida. Molec. Plant-Microbe 988 Interact., 8, 916-928. 989
Holub, E. B. (2008) Natural history of Arabidopsis thaliana and oomycete symbioses. Eur J Plant Pathol, 990 122, 91-109. 991
Hüberli, D., Hayden, K. J., Calver, M., and Garbelotto, M. 2012. Intraspecific variation in host 992 susceptibility and climatic factors mediate epidemics of sudden oak death in western US forests. Plant 993 Pathol. 61:579–592 994
Hu, C. H., Perez, F. G., Donahoo, R., McLeod, A., Myers, K., Ivors, K., Secor, G., Roberts, P. D., Fry, 995 W. E., Deahl, K. L., and Ristaino. J. B. (2012). Recent genotypes of Phytophthora infestans in 996 eastern USA reveal clonal populations and reappearance of mefenoxam sensitivity. Plant Dis. 96: 997 1323-1330. 998
Hu, J., Diao, Y., Zhou, Y., Lin, D., Bi, Y., Pang, Z., Trout Fryxell, R., Liu, X. and Lamour, K. (2013) Loss 999 of heterozygosity drives clonal diversity of Phytophthora capsici in China. PloS ONE, 8, e82691. 1000 Acc
epte
d A
rticl
e

Page 21
This article is protected by copyright. All rights reserved. 21
Hulvey, J., Young, J., Finley, L. and Lamour, K. (2010) Loss of heterozygosity in Phytophthora capsici 1001 after N-ethyl-nitrosourea mutagenesis. Mycologia, 102, 27-32. 1002
Hurtado-Gonzales, O., Aragon-Caballero, L., Apaza-Tapia, W., Donahoo, R. and Lamour, K. (2008) 1003 Survival and spread of Phytophthora capsici in coastal Peru. Phytopathology, 98, 688-694. 1004
Hussain, H.I., Yi, Z., Rookes, J.E., Kong, L.X. and Cahill, D.M. (2013) Mesoporous silica nanoparticles as 1005 a biomolecule delivery vehicle in plants, Journal of Nanoparticle Research, 15, 1-15 1006
Jiang, R. H. Y., Tripathy, S., Govers, F. and Tyler, B. M. (2008) RXLR effector reservoir in two 1007 Phytophthora species is dominated by a single rapidly evolving super-family with more than 700 1008 members. Proc. Natl. Acad. Sci. USA, 105, 4874-4879. 1009
Jiang, R. H. Y. and Tyler, B. M. (2012) Mechanisms and evolution of virulence in oomycetes. Ann. Rev. 1010 Phytopath., 50, 295-318. 1011
Jones, J.T., Haegeman, A., Danchin, E.G., Gaur, H.S., Helder, J., Jones, M.G., Kikuchi, T., Manzanilla-1012 López, R., Palomares-Rius, J.E., Wesemael, W.M., Perry, R.N. (2013) Top 10 plant-parasitic 1013 nematodes in molecular plant pathology. Mol Plant Pathol 14, 946-61. 1014
Judelson, H.S. (1997) The genetics and biology of Phytophthora infestans: modern approaches to a 1015 historical challenge. Fungal Genet. Biol. 22, 65-76. 1016
Judelson, H.S. and Blanco, F.A. (2005) The spores of Phytophthora: weapons of the plant destroyer. Nat. 1017 Rev. Microbiol. 3, 47-58. 1018
Judelson, H.S., Tyler, B.M. and Michelmore, R.W. (1991) Transformation of the oomycete pathogen, 1019 Phytophthora infestans. Mol. Plant–Microbe Interact. 4, 602-607. 1020
Judelson, H.S. and Whittaker, S.L. (1995) Inactivation of transgenes in Phytophthora infestans is not 1021 associated with their deletion, methylation, or mutation. Curr. Genet. 28, 571-579. 1022
Jung, T., Colquhoun, I.J. and Hardy, G. E. St. J. (2013) New insights into the survival strategy of the 1023 invasive soilborne pathogen Phytophthora cinnamomi in different natural ecosystems in Western 1024 Australia. Forest Pathology 43, 266-288 1025
Jupe, J., Stam, R., Howden, A. J., Morris, J. A., Zhang, R., Hedley, P. E. and Huitema, E. (2013) 1026 Phytophthora capsici-tomato interaction features dramatic shifts in gene expression associated with a 1027 hemi-biotrophic lifestyle. Genome biology, 14, R63. 1028
Kale, S. D., Gu, B., Capelluto, D. G. S., Dou, D.-L., Feldman, E., Rumore, A., et al. (2010) External lipid 1029 PI-3-P mediates entry of eukaryotic pathogen effectors into plant and animal host cells. Cell, 142, 284–1030 295. 1031
Kamoun, S. (2006) A catalogue of the effector secretome of plant pathogenic oomycetes. Annu. Rev. 1032 Phytopathol. 44, 41-60. 1033
Kamoun, S., Hraber, P., Sobral, B., Nuss, D. and Govers, F. (1999) Initial assessment of gene diversity 1034 for the oomycete pathogen Phytophthora infestans based on expressed sequences. Fungal Genet. 1035 Biol. 28, 94-106. 1036
Kasuga, T., Kozanitas, M., Bui, M., Hüberli, D., Rizzo, D. M., and Garbelotto, M. 2012. Phenotypic 1037 diversification is associated with host-induced transposon derepression in the sudden oak death 1038 pathogen Phytophthora ramorum. PLoS One. 7:e34728 1039
Kaufmann, M. J. and Gerdemann, J. W. (1958) Root and stem rot of soybeans caused by Phytophthora 1040 sojae n. sp. . Phytopathology, 48, 201-208. 1041
Kaur, Parwinder, K. Sivasithamparam, and M. J. Barbetti. (2008). Pathogenic behaviour of strains of 1042 Albugo candida from Brassica juncea (Indian mustard) and Raphanus raphanistrum (wild radish) in 1043 Western Australia. Australasian Plant Pathology 37:353-356. 1044
Kemen, Eric, Torsten Schultz-Larsen, Ariane C. Kemen, Alexi Balmuth, Alexandre Robert-1045 Seilaniantz, Kate Bailey, Eric Holub, David Studholme, Dan MacLean, and Jonathan D. G. Jones. 1046 (2011). Gene gain and loss during evolution of obligate parasitism in the white rust pathogen of 1047 Arabidopsis. PLoS Biol 9:e1001094. 1048
Kennelly, M. M., Gadoury, D. M., Wilcox, W. F., Magarey, P. A. and Seem, R. C. (2007) Primary 1049 infection, lesion productivity, and survival of sporangia in the grapevine downy mildew pathogen 1050 Plasmopara viticola. Phytopathology, 97, 512-522. 1051 Acc
epte
d A
rticl
e

Page 22
This article is protected by copyright. All rights reserved. 22
Kida, K., Tojo, M., Yano, K. and Kotani, S. (2006) First report of Pythium ultimum var. ultimum causing 1052 damping-off on okra in Japan. Plant Pathol. 56, 1042-1042. 1053
Krings, M., Taylor, T.N. and Dotzler, N. (2011) The fossil record of the Peronosporomycetes (Oomycota). 1054 Mycologia, 103, 455-7. 1055
Lalancette, N., Madden, L. V. and Ellis, M. A. (1987) A Model for Predicting the Sporulation of Plasmopara 1056 viticola Based on Temperature and Duration of High Relative-Humidity. Phytopathology, 77, 1699-1699. 1057
Lamour, K. H. and Hausbeck, M. K. (2000) Mefenoxam insensitivity and the sexual stage of Phytophthora 1058 capsici in Michigan cucurbit fields. Phytopathology, 90, 396-400. 1059
Lamour, K. H. and Hu, J. (2013) Diversity and Phytophthora: a threat to forests, crops and traditional 1060 laboratory research - Mini Review. CAB Reviews, 8. 1061
Lamour K. H. Kamoun, S. (2009) Oomycete Genetics and Genomics: Diversity, Interactions, and Research 1062 Tools. New Jersey: John Wiley & Sons. 1063
Lamour, K. H., Mudge, J., Gobena, D., Hurtado-Gonzales, O. P., Schmutz, J., Kuo, A., Miller, N. A., 1064 Rice, B. J., Raffaele, S., Cano, L. M., Bharti, A. K., Donahoo, R. S., Finley, S., Huitema, E., Hulvey, 1065 J., Platt, D., Salamov, A., Savidor, A., Sharma, R., Stam, R., Storey, D., Thines, M., Win, J., Haas, 1066 B. J., Dinwiddie, D. L., Jenkins, J., Knight, J. R., Affourtit, J. P., Han, C. S., Chertkov, O., 1067 Lindquist, E. A., Detter, C., Grigoriev, I. V., Kamoun, S. and Kingsmore, S. F. (2012) Genome 1068 sequencing and mapping reveal loss of heterozygosity as a mechanism for rapid adaptation in the 1069 vegetable pathogen Phytophthora capsici. Mol Plant Microbe Interact, 25, 1350-1360. 1070
Lapin, D. and Van den Ackerveken, G. (2013) Susceptibility to plant disease: more than a failure of host 1071 immunity. Trends Plant Sci. 1072
Leonian, L. H. (1922) Stem and fruit blight of peppers caused by Phytophthora capsici sp. nov. 1073 Phytopathology, 12, 401-408. 1074
Levesque, C. A., Brouwer, H., Cano, L., Hamilton, J. P., Holt, C., Huitema, E., et al. (2010) Genome 1075 sequence of the necrotrophic plant pathogen Pythium ultimum reveals original pathogenicity 1076 mechanisms and effector repertoire. Genome Biol. 11, R73. 1077
Li, C., Wang, J., Chien, D., Chujoy, E., Song, B. and VanderZaag, P. (2011) Cooperation-88: A high 1078 yielding, multi-purpose, late blight resistant cultivar growing in southwest China. Am. J. Pot. Res. 88, 1079 190-194. 1080
Links, M. G., E. Holub, R. H. Jiang, A. G. Sharpe, D. Hegedus, E. Beynon, D. Sillito, W. E. Clarke, S. 1081 Uzuhashi, and M. H. Borhan. 2011. De novo sequence assembly of Albugo candida reveals a small 1082 genome relative to other biotrophic oomycetes. BMC Genomics no. 12:503. doi: 10.1186/1471-2164-1083 12-503. 1084
Liu, Z., Bos, J.I., Armstrong, M., Whisson, S.C., da Cunha, L., Torto-Alalibo, T., Win, J., Avrova, A.O., 1085 Wright, F., Birch, P.R. and Kamoun, S. (2005) Patterns of diversifying selection in the phytotoxin-like 1086 scr74 gene family of Phytophthora infestans. Mol. Biol. Evol. 22, 659-672. 1087
Lucas, B. and Griffiths, P. D. (2004) Evaluation of common bean accessions for resistance to Pythium 1088 ultimum. Hortscience 39, 1193-1195. 1089
Lucas, J., Hayter, J. and Crute, I. (1995) The Downy Mildews: Host Specificity and Pathogenesis. In: 1090 Pathogenesis and Host Specificity in Plant Diseases. (Singh, U. and Singh, R., eds.). UK: Permagon, 1091 pp. 217-234. 1092
Madden, L. V., Ellis, M. A., Lalancette, N., Hughes, G. and Wilson, L. L. (2000) Evaluation of a disease 1093 warning system for downy mildew of grapes. Plant Dis, 84, 549-554. 1094
Madden, L. V., Hughes, G. and Ellis, M. A. (1995) Spatial Heterogeneity of the Incidence of Grape Downy 1095 Mildew. Phytopathology, 85, 269-275. 1096
Mallard, S., Cantet, M., Massire, A., Bachellez, A., Ewert, S. and Lefebvre, V. (2013) A key QTL cluster 1097 is conserved among accessions and exhibits broad-spectrum resistance to Phytophthora capsici: a 1098 valuable locus for pepper breeding. Mol Breeding, 32, 349-364. 1099
Mammella, M. A., Cacciola, S. O., Martin, F. and Schena, L. (2011) Genetic characterization of 1100 Phytophthora nicotianae by the analysis of polymorphic regions of the mitochondrial DNA. Fungal Biol., 1101 115, 432-442. 1102 Acc
epte
d A
rticl
e

Page 23
This article is protected by copyright. All rights reserved. 23
Mammella, M. A., Martin, F. N., Cacciola, S. O., Coffey, M. D., Faedda, R. and Schena, L. (2013) 1103 Analyses of the population structure in a global collection of Phytophthora nicotianae isolates inferred 1104 from mitochondrial and nuclear DNA sequences. Phytopathology, 103, 610-622. 1105
Mansfield, J., Genin, S., Magori, S., Citovsky, V., Sriariyanum, M., Ronald, P., Dow, M., Verdier, V., 1106 Beer, S.V., Machado, M.A., Toth, I., Salmond, G. and Foster, G.D. (2012) Top 10 plant pathogenic 1107 bacteria in molecular plant pathology. Mol. Plant Pathol. 13, 614–629. 1108
Martin, F. N. (1995) Electrophoretic karyotype polymorphisms in the genus Pythium. Mycologia, 87, 333-1109 353. 1110
Martin, F. N. and Loper, J. E. (1999) Soilborne plant diseases caused by Pythium spp: Ecology, 1111 epidemiology, and prospects for biological control. Crit. Rev. Plant Sci. 18, 111-181. 1112
Martin, M.D., Cappellini, E., Samaniego, J.A., Zepeda, M.L., Campos, P.F., Seguin-Orlando, A., Wales, 1113 N., Orlando, L., Ho, S.Y.W., Dietrich, F.S., Mieczkowski, P.A., Heitman, J., Willerslev, E., Krogh, 1114 A., Ristaino, J.B. and Gilbert, M.T.P. (2013) Reconstructing genome evolution in historic samples of 1115 the Irish potato famine pathogen. Nat. Commun. 4, 2172. 1116
Martin, M.D., Ho, S.Y,, Wales, N., Ristaino, J.B. and Gilbert, M.T.P. (2014) Persistence of the 1117 mitochondrial lineage responsible for the Irish potato famine in extant new world Phytophthora 1118 infestans. Mol Biol Evol 31, 1414-20. 1119
Matheron, M. E. and Matejka, J. C. (1990) Differential virulence of Phytophthora parasitica recovered from 1120 Citrus and other plants to rough lemon and tomato. Plant Dis., 74, 138-140. 1121
McDowell, J. M. (2011) Genomes of obligate plant pathogens reveal adaptations for obligate parasitism. 1122 Proc Natl Acad Sci U S A, 108, 8921-8922. 1123
Meijer, H.J.G., Hua, C., Kots, K., Ketelaar, T. and Govers, F. (2014) Actin dynamics in Phytophthora 1124 infestans; rapidly reorganizing cables and immobile, long-lived plaques. Cell. Microbiol., 16. 948-961. 1125
Meitz, J. C., Linde, C. C., Thompson, A., Langenhoven, S. and McLeod, A. (2010) Phytophthora capsici 1126 on vegetable hosts in South Africa: distribution, host range and genetic diversity. Australas Plant 1127 Pathol, 39, 431-439. 1128
Millardet, A. (1881) Notes sur les vignes américaines et opuscules divers sur le même sujet. Bordeaux. 1129 Moralejo, E. P.-S., A. M., Alvarez, L. A., Belbahri, L., Lefort, F. and Descals, E. (2009) Multiple alien 1130
Phytophthora taxa discovered on diseases ornamental plants in Spain. Plant Pathol., 58, 100-110. 1131 Morris, P. F. and Ward, E. W. B. (1992) Chemoattraction of zoospores of the soybean pathogen 1132
phytophthora-sojae by isoflavones. Physiol. Molec. Plant Pathol., 40, 17-22. 1133 Mukhtar, M. S., Carvunis, A. R., Dreze, M., Epple, P., Steinbrenner, J., Moore, J., et al. (2011) 1134
Independently evolved virulence effectors converge onto hubs in a plant immune system network. 1135 Science, 333, 596-601. 1136
Nadiminti, P., Dong Y., Sayer C., Hay P., Rookes J.E., Boyd B. and Cahill D.M. (2013) Nanostructured 1137 liquid crystalline particles as an alternative delivery vehicle for plant agrochemicals. ACS Advanced 1138 Materials and Interfaces 5, 1818-1826 1139
Naseby, D. C., Pascual, J. A. and Lynch, J. M. (2000) Effect of biocontrol strains of Trichoderma on plant 1140 growth, Pythium ultimum populations, soil microbial communities and soil enzyme activities. J. Appl. 1141 Microbiol. 88, 161-169. 1142
Olson, H. A. and Benson, D. M. (2011) Characterization of Phytophthora spp. on floriculture crops in North 1143 Carolina. Plant Dis., 95, 1013-1020. 1144
Olson, H. A., Jeffers, S. N., Ivors, K. L., Steddom, K. C., Williams-Woodward, J. L., Mmbaga, M. T., et 1145 al. (2013) Diversity and mefenoxam sensitivity of Phytophthora spp. associated with the ornamental 1146 horticulture industry in the Southeastern United States. Plant Dis., 97, 86-92. 1147
Pais, M., Win, J., Yoshida, K., Etherington, G.J., Cano, L.M., Raffaele, S., Banfield, M.J., Jones, A., 1148 Kamoun, S. and Saunders, D.G.O. (2013) From pathogen genomes to host plant processes: the 1149 power of plant parasitic oomycetes. Genome Biol 14, 211. 1150
Peressotti, E., Wiedemann-Merdinoglu, S., Delmotte, F., Bellin, D., Di Gaspero, G., Testolin, R., et al. 1151 (2010) Breakdown of resistance to grapevine downy mildew upon limited deployment of a resistant 1152 variety. Bmc Plant Biology, 10. 1153 Acc
epte
d A
rticl
e

Page 24
This article is protected by copyright. All rights reserved. 24
Peters, R.D., Al-Mughrabi, K.I., Kalischuk, M.L., Dobinson, K., Conn, K., et al. (2014) Migration and 1154 recombination increases Phytophthora infestans population diversity and independent segregation of 1155 genotype specific loci. Can J Plant Pathol: in press. 1156
Petrie, G. A. 1975. Prevalence of oospores of Albugo cruciferarum in Brassica seed samples from western 1157 Canada 1967-1973. Canadian Plant Disease Survey no. 55 (1):19-24. 1158
Pieterse, C.M.J., Risseeuw, E.P. and Davidse, L.C. (1991) An in planta induced gene of Phytophthora 1159 infestans codes for ubiquitin. Plant Mol. Biol. 17, 799-811. 1160
Ploch, S., Y. J. Choi, C. Rost, H. D. Shin, E. Schilling, and M. Thines. 2010. Evolution of diversity in 1161 Albugo is driven by high host specificity and multiple speciation events on closely related Brassicaceae. 1162 Molecular Phylogenetics and Evolution no. 57 (2):812-820. doi: 10.1016/j.ympev.2010.06.026. 1163
Quesada-Ocampo, L. M. and Hausbeck, M. K. (2010) Resistance in tomato and wild relatives to crown 1164 and root rot caused by Phytophthora capsici. Phytopathology, 100, 619-627. 1165
Qutob, D., Hraber, P., Sobral, B. and Gijzen, M. (2000) Comparative analysis of expressed sequences in 1166 Phytophthora sojae. Plant Phys., 123, 243-253. 1167
Qutob, D., Kemmerling, B., Brunner, F., Kufner, I., Engelhardt, S., Gust, A. A., et al. (2006) 1168 Phytotoxicity and innate immune responses induced by Nep1-like proteins. Plant Cell, 18, 3721-3744. 1169
Qutob, D., Patrick Chapman, B. and Gijzen, M. (2013) Transgenerational gene silencing causes gain of 1170 virulence in a plant pathogen. Nat Commun, 4, 1349. 1171
Raffaele, S., Farrer, R.A., Cano, L.M., Studholme, D.J., MacLean, D., Thines, M., Jiang, R.H.Y., Zody, 1172 M.C., Kunjeti, S.G., Donofrio, N.M., Meyers, B.C., Nusbaum, C., and Kamoun, S. (2010). Genome 1173 evolution following host jumps in the Irish potato famine pathogen lineage. Science, 330,1540-1543. 1174
Randall, E., Young, V., Sierotzki, H., Scalliet, G., Birch, P.R.J., Cooke, D.E.L., Csukai, M. and 1175 Whisson, S.C. (2014) Sequence diversity in the large subunit of RNA polymerase I contributes to 1176 Mefenoxam insensitivity in Phytophthora infestans. Mol. Plant Pathol., Feb 2014. doi: 1177 2010.1111/mpp.12124. 1178
Reeksting B.J., Coetzer, N., Mahomed, W., Engelbrecht, J. and van den Berg, N. (2014) De Novo 1179 sequencing, assembly, and analysis of the root transcriptome of Persea americana (Mill.) in response 1180 to Phytophthora cinnamomi and flooding. PloS One Published: February 10, 2014, DOI: 1181 10.1371/journal.pone.0086399 1182
Rehmany, A.P., Gordon, A., Rose, L.E., Allen, A.E., Armstrong, M.R., Whisson, S.C., Kamoun, S., 1183 Tyler, B.M., Birch, P.R. and Beynon, J.L. (2005) Differential recognition of highly divergent downy 1184 mildew avirulence gene alleles by RPP1 resistance genes from two Arabidopsis lines. Plant Cell, 17, 1185 1839-1850. 1186
Rimmer, S. R., S. Mathur, and C. R. Wu. 2000. Virulence of isolates of Albugo candida from western 1187 Canada to Brassica species. Canadian Journal of Plant Pathology 22:235. 1188
Rizzo, D. M., Garbelotto, M., and Hansen, E. M. 2005. Phytophthora ramorum: Integrative research and 1189 management of an emerging pathogen in California and Oregon forests. Annu. Rev. Phytopathol. 1190 43:309–335 1191
Rodewald, J. and Trognitz, B. (2013) Solanum resistance genes against Phytophthora infestans and their 1192 corresponding avirulence genes. Mol. Plant Pathol. 14, 740-757. 1193
Rookes, J.E., Wright, M.L. and Cahill D.M. (2008) Elucidation of defence responses and signalling 1194 pathways induced in Arabidopsis thaliana following challenge with Phytophthora cinnamomi 1195 Physiological and Molecular Plant Pathology 72, 151–161 1196
Rossi, V. and Caffi, T. (2012) The Role of Rain in Dispersal of the Primary Inoculum of Plasmopara viticola. 1197 Phytopathology, 102, 158-165. 1198
Rossi, V., Caffi, T. and Gobbin, D. (2013) Contribution of molecular studies to botanical epidemiology and 1199 disease modelling: grapevine downy mildew as a case-study. Eur J Plant Pathol, 135, 641-654. 1200
Rossi, V., Giosue, S. and Caffi, T. (2009) Modelling the Dynamics of Infections Caused by Sexual and 1201 Asexual Spores during Plasmopara viticola Epidemics. J Plant Pathol, 91, 615-627. 1202 Acc
epte
d A
rticl
e

Page 25
This article is protected by copyright. All rights reserved. 25
Rouxel, M., Mestre, P., Comont, G., Lehman, B., Schilder, A. and Delmotte, F. (2013) Phylogenetic and 1203 experimental evidence for host-specialized cryptic species in a biotrophic oomycete. New Phytologist, 1204 197, 251-263. 1205
Roy, S., Poidevin, L., Jiang, T. and Judelson, H. S. (2013) Novel core promoter elements in the oomycete 1206 pathogen Phytophthora infestans and their influence on expression detected by genome-wide analysis. 1207 BMC Genom. 14, 106. 1208
Saadoun, M. and Allagui, M. B. (2008) Pathogenic variability of Phytophthora nicotianae on pepper in 1209 tunisia. J. Plant Pathol., 90, 351-355. 1210
Scholthof, K.B.G., Adkins, S., Czosnek, H., Palukaitis, P., Jacquot, E., Hohn, T., Hohn, B., Saunders, 1211 K., Candresse, T., Ahlquist, P., Hemenway, C. and Foster, G.D. (2011) Top 10 plant viruses in 1212 molecular plant pathology. Mol. Plant Pathol. 12, 938–954. 1213
Schornack S, Huitema E, Cano LM, Bozkurt TO, Oliva R, et al. (2009) Ten things to know about 1214 oomycete effectors. Mol Plant Pathol, 10: 795–803. 1215
Schornack, S., van Damme, M., Bozkurt, T.O., Cano, L.M., Smoker, M., Thines, M., Gaulin, E., 1216 Kamoun, S. and Huitema, E. (2010) Ancient class of translocated oomycete effectors targets the host 1217 nucleus. Proc. Natl. Acad. Sci. USA, 107, 17421-17426. 1218
Schroeder, K. L., Martin, F. N., de Cock, A. W. A. M., Levesque, C. A., Spies, C. F. J., Okubara, P. A., 1219 et al. (2013) Molecular detection and quantification of Pythium species: evolving taxonomy, new tools, 1220 and challenges. Plant Dis. 97, 4-20. 1221
Seidl, M.F., Van den Ackerveken, G., Govers, F. and Snel, B. (2011) A domain-centric analysis of 1222 oomycete plant pathogen genomes reveals unique protein organization. Plant Physiol. 155, 628-644. 1223
Serrano, M.S., Fernández-Rebollo, P., De Vita, P. and Esperanza Sánchez M. (2013) Calcium mineral 1224 nutrition increases the tolerance of Quercus ilex to Phytophthora root disease affecting oak rangeland 1225 ecosystems in Spain. Agroforestry Systems 87, 173-179. 1226
Shaw, D.S. (1983) The perenosporales: a fungal geneticist’s nightmare. In: Zoosporic Plant Pathogens 1227 (Buczacki, S.T., ed.), pp. 85–121. London: Academic Press. 1228
Song, J., Win, J., Tian, M., Schornack, S., Kaschani, F., Ilyas, M., van der Hoorn, R.A.L., and Kamoun, 1229 S. (2009). Two effectors secreted by unrelated eukaryotic plant pathogens target the tomato defense 1230 protease Rcr3. Proc. Natl. Acad. Sci. USA, 106,1654-1659. 1231
Soylu, S., M. Keshavarzi, I. Brown, and J. W. Mansfield. 2003. Ultrastructural characterisation of 1232 interactions between Arabidopsis thaliana and Albugo candida. Physiological and Molecular Plant 1233 Pathology no. 63 (4):201-211. doi: 10.1016/j.pmpp.2003.12.002. 1234
Stam, R., Howden, A. J., Delgado-Cerezo, M., TM, M. M. A., Motion, G. B., Pham, J. and Huitema, E. 1235 (2013a) Characterization of cell death inducing Phytophthora capsici CRN effectors suggests diverse 1236 activities in the host nucleus. Frontiers in plant science, 4, 387. 1237
Stam, R., Jupe, J., Howden, A. J., Morris, J. A., Boevink, P. C., Hedley, P. E. and Huitema, E. (2013b) 1238 Identification and characterisation CRN effectors in Phytophthora capsici shows modularity and 1239 functional diversity. PloS one, 8, e59517. 1240
Stanghellini, M. E. and Hancock, J. G. (1971) Sporangium of Pythium ultimum as survival structure in soil. 1241 Phytopathology, 61, 157-164. 1242
Stredansky, M., Conti, E. and Salaris, A. (2000) Production of polyunsaturated fatty acids by Pythium 1243 ultimum in solid-state cultivation. Enzyme Microb. Tech. 26, 304-307. 1244
Strullu-Derrien, C., Kenrick, P., Rioult, J.P. and Strullu, D.G. (2011) Evidence of parasitic Oomycetes 1245 (Peronosporomycetes) infecting the stem cortex of the Carboniferous seed fern Lyginopteris oldhamia. 1246 Proc Royal Soc B, 278,675-680 1247
Sun, W., Jia, Y., O'Neill, N., Feng, B. and Zhang, X. (2008) Genetic diversity in Phytophthora capsici from 1248 eastern China. Can J Plant Pathol, 30, 414-424. 1249
Sun, W. X., Jia, Y. J., Feng, B. Z., O'Neill, N. R., Zhu, X. P., Xie, B. Y. and Zhang, X. G. (2009) Functional 1250 analysis of Pcipg2 From the straminopilous plant pathogen Phytophthora capsici. Genesis, 47, 535-1251 544. 1252 Acc
epte
d A
rticl
e

Page 26
This article is protected by copyright. All rights reserved. 26
Tan, M.Y.A., Hutten, R.C.B., Visser, R.G.F. and Eck, H.J. (2010) The effect of pyramiding Phytophthora 1253 infestans resistance genes RPi-mcd1 and RPi-ber in potato. Theor. Appl. Genet. 121, 117-125. 1254
Taylor, R. J., Pasche, J. S., Gallup, C. A., Shew, H. D. and Gudmestad, N. C. (2008) A foliar blight and 1255 tuber rot of potato caused by Phytophthora nicotianae: new occurrences and characterization of 1256 isolates. Plant Dis., 92, 492-503. 1257
Thines, M. (2014) Phylogeny and evolution of plant pathogenic oomycetes‚ a global overview. Eur J Plant 1258 Pathol, 1-17. 1259
Thines, M., Y. J. Choi, E. Kemen, S. Ploch, E. B. Holub, H. D. Shin, and J. D. Jones. 2009. A new 1260 species of Albugo parasitic to Arabidopsis thaliana reveals new evolutionary patterns in white blister 1261 rusts (Albuginaceae). Persoonia no. 22:123-8. doi: 10.3767/003158509X457931. 1262
Thines, M., and S. Kamoun. 2010. Oomycete-plant coevolution: recent advances and future prospects. 1263 Curr Opin Plant Biol no. 13 (4):427-33. doi: S1369-5266(10)00023-3 [pii] 1264
10.1016/j.pbi.2010.04.001. 1265 Thines, M., and O. Spring. 2005. A revision of Albugo (chromista, peronosporomycetes). Mycotaxon no. 1266
92:443-458. 1267 Thompson, S.E., Levin, S. and Rodriguez-Iturbe, I. (2104) Rainfall and temperatures changes have 1268
confounding impacts on Phytophthora cinnamomi occurrence risk in the southwestern USA under 1269 climate change scenarios. Global Change Biology 20, 1299-1312 1270
Tian, M., Huitema, E., Da Cunha, L., Torto-Alalibo, T. and Kamoun, S. (2004) A Kazal-like extracellular 1271 serine protease inhibitor from Phytophthora infestans targets the tomato pathogenesis-related protease 1272 P69B. J. Biol. Chem. 279, 26370-26377. 1273
Töpfer, R., Hausmann, L. and Eibach, R. (2011) Molecular breeding, in: Adam-Blondon AF, Martinez-1274 Zapater JM, Kole C (eds) Genetics, genomics and breeding of grapes. Science Publishers, Enfield, pp 1275 160–185. 1276
Torto-Alalibo, T., Tripathy, S., Smith, B. M., Arredondo, F., Zhou, L., Li, H., et al. (2007) Expressed 1277 sequence tags from Phytophthora sojae reveal genes specific to development and infection. Mol. Plant-1278 Microbe Interact., 20, 781-793. 1279
Torto, T.A., Li, S., Styer, A., Huitema, E., Testa, A., Gow, N.A., van West, P. and Kamoun, S. (2003) 1280 EST mining and functional expression assays identify extracellular effector proteins from the plant 1281 pathogen Phytophthora. Genome Res. 13, 1675-1685. 1282
Tyler, B. M. (2007) Phytophthora sojae: root rot pathogen of soybean and model oomycete. Molec. Plant 1283 Pathol., 8, 1-8. 1284
Tyler, B. M., Tripathy, S., Zhang, X., Dehal, P., Jiang, R. H. Y., Aerts, A., Arredondo, F. D., Baxter, L., 1285 Bensasson, D., Beynon, J. L., Chapman, J., Damasceno, C. M. B., Dorrance, A. E., Dou, D., 1286 Dickerman, A. W., Dubchak, I. L., Garbelotto, M., Gijzen, M., Gordon, S. G., Govers, F., Grünwald, 1287 N. J., Huang, W., Ivors, K. L., Jones, R. W., Kamoun, S., Krampis, K., Lamour, K. H., Lee, M.-K., 1288 McDonald, W. H., Medina, M., Meijer, H. J. G., Nordberg, E. K., Maclean, D. J., Ospina-Giraldo, M. 1289 D., Morris, P. F., Phuntumart, V., Putnam, N. H., Rash, S., Rose, J. K. C., Sakihama, Y., Salamov, 1290 A. A., Savidor, A., Scheuring, C. F., Smith, B. M., Sobral, B. W. S., Terry, A., Torto-Alalibo, T. A., 1291 Win, J., Xu, Z., Zhang, H., Grigoriev, I. V, Rokhsar, D. S., and Boore, J. L. 2006. Phytophthora 1292 genome sequences uncover evolutionary origins and mechanisms of pathogenesis. Science (80-. ). 1293 313:1261–1266 1294
Uzuhashi, S., Tojo, M. and Kakishima, M. (2010) Phylogeny of the genus Pythium and description of new 1295 genera. Mycoscience 51, 337-365. 1296
Vallance, J., Deniel, F., Barbier, G., Guerin-Dubrana, L., Benhamou, N. and Rey, P. (2012) Influence of 1297 Pythium oligandrum on the bacterial communities that colonize the nutrient solutions and the 1298 rhizosphere of tomato plants. Can. J. Microbiol. 58, 1124-1134. 1299
Van der Lee, T., De Witte, I., Drenth, A., Alfonso, C. and Govers, F. (1997) AFLP linkage map of the 1300 oomycete Phytophthora infestans. Fungal Genet. Biol. 21, 278-291. 1301 Acc
epte
d A
rticl
e

Page 27
This article is protected by copyright. All rights reserved. 27
Van Poucke, K., Franceschini, S., Webber, J. F., Vercauteren, A., Turner, J. A., McCracken, A. R., 1302 Heungens, K., and Brasier, C. M. 2012. Discovery of a fourth evolutionary lineage of Phytophthora 1303 ramorum: EU2. Fungal Biol. 116:1178–91 1304
Van West, P., Kamoun, S., van 't Klooster, J.W. and Govers, F. (1999) Internuclear gene silencing in 1305 Phytophthora infestans. Mol. Cell, 3, 339-348. 1306
Vercesi, A., Toffolatti, S. L., Zocchi, G., Guglielmann, R. and Ironi, L. (2010) A new approach to 1307 modelling the dynamics of oospore germination in Plasmopara viticola. Eur J Plant Pathol, 128, 113-1308 126. 1309
Viennot-Bourgin, G. (1949) Les champignons parasites des plantes cultivées. Paris, France: Librairies de 1310 l’Académie de Médecine. 1311
Vleeshouwers, V.G.A.A., Raffaele, S., Vossen, J., Champouret, N., Oliva, R., Segretin, M.E., Rietman, 1312 H., Cano, L., Lokossou, A.A., Kessel, G., Pel, M.A. and Kamoun, S. (2011) Understanding and 1313 exploiting late blight resistance in the age of effectors. Annu. Rev. Phytopathol. 49, 507-531. 1314
Vleeshouwers, V.G.A.A., Rietman, H., Krenek, P., Champouret, N., Young, C., Oh, S.-K., Wang, M.C., 1315 Bouwmeester, K., Vosman, B., Visser, R.G.F., Jacobsen, E., Govers, F., Kamoun, S. and Van der 1316 Vossen, E.A.G. (2008) Effector genomics accelerates discovery and functional profiling of potato 1317 disease resistance and Phytophthora infestans avirulence genes. PLoS ONE, 3, e2875. 1318
Wang, H. P. and Davis, R. M. (1997) Susceptibility of selected cotton cultivars to seedling disease 1319 pathogens and benefits of chemical seed treatments. Plant Dis. 81, 1085-1088. 1320
Wang, Q., Han, C., Ferreira, A. O., Yu, X., Ye, W., Tripathy, S., et al. (2011) Transcriptional programming 1321 and functional interactions within the Phytophthora sojae RXLR effector repertoire. Plant Cell, 23, 2064-1322 2086. 1323
Weiland, J. J. (2003) Transformation of Pythium aphanidermatum to geneticin resistance. Curr. Genet. 42, 1324 344-352. 1325
Werres, S., Marwitz, R., Veld, W., De Cock, A., Bonants, P. J. M., De Weerdt, M., Themann, K., Ilieva, 1326 E., and Baayen, R. P. 2001. Phytophthora ramorum sp. nov., a new pathogen on Rhododendron and 1327 Viburnum. Mycol. Res. 105:1155–1165 1328
Whisson, S.C., Boevink, P.C., Moleleki, L., Avrova, A.O., Morales, J.G., Gilroy, E.M., Armstrong, M.R., 1329 Grouffaud, S., van West, P., Chapman, S., Hein, I., Toth, I.K., Pritchard, L. and Birch, P.R.J. (2007) 1330 A translocation signal for delivery of oomycete effector proteins into host plant cells. Nature, 450, 115-1331 118. 1332
Wirthmueller, L., Maqbool, A. and Banfield, M.J. (2013) On the front line: structural insights into plant-1333 pathogen interactions. Nat. Rev. Microbiol. 11, 761-776. 1334
Wong, F., Burr, H. and Wilcox, W. (2001) Heterothallism in Plasmopara viticola. Plant Pathology, 50, 427-1335 432. 1336
Wrather, J. A. and Koenning, S. R. (2006) Estimates of disease effects on soybean yields in the United 1337 States 2003 to 2005. J. Nematology, 38, 173-180. 1338
Yoshida, K., Burbano, H.A., Krause, J., Thines, M., Weigel, D. and Kamoun S. (2014) Mining herbaria 1339 for plant pathogen genomes: back to the future. PLoS Pathog,10, e1004028. 1340
Yoshida, K., Schuenemann, V., Cano, L., Pais, M., Mishra, B., Sharma, R., Kamoun, S., Krause, J., 1341 Thines, M., Weigel, D. and Burbano, H. (2013) The rise and fall of the Phytophthora infestans lineage 1342 that triggered the Irish potato famine. eLife 2, e00731. 1343
Zadoks, J.C. (2008) The potato murrain on the European continent and the revolutions of 1848. Potato 1344 Res. 51, 5-45. 1345
Zerillo, M. M., Adhikari, B. N., Hamilton, J. P., Buell, C. R., Levesque, C. A. and Tisserat, N. (2013) 1346 Carbohydrate-active enzymes in Pythium and their role in plant cell wall and storage polysaccharide 1347 degradation. PloS One 8, e72572. 1348
1349 1350 A
ccep
ted
Arti
cle

Page 28
This article is protected by copyright. All rights reserved. 28
FIGURE LEGENDS 1351
1352
Figure 1. Potato plants with typical late blight lesions. Infection starts when a spore lands on the leaf and 1353 germinates. The germ tube forms an appressorium and an emerging penetration peg pushes into an 1354 epidermal cell. Then the inner cell layers are colonized. During the biotrophic phase, hyphae grow in the 1355 intercellular space while haustoria enter plant cell cavities and invaginate host cell plasma membrane. Later, 1356 P. infestans switches to necrotrophic growth resulting in the death of plant cells and the appearance of 1357 necrotic lesions on the infected tissues. In this phase, hyphae escape through the stomata and produce 1358 numerous asexual spores named sporangia that easily detach and disperse by wind or water. A sporangium 1359 that finds a new host can either germinate directly and initiate a new cycle or, at lower temperatures, 1360 undergo cleavage resulting in a zoosporangium from which 6-8 flagellated spores are released. These 1361 zoospores can swim for several hours but once they touch a solid surface they encyst and germinate to 1362 initiate new infections. Under favourable conditions the pathogen can complete the cycle from infection to 1363 sporulation in 4 days. In the field this cycle is repeated multiple times during one growing season resulting in 1364 billions of spores and a continuous increase of disease pressure. Besides leaves, stems and tubers also get 1365 infected and P. infestans can continue to flourish on the decaying plant material. If not managed properly, 1366 infected seed potatoes or waste on refuse piles are often the sources of inoculum for new infections in 1367 spring. An alternative route for surviving the winter is via oospores, sexual spores that can survive in soil for 1368 many years. P. infestans is heterothallic; isolates are either A1 or A2 mating type and sex organs only 1369 develop when isolates of opposite mating type sense the sex hormone produced by the mate. 1370 1371 Figure 2. One of the many Great Famine memorials around the world. These sculptures on Customs House 1372 Quays in Dublin, by artist Rowan Gillespie, stand as if walking towards the emigration ships on the Dublin 1373 Quayside (courtesy of Michael Seidl). 1374 1375 Figure 3. Hyaloperonospora arabidopsidis disease symptoms on a two-week old Arabidopsis seedling. 1376 Mature sporangiophores are visible as white structures on the right side of the leaf. 1377 1378 Figure 4. Diagram depicting a compatible interaction between H. arabidopsidis and Arabidopsis initiated by 1379 sporangiospore landing on the leaf surface. (S) sporangiospore; (A) appressorium; (N) nucleus; (Hy) 1380 hyphae; (Ha) haustorium; (C) cuticle; (UE) upper epidermis; (PM) palisade mesophyll cells; (SM) spongy 1381 mesophyll cells; (LE) lower epidermis; (Sp) mature sporangiophore. Note: Sporangiophore not drawn to 1382 scale. 1383 1384 Figure 5. Impact of sudden oak death in California. Tanoak mortality evidenced by defoliated or wilted 1385 canopies on the Bolinas Ridge at Mt. Tamalpais, Marin County, California. Photo courtesy of Janet Klein 1386 (Marin Municipal Open Space District). 1387 1388 Figure 6. Inferred pattern of migration of the four clonal lineages of Phytophthora ramorum. Modified from 1389 Grünwald et al. (2012). 1390 1391 Figure 7. Silencing machinery in Phytophthora. (A). Phylogenetic placement of dicer-like (DCL) and (B). 1392 argonaute (Ago) proteins in the genus Phytophthora. See Fahlgren et al. (2013) for more details. Species 1393 correspond to: Arabidopsis thaliana, Paramecium tetraurelia, Phytophthora infestans, Phytophthora 1394 ramorum, Phytophthora sojae, Tetrahymena thermophila, and Toxoplasma gondii. 1395 1396 Figure 8. Phytophthora sojae. A, Diseased soybean plants in the field, infected with P. sojae. Plant height is 1397 20 to 30 cm. B, Susceptible (left) and resistant (right) soybean plants inoculated in the stem with P. sojae, 1398 seven days after infection, illustrating R-gene mediated resistance. Pots are 10 cm in diameter. C, 1399 Germinating oospore of P. sojae growing on water agar. Oospore is 35 µm in diameter. D, Germinating P. 1400 A
ccep
ted
Arti
cle

Page 29
This article is protected by copyright. All rights reserved. 29
sojae cysts growing on water agar. Cysts are 15 µM in diameter. E, Etiolated soybean hypocotyls 1401 inoculated with a 10 µL water droplet containing 103 zoospores from a virulent (upper) or avirulent (lower) 1402 strain of P. sojae, 48 h after infection, illustrating strain specific variation in avirulence effectors and the 1403 hypersensitive response. Soybean hypocotyls are 5 mm in diameter. 1404 1405 Figure 9. Large diverse families of virulence proteins encoded by the P. sojae genome. (A) P. sojae 1406 genome contains clusters of conserved housekeeping genes (brown) that have conserved orders among 1407 Phytophthora species, separated by dynamic transposon-rich regions that contain genes (red) encoding 1408 virulence proteins, many of which are secreted. (B) Secreted virulence proteins (effectors) may act in the 1409 apoplast, or be transported inside the cell. Cell-entering effectors may have targets in the nucleus or 1410 cytoplasm, and may be detected by resistance proteins (Rps; resistance against P. sojae). 1411 1412 Figure 10. Heavy sporulation and spontaneous morphological variation in the vegetable pathogen P. 1413 capsici. (A) Naturally infected tomato fruit with sporangium production on the surface of the fruit and (B) a 1414 single-zoospore derived field isolate of P. capsici sectoring on V8 agar media following long term storage. 1415 1416
Figure 11. Downy mildew symptoms with well evident sporangiophores on the lower side of a grape leaf 1417 (A) and a young cluster (B). 1418
1419 Figure 12. Cryo-SEM micrograph of P. cinnamomi cyst between two epidermal cells on a root of tobacco. 1420 Note the adhesive material that surrounds the cyst that has been expelled by zoospore peripheral vesicles. 1421 Photo courtesy of Adrienne Hardham, Australian National University. 1422 1423 Figure 13. (A) Individual plants of Xanthorrheoa australis (austral grass tree), a highly suscpetible native 1424 Australian species, infected by Phytophthora cinnamomi within a dry sclerophyll eucalypt forest at Anglesea, 1425 Victoria, Australia. Note the dead and dying plants that have brown, collapsed leaves compared with the 1426 healthy green and erect leaves of plants which are yet to be killed. These individual plants range in age from 1427 approximately 20 years (the smallest in the centre) to around 70 years (green individual on the right of the 1428 image). (B) Advancing disease front caused by invasion by P. cinnamomi in Xanthorrhoea australis- 1429 dominated understorey in eucalyptus open forest at Wilsons Promontory, Victoria, Australia. The disease 1430 has moved from the foregound of the picture, where all susceptible vegetation including X. australis has 1431 been killed and its progress can be seen as a line of dead and dying X. australis (brown collapsed plants) at 1432 the disease margin. Healthy green plants behind them will soon be killed. Loss of the major understorey 1433 components, as in the forground, results in complete structural change and loss of all susceptible species. 1434 1435 Figure 14. Impact of Phytophthora parasitica-infection on citrus plants. A and B: five year-old citrus plants 1436 non-infected and infected with P. parasitica respectively; C and D: symptoms of P. parasitica on stems; C: 1437 non-infected, D: infected plant displaying gummosis symptoms; E and F: leaves and fruits of infected (left) 1438 and healthy (right) plants; G and H: scanning electron microscope images of citrus fine roots infected with P. 1439 parasitica. Yellow arrows show encysted zoospores and germ tube of P. parasitica. Bar represents 20 µm. 1440 A- F: photograph courtesy R. J. D. Dalio; G and H: photographs courtesy of M. E. Escanferla. 1441 1442 Figure 15. Severe infection of Brinjal fruit with P. parasitica. Typical symptoms are brown, soft water soaked 1443 patches which rapidly cover the whole fruit. Brinjal is also known as eggplant (Solanum melongena). 1444 Photograph courtesy of S. Guha Roy. 1445 1446 Figure 16. Pythium ultimum var. ultimum. (A) Pre-emergence damping-off in okra, resulting in the death of 1447 most plants in the front of the row. In the back, disease was controlled using metalaxyl. (B) Terminal 1448 hyphal swellings, bar = 10 µm. (C) Oospore within oogonium, bar = 10 µm. Images adapted from Kida et 1449 al. (2006) with permission. 1450 A
ccep
ted
Arti
cle

Page 30
This article is protected by copyright. All rights reserved. 30
1451 Figure 17. Disease symptoms of Albugo infections. (A) Disease resulting from the infection of Brassica 1452 juncea with an unknown Albugo species. The misshapen inflorescence phenotype is known as a 1453 “staghead”. (B) An example of immunity suppression by Albugo laibachii: A. thaliana Col-0 is resistant to H. 1454 arabidopsidis Emoy2 via RPP1, but when pre-infected with Al can support the growth of both pathogens. 1455 1456 Figure 18. Albugo spp. have compact genomes. Synteny between A. laibachii, Py. ultimum, H. 1457 arabidopsidis, and P. infestans. The region shown is an example of the relatively dense clustering of genes 1458 in Albugo species. With increasing genome size the distance between both genes increases and re-1459 organisations occur (red, synteny without inversion; blue, inverted regions). Reproduced from Kemen et al, 1460 (2011). 1461
1462
Acc
epte
d A
rticl
e

Page 31
This article is protected by copyright. All rights reserved. 31
Table 1. Top 10 oomycetes in molecular plant pathology. 1463 1464 Rank species Common disease
name(s) Number of papers (2005-2014)
Number of votes
1 Phytophthora infestans Late blight 1230 51 =2 Hyaloperonospora
arabidopsidis Downy mildew 137 25
=2 Phytophthora ramorum Sudden oak death, Ramorum disease
378 25
4 Phytophthora sojae Stem and root rot 276 22 5 Phytophthora capsici Blight; stem and fruit
rot; various others 541 17
6 Plasmopara viticola Downy mildew 326 15 7 Phytophthora cinnamomi Root rot; dieback 315 13 =8 Phytophthora parasitica Root and stem rot;
various others 142 10
=8 Pythium ultimum Damping off; root rot 319 10 10 Albugo candida White rust 65 9
1465 The “=” sign before the ranking indicates that the species tied for that position. The number of papers 1466 published in 2005-2014 is based on searches of the Scopus database (http://www.scopus.com) using the 1467 species names as a query. For H. arabidopsidis, a search for the alternative name “Peronospora parasitica” 1468 was also performed and the combined number is shown. Searches with the terms “oomycete*” and 1469 “Phytophthora” yielded 2068 and 4059 articles, respectively. 1470 1471 1472
1473
Acc
epte
d A
rticl
e

Page 32
This article is protected by copyright. All rights reserved. 32
1474 Table 2. Other oomycetes species that received votes. 1475 1476 Rank species
11 Aphanomyces euteiches 12 Albugo laibachii 13 Bremia lactucae 14 Phytophthora palmivora 15 Pseudoperonospora cubensis 16 Plasmopara halstedii 17 Peronophythora litchi 18 Peronosclerospora sorghi 19 Peronospora belbahrii 20 Phytophthora alni 21 Phytophthora brassicae 22 Phytophthora cactorum 23 Phytophthora meadii 24 Phytophthora phaseoli 25 Phytophthora plurivora (formerly P. citricola) 26 Plasmopara obducens 27 Pythium aphanidermatum 28 Pythium oligandrum 29 Sclerophthora rayssiae 30 Hyaloperonospora brassicae NR Saprolegnia parasitica (fish parasite) NR Lagenidium giganteum (mosquito parasite) NR Pythium insidiosum (mammalian parasite)
1477 NR, not ranked because the species is not associated to plants. 1478 1479
Acc
epte
d A
rticl
e

Page 33
This article is protected by copyright. All rights reserved. 33
1480
mpp_12190_f1 1481
1482
1483
Acc
epte
d A
rticl
e

Page 34
This article is protected by copyright. All rights reserved. 34
1484
mpp_12190_f2 1485
1486
Acc
epte
d A
rticl
e

Page 35
This article is protected by copyright. All rights reserved. 35
1487
mpp_12190_f3 1488
1489 Acc
epte
d A
rticl
e

Page 36
This article is protected by copyright. All rights reserved. 36
1490
mpp_12190_f4 1491
1492
Acc
epte
d A
rticl
e

Page 37
This article is protected by copyright. All rights reserved. 37
1493
mpp_12190_f5 1494
1495
Acc
epte
d A
rticl
e

Page 38
This article is protected by copyright. All rights reserved. 38
1496
1497
mpp_12190_f6 1498
1499
Acc
epte
d A
rticl
e

Page 39
This article is protected by copyright. All rights reserved. 39
1500
1501
mpp_12190_f7 1502
1503
Acc
epte
d A
rticl
e

Page 40
This article is protected by copyright. All rights reserved. 40
1504
1505
mpp_12190_f8_ 1506
1507
Acc
epte
d A
rticl
e
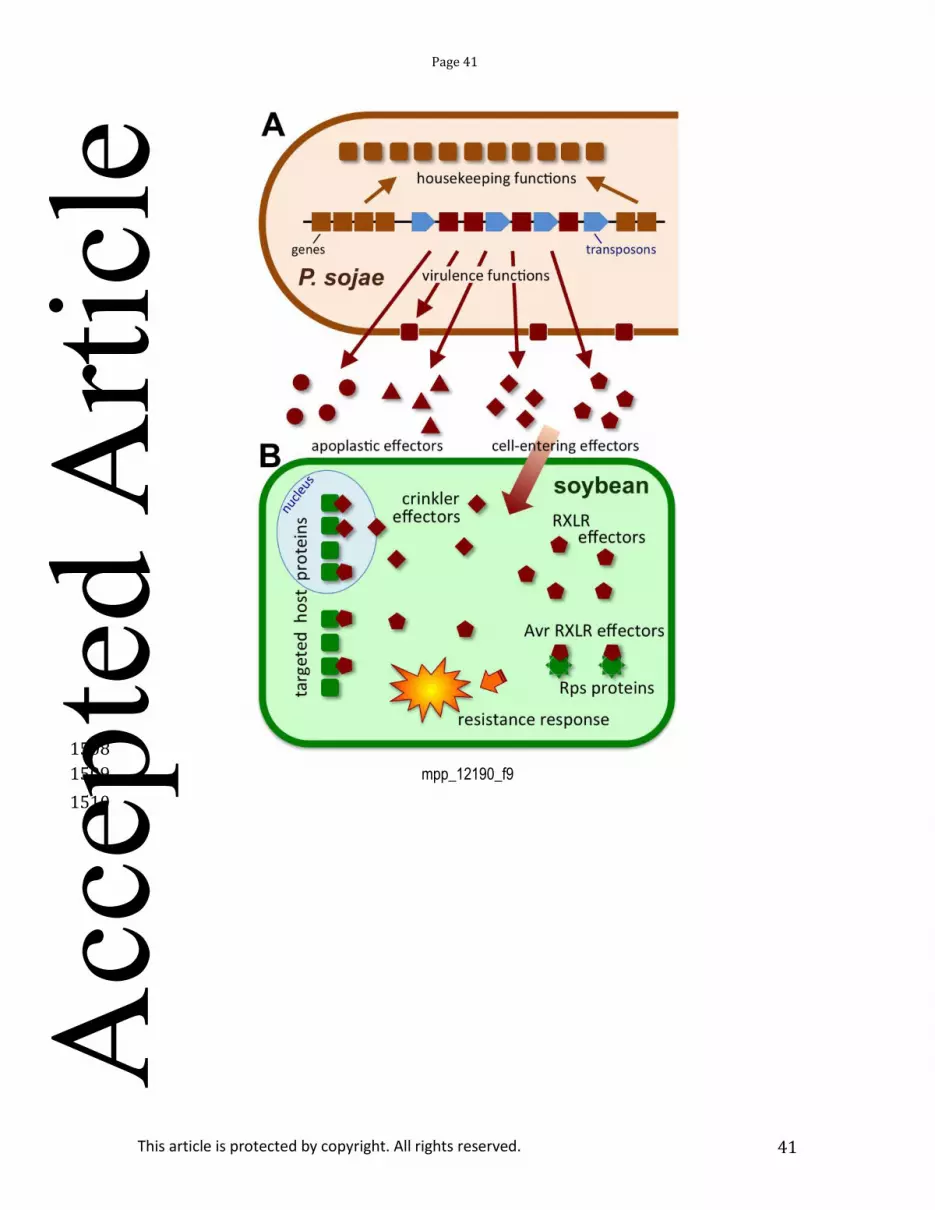
Page 41
This article is protected by copyright. All rights reserved. 41
1508
mpp_12190_f9 1509
1510
Acc
epte
d A
rticl
e

Page 42
This article is protected by copyright. All rights reserved. 42
1511
1512
mpp_12190_f10 1513
1514
Acc
epte
d A
rticl
e

Page 43
This article is protected by copyright. All rights reserved. 43
1515
1516
mpp_12190_f11 1517
1518
Acc
epte
d A
rticl
e

Page 44
This article is protected by copyright. All rights reserved. 44
1519
1520
mpp_12190_f12 1521
1522
Acc
epte
d A
rticl
e

Page 45
This article is protected by copyright. All rights reserved. 45
1523
1524
mpp_12190_f13 1525
1526
Acc
epte
d A
rticl
e

Page 46
This article is protected by copyright. All rights reserved. 46
1527
mpp_12190_f14 1528 Acc
epte
d A
rticl
e

Page 47
This article is protected by copyright. All rights reserved. 47
1529
1530
mpp_12190_f15 1531
1532
Acc
epte
d A
rticl
e

Page 48
This article is protected by copyright. All rights reserved. 48
1533
1534
mpp_12190_f16 1535
1536
Acc
epte
d A
rticl
e
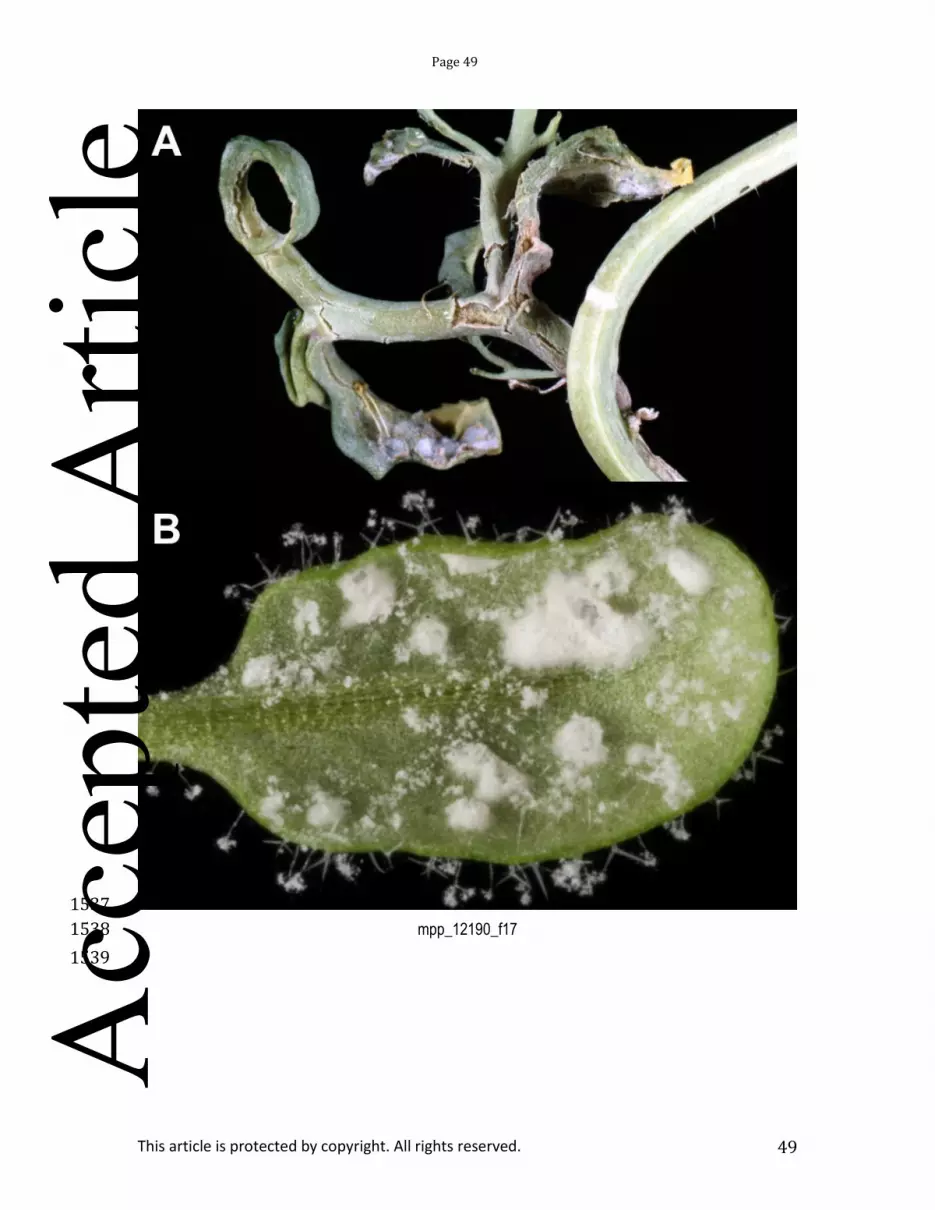
Page 49
This article is protected by copyright. All rights reserved. 49
1537
mpp_12190_f17 1538
1539
Acc
epte
d A
rticl
e

Page 50
This article is protected by copyright. All rights reserved. 50
1540
mpp_12190_f18 1541
1542
Acc
epte
d A
rticl
e





























