Embed Size (px)
Citation preview

The TACC proteins: TACC-lingmicrotubule dynamics and centrosomefunctionIsabel Peset1 and Isabelle Vernos1,2
1 Cell and Developmental Biology Program, Centre for Genomic Regulation (CRG), University Pompeu Fabra (UPF), Dr Aiguader 88,
Barcelona 08003, Spain2 Institucio Catalana de Recerca i Estudis Avancats (ICREA), Passeig Lluis Companys 23, 08010 Barcelona, Spain
Review
Abbreviations
AINT: ARNT interacting protein
AKAP350: A kinase (PRKA) anchor protein
Alp7: Altered growth polarity 7
Ark1: aurora-related kinase
ARNT: aryl hydrocarbon nuclear translocator protein
AZU-1: anti-zuai-1
CBP: calcium-binding protein
CPEB: cytoplasmic polyadenylation element binding protein
DCLK: doublecortin-like kinase
ECTACC: endothelial cell TACC
E1F4E: eukaryotic initiation factor 4E
ERIC: erythropoietin-induced cDNA
FOG-1: Friend of Gatal
GAS41: glioma amplified sequence 41
GCN5L2: general control of amino-acid synthesis 5-like 2
g-TURC: g-tubulin related complex
HEAT: huntingtin, elongation factor 3, A subunit of protein
phosphatase 2A and TOR1
INI-1: SWI/SNF core subunit
Ipl1: Increase-in-ploidy 1
ISREC: Swiss Institute for Experimental Cancer Research
KIF2C: kinesin family member 2C
LIS1: Lissencephaly-1
LSM7: U6 small nuclear NRA associated
MBD2: methyl-CpG binding domain protein 2
Mial: melanoma inhibitory activity 1
Mps1: MonoPolar Spindle 1
NDEL1 and NUDEL: nude nuclear distribution gene E homolog
(A. nidulans)-like 1 pCAF, p300/CBP-associated factor
SmG: snRNP Sm protein G
A major quest in cell biology is to understand themolecular mechanisms underlying the high plasticityof the microtubule network at different stages of thecell cycle, and during and after differentiation. Initialreports described the centrosomal localization ofproteins possessing transforming acidic coiled-coil(TACC) domains. This discovery prompted severalgroups to examine the role of TACC proteins during celldivision, leading to indications that they are importantplayers in this complex process in different organisms.Here, we review the current understanding of the role ofTACC proteins in the regulation of microtubuledynamics, and we highlight the complexity of centro-some function.
IntroductionCell proliferation and differentiation require dramaticrearrangements of the cytoskeleton that rely on the highlydynamic nature of the cytoskeletal components. Microtu-bules are dynamic filaments with fundamental roles ineukaryotic cell organization and function. During celldivision, they form the bipolar spindle, which segregatesthe chromosomes into the two daughter cells. Microtubulesshow prolonged states of polymerization and depolymer-ization that interconvert stochastically, exhibiting fre-quent transitions between growing and shrinkingphases, a property called ‘dynamic instability’ [1]. In thecell, multiple factors modulate this property by actingpositively or negatively on the nucleation, elongation ordestabilization of microtubules [1–3]. The relative activityof all these factors determines the steady-state length andstability of microtubules, in addition to their organization,and it is largely dictated by global and local phosphoryl-ation–dephosphorylation reactions [2,3]. In addition, othertypes of factors that have microtubule-severing and -anchoring activities also influence the microtubule net-work. The main microtubule-organizing centre (MTOC)of animal cells, the centrosome, acts as a platform uponwhich the different factors and activities accumulate in aregulated manner. It therefore exerts a tight local andtemporal control on the number, distribution and polarityof microtubules [4,5].
Corresponding authors: Peset, I. ([email protected]);Vernos, I. ([email protected]).
0962-8924/$ – see front matter � 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.tcb.2008.0
Transforming acidic coiled-coil (TACC) proteins emergedinitially as a group of proteins implicated in cancer. Thefirst member of the TACC family to be discovered wasidentified in a search of genomic regions that are amplifiedin breast cancer. It was named transforming acidic coiled-coil 1 (TACC1) because of its highly acidic nature, thepresence of a predicted coiled-coil domainat itsC terminus(now known as the TACC domain), and its ability topromote cellular transformation [6]. TACC proteins arepresent in different organisms, ranging from yeasts tomammals. There is only one TACC protein in the nema-tode Caenorhabditis elegans (TAC-1), in Drosophila mel-anogaster (D-TACC), in Xenopus laevis (Maskin), and
TTK: TTK protein kinase
Zyg-8: ZYGote defective
Zyg-9: ZYGote defective
6.005 Available online 23 July 2008 379

Review Trends in Cell Biology Vol.18 No.8
in the fission yeast Schizosaccharomyces pombe (Alp7also known as Mia1p); by contrast, mammals have threesuch proteins (TACC1, TACC2 [also known as AZU-1 andECTACC] and TACC3 [also known as AINT and ERIC1])[7–11]. Alternative splicing further increases the comp-lexity of the TACC protein family in mammals and flies[12–16].
The three human genes encoding TACC proteins are allin genomic regions that are rearranged in certain cancers,and their expression is altered in cancers from differenttissues. TACC1 and TACC2 are located in chromosomes8p11 and 10q26, respectively, two regions that are impli-cated in breast cancer and other tumors [6], and TACC3maps to 4p16, within a translocation breakpoint regionassociated with the disease multiple myeloma [17].
Figure 1. The TACC family of proteins: structural organization and regions of interactio
and the position of domains that interact with binding partners (underlined regions). TA
box). In addition, some members have highly acidic, imperfect repeats of 33 amino acids
boxes]) or a Ser–Pro Azu-1 motif (SPAZ) [24] (dark-grey boxes). Yellow lines indicate the
for AurA phosphorylation are shown as orange bars. The conserved Ser residue is highli
position of the Leu residue, which is important for the C. elegans TAC-1–Zyg-9 interac
proteins that have mapped interactions are shown.
380
Although TACC1 was originally found to be upregulatedin breast cancer [6], subsequent studies found that itsexpression is reduced in ovarian and breast cancer tissues[18,19]. TACC3 is also upregulated in several cancer celllines, including lung cancer [17,20]; but, again, it wasreported as being absent or reduced in ovarian and thyroidcancer tissues [21]. Initially, it was suggested that theTACC2 splice variant AZU-1 is a tumor suppressor inbreast cancer. However, the lack of any tumor phenotypein Tacc2-knockout mice did not support this idea [22]. Ittherefore appears that these proteins can be upregulatedor downregulated in different types of cancer or, surpris-ingly, even in the same type [14,18–25]; as such, theirputative involvement in cancer development and/or pro-gression is unclear.
n with binding partners. The figure shows alignment of the key structural features,
CC proteins have the conserved coiled-coil TACC domain at their C terminus (blue
(termed SPD repeats [28] owing to their specific amino acid composition [pale-blue
position of nuclear localization signals (NLSs). The conserved consensus sequences
ghted in orange, and additional consensus sites in Maskin are indicated in grey. The
tion, is shown with a white line [44]. For the sake of simplicity, only TACC family

Review Trends in Cell Biology Vol.18 No.8
Almost at the same time as the identification ofTACC1 in humans, Maskin was identified and extensivelycharacterized as a factor involved in the regulation ofmRNA translation during maturation of Xenopus oocytes[26]. Other TACC family members have also been impli-cated in various events related to gene regulation, in-cluding the regulation of translation, RNA maturationand gene expression (Figure 1, Table 1) [13,25,27–31].However, to date, no major common role has emergedfor TACC proteins in these processes. By contrast, a majorbreakthrough came with the identification of D-TACC as aDrosophila microtubule-associated and centrosomalprotein required for centrosome activity and microtubuleassembly during mitosis [12]. Since then, the idea thatTACC proteins have a role in regulating microtubuleassembly has gained solid support through various studiesperformed in different experimental systems. In the lightof these data, we review here our current understanding of
Table 1. Partners of TACC proteins, and the putative functions of
The table summarizes all the interreactions described in the literature for some TACC pro
red; proteins involved in RNA regulation are indicated in green; proteins involved in g
transport are indicated in black.
Abbreviations: aa, amino acids; AKAP350, A kinase (PRKA) CPC, chromosomal passen
acetyltransferase; IF, immunofluorescence; IP, immunoprecipitation; MT, microtubule; n.
the role of TACC proteins at the centrosome, and wediscuss some of the issues that still remain to be addressed.
The TACC proteinsThe TACC domain is the signature of this protein family.This coiled-coil domain is found at the C terminus of allthe family members, which have otherwise very diverseN-terminal domains (Figure 1) [7,16]. The TACC domainshows a high level of conservation throughout evolution,and the shorter member of the family, C. elegans TAC-1,consists of basically one TACC domain [8–10]. Together,this suggests that the TACC domain carries most ofthe common functional properties of this family ofproteins.
The temporal and tissue-specific expression patterns ofthe three mammalian TACC proteins have been moreextensively studied. TACC1 can be detected in severaladult tissues, but relatively high levels of expression occur
their interactions
teins. Proteins involved in MT dynamics and centrosomal functions are indicated in
ene regulation are indicated in blue and proteins involved in nucleo-cytoplasmic
ger complex; CRC, chromatin-remodeling complex; CT, centrosome; HAT, histone
d., not described; PCM, pericentriolar material; PD, pulldown; Y2H, Yeast two hybrid.
381

Review Trends in Cell Biology Vol.18 No.8
only at the beginning of development, after which itbecomes dramatically downregulated [6,32]. TACC2 is alsowidely expressed, showing the highest levels in heart andmuscle [32,33]. In mice, expression was detected at alldevelopmental stages [13]. By contrast, TACC3 isexpressed in relatively few adult tissues, but it showselevated levels in testis and ovary, and in the hematopoie-tic lineages [17,32,34]. Duringmouse development, TACC3is present in all the embryonic stages and particularly inproliferating tissues [15,32,35]. These data suggest thatTACC3 has a role during cell division, in particular duringdevelopment. Indeed, TACC3-deficient mice show embryo-nic lethality, associated with a greatly reduced cell num-ber, widespread apoptotic cell death, and mitotic defects[36,37]. Interestingly, this phenotypewas partially rescuedin mice that had reduced levels of the tumor suppressorprotein p53 [36]. However, to date, no clear picture hasemerged to describe the molecular mechanism linking p53activity and TACC3.
Intracellular localization of TACC proteinsLittle information is available concerning the cellularlocalization of TACC family members in interphase,although studies have revealed that some of them – thethree humanmembers andMaskin – are nuclear [38,39]. Itis during cell division that TACC proteins show their mostcharacteristic localization – within the centrosome(Figure 2, Box 1) [8–12,38,40–43]. In humans, the threefamily members show slightly different distribution pat-terns. TACC2 is strongly associated with the centrosomethroughout the cell cycle, whereas TACC1 and TACC3 onlylocalize to the centrosome during mitosis – TACC1 weakly,and TACC3 covering a larger area [38]. These differencesin localizations suggest that the three human TACCproteins have non-overlapping functions.
The centrosomal localization of TACC proteins is highlydynamic. However, given that microtubules do not modu-late the rapid exchange between centrosomal and cyto-plasmic pools in C. elegans, and considering that thecentrosomal localization of TAC-1, D-TACC and humanTACC proteins is insensitive to microtubule-depolymeriz-ing drugs, this class of proteins can be considered as corecomponents of the centrosome [8,9,12,38].
The characteristic centrosomal targeting of TACCproteins relies on their conserved TACC domain. Indeed,this domain alone was shown to localize strongly to thespindle poles and to the centre of centrosomal asters[12,42]. Recently, it was demonstrated that two residues(L229 and M581) in the TACC domain of C. elegans TAC-1are important for targeting TAC-1 to the centrosome [44].It is unclear at the moment whether these residues areimportant for structural reasons or for protein–proteininteractions.
However, the localization of TACC proteins is notrestricted to the centrosome, and most of them also associ-ate with microtubules during cell division to variousextents (Figure 2). Human TACC3 shows only restrictedassociation with spindle microtubules, whereas Maskinand C. elegans TAC-1 localize all along spindle microtu-bules, and D-TACC associates with both spindle and astralmicrotubules (Figure 2a,b) [8,12,38,40,42,45–47]. These
382
microtubule localization patterns are therefore specificfor each protein and are determined by sequences outsidethe TACC domain. It is interesting to note that truncatedproteins lacking the TACC domain do not localize to cen-trosomes ormicrotubules inDrosophila orXenopus [12,42].
Interestingly, some data suggest that TACC proteinsbind to the ends of microtubules. On the one hand, the highdegree of accumulation to the spindle poles suggests thatTACC proteins bind to microtubule minus-ends. In fact,this localization was directly observed for D-TACC inDrosophila embryos, in which microtubule minus-endscan be distinguished from the centrosomal aster at thespindle poles (Figure 2a) [12]. Consistently, Maskin loca-lizes to the centre of taxol-induced asters in Xenopus eggextracts [42]. On the other hand, green fluorescent protein(GFP)-labeled D-TACC proteins were visualized in livingDrosophila embryos as dots moving towards and awayfrom the centrosome, presumably associated to shrinkingor growing microtubule plus-ends [48]. Immunolocaliza-tion studies also suggest that TACC proteins are associ-ated with microtubule plus-ends in the vicinity of thechromosomes [8,9,11,12,48]. The mechanism underlyingthe preferential microtubule end localization of TACCproteins is still unclear. In vitro studies did not revealany preference for binding of Maskin to either the plus-ends or theminus-ends [40], suggesting that other proteinsmediate these end localizations.
Function(s) of the TACC proteins during cell divisionTo date, all the phenotypes described for situations inwhich the expression of TACC proteins is altered arerelated to defects in microtubule stability. In C. elegans,TAC-1-depleted embryos show defects in pronuclearmigration, shorter spindles and defective spindleelongation in anaphase. They also have shorter astralmicrotubules and, as a consequence, spindle-positioningdefects. Interestingly, microtubules do form in the cyto-plasm of TAC-1-depleted embryos, suggesting that TAC-1is required for microtubule assembly only at the centro-some. Consistently, the recovery of fluorescent tubulin atthe centrosome after photobleaching is slower in TAC-1-depleted embryos [8–10]. In Drosophila, d-tacc mutantsare female-sterile and have failures of pronuclear fusion.The majority of embryos appear to be arrested in the firstmitotic division, and those that develop have abnormallyshort centrosomal microtubules at all stages of the cellcycle and eventually die as a consequence of the accumu-lation of mitotic defects [12]. In Xenopus, spindlesassembled in egg extracts depleted of Maskin havereduced size and microtubule content, and the centro-somes nucleate fewer and shorter microtubules(Figure 2) [40–42]. In HeLa cells, depletion of TACC1 doesnot affect the cell cycle, althoughmultipolar spindles formand the cells also show proliferation defects [18]. By con-trast, the silencing of TACC3 results in partially destabi-lized microtubules, spindles with reduced microtubulecontent, and defects in chromosomealignment, in additionto a high mitotic index due to mitotic arrest [46,49].Consistently, increasing the concentration of D-TACCand Maskin results in the accumulation of these proteinsat the spindle poles and an increase in spindlemicrotubule
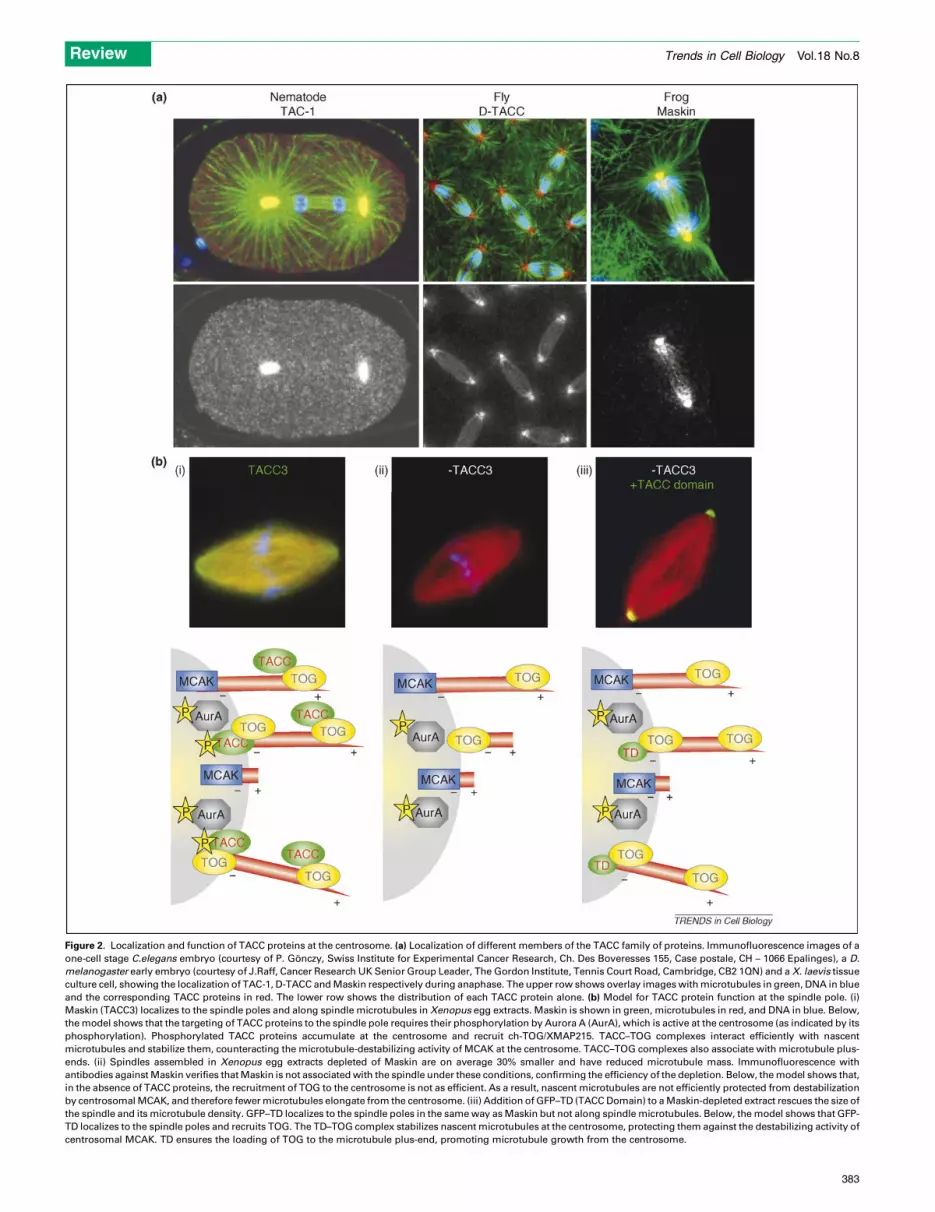
Figure 2. Localization and function of TACC proteins at the centrosome. (a) Localization of different members of the TACC family of proteins. Immunofluorescence images of a
one-cell stage C.elegans embryo (courtesy of P. Gonczy, Swiss Institute for Experimental Cancer Research, Ch. Des Boveresses 155, Case postale, CH – 1066 Epalinges), a D.
melanogaster early embryo (courtesy of J.Raff, Cancer Research UK Senior Group Leader, The Gordon Institute, Tennis Court Road, Cambridge, CB2 1QN) and a X. laevis tissue
culture cell, showing the localization of TAC-1, D-TACC and Maskin respectively during anaphase. The upper row shows overlay images with microtubules in green, DNA in blue
and the corresponding TACC proteins in red. The lower row shows the distribution of each TACC protein alone. (b) Model for TACC protein function at the spindle pole. (i)
Maskin (TACC3) localizes to the spindle poles and along spindle microtubules in Xenopus egg extracts. Maskin is shown in green, microtubules in red, and DNA in blue. Below,
the model shows that the targeting of TACC proteins to the spindle pole requires their phosphorylation by Aurora A (AurA), which is active at the centrosome (as indicated by its
phosphorylation). Phosphorylated TACC proteins accumulate at the centrosome and recruit ch-TOG/XMAP215. TACC–TOG complexes interact efficiently with nascent
microtubules and stabilize them, counteracting the microtubule-destabilizing activity of MCAK at the centrosome. TACC–TOG complexes also associate with microtubule plus-
ends. (ii) Spindles assembled in Xenopus egg extracts depleted of Maskin are on average 30% smaller and have reduced microtubule mass. Immunofluorescence with
antibodies against Maskin verifies that Maskin is not associated with the spindle under these conditions, confirming the efficiency of the depletion. Below, the model shows that,
in the absence of TACC proteins, the recruitment of TOG to the centrosome is not as efficient. As a result, nascent microtubules are not efficiently protected from destabilization
by centrosomal MCAK, and therefore fewer microtubules elongate from the centrosome. (iii) Addition of GFP–TD (TACC Domain) to a Maskin-depleted extract rescues the size of
the spindle and its microtubule density. GFP–TD localizes to the spindle poles in the same way as Maskin but not along spindle microtubules. Below, the model shows that GFP-
TD localizes to the spindle poles and recruits TOG. The TD–TOG complex stabilizes nascent microtubules at the centrosome, protecting them against the destabilizing activity of
centrosomal MCAK. TD ensures the loading of TOG to the microtubule plus-end, promoting microtubule growth from the centrosome.
Review Trends in Cell Biology Vol.18 No.8
383

Box 1. The centrosome
The centrosome is the main microtubule-organizing centre (MTOC)
of animal cells [4,5]. Centrosomes are small cellular organelles with
diverse morphologies, but they each consist of a pair of centrioles
surrounded by pericentriolar material (PCM), an electron-dense
material comprising core resident proteins and several non-
permanent structural and regulatory proteins. One characteristic
component of the PCM is g-tubulin, which promotes efficient
nucleation of microtubules at the centrosome. This imposes a
natural polarity on the resulting microtubule network, with micro-
tubule minus-ends focused at the PCM, and microtubule plus-ends
reaching out into the surrounding cytoplasm. In addition, centro-
somes act as platforms for the recruitment of multiple structural and
regulatory proteins. These include various microtubule nucleation
factors (e.g. pericentrin and centrosomin), microtubule-stabilizing
factors (e.g. ch-TOG/XMAP215 and TACC proteins), and microtu-
bule-destabilizing or -severing factors (e.g. the kinesin-like protein
MCAK, and katanin) [59]. The efficiency of microtubule elongation
from the centrosome is therefore determined by the relative
abundance and activity of all of these factors at any given time
[65]. The dynamic changes in centrosome morphology and activity
are tightly regulated by several centrosomal kinases, including
members of the Aurora, Polo-like kinase (Plk) and Never In Mitosis A
(NIMA) [4,5] families. Remarkably, many centrosomal proteins also
exist in a soluble, cytoplasmic pool, indicating that centrosomes are
highly dynamic organelles [5].
In resting mammalian cells, the centrosome migrates to the cell
surface, and one of the centrioles differentiates into the basal body
of a cilium [4], which functions as a sensory organelle or as a fluid
propeller. The presence of a cilium is transient in proliferating cells
in which the activity and number of centrosomes varies in tight
coordination with the cell cycle. The primary cilium present in G1
disassembles before the cell progresses into the cell cycle, and the
two centrioles duplicate during S phase. During G2, the two newly
formed centrosomes undergo a process called maturation, and they
integrate the control of entry into M phase with an increase in their
microtubule-nucleation capacity. This generates a robust aster of
highly dynamic microtubules, which are involved in centrosome
separation and spindle assembly. During mitosis, the centrosomes
are positioned at each spindle pole and have an important role in
determining spindle orientation and the plane of cell division [4].
Box 2. The ch-TOG/XMAP215 family
XMAP215 is the founding member of a large family of microtubule-
binding proteins. It was originally purified in the late 1980s from
Xenopus egg extracts. It was characterized as a factor that increases
the elongation rate of microtubules in vitro [66], and was found to
be related to chTOG (for colonic and hepatic tumor overexpressed
protein), a human protein overexpressed in tumor cells.
XMAP215 shows very different effects on microtubule dynamics
in vitro compared with other microtubule-associated proteins
(MAPs) such as tau and MAP2. Indeed, although these brain MAPs
promote a strong reduction in the frequency of microtubule
catastrophe without substantially altering the growth rate,
XMAP215 stimulates the growth rate of microtubules without
changing the catastrophe frequency. XMAP215 has also been found
to increase the depolymerization rate and to reduce the frequency of
rescues (i.e. switching between depolymerization and polymeriza-
tion phases). As a result, XMAP215 promotes an increase in
microtubule length and mass but, in so doing, promotes the
formation of microtubules of a highly dynamic nature, a property
that can be particularly important during M phase. XMAP215 has
also been shown to counteract the activity of the microtubule-
destabilizing motor MCAK [51]. Consistently, all of the phenotypes
associated with the disruption of ch-TOG/XMAP215 family members
are related to changes in microtubule stability, including decreased
microtubule growth and defects in spindle function [67].
ch-TOG/XMAP215 family proteins have a C-terminal domain
involved in MT binding and a N-terminal domain consisting of a
variable number of TOG domains. Each TOG domain contains six
HEAT repeats, which fold into a paddle-like domain, and wrap itself
around one tubulin dimer [68]. Recently, it was proposed that
XMAP215 acts as a processive polymerase, catalyzing the addition
of�25 tubulin dimers while moving with the assembling microtubule
tip. Under some circumstances, XMAP215 can also catalyze the
reverse reaction, therefore modulating microtubule dynamics [69].
Although, Drosophila Msps also localizes to the acentrosomal
spindle poles during female meiosis [50,70], a universal feature of this
family of proteins is their localization to the centrosome of metazoan
cells and to the spindle pole bodies of yeast. The centrosomal
localization of XMAP215 is mediated by its C-terminal microtubule-
binding domain [67]. In some organisms, this domain interacts with
TACC proteins [11,18]. Given that the localization of Msps is also
dependent on AurA in Drosophila [47], these data suggest that the
localization of XMAP215 family members to the centrosome relies on
their interaction with members of the TACC family.
Review Trends in Cell Biology Vol.18 No.8
length and number, which are effects opposite to thosecaused by depletion. A similar phenotype – accumulationat spindle poles and increase in microtubule length andnumber – arises upon overexpression of human TACC3but not TACC1 or TACC2 [38,42,48]. Altogether, thesedata clearly indicate that the TACC proteins have a con-served function in promoting centrosomal microtubuleassembly.
How do TACC proteins participate in microtubulestabilization?Several observations strongly suggest that TACC proteinsfunction not at the level of nucleation of microtubules but,rather, in the stabilization of microtubules. Experimentsperformed inXenopus egg extracts have clearly shown thatMaskin has no role in centrosomal microtubule nucleationactivity [42]. In C. elegans, TAC-1 mutant embryos do notshow defects in the distribution of the microtubule-nuclea-tor g-tubulin [8]. InDrosophila d-taccmutant embryos, thelocalization of g-tubulin and the centrosomal proteinsCP190 and CP60 to the centrosome is normal, suggestingthat microtubule nucleation at centrosomes is also unaf-fected [12]. Therefore, TACC proteins promotemicrotubulegrowth from the centrosome without altering the nuclea-tion of microtubules.
384
The TACC proteins only interact very weakly withpolymerized microtubules in vitro, but they do co-pelletvery efficiently with microtubules in Drosophila embryosand Xenopus egg extracts, suggesting that other factor(s)are involved [12,42,48]. The first clue to shed some lightupon this issue was a report describing an interactionbetween D-TACC andMinispindles (Msps), theDrosophilamember of the colonic and hepatic tumor-overexpressedgene (ch-TOG; also known as XMAP215) family of micro-tubule-associated proteins (Box 2) [48]. The functionalrelevance of this interaction is underscored by the obser-vation that TACC and ch-TOG/XMAP215 proteins havesimilar localizations. Furthermore, perturbing any of theseproteins produces similar phenotypes [8,49,50]. Interest-ingly, this interaction turned out to be conserved through-out evolution (Table 1) and to be mediated by the TACCdomain (Figure 1) [8,10,11,18,40–42,48]. This is consistentwith the idea that this domain has a crucial role in promot-ing microtubule assembly, and it also agrees with exper-imental data showing that the TACC domain is sufficientto rescue the phenotype of Maskin depletion in Xenopusegg extracts (Figure 2b) [40,42]. Overexpression studies

Box 3. The Aurora kinase family
The Aurora kinase family is an evolutionarily conserved family of
serine–threonine kinases. Although there is only one Aurora kinase
in yeasts (Ipl1p in S. cerevisiae, and Ark1 in S. pombe), metazoans
have three Aurora kinases: Aurora A (also known as STK6), Aurora B
(STK12) and Aurora C (STK13). Each of these proteins shows its
highest levels and activity during the G2 and M phases of the cell
cycle. The initial discovery of the Aurora mutation in Drosophila
implicated the Aurora protein in spindle assembly, but extensive
studies have shown that these kinases have more functions [71].
AurA and AurB have been more extensively studied. During cell
division, they have non-overlapping roles related to their distinctive
localizations. AurB is a chromosomal passenger protein required for
phosphorylation of histone H3, chromosome bi-orientation, the
spindle assembly checkpoint, and cytokinesis. The centrosomal
AurA, by contrast, has emerged as a major regulator of centrosome
activity, participating in centrosome maturation and separation, and
in spindle assembly. In addition, AurA has been implicated in entry
to M phase, and in mRNA translation, cilia disassembly, and
asymmetric cell division [72].
One clear function of AurA is the recruitment and regulation of
proteins at the centrosome, including centrosomin, g-TURC and
TACC proteins [71]. Relatively few substrates of the AurA kinase
have been identified, but several are spindle-assembly factors, such
as TACC proteins and NDEL1. Although AurA kinase can self-
activate by autophosphorylation, several activators have been
reported. One of them is TPX2, a RanGTP-regulated factor involved
in spindle assembly. TPX2 also targets AurA to spindle microtubules
[73,74].
In humans, the three Aurora kinases are overexpressed in a
variety of human cancers and are believed to have multiple roles in
the development and progression of cancer. Moreover, the gene
encoding the Aurora A kinase (AURKA) maps to the chromosome
region 20q13, which is frequently amplified in many human cancers.
The overexpression of AURKA induces centrosome amplification
and aneuploidy. It also confers resistance to taxol-dependent
apoptosis in cancer cells. In this context, it is also interesting to
note that AurA interacts with and inactivates the tumor suppressor
p53, and that it also interacts with the breast cancer susceptibility
gene BRCA1 and colocalizes with it at the centrosome. Given that
the overexpression of AurA has been shown to cause tumorigenic
transformation of human and rodent cells in vitro and in vivo [72], it
has been proposed that AurA acts as an oncogene.
Review Trends in Cell Biology Vol.18 No.8
have also provided some additional insights into the func-tional role of this domain. InHeLa cells, the overexpressionof any of the three TACC domains results in the formationof highly ordered, cytoplasmic polymers that interact withbundled microtubules but not with tubulin oligomers [38].In Drosophila embryos, overexpression of the C-terminalpart of D-TACC results in the formation of microtubuleasters in the cytoplasm – but only if Msps is also present[48]. All these data support the idea that the function of theTACC domain in promoting microtubule assembly ishighly dependent on its interaction with ch-TOG/XMAP215.
So what is the underlying mechanism promoting theassembly of microtubules? Experiments performed indifferent systems have shown that reducing TACC proteinlevels impairs the correct localization of ch-TOG/XMAP125to the centrosome [8–11,41,48,50]. One exception concernsTACC3 in HeLa cells, but this might have been due toincomplete TACC3 depletion or because any of the otherTACC proteins were compensating for the lack of TACC3by targeting ch-TOG to the centrosome [49]. In any case,increasing the concentration of TACC proteins results inan increase in the recruitment of ch-TOG/XMAP215 to thespindle poles [38,42,48]. By contrast, ch-TOG/XMAP215 isnot required for the localization of TACC proteins to thecentrosome [11,44,49].
Although all of these data strongly support the idea thatTACC proteins are required for the efficient recruitment ofch-TOG/XMAP215 proteins to the centrosome, it is stillunclear whether this targeting function is sufficient toexplain the function of TACC proteins. It is also possiblethat a functional relationship exists between TACCproteins and ch-TOG/XMAP215. In this context, it is inter-esting to recall that Msps is required for the formation ofmicrotubule asters in Drosophila embryo extracts contain-ing the C-terminal part of D-TACC [48]. Although there areonly a few clues concerning the mechanism involved at themolecular level, gel filtration experiments have shown thatXMAP215 and Maskin form a one-to-one complex in vitro,and this complex possesses a higher affinity for microtu-bules than do each protein on its own [41,42]. TACCproteins might therefore promote a conformational changein ch-TOG/XMAP215 that renders the molecule more effi-cient for microtubule binding. Interestingly, under theseconditions, microtubules are more resistant to depolymer-ization by a destabilizing factor, the mitotic centromere-associated kinesin MCAK (a kinesin-13 also known asKIF2C) [41,42].
The model that emerges is one in which TACC proteinsrecruit ch-TOG/XMAP215 to the centrosome and enhancesits microtubule binding and stabilizing activity. This coun-teracts the destabilizing activity of MCAK and therebypromotes microtubule growth from the centrosome(Figure 2) [41,42,51].
Regulation of TACC proteins by Aurora AAnother conserved partner of TACC proteins is the Ser–Thr kinase Aurora A (AurA–STK6) (Table 1, Box 3)[18,41,42,45,47,52]. In vitro pull-down experiments haveshown that Maskin interacts directly with AurA [42].Moreover, TACC proteins are good substrates for this
kinase in vitro, and most of them have one or moresequences that conform to a consensus motif for phos-phorylation by AurA. In all cases, these sites are locatedoutside the TACC domain (Figure 1, Table 1)[41,42,45,47,52,53]. One of them is conserved in severalTACC orthologs, indicating that it is functionally import-ant [45,52,53]. There is strong experimental support inseveral systems (i.e. nematode, fly, frog and human cells)indicating that phosphorylation has a role in TACC func-tion at the centrosome [8,41,42,47,53].
Experiments with purified proteins have shown thatMaskin can bind simultaneously to AurA and XMAP215,suggesting that the binding sites for these two proteins donot overlap (Figure 1) [42]. Moreover, phosphorylationdoes not appear to have any positive or negative influenceon these interactions. Indeed, in Xenopus egg extract,XMAP215 is pulled down as efficiently by phosphorylatedwild type Maskin as it is by an unphosphorylatable,mutated Maskin protein [41]. Finally, AurA does notenhance the microtubule binding affinity achieved bythe Maskin–XMAP215 complex in comparison withMaskin or XMAP215 proteins in isolation [41]. All of thesedata suggest that phosphorylation does not regulate the
385

Review Trends in Cell Biology Vol.18 No.8
interaction or functionality of TACC–XMAP215 com-plexes. It also seems unlikely that phosphorylation inducesa change in the oligomerization state of Maskin, becauseMaskin and unphosphorylatable mutants of Maskin arefound at the same position on sucrose gradients of variousprotein combinations [41].
What then does phosphorylation regulate? One possib-ility is that it regulates the turnover of the protein at thecentrosome, favoring the retention of the phosphorylatedform. Another possibility is that a direct interaction withthe AurA is involved, resulting in the phosphorylation ofthe TACC protein at the centrosome. In fact, in Droso-phila, phosphorylation of D-TACC seems to be requirednot for its localization to the centrosome but, rather, toenable it to load at the microtubule minus-ends [53].However, the inhibition of AurA kinase activity by a smallmolecule results in TACC3 failing to localize to centro-somes in human cells [45], suggesting that kinase activityrather than the protein itself is required. In any case,various data support the idea that AurA regulates thecentrosomal recruitment of TACC. Immunofluorescencestudies performed with an antibody against the phos-phorylated form of TACC3 have shown that phosphory-latedTACCproteins aremostly found at the centrosome inHeLa cells andDrosophila embryos [41,53,54]. Consistentwith this, a non-phosphorylatable mutant of TACC3 doesnot localize efficiently to the centrosome in transfectedHeLa cells or in Xenopus egg extracts [41,53,54]. In Dro-sophila embryos, a similar, although less pronounced,effect is observed for the localization of a non-phosphor-ylatable mutant of D-TACC [53]. In further agreement,TACC proteins do not localize to the centrosome in AurAmutants in C. elegans or Drosophila, and there is a strongreduction in the association of Maskin with spindlesassembled in Xenopus egg extracts depleted of AurA[8,42,47]. However, these observations cannot excludethe possibility that thesemislocalizations are attributableto indirect effects on either centrosome content or func-tionality due to the absence of this important centrosomalkinase. Overall, AurA phosphorylation of TACC proteinsappears to contribute substantially to their recruitment toor accumulation at the centrosome, and, as a consequence,to the recruitment of ch-TOG/XMAP215 (Figure 2) [41].The precise molecular mechanism involved remains to beelucidated. In this context, it is noteworthy that alterna-tive pathways to target TACC proteins to the centrosomehave been described recently. Indeed, the centrosomallocalization of D-TACC relies on its interaction with motif1 of theDrosophila protein centrosomin [55], and the dual-specificity kinase TTK is essential for the centrosomallocalization of human TACC2 [56]. Recently, it was alsoreported that the centrosomal proteinNDEL1 (also knownas NUDEL), another substrate of AurA, is required for thetargeting of TACC3 to the centrosome in human cells [57].Interestingly, NDEL1 is a binding partner of LIS1, aprotein that participates in the regulation of cytoplasmicdynein function and microtubule organization during celldivision and neuronal migration [58]. NDEL1 is alsorelated to microtubule-remodeling mechanisms throughthe recruitment of the microtubule-severing factor kata-nin [59].
386
Clearly, additional work is needed to address some keyquestions and to understand how TACC proteins functionto promote the assembly of microtubules.
ConclusionsTACC proteins have recently emerged as importantplayers in the complex process of regulating microtubuledynamics during cell division. Although it is now clearlyestablished that they have amajor role at the centrosome –promoting microtubule elongation together with ch-TOG/XMAP215 proteins – the molecular mechanism underlyingtheir activity is still unclear. Solid data support the factthat they interact with, and are substrates of, the kinaseAurA; but, again, although phosphorylation is essential fortheir localization to and function at the centrosome, themolecular mechanism involved is far from being under-stood. In fact, it is noteworthy that TACC proteins alsofunction in pathways that do not involve the centrosome.Indeed, D-TACC and Msps both localize at the acentroso-mal poles of Drosophila female meiotic spindles, and bothare required for maintaining the bipolarity of these spin-dles [50]. In addition, some data from frog and fission yeastsupport the idea that TACC proteins participate in theRanGTP-dependent spindle assembly pathway [39,40,60].Furthermore, in the fission yeast S. pombe, Alp7 functionsin the maintenance of microtubule organization duringinterphase [61]. It is therefore very likely that TACCproteins have a more general role in microtubule stabiliz-ation than currently appreciated.
Finally, it is still unclear whether the quite distinct rolesof TACC proteins in different aspects of gene regulationand inmicrotubule assembly are connected in someway. Inthis context, it is noteworthy that recent studies haverevealed that different classes of RNAs are associated withthe mitotic spindle [62,63]. Interestingly, both Maskin andAurA are involved in the regulation of mRNA translationin Xenopus oocytes [26].
It is quite apparent that much remains to be done tounderstand more clearly how TACC proteins function. In awider context, it will be interesting to elucidate the func-tional connections between their roles in microtubulestabilization and in RNA control. In addition, future workshould reveal whether their role is only restricted to eventsoccurring during cell division.
As a final remark, it should be noted that the study ofTACC proteins is starting to offer a variety of promisingapplications related to cancer therapies. Recently, it wasshown that monitoring phospho-TACC3 is an efficient wayof evaluating the effectiveness of AurA inhibitors that arepromising anti-cancer drugs, some of which have recentlyentered clinical trials [45]. The observation that TACC3depletion sensitizes cells to paclitaxel-induced cell deathalso suggests that TACC3 itself is a promising target forthe treatment of the tumors resistant to this widely usedtherapy [64]. Finally, TACC3 is also emerging as a goodprognostic marker for some cancers [20].
AcknowledgementsWe thank all members of the Vernos Laboratory, especially LuisBejarano, Teresa Sardon and Roser Pinyol, for critically reading themanuscript and providing helpful comments. We thank P. Gonczy(ISREC, CH) and J. Raff (The Gurdon Institute, UK) for the kind gift of

Review Trends in Cell Biology Vol.18 No.8
the immunofluorescence images of C. elegans and D. melanogasterembryos, respectively. We also thank Christoph Spinzig for creativesuggestions. Work in the I.V. laboratory is supported by the CRG, theEuropean Union MRTN/CT 2004 512348 and the Spanish Ministrygrants BFU2006–04694 and CSD2006–0023.
References1 Desai, A. and Mitchison, T.J. (1997) Microtubule polymerization
dynamics. Annu. Rev. Cell Dev. Biol. 13, 83–1172 Kline-Smith, S.L. and Walczak, C.E. (2004) Mitotic spindle assembly
and chromosome segregation: refocusing on microtubule dynamics.Mol. Cell 15, 317–327
3 Gadde, S. and Heald, R. (2004) Mechanisms and molecules of themitotic spindle. Curr. Biol. 14, R797–R805
4 Bettencourt-Dias, M. and Glover, D.M. (2007) Centrosome biogenesisand function: centrosomics brings new understanding. Nat. Rev. Mol.Cell Biol. 8, 451–463
5 Nigg, E.A. (2002) Centrosome aberrations: cause or consequence ofcancer progression? Nat. Rev. Cancer 2, 815–825
6 Still, I.H. et al. (1999) Cloning of TACC1, an embryonically expressed,potentially transforming coiled coil containing gene, from the 8p11breast cancer amplicon. Oncogene 18, 4032–4038
7 Gergely, F. (2002) Centrosomal TACCtics. Bioessays 24, 915–9258 Bellanger, J.M. andGonczy, P. (2003) TAC-1 and ZYG-9 form a complex
that promotes microtubule assembly in C. elegans embryos. Curr. Biol.13, 1488–1498
9 Le Bot, N. et al. (2003) TAC-1, a regulator of microtubule length in theC. elegans embryo. Curr. Biol. 13, 1499–1505
10 Srayko, M. et al. (2003) Caenorhabditis elegans TAC-1 and ZYG-9 forma complex that is essential for long astral and spindle microtubules.Curr. Biol. 13, 1506–1511
11 Sato, M. et al. (2004) Interdependency of fission yeast Alp14/TOG andcoiled coil protein Alp7 in microtubule localization and bipolar spindleformation. Mol. Biol. Cell 15, 1609–1622
12 Gergely, F. et al. (2000) D-TACC: a novel centrosomal protein requiredfor normal spindle function in the early Drosophila embryo. EMBO J.19, 241–252
13 Lauffart, B. et al. (2003) Molecular cloning, genomic structure andinteractions of the putative breast tumor suppressor TACC2.Genomics81, 192–201
14 Line, A. et al. (2002) Altered splicing pattern of TACC1 mRNA ingastric cancer. Cancer Genet. Cytogenet. 139, 78–83
15 Hao, Z. et al. (2002) TACC3 expression and localization in the murineegg and ovary. Mol. Reprod. Dev. 63, 291–299
16 Still, I.H. et al. (2004) Structure–function evolution of the transformingacidic coiled coil genes revealed by analysis of phylogenetically diverseorganisms. BMC Evol. Biol. 4, 16
17 Still, I.H. et al. (1999) The third member of the transforming acidiccoiled coil-containing gene family, TACC3, maps in 4p16, close totranslocation breakpoints in multiple myeloma, and is upregulatedin various cancer cell lines. Genomics 58, 165–170
18 Conte, N. et al. (2003) TACC1–chTOG–Aurora A protein complex inbreast cancer. Oncogene 22, 8102–8116
19 Lauffart, B. et al. (2005) Aberrations of TACC1 and TACC3 areassociated with ovarian cancer. BMC Womens Health 5, 8
20 Jung, C.K. et al. (2006) Expression of transforming acidic coiled-coilcontaining protein 3 is a novel independent prognostic marker in non-small cell lung cancer. Pathol. Int. 56, 503–509
21 Ulisse, S. et al. (2007) Transforming acidic coiled-coil 3 and Aurora-Ainteract in human thyrocytes and their expression is deregulated inthyroid cancer tissues. Endocr. Relat. Cancer 14, 827–837
22 Schuendeln, M.M. et al. (2004) The centrosomal, putative tumorsuppressor protein TACC2 is dispensable for normal development,and deficiency does not lead to cancer. Mol. Cell. Biol. 24, 6403–6409
23 Raff, J.W. (2002) Centrosomes and cancer: lessons from a TACC.Trends Cell Biol. 12, 222–225
24 Chen, H.M. et al. (2000) AZU-1: a candidate breast tumor suppressorand biomarker for tumor progression. Mol. Biol. Cell 11, 1357–1367
25 Conte, N. et al. (2002) Carcinogenesis and translational controls:TACC1 is down-regulated in human cancers and associates withmRNA regulators. Oncogene 21, 5619–5630
26 Richter, J.D. (2007) CPEB: a life in translation. Trends Biochem. Sci.32, 279–285
27 Garriga-Canut, M. and Orkin, S.H. (2004) Transforming acidic coiled-coil protein 3 (TACC3) controls friend of GATA-1 (FOG-1) subcellularlocalization and regulates the association between GATA-1 and FOG-1during hematopoiesis. J. Biol. Chem. 279, 23597–23605
28 Lauffart, B. et al. (2002) Interaction of the transforming acidic coiled-coil 1 (TACC1) protein with ch-TOG and GAS41/NuBI1 suggestsmultiple TACC1-containing protein complexes in human cells.Biochem. J. 363, 195–200
29 Sadek, C.M. et al. (2000) Isolation and characterization of AINT: anovel ARNT interacting protein expressed during murine embryonicdevelopment. Mech. Dev. 97, 13–26
30 Gangisetty, O. et al. (2004) The transforming acidic coiled coil proteinsinteract with nuclear histone acetyltransferases. Oncogene 23, 2559–2563
31 Angrisano, T. et al. (2006) TACC3 mediates the association of MBD2with histone acetyltransferases and relieves transcriptional repressionof methylated promoters. Nucleic Acids Res. 34, 364–372
32 Sadek, C.M. et al. (2003) TACC3 expression is tightly regulated duringearly differentiation. Gene Expr. Patterns 3, 203–211
33 Pu, J.J. et al. (2001) Cloning and structural characterization ofECTACC, a new member of the transforming acidic coiled coil(TACC) gene family: cDNA sequence and expression analysis inhuman microvascular endothelial cells. Cytokine 13, 129–137
34 McKeveney, P.J. et al. (2001) Characterization and localization ofexpression of an erythropoietin-induced gene, ERIC-1/TACC3,identified in erythroid precursor cells.Br. J. Haematol. 112, 1016–1024
35 Aitola, M. et al. (2003) Aint/Tacc3 is highly expressed in proliferatingmouse tissues during development, spermatogenesis, and oogenesis.J. Histochem. Cytochem. 51, 455–469
36 Piekorz, R.P. et al. (2002) The centrosomal protein TACC3 is essentialfor hematopoietic stem cell function and genetically interfaces withp53-regulated apoptosis. EMBO J. 21, 653–664
37 Yao, R. et al. (2007) TACC3 is required for the proper mitosis ofsclerotome mesenchymal cells during formation of the axialskeleton. Cancer Sci. 98, 555–562
38 Gergely, F. et al. (2000) The TACC domain identifies a family ofcentrosomal proteins that can interact with microtubules. Proc.Natl. Acad. Sci. U. S. A. 97, 14352–14357
39 Albee, A.J. et al. (2006) Phosphorylation of Maskin by Aurora-A isregulated by RanGTP and importin beta. J. Biol. Chem. 281, 38293–38301
40 O’Brien, L.L. et al. (2005) The Xenopus TACC homologue, Maskin,functions in mitotic spindle assembly. Mol. Biol. Cell 16, 2836–2847
41 Kinoshita, K. et al. (2005) Aurora A phosphorylation of TACC3/Maskinis required for centrosome-dependentmicrotubule assembly inmitosis.J. Cell Biol. 170, 1047–1055
42 Peset, I. et al. (2005) Function and regulation ofMaskin, a TACC familyprotein, in microtubule growth during mitosis. J. Cell Biol. 170, 1057–1066
43 Steadman, B.T. et al. (2002) Transforming acidic coiled-coil-containingprotein 4 interacts with centrosomal AKAP350 and the mitotic spindleapparatus. J. Biol. Chem. 277, 30165–30176
44 Bellanger, J.M. et al. (2007) ZYG-9, TAC-1 and ZYG-8 together ensurecorrect microtubule function throughout the cell cycle of C. elegansembryos. J. Cell Sci. 120, 2963–2973
45 LeRoy, P.J. et al. (2007) Localization of human TACC3 to mitoticspindles is mediated by phosphorylation on Ser558 by Aurora A: anovel pharmacodynamic method for measuring Aurora A activity.Cancer Res. 67, 5362–5370
46 Schneider, L. et al. (2007) The transforming acidic coiled coil 3 proteinis essential for spindle-dependent chromosome alignment and mitoticsurvival. J. Biol. Chem. 282, 29273–29283
47 Giet, R. et al. (2002) Drosophila Aurora A kinase is required to localizeD-TACC to centrosomes and to regulate astral microtubules. J. CellBiol. 156, 437–451
48 Lee, M.J. et al. (2001) Msps/XMAP215 interacts with the centrosomalprotein D-TACC to regulate microtubule behaviour. Nat. Cell Biol. 3,643–649
49 Gergely, F. et al. (2003) The ch-TOG/XMAP215 protein is essential forspindlepole organization inhumansomatic cells.GenesDev.17, 336–341
50 Cullen, C.F. and Ohkura, H. (2001) Msps protein is localized toacentrosomal poles to ensure bipolarity of Drosophila meioticspindles. Nat. Cell Biol. 3, 637–642
387

Review Trends in Cell Biology Vol.18 No.8
51 Tournebize, R. et al. (2000) Control of microtubule dynamics by theantagonistic activities of XMAP215 and XKCM1 in Xenopus eggextracts. Nat. Cell Biol. 2, 13–19
52 Pascreau, G. et al. (2005) Phosphorylation of Maskin by Aurora-Aparticipates in the control of sequential protein synthesis duringXenopus laevis oocyte maturation. J. Biol. Chem. 280, 13415–13423
53 Barros, T.P. et al. (2005) Aurora A activates D-TACC–Msps complexesexclusively at centrosomes to stabilize centrosomal microtubules.J. Cell Biol. 170, 1039–1046
54 Barnard, D.C. et al. (2005) Differential phosphorylation controlsMaskinassociation with eukaryotic translation initiation factor 4E andlocalization on the mitotic apparatus. Mol. Cell. Biol. 25, 7605–7615
55 Zhang, J. and Megraw, T.L. (2007) Proper recruitment of gamma-tubulin and D-TACC/Msps to embryonic Drosophila centrosomesrequires Centrosomin Motif 1. Mol. Biol. Cell 18, 4037–4049
56 Dou, Z. et al. (2004) TTK kinase is essential for the centrosomallocalization of TACC2. FEBS Lett. 572, 51–56
57 Mori, D. et al. (2007) NDEL1 phosphorylation by Aurora-A kinase isessential for centrosomal maturation, separation, and TACC3recruitment. Mol. Cell. Biol. 27, 352–367
58 Wynshaw-Boris, A. (2007) Lissencephaly and LIS1: insights into themolecular mechanisms of neuronal migration and development. Clin.Genet. 72, 296–304
59 Toyo-Oka, K. et al. (2005) Recruitment of katanin p60 byphosphorylated NDEL1, an LIS1 interacting protein, is essential formitotic cell division and neuronal migration. Hum. Mol. Genet. 14,3113–3128
60 Sato, M. and Toda, T. (2007) Alp7/TACC is a crucial target in Ran-GTPase-dependent spindle formation in fission yeast.Nature 447, 334–337
61 Zheng, L. et al. (2006) The fission yeast transforming acidic coiled coil-related protein Mia1p/Alp7p is required for formation andmaintenance of persistent microtubule-organizing centers at thenuclear envelope. Mol. Biol. Cell 17, 2212–2222
388
62 Blower, M.D. et al. (2007) Genome-wide analysis demonstratesconserved localization of messenger RNAs to mitotic microtubules.J. Cell Biol. 179, 1365–1373
63 Blower, M.D. et al. (2005) A Rae1-containing ribonucleoproteincomplex is required for mitotic spindle assembly. Cell 121,223–234
64 Schneider, L. et al. (2008) TACC3 depletion sensitizes to paclitaxel-induced cell death and overrides p21(WAF)-mediated cell cycle arrest.Oncogene 27, 116–125
65 Howard, J. and Hyman, A.A. (2007) Microtubule polymerases anddepolymerases. Curr. Opin. Cell Biol. 19, 31–35
66 Gard, D.L. and Kirschner, M.W. (1987) A microtubule-associatedprotein from Xenopus eggs that specifically promotes assembly atthe plus-end. J. Cell Biol. 105, 2203–2215
67 Kinoshita, K. et al. (2002) XMAP215: a key component of the dynamicmicrotubule cytoskeleton. Trends Cell Biol. 12, 267–273
68 Al-Bassam, J. et al. (2007) Crystal structure of a TOG domain:conserved features of XMAP215/Dis1-family TOG domains andimplications for tubulin binding. Structure 15, 355–362
69 Brouhard, G.J. et al. (2008) XMAP215 is a processive microtubulepolymerase. Cell 132, 79–88
70 Cullen, C.F. et al. (1999) mini spindles: A gene encoding a conservedmicrotubule-associated protein required for the integrity of the mitoticspindle in Drosophila. J. Cell Biol. 146, 1005–1018
71 Barr, A.R. and Gergely, F. (2007) Aurora-A: the maker and breaker ofspindle poles. J. Cell Sci. 120, 2987–2996
72 Giet, R. et al. (2005) Aurora kinases, aneuploidy and cancer, acoincidence or a real link? Trends Cell Biol. 15, 241–250
73 Kufer, T.A. et al. (2002) Human TPX2 is required for targeting Aurora-A kinase to the spindle. J. Cell Biol. 158, 617–623
74 Bayliss, R. et al. (2003) Structural basis of Aurora-A activation by TPX2at the mitotic spindle. Mol. Cell 12, 851–862
75 Delaval, B. et al. (2004) Aurora B -TACC1 protein complex incytokinesis. Oncogene 23, 4516–4522




























