Embed Size (px)
Citation preview

The role of sexual and clonal reproduction inmaintaining population in Fritillaria camtschatcensis (L.)
Ker-Gawl. (Liliaceae)
TAKAAKI SHIMIZU,1* YASUO HATANAKA,2 HIROKO ZENTOH,1 TAKESHI YASHIMA,1
EIICHIRO KINOSHITA,3à YASUYUKI WATANO1
AND TATEMI SHIMIZU1§
1Laboratory for Plant Natural History, Faculty of Science, Kanazawa University, Kanazawa 920±11,Japan, 2Graduate School of Natural Science and Technology, Kanazawa University, Kanazawa 920±11,
Japan and 3Botanic Garden, Faculty of Science, Kanazawa University, Kanazawa 920±11, Japan
Fritillaria camtschatcensis can reproduce by means of both sexual reproduction and clonal multiplication.Despite proli®c seed production, no seedlings have been found for several years in natural populationson Mt Hakusan. The purposes of this paper are to clarify: (i) whether population size is maintainedmainly by clonal multiplication; and (ii) if this is the case, to what extent occasional seedling estab-lishment affects population growth rate and population structure. Two permanent quadrats wereplaced in subalpine meadows in 1992 on Mt Hakusan. Plant size, location and reproductive states forall ramets in the quadrats were recorded every year. Projection matrices were created based on ®eldcensus, and computer simulation experiments were performed. Fritillaria camtschatcensis had two types of¯ower, male ¯ower and cosexual ¯ower, and they were changeable. This is the ®rst report on sexlability in Fritillaria. Clonal growth was more closely correlated with life-history stages, especially withsexual states than with plant size. The population growth rate, k, was 1.006 for the Mizuyajiri popu-lation and 1.047 for the Nanryu population, respectively. Seedlings were found in 1996 for the ®rsttime. These facts indicate that populations of F. camtschatcensis on Mt Hakusan can usually be main-tained by clonal multiplication. However, it is not yet certain whether seeds germinate every year orwhether a ¯ush of seedling emergence occurs once in every few years in natural populations. Computersimulation revealed that: (i) there was a critical germination rate above which population growth ratesuddenly increased; and (ii) occasional seedling establishment could provide almost the same contri-bution to population growth rate as that of annual seedling establishment. These results suggest thatpopulation size can be maintained mainly by clonal multiplication, and the role of sexual reproductionlies beyond maintaining the population size in F. camtschatcensis.
Key words: clonal reproduction; Fritillaria camtschatcensis; life history; sex lability; sexual reproduc-tion.
INTRODUCTION
Many perennial herbs and shrubs have beenknown to maintain populations simultaneously bymeans of sexual reproduction and clonal multi-plication. These two modes of reproduction havetheir own short-term and long-term advantages(cf. Silander 1985). Sexual reproduction has beenthought to create a new combination of genomesand purge accumulated deleterious mutations,and it leads to acceleration of adaptation andspeciation in response to selection (Maynard
*Present address: MTS Institute Inc., Kandasuda-cho, 1±5, Tokyo 101, Japan.
Present address: Ishikawa Prefectural Of®ce, Hir-osaka, Kanazawa 920, Japan.
àAuthor to whom correspondence should be ad-dressed.
§Present address: Matsuoka 211±3, Okada, Matsu-moto 390±03, Japan.
Received 26 March 1997.Accepted 17 November 1997.
Ecological Research (1998) 13, 27±39

Smith 1978). Clonal reproduction, on the otherhand, has been assumed to insure maintenance ofa local population even when recruitment byseedling is dif®cult (Abrahamson 1980; Pitelkaet al. 1985; Angevine & Handel 1986; Kawanoet al. 1987; Eriksson 1988, 1989; Nault & Gagnon1993).
Recently, much attention has been paid to howthe population size and genetic diversity can bemaintained in clonal species. Vegetatively repro-ducing species form a clone comprising a lot ofmore-or-less independent ramets, which some-times occupy a large area and/or persist for along time (Cook 1985). This causes some altera-tions to population structures of clonal species.Bimodal distribution is sometimes observed inclonally reproducing species (Eriksson & Bremer1993; Nault & Gagnon 1993). Sex expression andbreeding system also have a great in¯uence onseed production, seed dispersal and eventually onthe population structure (Silander 1985; Lovett-Doust & Laporte 1991; Susko et al. 1994).
Occasional or periodic recruitment of seedlingshas been known in some clonal species, and thepopulation size has been known to be maintainedby clonal multiplication in many of these species(Harper 1977; Cook 1985; Parker & Hamrick1992; Eriksson & Bremer 1993). Eriksson (1989)reported that seedling recruitment occurred inestablished populations of only 40% of 68 clonalspecies. Cook (1985) and Sebens and Thorne(1985) pointed out that new empty habitats (e.g.those created by catastrophic disturbance) werenecessary for seedlings to establish successfully.Watkinson and Powell (1993) revealed by com-puter simulation that a low ratio of seedlings toramet recruits was suf®cient to maintain popu-lation size, size structure of genets and geneticdiversity in Ranunculus repens.
Throughout the ®eld surveys from 1992 to1995, no seedlings were observed in naturalpopulations of F. camtschatcensis on Mt Hakusan,while a considerable number of seeds were pro-duced every year. We ®rst found seedlings in1996 and assumed that a natural population ofF. camtschatcensis on Mt Hakusan had usually beenmaintained by clonal multiplication, and seedlingrecruitment had occasionally occurred in a par-ticular year when adequate conditions for seed-ling emergence were ful®lled.
This paper presents the life history of F. cam-tschatcensis based on ®eld data from 1992 to 1995 onMt Hakusan. In the ®rst place, current states ofindependent ramets in the study site are described,and sexual types of ¯ower are also described. Be-cause life-cycle stages in¯uence survival and re-production of independent ramets in clonal plants,it is important in the study of the life history of F.camtschatcensis to know the stage distribution andpopulation dynamics. The size-dependent projec-tion matrix model (Caswell 1989) is adopted, andthe population growth rate (k) and expected stablestage distributions are calculated. Second, effectsof various situations of occasional seedling estab-lishment on the population growth rate and stageclass distribution are studied by some computersimulations to evaluate the role of seedling estab-lishment in maintaining the natural population.
METHODS
Study sites
This study was conducted on Mt Hakusan in thecentral Honshu, Japan. Two sites were chosen for®eld study. One was settled on ¯at ground about2450 m a.s.l. near Mizuyajiri snow valley in Mu-rodo-Daira, where a lot of dense patches ofF. camtschatcensis scattered. Snow cover usuallydisappears by early July around the study site. Thevegetation is sparse, and mainly composed ofF. camtschatcensis, Empetrum nigrum var. japonicum,Deschampsia caespitosa var. festicaefolia and Polygonumweyrichii var. alpinum (cf. Suzuki 1970). This site isdesignated as `Mizuyajiri' population in thispaper. The other was settled at Nanryu-Teien,about 2050 m a.s.l., south-east about 2 km awayfrom the Mizuyajiri population, and designated as`Nanryu' population. The topography of Nanryu-Teien is like a shallow basin surrounded by dwarfAbies mariessi forest. Snow deposition reaches asdeep as 6 m in winter, and snow usually remainsin early July. Vegetation gradually changes alongwith soil moisture. The center of Nanryu-Teien isalways wet, and the vegetation is composed ofPrimula cuneifolia var. hakusanensis, Plantago haku-sanensis and Juncus beringensis. The marginal regionis mesic, and the vegetation is dense. It is mainlycomposed of Carex blepharicarpa, Fauria crista-gallissp. japonica. and Fritillaria camtschatcensis.
28 T. Shimizu et al.

The plant
Fritillaria camtschatcensis (L.) Ker-Gawl. is a pe-rennial herb growing in the northern part ofJapan, Sakhalin, the Kurile Islands, Kamchatkaand the north-western part of North America(Shimizu 1983). Of the approximate 100 speciesof the genus Fritillaria distributed in the temper-ate zone from western Europe and the Mediter-ranean region to east Asia (Mabberley 1987),F. camtschatcensis is the species that can spread itshabitats at the highest latitude and altitude. Thehabitat of this species in central Honshu is re-stricted to the subalpine and alpine meadow.
Fritillaria camtschatcensis grows in sunny, mesicmeadows more than 2000 m a.s.l. on Mt Haku-san. Shoot elongation or radical leaf expansiontakes place soon after thawing, and ¯oweringusually occurs at the middle of July. Fritillariacamtschatcensis inhabiting central Honshu is diploid(Shimizu 1983), and usually does not bear morethan two ¯owers.
Bulbs are subterranean storage organs com-posed of a thickened short axis and with nu-merous bulb leaves attached spirally. One or twonew buds are sometimes formed at the axil of theupper bulb leaves when a bulb grows largeenough. When a new bulb is formed, the old onebegins to decompose. Thus, this plant can mul-tiply clonally by two means: one is the division ofa bulb itself into two independent bulbs afterproducing two axillary buds, and the other isseparation of bulb leaves from a mother bulb andthe formation of new independent ramets(Shimizu & Umebayashi 1995). Some morpho-logical characters, such as total leaf area and totalweight of a whole plant, are well correlated withthe diameter of an aboveground stem at groundlevel (Y. Hatanaka, unpubl. data).
Fritillaria camtschatcensis has two types of ¯ower.Male ¯owers have only stamina with a degener-ated pistil, and cosexual ¯owers have bothstamina and a pistil. Reciprocal cross-pollinationexperiments reveal that a morphological cosexual¯ower is functionally cosexual, and this species isself-incompatible (Yashima et al. 1997).
Field census
Two permanent quadrats were established in1992 in the Mizuyajiri population, and in 1993 in
the Nanryu population, respectively. The totalarea of the permanent quadrats was 8 m2 for theMizuyajiri population and 13.4 m2 for the Nan-ryu population. Field census was carried out twiceeach growing season. At ¯owering, ramet posi-tion in the quadrat, the sexual type of the ¯owerand the diameter of an aboveground stem atground level (designated as D in this paper) wererecorded. A few weeks after the ¯owering period,the number of capsules in the permanent quad-rats was recorded. The number of mature seedsper capsule in the quadrats was estimated fromthe seed number in capsules of various size,which were picked outside the quadrats(n � 30±50).
It was hardly possible to mark small ramets(one-leaf ramets) precisely in order to follow theirfate because they grew too densely in the quadrat.Therefore, the quadrats were divided into smallsubquadrats (20 cm ´ 20 cm), and the totalnumber of one-leaf ramets in the subquadratswas recorded. However, this method can only beused to ®nd out the net changes in ramet num-ber. In order to follow the fate of one-leaf ra-mets, some quadrats were set in 1994 around themain quadrat in the Mizuyajiri and the Nanryupopulations. All ramets were numbered andmapped. Based on Shimizu and Umebayashi(1995), we assumed that new-born ramets wereproducts of clonal reproduction of the nearestramet that bore an aboveground stem in theprevious year. When such a big ramet was notfound within a radius of 5 cm of the new ramet,the nearest one-leaf ramet was assumed to be themother.
Projection matrix model and its application
The projection matrix model is shown as follows,
Nt�1 � ANt;
where Nt is a column vector representing thedistribution of a population classi®ed by stagecriteria at time t, and A is a projection matrixdescribing the fate of ramets from its birth todeath. The maximum eigen value represents anapproximate value of the population growth rate,k. The right eigen vector, u, corresponding to themaximum eigen value, describes the stable stagedistribution (Caswell 1989).
Sexual and clonal reproduction in Fritillaria 29

In stochastic simulation, some matrices wereprepared, and they were assigned a samplingprobability. In this study, four matrices were pre-pared. One matrix represents no seedling emer-gence (the original matrix created on ®eld census),and the other three matrices represent situationswith different seedling recruitment. The ®rst rowof the projection matrix represents the expectednumber of seedlings, but there were no actual datafor seed germination. Thus, the number of seedsand seedlings for each reproductive stage was es-timated in the following ways: the number of ma-ture seeds per fruit was counted in theexperimental quadrats and the expected seednumber per reproductive individual was calcu-lated; overall fruit/¯ower ratios were estimated foreach population; and three different germinationrates were assumed, 1%, 3% and 5%. In thosematrices, the survival rate of seedlings is 0.50, andother elements are ®xed at the same values as thoseof the original matrix. Average maximum
likelihood estimators of population growth rateand stage distributions after 50 years are calculatedfrom 100 iterations (Caswell 1989).
RESULTS
Life cycle stage and sexual type
Three major life cycle stages were recognized:seedling stage, pre-reproductive stage, and re-productive stage. Seedlings could be easily dis-tinguished by a linear leaf and seed coatsometimes remaining on top of the leaf for a fewweeks after germination (Fig. 1). Pre-reproduc-tive stage and reproductive stage could be furthersubdivided. In the pre-reproductive stage, twotypes were recognized: one with only one radicalleaf, and the other with a sterile abovegroundstem. The latter was very similar in appearance tothe reproductive ramets. Reproductive ramets
Fig. 1. Diagrammatic illustrations of life cycle stages. S, seedling; L, ramets with one radical leaf; A, ramets producingsterile aerial stem; M, ramets bearing a male ¯ower; C, ramets bearing a cosexual ¯ower; T, ramets bearing two ¯owersirrespective of sexual types.
30 T. Shimizu et al.
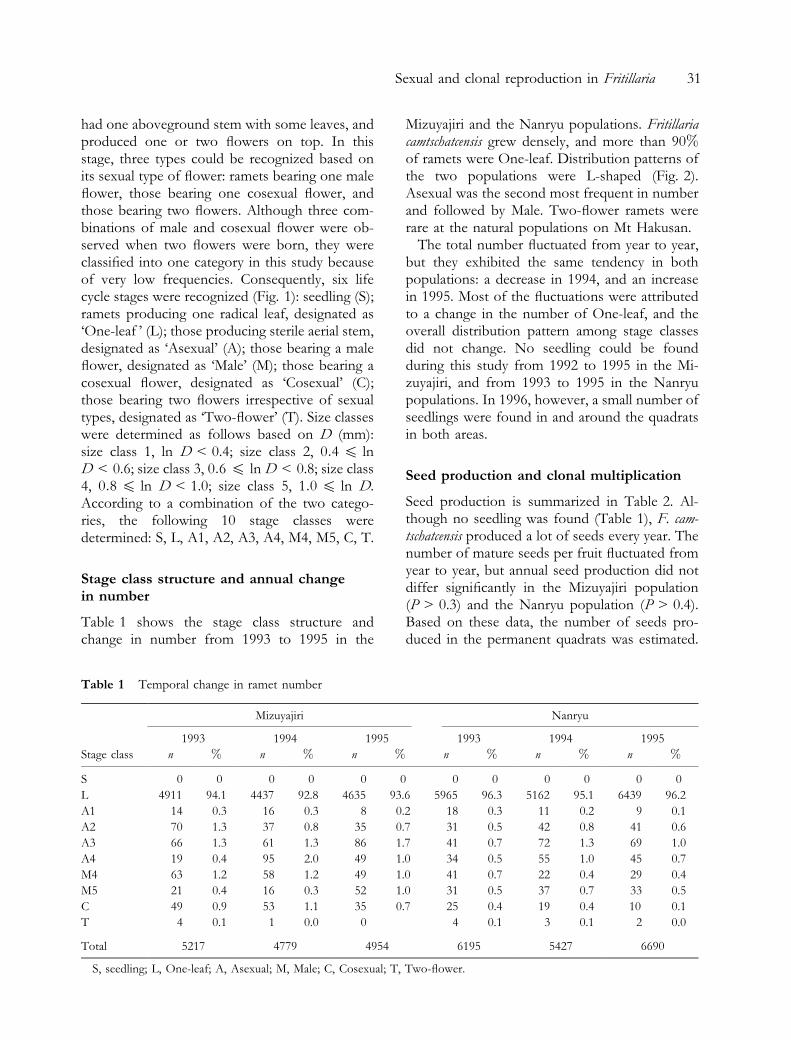
had one aboveground stem with some leaves, andproduced one or two ¯owers on top. In thisstage, three types could be recognized based onits sexual type of ¯ower: ramets bearing one male¯ower, those bearing one cosexual ¯ower, andthose bearing two ¯owers. Although three com-binations of male and cosexual ¯ower were ob-served when two ¯owers were born, they wereclassi®ed into one category in this study becauseof very low frequencies. Consequently, six lifecycle stages were recognized (Fig. 1): seedling (S);ramets producing one radical leaf, designated as`One-leaf ' (L); those producing sterile aerial stem,designated as `Asexual' (A); those bearing a male¯ower, designated as `Male' (M); those bearing acosexual ¯ower, designated as `Cosexual' (C);those bearing two ¯owers irrespective of sexualtypes, designated as `Two-¯ower' (T). Size classeswere determined as follows based on D (mm):size class 1, ln D < 0.4; size class 2, 0:4 O lnD < 0.6; size class 3, 0:6 O ln D < 0.8; size class4, 0:8 O ln D < 1.0; size class 5, 1:0 O ln D.According to a combination of the two catego-ries, the following 10 stage classes weredetermined: S, L, A1, A2, A3, A4, M4, M5, C, T.
Stage class structure and annual changein number
Table 1 shows the stage class structure andchange in number from 1993 to 1995 in the
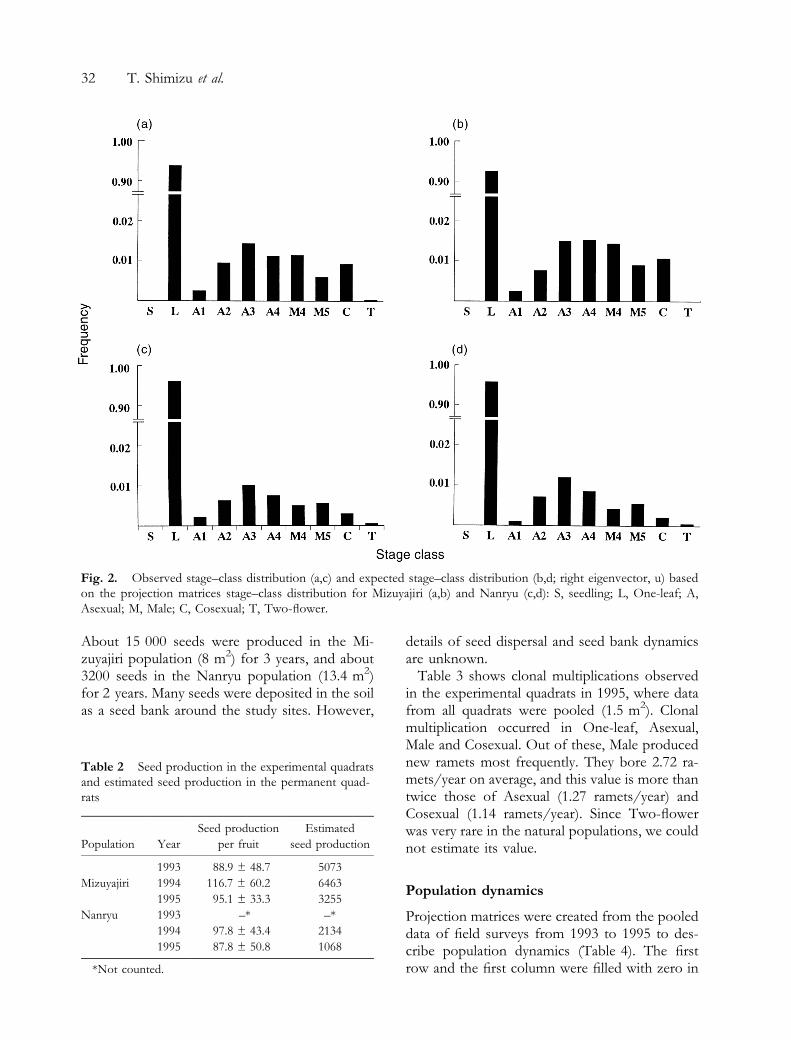
Mizuyajiri and the Nanryu populations. Fritillariacamtschatcensis grew densely, and more than 90%of ramets were One-leaf. Distribution patterns ofthe two populations were L-shaped (Fig. 2).Asexual was the second most frequent in numberand followed by Male. Two-¯ower ramets wererare at the natural populations on Mt Hakusan.
The total number ¯uctuated from year to year,but they exhibited the same tendency in bothpopulations: a decrease in 1994, and an increasein 1995. Most of the ¯uctuations were attributedto a change in the number of One-leaf, and theoverall distribution pattern among stage classesdid not change. No seedling could be foundduring this study from 1992 to 1995 in the Mi-zuyajiri, and from 1993 to 1995 in the Nanryupopulations. In 1996, however, a small number ofseedlings were found in and around the quadratsin both areas.
Seed production and clonal multiplication
Seed production is summarized in Table 2. Al-though no seedling was found (Table 1), F. cam-tschatcensis produced a lot of seeds every year. Thenumber of mature seeds per fruit ¯uctuated fromyear to year, but annual seed production did notdiffer signi®cantly in the Mizuyajiri population(P > 0.3) and the Nanryu population (P > 0.4).Based on these data, the number of seeds pro-duced in the permanent quadrats was estimated.
Table 1 Temporal change in ramet number
Mizuyajiri Nanryu
1993 1994 1995 1993 1994 1995
Stage class n % n % n % n % n % n %
S 0 0 0 0 0 0 0 0 0 0 0 0
L 4911 94.1 4437 92.8 4635 93.6 5965 96.3 5162 95.1 6439 96.2
A1 14 0.3 16 0.3 8 0.2 18 0.3 11 0.2 9 0.1
A2 70 1.3 37 0.8 35 0.7 31 0.5 42 0.8 41 0.6
A3 66 1.3 61 1.3 86 1.7 41 0.7 72 1.3 69 1.0
A4 19 0.4 95 2.0 49 1.0 34 0.5 55 1.0 45 0.7
M4 63 1.2 58 1.2 49 1.0 41 0.7 22 0.4 29 0.4
M5 21 0.4 16 0.3 52 1.0 31 0.5 37 0.7 33 0.5
C 49 0.9 53 1.1 35 0.7 25 0.4 19 0.4 10 0.1
T 4 0.1 1 0.0 0 4 0.1 3 0.1 2 0.0
Total 5217 4779 4954 6195 5427 6690
S, seedling; L, One-leaf; A, Asexual; M, Male; C, Cosexual; T, Two-¯ower.
Sexual and clonal reproduction in Fritillaria 31
About 15 000 seeds were produced in the Mi-zuyajiri population (8 m2) for 3 years, and about3200 seeds in the Nanryu population (13.4 m2)for 2 years. Many seeds were deposited in the soilas a seed bank around the study sites. However,
details of seed dispersal and seed bank dynamicsare unknown.
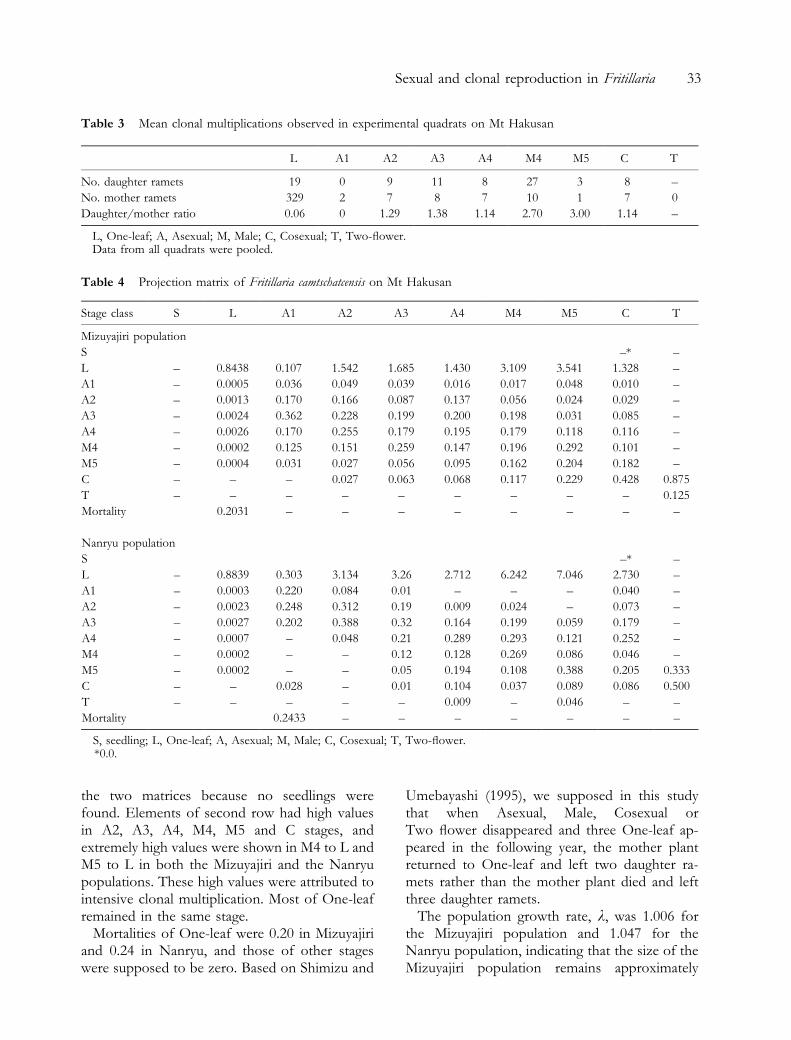
Table 3 shows clonal multiplications observedin the experimental quadrats in 1995, where datafrom all quadrats were pooled (1.5 m2). Clonalmultiplication occurred in One-leaf, Asexual,Male and Cosexual. Out of these, Male producednew ramets most frequently. They bore 2.72 ra-mets/year on average, and this value is more thantwice those of Asexual (1.27 ramets/year) andCosexual (1.14 ramets/year). Since Two-¯owerwas very rare in the natural populations, we couldnot estimate its value.
Population dynamics
Projection matrices were created from the pooleddata of ®eld surveys from 1993 to 1995 to des-cribe population dynamics (Table 4). The ®rstrow and the ®rst column were ®lled with zero in
Fig. 2. Observed stage±class distribution (a,c) and expected stage±class distribution (b,d; right eigenvector, u) basedon the projection matrices stage±class distribution for Mizuyajiri (a,b) and Nanryu (c,d): S, seedling; L, One-leaf; A,Asexual; M, Male; C, Cosexual; T, Two-¯ower.
Table 2 Seed production in the experimental quadratsand estimated seed production in the permanent quad-rats
Population Year
Seed production
per fruit
Estimated
seed production
1993 88.9 � 48.7 5073
Mizuyajiri 1994 116.7 � 60.2 6463
1995 95.1 � 33.3 3255
Nanryu 1993 ±* ±*
1994 97.8 � 43.4 2134
1995 87.8 � 50.8 1068
*Not counted.
32 T. Shimizu et al.
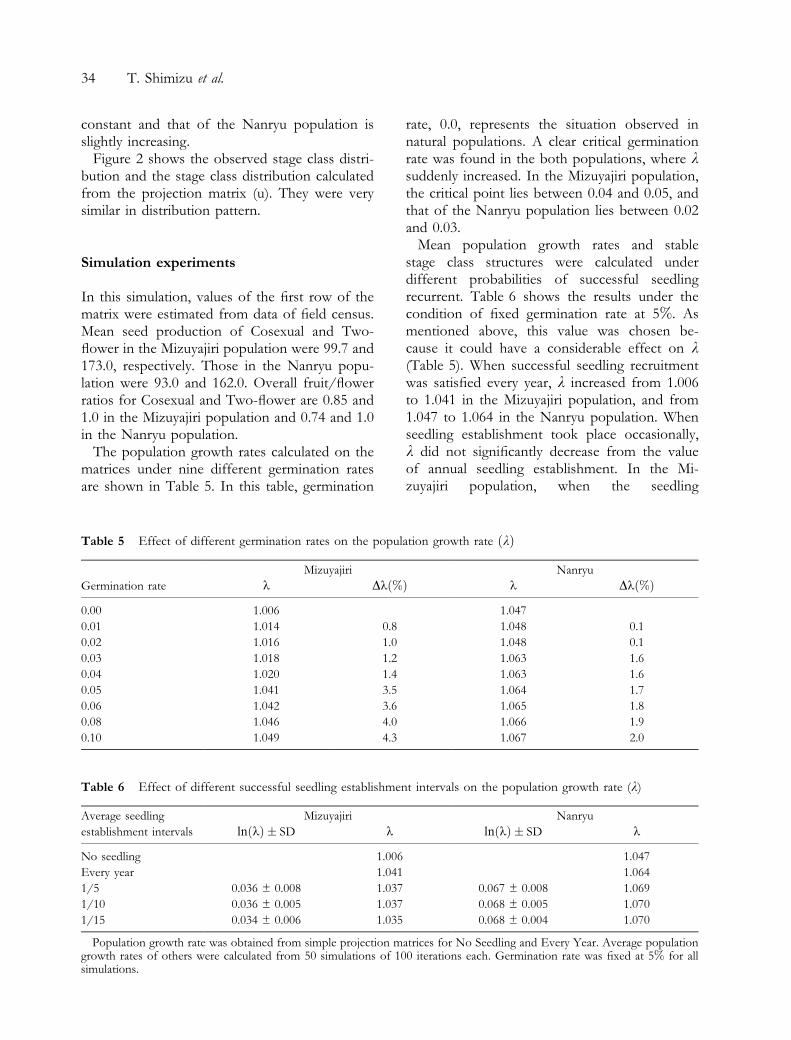
the two matrices because no seedlings werefound. Elements of second row had high valuesin A2, A3, A4, M4, M5 and C stages, andextremely high values were shown in M4 to L andM5 to L in both the Mizuyajiri and the Nanryupopulations. These high values were attributed tointensive clonal multiplication. Most of One-leafremained in the same stage.
Mortalities of One-leaf were 0.20 in Mizuyajiriand 0.24 in Nanryu, and those of other stageswere supposed to be zero. Based on Shimizu and
Umebayashi (1995), we supposed in this studythat when Asexual, Male, Cosexual orTwo ¯ower disappeared and three One-leaf ap-peared in the following year, the mother plantreturned to One-leaf and left two daughter ra-mets rather than the mother plant died and leftthree daughter ramets.
The population growth rate, k, was 1.006 forthe Mizuyajiri population and 1.047 for theNanryu population, indicating that the size of theMizuyajiri population remains approximately
Table 3 Mean clonal multiplications observed in experimental quadrats on Mt Hakusan
L A1 A2 A3 A4 M4 M5 C T
No. daughter ramets 19 0 9 11 8 27 3 8 ±
No. mother ramets 329 2 7 8 7 10 1 7 0
Daughter/mother ratio 0.06 0 1.29 1.38 1.14 2.70 3.00 1.14 ±
L, One-leaf; A, Asexual; M, Male; C, Cosexual; T, Two-¯ower.Data from all quadrats were pooled.
Table 4 Projection matrix of Fritillaria camtschatcensis on Mt Hakusan
Stage class S L A1 A2 A3 A4 M4 M5 C T
Mizuyajiri population
S ±* ±
L ± 0.8438 0.107 1.542 1.685 1.430 3.109 3.541 1.328 ±
A1 ± 0.0005 0.036 0.049 0.039 0.016 0.017 0.048 0.010 ±
A2 ± 0.0013 0.170 0.166 0.087 0.137 0.056 0.024 0.029 ±
A3 ± 0.0024 0.362 0.228 0.199 0.200 0.198 0.031 0.085 ±
A4 ± 0.0026 0.170 0.255 0.179 0.195 0.179 0.118 0.116 ±
M4 ± 0.0002 0.125 0.151 0.259 0.147 0.196 0.292 0.101 ±
M5 ± 0.0004 0.031 0.027 0.056 0.095 0.162 0.204 0.182 ±
C ± ± ± 0.027 0.063 0.068 0.117 0.229 0.428 0.875
T ± ± ± ± ± ± ± ± ± 0.125
Mortality 0.2031 ± ± ± ± ± ± ± ±
Nanryu population
S ±* ±
L ± 0.8839 0.303 3.134 3.26 2.712 6.242 7.046 2.730 ±
A1 ± 0.0003 0.220 0.084 0.01 ± ± ± 0.040 ±
A2 ± 0.0023 0.248 0.312 0.19 0.009 0.024 ± 0.073 ±
A3 ± 0.0027 0.202 0.388 0.32 0.164 0.199 0.059 0.179 ±
A4 ± 0.0007 ± 0.048 0.21 0.289 0.293 0.121 0.252 ±
M4 ± 0.0002 ± ± 0.12 0.128 0.269 0.086 0.046 ±
M5 ± 0.0002 ± ± 0.05 0.194 0.108 0.388 0.205 0.333
C ± ± 0.028 ± 0.01 0.104 0.037 0.089 0.086 0.500
T ± ± ± ± ± 0.009 ± 0.046 ± ±
Mortality 0.2433 ± ± ± ± ± ± ±
S, seedling; L, One-leaf; A, Asexual; M, Male; C, Cosexual; T, Two-¯ower.*0.0.
Sexual and clonal reproduction in Fritillaria 33
constant and that of the Nanryu population isslightly increasing.
Figure 2 shows the observed stage class distri-bution and the stage class distribution calculatedfrom the projection matrix (u). They were verysimilar in distribution pattern.
Simulation experiments
In this simulation, values of the ®rst row of thematrix were estimated from data of ®eld census.Mean seed production of Cosexual and Two-¯ower in the Mizuyajiri population were 99.7 and173.0, respectively. Those in the Nanryu popu-lation were 93.0 and 162.0. Overall fruit/¯owerratios for Cosexual and Two-¯ower are 0.85 and1.0 in the Mizuyajiri population and 0.74 and 1.0in the Nanryu population.
The population growth rates calculated on thematrices under nine different germination ratesare shown in Table 5. In this table, germination
rate, 0.0, represents the situation observed innatural populations. A clear critical germinationrate was found in the both populations, where ksuddenly increased. In the Mizuyajiri population,the critical point lies between 0.04 and 0.05, andthat of the Nanryu population lies between 0.02and 0.03.
Mean population growth rates and stablestage class structures were calculated underdifferent probabilities of successful seedlingrecurrent. Table 6 shows the results under thecondition of ®xed germination rate at 5%. Asmentioned above, this value was chosen be-cause it could have a considerable effect on k(Table 5). When successful seedling recruitmentwas satis®ed every year, k increased from 1.006to 1.041 in the Mizuyajiri population, and from1.047 to 1.064 in the Nanryu population. Whenseedling establishment took place occasionally,k did not signi®cantly decrease from the valueof annual seedling establishment. In the Mi-zuyajiri population, when the seedling
Table 5 Effect of different germination rates on the population growth rate �k�Mizuyajiri Nanryu
Germination rate k Dk�%� k Dk�%�0.00 1.006 1.047
0.01 1.014 0.8 1.048 0.1
0.02 1.016 1.0 1.048 0.1
0.03 1.018 1.2 1.063 1.6
0.04 1.020 1.4 1.063 1.6
0.05 1.041 3.5 1.064 1.7
0.06 1.042 3.6 1.065 1.8
0.08 1.046 4.0 1.066 1.9
0.10 1.049 4.3 1.067 2.0
Table 6 Effect of different successful seedling establishment intervals on the population growth rate (k)
Average seedling Mizuyajiri Nanryu
establishment intervals ln�k� � SD k ln�k� � SD k
No seedling 1.006 1.047
Every year 1.041 1.064
1/5 0.036 � 0.008 1.037 0.067 � 0.008 1.069
1/10 0.036 � 0.005 1.037 0.068 � 0.005 1.070
1/15 0.034 � 0.006 1.035 0.068 � 0.004 1.070
Population growth rate was obtained from simple projection matrices for No Seedling and Every Year. Average populationgrowth rates of others were calculated from 50 simulations of 100 iterations each. Germination rate was ®xed at 5% for allsimulations.
34 T. Shimizu et al.
establishment occurred on average once in 5years, k changed from 1.041 to 1.037. Evenwhen the seedling establishment occurred onaverage once in 15 years, k changed from 1.041to 1.035. In the Nanryu population, k tendedto increase as the interval of seedling estab-lishment became longer. Even if different ger-
mination rates, 1% and 3%, were given in thesimulations, the results were almost the same asthat of the germination rate of 5% (Table 7).
These results (Tables 5±7) are summarized asfollows: (i) when germination takes place everyyear, a critical germination rate exists abovewhich seedling recruits provide considerable
Table 7 Comparison of effect of different successful seedling establishment intervals on the population growth rate,k, under the different germination rates
Germination rate
Average seedling Mizuyajiri Nanryu
establishment intervals 1% 3% 1% 3%
1/5 1.027 1.029 1.066 1.067
1/10 1.028 1.029 1.068 1.069
1/15 1.030 1.029 1.068 1.068
Average population growth rates were calculated from 50 simulations of 100 iterations each.
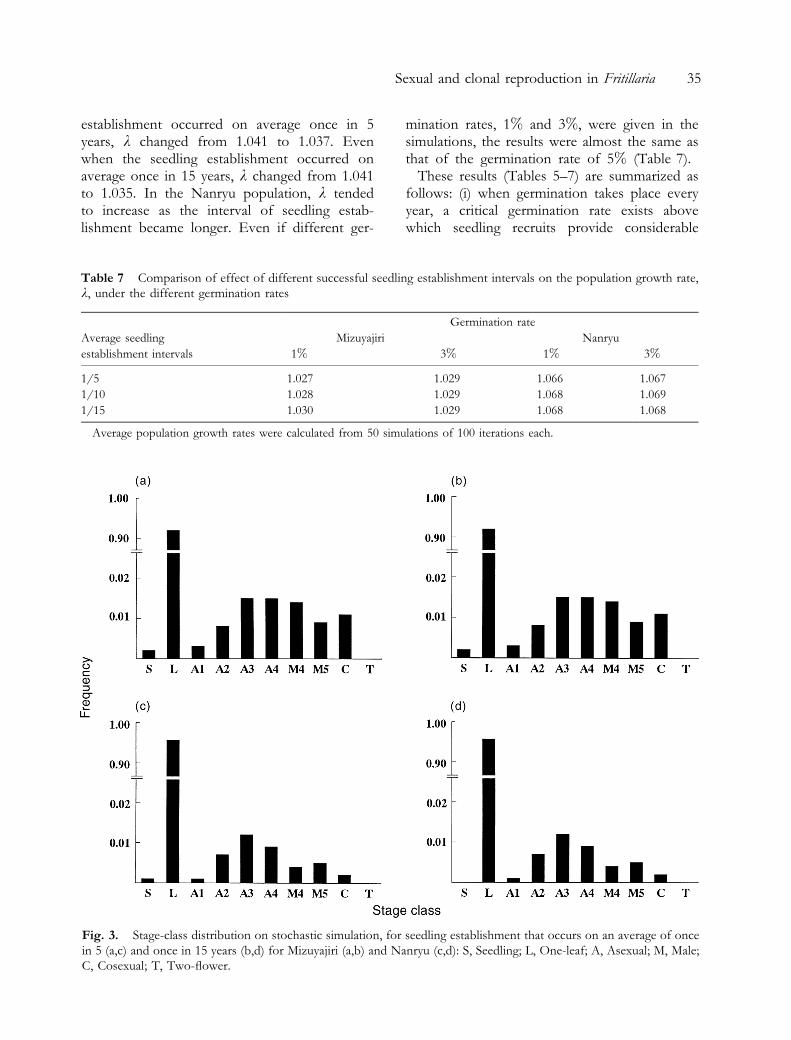
Fig. 3. Stage-class distribution on stochastic simulation, for seedling establishment that occurs on an average of oncein 5 (a,c) and once in 15 years (b,d) for Mizuyajiri (a,b) and Nanryu (c,d): S, Seedling; L, One-leaf; A, Asexual; M, Male;C, Cosexual; T, Two-¯ower.
Sexual and clonal reproduction in Fritillaria 35

effects on k; (ii) even a low recurrent probabilityof seedling establishment has almost the samecontribution to k as high recurrent probabilities;(iii) occasional germination and the subsequentseedling establishment sometimes provide asmuch contribution to k as annual seedling es-tablishment.
Figure 3 shows the stable stage class distribu-tions under two different conditions of seedlingemergence. Stable stage-class distributions werevery similar between both the Mizuyajiri and theNanryu populations. In addition, they were verysimilar to those actually observed (Fig. 2).
DISCUSSION
One of the conspicuous results is that male andcosexual ¯owers were changeable between them(Table 4). This is the ®rst report on sex lability inthis species. Plants show a variety of sex ex-pressions (Lloyd & Bawa 1984), and sex labilityhas been known in some trees and herbs(Schlessman 1988; Policansky 1982).
Liliaceae is a large family distributed all over theworld, to which the genus Fritillaria belongs. Inthis family, ¯owers are usually bisexual (Dahlgrenet al. 1985; Mabberley 1987), while some excep-tions have been known. Like F. camtschatcensis, forexample Fritillaria verticillata var. thunbergii (Naru-hashi 1977) and Zigadenus paniculatus (Emms 1993)have male and cosexual ¯owers, but sex labilityhas not been reported. At present, sex lability in F.camtschatcensis may be a peculiar case in this genus,and also may be the only example in Liliaceae.
Active clonal multiplications were observed inMale compared with Asexual and Cosexual(Table 3). Degree of clonal growth was moreclosely correlated with life-history stages, espe-cially with sexual states than with plant size in F.camtschatcensis. Trade-offs between clonal andsexual reproduction have been assumed (Caswell1985), and Table 3 may show the trade-offs be-tween clonal and sexual reproduction. Male boreonly a staminate ¯ower, and this means that manyresources are available for clonal reproduction.Thus, the evolution of a breeding system in thisspecies, so-called androdioecism, and the evolu-tion of sex lability may have a close relationship
with intensive clonal growth. Although cosexualplants are statistically larger than male plants(Y. Hatanaka, unpubl. data), phenomena andlogic of sex lability in F. camtschatcensis may differfrom those well known in Arisaema (Schaffner1922; Maekawa 1924; Bierzychudek 1981; Kino-shita 1987). Further ecological comparativestudies of this species with other Fritillaria speciesand theoretical studies will be necessary.
In this species, the L stage works as a rametbank. Most of the One-leaf ramets remained inthe same stage next year and other ramets fre-quently reproduced One-leaf by clonal repro-duction (Table 4). More than 90% of ramets wereOne-leaf, and the dynamics of L stage governedwhole dynamics of the population (Tables 1, 4).The role of Asexual is unclear. In many perennialliliaceous species, for example Erythronium (Ka-wano et al. 1987) and Allium (Nault & Gagnon1993), only reproductive individuals have above-ground stems and bear fruit. We supposed at ®rstthat Asexual was a reproductive ramet whichaborted ¯ower buds for some reason. However,Asexual was more frequent than Male and Co-sexual, and this fact suggests that Asexual is notmerely a failure of reproductive ramet but hassome signi®cance in the life history of this species.
Before this study had started, no seedlings wereunexpected in the F. camtschatcensis population onMt Hakusan because a large number of seedswere produced there (Table 2). However, arti®-cial germination in the laboratory was quite dif-®cult, and the majority of seeds would notgerminate (T. Shimada, pers. comm.). This sug-gests that this species may require peculiar con-ditions for seed germination.
In 1996, a small number of seedlings werefound in and around the two populations. Somepossibilities for sudden seedling emergence in1996 can be assumed: (i) seedling emergenceactually occurred in and/or around the quadratsevery year, but they were overlooked; (ii) noseedling emergence occurred around the studysites, whereas seedling emergence actually oc-curred in other places on Mt Hakusan; (iii)seedling emergence occurred every year beforethis study, but could not happen in the wholerange of Mt Hakusan because of the unusualweather; (iv) seedling emergence took place onceevery few or more years when favorable
36 T. Shimizu et al.

conditions were ful®lled. Assumptions (i) and (ii)may be ruled out, since careful attention was paidby many persons to the search for seedlingsaround the permanent quadrats in the wholerange of Mt Hakusan throughout this study. Withrespect to the third assumption, there is a smallpossibility that this could be the case. In 1993 and1994, the weather was extraordinarily bad duringthe past three dozens of years: continuous rain-fall, not very much sunshine and low temperaturein July and August 1993, and quite little rainfalland drought in July and August in 1994.
The fourth assumption may be the most plau-sible explanation, although the third one cannotbe fully ruled out. In any case, long-term obser-vations on seedling emergence are necessary. Inmany other species, no or few seedling recruitsare observed, where population size is mainlymaintained by clonal multiplication (Harper 1977;Cook 1985; Eriksson 1989; Parker & Hamrick1992; Eriksson & Bremer 1993). For example,seedlings of Rubus saxatilis were carefully searchedduring the observation period for 4 years, butseedlings were found in 1 year and only at onesite (Eriksson & Bremer 1993). It was concludedthat a very low rate of seedling recruitment mightbe suf®cient to maintain genetic diversity in apopulation. Such a situation in R. saxatilis is verysimilar to that in F. camtschatcensis.
The fate of seeds is still a black box. We haveno knowledge about seed dispersal, seed-bankdynamics and the conditions of seed germina-tion. Ramet density is very high (about 5000ramets per 8 m2 in Mizuyajiri and 6000 rametsper 13.4 m2 in Nanryu), and there is virtually nospace for seedlings to germinate. As Cook(1985) and Sebens and Thorne (1985) pointedout, new empty habitats were necessary forseedlings to establish successfully, and highdensity may be one of the reasons that seedgermination was not observed for several yearson Mt Hakusan.
We assume that the population size of F. cam-tschatcensis on Mt Hakusan can be maintainedsteadily by clonal reproduction, because k wasmore than 1.0 in both populations: 1.006 for theMizuyajiri and 1.047 for the Nanryu, respectively.Actual data on seed production were obtainedfrom the ®eld census, but there was no reliabledata on seed germination in natural populations.
Thus, some simulation experiments were con-ducted to evaluate the effects of seedling estab-lishment on population growth rate. As we hadno knowledge about seed-bank dynamics, weignored the seed bank and assumed in the sim-ulations that seeds were alive for 1 year and thatthe seed bank was replaced every year.
Simulation experiments demonstrated thatthere was a critical germination rate above whichk suddenly increased (Table 4). The critical valueslay between 2% and 3% in the Nanryu popula-tion and between 4% and 5% in the Mizuyajiripopulation. This means that if germination rate isbelow the critical value, successful seedling es-tablishment can make little contribution tomaintain population; even if a small amount ofseeds germinate every year in natural populations,they have virtually no in¯uence on populationgrowth rate and size distribution, etc.
It was an unexpected result that occasionalseedling establishment (e.g. once in 15 years)could provide almost the same contribution to kas that of annual seedling establishment(Tables 6, 7). In the Nanryu population, k tendedto increase as the interval of seedling establish-ment became longer. Whether or not this iscommon in clonal species is not yet certain.Watkinson and Powell (1993) showed by com-puter simulation experiments that under thecondition of no seedling input, the number ofgenets declined, while the number of rametsslowly converged toward an equilibrium value.They also showed that by year 20 the range ofpopulation size had stabilized depending on thelevel of seedling input. They also argued that itwas the ratio of seedling to ramet recruitmentsrather than the mortality rate of clonal growth orseedling number that determined the architectureof the population of clonal species. Our results,like those by Watkinson and Powell (1993),supported that occasional seedling establishmentin clonal plants played a very small role in de-termining the number and the stage structure of apopulation.
In conclusion, the natural populations ofF. camtschatcensis on Mt Hakusan can be mainlymaintained by clonal reproduction. However, it isnot yet certain whether seeds germinate everyyear or whether there is a ¯ush seedling emer-gence occurring once every a few or more years
Sexual and clonal reproduction in Fritillaria 37

in the natural populations. In any case, seedlingsin this species contribute little to maintainingpopulation size compared to that usually seen inmany plants. Maybe the role of sexual repro-duction lies beyond maintaining population size.To ascertain these problems, long-term ®eldcensus on seedling number will be necessary.
ACKNOWLEDGEMENT
We are very grateful to the authorities of Shir-amine-mura and Hakusan Kankoh-Kyokai, es-pecially to all those who were concerned withNanryu-Sanso and Murodo, for providing us withvarious facilities during the ®eld survey onMt Hakusan, 1993±1996. Shirayamahime-Jinjaand Hakusan National Park Administrative Of-®ce of the Environmental Agency offeredpermissions and facilities for collecting sampleplants for which we are grateful. We thankmembers of the Laboratory for Plant NaturalHistory, the Laboratory for Animal Ecology, andthe Botanic Garden of Kanazawa University,Kanazawa school of Nihon Dobutsu-ShokubutsuSenmon Gakuin for various support throughoutthis study. This study was supported by the JointResearch Utilizing Science and TechnologyPotential in Region from the Science and Tech-nology Agency, Japan.
REFERENCES
ABRAHAMSON W. G. (1980) Demography and vegeta-tive reproduction. In: Demography and Evolution inPlant Populations (ed. O. T. Solbrig) pp. 89±106.Blackwell Science, Oxford.
ANGEVINE M. W. & HANDEL S. N. (1986) Invasion offorest ¯oor space, clonal architecture, andpopulation growth in the perennial herb Clintoniaborealis. Journal of Ecology 74: 547±560.
BIERZYCHUDEK P. (1981) The demography of jack-in-the-pulpit, a forest perennial that changes sex.Ecological Monographs 52: 335±351.
CASWELL H. (1985) The evolutionary demography ofclonal reproduction. In: Population Biology andEvolution of Clonal Organisms (eds J. B. C. Jackson,L. W. Buss & R. E. Cook) pp. 187±224. YaleUniversity Press, New Haven.
CASWELL H. (1989) Matrix Population Models. Sinauer,Sunderland, MA.
COOK R. E. (1985) Growth and development in clonalplants. In: Population Biology and Evolution of ClonalOrganisms (eds J. B. C. Jackson, L. W. Buss & R. E.Cook) pp. 259±296. Yale University Press, NewHaven.
DAHLGREN R. M. T., CLIFFORD H. T. & YEO P. F.(1985) The Families of the Monocotyledons. Springer-Verlag, Berlin.
EMMS S. K. (1993) Andromonoecy in Zigadenuspaniculatus (Liliaceae): spatial and temporal patternsof sex allocation. American Journal of Botany 76: 30±39.
ERIKSSON O. (1988) Ramet behavior and populationgrowth in the clonal herb Potentilla anserina. Journal ofEcology 76: 522±536.
ERIKSSON O. (1989) Seedling dynamics and lifehistories in clonal plants. Oikos 55: 231±238.
ERIKSSON O. & BREMER B. (1993) Genet dynamics ofthe clonal plant Rubus saxatilis. Journal of Ecology 81:533±542.
HARPER J. L. (1977) Population Biology of Plants.Academic Press, London.
KAWANO S., TAKADA T., NAKAYAMA S. & HIRATSUKA
A. (1987) Demographic differentiation and life-history evolution in temperate woodland plants. In:Differentiation Pattern in Higher Plants (eds M.Urbanska) pp. 152±181. Academic Press, London.
KINOSHITA E. (1987) Sex change and populationdynamics in Arisaema (Araceae). I. Arisaema serratum(Thunb.) Schott. Plant Species Biology 2: 15±28.
LLOYD D. G. & BAWA K. S. (1984) Modi®cations ofthe gender of seed plants in varying conditions.Evolutionary Biology 17: 255±338.
LOVETT-DOUST J. & LAPORTE G. (1991) Populationsex ratios, population mixtures and fecundity in aclonal dioecious macrophyte, Vallisneria americana.Journal of Ecology 79: 477±489.
MABBERLEY D. J. (1987) The Plant Book. CambridgeUniversity Press, Cambridge.
MAEKAWA T. (1924) On the phenomena of sextransition in Arisaema japonica. Journal of College ofAgriculture Hokkaido Imperial University 13: 217±305.
MAYNARD SMITH J. (1978) The Evolution of Sex.Cambridge University Press, Cambridge.
NARUHASHI N. (1977) Andromonoecium of Fritillariaverticillata var. thunbergii. Journal of Geobotany 25: 27±32(in Japanese with English summary).
NAULT A. & GAGNON D. (1993) Ramet demographyof Allium tricoccum, a spring ephemeral, perennialherb. Journal of Ecology 81: 101±119.
PARKER K. C. & HAMRICK J. L. (1992) Geneticdiversity and clonal structure in a Cilumnar cactus,
38 T. Shimizu et al.

Lophocereus schottii. American Journal of Botany 79: 86±96.
PITELKA L. F., HANSEN S. B. & ASHMUN J. W. (1985)Population biology of Clintonia borealis. I. Ramet andpatch dynamics. Journal of Ecology 73: 169±183.
POLICANSKY D. (1982) Sex change in plants andanimals. Annual Review of Ecology and Systematics 13:471±495.
SCHAFFNER J. H. (1922) Control of sexual state inArisaema triphyllum and A. dracontium. American Journalof Botany 9: 72±78.
SCHLESSMAN M. A. (1988) Gender diphasy (`SexChoice'). In: Plant Reproductive Ecology (eds J. LovettDoust & L. Lovett Doust) pp. 139±153. OxfordUniversity Press, New York.
SEBENS K. P. & THORNE B. L. (1985) Coexistenceof clones, clonal diversity, and the effects ofdisturbance. In: Population Biology and Evolution ofClonal Organisms (eds J. B. C. Jackson, L. W. Buss &R. E. Cook) pp. 357±398. Yale University Press,New Haven.
SHIMIZU T. (1983) The New Alpine Flora of Japan in ColorVol. II. Hoikusha, Osaka (in Japanese).
SHIMIZU T. & UMEBAYASHI M. (1995) Supplement to`Underground Organs of Herbaceous Angiosperms'
(1). Annual Report of Botanic Garden, Faculty of Science,Kanazawa University 18: 1±6 (in Japanese).
SILANDER J. A. (1985) Micro evolution in clonalplants. In: Population Biology and Evolution of ClonalOrganisms (eds J. B. C. Jackson, L. W. Buss & R. E.Cook) pp. 107±152. Yale University Press, NewHaven.
SUSKO C. L. D., LOVETT-DOUST L. & LOVETT-DOUST
J. (1994) Population genetic structure of Vallisneriaamericana, a dioecious clonal macrophyte. AmericanJournal of Botany 81: 1004±1012.
SUZUKI T. (1970) Die P¯anzengesellschaften und dievertikale Vegetationsstufe vom Hakusan-Gebirge.In: Hakusan no Shizen. pp. 114±156. IshikawaPrefectural Of®ce, Kanazawa.
WATKINSON A. R. & POWELL J. C. (1993) Seedlingestablishment and the maintenance of clonaldiversity in plant populationsÐa computersimulation of Ranunculus repens. Journal of Ecology 81:707±717.
YASHIMA T., KINOSHITA E. & SHIMIZU T. (1997)Flowering phenology and self-incompatibility inFritillaria camtschatcensis (L.) Ker-Gawl. Journal ofPhytogeography and Taxonomy 45: 129±133.
Sexual and clonal reproduction in Fritillaria 39






























