Embed Size (px)
Citation preview

.sciencedirect.com
f u n g a l e c o l o g y 6 ( 2 0 1 3 ) 2 8 1e2 9 2
available at www
journal homepage: www.elsevier .com/locate/ funeco
The cultivable endophytic community of Norway spruceectomycorrhizas from microhabitats lacking ericaceous hostsis dominated by ericoid mycorrhizal Meliniomyces variabilis
Martin VOHN�IKa,b,*, Libor MRNKAa, Tereza LUKE�SOV�Aa,b, Mar�ıa Clara BRUZONEc,Petr KOHOUTa,b,1, Judith FEHRERd
aDepartment of Mycorrhizal Symbioses, Institute of Botany ASCR, Pr�uhonice 252 43, Czech RepublicbDepartment of Plant Experimental Biology, Faculty of Science, Charles University in Prague, Prague 128 44, Czech RepubliccLaboratorio de Microbiolog�ıa Aplicada y Biotecnolog�ıa, Centro Regional Universitario Bariloche, INIBIOMA (Universidad Nacional del
Comahue e CONICET), San Carlos de Bariloche, R�ıo Negro, ArgentinadDNA Laboratory, Institute of Botany ASCR, Pr�uhonice 252 43, Czech Republic
a r t i c l e i n f o
Article history:
Received 21 August 2012
Revision received 7 February 2013
Accepted 22 February 2013
Available online 14 May 2013
Corresponding editor:
Kevin K. Newsham
Keywords:
Cadophora finlandica
Dark septate endophytes
Ectomycorrhiza
Endophytic fungi
Ericaceae
Ericoid mycorrhiza
Meliniomyces bicolor
Rhizoscyphus ericae
Root associated fungi
* Corresponding author. Department of Myco271015331.
E-mail addresses: [email protected] Present address: Group of Mycology, Inst
1754-5048/$ e see front matter ª 2013 Elsevhttp://dx.doi.org/10.1016/j.funeco.2013.03.006
a b s t r a c t
Most of the temperate conifers associate with ectomycorrhizal fungi, but their roots also
harbour a wide range of endophytes. We focused on ascomycetes associating with basidio-
mycetous ectomycorrhizasofNorway spruce ina temperatemontane forest in central Europe
and found that the majority of the co-associated fungi belonged to the Rhizoscyphus ericae
aggregate (REA), being dominated by Meliniomyces variabilis. We further tested the ability of
representative isolates to colonize spruce root tips and European blueberry (Vaccinium myr-
tillus) hair roots in an agar system as well as their effect on blueberry growth in a peat-agar
system. M. variabilis intracellularly colonized spruce (Picea abies) root tip cortex, formed eri-
coidmycorrhizas in blueberry and enhanced blueberry shoot and root growth in comparison
with non-inoculatedplants. Our findings suggest that spruce ectomycorrhizasmay represent
selective niches for ericoid mycorrhizal fungi in habitats lacking suitable ericaceous hosts.
ª 2013 Elsevier Ltd and The British Mycological Society. All rights reserved.
Introduction range from facultative saprobes and parasites to neutral
Any healthy or dead plant tissue contains fungal endobionts
(hereafter referred to as mycobionts), both above- and below-
ground (Sieber 2002; Summerbell 2005). These mycobionts
rrhizal Symbioses, Instit
s.cz, [email protected] (itute of Ecology and Earthier Ltd and The British M
endophytes to mutualistic symbionts positively affecting host
plant fitness (Rodriguez et al. 2009; Newsham 2011). While
there seems to be only a small overlap between the fungi
inhabiting plant stems and roots (Sieber 2002, 2007), some
ute of Botany ASCR, Pr�uhonice 252 43, Czech Republic. Tel.: þ420
M. Vohn�ık).Sciences, University of Tartu, 14a Ravila, Tartu 50411, Estonia.
ycological Society. All rights reserved.

282 M. Vohn�ık et al.
widespread mycobionts apparently colonize both aerial and
underground plant tissues (Menkis et al. 2004; Rodriguez et al.
2009). Aerial mycobionts can influence longevity of plant tis-
sues by protection against water stress and herbivores and
may accelerate their senescence (Carroll 1988; Arnold et al.
2003; Rodriguez et al. 2009), thus directly affecting plant bio-
mass available for carbon capture through photosynthesis. In
contrast, root mycobionts directly influence plant mineral
nutrition through scavenging nutrients from recalcitrant
substrates unavailable to host plants (mycorrhizal fungi),
protecting roots from biotic and abiotic stresses (mycorrhizal
and endophytic fungi) and/or influencing their longevity
(mycorrhizal fungi, fungal root endophytes, parasites and
casual saprobes) (Addy et al. 2005; Smith & Read 2008).
Roots of the large majority of terrestrial plants form
mutualistic symbioses with mycorrhizal fungi which pro-
foundly affect their anatomy, morphology, distribution and
metabolism (Brundrett 2002). In temperate and boreal forests,
most dominant trees form ectomycorrhizas (EcM). EcM fungi
mostly belong to the Basidiomycota with a relatively smaller
part belonging to the Ascomycota (LoBuglio et al. 1996;
Tedersoo et al. 2006). Ectomycorrhizas additionally harbour
diverse spectra of co-associated, mostly ascomycetous fungi
(Kernaghan et al. 2003; Urban et al. 2008; Tedersoo et al. 2009;
Kernaghan & Patriquin 2011). Due to their fast growth on
nutrient media, these may be preferentially isolated from the
respective ectomycorrhizas and mistaken for true EcM fungi
(Summerbell 2005 and references therein). Likewise, DNA of
the co-associated fungi may interfere with DNA of true EcM
mycobionts during PCR and/or may be preferentially ampli-
fied by the commonly used fungal primers (Gr€unig et al. 2008a;
Tedersoo et al. 2009). Significance of the EcM co-associated
fungi is largely unknown, but was hypothesized to include
specific inter-fungal interactions helping to maintain EcM
fungi diversity (Urban et al. 2008) or alteration of nutrient
uptake by the respective ectomycorrhizas (Jones et al. 2009).
Intriguingly, EcM co-associated fungi often display affinities to
Helotiales which commonly inhabit roots of Ericaceae
(Kernaghan et al. 2003; Addy et al. 2005; Tedersoo et al. 2009)
and at least some of them seem to be capable of forming eri-
coid mycorrhizas under controlled conditions (Vohn�ık et al.
2007a; Grelet et al. 2009).
Although EcM co-associated fungi comprise wide spectra
of fungal symbionts, two assemblages seem to be more fre-
quent in temperate and boreal forests of the Northern Hemi-
sphere: members of the Rhizoscyphus ericae aggregate (REA)
(Vr�alstad et al. 2000) and the dark septate endophytes (DSE)
displaying affinities or belonging to the Phialocephala fortinii
s.l.eAcephala applanata species complex (PAC) (Gr€unig et al.
2008a). Both groups contain species able to form ectomycor-
rhizas with developed Hartig net; Meliniomyces bicolor
(belonging to REA) produces the Piceirhiza bicolorata EcM mor-
photype (Vr�alstad et al. 2000; Villarreal-Ruiz et al. 2004) and
Acephala macrosclerotiorum (related to PAC) forms the Pinirhiza
sclerotia EcM morphotype (M€unzenberger et al. 2009). In addi-
tion, REA comprises the well-known ericoidmycorrhizal (ErM)
fungus R. ericae, and there is evidence supporting ericoid
mycorrhizal status of M. bicolor and M. variabilis (Grelet et al.
2009). On the other hand, despite DSE being frequently
observed, isolated and molecularly detected in ericaceous
roots, and that some of them seem to form intracellular
hyphal coils resembling ericoid mycorrhizas (Usuki &
Narisawa 2005; Vohn�ık & Albrechtov�a 2011), it is so far
unclear whether they form functional mycorrhizal symbioses
with Ericaceae.
In contrast to arbuscular mycorrhizal and EcM fungi, we
know very little about the extent, distribution and fate of the
mycelium of ErM fungi in the soil environment. It is believed
that the ErM extraradical mycelium reaches only a few milli-
metre from the root surface into the surrounding substratum
(Smith & Read 2008), and it has been suggested that genets of
M. variabilis are relatively small (<13 cm; Grelet et al. 2010).
While Oidiodendron maius, a typical ErM fungus, is an efficient
saprobe which can persist in the soil without contact with
Ericaceae (Rice & Currah 2006), the REAmembers are typically
detected in plant roots. Congruently, Bergero et al. (2003) found
that a consortium of ErM fungi persisted and maintained
mycorrhizal ability in the soil in a Mediterranean habitat
lacking ericaceous hosts but the authors failed to detect any
REA members. On the other hand, there is conflicting infor-
mation about the effect of Ericaceae on the REA members
interacting with EcM roots. For example, Kohout et al. (2011)
reported that the presence of Vaccinium spp. inhibited the
ability of M. bicolor to form ectomycorrhizas with Pinus spp.
while genetically identical isolates of Meliniomyces variabilis
were commonly detected in neighbouring Pinus sylvestris
ectomycorrhizas and Vaccinium vitis-idaea hair roots (Grelet
et al. 2010). Although several authors have hypothesized that
Ericaceae might be interconnected with neighbouring EcM
conifers through shared mycelium of some REA mycobionts,
especially Cadophora finlandica and M. bicolor (e.g., Villarreal-
Ruiz et al. 2004; Vr�alstad 2004), annotations of ITS nrDNA
sequences deposited in GenBank suggest that these two spe-
cies are only rarely detected in Ericaceae roots, in contrast to
M. variabilis and R. ericae (cf. Suppl. Fig 1). These findings
suggest possible differences within the R. ericae aggregate in
preferences for different host plants as well as in the ability to
develop and persist in the soil environment outside host roots
(cf. Baldrian et al. 2012).
The Bohemian Forest NP lies in a mountain range
extending fromSouth Bohemia, the Czech Republic, to Austria
and Germany. It comprises forested hills/mountains at an
elevation between 800 and 1 400 m a.s.l. and covers an area of
approx. 700 km2. Most of its montane ecosystems are formed
by secondary Norway spruce (Picea abies) forests with erica-
ceous, grass and hygrophilous understorey, such vegetation
being characteristic for many European temperate and boreal
regions. Detailed information on fungi associating with ecto-
mycorrhizas in this ecosystem type is lacking; to our knowl-
edge, the only local report is from serpentinite sites in Austria
(Urban et al. 2008). We, therefore, investigated the diversity of
ascomycetous fungi living in endorhizal symbiosis with Nor-
way spruce basidiomycetous ectomycorrhizas at two sites in
the Bohemian Forest NP. Basidiomycetous EcMs were chosen
as they form the great majority of spruce ectomycorrhizas at
the two sites (M. Vohn�ık, personal observation) and basidio-
mycetous EcM mycelia seem to dominate the fungal soil
community detected at a nearby experimental site (Baldrian
et al. 2012). We specifically focused on microsites lacking eri-
caceous understory which led us to the expectation that the

Fig 1 e Symbiotic potential of selected isolates obtained in this study in roots of Picea abies and Vaccinium myrtillus
seedlings. (A) Acephala applanata EF093158 D P. abies: intercellular hyphae in the cortex (arrows) accompanied by a circle of
microsclerotia (asterisks) around the endodermis. Hand-sectioned root tip, bar [ 50 mm. (B) Meliniomyces variabilis
EF093169 D P. abies: intercellular hyphae in the cortex cells (arrows). Hand-sectioned root tip stained with trypan blue,
bar [ 50 mm. (C) Typical morphology of P. abies roots colonized by Cadophora finlandica EF093179. Bar [ 500 mm. (D)
Cadophora finlandica EF093179 D P. abies: intercellular hyphae forming Hartig net (arrows) accompanied by intracellular
hyphae (arrowheads) in the cortex. Hand-sectioned root tip stained with aniline blue, bar [ 12.5 mm. (E) Meliniomyces
variabilis EF093169 D V. myrtillus: an intracellular hyphal coil in a rhizodermal cell, morphologically identical to ericoid
mycorrhiza. Stained with trypan blue, bar [ 25 mm. (F) Cadophora finlandica EF093179 D V. myrtillus: the rhizodermal cells
are filled with dense hyphal loops identical to ericoid mycorrhizas. Stained with trypan blue, bar [ 25 mm. (G) Cadophora
finlandica EF093155 D V. myrtillus: melanized dark septate hyphae forming loose intracellular loops while spreading from
cell to cell in the rhizodermis. An arrow points at a thin hypha penetrating the host cell wall. Stained with trypan blue,
bar [ 20 mm. (H) Meliniomyces bicolor EF093180 D V. myrtillus: rhizodermal cells filled with dense hyphal coils
morphologically identical to ericoid mycorrhiza. Stained with trypan blue, bar [ 25 mm. (I) Acephala applanata
EF093158 D V. myrtillus: dark septate hyphae occasionally forming intracellular microsclerotia (asterisks). Bar [ 30 mm.
Meliniomyces variabilis in spruce ectomycorrhizas 283

284 M. Vohn�ık et al.
endophytic fungal community would be dominated by non-
ericaceous mycobionts. As we intended to screen endo-
phytes living inside the screened ectomycorrhizas, rather
than their superficial associates which are often detected by
culture-independent approaches, we employed a culture-
dependent approach which additionally favoured growth of
ascomycetes over basidiomycetes. Isolates of EcM co-
associated fungi were also needed to test their symbiotic
interactions with Norway spruce and European blueberry
(Vaccinium myrtillus). In the symbiotic tests, we wanted to find
out whether: (1) the isolates have endophytic ability, i.e., they
can intracellularly colonize spruce root tips without apparent
negative effects e this would mean that they are likely true
endophytes of the respective ectomycorrhizas from which
they were isolated; (2) at least some isolates have ectomy-
corrhizal ability, i.e., they form the Hartig net e this would
mean that they could participate in formation of the respec-
tive ectomycorrhizas; and (3) at least some isolates have the
ericoid mycorrhizal ability, i.e., they form the characteristic
morphological structures in Ericaceae hair roots and pos-
itively affect the colonized host plants e this wouldmean that
ectomycorrhizas could serve as micro-reservoirs of functional
ericoid mycorrhizal inoculum in habitats lacking suitable
ericaceous hosts.
Materials and methods
Field site and root sampling
The two investigated sites are located in the central part
of the Bohemian Forest NP at an elevation of approx.
1 075 m a.s.l (N49�00, E13�290 and N48�590, E13�290). The valley
is dominated by a secondary Norway spruce (P. abies) forest
(>80 yr old) with understory consisting of ericaceous plants
(V. myrtillus and V. vitis-idaea), grass (mostly Avenella flexuosa
and Calamagrostis villosa), and bryophytes (Sphagnum spp.);
the prevailing soil type is podzol. In total, 19 3e5 yr old spruce
seedlings inhabiting different niches within the two sites
were sampled on two occasions during the vegetation sea-
son. The different niches included dead Norway spruce
stumps, mosses, fallen decaying spruce trunks, the forest
floor covered by spruce needles and waterlogged Sphagnum
undergrowth. These microhabitats were chosen to capture
the heterogeneous ecological amplitude of spruce seedlings
regenerating within the National Park. However, we omitted
spruce seedlings regenerating in the vicinity of Ericaceae or
directly within ericaceous shrubs as these situations are a
subject of another ongoing study. Seedlings were carefully
dug up not to destroy their fine ectomycorrhizal roots,
packed in plastic bags and kept refridgerated until the iso-
lation of the ectomycorrhiza co-associated fungi on the fol-
lowing day.
Isolation of fungal cultures
The roots of the seedlings were thoroughly washed with tap
water, transferred to250mlflaskswith100mlofdistilledwater
with two drops of Tween 20 and shaken for 10 min. Basidio-
mycetous EcM root tips were recognized by the presence of
clamped hyphal mantles and/or emanating clamped hyphae
and typical EcM root tip morphology. According to their mor-
phology (Agerer 1987e2002, A. Lep�sov�a pers. com.) the collected
ectomycorrhizas belonged to Amphinema byssoides, Hebeloma
velutipes, Lactarius spp., Piceirhiza guttata (¼Tylospora fibrillosa),
Russula ochroleuca and Thelephora terrestris. In total, 360 basi-
diomycetous EcM tips of healthy appearance were randomly
selected from the collected seedlings, cut from the main roots
(length approx. 3 mm), surface-sterilized (0.5 % NaClO for
10 min), serially washed in sterile deionized water, aseptically
placed onto modified Melin Norkrans medium (MMN), 15 EcM
tips per 9 cm diam. Petri dish, and incubated for 2 months at
20 �C in the dark. The dishes were periodically checked for
mycelial growth and possible occurrence of contaminants.
According to Schild et al. (1988) and our previous testing, this
medium favours the growth of ascomycetes over basidiomy-
cetes, while addition of benomyl leads to a higher yield of
putatively EcM basidiomycetes. Fast growing and intensively
sporulating fungi were discarded and the remaining isolates
were transferred onto new MMN media for subculture. Their
radial growth onmalt extract agar (SigmaeAldrich, Germany),
MMN, corn meal agar, oatmeal agar and potato dextrose agar
(all HiMedia Laboratories, India) was measured each second
day for 3 weeks, and mycelial characteristics were recorded
after 2 weeks of growth. The isolates were then conservatively
grouped intodistinct culturemorphotypesandrepresentatives
of eachmorphotype (2e3 isolates if available)were subjected to
molecular identification.
DNA extraction and amplification
Fungal DNA was extracted from fresh mycelia using the sor-
bitol method (�Storchov�a et al. 2000). DNA was dissolved in
10 mM Tris/HCl (pH 8.0) and kept in a freezer (�20 �C). DNA
amplifications of the ITS nrDNA regionswere performed using
an Eppendorf Master Cycler (Eppendorf AG, Germany) using
the primers ITS1F and ITS4. The PCR mix consisted of 5 ml of
10� PCR buffer without MgCl2 (Fermentas International Inc,
Canada), 200 nM concentration of dNTPmixture, 2 mMMgCl2,
0.8 mM of each primer, 1 U of Taq DNA polymerase, 32 ml of
sterile ddH2O and 4 ml of 1:4 diluted template DNA. The fol-
lowing cycling parameters were used: initial denaturation
step for 4 min at 94 �C, 35 cycles consisting of a denaturation
step at 94 �C for 30 s, annealing at 55 �C for 30 s, extension at
72 �C for 70 s, with a final extension at 72 �C for 10 min. PCR
products were checked for length and quality/quantity by gel
electrophoresis (1.5 % agarose) and purified using the High
Pure PCR product purification kit (Roche Holding Ltd, Swit-
zerland). Sequencing was done by GATC Biotech AG (Ger-
many) using the PCR primers, sequence electropherograms
were edited manually. The obtained sequences were sub-
mitted to the GenBank database (accession numbers
EF093147eEF093185, Table 1).
Species identification
As a first step towards species identification, the obtained
ITS sequences were subjected to BLAST searches (GenBank,
NCBI) to retrieve the most similar sequences. Most of them
could be assigned to particular species (sequence identity
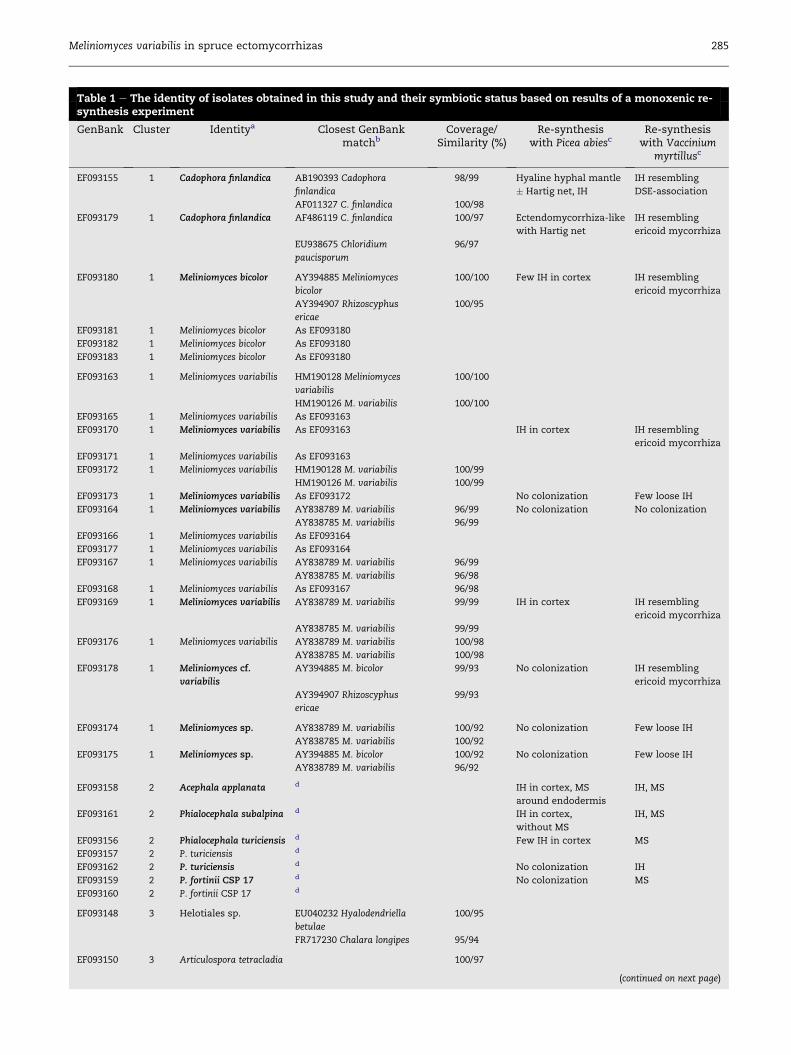
Table 1 e The identity of isolates obtained in this study and their symbiotic status based on results of a monoxenic re-synthesis experiment
GenBank Cluster Identitya Closest GenBankmatchb
Coverage/Similarity (%)
Re-synthesiswith Picea abiesc
Re-synthesiswith Vaccinium
myrtillusc
EF093155 1 Cadophora finlandica AB190393 Cadophora
finlandica
98/99 Hyaline hyphal mantle
� Hartig net, IH
IH resembling
DSE-association
AF011327 C. finlandica 100/98
EF093179 1 Cadophora finlandica AF486119 C. finlandica 100/97 Ectendomycorrhiza-like
with Hartig net
IH resembling
ericoid mycorrhiza
EU938675 Chloridium
paucisporum
96/97
EF093180 1 Meliniomyces bicolor AY394885 Meliniomyces
bicolor
100/100 Few IH in cortex IH resembling
ericoid mycorrhiza
AY394907 Rhizoscyphus
ericae
100/95
EF093181 1 Meliniomyces bicolor As EF093180
EF093182 1 Meliniomyces bicolor As EF093180
EF093183 1 Meliniomyces bicolor As EF093180
EF093163 1 Meliniomyces variabilis HM190128 Meliniomyces
variabilis
100/100
HM190126 M. variabilis 100/100
EF093165 1 Meliniomyces variabilis As EF093163
EF093170 1 Meliniomyces variabilis As EF093163 IH in cortex IH resembling
ericoid mycorrhiza
EF093171 1 Meliniomyces variabilis As EF093163
EF093172 1 Meliniomyces variabilis HM190128 M. variabilis 100/99
HM190126 M. variabilis 100/99
EF093173 1 Meliniomyces variabilis As EF093172 No colonization Few loose IH
EF093164 1 Meliniomyces variabilis AY838789 M. variabilis 96/99 No colonization No colonization
AY838785 M. variabilis 96/99
EF093166 1 Meliniomyces variabilis As EF093164
EF093177 1 Meliniomyces variabilis As EF093164
EF093167 1 Meliniomyces variabilis AY838789 M. variabilis 96/99
AY838785 M. variabilis 96/98
EF093168 1 Meliniomyces variabilis As EF093167 96/98
EF093169 1 Meliniomyces variabilis AY838789 M. variabilis 99/99 IH in cortex IH resembling
ericoid mycorrhiza
AY838785 M. variabilis 99/99
EF093176 1 Meliniomyces variabilis AY838789 M. variabilis 100/98
AY838785 M. variabilis 100/98
EF093178 1 Meliniomyces cf.
variabilis
AY394885 M. bicolor 99/93 No colonization IH resembling
ericoid mycorrhiza
AY394907 Rhizoscyphus
ericae
99/93
EF093174 1 Meliniomyces sp. AY838789 M. variabilis 100/92 No colonization Few loose IH
AY838785 M. variabilis 100/92
EF093175 1 Meliniomyces sp. AY394885 M. bicolor 100/92 No colonization Few loose IH
AY838789 M. variabilis 96/92
EF093158 2 Acephala applanata d IH in cortex, MS
around endodermis
IH, MS
EF093161 2 Phialocephala subalpina d IH in cortex,
without MS
IH, MS
EF093156 2 Phialocephala turiciensis d Few IH in cortex MS
EF093157 2 P. turiciensis d
EF093162 2 P. turiciensis d No colonization IH
EF093159 2 P. fortinii CSP 17 d No colonization MS
EF093160 2 P. fortinii CSP 17 d
EF093148 3 Helotiales sp. EU040232 Hyalodendriella
betulae
100/95
FR717230 Chalara longipes 95/94
EF093150 3 Articulospora tetracladia 100/97
(continued on next page)
Meliniomyces variabilis in spruce ectomycorrhizas 285

Table 1 e (continued )
GenBank Cluster Identitya Closest GenBankmatchb
Coverage/Similarity (%)
Re-synthesiswith Picea abiesc
Re-synthesiswith Vaccinium
myrtillusc
EU998918 Articulospora
tetracladia
EU998917 A. tetracladia 100/97
EF093184 3 Varicosporium elodeae GQ152148 V. elodeae 95/100 No colonization No colonization
GQ411275 V. elodeae 93/100
EF093185 3 V. elodeae As EF093184 No colonization No colonization
EF093147 3 cf. Neobulgaria HM051080 N. pura 89/97
HM116745 Neobulgaria sp. 99/93
EF093149 3 cf. Neobulgaria As EF093147
EF093153 3 Tolypocladium inflatum AB255606 Tolypocladium
inflatum
100/100
AB103381 T. inflatum 100/100
EF093154 3 Thysanophora
penicillioides
AB213268 Thysanophora
penicillioides
99/99
AB213265 T. penicillioides 99/99
EF093151 4 Hebeloma sp. AF430291 Hebeloma
incarnatulum
98/99
(Basidiomycota) AY309960 Hebeloma bryogenes 96/100
EF093152 4 Mycena galopus HM240534 M. galopus 96/99
(Basidiomycota) JF908484 M. galopus 94/99
a According to BLAST and/or phylogenetic analyses; the strains in bold were used in monoxenic cultures with Picea abies and Vaccinium
myrtillus.
b Preference was given for sequences derived from cultures (with deposited vouchers if available) with scientific names.
c IH ¼ intracellular hyphae; MS ¼ microsclerotia; DSE ¼ dark septate endophytes.
d These sequences fell within the Phialocephala fortiniieAcephala applanata species complex. As ITS nrDNA sequences are not suitable for
classification of Phialocephala-like strains to cryptic species, they were identified according to Gr€unig et al. (2008b).
286 M. Vohn�ık et al.
�97 %). For a more reliable identification, the isolates
belonging to the R. ericae aggregate were subjected to phy-
logenetic analyses. Further sequences, derived preferentially
from cultured isolates deposited in international culture
collections, as well as suitable outgroup taxa were retrieved
from GenBank to represent the genetic variability of the
isolates as comprehensively as possible (Suppl. Table 1).
Sequences were aligned manually using BioEdit V7.0.0 (Hall
1999). Maximum parsimony (MP) and minimum evolution
(ME) analyses were done with PAUP* V4.0b10 (Swofford
2002). MP analysis was performed as a heuristic search
with 100 random addition sequence replicates, saving no
more than 100 trees of length greater than or equal to 1 per
replicate, and TBR branch swapping. Bootstrapping was
done with the same settings and 1 000 replicates, but with-
out branch swapping. ME analysis was run as a heuristic
search using HKY85 distances with TBR branch swapping,
bootstrapping was done as for MP analysis. Bayesian analy-
sis was done using MrBayes V3.1 (Ronquist & Huelsenbeck
2003) with six substitution rates, gamma distribution and a
proportion of invariable sites as priors. MCMC analyses were
performed in two parallel runs with four chains each for 4
million generations, sampling every 1 000th tree. All stat-
istical parameters indicated that convergence was reached.
The first 25 % of the trees were discarded as burn-in, and the
remaining 75 % trees were summarized. Fungal isolates
belonging to the P. fortinii s.l.eA. applanata species complex
were kindly identified to cryptic species by C. Gr€unig
according to Gr€unig et al. (2008b).
Symbiotic potential of selected isolates in Picea andVaccinium
P. abies and V. myrtillus seedswere surface-sterilizedwith 30 %
hydrogen peroxide for 10 and 3 min, respectively, and placed
on MMN agar to check for possible contamination. Non-
contaminated seeds were left to germinate for 3 weeks.
Lower compartments of split Petri dishes (9 cm in diam.) with
perforated central septa were filled with MMN (10� diluted
sugars), overlaid with a sterile cellophane membrane to pre-
vent growth of roots into the medium and inoculated with
agar plugs cut from margins of colonies of 17 selected fungal
isolates actively growing on MMN (Table 1). The dishes with
the plugs were left for 3 weeks at room temperature in the
dark to produce vegetative mycelium. Then, 3-week-old
seedlings were transferred to the dishes (five seedlings per
each isolate/host combination), the empty upper compart-
ments accommodating their shoots and the lower compart-
ments their roots. The roots of the seedlings were covered
with a piece of sterile moistened filter paper. The dishes were
sealed with air-permeable film, lower compartments covered
with aluminium foil, and placed in a vertical position in a
growth chamber (16/8 hr and 21/15 �C day/night cycle, irra-
diation 250 m mol m�2 s�1). After 5 months, the plants were
extracted and their roots were separated and processed as
follows: V. myrtillus roots were cleared with 10 % KOH (20 min
at 121 �C), washed with tap water, acidified (1 min in 3 % HCl),
washed with tap water, stained with trypan blue (1 hr at
121 �C) and de-stained overnight in lactoglycerol; P. abies roots

Meliniomyces variabilis in spruce ectomycorrhizas 287
were hand-sectioned and thin sections were stained with
aniline or trypan blue. Both dyes were prepared in lactogly-
cerol (lactic acid:glycerol:deionized water ¼ 1:1:3) as 0.05 %
solutions. The stained roots and thin sections were observed
using an Olympus BX60microscope equippedwith DIC at high
magnifications (400� and 1 000�). Pictures were taken with an
Olympus DP70 camera, graphic documentation was modified
for clarity in Paint.NET (Brewster, Jackson and contributors þMicrosoft Corporation) when needed.
Effect of selected isolates on V. myrtillus growth
The potential ericoid mycorrhizal status of selected REA iso-
lateswas also tested in a peat-agar system. Briefly, 10ml of 1 %
(w/v) autoclaved water agar was poured into sterile 50 ml
plastic vials and a small piece of mycelium of each of the
selected isolates was subsequently transferred onto its sol-
idified surface. Sealed vials were kept at room temperature in
the dark and periodically checked for new hyphal growth.
After 28 d new mycelium covered the agar surface in all ino-
culated vials; at this point approx. 10 ml of twice-autoclaved
white peat (Br�at-Velkoobchod s.r.o., Czech Republic; pH 4.0)
were added into each vial, which were then treated as above.
After 29 d, one axenic 48-d-old V. myrtillus seedling was
aseptically inserted into peat, the vials were covered and
sealed with an air-permeable film and incubated in a growth
chamber (modified from Vohn�ık et al. 2012). Due to space
limitations it was not possible to use all REA isolates; there-
fore, we tested one representative of each REA clade (two in
the case of the new Meliniomyces sp. clade, see Suppl. Fig 1)
randomly selected from the isolates used in monoxenic cul-
tures with P. abies and V. myrtillus (Table 2). An isolate of
R. ericae, obtained in an unpublished study from surface-
sterilized ErM roots of Calluna vulgaris collected in the same
area, and an O. maius isolate JF304942 were used as positive
controls. Negative control vials were treated as above but
without inoculation with fungal mycelium (we assumed that
possible nutrient input to the experimental systems through
the inoculation was minimal). There were five vials with
healthy V. myrtillus seedlings for each treatment, including
the positive and negative control treatments.
Table 2 e The effect of selected isolates on the growth of Vacci
Isolatea Fresh root weight (mg)H7,37 ¼ 15.83; p ¼ 0.027
Cadophora finlandica EF093179 [n ¼ 4] 6.43 � 1.26 ab
Meliniomyces bicolor EF093180 [3] 6.70 � 1.46 ab
Meliniomyces variabilis EF093169 [5] 9.06 ± 1.13 b
Meliniomyces sp. EF093174 [5] 7.44 � 1.13 ab
Meliniomyces sp. EF093175 [5] 6.50 � 1.13 ab
Oidiodendron maius JF304942 [5] 3.62 � 1.13 ab
Rhizoscyphus ericaeb [5] 7.44 � 1.13 ab
Control (not inoculated) [5] 2.72 ± 1.13 a
The values are means � SE, different letters indicate significantly differen
comparisons of means ( p ¼ 0.05).
a Identity according to Table 1.
b This isolate was obtained from an ericoid mycorrhizal Calluna vulgaris
c Not included in the statistical analysis.
After 132 d, seedlings were carefully pulled from peat,
washed with tap water and their roots were separated from
shoots, dried with paper towels and immediately weighed.
Roots were then cleared in 10 % KOH, acidified in 3 % HCl,
stained with trypan blue at room temperature and de-stained
in lactoglycerol. ErM colonization was assessed by a single
person (C.M.B.), using the samemicroscope as above at 1 000�magnification, in 25 consecutive rhizodermal cells observed in
20 random fields of view per each root system (i.e., 500 cells
per each seedling), including negative control plants. Root
colonization was documented as above. Shoots were dried for
2 hr at 60 �C, and weighed. The collected data (fresh root
weight, dry shoot weight and ericoid mycorrhizal colo-
nization) were checked for homogeneity of variances and
normal distribution; fresh root weight and dry shoot weight
were not normally distributed while ericoid mycorrhizal col-
onization had neither homogeneity of variances nor normal
distribution, even after log and sqrt transformations. We,
therefore, used non-parametric KruskaleWallis ANOVA fol-
lowed by post-hoc multiple comparisons of means. STATIS-
TICA 10 (StatSoft, USA) was used for the statistical analyses. A
piece of peat was aseptically picked from several randomly
chosen vials and placed in Petri dishes with MMN to verify
viability of the inoculated strains at the end of the experiment.
Results
Isolation and identification of fungal cultures
The 360 surface-sterilized basidiomycetous ectomycorrhizas
yielded 128 relatively slow-growing non-sporulating isolates
which were sorted into 39 morphotypes differing in growth
and mycelial characteristics. The rest of the ectomycorrhizas
(ca. 64 %) either yielded fast-growing sporulating con-
taminants or produced no mycelium within the cultivation
period. By comparing their ITS nrDNA sequences with public
databases and with the help of phylogenetic analyses, we
were able to assign at least putative genus names to 38 mor-
photypes (36 ascomycetous, two basidiomycetous) belonging
to four distinct clusters (Table 1). The first cluster comprised
nium myrtillus in a peat-agar system
Dry shoot weight (mg)H7,37 ¼ 27.57; p ¼ 0.0003
Ericoid mycorrhizalcolonization (%) H5,28 ¼ 21.46;
p ¼ 0.0007
11.13 � 1.17 ab 0c
11.37 � 1.36 ab 30.27 � 7.79 ab
15.20 ± 1.05 b 69.20 ± 6.04 b
11.82 � 1.05 ab 22.76 � 6.04 ab
10.06 � 1.05 ab 8.80 ± 6.04 a
2.70 ± 1.05 a 26.08 � 5.91 ab
8.78 � 1.05 ab 69.16 ± 6.04 b
2.64 ± 1.05 a 0c
t groups according to KruskaleWallis ANOVA followed bymultiple
hair root and served as positive control.

288 M. Vohn�ık et al.
the R. ericae aggregate (Suppl. Fig 1 and Suppl. Table 1)
encompassing C. finlandica, M. bicolor, M. variabilis and two
unidentified ascomycetous isolates (EF093174 and
EF093175 ¼ a putatively new Meliniomyces species, referred to
as “Meliniomyces sp. Bohemian Forest”). The second cluster
comprised the P. fortinii s.l.eA. applanata species complex
encompassing A. applanata, Phialocephala subalpina, Phialoce-
phala turiciensis and two unidentified Phialocephala-related
isolates (EF093159 and EF093160¼ putative P. fortinii s.l. cryptic
species CSP 17, C. Gr€unig, pers. com.). The third cluster com-
prised various ascomycetes from the Eurotiales, Helotiales
and Hypocreales, including those with affinities to known
aquatic hyphomycetes which sometimes inhabit terrestrial
roots. The fourth cluster comprised the only two basidiomy-
cetes isolated, the ectomycorrhizal Hebeloma cf. bryogenes/
incarnatum and the saprotrophic Mycena galopus, suggesting
that our approach indeed favoured ascomycetes over
basidiomycetes.
All methods of phylogenetic analyses resulted in generally
the same trees with similar branch support (given that pos-
terior probabilities of �0.94 in Bayesian analyses and boot-
strap support of �70 % in all other analyses are significant). As
expected, phylogenetic relationships were largely independ-
ent of the isolates’ geographic origin or host species. The REA
isolates (Suppl. Fig 1) consisted of several more or less well-
defined (putative) Meliniomyces species in which C. finlandica
and R. ericae were embedded as monophyletic clades.
According to their placement in the tree and significant stat-
istical support, the two Meliniomyces-related morphotypes
indeed represent a new species within REA; no other
sequences represented in GenBank to date aremore similar to
these isolates than those included in the tree (maximal iden-
tity 93 %). In contrast, the rest of our REA morphotypes clus-
tered with well-defined species in this clade which were,
again, independent of host species or geographic origin. Our
analyses, thus, show that central European REA samples,
whichwere previously underrepresented in GenBank, fall into
the range of intraspecific variation for most REA species
(provided that they were correctly identified by their authors).
Symbiotic potential of selected isolates in Picea andVaccinium
In the monoxenic cultures with P. abies seedlings, none of the
five tested PAC isolates formed ectomycorrhizas with Hartig
net and hyphal mantle. However, some isolates (A. applanata
EF093158, P. turiciensis EF093156 and P. subalpina EF093161)
produced intracellular hyphae in the cortex cells of P. abies
roots. Additionally, A. applanata formed a circle of dense
microsclerotia around the endodermis, which was accom-
panied by dense intra- and intercellular colonization (Fig 1A).
Generally, roots of the plants inoculated with DSE isolates
lacked hair roots and resembled “pseudomycorrhizal” roots as
described by Melin (1922).
Several REA isolates (M. bicolor EF093180, M. variabilis
EF093169 and EF093170) produced intracellular colonization in
the P. abies cortex cells which was usually accompanied with
apparent darkening and thickening of their cell walls (cf.
Fig 1B and D). C. finlandica EF093155 and EF093179-inoculated
roots lacked root hairs and superficially resembled
ectomycorrhizas, but not the EcM morphotype P. bicolorata
(Fig 1C). Microscopic observation of these roots showed that
both strains formed an intraradical association resembling
ectendomycorrhiza, i.e., intercellular cortical Hartig net
accompanied by a few intracellular hyphae (Fig 1D). Spruce
roots with such ectendomycorrhizal association were
embedded in loose wefts of hyphae, but without dense hyphal
mantles. None of theM. bicolor norM. variabilis isolates formed
ectomycorrhiza or ectendomycorrhiza with spruce seedlings.
About one-half of the tested REA isolates, including the
putatively new Meliniomyces species, produced no inter- or
intracellular colonization. Similarly, neither Varicosporium
elodeae isolates (EF093184 and EF093185) colonized spruce
roots (Table 1).
Except for one M. variabilis isolate (EF093164) and both
V. elodeae isolates, all the remaining isolates colonized roots of
V. myrtillus intracellularly. Most of the REA isolates formed
dense intracellular hyphal coils resembling ericoid mycor-
rhiza, including C. finlandica EF093179 and M. bicolor EF093180
(Fig 1E, F and H). However, loose intracellular loops formed by
melanized and relatively thick hyphae of C. finlandica EF093155
resembled colonization typical for DSE-association rather
than ericoid mycorrhiza (Fig 1G). The putative new Melinio-
myces species infrequently formed loose to dense intracellular
hyphal loops and coils. Further in vitro experiments are nee-
ded to decide whether this colonization pattern represents
ericoid mycorrhiza. On the other hand, the typical DSE colo-
nization pattern was produced by all PAC isolates; most of
them formed vigorous intraradical colonization and often
produced intracellular microsclerotia which usually filled the
whole lumen of the colonized rhizodermal or cortical cells
(Fig 1I). None of the tested PAC strains formed intracellular
coils typical of ericoid mycorrhiza (Table 1).
Effect of selected isolates on V. myrtillus growth
All blueberry seedlings were green and healthy-looking at the
time of harvest except two inoculated withM. bicolor EF093180
and one inoculated with C. finlandica EF093179, which died
during the course of the experiment. Ericoid mycorrhizas
were formed in the roots inoculated with M. bicolor EF093180,
M. variabilis EF093169,Meliniomyces sp. EF093174 and EF093175,
O. maius JF304942 and R. ericae, though at different levels (see
below); non-inoculated control plants as well as those inocu-
lated with C. finlandica EF093179 were free of ericoid mycor-
rhizal colonization and were, therefore, excluded from the
statistical analysis of ErM colonization. All peat pieces trans-
ferred onto MMN producedmycelial colonies morphologically
identical to the respective inoculated isolates, including
C. finlandica EF093155; we did not detect any fast-growing
sporulating contaminants. This test, therefore, showed that at
the end of the experiment the screened vials contained viable
non-contaminated inoculum.
Inoculation had significant effects on all measured
parameters, i.e., fresh root weight (H7,37 ¼ 15.83, p¼ 0.027), dry
shoot weight (H7,37 ¼ 27.57, p ¼ 0.0003) and ErM colonization
(H5,28 ¼ 21.46, p ¼ 0.0007). Post-hoc multiple comparisons of
means indicated that plants forming ericoid mycorrhiza with
M. variabilis EF093169 produced significantly more fresh root
biomass than the non-inoculated plants, and their dry shoot

Meliniomyces variabilis in spruce ectomycorrhizas 289
weightwas significantly higher than in the case of theO.maius
JF304942-inoculated and non-inoculated plants. Additionally,
roots of the plants inoculated with M. variabilis EF093169 and
R. ericae had significantly higher ericoid mycorrhizal colo-
nization than those inoculated withMeliniomyces sp. EF093175
(Table 2).
Discussion
The EcM co-associated fungi only recently gained systematic
attention and their ecophysiological function remains largely
unclear. We still do not have answers for some basic ques-
tions, e.g., whether each single ectomycorrhiza possesses
endophytes. In our study from central Europe, approx. 36 % of
the screened ectomycorrhizal root tips yielded an endophyte.
This is comparable to 33.5 % obtained by Tedersoo et al. (2009),
who screened ectomycorrhizas from two Tasmanian wet
sclerophyll forests and a bit less than 48 % obtained by
Kernaghan & Patriquin (2011), who focused on ectomycor-
rhizas formed by the ascomycete Cenococcum geophilum at two
boreal sites in east Canada. It remains to be established where
is the actual living space of the co-associates in the respective
ectomycorrhizas, as they may dynamically inhabit hyphal
mantles and/or root intercellular and intracellular spaces;
intracellular colonization by apparently non-EcM mycelium
may be high in senescing ectomycorrhizas but is usually
absent in young turgescent EcM root tips (cf. Fig 2A and B). It is
plausible that intraradical competition for space and/or
resources ultimately leads to antagonism between EcM fungi
and their co-associates with yet unknown consequences to
Fig 2 e Typical anatomy of basidiomycetous ectomycorrhizas c
ectomycorrhiza with frequent intracellular hyphal colonization
(arrowheads) and a hyphal mantle (hm). Bar [ 50 mm. (B) Healt
(arrowheads) and a hyphal mantle (hm), with no intracellular c
permission of A. Lep�sov�a.
the host plant. Furthermore, ectomycorrhiza ageing ulti-
mately leads to termination of the active carbon flow from the
host plant to the EcM fungus which might give a competitive
advantage to its co-associate. However, these hypotheses
need to be further investigated.
In this study the frequency of mycobionts from the R. ericae
aggregate exceeded that of DSE and four different REA species
were obtained: C. finlandica, M. bicolor, the putatively new
Meliniomyces sp. and the dominant M. variabilis. Congruently,
Kernaghan & Patriquin (2011) detected M. bicolor and M. vari-
abilis in C. geophilum ectomycorrhizas in eastern Canada, and
Grelet et al. (2010) detected M. variabilis as the most frequent
co-associate of P. bicolorata-like ectomycorrhizas in two
P. sylvestris forests in northeeast Scotland. However, none of
our isolates belonged to the prominent ErM fungus R. ericae,
similarly to Kernaghan & Patriquin (2011) and Grelet et al.
(2010). This lack cannot be simply explained by absence in
the studied ecosystem because R. ericae, as well asM. variabilis,
were regularly isolated from hair roots of Ericaceae growing in
the investigated area in the Bohemian Forest NP (M. Vohn�ık,
unpublished). On the other hand, M. variabilis (similarly to
R. ericae and in contrast to Meliniomyces vraolstadiae) did not
belong among the major soil fungal OTUs found in the same
area by Baldrian et al. (2012). These observations suggest that
M. variabilis is an obligate biotroph rather than a free-living
soil dweller. Additionally, this species seems to have the
broadest ecological niche among the R. ericae aggregate
members e it has been detected as an endophyte in the roots
of a taxonomically diverse spectrum of plants including
Fagaceae, Orchidaceae and Salicaceae (Hambleton & Sigler
2005), Betulaceae and Pinaceae (Kernaghan & Patriquin
ollected in the vicinity of the sampling sites. (A) Senescing
in the cortex cells (arrows), a senescing Hartig net
hy turgescent ectomycorrhiza with a developed Hartig net
olonization in the cortex. Bar [ 50 mm. Photos used with

290 M. Vohn�ık et al.
2011), and Poaceae and Primulaceae (Tejesvi et al. 2013), but
seems to prefer Ericaceae, the only group which forms true
mycorrhizas with M. variabilis (Othaka & Narisawa 2008).
Thus, given its ability to intracellularly colonize conifer root
tips (Piercey et al. 2002; Vohn�ık et al. 2007b; the present study),
these most likely represent suitable refuges for M. variabilis in
microhabitats lacking ericaceous hosts. The outcome of col-
onization by M. variabilis for the EcM plant remains unclear
but the colonized roots often show signs of defence against
penetrating hyphae (Othaka & Narisawa 2008; the present
study).
C. finlandica and M. bicolor have been detected as EcM co-
associated fungi but they also form characteristic ecto- and
ectendomycorrhizas (Wang&Wilcox 1985; Vr�alstad et al. 2000,
2002; Villarreal-Ruiz et al. 2004). Additionally, both species
have been shown to form structures resembling ericoid
mycorrhizas in vitro (Vr�alstad et al. 2002; Villarreal-Ruiz et al.
2004; the present study), but they are only rarely detected in
Ericaceae roots under natural conditions (cf. Suppl. Fig 1).
These findings indicate an intriguing difference in life strat-
egies of the closely related REA mycobionts where some spe-
cies seem to prefer roots of Ericaceae (ericoid mycorrhizal
R. ericae), some roots of conifers (endophytic/ectomycorrhizal
C. finlandica and M. bicolor), some seem to be endophytic gen-
eralists as well as mycorrhizal specialists (M. variabilis, pos-
sibly also the newly discovered Meliniomyces sp.) while others
possibly form ectomycorrhizas with conifers and at the same
time represent a significant part of the soil fungal mycelium
(M. vraolstadiae, cf. Baldrian et al. 2012).
The in vitro potential of some REA members to form ecto-
mycorrhizas as well as ericoid mycorrhizas might be of great
ecological significance because Ericaceae often form a domi-
nant part of understory of EcM trees. In this study, both DSE
and REA isolates colonized spruce and blueberry seedlings;
however, DSE failed to form ectomycorrhizas or ericoid
mycorrhizas and M. bicolor, M. variabilis and Meliniomyces sp.
formed only ericoid mycorrhizas. Congruently, Vr�alstad et al.
(2002) reported a delimitation of EcM- and ErM-forming iso-
lates within REA. Intriguingly, in the present study one
C. finlandica strain formed ectendomycorrhizawith spruce and
associated with blueberry roots in a manner morphologically
resembling ericoidmycorrhiza in the agar-based system. Prior
to our report, there was only one re-synthesis study showing a
REA isolate with dual mycorrhizal ability (Villarreal-Ruiz et al.
2004); this isolate (M. bicolor LVR4069) formed characteristic P.
bicolorata ectomycorrhizas with pine, and the authors noted
that while interacting with blueberry, its “hyphae entered the
cells of the hair roots and formed coils characteristic of ericoid
mycorrhizas”. However, Tedersoo et al. (2009) noted that soil
saprobes belonging to Helotiales frequently display root
endophytic lifestyle and suggested that Ericaceae evolved
capacities to host them and to stimulate the formation of
intracellular hyphal coils for improved nutrient exchange.
This most probably led to the evolution of ErM symbiosis, yet
Ericaceae apparently kept the ability to host a wide range of
helotialean endophytes (Walker et al. 2011). Although C. fin-
landica EF093179 and M. bicolor LVR4069 formed structures
morphologically corresponding to ericoid mycorrhizas in vitro
on agar media, bi-directional nutrient transfer similar to that
observed by Grelet et al. (2009) needs to be demonstrated to
prove their ErM capabilities. Additionally, the incidence of C.
finlandica and M. bicolor in ericaceous roots under natural
conditions should be investigated to decide on ecophysio-
logical significance of their realized ericoid mycorrhizal
potential.
In the present study, Phialocephala-related DSE fungi repre-
sented the second most frequent group of EcM co-associated
fungi. These mycobionts generally inhabit living, decaying or
dead, mycorrhizal or non-mycorrhizal roots as well as stumps,
living stems, needles and fine or coarse woody debris (Menkis
et al. 2004, 2006; Gr€unig et al. 2009) although they seem to pre-
fer the cortex of primary roots and the periderm of lignified
roots of conifers (Gr€unig et al. 2008a). In the early period of
ectomycorrhiza research, DSE were often isolated from
healthy-looking roots or ectomycorrhizas (Mycelium radicis
atrovirens) and considered as true EcM fungi. However, inocu-
lation experiments showed that despite intraradical colo-
nization, they were not capable of forming EcM symbiosis and
often suppressed growth of the inoculated plants; hence their
early collective name “pseudomycorrhizal” fungi. Congruently,
none of the DSE isolates tested in the present study formed
ectomycorrhizaswith spruce seedlings. Instead, they colonized
spruce root cortex intracellularly, often formed typical micro-
sclerotia and the colonized roots morphology corresponded to
pseudomycorrhiza (Wang & Wilcox 1985). These results are in
agreement with a large body of literature showing neutral/
negative effects of P. fortinii-related DSE when inoculated in
non-mycorrhizal (but otherwise mycotrophic) coniferous
plants (e.g., Tellenbach et al. 2011). On the other hand, com-
parably less is known about physiological consequences of
their interactions with hosts possessing mycorrhizas. Based on
the few published experimental reports available, it appears
that they are neutral or rather negative (e.g., Hashimoto &
Hyakumachi 2001; Vohn�ık et al. 2005; Jones et al. 2009). While
in conifer plants from temperate regions most of the so far
investigated DSE seem to form harmful pseudomycorrhiza, the
effects of DSE may shift to positive in other plant guilds and/or
other climatic regions (e.g., Newsham 1999; Newsham et al.
2009). Additionally, the apparent ectomycorrhizal ability of
some DSE (Hashimoto & Hyakumachi 2001; Kaldorf et al. 2004)
deserves further attention. However, to our knowledge, A.
macrosclerotiorum is so far the only DSE observed to form ecto-
mycorrhizas in nature and at the same time confirmed to be an
EcM fungus in vitro (M€unzenberger et al. 2009).
In addition to the Phialocephala- andR. ericae-related isolates,
we obtained several other Helotiales with affinities to known
aquatic hyphomycetes (Articulospora tetracladia, V. elodeae, Neo-
bulgaria sp.) reported as common endophytes in submerged
aquatic plants (e.g., Kohout et al. 2012). Here, we obtained
aquatic species as endophytes in ectomycorrhizas but these
subsequently failed to colonize spruce and blueberry roots.
Further experiments, possibly in conditionsmore reflecting the
aquatic environment, are therefore needed to experimentally
confirm their endophytic lifestyle.
To conclude, our study showed that at least 36 % of appa-
rently healthy basidiomycetous ectomycorrhizas harboured
mostly ascomycetous endophytes. In contrast to our hypoth-
esis, the endophytic community was dominated by ericoid
mycorrhizal M. variabilis. The ericoid mycorrhizal potential of
this mycobiont was demonstrated in two independent trials

Meliniomyces variabilis in spruce ectomycorrhizas 291
where it formed characteristic intracellular structures in hair
roots and enhanced growth of an ericaceous host. The total
lack of the typical ErM fungus R. ericae in the screened ecto-
mycorrhizas suggests that these may represent selective
niches for certain ErM fungi andmay serve asmicro-reservoirs
of ErM inoculum in habitats lacking suitable ericaceous hosts.
It remains to be demonstrated where is the actual living space
ofM. variabilis inside the colonized ectomycorrhizas, andwhat
is the outcome of such colonization for the respective ecto-
mycorrhizas and consequently for the host plant.
Acknowledgements
This study was financially supported by the Czech Science
Foundation (GACR 206/09/P340) and the Czech Ministry of
Education, Youth and Sports (OC10058) and is a part of the
long-term research projects of the Institute of Botany ASCR
(RVO 67985939) and Charles University in Prague (SVV265203/
2012). Authors thank C. Gr€unig for identification of the Phia-
locephala-related isolates, O. Koukol for help with collection of
the field samples and A. Lep�sov�a for help with morphotyping
and permission to use her photos. Valuable comments of
anonymous reviewers and Kevin K. Newsham (editor) are
greatly appreciated. M.C.B. is grateful for a travel stipendium
from the Czech Ministry of Education, Youth and Sports
(AMB12AR014).
Supplementary material
Supplementary data related to this article can be found online
at http://dx.doi.org/10.1016/j.funeco.2013.03.006.
r e f e r e n c e s
Addy HD, Piercey MM, Currah RS, 2005. Microfungal endophytesin roots. Canadian Journal of Botany 83: 1e13.
Agerer R (ed), 1987e2002. Colour Atlas of Ectomycorrhizae. 1ste12thDelivery. Einhorn-Verlag, Schw€abisch Gm€und, Germany, ISBN3-921703-77-8.
Arnold AE, Mejia LC, Kyllo D, Rojas EI, Maynard Z, Robbins N,Herre EA, 2003. Fungal endophytes limit pathogen damage in atropical tree. Proceedings of the National Academy of Sciences ofthe United States of America 100: 15649e15654.
Baldrian P, Kola�r�ık M, �Stursov�a M, Kopeck�y J, Val�a�skov�a V,V�etrovsk�y T, �Zif�c�akov�a L, �Snajdr J, Ridl J, Vl�cek C, Vo�r�ı�skov�a J,2012. Active and total microbial communities in forest soil arelargely different and highly stratified during decomposition.ISME Journal 6: 248e258.
Bergero R, Girlanda M, Bello F, Luppi AM, Perotto S, 2003. Soilpersistence and biodiversity of ericoid mycorrhizal fungi inthe absence of the host plant in a Mediterranean ecosystem.Mycorrhiza 13: 69e75.
Brundrett MC, 2002. Coevolution of roots and mycorrhizas of landplants. New Phytologist 154: 275e304.
Carroll G, 1988. Fungal endophytes in stems and leaves: fromlatent pathogen to mutualistic symbiont. Ecology 69: 2e9.
Grelet GA, Johnson D, Paterson E, Anderson IC, Alexander IJ, 2009.Reciprocal carbon and nitrogen transfer between an
ericaceous dwarf shrub and fungi isolated from Piceirhizabicolorata ectomycorrhizas. New Phytologist 182: 359e366.
Grelet GA, Johnson D, Vr�alstad T, Alexander IJ, Anderson IC, 2010.New insights into the mycorrhizal Rhizoscyphus ericaeaggregate: spatial structure and co-colonization ofectomycorrhizal and ericoid roots. New Phytologist 188:210e222.
Gr€unig CR, Queloz V, Sieber TN, Holdenrieder O, 2008a. Darkseptate endophytes (DSE) of the Phialocephala fortinii s.l.eAcephala applanata species complex in tree roots:classification, population biology, and ecology. Botany 86:1355e1369.
Gr€unig CR, Duo A, Sieber TN, Holdenrieder O, 2008b. Assignmentof species rank to six reproductively isolated cryptic species ofthe Phialocephala fortinii s. l.eAcephala applanata speciescomplex. Mycologia 100: 47e67.
Gr€unig CR, Queloz V, Duo A, Sieber TN, 2009. Phylogeny ofPhaeomollisia piceae gen. sp. nov.: a dark, septate, conifer-needle endophyte and its relationships to Phialocephala andAcephala. Mycological Research 113: 207e221.
Hall TA, 1999. BioEdit: a user-friendly biological sequencealignment editor and analysis program forWindows 95/98/NT.Nucleic Acids Symposium Series 41: 95e98.
Hambleton S, Sigler L, 2005. Meliniomyces, a new anamorph genusfor root-associated fungi with phylogenetic affinities toRhizoscyphus ericae (¼Hymenoscyphus ericae), Leotiomycetes.Studies in Mycology 53: 1e27.
Hashimoto Y, Hyakumachi M, 2001. Effects of isolates ofectomycorrhizal fungi and endophytic Mycelium radicisatrovirens that were dominant in soil from disturbed sites ongrowth of Betula platyphylla var. japonica seedlings. EcologicalResearch 16: 117e125.
Jones MD, Grenon F, Peat H, Fitzgerald M, Holt L, Philip LJ,Bradley R, 2009. Differences in N-15 uptake amongst spruceseedlings colonized by three pioneer ectomycorrhizal fungi inthe field. Fungal Ecology 2: 110e120.
Kaldorf M, Renker C, Fladung M, Buscot F, 2004. Characterisationand spatial distribution of ectomycorrhizas colonizing aspenclones released in an experimental field. Mycorrhiza 14:295e306.
Kernaghan G, Sigler L, Khasa D, 2003. Mycorrhizal and rootendophytic fungi of containerized Picea glauca seedlingsassessed by rDNA sequence analysis. Microbial Ecology 45:128e136.
Kernaghan G, Patriquin G, 2011. Host associations between fungalroot endophytes and boreal trees. Microbial Ecology 62:460e473.
Kohout P, S�ykorov�a Z, Bahram M, Hadincov�a V, Albrechtov�a J,Tedersoo L, Vohn�ık M, 2011. Ericaceous dwarf shrubs affectectomycorrhizal fungal community of the invasive Pinusstrobus and native Pinus sylvestris in a pot experiment.Mycorrhiza 21: 403e412.
Kohout P, S�ykorov�a Z, �Ctvrtl�ıkov�a M, Rydlov�a J, Suda J, Vohn�ık M,Sudov�a R, 2012. Surprising spectra of root associated fungi insubmerged aquatic plants. FEMS Microbiology Ecology 80:216e235.
LoBuglio KF, Berbee ML, Taylor JW, 1996. Phylogenetic origins ofthe asexual mycorrhizal symbiont Cenococcum geophilum Fr.and other mycorrhizal fungi among the ascomycetes.Molecular Phylogenetics and Evolution 6: 287e294.
Melin E, 1922. On the mycorrhizas of Pinus sylvestris L. and Piceaabies Karst. Journal of Ecology 9: 254e257.
Menkis A, Allmer J, Vasiliauskas R, Lygis V, Stenlid J, Finlay R,2004. Ecology and molecular characterization of dark septatefungi from roots, living stems, coarse and fine woody debris.Mycological Research 108: 965e973.
Menkis A, Vasiliauskas R, Taylor AFS, Stenstrom E, Stenlid J,Finlay R, 2006. Fungi in decayed roots of conifer seedlings in

292 M. Vohn�ık et al.
forest nurseries, afforested clear-cuts and abandonedfarmland. Plant Pathology 55: 117e129.
M€unzenberger B, Bubner B, W€ollecke J, Sieber TN, Bauer R,Fladung M, H€uttl RF, 2009. The ectomycorrhizal morphotypePinirhiza sclerotia is formed by Acephala macrosclerotiorum sp.nov., a close relative of Phialocephala fortinii. Mycorrhiza 19:481e492.
Newsham KK, 1999. Phialophora graminicola, a dark septate fungus,is a beneficial associate of the grass Vulpia ciliata ssp. ambigua.New Phytologist 144: 517e524.
Newsham KK, Upson R, Read DJ, 2009. Mycorrhizas and darkseptate root endophytes in polar regions. Fungal Ecology 2:10e20.
Newsham KK, 2011. A meta-analysis of plant responses to darkseptate root endophytes. New Phytologist 190: 783e793.
Othaka N, Narisawa K, 2008. Molecular characterization andendophytic nature of the root-associated fungus Meliniomycesvariabilis (LtVB3). Journal of General Plant Pathology 74: 24e31.
Piercey MM, Thormann MN, Currah RS, 2002. Saprobiccharacteristics of three fungal taxa from ericalean roots andtheir association with the roots of Rhododendron groenlandicumand Picea mariana in culture. Mycorrhiza 12: 175e180.
Rice AV, Currah RS, 2006. Oidiodendron maius: saprobe in Sphagnumpeat, mutualist in ericaceous roots? In: Schulz BJE, Boyle CJC,Sieber TN (eds),Microbial Root Endophytes. Springer, Heidelberg,Germany.
Rodriguez RJ, White JF, Arnold AE, Redman RS, 2009. Fungalendophytes: diversity and functional roles. New Phytologist182: 314e330.
Ronquist F, Huelsenbeck JP, 2003. MRBAYES 3: Bayesianphylogenetic inference under mixed models. Bioinformatics 19:1572e1574.
Schild DE, Kennedy A, Stuart MR, 1988. Isolation of symbiont andassociated fungi from ectomycorrhizas of Sitka spruce.European Journal of Forest Pathology 18: 51e61.
Sieber TN, 2002. Fungal root endophytes. In: Waisel EA, Kafkafi U(eds), Plant Roots, the Hidden Half, 3rd edn. Marcel Dekker Inc.,Basel, Switzerland.
Sieber TN, 2007. Endophytic fungi in forest trees: are theymutualists? Fungal Biology Reviews 21: 75e89.
Smith SE, Read DJ, 2008. Mycorrhizal Symbiosis. Academic Press,London, UK, 804 pp.
�Storchov�a H, Hrdli�ckov�a R, Chrtek J, Tetera M, Fitze D, Fehrer J,2000. An improved method of DNA isolation from plantscollected in the field and conserved in saturated NaCl/CTABsolution. Taxon 49: 79e84.
Summerbell RC, 2005. From Lamarckian fertilizers to fungalcastles: recapturing the pre-1985 literature on endophytic andsaprotrophic fungi associated with ectomycorrhizal rootsystems. Studies in Mycology 53: 191e256.
Swofford DL, 2002. PAUP*. Phylogenetic Analysis Using Parsimony(*and Other Methods). Version 4. Sinauer, Sunderland, MA, USA.
Tedersoo L, Hansen K, Perry BA, Kjøller R, 2006. Molecular andmorphological diversity of pezizalean ectomycorrhiza. NewPhytologist 170: 581e596.
Tedersoo L, P€artel K, Jairus T, Gates G, P~oldmaa K, Tamm H, 2009.Ascomycetes associated with ectomycorrhizas: moleculardiversity and ecology with particular reference to theHelotiales. Environmental Microbiology 11: 3166e3178.
Tejesvi MV, Sauvola T, Pirttil€a AM, Ruotsalainen AL, 2013.Neighboring Deschampsia flexuosa and Trientalis europaeaharbor contrasting root fungal endophytic communities.Mycorrhiza 23: 1e10.
Tellenbach C, Gr€unig CR, Sieber TN, 2011. Negative effects onsurvival and performance of Norway spruce seedlingscolonized by dark septate root endophytes are primarilyisolate-dependent. Environmental Microbiology 13: 2508e2517.
Urban A, Puschenreiter M, Strauss J, Gorfer M, 2008. Diversity andstructure of ectomycorrhizal and co-associated fungalcommunities in a serpentine soil. Mycorrhiza 18: 339e354.
Usuki F, Narisawa K, 2005. Formation of structures resemblingericoid mycorrhizas by the root endophytic fungusHeteroconium chaetospira within roots of Rhododendron obsutumvar. kaempferi. Mycorrhiza 15: 61e64.
Villarreal-Ruiz L, Anderson IC, Alexander IJ, 2004. Interactionbetween an isolate from the Hymenoscyphus ericae aggregateand roots of Pinus and Vaccinium. New Phytologist 164:183e192.
Vohn�ık M, Albrechtov�a J, Vos�atka M, 2005. The inoculation withOidiodendron maius and Phialocephala fortinii alters phosphorusand nitrogen uptake, foliar C: N ratio and root biomassdistribution in Rhododendron cv. Azurro. Symbiosis 40: 87e96.
Vohn�ık M, Fendrych M, Albrechtov�a J, Vos�atka M, 2007a.Intracellular colonization of Rhododendron and Vaccinium rootsby Cenococcum geophilum, Geomyces pannorum and Meliniomycesvariabilis. Folia Microbiologica 52: 407e414.
Vohn�ık M, Fendrych M, Kola�r�ık M, Gryndler M, Hr�selov�a H,Albrechtov�a J, Vos�atka M, 2007b. The ascomycete Meliniomycesvariabilis isolated from a sporocarp of Hydnotryatulasnei(Pezizales) intracellularly colonizes roots of ecto- and ericoidmycorrhizal host plants. Czech Mycology 59: 215e226.
Vohn�ık M, Albrechtov�a J, 2011. The co-occurrence andmorphological continuum between ericoid mycorrhiza anddark septate endophytes in roots of six European Rhododendronspecies. Folia Geobotanica 46: 373e386.
Vohn�ık M, Sadowsky JJ, Kohout P, Lhot�akov�a Z, Nestby R,Kola�r�ık M, 2012. Novel root-fungus symbiosis in Ericaceae:sheathed ericoid mycorrhiza formed by a hitherto undescribedbasidiomycete with affinities to Trechisporales. PLoS ONE 7:e39524. http://dx.doi.org/10.1371/journal.pone.0039524.
Vr�alstad T, Fossheim T, Schumacher T, 2000. Piceirhiza bicolorata e
the ectomycorrhizal expression of the Hymenoscyphus ericaeaggregate? New Phytologist 145: 549e563.
Vr�alstad T, Schumacher T, Taylor AFS, 2002. Mycorrhizalsynthesis between fungal strains of the Hymenoscyphus ericaeaggregate and potential ectomycorrhizal and ericoid hosts.New Phytologist 153: 143e152.
Vr�alstad T, 2004. Are ericoid and ectomycorrhizal fungi part of acommon guild? New Phytologist 164: 7e10.
Walker JF, Aldrich-Wolfe L, Riffel A, Barbare H, Simpson NB,Trowbridge J, Jumpponen A, 2011. Diverse Helotialesassociated with the roots of three species of Arctic Ericaceaeprovide no evidence for host specificity. New Phytologist 191:515e527.
Wang CJK, Wilcox HE, 1985. New species of ectendomycorrhizaland pseudomycorrhizal fungi e Phialophora finlandia,Chloridium paucisporum and Phialocephala fortinii. Mycologia 77:951e958.
























