Embed Size (px)
Citation preview

276 BIOCHIMICA ET BIOPHYSICA ACTA
BBA 25787
T H E CATABOLISM OF L-ARGINtNE BY N E U R O S P O R A C R A S S A
M. CASTANEDA *, J. MARTUSCELLI *~ AND J. MORA *~"
Departamento de Bioquimica, I•stilulo Nacional de la Nutricidn. 2VHxico. D.F. Mexico)
(Received December 2oth, I966)
SUMMARY
I. The catabolism of exogenous and endogenously biosynthesized L-arginine has been studied in Neurospora erassa by growing the strains wild-type 74 A and the mutant auxotroph for L-proline, arg-g, in L-arginine or L-citrulline, with or withou~ NH~NO a.
2. A culture of N. crassa 74 A or arg-8 growing in the presence of L-citrulline (as nitrogen source, and nitrogen/L-proline source, respectively) develops an am- moniurn deficiency, as judged by a lower pool of almost all the amino acids, an in- crease in the ratio of dry weight to protein, and a low rate of growth.
3- The ammonium starvation occurs in the presence of a highly elevated pool of L-citrulline and L-arginine, and of a ful!y induced arginase, ornithine transaminase and urease.
4. NH4NOa or L-arginine when present in the medium reverse the ammonium deficiency. The citrulline and arginine pool, and the arginase, ornithine transaminase and urease activities, are then restored to basal levels and the catabolism of endo- genous arginine is accelerated.
5. The basis for the preferential utilization of exogenous arginme versus endo- genous arginine, that occurs in the absence of NH4NO a, is discussed in relation to channeling and cell organization.
INTRODUCTION
Because all of the enzymes that participate in the ornithine cycle are present in the mold Neurospora crassa, it has been assumed that the cycle actually operates in this microorganism 1. However, in contrast with ureote!ic organisms 2-~, N. crassa wild type does not require L-arginine for optimal growth 6, the L-arginme pool is easily detectable, ammonia is not a toxic compound, and a glutamine-dependent carbamyl phosphate synthetase 7 instead of an ammonia-dependent carbamyl phosphate syn- thetase and urease 1 are present in cell-free extracts. Moreover, recently we have found that the fungal arginase is a protein very similar to an argmase found in the
* Present address: Division of Biology, California Ins t i tu te of Technology, Pasadena, Catif.. U.S.A.
** Present address: Ins t i tu to de Estudios M6dicos y Bioldgicos, Universi ty of rVI6xico, M6xico, D .F . , Mexico.
*** Present address: Depa r tmen t oI Botany, Universi ty of Michigan, Ann Arbor, Mich.. U.S A.
Biochim. Biophys. Acta, 14I (1967) 276-286

CATABOLISM OF L-ARGININE BY N . ¢rassa 277
liver of uricotelic animals and different from that present in the liver of ureotelic animalsS, 9. These facts, together with the existence of a mutant strain of N. crassa unable to catabolize endogenous L-arginine 1°, suggest that the role of the ureotelic arginase is to hydrolyze endogenous arginine, supplying L-ornithine for further ammonia fixation in the ornithine cycle, while the function of the fungal arginase is the hydrolysis of exogenous L-arginine, providing L-omithine and urea as L-glutamic acid, L-proline and ammonia sources. The endogenously synthesized arginine in this fungus is thus the source mainly available for protein biosynthesis. To test this hypothesis, the catabolism of exogenous and endogenously biosynthesized L-arginine was studied in N. crassa. This paper reports the results.
MATERIAL AND METHODS
Strains N. crassa 74 A and arg-8 (44207) were obtained from H. J. VOGEL (Institute of
Microbiology of Rutgers, N.J.). Arg-I2 was obtained from V. W. WOODWARD (Rice University, Houston, Texas). am I was obtained from the Fungal Genetics Stock Center (Darmouth College, Hanover, N.H.).
Fungal growth The mold was grown in 5oo ml of Medium N (see ref. 6), contained in erlenmeyer
flasks shaken (12o strokes/rain) at 29 °. The inocumm for growth experiments was obtained by collecting a I6-h growth of an aliquot from a stock solution of mycelia prepared according to DO~IACHIE 11. When the inoculum was blended, this was carried out in a Waring blendor for 20 sec twice, at high speed. When CO 2 was collected, air was bubbled into the culture flasks in addition to shaking.
Cell-free extracts The mycelium was filtered, lyophilized and ground in the cold with glass powder
(30 mesh) in 2.5 mM potassium phosphate buffer (pH 7.4); after centrifugation, the supernatant was collected, the precipitate was extracted again and the supernatants were combined.
Enzyme assays Arginase (L-arginine amidinohydrolase, EC 3.5.3.1) was assayed by the for-
mation of ornithine; it was found that 50 mM of L-arginine in the assay mixture does not interfere seriously with the determination of ornithine; other conditions were as previously describedS, ". Ornithine transaminase (L-ornithine:2-oxo-glutarate aminotransferase, EC 2.6.1.13) was measured by the method of MIDDELHOVEN 1°',
urease (urea amidohydrolase, EC 3.5.1.5) according to SUMNER AND HAND 13, glycine transamidinase (L-arginine :glycine amidinotransferase, EC 2.1.4.1 ) according to WALKER 14 and also by using L-[guanido-14C~arginine as substrate and assaying for labelled guanidinoacetic acid. Arginine deiminase (L-arginine iminohydrolase, EC 3.5.3.6) was assayed according to OGINSKY AND GEHRIG 1~, L-amino-acid oxidase (L-amino-acid:oxygen oxidoreductase (deaminating), ECI.4.3.2) according to BURTON TM, with L-citrulline as substrate, and (after treating the cell-free extracts with Sephadex G-25) ornithine transcarbamylase (carbamoylphosphate:L-ornithine car-
L
Biochim. Biophys. Acta, i 4 i (1967) 276-286

278 ~. GASTANEDA, J~ I~ARTUSCELL[, J. _~fORA
bamoyltransferase, EC 2.L3.3) according to DA-WS~L Guanidobatyric acid ureo-., hydrolase was measured as previously described 8, except that because of the presence of urease the p~oduet 7-aminobutyric acid was assayed instead of urea. The former compound was isolated from the assay mixture by paper chromatography in 8o % phenol. Care was taken to assay all the enzymatic activities under optimal conditions at 37 °. Protein was determined by the method of LowRY et al. 18. The specific activities of the enzymes studied are expressed as /zmoies of product per h per mg of protein,
Amino acid analysis A sample for amino acid analysis of the protein of N. cfassa was obtained in
the following way: an aliquot of cell-free extract was precipitated with cold xo % trichloroacetic acid for I h; the precipitate was resuspended in 6 M HC1, heated at IOO ° under vacuum for 24 h, and, after evaporating the HCI, the sample was resus- pended in Lo M sodium acetate buffer (pH 5.o). Analysis of the free amino acids present in the cell-free extract was performed on a sample obtained after precipitating an aliquot with 1% picric acid, pouring the supernatant over a column (z5 mm × 4o mm) of resin Dowex AG2-X8 in the chloride form, and collecting the eluate after washing the column with i ml of o.o2 M HC1. The amino acid analyses were carried out on neutralized samples with a Spinco amino acid analyzer. The ornithine and arginine pools were determined by the methods of CPIINaRV 19 and VA~ P I L S ~ et al. 2°, respectively, after precipitating the proteins of the cell-free extracts with 5 % trichloroacetic acid. The pool of eitrullinewas assayed by the ARCm'BALD method 2~, after precipitating the protein with o.5 M HCIO,. The pools are expressed as ~moles of amino acid per mg of protein.
Radioactivity experiments When L-[ureido-14C~citrulline was added to the culture, labelled arginine was
isolated from the cell-free extract by treating an aliquot with 1% picric acid as mentioned above. The eluate was adjusted to pH 5 with NaOH and poured over a column (IO mm ~ 5o mm~ of Amber!ite CG-z2o previously equilibrated with o.I N sodium citrate (pH 5.o). The arginine was e!uted with 2o ml of o.2 N NaOH and an aliquot counted and/or assayed by the method of VA.N PILSUX et al. 2°. The incorpo- ration of radioactivity into protein was measured according to CAMPBELL, COOPER AI~I) HICKS z2. The evolved radioactive CO 2 was trapped during growth by connecting a 25-ml erlenmever flask containing 8 ml of 2.5 M NaOH to the culture flask. At the desired times the mold was filtered, 2.5 ml of 25 % trichloroacetic acid was added to the medium, and the CO 2 was trapped again during further incubation on a shaker for 3 h at 29 °. BaCO a was precipitated after adding 2.5 ml of 2.o M NH4CI and x.5 mi of o.75 M BaC12 to a i-ml atiquoz of the trapping solution. The precipitate was diluted and plancheted ~o infinite thickness and counted in a Nuclear-Chicago D-47 gas-flow detector witlh Micromil window.
Chemicals L-Arginine, L-ornithine and L-citrnlline for growth experiments were obtained
from Nutritional Biockemicals Co., and from California Corporation for Biochemical Research when used as substrates in enzymic assays. L-Eumido-14ClCitrutline, L- [guanido-14C~arginine and z-aspartic acid were also obtained from_ the latter company.
Biochim. Biophys° Ac~c~, x4i (1967) 276-286
CATABOLISM OF L-ARGININE BY N . ClzCtssa 279
o-Aminobenzaldehyde was obtained from Fluka, urea from J. T. Baker Chemical Co. and pyridoxal phosphate and carbamyl phosphate from Sigma Chemical Co.
RESULTS
Catabolism of exogenous L-arginine The catabolism of exogenous L-arginine was studied by growing N. crassa 74 A
and the mutant auxotroph for L-proline, arg-8, in a medium containing L-arginine in the presence and in the absence of NH4NO ~. L-Arginine is a precursor of L-proline through the intermediary formation of L-ornithine and glutamic acid-~/-semialde- hyde 1°. At different times, the two types of cultures were compared in regard to the arginine pool and to the activities of arginase, ornithine transaminase, urease, ornithine transcarbamylase and L-amino-acid oxidase. The cultures were also com- pared to those grown in different concentrations of NH4NO ~ as the sole nitrogen source in N. crassa 74A.
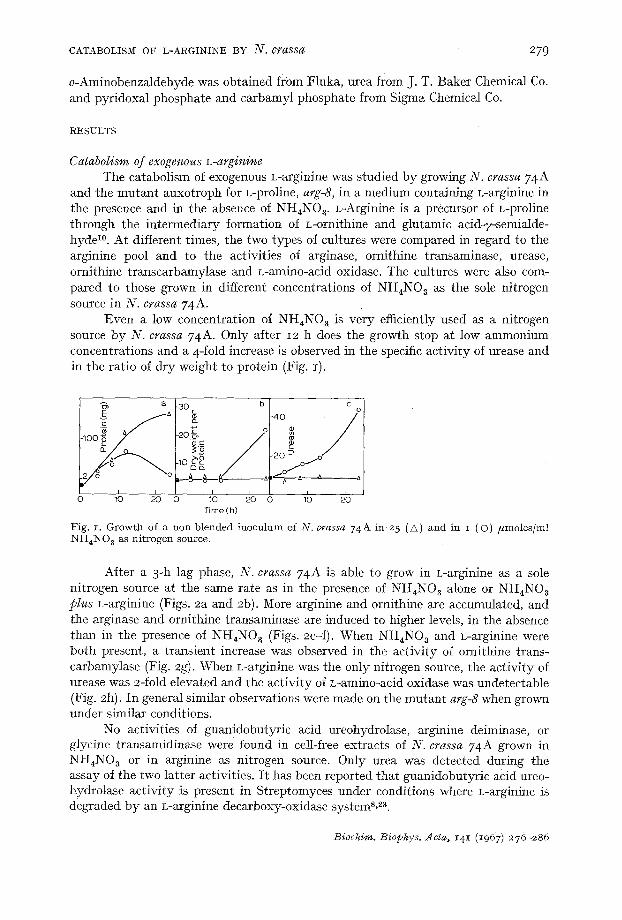
Even a low concentration of NH4NO a is very efficiently used as a nitrogen source by N. crassa 74A. Only after 12 h does the growth stop at low ammonium concentrations and a 4-fold increase is observed in the specific activity of urease and in the ratio of dry weight to protein (Fig. I).
{ lOOo/
-30 b
-20 f~ /
~ o -10 ',-- ~- 0o_
,_-4_~--~ /
c o
-4.0
I gX
T i m e ( h )
Fig. z. Growth of a non-blended inoculum of N. crassa 74 A in 25 (A) and in I (O) /zmoles/ml NI I4NO a as ni t rogen source.
After a 3-h lag phase, N. crassa 74A is able to grow in L-arginine as a sole nitrogen source at the same rate as in the presence of NH4NO ~ alone or NH4NO ~ plus L-arginine (Figs. 2a and 2b). More arginine and ornithine are accumulated, and the arginase and ornithine transaminase are induced to higher levels, in the absence than in the presence of NH4NO a (Figs. 2c-f). When NH~NO a and L-arginine were both present, a transient increase was observed in the activity of ornithine trans- carbamylase (Fig. 2g). When L-arginine was the only nitrogen source, the activity of urease was 2-fold elevated and the activity of L-amino-acid oxidase was undetectable (Fig. 2h). In general similar observations were made on the mutant arg-8 when grown under similar conditions.
No activities of guanidobutyric acid ureohydrolase, arginine deiminase, or glycine transamidinase were found in cell-free extracts of N. crassa 74A grown in NH4NO a or in arginine as nitrogen source. Only urea was detected during the assay of the two latter activities. It has been reported that guanidobutyric acid ureo- hydrolase activity is present in Streptomyces under conditions where L-arginine is degraded by an L-arginine deearboxy-oxidase systemSm.
Biochim. Biophys. Acta, x4i (1967) 276-286

2 8 0 No CASTAINEDA, J o X[ARTUSCELLI, J. )~IORA
Catabolism of endogenously biosynthesized L-arginine The rationale for studying the catabolism of endogenous L-arginine was to
supply a precursor of this amino acid (L-citruiline) as a nitrogen source in N. crosse
Protein a ~rvW~rIo.tei..~ightper 30 /a? °
/ -4,0
Arginine pool d Arginase /o
Ornithine U pease traBscarbamy/ase g -4Q -4
20~, , '~ \ [/_ o~
~= Opnithine o I b -'.~ pool o ~ / ~ c,!
l 0.e ~ - ~ - ~
] D i r e-00
002~- °/transami/OrnitNnease
• ! t ? / ~ ~x..x.z 10.01
h
o~ 1o 2'o o 4o 20 Time(h)
Fig. 2. Growth of a non-bIended i n o c u l u m of N . crassc~ 74 A in 2000 ffg/ml L-arginine wi th [A} and w i t h o u t ( 0 ) 25 f fmoles /ml NH~NO a.
~rotein ̂ J a -30 o protetn - ' a Dry weight per b 30 Citrulline pool c
/ j o o y O & &
~o/O / A. ~ - ~ a -° ' . d Arginase e O03F ~o Ornithine f
' J / ~transaminase o ° f ° - ° ~ / / ~ °'-o
rgnine poo[ i i ! a / ~ . ~ ~/~/// x ~ _ 6 0 ~ ° Qo2LA 0.01 xXa ~ o
#, &--~ &
IOrnithlne g ] Urease h I jtranscarbamylase I po p.o
20 o o D t k I 1 / f 0 20 " 40 0 20 40
Time(h)
Fig. 3. Growth of a non-b lended i n o c u l u m of N . cvassa 74 A in 2ooo f ig/m[ L-citrull ine wi th (A) and without (O) 25 ffmoles/mi NH4NO 3.
Biochim. Biophys . Acta, 141 (1967) 276-286

CATABOLISM OF L-ARGININE BY N . Gfctssa 281
74 A and as a nitrogen and/or L-proline source in the proline auxotroph, arg-8. Since the requirement for z-proline can be substituted by exogenous z-citrulline or L- arginine in the mutant arg-8 (ref. 24)--and in am D a strain that lacks glutamic de- hydrogenase 25, L-citrulline is not an L-glutamic acid source2G--it was assumed that the catabolism of endogenous z-arginine is a necessary condition for growth under the circumstances described.
In Fig. 3 are presented the results obtained when a non-blended inoculum of N . crassa 74Awas grown in a medium containing L-citrulline with or without NH4NO 3. After a 5-h lag phase, growth starts at a very low rate in the presence of L-citrnlline as a nitrogen source. Under these conditions the ratio of dry weight to protein, the citrulline and arginine pools, and the specific activities of arginase and ornithine transaminase and urease increase at levels higher than when the mold is growing in the presence of exogenous L-arginine as nitrogen source. Again all these changes are greatly minimized when the mycelium is growing in the presence of L-citrulline and NH4NO a. In Table I the concentrations of amino acids in cell-free extracts of N . crassa 74 A grown in L-citrulline (A) or in NH,NOa (B) as the nitrogen source are presented. The decrease in the concentrations of all the amino acids except citrul- line, arginine and ornithine is readily apparent in (A).
T A B L E I
AlUlNO ACID POOLS OF N . crassa 74 A
A non-b lended inocu lum of N. crassa 74 A was grown 12 11 in 2000 # g / m l of L-citruUine (A) or 25 #moles / rn l of NIK4NO a (B), as n i t rogen source.
A B (#moles/mg of protein)
Aspar t i c acid o.o o.271 Threonine o .o i I4 o.28I Serine o .o i 75 o.69I G lu ta ln ine o.o456 0. 315 Pro l ine o.o o.oi 73 G l u t a m i c ac id 0.0585 0.25 Ci t ru l l ine 2.95 o.oo Glycine o.oi43 o.o74 Alan ine 0.0427 0.59 Val ine 0.00856 o.089 Orn i th ine o . i8 o.o25 Meth ion ine o.oo o.o271 Isoleucine o.oo o.o136 Leuc ine o.oo 0.00865 Lys ine o.o214 0.0383 H i s t i d ine o.o14 o.o195 Arg in ine I. 33 o. 0259
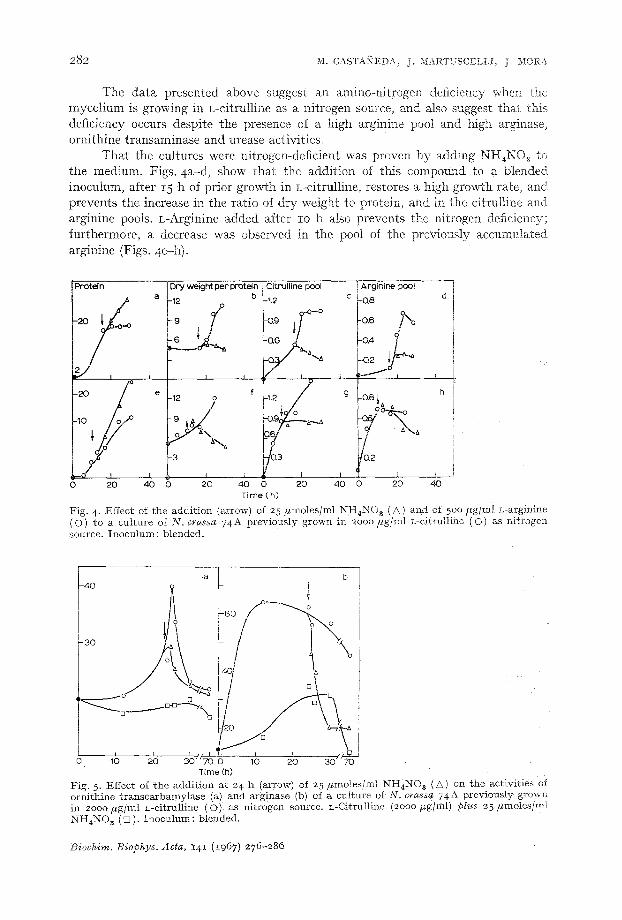
These changes also occur when blended mycelia of N . crassa 74A were used as inoculum; however, under these conditions L-citrulline is a better nitrogen source than when a non-blended inoculum was used (Figs. 4a-d). Noteworthy was the in- crease in the specific activity of ornithine transcarbamylase (Fig. 5a) in the presence of a high pool of L-arginine, and the induction of L-amino-acid oxidase, in the presence of L-citrulline without NH4NO~, when a blended inoculnm was used.
Bioehim. Biophys. Acta, 141 (1967) 276-286
282 M. CASTAX-EDA, ~, MARTUSCELLX, J. }~ORA
The data presented above suggest an amino-nitrogen deficiency when the mycelium is growing in L-citrulline as a nitrogen source, and also suggest that this deficiency occurs despite the presence of a high arginine pool and high arginase, ornithine transaminase and urease activities.
That the cultures were nitrogen-deficient was proven by adding NH~NOa tc the medium. Figs. 4a-d, show that the addition of this compound to a blended inoculum, after I5 h of prior growth in L-citmlline, restores a high growth rate, and prevents the increase in the ratio of dry weight to protein, and in the citrulline and arginine pools. L=&rginine added after i o h also prevents the nitrogen deficiency; furthermore, a decrease was observed in the pool of the previously accumulated arginine (Figs. 4e-h).
PrOtein /
I a
-10
0 2O
I
e
Dry weight per protein
-12 o b
e 12 o f 1,2
~ \~ ).6/
-3
40 0 20 40 0 20 40 0 20 40 Ti me (h)
Citr'ulline pool
-1.2 o..O o
-0.9 1] --
0.6 c~, z~&x &
Arginine gx~ol c -0.8
-0.6 7No o
-ON !t ~ -O2 -a
i I
g i-o.8, !
Fig. 4- Effect of the addi t ion (arrow) of 25 #moles /ml NH4NO ~ (A) and of 500 Fg/ml L-arginine (O) to a cul ture of N. crassa 74A previousIy grown in 2ooo #g/ml L-citrulline (O) as ni t rogen source. Inoculum: blended.
/ ° i
b
Time (h) Fig. 5. ]Effect of the addition at 24 h (arrow) of 25 #moies/ml NH~NO 3 (A) on the activities of orni tbine t ranscarbamylase (a) and arginase (b) of a cul ture of N. c~assc~ 74 A previouMy grown in 2ooo #g/ml L-citrulline (O) as ni t rogen source, z-Citrull ine (2ooo/~g/ml) plus 25 Fmoles/m! NH~NO 3 ( [] ). Inoculum : b lended,
Biochim. Biophys. Acla, :E4I (1"967) 276-286
CATABOLISM OF L-ARGININE BY N . c~'assa 283
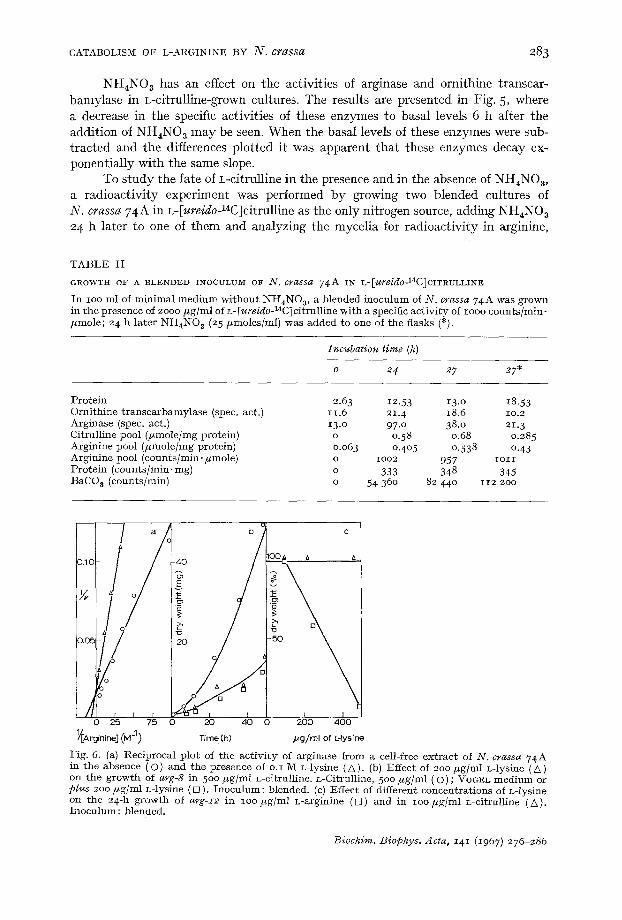
NH4NO ~ has an effect on the activities of arginase and ornithine transcar- bamylase in L-citrulline-grown cultures. The results are presented in Fig. 5, where a decrease in the specific activities of these enzymes to basal levels 6 h after the addition of NH4NOa may be seen. When the basal levels of these enzymes were sub- tracted and the differences plotted it was apparent that these enzymes decay ex- ponentially with the same slope.
To study the fate of L-citrulline in the presence and in the absence of NH4NO 3, a radioactivity experiment was performed by growing two blended cultures of N. crassa 74A in L-[ureido-~4C~citrulline as the only nitrogen source, adding NH4NO a 24 h later to one of them and analyzing the mycelia for radioactivity in arginine,
T A B L E II
G R O W T H OF A D L : E N D ~ D I N O C U L U M OF N. cfassa 74 A I N L-[uFeido-14C]CITRULLINE
In IOO ml of minimal med ium wi thou t NI taNO 3, a b lended inoculum of N. crassa 74 A was grown in t he presence of 2ooo ffg/ml of L-[ureido-14C]citrulline with a specific ac t iv i ty of iooo counts/rain- #mole ; 24 h la ter NH4NO 3 (25 ffmoles/ml) was added to one of the flasks (*).
Incubation time (h)
o 24 27 27*
Pro te in Orni thine t ranscarbaruylase (spec. act.) Arginase (spec. act.) Citrulline pool (ffmole/mg protein) Arginine pool (ffmole/mg protein) Arginine pool (counts/miu-ffmole) Pro te in (counts /min. mg) BaCO 3 (counts/rain)
2.63 12.53 13.o 18.53 11.6 21. 4 18.6 lO.2 13.o 97.0 38.0 21.3 o 0.58 0.68 0.285 0.063 0.405 0.538 0.43 o lOO2 957 i o l i o 333 348 345 o 54 360 82 44 ° 112 200
o
/? ;o
25
b
/ -20 o
75 0 20 40
b -50
0 200 400
~Arginine] (M -1) Time(h) /~g/ml of L4ysine
Fig. 6. (a) Reciprocal p lo t ' of the ac t iv i ty of arginase f rom a cell-free ex t rac t of N. crassa 74A in the absence (O) and the presence of o.I M L-lysine (A) . (b) ]Effect of 2oo pg/ml L-lysine (A) on the growth of arg-8 in 50o #g/ inl L-citrulline. L-Citrulline, 5oo ffg/ml (O) ; VOGEL medium or plus 2o0 ffg/ml L-lysine ([]) . Inoculum: blended. (c) Effect of different concentra t ions of L-lysine on the 24-h g rowth of arg-I2 in IOOffg/ml L-arginine ( ~ ) and in i oo f fg /ml L-citrulline (G). Inoculum : blended.
Biochim. Biophys. Acta, 141 (1967) 276-286

284 ivr. CASTANEDA, J. NARTUSCELLI, J NORA
protein and CO S (see METHODS). The results of such experiments are presented in Table II . The addition of NH4NO a leads to a 2-fold increase of evoived radioactive COs over the control. This is accomplished in the presence of a relatively low activity of arginase and low citrulline and arginine pools, when compared to values obtained in the absence of NH4NO a. The arginine pool was Ioo % ldoelled as was also the arginine of the proteins° This was calculated after determining the percentage of arginine in the protein of N. crassa 74A growing in L-citmlline with (6 %) and without (4.5 %) NH~NOa-
Enough evidence is available to suggest that L-citrulline is a proline source through the catabolism of endogenously biosynthesized arginine by arginase~,2<2L If this is so, the inhibition of the arginase i~ vivo should interfere with the growth of the mutant a~'g-8 in z-citrulline. For this purpose z-lysine, a welI-known inhibitor of arginase in other species ~s was used. In Figs. 6a and 6b, evidence is presented for the inhibition of N. crassa arginase by L-lysine i~ vitro, and of the growth of the mutan t arg-8 when growing in the presence of L-citrulline. The residual growth of the mutant arg-8 in minimal medium was not inhibited by L-lysine {Fig. 6b). L-Lysine does not inhibit the growth of the mutant arg-:r2 (a strain that lacks ornithine trans- carbamylase2"), in L-citrulline; it does, however, inhibit the growth of this strain in L-arginine (Fig. 6c).
When L-citrulline was used as a L-proline source in the presence and in the absence of NH~NO 3 in arg-8, the results resemble those found in N. cfassct 74 A when grown under such conditions.
DISCUSSION
In N. cmssa, NH4NO a plays an important role in the accumulation and in the catabolism of exogenous and endogenous L-arginine.
More arginine is accumulated from the medium by N. crassa 74 A in the absence than in the presence of NH4NO a. An effect of NH4NO a has also been observed_ m this laboratory, on the accumulation of z-arginine by the mutant am1, a strain unable to utilize ammonium sG.
Even though exogenous arginine is available for catabolism in the presence and in the absence of ammonium in arg-8, a mutant of this strain has been found which is able to catabolize exogenous arginine only in the absence of ammonium as Of great interest is the recent isolation of mutants of Saccharomyces cemv,siae sped- fically impaired in their permeabili ty for L-arginine; this is manifested only when ammonimn is present a.
In the absence of ammonia, exogenous arginine is catabolized by arginase in preference to endogenously biosynthesized arginine. The ammonia deficiency that occurs in the presence of L-citrulline as nitrogen source would be the result of an imbalance between the formation of L-arginine from z-citru!Iine, on the one hand. and the catabolism of this endogenous arginine on the other. Very possib!y, as the ammonia deficiency progresses, further limitations are imposed upon the catabolism of endogenous arginine. When NH~NO a or L-arginine is added, the deficiency dis- appears: first, because of the availability of ammonia, but also since in the presence of these compounds fess citrulline accumulates and less trapping of ammonia occurs
and at the same time, endogenously biosynthesized arginine becomes more available
Biochim. Biophys. Acla, i4i (.1967) 276-286

CATABOLISM OF L-ARGININE BY N . crassa 285
for catabolism. These assumptions are supported by the radioactivity experiments in which it was found that after the addition of NHhN0 3 there is a significant increase in the labelled (endogenous) arginine catabolized to C0 2, concomitant with a decrease in the citrulline and arginine pools, and the arginase activity.
The results presented here bear some similarity with those reported by GRoss 32, in relation to mutant strains of N . crassa that are not able to carry out some single reaction subsequent to the synthesis of 5-dehydroshikimic acid. These mutants ac- cumulate protocatechuic acid in their growth medium despite having induced a proto- catechuic acid oxidase. 0n ly when the glucose or the sucrose of the medium is ex- hausted is the accumulated protocatechuic acid quantitatively metabolized.
The physical basis for the preferential utilization of exogenous as against endogenous arginine that occurs in the absence of NHhN0 8, can only be a matter of speculation at the present time.
Recently, evidence has been presented that establishes metabolic channeling as an important process in cell organization. DAVIS 33 has reported in N . crassa
segregation of carbamyl phosphate for arginine biosynthesis from that for pyrimidine biosynthesis; two different specific enzymes are involved in the synthesis of this compound. SERCARZ AND GORINI ~4 have found a mutant in Escher i ch ia colt that segregates the arginine for protein synthesis from the arginine that is involved in repression. 1V~UNKRES AND WOODWARD 3s have presented evidence for the locational specificity of mitochondrial malate dehydrogenase in N . crassa, and KINSEY AND WAGNER 36 have found a particulate system in the same fungi that synthesizes different amino acids, among others, arginine.
ACKNOWLEDGEMENT
I t is a pleasure to acknowledge the criticisms and help with the manuscript of Dr. R. H. DAvIs.
R E F E R E N C E S
i A. SRB AND 1~. ]-I. I-IoRoWITZ, J. Biol. Chem., 154 (1944) 129. 2 A. BORMAN, T. R. WOOD, H. C. BLACK, E. G. AND1~RSON, J. M. OEST]~RLING, M. WOMACK
AND W. C. ROSE, dr. Biol. Chem., 166 (1946) 585 . 3 S. A. I~APLAN AND C. S. NAGAREDA, Am. d r. Physiol., 202 (1962) 695. 4 P ' P" COHEN AND C.TvV. BROWN, JR., in M. ELORKIN AND H. S. MASON, Comparative Biochemistry,
Vol. 2, Academic , New York, 196o, p. I6 i . 5 J . 1VioRA, J . MARTUSCELLI, J. ORTIZ PINEDA AND G. SOB~RCN, Biochem. J., 96 (1965) 28. 6 H. J. VOGEL, Microbiol. Genet. Bull., 13 (1956) 42. 7 R. H. DAvis, Genetics, 54 (1966) 33 °. 8 J. 1ViORA, R. TARRAB, J . MARTUSCELLI AND G. SOB~RbN, Biochem. J., 96 (1965) 588. 9 J- 1VIoRA, R. TARRAB AND L. F. BOJALIL, Biochim. Biophys. Acta, 118 (1966) 206.
io R. I-I. VOGXL AND M. J . I<OPAC, Biochim. Biophys. Acta, 36 (1959) 505 • i i W. D. DONACHIE, Biochim. Biophys. Acta, 82 (1964) 284. 12 W. J. MIDDELHOVEN, Biochim. Biophys. Acta, 93 (1964) 650. 13 J. ]3. SUMNER AND D. B. HAND, d r. Biol. Chem., 76 (1928) 149. 14 J . B. WALKER, J. Biol. Chem., 234 (1959) 1481. 15 E. L. OGI:NSKY AND R. ~'. GErlRIG, J. Biol. Chem., 198 (1952) 799. 16 }<. BURTON, Bioehem. ,f., 5 ° (1951) 258. 17 R. H. DAvis, Arch. Biochem. Biophys., 97 (1962) 185. 18 O. }-I. LOWRY, N. J. ROSEBROUGI-I, A. L. FARR AND R. J. RANDALL, J. Biol. Chem., 193 (1951)
265. 19 E. P. CHINARD, J. Biol. Chem., 199 (1952) 91.
Biochim. Biophys. Acta, 141 (1967) 276-286

286 M. CASTAI~EDA, J~ MARTUSCELLI, J. MORA
20 J. F. VAN PILSUM, R. P. MART~N, E. KITO AND H. HESS, d r. Biol. Chem., 222 (I956) 225, 2I R. M. ARCHIBALD, f. Biol. Chem., z56 (I946) 121. 22 P. N. CAMPBELL, C. COOPER AND M. HICKS, Biochem. f . , 92 (1964) 225. 23 N. V. THOAI, F. THOME-BEAu AND A. OLIJMUCI~L Bioehim. Biophyso Acta, H5 (z966) 73. 24 A. M. SRB, J. R. S. FINCHAM AND D. BONNER, Am. J. Botany, 37 (195 °) 533. 25 J. R. S. ~'INCHAM, d r. GeE. Microbiol., 2I (I959) 60o. 26 J. R. S. FINCHAM, dr. Biol. Chem., I82 (I95 o) 6I. 27 P. H. ABELSON AND H. j . VO~EL, J . Biol. Chore., 213 (1955) 355. 28 A. HUNTER AND C. E. DOWNS, J . Biol. Chem., I57 (I945) 427 • 29 V. W. WOODWARD AND P. SCHWARZ, Genetics, 49 (1964) 845. 30 L. CAI~EDO, J. MARTUSCELLI AND J . MORA, Natl. Cancer Inst..Zvfonograph, i n t h e press. 31 M. GRENSON, M. MOUSSET, J. H. WIAME AND J. BECHET, Biochim. Biophys. Acfa, 127 (I966)
325 . 32 S. R. GROSS, Trans. N . Y . Acad. Sci., 22 (i959) 44. 33 R. H. DAVIS, Science, I42 (I963) i652. 34 E. E. SERCARZ AND L. GORINI, jr. Mol. Biol., 8 (I964) 254. 35 I42. D. MUNKRES AND D. O. WOODWARD, Proc. Natl. Acad. Sci. U.S., 55 (z966) I217o 36 J. A. KINSEY AND R. P. WAGNER, Proc. Natl. Acad. Sci. U.S., 55 (1966) 4o4 .
Biochim. Biophys. Acta, I4 I (I967) 276-286





























