Embed Size (px)
Citation preview

DOI: 10.1126/science.1204592, 1429 (2011);332 Science
et al.Carmine SettembreTFEB Links Autophagy to Lysosomal Biogenesis
This copy is for your personal, non-commercial use only.
clicking here.colleagues, clients, or customers by , you can order high-quality copies for yourIf you wish to distribute this article to others
here.following the guidelines
can be obtained byPermission to republish or repurpose articles or portions of articles
): May 14, 2012 www.sciencemag.org (this information is current as of
The following resources related to this article are available online at
http://www.sciencemag.org/content/332/6036/1429.full.htmlversion of this article at:
including high-resolution figures, can be found in the onlineUpdated information and services,
http://www.sciencemag.org/content/suppl/2011/05/25/science.1204592.DC1.html can be found at: Supporting Online Material
http://www.sciencemag.org/content/332/6036/1429.full.html#relatedfound at:
can berelated to this article A list of selected additional articles on the Science Web sites
http://www.sciencemag.org/content/332/6036/1429.full.html#ref-list-1, 10 of which can be accessed free:cites 29 articlesThis article
http://www.sciencemag.org/content/332/6036/1429.full.html#related-urls8 articles hosted by HighWire Press; see:cited by This article has been
http://www.sciencemag.org/cgi/collection/cell_biolCell Biology
subject collections:This article appears in the following
registered trademark of AAAS. is aScience2011 by the American Association for the Advancement of Science; all rights reserved. The title
CopyrightAmerican Association for the Advancement of Science, 1200 New York Avenue NW, Washington, DC 20005. (print ISSN 0036-8075; online ISSN 1095-9203) is published weekly, except the last week in December, by theScience
on
May
14,
201
2w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

21. We use a density r = 2800 kg/m3. Using r = 3000 kg/m3,which is a common choice in other energy studies, wouldhave reduced our radiated energy by 6.5% to ES = 8.5 ×1017 J.
22. S. Ide, G. C. Beroza, Geophys. Res. Lett. 28, 3349(2001).
23. H. Kanamori, Phys. Earth Planet. Inter. 6, 346 (1972).24. H. Kanamori, M. Kikuchi, Nature 361, 714 (1993).25. A. Venkataraman, H. Kanamori, J. Geophys. Res. 109,
B05302 (2004).26. Y. Tanioka, K. Sataka, Geophys. Res. Lett. 23, 1549
(1996).27. W. Nakayama, M. Takeo, Bull. Seismol. Soc. Am. 87, 918
(1997).28. T. Sato, K. Imanishi, M. Kosuga, Geophys. Res. Lett. 23,
33 (1996).
29. N. Uchida et al., abstract S34B‐04 presented at the 2010American Geophysical Union Fall Meeting, San Francisco,CA, 13 to 17 December 2010.
30. A. Baltay, S. Ide, G. Prieto, G. Beroza, Geophys. Res. Lett.38, L06303 (2011).
31. P. Wessel, W. H. F. Smith, Eos Trans. AGU 72, 441(1991).
Acknowledgments: This work was supported by the Ministry ofEducation, Culture, Sports, Science and Technologyof Japan KAKENHI (grant 21107007) and the Japan Societyfor the Promotion of Science KAKENHI (grant 23244090).GMT software (31) was used to draw figures. A.B. wassupported at Stanford University by the Gabilan StanfordGraduate Fellowship. GSN data are available online(www.iris.edu/hq/programs/gsn/data) through a cooperativescientific facility operated jointly by the Incorporated
Research Institutions for Seismology, the USGS, and theNSF. KiK-net data are also available online (www.kik.bosai.go.jp/kik/) through National Research Institute for EarthScience and Disaster Prevention.
Supporting Online Materialwww.sciencemag.org/cgi/content/full/science.1207020/DC1Materials and MethodsSOM TextFigs. S1 to S8Table S1References (32–34)
4 April 2011; accepted 12 May 2011Published online 19 May 2011;10.1126/science.1207020
TFEB Links Autophagy toLysosomal BiogenesisCarmine Settembre,1,2,3 Chiara Di Malta,1 Vinicia Assunta Polito,1,2,3 Moises Garcia Arencibia,4
Francesco Vetrini,2 Serkan Erdin,2,3 Serpil Uckac Erdin,2,3 Tuong Huynh,2,3 Diego Medina,1
Pasqualina Colella,1 Marco Sardiello,2,3 David C. Rubinsztein,4 Andrea Ballabio1,2,3,5*
Autophagy is a cellular catabolic process that relies on the cooperation of autophagosomes andlysosomes. During starvation, the cell expands both compartments to enhance degradationprocesses. We found that starvation activates a transcriptional program that controls major steps ofthe autophagic pathway, including autophagosome formation, autophagosome-lysosome fusion,and substrate degradation. The transcription factor EB (TFEB), a master gene for lysosomalbiogenesis, coordinated this program by driving expression of autophagy and lysosomal genes.Nuclear localization and activity of TFEB were regulated by serine phosphorylation mediated by theextracellular signal–regulated kinase 2, whose activity was tuned by the levels of extracellularnutrients. Thus, a mitogen-activated protein kinase–dependent mechanism regulates autophagy bycontrolling the biogenesis and partnership of two distinct cellular organelles.
Macro-autophagy is an evolutionarilyconserved mechanism that targets in-tracytoplasmic material to lysosomes,
thus providing an energy supply during nutrientstarvation (1, 2). Autophagy activation duringstarvation is negatively regulated by mammaliantarget of rapamycin complex 1 (mTORC1), whoseactivity is dependent on cellular energy needs (3).The observation that starvation induced the tran-scription of several autophagy genes, whereas in-hibition of mTORC1 did not, suggests the presenceof alternative transcriptionalmTORC1-independentregulation of autophagy (4, 5) (fig. S1, A and B).
We tested whether TFEB, a transcription fac-tor (“EB”) that controls lysosomal biogenesis bypositively regulating genes belonging to the Co-
ordinated Lysosomal Expression and Regulation(CLEAR) network, also regulated autophagy(6, 7). Stable TFEB overexpression in HeLa cellssignificantly increased the number of autophago-somes detected by immunofluorescence and im-munoblotting of the LC3 protein, which specificallyassociates with autophagosomes (8) (Fig. 1, Aand B). Similar data were obtained by transientoverexpression of TFEB in HeLa and COS7(monkey kidney fibroblast) cells (fig. S2, A to C)and from electron microscopy on mouse embry-onic fibroblasts (MEFs) infected with a lentivirusoverexpressing TFEB (fig. S3, A to D). This in-crease persisted in cells treated with bafilomycinand pepstatin, as well as the cysteine proteinaseinhibitor E64, which are lysosomal inhibitors ofautophagosome and LC3-II degradation; the sus-tained increase indicated that TFEB activates theformation of autophagosomes (Fig. 1A and fig.S4, A and B).
RNA interference (RNAi) of TFEB in HeLacells resulted in decreased levels of LC3-II bothin normal and starved conditions, in either thepresence or absence of bafilomycin (Fig. 1, Cand D). The decrease of LC3-II correlated withthe levels of TFEB down-regulation achieved bythe different RNAi oligomers (fig. S4, C and D).These gain- and loss-of-function data suggest
that the biogeneses of autophagosomes and lyso-somes are co-regulated by TFEB. We next mea-sured the rate of delivery of autophagosomes tolysosomes using an RFP-GFP (red fluorescentprotein–green fluorescent protein) tandem taggedLC3 protein, which discriminates early autophagicorganelles [GFP-positive and monomeric RFP(mRFP)–positive] from acidified autophagolyso-somes (GFP-negative and mRFP-positive), be-cause of quenching of the GFP signal (but not ofmRFP) inside acidic compartments. The numberof autophagolysosomes was higher in TFEB over-expressing cells than in control cells, which indi-cated that TFEB enhanced the autophagic flux(Fig. 1E). Consistently, degradation of long-livedproteins (9) was enhanced by TFEB overexpres-sion and reduced by TFEB depletion (knockdown)(fig. S5A).
To test whether TFEB regulated the expres-sion of autophagy genes, we analyzed themRNAlevels of a group of 51 genes reported to be in-volved in several steps of the autophagic pathway(1, 10, 11). The enhancement of the expressionlevels of autophagy genes in cells overexpressingTFEBwas similar to that of starved cells (Fig. 2Aand table S1) [Pearson correlation coefficient (r) =0.42; P = 0.001]. Eleven of the analyzed geneswere significantly up-regulated after transient,stable, and tetracycline-dependent TFEB expres-sion, whereas they were down-regulated afterTFEB silencing in both normal and starved con-ditions (Fig. 2A, fig. S5B, and tables S2 and S3).The expression of UVRAG, WIPI, MAPLC3B,SQSTM1, VPS11, VPS18, and ATG9B was mostsignificantly affected by TFEB overexpression(tables S2 and S3). These genes are known toplay a role in different steps of autophagy (fig.S6) and appeared to be direct targets of TFEBbecause they carry at least one TFEB target site(6) in their promoters (fig. S7). In addition, wevalidated the binding of TFEB to the target se-quence by quantitative chromatin immunopre-cipitation assay (QChip) and observed that thisbinding is further enhanced during starvation(fig. S8, A and B).
In normal conditions, TFEB is localized tothe cytoplasm (6). Nutrient starvation rapidlyinduced TFEB nuclear translocation (Fig. 2, Band C), and cytosolic TFEB from starved cells
1Telethon Institute of Genetics and Medicine (TIGEM), Via PietroCastellino 111, 80131 Naples, Italy. 2Department of Molecularand Human Genetics, Baylor College of Medicine, Houston, TX77030, USA. 3Jan and Dan Duncan Neurological ResearchInstitute, Texas Children’s Hospital, Houston, TX 77030, USA.4Cambridge Institute for Medical Research, Wellcome Trust–Medical Research Council Building, Addenbrooke’s Hospital,Hills Road, Cambridge CB2 0XY, UK. 5Medical Genetics, De-partment of Pediatrics, Federico II University, Via Pansini 5,80131 Naples, Italy.
*To whom correspondence should be addressed. E-mail:[email protected]
www.sciencemag.org SCIENCE VOL 332 17 JUNE 2011 1429
REPORTS
on
May
14,
201
2w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
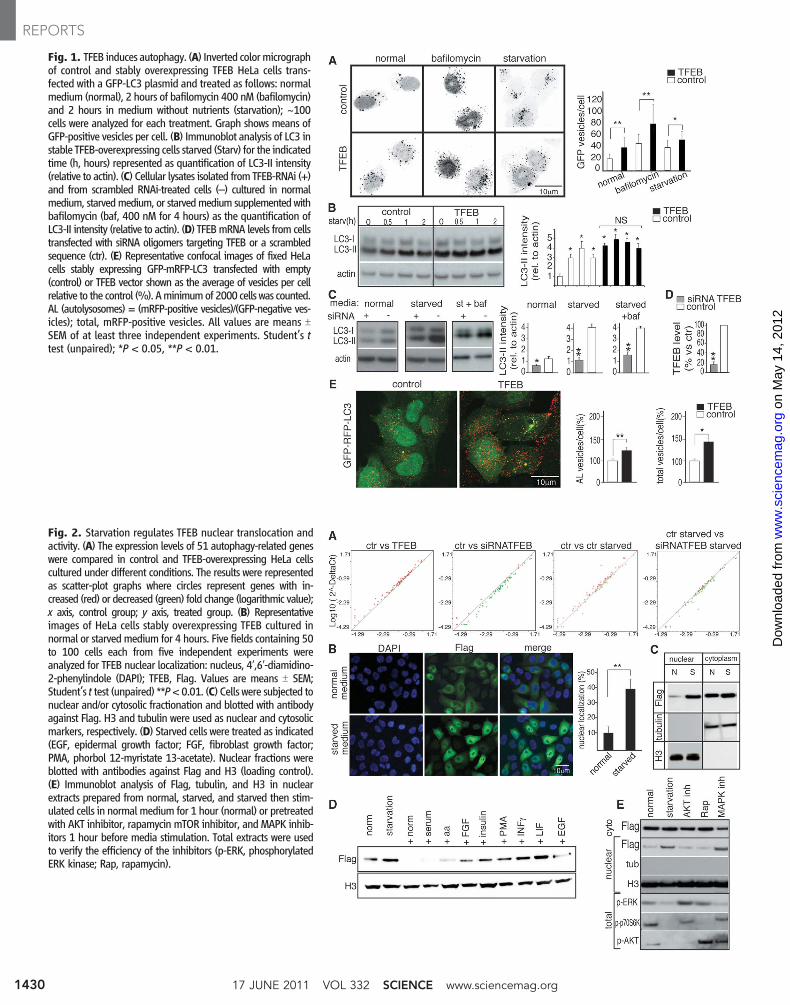
Fig. 1. TFEB induces autophagy. (A) Inverted colormicrographof control and stably overexpressing TFEB HeLa cells trans-fected with a GFP-LC3 plasmid and treated as follows: normalmedium (normal), 2 hours of bafilomycin 400 nM (bafilomycin)and 2 hours in medium without nutrients (starvation); ~100cells were analyzed for each treatment. Graph shows means ofGFP-positive vesicles per cell. (B) Immunoblot analysis of LC3 instable TFEB-overexpressing cells starved (Starv) for the indicatedtime (h, hours) represented as quantification of LC3-II intensity(relative to actin). (C) Cellular lysates isolated from TFEB-RNAi (+)and from scrambled RNAi-treated cells (–) cultured in normalmedium, starvedmedium, or starvedmedium supplemented withbafilomycin (baf, 400 nM for 4 hours) as the quantification ofLC3-II intensity (relative to actin). (D) TFEBmRNA levels from cellstransfected with siRNA oligomers targeting TFEB or a scrambledsequence (ctr). (E) Representative confocal images of fixed HeLacells stably expressing GFP-mRFP-LC3 transfected with empty(control) or TFEB vector shown as the average of vesicles per cellrelative to the control (%). Aminimumof 2000 cells was counted.AL (autolysosomes) = (mRFP-positive vesicles)/(GFP-negative ves-icles); total, mRFP-positive vesicles. All values are means TSEM of at least three independent experiments. Student’s ttest (unpaired); *P < 0.05, **P < 0.01.
Fig. 2. Starvation regulates TFEB nuclear translocation andactivity. (A) The expression levels of 51 autophagy-related geneswere compared in control and TFEB-overexpressing HeLa cellscultured under different conditions. The results were representedas scatter-plot graphs where circles represent genes with in-creased (red) or decreased (green) fold change (logarithmic value);x axis, control group; y axis, treated group. (B) Representativeimages of HeLa cells stably overexpressing TFEB cultured innormal or starvedmedium for 4 hours. Five fields containing 50to 100 cells each from five independent experiments wereanalyzed for TFEB nuclear localization: nucleus, 4′,6′-diamidino-2-phenylindole (DAPI); TFEB, Flag. Values are means T SEM;Student’s t test (unpaired) **P<0.01. (C) Cells were subjected tonuclear and/or cytosolic fractionation and blotted with antibodyagainst Flag. H3 and tubulin were used as nuclear and cytosolicmarkers, respectively. (D) Starved cells were treated as indicated(EGF, epidermal growth factor; FGF, fibroblast growth factor;PMA, phorbol 12-myristate 13-acetate). Nuclear fractions wereblotted with antibodies against Flag and H3 (loading control).(E) Immunoblot analysis of Flag, tubulin, and H3 in nuclearextracts prepared from normal, starved, and starved then stim-ulated cells in normal medium for 1 hour (normal) or pretreatedwith AKT inhibitor, rapamycin mTOR inhibitor, and MAPK inhib-itors 1 hour before media stimulation. Total extracts were usedto verify the efficiency of the inhibitors (p-ERK, phosphorylatedERK kinase; Rap, rapamycin).
17 JUNE 2011 VOL 332 SCIENCE www.sciencemag.org1430
REPORTS
on
May
14,
201
2w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
appeared to have a lower molecular weight thanthat of normally fed cells (fig. S9A). This molec-ular weight shift occurred rapidly, but transiently,and was abolished within 1 hour after we addedback normal media to starved cells, concomitantwith a decrease of nuclear TFEB (fig. S9A).After starvation medium was supplemented withserum, amino acids, or growth factors [i.e., insulinor epidermal growth factor (EGF)], TFEB nucleartranslocation was inhibited comparedwith resultsfrom starvation medium alone (Fig. 2D), where-as no effect was observed with cytokines [i.e.,interferon (INF) or leukemia inhibitory factor(LIF)] (Fig. 2D), which suggested that activationof TFEB is controlled by a signaling mechanismthat is sensitive to nutrient and growth factors.We stimulated starved cells with normal mediumsupplemented with drugs inhibiting the mTORC,phosphatidylinositol 3-kinase–AKT (PI3K-AKT),and mitogen-activated protein kinases (MAPKs).MAPK inhibition resulted in TFEB nuclear lo-calization, whereas AKT and mTOR inhibitionhad no effect (Fig. 2E and fig. S9B).
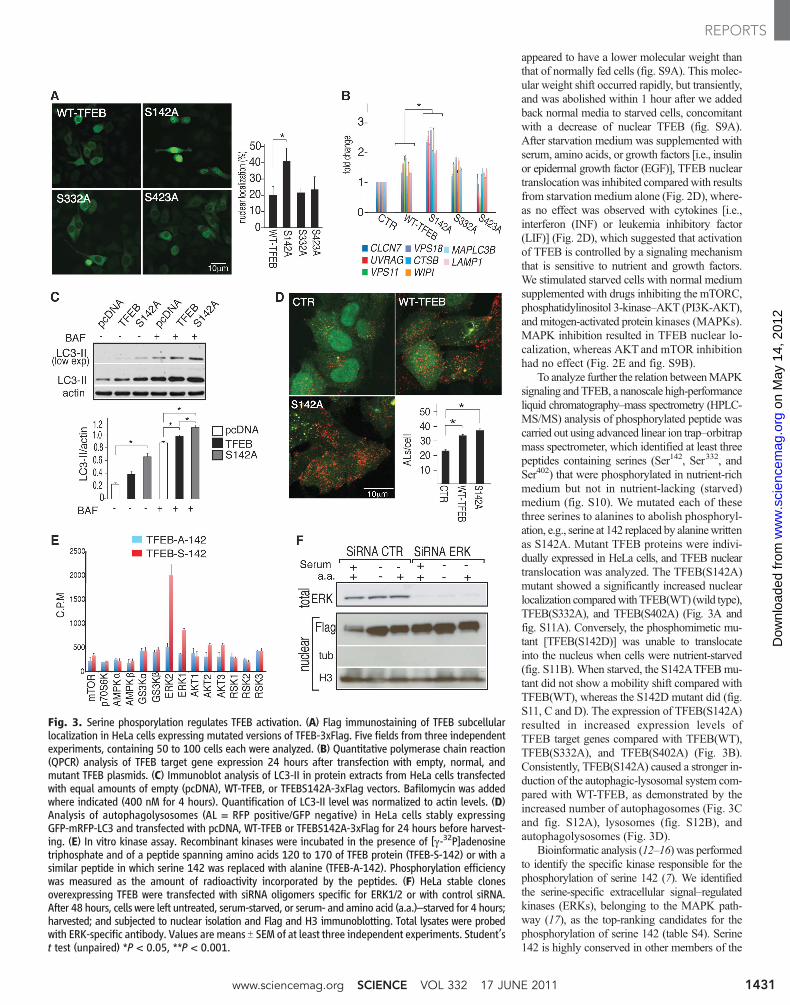
To analyze further the relation betweenMAPKsignaling and TFEB, a nanoscale high-performanceliquid chromatography–mass spectrometry (HPLC-MS/MS) analysis of phosphorylated peptide wascarried out using advanced linear ion trap–orbitrapmass spectrometer, which identified at least threepeptides containing serines (Ser142, Ser332, andSer402) that were phosphorylated in nutrient-richmedium but not in nutrient-lacking (starved)medium (fig. S10). We mutated each of thesethree serines to alanines to abolish phosphoryl-ation, e.g., serine at 142 replaced by alaninewrittenas S142A. Mutant TFEB proteins were indivi-dually expressed in HeLa cells, and TFEB nucleartranslocation was analyzed. The TFEB(S142A)mutant showed a significantly increased nuclearlocalization comparedwithTFEB(WT) (wild type),TFEB(S332A), and TFEB(S402A) (Fig. 3A andfig. S11A). Conversely, the phosphomimetic mu-tant [TFEB(S142D)] was unable to translocateinto the nucleus when cells were nutrient-starved(fig. S11B). When starved, the S142ATFEBmu-tant did not show a mobility shift compared withTFEB(WT), whereas the S142D mutant did (fig.S11, C and D). The expression of TFEB(S142A)resulted in increased expression levels ofTFEB target genes compared with TFEB(WT),TFEB(S332A), and TFEB(S402A) (Fig. 3B).Consistently, TFEB(S142A) caused a stronger in-duction of the autophagic-lysosomal system com-pared with WT-TFEB, as demonstrated by theincreased number of autophagosomes (Fig. 3Cand fig. S12A), lysosomes (fig. S12B), andautophagolysosomes (Fig. 3D).
Bioinformatic analysis (12–16) was performedto identify the specific kinase responsible for thephosphorylation of serine 142 (7). We identifiedthe serine-specific extracellular signal–regulatedkinases (ERKs), belonging to the MAPK path-way (17), as the top-ranking candidates for thephosphorylation of serine 142 (table S4). Serine142 is highly conserved in other members of the
Fig. 3. Serine phosporylation regulates TFEB activation. (A) Flag immunostaining of TFEB subcellularlocalization in HeLa cells expressing mutated versions of TFEB-3xFlag. Five fields from three independentexperiments, containing 50 to 100 cells each were analyzed. (B) Quantitative polymerase chain reaction(QPCR) analysis of TFEB target gene expression 24 hours after transfection with empty, normal, andmutant TFEB plasmids. (C) Immunoblot analysis of LC3-II in protein extracts from HeLa cells transfectedwith equal amounts of empty (pcDNA), WT-TFEB, or TFEBS142A-3xFlag vectors. Bafilomycin was addedwhere indicated (400 nM for 4 hours). Quantification of LC3-II level was normalized to actin levels. (D)Analysis of autophagolysosomes (AL = RFP positive/GFP negative) in HeLa cells stably expressingGFP-mRFP-LC3 and transfected with pcDNA, WT-TFEB or TFEBS142A-3xFlag for 24 hours before harvest-ing. (E) In vitro kinase assay. Recombinant kinases were incubated in the presence of [g-32P]adenosinetriphosphate and of a peptide spanning amino acids 120 to 170 of TFEB protein (TFEB-S-142) or with asimilar peptide in which serine 142 was replaced with alanine (TFEB-A-142). Phosphorylation efficiencywas measured as the amount of radioactivity incorporated by the peptides. (F) HeLa stable clonesoverexpressing TFEB were transfected with siRNA oligomers specific for ERK1/2 or with control siRNA.After 48 hours, cells were left untreated, serum-starved, or serum- and amino acid (a.a.)–starved for 4 hours;harvested; and subjected to nuclear isolation and Flag and H3 immunoblotting. Total lysates were probedwith ERK-specific antibody. Values are means T SEM of at least three independent experiments. Student’st test (unpaired) *P < 0.05, **P < 0.001.
www.sciencemag.org SCIENCE VOL 332 17 JUNE 2011 1431
REPORTS
on
May
14,
201
2w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
helix-loop-helix leucine zipper gene family suchas the microphthalmia transcription factor (fig.S13), which is phosphorylated by ERK2 (18). Evi-dence for ERK2-mediated TFEB phosphorylationcame from ERK2-TFEB coimmunoprecipitation(fig. S12C) in normal but not in starved mediumand from a peptide-based kinase assay showingthat mutation of Ser142 to alanine abolished ERK2-mediated phosphorylation (Fig. 3E and table S5).Small interfering RNA (siRNA)–mediated knock-down of ERK1/2 proteins induced TFEB nucleartranslocation to a similar extent as serum star-vation (Fig. 3F). Furthermore, the expression ofconstitutively active MEK in HeLa cells resultedin down-regulation of TFEB target gene expres-sion during starvation, and this effect was sim-ilar whether TFEB was knocked down or not(fig. S14A).
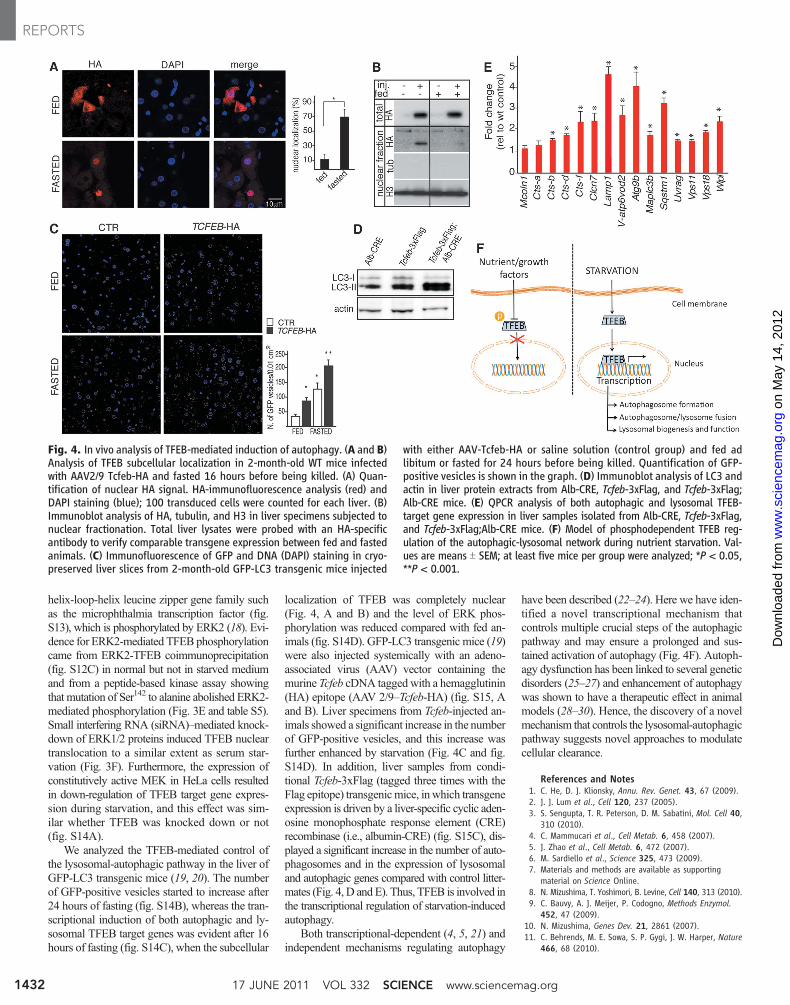
We analyzed the TFEB-mediated control ofthe lysosomal-autophagic pathway in the liver ofGFP-LC3 transgenic mice (19, 20). The numberof GFP-positive vesicles started to increase after24 hours of fasting (fig. S14B), whereas the tran-scriptional induction of both autophagic and ly-sosomal TFEB target genes was evident after 16hours of fasting (fig. S14C), when the subcellular
localization of TFEB was completely nuclear(Fig. 4, A and B) and the level of ERK phos-phorylation was reduced compared with fed an-imals (fig. S14D). GFP-LC3 transgenic mice (19)were also injected systemically with an adeno-associated virus (AAV) vector containing themurine Tcfeb cDNA tagged with a hemagglutinin(HA) epitope (AAV 2/9–Tcfeb-HA) (fig. S15, Aand B). Liver specimens from Tcfeb-injected an-imals showed a significant increase in the numberof GFP-positive vesicles, and this increase wasfurther enhanced by starvation (Fig. 4C and fig.S14D). In addition, liver samples from condi-tional Tcfeb-3xFlag (tagged three times with theFlag epitope) transgenicmice, in which transgeneexpression is driven by a liver-specific cyclic aden-osine monophosphate response element (CRE)recombinase (i.e., albumin-CRE) (fig. S15C), dis-played a significant increase in the number of auto-phagosomes and in the expression of lysosomaland autophagic genes compared with control litter-mates (Fig. 4, D and E). Thus, TFEB is involved inthe transcriptional regulation of starvation-inducedautophagy.
Both transcriptional-dependent (4, 5, 21) andindependent mechanisms regulating autophagy
have been described (22–24). Here we have iden-tified a novel transcriptional mechanism thatcontrols multiple crucial steps of the autophagicpathway and may ensure a prolonged and sus-tained activation of autophagy (Fig. 4F). Autoph-agy dysfunction has been linked to several geneticdisorders (25–27) and enhancement of autophagywas shown to have a therapeutic effect in animalmodels (28–30). Hence, the discovery of a novelmechanism that controls the lysosomal-autophagicpathway suggests novel approaches to modulatecellular clearance.
References and Notes1. C. He, D. J. Klionsky, Annu. Rev. Genet. 43, 67 (2009).2. J. J. Lum et al., Cell 120, 237 (2005).3. S. Sengupta, T. R. Peterson, D. M. Sabatini, Mol. Cell 40,
310 (2010).4. C. Mammucari et al., Cell Metab. 6, 458 (2007).5. J. Zhao et al., Cell Metab. 6, 472 (2007).6. M. Sardiello et al., Science 325, 473 (2009).7. Materials and methods are available as supporting
material on Science Online.8. N. Mizushima, T. Yoshimori, B. Levine, Cell 140, 313 (2010).9. C. Bauvy, A. J. Meijer, P. Codogno, Methods Enzymol.
452, 47 (2009).10. N. Mizushima, Genes Dev. 21, 2861 (2007).11. C. Behrends, M. E. Sowa, S. P. Gygi, J. W. Harper, Nature
466, 68 (2010).
Fig. 4. In vivo analysis of TFEB-mediated induction of autophagy. (A and B)Analysis of TFEB subcellular localization in 2-month-old WT mice infectedwith AAV2/9 Tcfeb-HA and fasted 16 hours before being killed. (A) Quan-tification of nuclear HA signal. HA-immunofluorescence analysis (red) andDAPI staining (blue); 100 transduced cells were counted for each liver. (B)Immunoblot analysis of HA, tubulin, and H3 in liver specimens subjected tonuclear fractionation. Total liver lysates were probed with an HA-specificantibody to verify comparable transgene expression between fed and fastedanimals. (C) Immunofluorescence of GFP and DNA (DAPI) staining in cryo-preserved liver slices from 2-month-old GFP-LC3 transgenic mice injected
with either AAV-Tcfeb-HA or saline solution (control group) and fed adlibitum or fasted for 24 hours before being killed. Quantification of GFP-positive vesicles is shown in the graph. (D) Immunoblot analysis of LC3 andactin in liver protein extracts from Alb-CRE, Tcfeb-3xFlag, and Tcfeb-3xFlag;Alb-CRE mice. (E) QPCR analysis of both autophagic and lysosomal TFEB-target gene expression in liver samples isolated from Alb-CRE, Tcfeb-3xFlag,and Tcfeb-3xFlag;Alb-CRE mice. (F) Model of phosphodependent TFEB reg-ulation of the autophagic-lysosomal network during nutrient starvation. Val-ues are means T SEM; at least five mice per group were analyzed; *P < 0.05,**P < 0.001.
17 JUNE 2011 VOL 332 SCIENCE www.sciencemag.org1432
REPORTS
on
May
14,
201
2w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

12. T. H. Dang, K. Van Leemput, A. Verschoren, K. Laukens,Bioinformatics 24, 2857 (2008).
13. Y. Xue et al., Mol. Cell. Proteomics 7, 1598 (2008).14. R. Amanchy et al., Nat. Biotechnol. 25, 285 (2007).15. R. Linding et al., Cell 129, 1415 (2007).16. F. Gnad et al., Genome Biol. 8, R250 (2007).17. W. Kolch, Nat. Rev. Mol. Cell Biol. 6, 827 (2005).18. T. J. Hemesath, E. R. Price, C. Takemoto, T. Badalian,
D. E. Fisher, Nature 391, 298 (1998).19. N. Mizushima, A. Yamamoto, M. Matsui, T. Yoshimori,
Y. Ohsumi, Mol. Biol. Cell 15, 1101 (2004).20. J. D. Rabinowitz, E. White, Science 330, 1344 (2010).21. J. Y. Yang et al., Nat. Cell Biol. 10, 138 (2008).22. C. He, B. Levine, Curr. Opin. Cell Biol. 22, 140 (2010).23. J. Kim, M. Kundu, B. Viollet, K. L. Guan, Nat. Cell Biol.
13, 132 (2011).24. D. F. Egan et al., Science 331, 456 (2011).25. E. Wong, A. M. Cuervo, Nat. Neurosci. 13, 805 (2010).26. C. Settembre et al., Hum. Mol. Genet. 17, 119 (2008).27. C. Settembre et al., Genes Dev. 22, 2645 (2008).
28. T. Hidvegi et al., Science 329, 229 (2010).29. B. Levine, G. Kroemer, Cell 132, 27 (2008).30. B. Ravikumar, D. C. Rubinsztein, Neuroreport 15, 2443
(2004).Acknowledgments: We thank H. Bellen, A. De Matteis,
G. Diez-Roux, M. Ferron, G. Karsenty, A. Luini, andH. Zoghbi for critical reading of the manuscript;R. Polishchuck for technical advice with the electronmicroscopy; K. Moreau, M. Renna, A. Ronza, andM. Palmieri for their contribution; and L. Naldini for thelentivirus production. We acknowledge the support ofthe Italian Telethon Foundation (C.S., C.D.M., V.P.,M.S., and A.B.); the Beyond Batten Disease Foundation(C.S., C.D.M., V.P., S.U.E., T.H., M.S., and A.B.);European Research Council Advanced Investigator grantno. 250154 (A.B.); a European Molecular BiologyOrganization long-term fellowship (C.S.); a WellcomeTrust Senior Research Fellowship (D.C.R.); and a U.K.Medical Research Council program grant (D.C.R.). Wewish to thank Cherie and James C. Flores for their
generous donation. We also thank S. Tooze for GFP-LC3cells, T. Yoshimori for the RFP-GFP, and P. Pandolfi forthe HA-ERK2 and ca-MEK; M. Krentz for the CAG-lox-LacZplasmid and for the useful suggestions during transgenegeneration; and the TIGEM AAV vector core for virusproduction. Confocal microscopy was supported bythe Intellectual and Developmental DisabilitiesResearch Center at the Baylor College of Medicine(5 P30 HD024064).
Supporting Online Materialwww.sciencemag.org/cgi/content/full/science.1204592/DC1Materials and MethodsFigs. S1 to S15Tables S1 to S5References
21 February 2011; accepted 10 May 2011Published online 26 May 2011;10.1126/science.1204592
AMPK Is a Direct AdenylateCharge-Regulated Protein KinaseJonathan S. Oakhill,* Rohan Steel, Zhi-Ping Chen, John W. Scott, Naomi Ling,Shanna Tam, Bruce E. Kemp
The adenosine monophosphate (AMP)–activated protein kinase (AMPK) regulates whole-body andcellular energy balance in response to energy demand and supply. AMPK is an abg heterotrimeractivated by decreasing concentrations of adenosine triphosphate (ATP) and increasing AMPconcentrations. AMPK activation depends on phosphorylation of the a catalytic subunit onthreonine-172 (Thr172) by kinases LKB1 or CaMKKb, and this is promoted by AMP binding to the gsubunit. AMP sustains activity by inhibiting dephosphorylation of a-Thr172, whereas ATP promotesdephosphorylation. Adenosine diphosphate (ADP), like AMP, bound to g sites 1 and 3 andstimulated a-Thr172 phosphorylation. However, in contrast to AMP, ADP did not directly activatephosphorylated AMPK. In this way, both ADP/ATP and AMP/ATP ratios contribute to AMPK regulation.
Matching energy supply with demand isessential for the survival and functionof organisms. In eukaryotes, strictmain-
tenance of the cellular energy status, as reflectedin relative concentrations of adenosine tri-, di-,and monophosphate (ATP, ADP, and AMP, re-spectively), is of paramount importance for thecontrol of all energy-requiringmetabolic processes.As energy demands increase, ATP is depleted,and increasing concentrations of ADP result in anincrease in the concentration of AMP due to theadenylate kinase equilibrium (1). Eukaryotes havea sensitive signaling mechanism that both moni-tors cellular energy status and acts to restoreenergy balance to protect the cell from metabolicstress caused by demand (e.g., muscle contrac-tion) or deprivation (e.g., ischemia). At the centerof this mechanism lies the AMP-activated pro-tein kinase (AMPK), an abg heterotrimeric serine-threonine kinase that senses the concentrations ofAMP.Once activated, AMPKphosphorylates andregulates key enzymes in all branches of metab-olism, as well as transcription factors that regulate
gene expression, to redirect cellular metabolismaway from anabolic, ATP-consuming pathwaysto energy-generating catabolic pathways.
Mammalian AMPK contains three adeninenucleotide-binding sites on the g subunit that areoccupied under physiological conditions: AMPand ATP bind interchangeably to sites 1 and 3,whereas site 4 binds AMP in a nonexchangeablemanner (2). In the Schizosaccharomyces pombeAMPKhomolog,AMPandATPbind interchange-ably at site 4 (3). AMPK is inactive unless phos-phorylated on Thr172 in the a-catalytic subunitactivation loop by the kinases LKB1 or CaMKKb(4). AMP binding stimulates phosphorylation ofa-Thr172, and this stimulation depends on the bsubunit being myristoylated (5). Once phospho-rylated on Thr172, AMPK is further directly acti-vated two- to fivefold byAMP, andAMP bindingalso suppresses inactivation due to dephospho-rylation of phosphorylated Thr172 (pThr172) byprotein phosphatases PP2a and PP2c (6). Oncephosphorylated on Thr172, neither direct acti-vation by AMP nor protection against dephos-phorylation requires b-subunit myristoylation (5).
Department of Protein Chemistry andMetabolism, St. Vincent’sInstitute of Medical Research, University of Melbourne, 41Victoria Parade, Fitzroy 3065, Victoria, Australia.
*To whom correspondence should be addressed. E-mail:[email protected]
pThr
172fo
ld in
crea
se
char
ge =
1.0
)
0
1
2
3
pThr172
αAdenylate charge
ATP (µM)ADP (µM)AMP (µM)
1.02000
00
0.9164630846
0.81349502149
0.71088624288
----
***
*****
0
1
2
3
pThr
172
fold
incr
ease
(v
sba
sal)
basal+AMP+ADP
α1β1γ1 α(1-392)
********
****
(vs
aden
ylat
e
172
----
----
******
**********
0
1
2
3
172
1 -
********
****
0
1
2
3
172
-
********
****
A B
Fig. 1. ADP stimulates CaMKKb-mediatedphosphorylation of AMPK a-Thr172. (A andB) Error bars denote mean T SEM of three toseven independent experiments. Statistical analyses weredone with Student’s t test. Thr172 phosphorylation. PP2c-dephosphorylated purified AMPK (a1b1g1, WT, or indi-cated mutants from COS7 cells) or purified Escherichia coli–expressed a1(1-392) fragments were phos-phorylated by CaMKKb T AMP or ADP. 200 mM adenosine nucleotides were used unless otherwise stated.Phosphorylation of Thr172 and total amount of a subunit were measured by simultaneous immunoblot. (A)Fold increase in pThr172 of a1b1g1 or a1(1-392) relative to basal (non–AMP- or –ADP-incubated) phos-phorylation for each protein (see fig. S1B). ****P < 0.0001 versus basal phosphorylation. (B) Regulationof phosphorylation of Thr172 by adenylate energy charge. Nucleotide concentrations were calculated (31)using a total adenine nucleotide pool of 2 mM and assumed equilibrium of the adenylate kinase reaction.5 mM MgCl2 was used to ensure ATP chelation. **P < 0.002, ***P < 0.001 versus phosphorylation atadenylate charge = 1. The immunoblot shown is a single representative experiment.
www.sciencemag.org SCIENCE VOL 332 17 JUNE 2011 1433
REPORTS
on
May
14,
201
2w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from





























