Embed Size (px)
Citation preview

Synthesis and Preliminary Evaluation of 18F-labeled 4-ThiaPalmitate as a PET Tracer of Myocardial Fatty Acid
OxidationTimothy R. DeGrado,1 Shuyan Wang,1 James E. Holden,2 R. Jerome Nickles,2
Michael Taylor2 and Charles K. Stone2
1DEPARTMENT OF RADIOLOGY, DUKE UNIVERSITY MEDICAL CENTER, DURHAM, NORTH CAROLINA USA; AND 2DEPARTMENTS
OF MEDICAL PHYSICS AND CARDIOLOGY, UNIVERSITY OF WISCONSIN-MADISON, MADISON, WISCONSIN, USA
ABSTRACT. Interest remains strong for the development of a noninvasive technique for assessment ofregional fatty acid oxidation rate in the myocardium. 18F-labeled 4-thia palmitate (FTP, 16-[18F]fluoro-4-thia-hexadecanoic acid) has been synthesized and preliminarily evaluated as a metabolically trapped probe ofmyocardial fatty acid oxidation for positron emission tomography (PET). The radiotracer is synthesized byKryptofix 2.2.2/K2CO3 assisted nucleophilic radiofluorination of an iodo-ester precursor, followed byalkaline hydrolysis and by purification by reverse phase high performance liquid chromatography. Biodistri-bution studies in rats showed high uptake and long retention of FTP in heart, liver, and kidneys consistentwith relatively high fatty acid oxidation rates in these tissues. Inhibition of carnitine palmitoyl-transferase-Icaused an 80% reduction in myocardial uptake, suggesting the dependence of trapping on the transport oftracer into the mitochondrion. Experiments with perfused rat hearts showed that the estimates of thefractional metabolic trapping rate (FR) of FTP tracked inhibition of oxidation rate of palmitate with hypoxia,whereas the FR of the 6-thia analog 17-[18F]fluoro-6-thia-heptadecanoic acid was insensitive to hypoxia. Invivo defluorination of FTP in the rat was evidenced by bone uptake of radioactivity. A PET imaging studywith FTP in normal swine showed excellent myocardial images, prolonged myocardial retention, and no boneuptake of radioactivity up to 3 h, the last finding suggesting a species dependence for defluorination of theomega-labeled fatty acid. The results support further investigation of FTP as a potential PET tracer forassessing regional fatty acid oxidation rate in the human myocardium. NUCL MED BIOL 27;3:221–231, 2000.© 2000 Elsevier Science Inc. All rights reserved.
KEY WORDS. Fatty acids, b-Oxidation, PET, Thia fatty acids
INTRODUCTION
Interest has remained strong for the development of a noninvasivetechnique for regional assessment of b-oxidation of long-chain fattyacids. b-Oxidation is depended upon for the majority of adenosinetriphosphate production in the heart in normal conditons. Nonin-vasive assessment of regional b-oxidation rate may allow earlydetection of metabolic abnormalities in the myocardium that maypresage irreversible tissue injury. However, the development of ameasurement technique using a gamma-emitter labeled fatty acidanalog has proved problematic in spite of extensive efforts withnumerous tracers.
[1-11C]Palmitate (CPA) has been studied with positron emissiontomography (PET) in animals and humans as a tracer of myocardialfatty acid metabolism. In nonischemic conditions, washout of theoxidative product 11CO2 predominates the early clearance phase,whereas turnover of CPA incorporated into complex lipids deter-mines the slower late clearance phase. The clearance rate and therelative size of the rapid phase correlate with cardiac work (11) andmyocardial oxygen consumption rate (27). Noninvasive delineationof regional fatty acid oxidation rates from dynamic PET imaging
studies with CPA were proposed on the basis of these findings. Arelatively complex tracer kinetic model is necessary to model CPAkinetics for quantitative measurement of fatty acid utilization thatincludes catabolite and lipid storage compartments (13). Bergmannet al. (2) developed a seven-parameter compartmental modelingtechnique for quantification of CPA kinetics that was in goodagreement with measured arteriovenous differences of fatty acidsacross the canine heart in nonischemic conditions. The modelingtechnique may not be applicable in ischemic regions due to lowsignal-to-noise ratio in the images. Furthermore, the enhancedback-diffusion of extracted nonmetabolized tracer from ischemicmyocardium is kinetically indistinct from clearance of 11CO2 duringthe early clearance phase (10). In this situation, the kinetics of theearly clearance phase do not quantitatively reflect the oxidative rateof fatty acids by the myocardium. Structurally modified fatty acidtracers have been developed to prolong the retention of oxidativemetabolites in tissues, allowing the application of more simplifiedimaging protocols and data analysis strategies. 11C-labeled b-methylheptadecanoic acid (BMHA) was developed as a metabolicallytrapped tracer (9, 19). BMHA showed prolonged retention inmyocardium due to incorporation mainly into triglycerides (1).Likewise, myocardial retention of a radioiodinated b-methylatedfatty acid was found to be insensitive to decreases of b-oxidationrate secondary to pharmacologic inhibition of carnitine palmitoyl-transferase-I (CPT-I) (5).
The 6-thia fatty acid analog 14(R,S)-[18F]fluoro-6-thia-heptade-
Address correspondence to: Timothy R. DeGrado, Ph.D., Duke UniversityMedical Center, Dept. of Radiology, Box 3949, Durham, NC 27710, USA;e-mail: [email protected].
Received 17 July 1999.Accepted 15 October 1999.
Nuclear Medicine & Biology, Vol. 27, pp. 221–231, 2000 ISSN 0969-8051/00/$–see front matterCopyright © 2000 Elsevier Science Inc. All rights reserved. PII S0969-8051(99)00101-8

canoic acid (FTHA; Fig. 1) was developed as a metabolicallytrapped tracer of b-oxidation in myocardium-based (4, 7) pharma-cology studies with 4-thia fatty acids (31). 4-Thia fatty acids areb-oxidized in the mitochondria to 4-thia-enoyl-CoAs, which ap-parently accumulate in the mitochondrion. Initial studies withFTHA have shown it to be metabolically retained in the myocar-dium and have excellent imaging properties in normal humansubjects (8, 21) and in patients with coronary artery disease (28).Experimental validation studies have shown good correspondenceof trapping of FTHA in the myocardium with fatty acid oxidationin various conditions; however, FTHA accumulation in the myo-cardium failed to track decreases in b-oxidation in hypoxic condi-tions (35). Evidently, b-oxidation is not solely responsible forintracellular retention of the radiolabel and the hypoxic conditionpresumably exposes retention of radiolabel in acyl intermediates orcomplex lipids. The suboptimal specificity of FTHA for b-oxidationmotivated further radiotracer development, specifically 4-thia fattyacid analogs.
The present study reports the synthesis and preliminary evalua-tion of 18F-labeled 4-thia palmitate (FTP) as a myocardial b-oxi-dation probe for PET.
METHODS AND MATERIALSSynthesis of Radiotracers
Chemical structures are shown in Figure 1 for the 18F-labeled thiafatty acids that were synthesized and evaluated: FTP and 17-[18F]fluoro-6-thia-heptadecanoic acid (17F6THA).
Chemicals were of analytical grade. Dry acetonitrile was obtainedcommercially (Pierce, Rockford, IL USA). 1H-Nuclear magneticresonance (NMR) spectra were recorded with a Varian (PaloAlto, CA USA) Unity 500 MHz spectrometer using CDCl3 as asolvent (Me4Si, 0.00 ppm). Rf values refer to thin layer chromatog-raphy (TLC) performed on silica gel with the solvent system noted.Routine column chromatography was performed under normalpressure with silica gel (100–200 mesh) and the solvent systemnoted.
Methyl 16-iodo-4-thia-hexadecanoate (1)
1,12-Diiodododecane (2.66 g, 6.3 mmol) was dissolved in 20 mLacetonitrile. Methyl 3-mercaptopropionate (0.76 g, 6.3 mmol) andK2CO3 (1.1 g, 8 mmol) were added and the mixture was allowed toreact at room temperature for 20 h. The mixture was acidified with
ice-diluted HCl and the solution was extracted twice with ether(20 mL). The combined ether fractions were washed successivelywith dilute NaHCO3, water, and brine; dried (MgSO4); andevaporated under reduced pressure. The product 1 (0.84 g, 32%yield) was isolated by column chromatography (hexane/ethyl ace-tate 9:1) and recrystallized in MeOH. TLC (hexane/ethyl acetate9:1) Rf 5 0.4. Melting point 5 33°C. 1H-NMR 1.4 (m, 20H, CH2),2.52 (t, 2H, C(2)H2), 2.61 (t, 2H, C(5)H2), 2.79 (t, 2H, C(3)H2),3.20 (t, 2H, C(16)H2), 3.71 (s, 3H, -COO-CH3).
16-Fluoro-4-thia-hexadecanoic Acid (2)
Fluorination of 1 (0.2 g, 0.5 mmol) was performed by addition oftetrabutylammonium fluoride (TBAF; 4 mmol as 1 M tetrahydro-furan solution). The mixture was allowed to react at room temper-ature for 20 h. The resulting flouro-ester was hydrolyzed by additionof 1 mL 1 N potassium hydroxide (KOH) and 1 mL ethanol for 2 hat room temperature. The product 2 (0.05 g, 34% yield) was isolatedby column chromatography (hexane:ether:acetic acid, 3:1:0.1) andrecrystallized in hexane. TLC (hexane/ether/acetic acid 3:1:0.1)Rf 5 0.25. Melting point 5 59°C. 1H-NMR 1.4 (m, 20H, CH2),2.55 (t, 2H, C(2)H2), 2.65 (t, 2H, C(5)H2), 2.80 (t, 2H, C(3)H2),4.44 (dt, 2H, C(16)H2, JHF 5 47.5, JHH 5 6.2 Hz).
Methyl 17-tosyloxy-6-thia-heptadecanoate (3)
To a solution of 11-bromo-1-undecanol (10 g, 40 mmol) indimethyl sulfoxide (DMSO; 80 mL) was added thiourea (3.64 g,48 mmol), and the mixture was allowed to react at room tempera-ture for 21 h. The mixture was extracted twice with hexane (20 mL)to remove the unreacted bromo alcohol. To the DMSO fraction wasadded 2 N KOH (50 mL) and the mixture was heated at 80°C for5 min, releasing the thiol 11-mercapto-undecanol. The mixture wasacidified (HCl) and extracted twice with ether (40 mL). Thecombined ether phases were washed successively with water andbrine, dried over MgSO4, and evaporated under reduced pressure.The resulting thiol (approximately 7.2 g) was made to react withmethyl 5-bromo-pentanoate (7 g, 37 mmol) according to theprocedure for compound 1. Crystallization of the product in hexaneyielded 5.5 g (18 mmol, 48% yield from the thiol) of methyl17-hydroxy-6-thia-heptadecanoate. TLC (hexane:ether, 1:1)showed a single product at Rf 5 0.3. Without further characteriza-tion, the hydroxy-ester (4 g, 13 mmol) was caused to react withTs-Cl (2.7 g, 14 mmol) and pyridine (14 mmol) in CH2Cl2 (40 mL)
FIG. 1. Chemical structures of18F-labeled thia fatty acid ana-logs. The (v-3) labeled, 6-thiaanalog 14(R,S)-[18F]fluoro-6-thia-heptadecanoic acid (FTHA) hasbeen previously developed (6).FTP, 18F-labeled 4-thia palmi-tate; 17F6THA, 17-[18F]fluoro-6-thia-heptadecanoic acid.
222 T. R. DeGrado et al.

at 5°C for 4 h. The mixture was acidified with ice-diluted HCl, andthe organic layer separated, dried with MgSO4, and evaporatedunder reduced pressure. The product 3 (2.4 g, 34% yield) wasisolated by column chromatography (hexane:ethyl acetate, 7:3) andrecrystallized in hexane. TLC (hexane:ethyl acetate, 7:3) Rf 5 0.6.Melting point 5 54°C. 1H-NMR 1.3 (m, 22H, CH2), 2.38 (t, 2H,C(2)H2), 2.5 (m, 7H, C(5)H2, C(7)H2, (tosyl) CH3), 3.7 (s, 3H,-COO-CH3), 4.0 (t, 2H, C(17)H2), 7.6 (m, 4H, aryl).
17-Fluoro-6-thia-heptadecanoic Acid (4)
Compound 3 (1 g, 1.8 mmol) was caused to react with TBAF(2.5 mmol) according to the procedure for compound 2. Theresultant fluoro-ester was hydrolyzed in KOH/ethanol as previouslydescribed and crystallized twice in ethanol/water to yield 0.4 g (70%yield) of product 4. Melting point 5 54°C. 1H-NMR 1.3 (m, 22H,CH2), 2.38 (t, 2H, C(2)H2), 2.5 (m, 4H, C(5)H2, C(7)H2), 4.44(dt, 2H, C(17)H2, JHF 5 47.3, JHH 5 6.2 Hz).
18F-Labeling Procedure
The precursors for FTP and 17F6THA were compounds 1 and 3,respectively. To a 2 mL glass vial was added Kryptofix 2.2.2(10 mg), acetonitrile (0.5 mL), and 20 mL of a 9% K2CO3 solutionin water. [18F]Fluoride, which was produced via proton bombard-ment of H2
18O (.95 atom %), was then added, the vessel placed inan aluminum heating block at 85°C, and the solvent evaporatedunder a stream of helium or nitrogen. The residue was further driedby azeotropic distillation with acetonitrile (2 3 0.3 mL). A solutionof the precursor (approximately 2 mg) in acetonitrile (0.5 mL) wasadded, and the vial was sealed and returned to the heating block.Reaction time was 15 min. The vial was briefly cooled by placing inice water. The incorporation of [18F]fluoride was monitored byradio-TLC (hexane/ethyl acetate 3:1). Rf values were 0.0 and .0.4for [18F]fluoride and [18F]fluoro-ester, respectively.
Subsequent hydrolysis of the resulting [18F]fluoro-ester was per-formed in the same vessel by the addition of 0.15 mL 0.2N KOHand continued heating at 90°C for 4 min. The mixture was cooled,acidified with concentrated acetic acid (25 mL), filtered, andapplied to the preparative high performance liquid chromatography(HPLC) column (Table 1). An in-line ultraviolet detector(210 nm) was used to monitor the elution of unlabeled materials.The [18F]fluoro-fatty acid fraction was collected, evaporated todryness, formulated in 1–2% albumin in isotonic NaCl solution,and filtered through a 0.22 mm filter (Millex-GS, Millipore,Bedford, MA USA).
Biodistribution Studies in Rats
Female Sprague-Dawley rats (180–225 g) were fasted overnight orfed ad libitum. The rats were anesthetized with pentobarbital
(75 mg/kg) before injection of radiotracer, remaining anesthetizedthroughout the study. The 18F-labeled radiotracer (20–40 mCi) wasinjected into the femoral vein. A prescribed duration of time wasallowed before procurement of heart, liver, lung, blood, kidney,bone (rib), brain (whole), and skeletal muscle. The tissues werecounted and weighed. Radiotracer uptake was calculated as:
Uptake (%dose kg/g) 5CPM(tissue) 3 Body Wt.(kg) 3 100
Tissue Wt.(g) 3 CPM (dose)(1)
Nonesterified fatty acid (NEFA) concentrations in plasma at timeof euthanasia were measured colorimetrically (WACO Bioproducts,Richmond,VA USA).
In one group of fasted rats, the CPT-I inhibitor etomoxir(40 mg/kg, Byk Gulden, Konstanz, Germany) was administeredintrperitoneally 2 h prior to FTP injection.
Crude analysis of the nature of the metabolites in the heart wasperformed by a Folch-type extraction procedure as previouslydescribed (7). The rat heart was excised 30 min postinjection andthoroughly homogenized and sonicated (20 s) in 7 mL chloroform/methanol (2:1) at 0°C. Urea (40%, 1.75 mL) and 5% sulfuric acid(1.75 mL) were added and the mixture was sonicated for anadditional 20 s. After centrifugation for 10 min, aqueous, organic,and protein interphase fractions were separated and counted. Theorganic phase was further analyzed by silica-gel TLC for radiola-beled diglycerides, fatty acids, triglycerides, and cholesterol ester aspreviously described (7).
Isolation and Perfusion of Rat Hearts
Hearts were excised from pentobarbital anesthetized femaleSprague-Dawley rats (225–250 g) that had been fed ad libitum.Following cannulation of the aorta, retrograde (Langendorff) per-fusion of the hearts was commenced using a peristaltic pump. Theperfusion medium was Krebs-Henseleit (K-H) bicarbonate buffer ofthe following composition (mmol/L): Na1 143; K1 5.9; Ca21 1.85;Mg21 1.0; Cl2 125.6; SO4
22 1.18; H2PO42 1.2; HCO3
2 25, glucose10; palmitate 0.15; and albumin 0.15. The perfusate was prefilteredusing 5 mm in-line filters (Millipore Corp., Bedford, MA USA).Under standard conditions, the perfusate was gassed with a 95%/5%oxygen (O2)/carbon dioxide CO2 mixture. To assess the tracerkinetics in conditions of hypoxia, the fraction of oxygen waslowered to 35% while maintaining the CO2 fraction at 5%. Themakeup gas was nitrogen. The apparatus utilized water-jacketedvessels and a heater/circulator to deliver the medium to the heart ata temperature of 37°C. Aortic pressure was monitored by a pressuretransducer and allowed measurement of heart rate and mean aorticpressure. Hearts were not externally paced. Coronary flow wasmeasured manually.
Bolus Injection of Radiotracer
For experiments using bolus administration of 18F-labeled fattyacids, a catheter (PE-30 tubing) was inserted into the aorticcannula. The opening of the catheter was situated near the root ofthe coronary artery of the perfused rat heart. The standard protocolfor bolus injection studies consisted of a 20-min stabilization period,after which the radiotracer (approximately 5 mCi in 50 mL) wasmanually injected through the catheter in less than 0.5 s. A 20-minwashout period followed.
TABLE 1. Semi-preparative Reverse Phase HPLC CapacityFactors (k*) of Fatty Acid Analogs (Nucleosil, C-18 (10m)250 3 10 mm, Flow 5 4.3 ml/min, Mobile PhaseMeOH/H2O/AcOH 90:9.5:0.5)
Compound k*
16-Iodo-4-thia-hexadecanoic acid 5.116-Fluoro-4-thia-hexadecanoic acid (2, FTP) 3.217-Tosyloxy-6-thia-heptadecanoic acid 3.017-Fluoro-6-thia-heptadecanoic acid (4,17F6THA) 3.5
18F-labeled 4-thia Palmitate in Heart 223

Pulse-perfusion of Radiotracer
For pulse-perfusion of radiotracer, two reservoirs of perfusate wereemployed. Before each study, one reservoir was filled with 500 mLof perfusion medium containing 100–200 mCi/L 18F-labeled fattyacid and approximately 50 mCi/L [9,10-3H]palmitate. A three-wayvalve was employed for switching the perfusate between the twosources. The standard pulse-perfusion protocol consisted of a20-min stabilization period without tracers, a 20-min perfusion withradiotracers, and a 30–40-min washout period (without tracers) insuccession.
The 18F radioactivity concentration of the wash-in medium Cp
(CPM/mL) was measured in a NaI well-counter. At the end ofperfusion, ventricles were opened and gently blotted on papertoweling. Hearts were then placed in preweighed tubes, weighed,and counted in the NaI well-counter. As a consequence of theprolonged washout period, no radioactivity remained in the leftheart chambers or perfusion cannulae at the end of perfusion. Theapparent distribution volume (ADV) of radioactivity in the wholeheart was calculated from the sum of radioactivity in the heart andthe paper toweling (Ah), the dry mass of the heart (Mh), and Cp:
ADV (tend) 5 Ah/(Cp 3 Mh) (2)
where tend was the time at the end of perfusion.
Estimation of Palmitate Oxidation Rate
Hearts were perfused with [9,10-3H]palmitate at a concentration of50 mCi/L. Arterial samples were collected from the reservoir used toperfuse the heart. Coronary effluent samples were collected after a10 min equilibration period. Tritiated water was separated fromnonmetabolized 3H-palmitate in the coronary samples by an organicsolvent extraction procedure described by Saddik and Lopaschuk(25). Fractional palmitate oxidation rate (mL/min/g dry) wascalculated as the product of mass-specific coronary flow (mL/min/gdry) and the fraction of coronary effluent 3H2O concentrationrelative to perfusate 3H-palmitate concentration at steady-state.Palmitate oxidation rate (nmol/min/g dry) was calculated by mul-tiplying the fractional rate by the concentration of palmitate in theperfusate (nmol/mL).
Acquisition and Normalization of 18F Time-activityCurves in Isolated Rat Hearts18F Radioactivity was externally monitored using two lead-colli-mated 2 3 2 inch NaI(Tl) scintillation probes configured electron-ically for g,g-coincidence detection. The true coincidence countrate in the heart (Yi [counts/s]) was calculated as the difference ofprompt coincidence rate and random coincidence rate, the latter ofwhich was estimated by the delayed signal technique. The truecoincidence count rate was corrected for radioactive decay.
For bolus injection studies, the time-activity curves were normal-ized by setting the peak count rate equal to 1.0. For studies in whichradiotracers were administered by pulse-perfusion, the resultanttime-activity curve was converted to units of ADV. This wasaccomplished by multiplication of each data point Yi by a calibra-tion factor (fc),
ADVi~ml/g dry! 5 Yi fc (3)
where fc was given by the equation:
fc Sml/g drycounts/sD 5
ADV~tend!
Ye(4)
where ADV(tend) is given by Eq. 3, and Ye is the average of Y overthe last 10 s of perfusion.
Tracer Kinetic Modeling
For bolus injection studies, the time-activity curves were least-squares fit to the three-exponent model:
y 5 A1 exp(2E1 t) 1 A2 exp(2E2 t) 1 A3 exp(2E3 t) (5)
For pulse-infusion studies, a two-compartment model (Fig. 2) wasfit to the time-activity curves. The first compartment (C1) repre-sents radiolabel in reversible pools in the tissue and is in exchangewith the blood (perfusate) through rate constants k1 and k2. Thesecond compartment (C2) represents “trapped” metabolites in thetissue. The production rate of trapped metabolites is represented byrate constant k3. The slow clearance of radiolabel from the heart isdue to loss of radioactivity from the trapped compartment. Thisclearance process is represented by the rate constant k4. A bloodvolume term was included in the model to account for radiotracerpresent in the vascular spaces. The differential equations describingthe kinetics of the radiolabel are:
dC1(t)dt
5 k1Cp~t! 2 ~k2 1 k3!C1~t! (6)
dC2(t)dt
5 k3C1~t! 2 k4 C2~t! (7)
where Cp(t) is the concentration of radiotracer in the perfusate.The total apparent distribution volume of radiolabel in the heart ismodeled as:
FIG. 2. Two-compartment model for describing the kineticsof 18F-labeled thia fatty acid analogs in isolated rat hearttissue. In addition to these compartments representing radio-activity in tissue compartments, a blood volume term isincluded in the modeling technique (Eq. 8) to account forradiolabel in the vascular spaces within the field of view of theradiation detectors.
224 T. R. DeGrado et al.

ADV(t) 5 BV Cp~t! 1 ~1 2 BV!~C1~t! 1 C2~t!! (8)
The compartmental model solutions were derived numerically usinga fourth order Runge-Kutta integrator. Parameter estimation uti-lized a nonlinear least-squares fitting routine (22).
An operational equation is derived for the fractional metabolicrate of tracer (FR) as the rate of entry of radiolabel into compart-ment C2 at quasi-steady-state (dC1/dt 5 0). In this case, thedistribution volume of nonmetabolized tracer in tissue is given as:
V1 5C1
CpU
ss
5k1
k2 1 k3(9)
The fractional tracer metabolic rate (FRFTP) (mL/min/g dry) intissue is estimated as:
FR 5dC2(t)/dt
CpU
ss
5 V1k3 5k1k3
k2 1 k3(10)
PET Imaging of FTP in Normal Swine
Dynamic PET imaging of two normal swine (45–52 kg) wasperformed using the Advance scanner (General Electric MedicalSystems, Milwaukee, WI USA). Emission scanning was preceededby a whole-body transmission scan. Emission scanning commencedwith a dynamic study of the heart until 20 min postinjection,followed by three consecutive whole-body scans for a total scantime of 3 h. The dynamic heart scan sequence was 12 3 10 s, 3 330 s, and 1 3 5 min. Image analysis was performed by manualdrawing of regions on the PET images. Regions were analyzed tocreate time-activity curves for left ventricular blood pool, myocar-dium (septum), lung, and liver. Results were expressed as %dose/mLof tissue.
Statistical Analysis
Data are expressed as mean 6 standard deviation. The t-test(two-tailed) for unpaired samples was used to compare means of twogroups of hearts.
RESULTSRadiotracer Syntheses
The omega-labeled 18F-thia fatty acids FTP and 17F6THA weresynthesized in decay-corrected radiochemical yields ranging from 25
to 60. Reverse phase HPLC purification successfully separated the18F fluoro fatty acids from their respective hydrolyzed precursorcompounds, although the tosyloxy fatty acid (hydrolyzed product ofprecursor 3) eluted nearest to the 18F product 17F6THA, necessi-tating the discarding of a small fraction of 17F6THA peak to avoidcontamination. Use of the iodo-ester precursor for FTP was advan-tageous because its corresponding fatty acid eluted long after theelution of the 18F fluoro fatty acid product. The correspondingbromo-ester was also evaluated (data not shown) but found lesspreferable to the iodo-ester because it had a lower melting point,leading to less practical storage and handling.
Biodistribution Studies in Rats
Table 2 show biodistribution of FTP in rats fed ad libitum or fastedovernight. Arterial nonesterified fatty acid concentrations were0.19 6 0.06 mM and 0.43 6 0.07 mM (p , 0.001) in fed and fastedrats, respectively. The highest uptake for all the tracers wasobserved in heart, liver, and kidney, regardless of dietary status. Thisbiodistribution is consistent with the relatively high b-oxidationactivity in these three organs. Radioactivity levels in blood, lung,kidneys, and skeletal muscle were higher in the fed state than in thefasted state. Considering the twofold higher concentration ofnonesterified fatty acids in the blood with fasting, the nearlyequivalent fractional uptake of FTP in the heart and liver agreeswith the expected increase of fatty acid utilization by these organsduring fasting. Myocardial and hepatic uptake was not significantlychanged for FTP in fasted rats from 30 to 120 min, although kidneyuptake was somewhat decreased over time. The v-labeled analogFTP showed significantly higher (p , 0.01) bone uptake thanpreviously observed using a (v-3)-labeled thia-substituted fatty acid(6), consistent with more extensive defluorination of v-18F-labelednon-b-oxidizable fatty acid analogs in rodents.
Pretreatment of fasted rats with the CPT-I inhibitor etomoxircaused an 80% reduction of myocardial uptake of FTP at 30 minpostinjection (Table 2) despite higher levels of radioactivity re-maining in blood. Although the heart:blood ratio was 11.0 incontrols at 30 min, the same ratio was 1.1 in etomoxir-treated rats.Kidney uptake was 37% lower in etomoxir-treated rats, but liveruptake was unchanged. Higher uptakes observed in lung, brain, andskeletal muscle of etomoxir-treated rats may be related to the higherlevel of radioactivity in the blood.
Table 3 shows the distribution of 18F radioactivity in heartsexcised from control and etomoxir-treated rats at 30 min postin-jection of FTP. The results show the predominant fate of the
TABLE 2. Uptake (% dose kg/g) of FTP in Sprague-Dawley Rats
Dietary StatusInterventionTime p.i. (min)n
Fed Control307
Fasted Control305
Fasted Control120
3
Fasted Etomoxir305
Heart 0.423 6 0.203 0.320 6 0.170 0.172 6 0.046 0.060 6 0.012*Blood 0.053 6 0.013** 0.032 6 0.008 0.026 6 0.003 0.054 6 0.012**Lung 0.167 6 0.051** 0.078 6 0.018 0.060 6 0.007 0.101 6 0.037Liver 1.24 6 0.44 1.15 6 0.16 1.11 6 0.15 1.12 6 0.07Kidney 0.39 6 0.04** 0.248 6 0.041 0.183 6 0.016* 0.156 6 0.023**Bone 0.152 6 0.048 0.238 6 0.073 0.577 6 0.147* 0.165 6 0.094Brain 0.021 6 0.002 0.022 6 0.003 0.018 6 0.003 0.048 6 0.016*Skeletal muscle 0.037 6 0.006** 0.020 6 0.005 0.017 6 0.002 0.035 6 0.006**
Note: Rats were female, 180–200 g, fasted overnight. Etomoxir-treated rats were given 40 mg/kg etomoxir 2 hr prior to tracer injection. Uptake is normalizedto body mass (kg). *p , 0.05 versus 30 min fasted controls. ** p , 0.01 versus FTP uptake at 30 min fasted controls in same organ.
18F-labeled 4-thia Palmitate in Heart 225

radiolabel in control hearts to be distribution into the proteininterphase layer containing precipitated protein and to a lesserextent undisturbed membrane fragments. This fraction is decreasedby 89% in etomoxir-treated hearts, which is consistent with thehypothesis that protein-bound metabolites of FTP are produced as aresult of its oxidation in the mitochondrion. Aqueous solubleradiolabeled metabolites of FTP are relatively minor. Organicsoluble metabolites are predominantly polar species that remainedunidentified by this assay. They could be phospholipids or fatty acylintermediates of FTP that did not remain protein-bound. Incorpo-ration of FTP into triglycerides was found to be very small in bothcontrol and CPT-I inhibited conditions.
Studies in Isolated Langendorff Perfused Rat Hearts
Studies were performed in Langendorff perfused rat hearts todelineate the kinetics of thia fatty acid analogs and determine therelationship of tracer retention to fatty acid oxidation rate innormoxic and hypoxic myocardium. The perfusate was K-H bufferwith 5 mM glucose, 0.15 mM albumin, and 0.15 mM palmitate. Theperfusate was gassed with mixtures of 95% O2/5% CO2 (normoxiccondition) or 35% O2/60% N2/5% CO2 (hypoxic condition). Flowwas maintained at 7 mL/min.
Figure 3 shows a representative time-activity curve in a Langen-dorff perfused heart given a rapid bolus injection of FTP at theaortic root. The time-activity curves were fit to a three-exponentmodel (Table 4). The most rapid phase represents clearance ofvascular spaces. The intermediate phase represents extracted butunretained radiolabel in tissue. The clearance half-time of thisphase is approximately 11 s. The slowest component evidencesturnover of the tracer in one or more slowly clearing compartments.On average, approximately 20% of initially extracted tracer is takenup into the slow turnover compartment(s). The mean clearancehalf-time of the slowest clearance phase is approximately 90 min.This “trapped” component is clearly delineated from the more rapidclearance components by 4 min after injection.
The relationship of the steady-state accumulation rate of FTPand 17F6THA to palmitate oxidation rate was determined inisolated rat hearts using simultaneous pulse-infusion of a 18F-labeledtracer and [9,10-3H]palmitate. Experiments were performed in
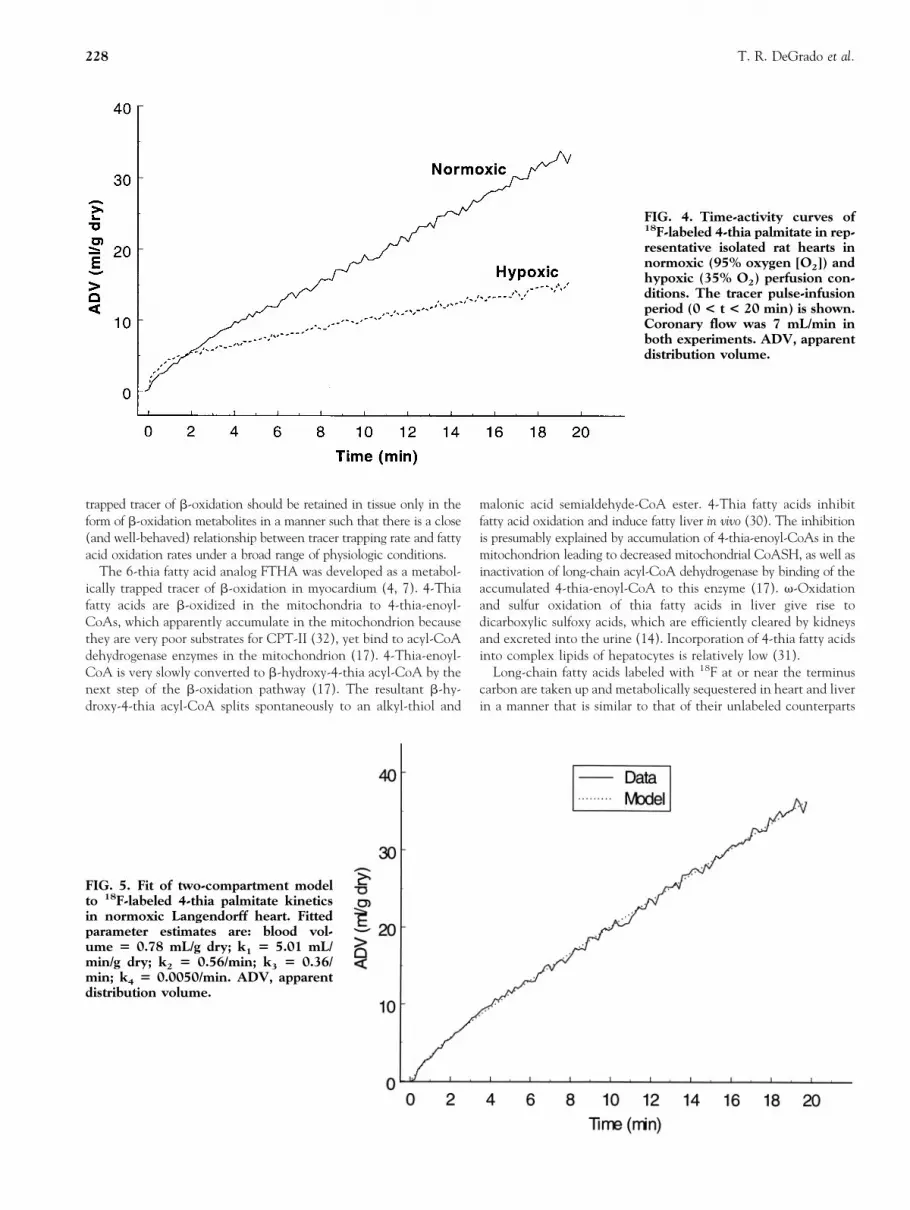
normoxic (95% O2) and hypoxic (35% O2) conditions, the latterrepresenting a condition in which fatty acid oxidation rate islimited by mitochondrial b-oxidation rate. After perfusion for20–30 min with radiotracers at constant concentration in theperfusate, perfusion without radiotracers (washout) followed for 40min. The heart rate of the unpaced heart was 25% lower in thehypoxic condition (205 6 33 beats/min [hypoxic] vs. 276 6 33beats/min [control]; p , 0.01). Mean aortic pressure was notsignificantly different in the hypoxic condition (47 6 8 mmHg[hypoxic] vs. 43 6 4 mmHg [control]). Therefore, changes in fattyacid oxidation may be partially due to the moderately lowerworkload in addition to intracellular hypoxia. Figure 4 showsrepresentative time-activity curves for FTP during the uptake phase,illustrating the markedly lower accumulation rate of tracer in thehypoxic condition. The time-activity curves were fit to a two-compartment model (Fig. 5) with compartments representing re-versible precursor (C1) and “trapped” metabolite (C2) compart-ments. Fitting of data from the last 25–30 min of the washout periodgave estimates of the slow clearance rate of “trapped” radiolabel(k4). This value of k4 was subsequently fixed in the least-squaresfitting of k1, k2, and k3 in the two-compartment model. From thefitted rate constants, FR was calculated from Eq. 10.
Figure 6 shows the correlation of fractional metabolic rate forFTP and 17F6THA with palmitate oxidation rates. Table 5 sum-marizes the FR estimates and palmitate oxidation rates. Metabolictrapping rates of the 4-thia analog FTP were found to correlate wellwith rates of palmitate oxidation in normal and hypoxic conditions(r 5 0.85). In contrast, 17F6THA metabolic trapping rate wasinsensitive to decreases in palmitate oxidation rate observed inhypoxia. The FR of FTP (FRFTP) was approximately 60% thefractional palmitate oxidation rate. Mean FRFTP was decreased 50%in the hypoxic group relative to the normoxic group, tracking adecrease of 40% in the oxidation rate of palmitate with hypoxia(Table 5).
PET Imaging of FTP Uptake in Normal Swine
Whole-body dynamic PET imaging studies of FTP (5.5–7.6 mCibolus) in two anesthetized normal swine showed rapid accumulationof tracer in heart, liver, and kidneys. No bone uptake of 18F was
TABLE 4. Parameter Estimates of 3-exponent Fits to FTP Kinetics in Langendorff Perfused Rat Heart
A1 E1 (min21) A2 E2 (min21) A3 E3 (min21)
0.63 6 0.14 35.4 6 9.0 0.28 6 0.15 3.67 6 0.79 0.078 6 0.023 0.0076 6 0.0016
Note: Fitted model is y 5 A1 exp (2E1t) 1 A2 exp (2E2t) 1 A3 exp (2E3t). Three hearts were perfused.
TABLE 3. Distribution of 18F in Rat Hearts at 30 Min after Injection of FTP. Rats were Fasted Overnight. Values areExpressed as % Dose kg (Body wt)/g (Heart) for Direct Comparison with Total Uptake in Same Units as Given in Table 2.Values are Corrected for Incomplete Recovery of Radioactivity from the Homogenization Procedure. The Organic SolubleMetabolites were Analyzed by Silica-gel TLC using Petroleum Ether/ether/acetic Acid (70:30:1). The Rf Values andDesignations (DG 5 diglycerides, FA 5 fatty acids, TG 5 triglycerides) of the TLC Fractions are Shown.
Group(n)
Aqueoussoluble
Proteininterphase
Organic soluble
TotalPolar
Rf 5 0–0.1‘‘DG’’
Rf 5 0.2–0.3‘‘FA’’
Rf 5 0.35–0.5“TG”
Rc 5 0.79–0.9
Control (5) 0.018 6 0.004 0.255 6 0.011 0.047 6 0.009 0.027 6 0.011 0.004 6 0.002 0.005 6 0.005 0.006 6 0.005Etomoxir (3) 0.003 6 0.001* 0.027 6 0.006* 0.030 6 0.006* 0.014 6 0.006* 0.002 6 0.001 0.010 6 0.001* 0.002 6 0.002
* p , 0.05 versus control.
226 T. R. DeGrado et al.

observed over the 3 h imaging period, suggesting negligible meta-bolic defluorination of the omega-[18F]fluoro fatty acid in the swine.Left ventricular myocardial uptake of FTP was high (Fig. 7).Time-activity curves for myocardium, left ventricular blood pool,and lung regions are shown in Figure 8. Blood clearance was rapid:Myocardium-to-blood ratio exceeded 7:1 at 20 min postinjection.Myocardium-to-lung ratio was .8:1 at 20 min postinjection.Myocardial clearance half-time of 18F radioactivity was approxi-mately 5 h after 20 min postinjection, evidencing effective trappingof the radiolabel in tissue. Liver:heart ratios were 0.4–0.8. Clear-ance rate of radioactivity from liver (T1/2 approximately 50 min)was faster than that from heart.
DISCUSSION
Mitochondrial b-oxidation of long-chain fatty acids is an importantprocess in tissues of the body because it supplies major proportionsof the energy requirement in heart, liver, and kidney. Noninvasiveassessment of b-oxidation may be a useful diagnostic tool becauseimpairment of b-oxidation may be interpreted as a feature of diseaseprogression in patients with coronary artery disease (18, 26, 29, 36)and various inherited (12 , 23) and acquired (15, 33) cardiomyop-athies. Pharmacologic inhibition of fatty acid oxidation has beensuggested as a potential therapy for enhancing recovery of reper-
fused ischemic myocardium (20), whereas gene therapy is a poten-tial aid for patients with inherited deficiencies of fatty acidoxidation enzymes. Noninvasive assessment of fatty acid oxidationwith PET could provide critical information for developing therapyprotocols and monitoring the effects of therapy in individualpatients.
The advantages of an 18F-labeled fatty acid analog that isspecifically retained as a consequence of b-oxidation would include1) the favorable 110-min half-life of 18F for PET imaging, synthesisof multiple doses, and possibility for commercial production anddistribution; 2) excellent imaging characteristics of 18F (shortpositron range in tissue); 3) simplified, “metabolic-trapping” kinet-ics in tissue such that b-oxidation rate is indicated by traceraccumulation, allowing quantitative estimates of b-oxidation ratein analogy to the estimation of glucose phosphorylation rate with2-[18F]fluoro-2-deoxy-glucose (24); and 4) prolonged retention ofradioactivity in tissue that allows imaging by low temporal resolu-tion collimated single photon emission computed tomography orcoincidence-capable gamma cameras, as well as higher signal-to-noise in dedicated PET scanners. Importantly, quantitative assess-ment of b-oxidation rate in ischemic tissue is feasible. Although theadvantages of a metabolically trapped tracer are compelling, heavydemands are place on radiotracer design, characterization, and valida-tion to establish the utility of a proposed tracer. A metabolically
TABLE 5. Comparison of Estimated Fractional 18F-labeled Thia Fatty Acid Trapping Rates and Palmitate Oxidation Ratesin Langendorff Perfused Rat Hearts. All Values have Units ml/min/g Dry
Tracer
18F Tracer metabolictrapping rates
[FR 5 k1k3/(k2 1 k3)]Fractional palmitate
oxidation rate
Normoxic (n) Hypoxic (n) % Decrease Normoxic Hypoxic % decrease
FTP 1.45 6 0.39 (5) 0.73 6 0.16 (5) 50* 2.73 6 0.23 1.64 6 0.30 40*17F6THA 1.28 6 0.18 (4) 1.17 6 0.35 (5) 9 2.37 6 0.13 1.47 6 0.39 39*
* p , 0.01. Oxygenation of perfusate: normoxic, 95% O2; hypoxic, 35% O2.
FIG. 3. Representative time-ac-tivity curve with bolus injectionof 18F-labeled 4-thia palmitate inLangendorff perfused rat heart.Rapid (“reversible”) and slow(“trapped”) clearance phases arereadily observed. Table 4 givesparameter estimates from three-exponent fits to the curves.
18F-labeled 4-thia Palmitate in Heart 227
trapped tracer of b-oxidation should be retained in tissue only in theform of b-oxidation metabolites in a manner such that there is a close(and well-behaved) relationship between tracer trapping rate and fattyacid oxidation rates under a broad range of physiologic conditions.
The 6-thia fatty acid analog FTHA was developed as a metabol-ically trapped tracer of b-oxidation in myocardium (4, 7). 4-Thiafatty acids are b-oxidized in the mitochondria to 4-thia-enoyl-CoAs, which apparently accumulate in the mitochondrion becausethey are very poor substrates for CPT-II (32), yet bind to acyl-CoAdehydrogenase enzymes in the mitochondrion (17). 4-Thia-enoyl-CoA is very slowly converted to b-hydroxy-4-thia acyl-CoA by thenext step of the b-oxidation pathway (17). The resultant b-hy-droxy-4-thia acyl-CoA splits spontaneously to an alkyl-thiol and
malonic acid semialdehyde-CoA ester. 4-Thia fatty acids inhibitfatty acid oxidation and induce fatty liver in vivo (30). The inhibitionis presumably explained by accumulation of 4-thia-enoyl-CoAs in themitochondrion leading to decreased mitochondrial CoASH, as well asinactivation of long-chain acyl-CoA dehydrogenase by binding of theaccumulated 4-thia-enoyl-CoA to this enzyme (17). v-Oxidationand sulfur oxidation of thia fatty acids in liver give rise todicarboxylic sulfoxy acids, which are efficiently cleared by kidneysand excreted into the urine (14). Incorporation of 4-thia fatty acidsinto complex lipids of hepatocytes is relatively low (31).
Long-chain fatty acids labeled with 18F at or near the terminuscarbon are taken up and metabolically sequestered in heart and liverin a manner that is similar to that of their unlabeled counterparts
FIG. 4. Time-activity curves of18F-labeled 4-thia palmitate in rep-resentative isolated rat hearts innormoxic (95% oxygen [O2]) andhypoxic (35% O2) perfusion con-ditions. The tracer pulse-infusionperiod (0 < t < 20 min) is shown.Coronary flow was 7 mL/min inboth experiments. ADV, apparentdistribution volume.
FIG. 5. Fit of two-compartment modelto 18F-labeled 4-thia palmitate kineticsin normoxic Langendorff heart. Fittedparameter estimates are: blood vol-ume 5 0.78 mL/g dry; k1 5 5.01 mL/min/g dry; k2 5 0.56/min; k3 5 0.36/min; k4 5 0.0050/min. ADV, apparentdistribution volume.
228 T. R. DeGrado et al.

(16). Therefore, it was expected that a terminally 18F-labeledeven-substituted thia fatty acid would trace fatty acid uptake andundergo b-oxidation to a potentially trapped metabolite in themitochondrion. Placement of the 18F atom at the (v-3) carbon wasfound to minimize defluorination of the tracer in mice as a conse-quence of cytochrome P-450 mediated v-oxidation mainly in liver (6).
Evaluation studies have shown the myocardial accumulation ofFTHA to follow expected responses of b-oxidation in the followingconditions: 1) reduced with pharmacologic inhibition of CPT-I inmouse (7); 2) unchanged by dipyridamole- or adenosine-inducedhyperemia (8, 34); 3) increased by exercise in normal human volun-teers correlating with increases in heart rate-pressure product (8); 4)decreased by lactate in pig heart with parallel decreases in oxidation ofpalmitate (34); 5) decreased by ischemia in pig heart with paralleldecreases in oxidation of palmitate (35); 6) decreased in humans withcoronary artery disease (28); and 7) decreased with glucose/insulinclamp in normal human myocardium (21). However, accumulationof FTHA was not sensitive to inhibition of fatty acid oxidation withhypoxia in extracorporeally perfused swine myocardium (35).
Our present results with the corresponding v-labeled 6-thiaanalog 17F6THA in perfused rat hearts confirm the insensitivity of6-thia fatty acid kinetics to hypoxia in the myocardium, suggestingthe existence of a retention mechanism in the heart for 18F-labeled6-thia fatty acid analogs that is not dependent on fatty acidoxidation. The placement of the sulfur atom at the sixth position ofFTHA rather than the fourth was motivated by concerns thatinterference by the sulfur atom with initial metabolic and transportsteps would be lower with the larger distance between the sulfuratom and the carboxylic group. The present results with 4-thia fattyacid analogs show this not to be the case. Accumulation of the6-thia (17F6THA) and 4-thia (FTP) long-chain fatty acids wereequivalent in normoxic perfused rat hearts, suggesting that bothradiotracers were equally accepted by the fatty acid binding pro-teins, carriers, and enzymes involved in the transport and metabolictrapping of the tracers in the myocyte. However, both 4-thia and6-thia analogs were more slowly oxidized by isolated heart thannatural palmitate. Their fractional oxidation rate was approximately60% that of palmitate in normal isolated rat hearts. It remains to bedetermined which steps of transport or metabolism are slower forthe thia fatty acid analogs relative to natural fatty acids.
In isolated rat heart, reduction in fatty acid oxidation rate withhypoxia was closely tracked by reduction in the trapping rate of4-thia FTP. In contrast, the accumulation of the 6-thia analog17F6THA was unchanged in hypoxia, confirming observations inhypoxic swine myocardium with FTHA (35). Evidently, the con-
FIG. 6. Correlation of estimated fractional tracer metabolictrapping rates of 18F-labeled thia fatty acids and fractionalpalmitate oxidation rate in perfused rat heart. Squares, nor-moxic (95% oxygen [O2]); triangles, hypoxic (35% O2). FTP,18F-labeled 4-thia palmitate; 17F6THA, 17-[18F]fluoro-6-thia-heptadecanoic acid.
FIG. 7. Positron emission tomographyimage of mid-ventricular short-axis sliceof normal swine heart acquired 10–20min postinjection with 18F-labeled 4-thiapalmitate.
18F-labeled 4-thia Palmitate in Heart 229

dition of hypoxia exposes mechanism(s) of retention in myocytesfor the 6-thia fatty acids that the 4-thia fatty acids do not share. Thechemical nature of the retained fraction of radioactivity with17F6THA in hypoxic myocardium was not identified in this study,but previous investigations in living mouse hearts showed incorpo-ration of FTHA into polar lipids and, to a lesser extent, triglycerides(7). In addition, radiolabeled acyl intermediates of 17F6THA couldpossibly accumulate in hypoxic myocardium to a greater degreethan those of FTP, depending on rates of formation and hydrolysis.
The metabolic trapping rates of the long-chain analogs17F6THA and FTP (1.3–1.5 mL/min/g dry) in relation to coronaryflows of approximately 40 mL/min/g dry in normal conditions implynet (steady-state) extractions of approximately 3%. The discrep-ancy in this figure with the 8% “trapped” fraction in bolus injectionexperiments with FTP (Table 4) is likely due to a systematicunderestimation of the total dose by the peak count rate in thebolus technique. At physiologic flows of 1–2 mL/min/g wet, the netextraction of FTP in vivo should be in the 10–25% range. At theseextraction levels, the tracer uptake is not likely to be influencedsignificantly by changes in myocardial perfusion per se.
The slow clearance rate of FTP in the normal swine heart isconsistent with the avid myocardial retention of FTP in living ratsat 2 h postinjection (Table 2). Analytical studies in extracts ofhearts from living rats showed that protein-bound metabolitescomprise .80% of the radioactivity in the heart in normalconditions. The protein-bound radioactivity is .90% reduced inCPT-I-inhibited rat hearts. This result is consistent with decrease infatty acid oxidation rates with CPT-I inhibition (3) and suggeststhat the accumulation of protein-bound metabolites in the myocar-dium is dependent on transport and oxidation of FTP in themitochondrion. A putative end-metabolite of FTP in the mitochon-drion that could bind to proteins is the product of the first step ofb-oxidation, 16-[18F]fluoro-4-thia-hexadec-2-enoyl-CoA. The nextstep of b-oxidation (i.e., hydration of the 4-thia-enoyl-CoA ester)has been shown to be very slow with a nonfluorinated 4-thia fatty
acid (17). Further analytical work is necessary to demonstrate thatthe chemical form of “trapped” species of FTP is a product ofb-oxidation and to validate the putative two-compartment modelused to fit the kinetics of FTP in isolated rat hearts and estimatefatty acid oxidation rate over a broad range of conditions. Untilsuch work is done, it cannot be firmly concluded that the trappingof FTP in the myocardium strictly measures b-oxidation rate.
The lack of evidence of in vivo defluorination of FTP in the livingswine was a surprising finding in light of the evidence for defluori-nation of FTP in rodents (Table 2). Initial studies with FTP inhumans also show neglible bone uptake of radioactivity (to bepublished elsewhere), suggesting that the metabolic handling ofFTP in swine and humans is different than in the rat. A speciesdependence of v-oxidative defluorination of FTP is suggested. Rodentspecies are known to have particularly strong hepatic cytochromeP-450–dependent v-oxidative metabolism of fatty acids (6).
The high liver uptake of FTP is undesirable for myocardialimaging, especially near the inferior wall of the left ventricle.Nevertheless, the spatial resolution of current state of the art PETscanners and image reconstruction algorithms may allow measure-ment of myocardial radioactivity concentration with minimal spill-over problems from liver. Whether dietary status could be manipulatedto optimize the heart:liver ratio of FTP uptake remains to be seen.
CONCLUSIONS
Our results show that the 4-thia palmitate analog FTP has potentialas a metabolically trapped tracer of fatty acid oxidation in heart.FTP shares the excellent imaging characteristics as the previouslydeveloped 6-thia analog FTHA but may possess higher specificityfor indication of fatty acid oxidation. Further studies are needed todefine the mechanism of retention and understand the relationshipof FTP trapping rate in the myocardium to oxidation rate oflong-chain fatty acids in various conditions and disease states.
FIG. 8. Time-activity curves for 18F-labeled 4-thia palmitate in normalswine myocardium, lung, liver, and leftventricular blood pool.
230 T. R. DeGrado et al.

This work was performed with the support of National Institutes ofHealth Grant HL-54882. The authors thank Larry F. Whitesell andKhristen J. Carlson for their excellent technical support. Etomoxir wasgenerously provided by Dr. H. Wolf of Byk Gulden, Konstanz,Germany.
References1. Abendschein D. R., Fox K. A. A., Ambos H. D., Sobel B. E. and
Bergmann S. R. (1987) Metabolism of beta-methyl[1-11C]heptade-canoic acid in canine myocardium. Nucl. Med. Biol. 14, 579–585.
2. Bergmann S. R., Weinheimer C. J., Markham J. and Herrero P. (1996)Quantitation of myocardial fatty acid metabolism using PET. J. Nucl.Med. 37, 1723–1730.
3. Bielefeld D. R., Vary T. C. and Neely J. R. (1985) Inhibition ofcarnitine palmitoyl-CoA transferase activity and fatty acid oxidation bylactate and oxfenicine in cardiac muscle. J. Mol. Cell. Cardiol. 17,619–625.
4. DeGrado T. R. (1991) Synthesis of 14(R,S)-[18F]fluoro-6-thia-heptade-canoic acid (FTHA). J. Label. Comp. Radiopharm. 29, 989–995.
5. DeGrado T. R., Holden J. E., Ng C. K., Raffel D. M. and Gatley S. J.(1989) b-Methyl-15-p-iodophenylpentadecanoic acid metabolism andkinetics in the isolated rat heart. Eur. J. Nucl. Med. 15, 78–80.
6. DeGrado T. R. and Moka D. C. (1992) Non-b-oxidizable T-[18F]fluorolong-chain fatty acid analogs show cytochrome P-450 mediated deflu-orination: Implications for the design of PET tracers of myocardial fattyacid utilization. Nucl. Med. Biol. 19, 389–397.
7. DeGrado T. R., Stocklin G. and Coenen H. H. (1991) 14(R,S)-[18F]Fluoro-6-thia-heptadecanoic acid (FTHA): Evaluation in mouse ofa new in vivo probe of myocardial utilization of long-chain fatty acids.J. Nucl. Med. 32, 1888–1896.
8. Ebert A., Herzog H., Stoecklin G., Henrich M., DeGrado T. R., CoenenH. H. and Feinendegen L. E. (1994) Kinetics of 14(R,S)-[F-18]fluoro-6-thia-heptadecanoic acid (FTHA) in normal human heart at rest,during exercise and after dipyridamole. J. Nucl. Med. 35, 51–56.
9. Elmaleh D. R., Livni E., Levy S., Varnum D., Strauss H. W. andBrownell G. L. (1983) Comparison of 11C and 14C-labeled fatty acidsand their b-methyl analogs. Nucl. Med. Biol. 10, 181–187.
10. Fox K. A. A., Abendschein D. R., Ambos H. D., Sobel B. E. andBergmann S. R. (1985) Efflux of metabolized and nonmetabolized fattyacid from myocardium. Implications for quantifying myocardial metab-olism tomographically. Circ. Res. 57, 232–243.
11. Goldstein R. A., Klein M. S., Welch M. J. and Sobel B. E. (1980)External assessment of myocardial metabolism with C-11 palmitate invivo. J. Nucl. Med. 21, 342–438.
12. Hale D. E. and Bennett M. J. (1992) Fatty acid oxidation disorders: Anew class of metabolic diseases. J. Pediatr. 121, 1–11.
13. Huang S. C. and Phelps M. E. (1986) Principles of tracer kineticmodeling in positron emission tomography and autoradiography. InPositron Emission Tomography and Autoradiography. Principles and Appli-cations for the Brain and Heart (Edited by Phelps M. E., Mazziotta J. C.and Schelbert H. R.), pp. 287–346. Raven Press, New York.
14. Hvattum E., Skrede S., Bremer J. and Solbakken M. (1992) Themetabolism of tetradecylthiopropionic acid, a 4-thia stearic acid, in therat. In vivo and in vitro studies. Biochem. J. 286, 879–887.
15. Kelly D. P., Mendelsohn N. J., Sobel B. E. and Bergmann S. R. (1993)Detection of impaired myocardial fatty acid utilization by positron-emission tomography indicative of genetic deficiency of long-chainacyl-CoA dehydrogenase. Am. J. Cardiol. 71, 738–744.
16. Knust E. J., Kuperfernagel C. and Stocklin G. (1979) Long-chain F-18fatty acids for the study of regional metabolism in heart and liver;odd-even effects in mice. J. Nucl. Med. 20, 1170–1175.
17. Lau S. M., Brantley R. K. and Thorpe C. (1989) 4-Thia-trans-2-alkenoyl-CoA derivatives: Properties and enzymatic reactions. Biochem-istry 28, 8255–8262.
18. Liedtke A. J. (1981) Alterations of carbohydrate and lipid metabolismin the acutely ischemic heart. Prog. Cardiovasc. Dis. 23, 321–336.
19. Livni E., Elmaleh D. R., Levy S., Brownell G. L. and Strauss W. H.(1982) Beta-methyl[1-11C]heptadecanoic acid: A new myocardial met-abolic tracer for positron emission tomography. J. Nucl. Med. 23,169–175.
20. Lopaschuk G. D. (1997) Alterations in fatty acid oxidation duringreperfusion of the heart after myocardial ischemia. Am. J. Cardiol. 80,11A–16A.
21. Maki M. T., Haaparanta M., Nuutila P., Oikonen V., Luotolahti M.,Eskola O. and Knuuti J. M. (1998) Free fatty acid uptake in themyocardium and skeletal muscle using fluorine-18-fluoro-6-thia-hepta-decanoic acid. J. Nucl. Med. 39, 1320–1327.
22. Marquardt D. W. (1963) An algorithm for least-squares estimation ofnonlinear parameters. J. Soc. Ind. Appl. Math. 11, 431–441.
23. North K. N., Hoppel C. L., De Girolami U., Kozakewich H. P. andKorson M. S. (1995) Lethal neonatal deficiency of carnitine palmitoyl-transferase II associated with dysgenesis of the brain and kidneys.J. Pediatr. 127, 414–420.
24. Phelps M. E., Hoffman E. J., Selin C., Sokoloff L. and Kuhl D. E. (1979)Tomographic measurement of local cerebral metabolic rate in humanswith (F-18) 2-fluoro-2-deoxy-D-glucose: Validation of method. Ann.Neurol. 6, 371–388.
25. Saddik M. and Lopaschuk G. (1992) Myocardial triglyceride turnoverduring reperfusion of isolated rat hearts subjected to a transient periodof global ischemia. J. Biol. Chem. 267, 3825–3831.
26. Schon H. R., Schelbert H. R., Najafi A., Hansen H., Huang S. C.,Barrio J. and Phelps M. E. (1982) C-11 labeled palmitic acid for thenoninvasive evaluation of regional myocardial fatty acid metabolismwith positron-computed tomography. II. Kinetics of C-11 palmitic acidin acutely ischemic myocardium. Am. Heart J. 103, 532–547.
27. Schon H. R., Schelbert H. R., Robinson G., Najafi A., Huang S. C.,Hansen H., Barrio J., Kuhl D. E. and Phelps M. E. (1982) C-11 labeledpalmitic acid for the noninvasive evaluation of regional myocardial fattyacid metabolism with positron-computed tomography. I. Kinetics ofC-11 palmitic acid in normal myocardium. Am. Heart J. 103, 532–547.
28. Schultz G., Vom Dahl J., Kaiser H. J., Koch K. C., Sabri O., BanneitzI., Cremerius U. and Buell U. (1996) Imaging of b-oxidation by staticPET with 14(R,S)-[18F]-fluoro-6-thiaheptadecanoic acid (FTHA) inpatients with advanced coronary heart disease: A comparison with18FDG-PET and 99mTc-MIBI SPECT. Nucl. Med. Commun. 17, 1057–1064.
29. Schwaiger M., Fishbein M. C., Block M., Wijns W., Selin C., PhelpsM. E. and Schelbert H. R. (1987) Metabolic and ultrastructuralabnormalities during ischemia in canine myocardium: Noninvasiveassessment by positron emission tomography. J. Mol. Cell. Cardiol. 19,259–269.
30. Skorve J., Asiedu D., Solbakken M., Gjestdal J., Songstad J. and BergeR. K. (1992) Comparative effects of oxygen and sulfur-substituted fattyacids on serum lipids and mitochondrial and peroxisomal fatty acidoxidation in rat. Biochem. Pharmacol. 43, 815–822.
31. Skrede S., Sorensen H. N., Larsen L. N., Steineger H. H., Hovik K.,Spydevold O. S., Horn R. and Bremer J. (1997) Thia fatty acids,metabolism and metabolic effects. Biochim. Biophys. Acta 1344, 115–131.
32. Skrede S., Wu P. and Osmundsen H. (1995) Effects of tetradecylthio-propionic acid and tetradecylthioacrylic acid on rat liver lipid metabo-lism. Biochem. J. 305, 591–597.
33. Sochor H., Schelbert H. R., Schwaiger M., Henze E. and Phelps M. E.(1986) Studies of fatty acid metabolism with positron emission tomog-raphy in patients with cardiomyopathy. Eur. J. Nucl. Med. 12, S66–S69.
34. Stone C. K., Pooley R. A., DeGrado T. R., Renstrom B., Nickles R. J.,Nellis S. H., Liedtke A. J. and Holden J. E. (1998) Myocardial uptakeof the fatty acid analog 14-18F-fluoro-6-thia-heptadecanoic acid(FTHA) in comparison to b-oxidation rates by tritiated palmitate.J. Nucl. Med. 39, 1690–1696.
35. Stone C. K., Pooley R. A., DeGrado T. R., Renstrom B., Nickles R. J.,Nellis S. H., Liedtke A. J. and Holden J. E. (1998) Comparison of fattyacid tracers FTHA and BMIPP retention during myocardial ischemiaand hypoxia. J. Nucl. Med. 39, 1684–1689.
36. Tamaki N., Kawamoto M., Takahashi N., Yonekura Y., Magata Y.,Torizuka T., Nohara R., Kambara H. and Konishi J. (1993) Assessmentof myocardial fatty acid metabolism with positron emission tomographyat rest and during dobutamine infusion in patients with coronary arterydisease. Am. Heart J. 125, 702–710.
18F-labeled 4-thia Palmitate in Heart 231
![Synthesis of 1-[18F]fluorodeoxyglucose: an unexpected rearrangement in the reaction of 2-O-methanesulfonyl-β-D-mannopyranose with [18f] fluoride](https://img.dokumen.tips/doc/110x75/63455e7d38eecfb33a06872d/synthesis-of-1-18ffluorodeoxyglucose-an-unexpected-rearrangement-in-the-reaction.jpg)
![An improved total synthesis of PET HSV-tk gene reporter probe 9-(4-[18F]fluoro-3-hydroxymethylbutyl)guanine ([18F]FHBG)](https://img.dokumen.tips/doc/110x75/634ff0b00ca35926a7094d4a/an-improved-total-synthesis-of-pet-hsv-tk-gene-reporter-probe-9-4-18ffluoro-3-hydroxymethylbutylguanine.jpg)
![N3-Substituted thymidine analogues V: Synthesis and preliminary PET imaging of N3-[18F]fluoroethyl thymidine and N3-[18F]fluoropropyl thymidine](https://img.dokumen.tips/doc/110x75/63528e46cb577625920dc5ec/n3-substituted-thymidine-analogues-v-synthesis-and-preliminary-pet-imaging-of-n3-18ffluoroethyl.jpg)




![[18F]EF3 is not superior to [18F]FMISO for PET-based hypoxia evaluation as measured in a rat rhabdomyosarcoma tumour model](https://img.dokumen.tips/doc/110x75/63615fcd8838ef044c074d10/18fef3-is-not-superior-to-18ffmiso-for-pet-based-hypoxia-evaluation-as-measured.jpg)





![In vivo quantification of hypoxic and metabolic status of NSCLC tumors using [18F]HX4 and [18F]FDG-PET/CT imaging](https://img.dokumen.tips/doc/110x75/635cbd98a3fa66b45c0e0f69/in-vivo-quantification-of-hypoxic-and-metabolic-status-of-nsclc-tumors-using-18fhx4.jpg)



![Imaging of FTLD-tau Using [18F]AV-1451 - TSpace](https://img.dokumen.tips/doc/110x75/6322a95baa6c954bc707cac6/imaging-of-ftld-tau-using-18fav-1451-tspace.jpg)











