Embed Size (px)
Citation preview

www.elsevier.com/locate/ybbrc
Biochemical and Biophysical Research Communications 341 (2006) 837–846
BBRC
Subcellular localization and regulation of type-1Cand type-5 phosphodiesterases q
Susanna Dolci a, Alessia Belmonte a,b, Rocco Santone d, Mauro Giorgi d,Manuela Pellegrini a, Eleonora Carosa e, Emilio Piccione b, Andrea Lenzi c,
Emmanuele A. Jannini e,*
a Department of Public Health and Cellular Biology, 2nd University of Rome ‘‘Tor Vergata’’, Rome, Italyb Department of Surgery, 2nd University of Rome ‘‘Tor Vergata’’, Rome, Italy
c Department of Medical Pathophysiology, 1st University of Rome ‘‘La Sapienza’’, Rome, Italyd Department of Basic and Applied Biology, University of L’Aquila, L’Aquila, Italye Department of Experimental Medicine, University of L’Aquila, L’Aquila, Italy
Received 28 December 2005Available online 23 January 2006
Abstract
We investigated the subcellular localization of PDE5 in in vitro human myometrial cells. We demonstrated for the first time thatPDE5 is localized in discrete cytoplasmic foci and vesicular compartments corresponding to centrosomes. We also found that PDE5intracellular localization is not cell- or species-specific, as it is conserved in different animal and human cells. PDE5 protein levels arestrongly regulated by the mitotic activity of the smooth muscle cells (SMCs), as they were increased in quiescent, contractile myometrialcultures, and conditions in which proliferation was inhibited. In contrast, PDE1C levels decreased in all conditions that inhibited pro-liferation. This mirrored the enzymatic activity of both PDE5 and PDE1C. Increasing cGMP intracellular levels by dbcGMP or sildenafiltreatments did not block proliferation, while dbcAMP inhibited myometrial cell proliferation. Together, these results suggest that PDE5regulation of cGMP intracellular levels is not involved in the control of SMC cycle progression, but may represent one of the markers ofthe contractile phenotype.� 2006 Elsevier Inc. All rights reserved.
Keywords: Phosphodiesterases; Signal transduction; Uterus; Sildenafil
In the mature animal, smooth muscle cells (SMCs)have a quiescent, contractile phenotype. Although theirprincipal function is contraction, different stimuli canshift them from the quiescent to the proliferative pheno-type [1]. Proliferation and contraction are both stronglyregulated by the intracellular second messengers cyclicAMP and cyclic GMP, which are degraded by a super-family of enzymes known as phosphodiesterases (PDEs)[2]. These are currently classified into 11 families based
0006-291X/$ - see front matter � 2006 Elsevier Inc. All rights reserved.
doi:10.1016/j.bbrc.2006.01.035
q This work was partially supported by a Pfizer Italia unrestricted grantand by Italian Ministry of Research and Education ‘‘CIPE’’ and ‘‘PRIN’’grants.* Corresponding author. Fax: +39 0 862 433523.E-mail address: [email protected] (E.A. Jannini).
on their substrate and inhibitor profiles and structuralcharacteristics [2–4]. PDE families differ in their molecu-lar weight, charge, affinity to substrates and inhibitors,subcellular localization, and tissue distribution. PDE het-erogeneity is dependent on a large number of genes,probably related to specific functional roles for each sub-type in a given cellular context [2,3]. Of the 11 PDE genefamilies, type 1C, a calmodulin-stimulated PDE, hydro-lyzes both cAMP and cGMP at the same Km [5]. ThisPDE1 isoform is the main cAMP hydrolytic PDE inhuman proliferating SMCs in the presence of calcium.Type-5 PDE selectively hydrolyzes cGMP and its specificinhibition by sildenafil exerts a relaxant effect on corporacavernosa of both sexes [6,7], as well as on uterine vas-cular smooth muscle [8] and myometrium [9,10].

838 S. Dolci et al. / Biochemical and Biophysical Research Communications 341 (2006) 837–846
Although PDE5 was discovered and shown to be phys-iologically important in 1996 [11] through the serendipitousfinding of its inhibitor sildenafil, its distribution wasinferred indirectly, by biochemical evidence of specificactivity. The first tissue localization studies of the PDE5protein were performed only recently, demonstrating itsanatomical distribution in animal tissues [12], in the humanvagina [13], and in the male corpora cavernosa [14]. Thedistribution of PDE1C protein and activity is betterknown. However, while it is becoming clear that cAMP-hy-drolyzing PDE4D is compartmentalized within expressingcells [15], little is known about the subcellular localizationof cGMP-hydrolyzing PDEs.
As uterine contractility control plays a pivotal role inthe maintenance of pregnancy, we studied the subcellularlocalization and regulation of PDE5 and PDE1C levels inhuman myometrial cultures. Here we demonstrate thatthese enzymes are differentially distributed within SMCand differentially regulated by the proliferative phenotype.
Materials and methods
Materials. All reagents were of analytical grade and purchased fromSigma (Sigma-Aldrich, St. Louis, MI), unless otherwise stated. Sildenafilwas generously donated by Pfizer (Sandwich, Kent, UK). We used apolyclonal antibody, which specifically recognizes the enzyme amino-ter-minal domain of PDE5 [12]. To obtain the PDE5 antibody, a recombinantfusion protein was produced based on the homologous region of thehuman and bovine PDE5 sequence [16]. As previously demonstrated [12],Ab specificity was tested in preliminary experiments by incubation prior toWestern blot and immunofluorescence staining with the peptide used toraise the antibody (not shown). Anti-human PDE1C rabbit Ab was pur-chased from FabGennix (Frisco, TX).
Cell cultures. Myometrial tissues were obtained, following informedconsent and Ethics Committee approval, from nine pregnant women whounderwent cesarian section at term of gestation. All cesarean sections wereperformed under regional anesthesia before the onset of labor. Thereasons for cesarian section were: previous cesarian cut (n = 4), breechpresentation (n = 2), and cephalo-pelvic disproportion (n = 3). Themedian parity was 1 (0–2) and the mean age of patients was 34.2 (range29–37). Thin fragments of myometrium (2 · 2 mm) were cut with a sterileblade and digested for 30 min in 1 mg/ml collagenase in DMEM and thenfor 15 min in trypsin/EDTA. After digestion, the cell suspension wasrinsed in DMEM/10% fetal calf serum (FCS, Gibco-BRL, Milan, Italy)and plated in 6 cm dishes in DMEM/10% FCS. At confluence, cells weresplit three times 1:3 and cultures were then subjected to stimulationexperiments. At the third cell passage we also confirmed smooth muscleactin and desmin expression by immunofluorescence (not shown).Peripheral blood mononuclear cells (PBMCs) were also used for immu-nofluorescence studies. These were obtained from venous blood samples ofhealthy volunteer donors and isolated on a Ficoll-diatrizoate gradient.C2C12 myoblasts, cultured in DMEM 10% FCS, were also used, as well asprimary cultures from 20 dpn rat corpora cavernosa obtained as describedby Krall et al. [17] with slight modifications. Muscular actin stain dem-onstrated that these cultures were enriched at 90% of SMC. For serumstarvation experiments, cell monolayers were cultured for 24 h in DMEMalone. For MEK1 (extracellular signal-regulated kinase1) pathway inhi-bition, DMSO as control vehicle or the MEK-inhibitor U0126 (Promega,Milan, Italy) was added to the cell cultures to the indicated final con-centration. Cells were collected after 24 h incubation at 37 �C.
Immunofluorescence experiments. Cells were grown onto glass cover-slips and fixed for 10 min in cold methanol. Coverslips were then rinsed inPBS and incubated with anti-PDE5, anti-PDE1C, anti-centrin 3, anti-c-tubulin antibodies or mouse anti-A-kinase anchoring protein 450
(AKAP-450) antibody (the last three kindly provided by Dr. M. Bornens,Curie Institute, Paris, France), diluted 1:100 in PBS 0.5% BSA or 1:10(anti-AKAP-450), and left overnight at 4 �C in a humidified chamber.Coverslips were then washed three times in PBS and incubated with acyanin3 anti-rabbit antibody (1:400). Co-localization immunofluorescence(anti-PDE5 and anti-AKAP-450) was performed using a FITC-conjugatedanti-mouse antibody (1:400). Nuclei were stained with Hoechst 33349. Inthe control immunofluorescence for PDE5, the recombinant GST-fusionpeptide was used to pre-incubate anti-PDE5 antibodies, as previouslydescribed [26]. Slides were examined by epifluorescence (Nikon, Tokyo,Japan) or confocal microscopy (Sarastro 2000, Molecular Dynamics,Sunnyvale, CA).
Western blot. Proteins extracted from myometrial cells in RIPA buffer(50 mM Tris–HCl, pH 8.0; 0.15 M NaCl; 1 mM EDTA; 1% NP-40, 0.5%deoxycholate, and 0.1% SDS) were electrophoretically separated in an 8%SDS polyacrylamide gel and then transferred onto a polyvinylidenedifluoride membrane (Millipore, Bedford, MA). After blocking with a 5%skim milk solution in PBS with 0.1% Tween 20, the membrane wasincubated with the primary antiserum, diluted 1:500 in PBS-Tween, 5%BSA. Immunoreactivity was subsequently revealed using a luminol-basedreagent. The immunoreactive band was of the expected size, confirmingthe cross-reactivity of human PDE5 with our antibody, due to the highidentity (97%) between human and bovine protein sequences within theregion used for raising the antibodies [12,18]. Similar results were observedusing the rabbit anti-cGMP binding cGMP-specific PDE5 polyclonalantibody (Chemicon Int., Temecula, CA) (not shown), further confirmingthe specificity of our antibody.
Cell fractionation studies were performed by scraping cells in PBSsupplemented with the Protease Inhibitor Cocktail (Sigma P8340) andthen incubating for 20 min on ice in a hypotonic buffer (10 mMHepes, pH7.9, 10 mM KCl, 1.5 mM MgCl2 0.1 mM EGTA, and 0.5 mM DTT)supplemented with protease inhibitor cocktail, 10 mM b-glycerophos-phate, and 0.1 mM sodium vanadate. Nuclear pellets, obtained after 1000gcentrifugation at 4 �C for 10 min, were solubilized in the hypotonic buffercontaining 1% Triton X-100. Supernatants were supplemented with 1%Triton. Both nuclear and cytoplasmic fractions were then centrifuged at20,000g for 30 min at 4 �C. The supernatants from both samples wereconsidered Triton-soluble, while the pellets were further solubilized with aRIPA buffer and were considered as the Triton-insoluble fraction. Nuclearpellet purity was verified by microscopic observation and cellular actin andanti-histone H3 antibodies (06-755, Upstate Biotechnology) were used tofurther confirm the purity of the fractions. Anti-phospho Erk1/2 (mousemonoclonal, #sc-7383) and anti-Erk2 (rabbit polyclonal, #sc-154, SantaCruz Biotechnology, Milan, Italy) antibodies were used to verify the MEKpathway inhibition. LY294002 came from Alexis (Milan, Italy) (#270-038-M005), and anti-phospho-Akt and anti-Akt rabbit polyclonal [Phospho-Plus Akt (Ser-473) antibody kit, #9270] were from New England Biolabs.
[3H]thymidine incorporation. 6 · 103 cells were plated in microwelldishes. At each experimental point 0.5 lCi [3H]thymidine was added forthe last four hours of a 24 h culture. Cells were then harvested using amanual harvester (Pharmacia, Milan, Italy) and radiolabeled DNA wasquantified in a b counter.
PDE enzyme assays. Cells were homogenized in Tris–HCl 20 mMbuffer, pH 7.2, containing 0.2 mM EGTA and 5 mM b-mercaptoethanol,supplemented with protease inhibitor cocktail and 5 mM MgCl2 using aglass homogenizer (15 strokes, 4 �C). The homogenates were centrifugedat 14,000g for 30 min at 4 �C. Pellets were re-suspended in the homoge-nization buffer and centrifuged at 14,000g for 30 min (4 �C). The first andsecond supernatants were then pooled and used for PDE activity evalu-ation. PDE activity was measured with the two-step method described byThompson and Appelman [19]. All assays were performed at 30 �C using1 lM tritiated cGMP as substrate, in the presence of either 1 mM EGTA(to block PDE1C activity) or 0.5 mM CaCl2 (basal activity) and 4 lg/mlcalmodulin (CaM) (to stimulate PDE1C activity). PDE5 changes wereassessed by the specific inhibition with sildenafil (60 nM).
Statistical analysis. The Student t test and ANOVA have been used toassess the significance of proliferation studies ([3H]thymidine incorpora-tion and colorimetric assays) and PDE5 enzymatic activity setting
S. Dolci et al. / Biochemical and Biophysical Research Communications 341 (2006) 837–846 839
significance at P < 0.05. All experiments were performed at least in trip-licate for each tissue sample.
Results
Subcellular distribution of PDE5 and PDE1C in human
myometrial cells
As PDE1C and PDE5 are the most abundant cGMPPDEs in human uterine smooth muscle cells [20], we con-centrated on cultured human myometrial cells to study
A
B
C
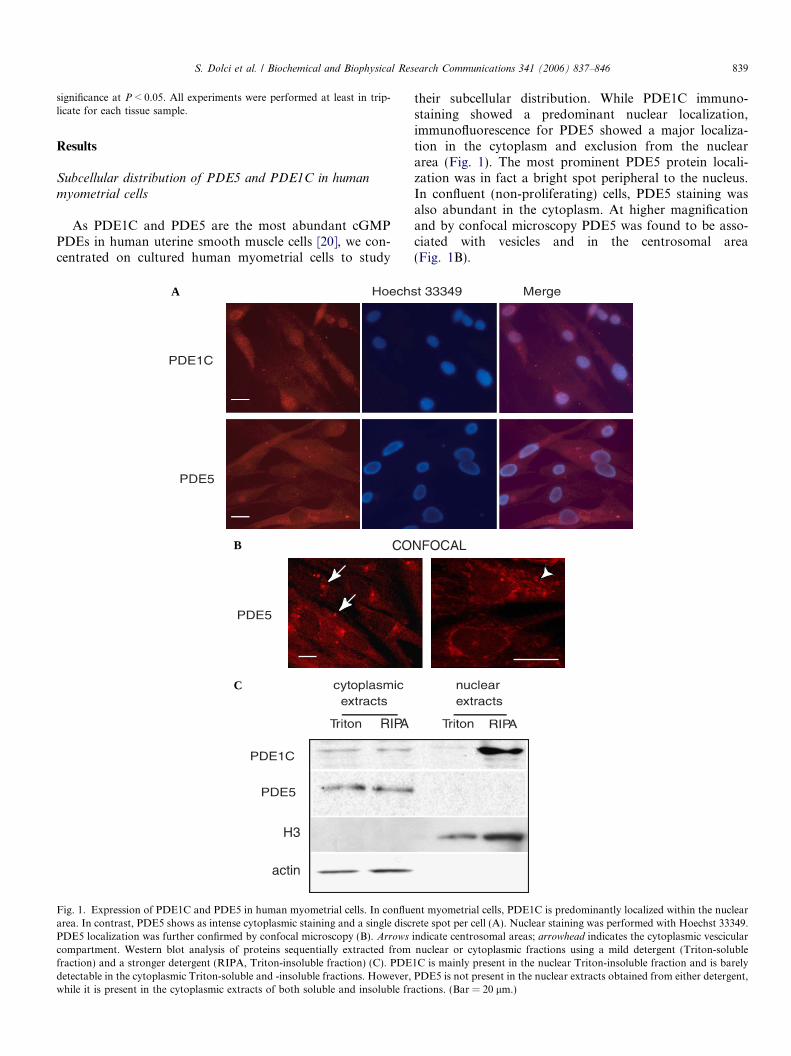
Fig. 1. Expression of PDE1C and PDE5 in human myometrial cells. In confluearea. In contrast, PDE5 shows as intense cytoplasmic staining and a single discPDE5 localization was further confirmed by confocal microscopy (B). Arrowscompartment. Western blot analysis of proteins sequentially extracted fromfraction) and a stronger detergent (RIPA, Triton-insoluble fraction) (C). PDEdetectable in the cytoplasmic Triton-soluble and -insoluble fractions. However,while it is present in the cytoplasmic extracts of both soluble and insoluble fra
their subcellular distribution. While PDE1C immuno-staining showed a predominant nuclear localization,immunofluorescence for PDE5 showed a major localiza-tion in the cytoplasm and exclusion from the nucleararea (Fig. 1). The most prominent PDE5 protein locali-zation was in fact a bright spot peripheral to the nucleus.In confluent (non-proliferating) cells, PDE5 staining wasalso abundant in the cytoplasm. At higher magnificationand by confocal microscopy PDE5 was found to be asso-ciated with vesicles and in the centrosomal area(Fig. 1B).
nt myometrial cells, PDE1C is predominantly localized within the nuclearrete spot per cell (A). Nuclear staining was performed with Hoechst 33349.indicate centrosomal areas; arrowhead indicates the cytoplasmic vescicularnuclear or cytoplasmic fractions using a mild detergent (Triton-soluble1C is mainly present in the nuclear Triton-insoluble fraction and is barelyPDE5 is not present in the nuclear extracts obtained from either detergent,ctions. (Bar = 20 lm.)

840 S. Dolci et al. / Biochemical and Biophysical Research Communications 341 (2006) 837–846
To confirm the immunofluorescence data, we obtainednuclear and cytoplasmic extracts from myometrial cell cul-tures using a subcellular fractionation procedure. Westernblot analysis of cytoplasmic and nuclear extracts showedthat PDE5 was present exclusively in the cytoplasmic frac-tion both in the Triton and RIPA extracts, while PDE1Cwas abundantly present in the nuclear extracts obtainedby RIPA buffer, but not in the Triton-extracted nuclearfraction. Low levels of PDE1C were present in cytoplasmicfractions extracted under both conditions (Fig. 1C). H3-histone and actin were probed to show nuclear and cyto-plasmic fractions, respectively (Fig. 1C).
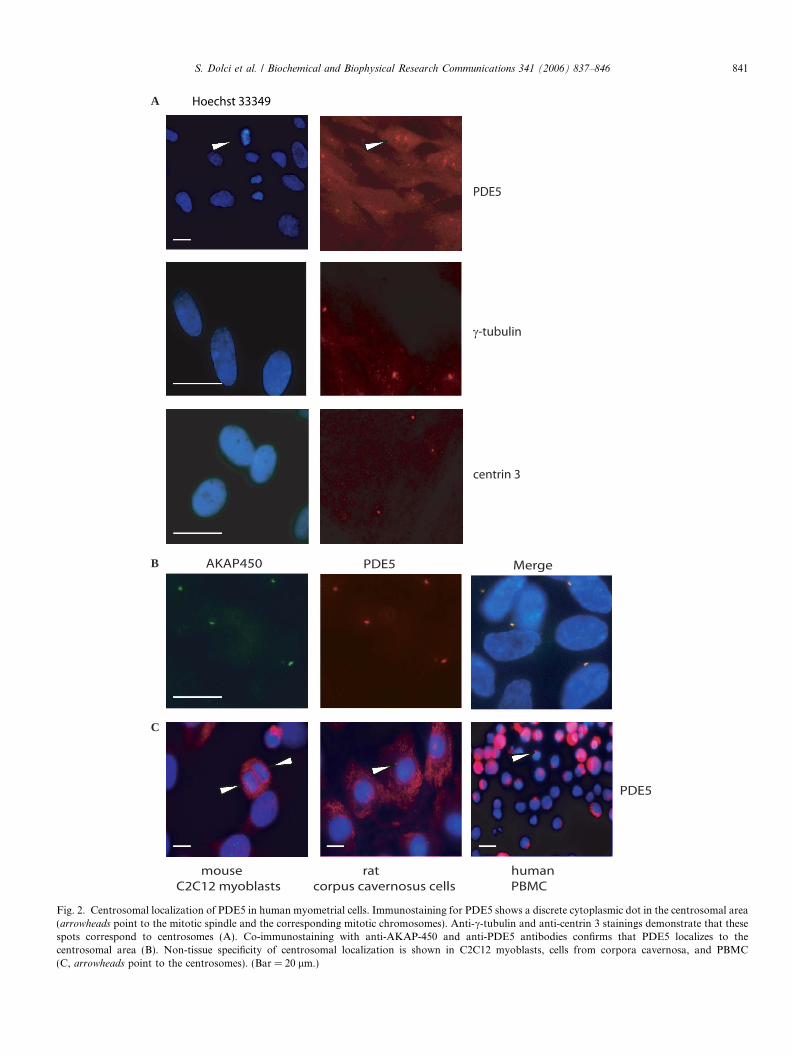
To verify if PDE5 was localized to centrosomes, we per-formed immuno-fluorescence microscopy on cultured myo-metrial cells using anti-c-tubulin and anti-centrin 3antibodies, which have been demonstrated to be specificmarkers for centrosomes [15,21,22]. As shown in Figs. 2Aand B the antibodies localized to centrosomes, as forPDE5. Since both antibodies were raised in rabbit as forPDE5, we could not co-label the same cells. However,co-localization experiments were performed using a mouseanti-AKAP-450 antibody, which has been shown to map tothe centrosomal area [15], and rabbit anti-PDE5 antibodyfurther confirming that PDE5 was localizing to the centro-somes (Fig. 2C). The specificity of localization was sup-ported by the evidence that centrosomal staining was nolonger observed when the anti-PDE5 antibody was pre-in-cubated with the recombinant PDE5 peptide used to raisethe antibody (not shown). The centrosomal localization isnot peculiar to smooth muscle cells, as it was conservedin mouse C2C12 myoblasts, in rat cells obtained from cor-pora cavernosa, and in human PBMCs (Fig. 2C).
Effect of cell confluence on PDE5 and PDE1C protein levels
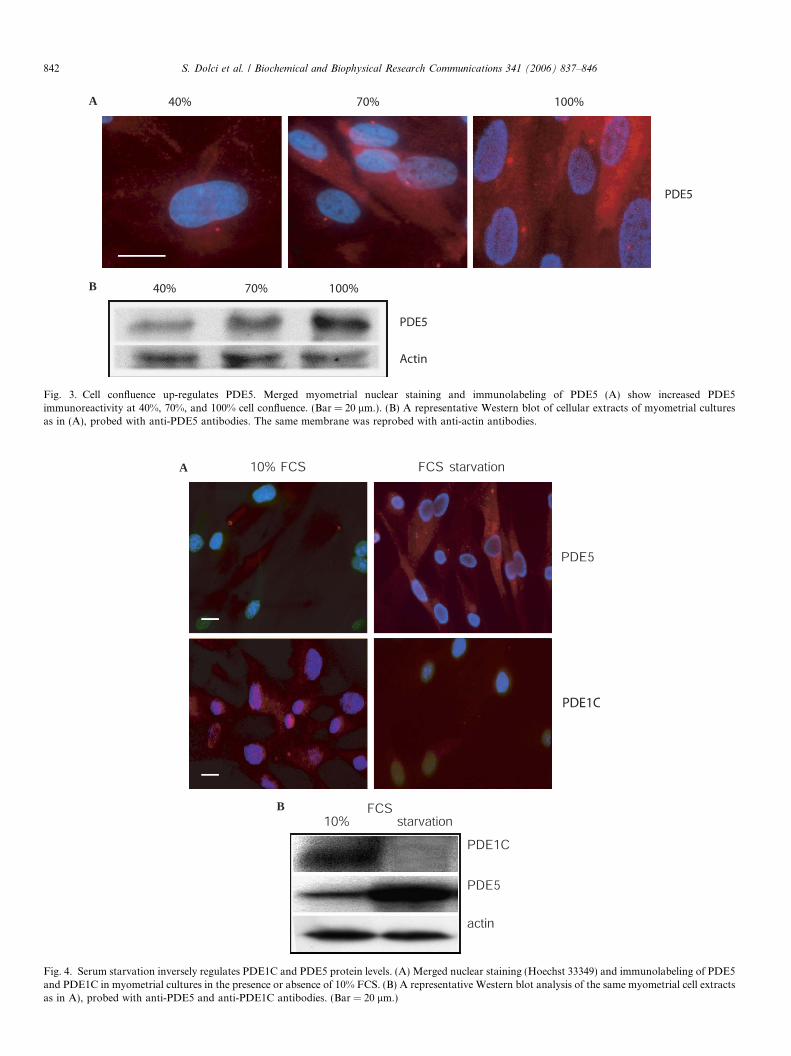
In the immuno-fluorescence experiments, we observedthat the intensity of PDE5 staining in the vesicle compart-ment, but not in the centrosomal area, was dependent onmyometrial cell confluence. Fig. 3A shows that when cellswere 40% confluent, PDE5 staining was fainter than for70% or 100% confluence. We therefore analyzed the pro-tein levels of cultures with the same cell confluences. Mir-roring the immuno-fluorescence experiments, PDE5protein levels sharply increased at 100% confluence(Fig. 3B).
To understand if PDE5 levels were regulated by growtharrest at confluence or by cell contacts occurring at conflu-ence, we plated non-confluent myometrial cells at 40% con-fluence and after 12 h starved part of the cultures fromFCS. The cultures were then incubated for 24 h and cellextracts were obtained. Figs. 4A and B show that serumstarvation up-regulated PDE5 levels even in non-confluentcultures and blocked cell proliferation (see Fig. 5C). Thisrules out the possibility that cell-to-cell contacts might beresponsible for PDE5 up-regulation.
As PDE1C has been shown to be up-regulated in prolif-erating aortic smooth muscle cells [5], we probed the same
filter for PDE1C. This was highly expressed in proliferatingcell cultures and down-regulated in serum starved cultures(Fig. 4B), as also observed in immunofluorescence experi-ments (Fig. 4A), confirming that PDE1C is specific forthe proliferative phenotype.
Blocking myometrial proliferation by MEK1 pathway
inhibition affects PDE5 and PDE1C levels
It has been shown that MEK inhibitors can blockPDGF-induced myometrial cell proliferation [23]. To blockserum-induced proliferation, myometrial cell cultures weregrown for 24 h in 10% FCS + 10 lM U0126, an MEKinhibitor. At the same time, parallel cultures were assayedfor [3H]thymidine incorporation (Fig. 5C). When prolifer-ating cell cultures were probed with anti-PDE5 antibodiesin the presence of FCS, faint levels of immunoreactivitywere observed compared to those of the U0126 treatment(FCS + 10 lM U0126). In contrast, PDE1C immunostain-ing revealed an intense nuclear localization in proliferatingcells, which sharply decreased in U0126-treated cells(Fig. 5A). To confirm these data, protein extracts fromtreated and untreated cells were then probed for PDE5and phospho-Erk1/2 (p-Erks) and Erk2 in Western blotanalysis (Fig. 5B). The U0126 addition increased PDE5levels over the control, as also occurred in the serumstarved cultures, and was able to block Erk1/2 phosphory-lation and cell proliferation (Fig. 5C). The same extractswere also probed for PDE1C, which has been shown tobe expressed in proliferating but not in quiescent smoothmuscle cells [24]. PDE1C was highly expressed in prolifer-ating myometrial cells and was dramatically down-regulat-ed by U0126 treatment.
To further evaluate the effect of proliferation on PDE1Cand PDE5 protein levels, we studied their enzymatic activ-ity in cells cultured in the presence or absence of FCS(Fig. 6). cGMP-PDE1C activity was the main enzymaticactivity present in proliferating myometrial cells (+FCS),as shown by the inducibility by CaM and the partial sensi-tivity to sildenafil. PDE5 activity was predominant in thestarving condition (�FCS), as demonstrated by theabsence of induction by CaM and the strong inhibitionby sildenafil.
High levels of cGMP do not affect myometrial cell
proliferation
Given that PDE5 levels were increased in quiescentmyometrial cells, we investigated if cGMP levels might reg-ulate myometrial proliferation. We performed [3H]thymi-dine incorporation in myometrial cells preincubated for24 h with dbcGMP and/or with sildenafil in the absenceor in presence of 10% FCS. One millimolar dbcAMP withor without sildenafil was also used as a control to inhibitproliferation [25]. While FCS induced a strong increase in[3H]thymidine incorporation over the serum-free control(Fig. 7), the presence of 1 mM dbcGMP or 60 nM sildenafil
A
B
C
Fig. 2. Centrosomal localization of PDE5 in human myometrial cells. Immunostaining for PDE5 shows a discrete cytoplasmic dot in the centrosomal area(arrowheads point to the mitotic spindle and the corresponding mitotic chromosomes). Anti-c-tubulin and anti-centrin 3 stainings demonstrate that thesespots correspond to centrosomes (A). Co-immunostaining with anti-AKAP-450 and anti-PDE5 antibodies confirms that PDE5 localizes to thecentrosomal area (B). Non-tissue specificity of centrosomal localization is shown in C2C12 myoblasts, cells from corpora cavernosa, and PBMC(C, arrowheads point to the centrosomes). (Bar = 20 lm.)
S. Dolci et al. / Biochemical and Biophysical Research Communications 341 (2006) 837–846 841
A
B
Fig. 3. Cell confluence up-regulates PDE5. Merged myometrial nuclear staining and immunolabeling of PDE5 (A) show increased PDE5immunoreactivity at 40%, 70%, and 100% cell confluence. (Bar = 20 lm.). (B) A representative Western blot of cellular extracts of myometrial culturesas in (A), probed with anti-PDE5 antibodies. The same membrane was reprobed with anti-actin antibodies.
A
B
Fig. 4. Serum starvation inversely regulates PDE1C and PDE5 protein levels. (A) Merged nuclear staining (Hoechst 33349) and immunolabeling of PDE5and PDE1C in myometrial cultures in the presence or absence of 10% FCS. (B) A representative Western blot analysis of the same myometrial cell extractsas in A), probed with anti-PDE5 and anti-PDE1C antibodies. (Bar = 20 lm.)
842 S. Dolci et al. / Biochemical and Biophysical Research Communications 341 (2006) 837–846
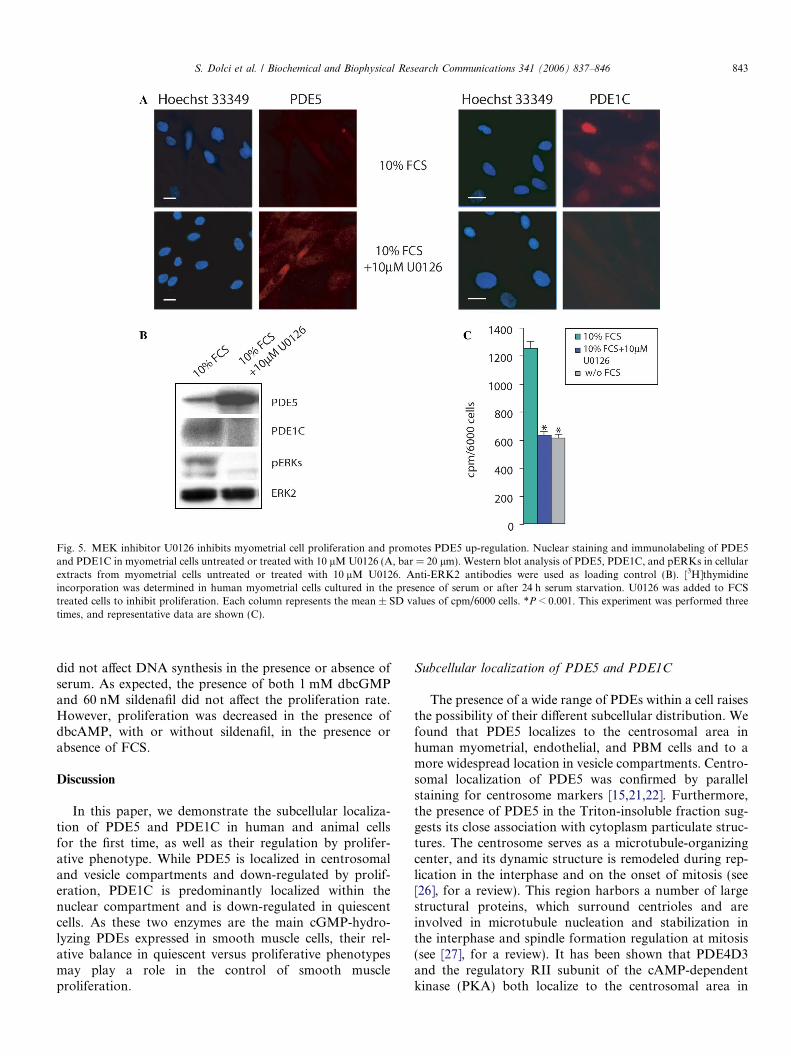
Fig. 5. MEK inhibitor U0126 inhibits myometrial cell proliferation and promotes PDE5 up-regulation. Nuclear staining and immunolabeling of PDE5and PDE1C in myometrial cells untreated or treated with 10 lMU0126 (A, bar = 20 lm). Western blot analysis of PDE5, PDE1C, and pERKs in cellularextracts from myometrial cells untreated or treated with 10 lM U0126. Anti-ERK2 antibodies were used as loading control (B). [3H]thymidineincorporation was determined in human myometrial cells cultured in the presence of serum or after 24 h serum starvation. U0126 was added to FCStreated cells to inhibit proliferation. Each column represents the mean ± SD values of cpm/6000 cells. *P < 0.001. This experiment was performed threetimes, and representative data are shown (C).
S. Dolci et al. / Biochemical and Biophysical Research Communications 341 (2006) 837–846 843
did not affect DNA synthesis in the presence or absence ofserum. As expected, the presence of both 1 mM dbcGMPand 60 nM sildenafil did not affect the proliferation rate.However, proliferation was decreased in the presence ofdbcAMP, with or without sildenafil, in the presence orabsence of FCS.
Discussion
In this paper, we demonstrate the subcellular localiza-tion of PDE5 and PDE1C in human and animal cellsfor the first time, as well as their regulation by prolifer-ative phenotype. While PDE5 is localized in centrosomaland vesicle compartments and down-regulated by prolif-eration, PDE1C is predominantly localized within thenuclear compartment and is down-regulated in quiescentcells. As these two enzymes are the main cGMP-hydro-lyzing PDEs expressed in smooth muscle cells, their rel-ative balance in quiescent versus proliferative phenotypesmay play a role in the control of smooth muscleproliferation.
Subcellular localization of PDE5 and PDE1C
The presence of a wide range of PDEs within a cell raisesthe possibility of their different subcellular distribution. Wefound that PDE5 localizes to the centrosomal area inhuman myometrial, endothelial, and PBM cells and to amore widespread location in vesicle compartments. Centro-somal localization of PDE5 was confirmed by parallelstaining for centrosome markers [15,21,22]. Furthermore,the presence of PDE5 in the Triton-insoluble fraction sug-gests its close association with cytoplasm particulate struc-tures. The centrosome serves as a microtubule-organizingcenter, and its dynamic structure is remodeled during rep-lication in the interphase and on the onset of mitosis (see[26], for a review). This region harbors a number of largestructural proteins, which surround centrioles and areinvolved in microtubule nucleation and stabilization inthe interphase and spindle formation regulation at mitosis(see [27], for a review). It has been shown that PDE4D3and the regulatory RII subunit of the cAMP-dependentkinase (PKA) both localize to the centrosomal area in
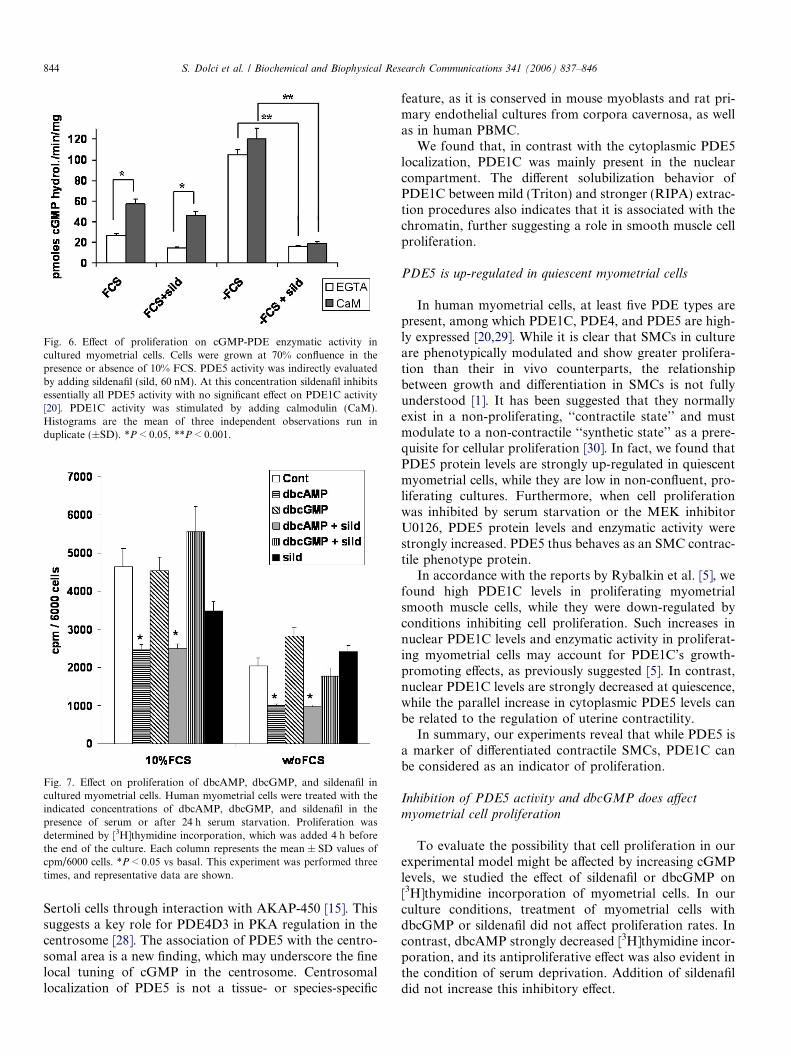
Fig. 6. Effect of proliferation on cGMP-PDE enzymatic activity incultured myometrial cells. Cells were grown at 70% confluence in thepresence or absence of 10% FCS. PDE5 activity was indirectly evaluatedby adding sildenafil (sild, 60 nM). At this concentration sildenafil inhibitsessentially all PDE5 activity with no significant effect on PDE1C activity[20]. PDE1C activity was stimulated by adding calmodulin (CaM).Histograms are the mean of three independent observations run induplicate (±SD). *P < 0.05, **P < 0.001.
Fig. 7. Effect on proliferation of dbcAMP, dbcGMP, and sildenafil incultured myometrial cells. Human myometrial cells were treated with theindicated concentrations of dbcAMP, dbcGMP, and sildenafil in thepresence of serum or after 24 h serum starvation. Proliferation wasdetermined by [3H]thymidine incorporation, which was added 4 h beforethe end of the culture. Each column represents the mean ± SD values ofcpm/6000 cells. *P < 0.05 vs basal. This experiment was performed threetimes, and representative data are shown.
844 S. Dolci et al. / Biochemical and Biophysical Research Communications 341 (2006) 837–846
Sertoli cells through interaction with AKAP-450 [15]. Thissuggests a key role for PDE4D3 in PKA regulation in thecentrosome [28]. The association of PDE5 with the centro-somal area is a new finding, which may underscore the finelocal tuning of cGMP in the centrosome. Centrosomallocalization of PDE5 is not a tissue- or species-specific
feature, as it is conserved in mouse myoblasts and rat pri-mary endothelial cultures from corpora cavernosa, as wellas in human PBMC.
We found that, in contrast with the cytoplasmic PDE5localization, PDE1C was mainly present in the nuclearcompartment. The different solubilization behavior ofPDE1C between mild (Triton) and stronger (RIPA) extrac-tion procedures also indicates that it is associated with thechromatin, further suggesting a role in smooth muscle cellproliferation.
PDE5 is up-regulated in quiescent myometrial cells
In human myometrial cells, at least five PDE types arepresent, among which PDE1C, PDE4, and PDE5 are high-ly expressed [20,29]. While it is clear that SMCs in cultureare phenotypically modulated and show greater prolifera-tion than their in vivo counterparts, the relationshipbetween growth and differentiation in SMCs is not fullyunderstood [1]. It has been suggested that they normallyexist in a non-proliferating, ‘‘contractile state’’ and mustmodulate to a non-contractile ‘‘synthetic state’’ as a prere-quisite for cellular proliferation [30]. In fact, we found thatPDE5 protein levels are strongly up-regulated in quiescentmyometrial cells, while they are low in non-confluent, pro-liferating cultures. Furthermore, when cell proliferationwas inhibited by serum starvation or the MEK inhibitorU0126, PDE5 protein levels and enzymatic activity werestrongly increased. PDE5 thus behaves as an SMC contrac-tile phenotype protein.
In accordance with the reports by Rybalkin et al. [5], wefound high PDE1C levels in proliferating myometrialsmooth muscle cells, while they were down-regulated byconditions inhibiting cell proliferation. Such increases innuclear PDE1C levels and enzymatic activity in proliferat-ing myometrial cells may account for PDE1C’s growth-promoting effects, as previously suggested [5]. In contrast,nuclear PDE1C levels are strongly decreased at quiescence,while the parallel increase in cytoplasmic PDE5 levels canbe related to the regulation of uterine contractility.
In summary, our experiments reveal that while PDE5 isa marker of differentiated contractile SMCs, PDE1C canbe considered as an indicator of proliferation.
Inhibition of PDE5 activity and dbcGMP does affect
myometrial cell proliferation
To evaluate the possibility that cell proliferation in ourexperimental model might be affected by increasing cGMPlevels, we studied the effect of sildenafil or dbcGMP on[3H]thymidine incorporation of myometrial cells. In ourculture conditions, treatment of myometrial cells withdbcGMP or sildenafil did not affect proliferation rates. Incontrast, dbcAMP strongly decreased [3H]thymidine incor-poration, and its antiproliferative effect was also evident inthe condition of serum deprivation. Addition of sildenafildid not increase this inhibitory effect.

S. Dolci et al. / Biochemical and Biophysical Research Communications 341 (2006) 837–846 845
Even if several reports have shown that increased cGMPlevels suppress SMC proliferation [31,32], the effect ofcGMP on SMC proliferation is a complex topic [33]. Ithas been shown that the antimitogenic effects of organicnitrates are potentiated by sildenafil, through inhibitionof PDE3 activity and subsequent elevation of cAMP [34].However, specific PDE3 inhibitors do not affect SMC pro-liferation rate [35,36], PDE3 is not the predominant PDEin the myometrial cells [20], and sildenafil and anotherPDE5 inhibitor, zaprinast, do not inhibit SMC DNA syn-thesis under conditions where cGMP synthesis is stimu-lated [24], in agreement with our findings. In fact, cGMPis not as powerful as cAMP in suppressing SMC prolifera-tion [25]. Furthermore, it has been shown that in primaryrat SMC, increased cGMP levels can amplify fibroblastgrowth factor- or epidermal growth factor-elicited cell pro-liferation [37,38], whereas these reagents inhibit prolifera-tion of subcultured SMC [39].
Taken together our results suggest that in proliferatingmyometrial cells, the nuclear localization of PDE1C con-trols cAMP accumulation therein to promote cell cycleprogression. In contrast, in the quiescent phenotype, thedistinct PDE5 localization and accumulation in the cyto-plasmic vesicular compartment may account for cGMPregulation during contraction.
The data presented here may also have direct relevanceto clinical studies involving the use of sildenafil as a toco-lytic in pre-term births [9] and preeclampsia [40] with orwithout treatment able to increase PDE5 [41].
Acknowledgments
We thank Drs. Pellegrino Rossi and Claudio Sette (2ndUniversity of Rome ‘‘Tor Vergata’’) for comments to themanuscript, Pfizer (New York, NY) for Sildenafil, Dr. M.Bornens (Curie Institute, Paris) for the anti-centrin,c-tubulin, and anti-AKAP-450 antibodies, and Ms.Marie-Helene Hayles for her help with the English. Thiswork is supported by an unrestricted grant by Pfizer Italia(Rome, Italy) and by the PRIN grants of the ItalianMinistry of University.
References
[1] G.K. Owens, Regulation of differentiation of vascular smooth musclecells, Physiol. Rev. 75 (1995) 487–517.
[2] J.A. Beavo, Cyclic nucleotide phosphodiesterases: functional impli-cations of multiple isoforms, Physiol. Rev. 75 (1995) 725–748.
[3] S.H. Soderling, J.A. Beavo, Regulation of cAMP and cGMPsignaling: new phosphodiesterases and new functions, Curr. Opin.Cell. Biol. 12 (2001) 174–179.
[4] L. Fawcett, R. Baxendale, P. Stacey, C. McGrouther, I. Harrow,S. Soderling, J. Hetman, J.A. Beavo, S.C. Phillips, Molecularcloning and characterization of a distinct human phosphodiester-ase gene family: PDE11A, Proc. Natl. Acad. Sci. USA 97 (2000)3702–3707.
[5] S.D. Rybalkin, K.E. Bornfeld, W.K. Sonnenburg, I.G. Rybalkina,K.S. Kwak, K. Hanson, E.G. Krebs, J.A. Beavo, Calmodulin-stimulated cyclic nucleotide phosphodiesterase (PDE1C) is induced in
human arterial smooth muscle cells of the synthetic, proliferativephenotype, J. Clin. Invest. 100 (1997) 2611–2621.
[6] I. Goldstein, T.F. Lue, H. Padma-Nathan, R.C. Rosen, W.D. Steers,P.A. Wicker, Oral sildenafil in the treatment of erectile dysfunction,N. Engl. J. Med. 338 (1998) 1397–1404.
[7] J.R. Berman, L.A. Berman, S.M. Toler, J. Gill, S. Haughies, Safetyand efficacy of sildenafil citrate for the treatment of female sexualarousal disorders: a double-blind, placebo controlled study, J. Urol.170 (2003) 2333–2338.
[8] G. Sher, J.D. Fish, Vaginal sildenafil (Viagra): a preliminary report ofa new method to improve uterine artery blood flow and endometrialdevelopment in patients undergoing IVF, Hum. Reprod. 15 (2000)806–809.
[9] A.M. Agha, R.A. Taha, Sildenafil inhibits agonist-evoked rat uterinecontractility: influence of guanylyl cyclase inhibition, Eur. J. Phar-macol. 428 (2001) 343–348.
[10] C.S. Buhimschi, R.E. Garfield, C.P. Weiner, I.A. Buhimschi, Thepresence and function of phosphodiesterase type 5 in the ratmyometrium, Am. J. Obstet. Gyncol. 190 (2004) 268–274.
[11] M. Boolell, M.J. Allen, S.A. Ballard, S. Gepi-Attee, G.J. Muirhead,A.M.Naylor, I.H. Osterloh, C. Gingell, Sildenafil: an orally active type5 cyclic GMP-specific phosphodiesterase inhibitor for the treatment ofpenile erectile dysfunction, Int. J. Impot. Res. 8 (1996) 47–52.
[12] D. Giordano, M.E. De Stefano, G. Citro, A. Modica, M. Giorgi,Expression of cGMP-binding c-GMP-phosphodiesterase (PDE5) inmouse tissues and cell lines using an antibody against the enzymeamino-terminal domain, Biochem. Biophys. Acta 1539 (2001) 16–27.
[13] G. d’Amati, C.R. di Gioia, M. Bologna, D. Giordano, M. Giorgi, S.Dolci, E.A. Jannini, Type 5 phosphodiesterase expression in thehuman vagina, Urology 60 (2002) 191–195.
[14] A. Morelli, S. Filippi, R. Mancina, M. Luconi, L. Vignozzi, M.Marini, C. Orlando, G.B. Vannelli, A. Aversa, A. Natali, G. Forti,M. Giorgi, E.A. Jannini, F. Ledda, M. Maggi, Androgens regulatephosphodiesterase type 5 expression and functional activity incorpora cavernosa, Endocrinology 145 (2004) 2253–2263.
[15] K.A. Tasken, P. Collas, W.A. Kemmner, O. Witczak, M. Conti, K.Tasken, Phosphodiesterase 4D and protein kinase A Type IIconstitute a signaling unit in the centrosomal area, J. Biol. Chem.276 (2001) 21999–22002.
[16] K. Loughney, T.R. Hill, V.A. Florio, L. Uher, G.J. Rosman, S.L.Wolda, B.A. Jones, M.L. Howard, L.M. McAllister-Lucas, W.K.Sonnenburg, S.H. Francis, J.D. Corbin, J.A. Beavo, K. Ferguson,Isolation and characterization of cDNAs encoding PDE5A, a humancGMP-binding cGMP-specific 3 0,5 0-cyclic nucleotide phosphodiester-ase, Gene 216 (1998) 139–147.
[17] J.F. Krall, M. Fittingoff, J. Rajfer, Characterization of cyclicnucleotide and inositol 1,4,5-trisphosphate-sensitive calcium-ex-change activity of smooth muscle cells cultured from the humancorpora cavernosa, Biol. Reprod. 39 (1988) 913–922.
[18] P. Stacey, S. Rulten, A. Dapling, S.C. Phillips, Molecular cloning andexpression of human cGMP-binding cGMP-specific phosphodiester-ase (PDE5), Biochem. Biophys. Res. Commun. 247 (1998) 249–254.
[19] W.J. Thomson, M.M. Appleman, Multiple cyclic nucleotide phos-phodiesterase activities from rat brain, Biochemistry 10 (1971)311–316.
[20] S.D. Rybalkin, I.G. Rybalkina, R. Feil, F. Hofman, J.A. Beavo,Regulation of cGMP-specific phosphodiesterase (PDE5) phosphory-lation in smooth muscle cells, J. Biol. Chem. 277 (2002) 3310–3317.
[21] E. Bailly, M. Doree, P. Nurse, M. Bornens, p34cdc2 is located in bothnucleus and cytoplasm; part is centrosomally associated at G2/M andenters vesicles at anaphase, EMBO J. 8 (1989) 3985–3995.
[22] T. Stearns, M. Kirschner, In vitro reconstitution of centrosomeassembly and function: the central role of gamma-tubulin, Cell 76(1994) 623–637.
[23] I. Boulven, B. Palmier, P. Robin, M. Vacher, S. Harbon, D. Leiber,Platelet-derived growth factor stimulates phospholipase C-gamma 1,extracellular signal-regulated kinase, and arachidonic acid releasein rat myometrial cells: contribution to cyclic 3 0,5 0-adenosine

846 S. Dolci et al. / Biochemical and Biophysical Research Communications 341 (2006) 837–846
monophosphate production and effect on cell proliferation, Biol.Reprod. 65 (2001) 496–506.
[24] S.D. Rybalkin, C. Yan, K.E. Bornfeldt, J.A. Beavo, Cyclic GMPphosphodiesterases and regulation of smooth muscle function, Circ.Res. 93 (2003) 280–291.
[25] S. Fukumoto, H. Koyama, M. Hosoi, K. Yamakawa, S. Tanaka, H.Morii, Y. Nishizawa, Distinct role of cAMP and cGMP in cell cyclecontrol of vascular smooth muscle cells: cGMP delays cell cycletransition to through suppression of cyclin D1 and cyclin-dependentkinase 4 activation, Circ. Res. 85 (1999) 985–991.
[26] T. Sterns, Centrosome duplication, a centriolar pas de deux, Cell 105(2001) 417–420.
[27] A.M. Tassin, M. Bornens, Centrosome structure and microtubulenucleation in animal cells, Biol. Cell 91 (1999) 343–354.
[28] D. Diviani, J.D. Scott, AKAP signaling complexes at the cytoskel-eton, J. Cell Sci. 114 (2001) 1431–1437.
[29] M.J. Leroy, C. Lugnier, J. Merezak, G. Tanguy, S. Olivier, A. Le Bec,F. Ferre, Isolation and characterization of the rolipram-sensitiveAMP-specific phosphodiesterase (type IV PDE) in human termmyometrium, Cell. Signal. 6 (1994) 405–412.
[30] J.H. Chamley-Campbell, G.R. Campbell, R. Ross, The smoothmuscle cell in culture, Physiol. Rev. 59 (1979) 1–6.
[31] U.C. Garg, A. Hassid, Nitric oxide-generating vasodilators and 8-bromo-cyclic guanosine monophosphate inhibit mitogenesis andproliferation of cultured rat vascular smooth muscle cells, J. Clin.Invest. 83 (1989) 1774–1777.
[32] J.W.Assender, K.M. Southgate,M.B. Hallett, A.C.Newby, Inhibitionof proliferation, but not of Ca2mobilization, by cyclic AMP andGMPin rabbit aortic smooth-muscle cells, Biochem. J. 288 (1992) 527–532.
[33] H. Koyama, K.E. Bornfeldt, S. Fukumoto, Y. Nishizawa, Molecularpathways of cyclic nucleotide-induced inhibition of arterial smoothmuscle cell proliferation, J. Cell. Physiol. 186 (2001) 1–10.
[34] M.T. Osinski, B.H. Rauch, K. Schror, Antimitogenic actions oforganic nitrates are potentiated by sildenafil and mediated viaactivation of protein kinase A, Mol. Pharmacol. 59 (2001) 1044–1050.
[35] X.L. Pan, E. Arauz, J.J. Krzanowski, D.F. Fitzpatrick, J.B. Polson,Sinergistic interactions between selective pharmacological inhibitorsof phosphodiesterase isozyme families PDE III and PDSE IV toattenuate proliferation of rat vascular smooth muscle cells, Biochem.Pharmacol. 48 (1994) 827–835.
[36] J.E. Souness, G.A. Hassall, D.P. Parrott, Inhibition of pig aorticsmooth muscle cell DNA synthesis by selective type III and type IVcyclic AMP phosphodiesterase inhibitors, Biochem. Pharmacol. 44(1992) 857–866.
[37] A. Hassid, H. Arabshahi, T. Bourcier, G.S. Dhaunsi, C. Matthews,Nitric oxide selectively amplifies FGF-2-induced mitogenesis inprimary rat aortic smooth muscle cells, Am. J. Physiol. 267 (1994)H1040–H1048.
[38] G.S. Dhaunsi, A. Hassid, Atrial and C-type natriuretic peptidesamplify growth factor activity in primary aortic smooth muscle cells,Cardiovasc. Res. 3 (1996) 37–47.
[39] G.S. Dhaunsi, C. Matthews, K. Kaur, A. Hassid, NO increasesprotein tyrosine phosphatase activity in smooth muscle cells:relationship to antimitogenesis, Am. J. Physiol. 272 (1997) H1342–H1349.
[40] J.W. Downing, R. Ramasubramanian, R..F. Johnson, B.H. Minzter,R.L. Paschall, H.W. Sundell, B. Engelhardt, R. Lewis, Hypothesis:selective phosphodiesterase-5 inhibition improves outcome in pre-eclampsia, Med. Hypotheses 63 (2004) 1057–1064.
[41] A. Belmonte, C. Ticconi, S. Dolci, M. Giorgi, A. Zicari, A. Lenzi,E.A. Jannini, E. Piccione, Regulation of phophodiesterase 5 expres-sion and activity in human pregnant and non-pregnant myometrialcells by human chorionic gonadotropin, J. Soc. Gynecol. Invest. 12(2005) 570–577.
















![Ir :irlrh i:i1r ( hi , 1c !iin, ilil| \iir\ rl1lg tIi]o. l:,, .i .ri1 1;:jr](https://img.dokumen.tips/doc/110x75/6327e299051fac18490e77c4/ir-irlrh-ii1r-hi-1c-iin-ilil-iir-rl1lg-tiio-l-i-ri1-1jr.jpg)












