Embed Size (px)
Citation preview

MOLECULAR AND DEVELOPMENTAL NEUROSCIENCE
Sonic hedgehog (Shh)-Gli signaling controls neuralprogenitor cell division in the developing tectum in zebrafish
Carmen G. Feijoo,1,2,* Maritza G. Onate,1,* Luis A. Milla1 and Veronica A. Palma1
1Center for Genomics of the Cell, Facultad de Ciencias, Universidad de Chile, Casilla 653, Santiago, Chile2Departamento de Ciencias, Facultad de Ciencias, Universidad Andres Bello, Santiago, Chile
Keywords: Gli, proliferation, Shh, tectum, zebrafish
Abstract
Despite considerable progress, the mechanisms that control neural progenitor differentiation and behavior, as well as their functionalintegration into adult neural circuitry, are far from being understood. Given the complexity of the mammalian brain, non-mammalianmodels provide an excellent model to study neurogenesis, including both the cellular composition of the neurogenic microenvi-ronment, and the factors required for precursor growth and maintenance. In particular, we chose to address the question of thecontrol of progenitor proliferation by Sonic hedgehog (Shh) using the zebrafish dorsal mesencephalon, known as the optic tectum(OT), as a model system. Here we show that either inhibiting pharmacologically or eliminating hedgehog (Hh) signaling by usingmutants that lack essential components of the Hh pathway reduces neural progenitor cell proliferation affecting neurogenesis in theOT. On the contrary, pharmacological gain-of-function experiments result in significant increase in proliferation. Importantly, Shh-dependent function controls neural progenitor cell behavior as sox2-positive cell populations were lost in the OT in the absence of Hhsignaling, as evidenced in slow-muscle-omitted (smu) mutants and with timed cyclopamine inhibition. Expressions of essentialcomponents of the Hh pathway reveal for the first time a late dorsal expression in the embryonic OT. Our observations argue stronglyfor a role of Shh in neural progenitor biology in the OT and provide comparative data to our current understanding of progenitor ⁄ stemcell mechanisms that place Shh as a key niche factor in the dorsal brain.
Introduction
During CNS development, undifferentiated progenitor cells proliferatefrom neural stem ⁄ precursor cells and gradually restrict their fatesaccording to environmental cues. Secreted and membrane-associatedmolecules convey the information between cells, with the secretedglycoprotein Sonic hedgehog (Shh) being one such signaling mole-cule. In contrast to its role in early neural patterning and differentiationof the ventral axis of the CNS, recent findings indicate that during bothlate development and adulthood Shh acts as a mitogen, modulatingneural progenitor proliferation in the dorsal brain (Dahmane et al.,2001; Machold et al., 2003; Palma & Ruiz i Altaba, 2004; Palmaet al., 2005; Ulloa & Briscoe, 2007).
The molecular mechanisms that govern compartmentalization andneuronal differentiation are phylogenetically conserved among avariety of vertebrate species (Puelles & Rubenstein, 1993). However,each vertebrate species has specific brain structures as a result ofadaptation to their environment. For example, mammals developed thesix-layered neocortex for processing numerous environmental stimuli,while a comparable degree of complexity arose in the fish and avianoptic tectum (OT), the dorsal derivative of the midbrain. Most neurons
in the mammalian brain are generated embryonically during restrictedphases, and the mature brain is characterized by a relatively constantnumber of neurons. In contrast, it is well known that the OT growscontinuously throughout the lifespan of teleosts. The cellular andmolecular mechanisms behind differential brain growth in fish are notyet known, but may be related to those mechanisms that would bedesirable to control in mammalian brains. Based on the finding thatShh regulates the behavior of cells with stem cell properties in thedeveloping and adult mammalian dorsal brain, we therefore set out toinvestigate its possible involvement in the control of stem cell lineagesin the zebrafish OT.Cellular responses to hedgehog (Hh) pathway activation are initiated
by the Shh ligand acting through the Patched 1 (Ptc1) and Smoothened(Smo) membrane proteins. In the absence of Shh, Ptc1 maintains Smoin an inactive state, thus silencing intracellular signaling. With thebinding of Shh, Ptc1 inhibition of Smo is released and the signal istransduced (Lum & Beachy, 2004). The transcriptional response to Shhsignaling is mediated by a family of zinc-finger transcription factors:Gli1, Gli2 and Gli3 in vertebrates (Kasper et al., 2006). In zebrafish,Gli1 is the major activator of Hh target genes, and Gli2, similarly toGli3, plays both activator and repressor roles in different regions of theembryo (Karlstrom et al., 2003; Tyurina et al., 2005).Our study took advantage of well-defined mutant loss-of-function
mutations in zebrafish, including slow-muscle-omitted (smu) thataffects smo (Barresi et al., 2000; Varga et al., 2001), detour (dtr) that
Correspondence: Dr V. A. Palma, as above.E-mail: [email protected]
*C.G.F. and M.G.O. contributed equally to this work.
Received 30 December 2009, revised 28 September 2010, accepted 11 November 2010
European Journal of Neuroscience, Vol. 33, pp. 589–598, 2011 doi:10.1111/j.1460-9568.2010.07560.x
ª 2011 The Authors. European Journal of Neuroscience ª 2011 Federation of European Neuroscience Societies and Blackwell Publishing Ltd
European Journal of Neuroscience

affects gli1, and you-too (yot) a potent dominant repressor form (DR)of gli2 (Karlstrom et al., 2003). Importantly, zebrafish carrying nullmutations for a number of members of the Hh pathway survive longenough to undertake an analysis at late embryogenesis. Here wedemonstrate that blockage of Shh signaling in vivo results indiminished proliferation of OT progenitors and decreased neurogen-esis. Pharmacological loss-of-function and gain-of-function experi-ments identify a specific time window in which neural progenitors areresponsive to Hh signaling. Importantly, we demonstrate for the firsttime that Shh and its downstream effectors are expressed in the OT inlate embryogenesis and postembryonic stages. Taken together, ourdata demonstrate a critical role of Shh signaling in the regulation ofembryonic and post-embryonic neural progenitor cell lineages in theOT of the zebrafish brain.
Materials and methods
Zebrafish strains and maintenance
Wild-type (WT) fish were raised in our facility according to standardprotocols (Westerfield, 1993). Embryos were grown at 28 �C in E3medium (in mm: NaCl, 5; KCl, 0.17; CaCl2, 0.33; MgSO4, 0.3; 0.1%Methylene Blue) and, when necessary, were anesthetized and fixed in4% paraformaldehyde overnight at 4 �C. Developmental time pointsare expressed as hours post-fertilization (hpf). The mutant lines dtr, yot(Karlstrom et al., 1996) and smu (Barresi et al., 2000; Varga et al.,2001) have been described previously. The transgenic zebrafish lineexpressing green fluorescent protein (GFP) under control of regulatorysequences of the zebrafish POMC gene (PomC::gfp) was described byDr Melmed (Liu et al., 2003).All animal procedures were in accordance with the Chilean
legislation, and were approved by Institutional Animal Care and UseCommittees.
Pharmacological treatments
Zebrafish embryos were treated with 10 lm cyclopamine (InfinityPharmaceuticals, Cambridge, MA, USA) by adding 1 lL of a 10 mm
stock solution (in 95% EtOH) to 1 mL of E3 medium at defined timepoints. Control embryos were treated simultaneously with an equalvolume (1 lL) of 95% EtOH (cyclopamine carrier). For purmorph-amine (Calbiochem, San Diego, CA, USA) treatments, embryos weregrown in E3 medium with 10 lm of the Hh agonist at indicated timepoints and control embryos were treated with 1% dimethylsulfoxide(DMSO) in E3 medium. Treatments were carried out in six-well platesin 2 mL E3 medium (30 embryos ⁄ well) at 28.5 �C. Embryos werefixed with 4% paraformaldehyde, dehydrated in MeOH, and processedfor in situ hybridization and ⁄ or antibody labeling.
Whole-mount in situ hybridization and immunohistochemistry
In situ hybridization was done as previously described (Jowett &Lettice, 1994). The following genes were obtained as cDNA clonesand were used as templates for making RNA probes: sox2 (Hernandezet al., 2007), ptc1 (Concordet et al., 1996), fgf8 (Furthauer et al.,1997), wnt1 (Lun & Brand, 1998), shh (Krauss et al., 1993), pax7(Hammond et al., 2007), neuroD (Korzh et al., 1998), otx2 (van deWater et al., 2001), and gli1 and gli2 (Karlstrom et al., 2003). Forimmunohistochemistry, the following antibodies were used: mouseZn-12 (1 : 200; Trevarrow et al., 1990); mouse Zrf-1 ⁄ glial fibrillaryacidic protein (GFAP; 1 : 100; Trevarrow et al., 1990; Bernardos &
Raymond, 2006); rabbit anti-phospho-histone H3 (H3-P; 1 : 500;Upstate 07-424, Billerica, MA, USA). The secondary antibodies usedwere anti-mouse peroxidase (1 : 1000) and anti-rabbit peroxidase(1 : 300; Jackson ImmunoResearch, West Grove, PA, USA) coupledto either Cy5 (1 : 250; Vector Labs, Burlingame, CA, USA) or Alexa488 (1 : 500; Molecular Probes, Carlsbad, CA, USA). Immunohisto-chemistry labeling was carried out essentially as described in Sarrazinet al. (2006). For double-staining with sox2, gli2 or otx2 probes andthe antibody H3-P, an in situ hybridization ⁄ immunohistochemistrycombined protocol was developed. For immunofluorescence, anti-GFPrabbit 1 : 500 and anti-rabbit-FITC 1 : 1000 (Molecular Probes) wereused. DAPI nuclear staining identified the total number of cells.Whole-mount embryos were cleared in 75% glycerol.
Acridine orange staining
For cell death characterization, embryos were stained according toWilliams et al. (2000). Briefly, embryos were incubated for 20 min in5 lg ⁄ mL Acridine orange (Sigma, St Louis, MO, USA) in E3medium, washed three times for 5 min, and observed under fluores-cence microscopy. A positive control consisting of incubation in10 lm CuSO4 was included (Hernandez et al., 2007).
Sectioning
For ultramicrotome sections, representative whole-mount immuno-stained embryos were fixed in 3% glutaraldehyde ⁄ 0.1 m sodiumcocadilate buffer pH 7.2 and postfixed in 1% osmium tetroxidepreceded by dehydration in a graded acetone series. The samples wereembedded in epon-acetone 1 ⁄ 1 followed by epon 100%, andpolymerization was made at 60 �C. Semi-thin sections (2 lm) wereobtained in a Sorvall MT-5000 ultramicrotome and stained with 1%toluidine. For microtome sections of whole-mount stained embryosprocessed for in situ hybridization, samples were dehydrated in 30%sucrose, then embedded in gelatin solution (5% sucrose, 6% type-Agelatin) and fixed in 4% paraformaldehyde. Sections of 20 lm wereobtained in a sliding microtome (Leica SM2010R). For cryostatsections, representative double-stained embryos for in situ hybridiza-tion ⁄ immunohistochemistry were dehydrated in 30% sucrose, embed-ded in OCT and stored overnight at )80 �C. Sections of 10 lm wereobtained in a Microm HM 525 cryostat.
Imaging and statistics
Photographs were either taken with a Leica DFC 300FX camera usinga Leica MZ12 dissecting microscope, a Confocal microscope ZeissLaser Meta (whole-mount) or in a Nikon Optishot II microscope withQ Imaging Micropublisher 3.3 RTV (sections). Images were processedwith Photoshop 7.0 for Macintosh and Image J (NIH). In the cell-counting experiments, 15 mid-sagittal sections of a minimum of eightdifferent individuals for each pharmacological treatment were counted.Results are presented as mean ± standard error; significance wasascertained by performing one tailed-Student’s t-test of each drugtreatment to control situation using Graphpad Prism 5 software, andsignificance was set at P < 0.05.
Results
The Shh signaling pathway regulates proliferation of OT neuralprogenitor cells during a critical developmental window
To address the question of whether Shh signaling modulates neuralproliferation in the OT in zebrafish, we treated WT embryos with the
590 C. G. Feijoo et al.
ª 2011 The Authors. European Journal of Neuroscience ª 2011 Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 33, 589–598

alkaloid cyclopamine to entirely block Hh signaling at the level of theSmo receptor (Taipale et al., 2000). We assayed proliferation by usingthe M phase mitotic marker anti-H3-P antibody that labels OTproliferation zones, as already described in medaka and zebrafish(Wullimann & Knipp, 2000; Nguyen et al., 2001a,b; Mueller &Wullimann, 2003). During the first week of development, the teleostOT is not differentiated into the characteristic adult layers. Consistentwith the terminology agreed upon by the Boulder Committee(Schmatolla, 1972), it consists of a gray-matter zone occupying theinner (ventricular) half of each tectal lobe – equivalent to theventricular and subventricular zones; a white-matter outer zone –equivalent to the intermediate and marginal zones; and a scattering of(marginal) cells, relatively few in number in the white matter, thoughtto have migrated from the subventricular zone.
Because by 30 hpf the body plan has been established and Shh mayfunction in roles other than patterning of the embryonic axes, weincubated embryos in cyclopamine from this developmental time pointonward at different development stages (30, 48, 60 and 72 hpf), fixingall embryos at 96 hpf for H3-P expression analysis (Fig. 1B–F). From30 hpf on, a clearly defined proliferation zone is observed in the OT,manifested as a ‘T’ shape that covers the tectal midline and themidbrain–hindbrain boundary (MHB). When treatment was initiatedat 30 and 48 hpf, cyclopamine completely blocked proliferation.Treatment begun at 60 hpf resulted in decreased proliferation, but anumber of cells retained H3-P-positive labeling (Fig. 1D), whilestarting at 72 hpf had no apparent effect on OT proliferation. Tospecify the time point in which Shh acts on OT proliferation, we nextrepeated the experiment narrowing the time frame of exposure bystarting at 30, 36, 48, 53 hpf, but fixing at 60 hpf (Fig. 1H–L).Overall, the T-shape proliferation profile in the marginal zone did notshow substantial difference when the treatment was started at 53 hpf(Fig. 1K), indicating that Shh is required prior to this developmental
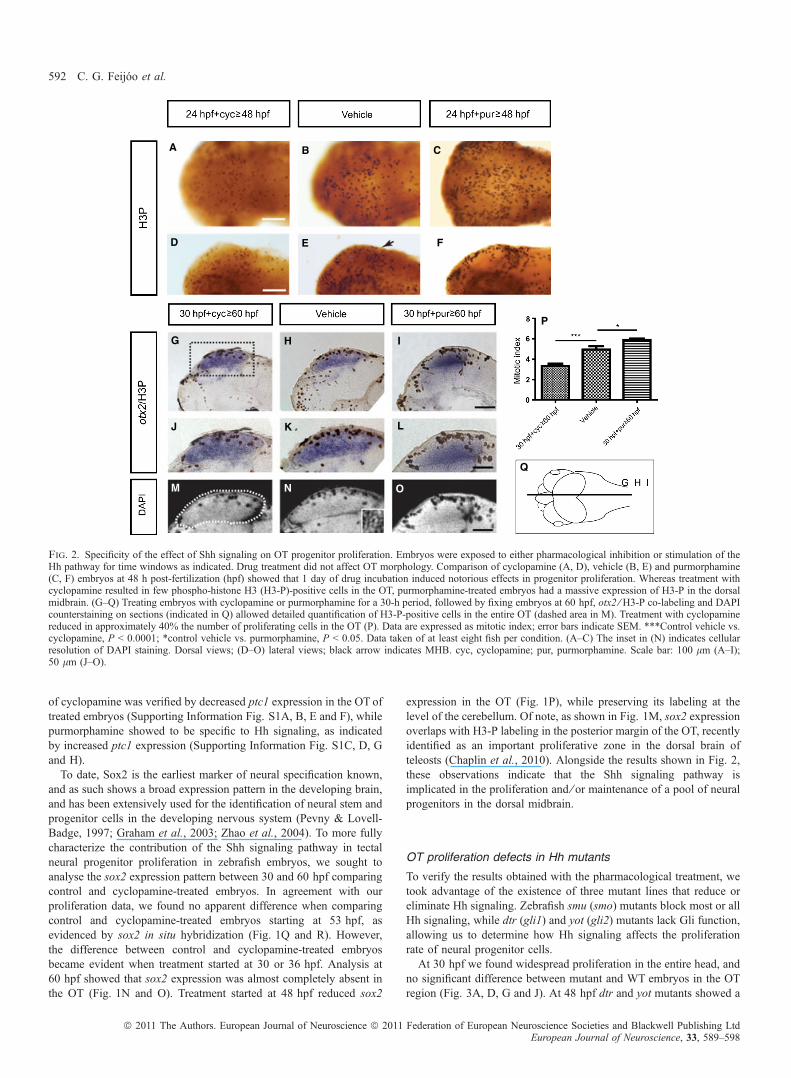
stage. Finally, we focused our examination of the proliferative markerin the OT after performing the inhibitor treatment starting at 24 hpfand fixing at 48 hpf, revealing a clear reduction in proliferation(Fig. 2A and D) in comparison to control siblings (Fig. 2B and E). Tofurther test the requirements of Shh signaling we performed atreatment with the small molecule agonist of Hh signaling purmorph-amine (Briscoe, 2006; Aanstad et al., 2009) and obtained the oppositeresult – an increase in the number of proliferative cells in the OT(Fig. 2C and F). Together, the time-lapse treatments suggest thatproliferating cells in the OT were sensitive to Shh signaling during aprecise time window.In order to quantify positive H3-P cells in the OT, we next treated
embryos with either cyclopamine or purmorphamine for a 30-h periodfixing embryos at 60 hpf. To provide detailed spatial information, weused the expression of otx2, reported as a specific OT marker, incombination with H3-P labeling (Fig. 2G–L). After counterstainingwith DAPI (Fig. 2M–O), we established the mitotic index (Fig. 2P).We found that H3-P-labeled cells were significantly reduced afterinhibitor treatment (mean ± standard error: control ⁄ vehi-cle = 4.951 ± 0.330, n = 20 sections; cyclopamine = 3.351 ± 0.211,n = 15 sections; P < 0.0001 by Student’s t-test; see Fig. 2K).Reinforcing a possible role for Shh signaling in OT growth, ourtreatment with purmorphamine resulted in an increase in proliferation(purmorphamine = 5.896 ± 0.1384, n = 15 sections; P < 0.05 byStudent’s t-test; see Fig. 2K). Similar results were obtained afterH3-P immunofluorescence labeling of treated embryos followed byoptical sectioning (Supporting Information Fig. S2A–D). To furtherconfirm dorsal location of positive H3-P cells we performed animmunostaining on PomC::gfp transgenic larvae. Taking advantage ofthe already described ventral expression of this pituitary anlage marker(Liu et al., 2003), we verified that indeed proliferating cells are mostlylocated dorsally (Supporting Information Fig. S2E and F). Effectiveness
A
B C D E F
LKJIH
G
M
N O P Q R
Fig. 1. Specific temporal inhibition of Shh signaling with cyclopamine results in reduced OT neural progenitor proliferation. Control phospho-histone H3 (H3-P)-positive labeling at 96 h post-fertilization (hpf; A) and 60 hpf (G) zebrafish embryos. The tectal proliferation region is demarcated with a dashed line. The blackrectangle indicates optical zooms shown from (B) to (L). In the first row (B–F) the effects of blocking the Shh signaling pathway between 30 and 96 hpf in tectalproliferation are shown. Treatments starting at 30 hpf (B) and 48 hpf (C) resulted in a complete loss of tectal precursor proliferation. (D) Embryo treatments started at60 hpf show decreased proliferation, while starting at 72 hpf (E) had no apparent effect on OT proliferation in comparison to the control (F). In the second row(H–L), a more restricted analysis was performed in order to accurately define the time point when Shh controls tectal proliferation. (M–R) Analysis of the expressionpattern of the neural progenitor marker sox2 after cyclopamine treatment at time points as indicated (M). Representative sagittal section of a 60 hpf embryo stainedfor both sox2 and H3P markers. The inset shows a cartoon of a lateral view of a zebrafish embryo illustrating the analysed OT region (dashed line) and expected sox2expression in black. (N–R) Whole-mount in situ hybridization of sox2 reveals a correlation between reduced proliferation and absence of a subpopulation of sox2 (+)neural progenitors in the OT after cyclopamine treatment. The white bracket shows progressive recovery of sox2 expression in the periventricular zone after delayedcyclopamine treatment. (A–L) Dorsal views and anterior up; (M–R) lateral views; black arrow points to MHB. cyc, cyclopamine; D, diencephalon; OT, optic tectum;T, telencephalon; tg, tegmentum. Scale bar: 50 lm.
Mitogenic Shh signaling in the developing tectum in zebrafish 591
ª 2011 The Authors. European Journal of Neuroscience ª 2011 Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 33, 589–598
of cyclopamine was verified by decreased ptc1 expression in the OT oftreated embryos (Supporting Information Fig. S1A, B, E and F), whilepurmorphamine showed to be specific to Hh signaling, as indicatedby increased ptc1 expression (Supporting Information Fig. S1C, D, Gand H).To date, Sox2 is the earliest marker of neural specification known,
and as such shows a broad expression pattern in the developing brain,and has been extensively used for the identification of neural stem andprogenitor cells in the developing nervous system (Pevny & Lovell-Badge, 1997; Graham et al., 2003; Zhao et al., 2004). To more fullycharacterize the contribution of the Shh signaling pathway in tectalneural progenitor proliferation in zebrafish embryos, we sought toanalyse the sox2 expression pattern between 30 and 60 hpf comparingcontrol and cyclopamine-treated embryos. In agreement with ourproliferation data, we found no apparent difference when comparingcontrol and cyclopamine-treated embryos starting at 53 hpf, asevidenced by sox2 in situ hybridization (Fig. 1Q and R). However,the difference between control and cyclopamine-treated embryosbecame evident when treatment started at 30 or 36 hpf. Analysis at60 hpf showed that sox2 expression was almost completely absent inthe OT (Fig. 1N and O). Treatment started at 48 hpf reduced sox2
expression in the OT (Fig. 1P), while preserving its labeling at thelevel of the cerebellum. Of note, as shown in Fig. 1M, sox2 expressionoverlaps with H3-P labeling in the posterior margin of the OT, recentlyidentified as an important proliferative zone in the dorsal brain ofteleosts (Chaplin et al., 2010). Alongside the results shown in Fig. 2,these observations indicate that the Shh signaling pathway isimplicated in the proliferation and ⁄ or maintenance of a pool of neuralprogenitors in the dorsal midbrain.
OT proliferation defects in Hh mutants
To verify the results obtained with the pharmacological treatment, wetook advantage of the existence of three mutant lines that reduce oreliminate Hh signaling. Zebrafish smu (smo) mutants block most or allHh signaling, while dtr (gli1) and yot (gli2) mutants lack Gli function,allowing us to determine how Hh signaling affects the proliferationrate of neural progenitor cells.At 30 hpf we found widespread proliferation in the entire head, and
no significant difference between mutant and WT embryos in the OTregion (Fig. 3A, D, G and J). At 48 hpf dtr and yot mutants showed a
A B C
D E F
LKJ
IHG
M N O
P
Q
Fig. 2. Specificity of the effect of Shh signaling on OT progenitor proliferation. Embryos were exposed to either pharmacological inhibition or stimulation of theHh pathway for time windows as indicated. Drug treatment did not affect OT morphology. Comparison of cyclopamine (A, D), vehicle (B, E) and purmorphamine(C, F) embryos at 48 h post-fertilization (hpf) showed that 1 day of drug incubation induced notorious effects in progenitor proliferation. Whereas treatment withcyclopamine resulted in few phospho-histone H3 (H3-P)-positive cells in the OT, purmorphamine-treated embryos had a massive expression of H3-P in the dorsalmidbrain. (G–Q) Treating embryos with cyclopamine or purmorphamine for a 30-h period, followed by fixing embryos at 60 hpf, otx2 ⁄ H3-P co-labeling and DAPIcounterstaining on sections (indicated in Q) allowed detailed quantification of H3-P-positive cells in the entire OT (dashed area in M). Treatment with cyclopaminereduced in approximately 40% the number of proliferating cells in the OT (P). Data are expressed as mitotic index; error bars indicate SEM. ***Control vehicle vs.cyclopamine, P < 0.0001; *control vehicle vs. purmorphamine, P < 0.05. Data taken of at least eight fish per condition. (A–C) The inset in (N) indicates cellularresolution of DAPI staining. Dorsal views; (D–O) lateral views; black arrow indicates MHB. cyc, cyclopamine; pur, purmorphamine. Scale bar: 100 lm (A–I);50 lm (J–O).
592 C. G. Feijoo et al.
ª 2011 The Authors. European Journal of Neuroscience ª 2011 Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 33, 589–598
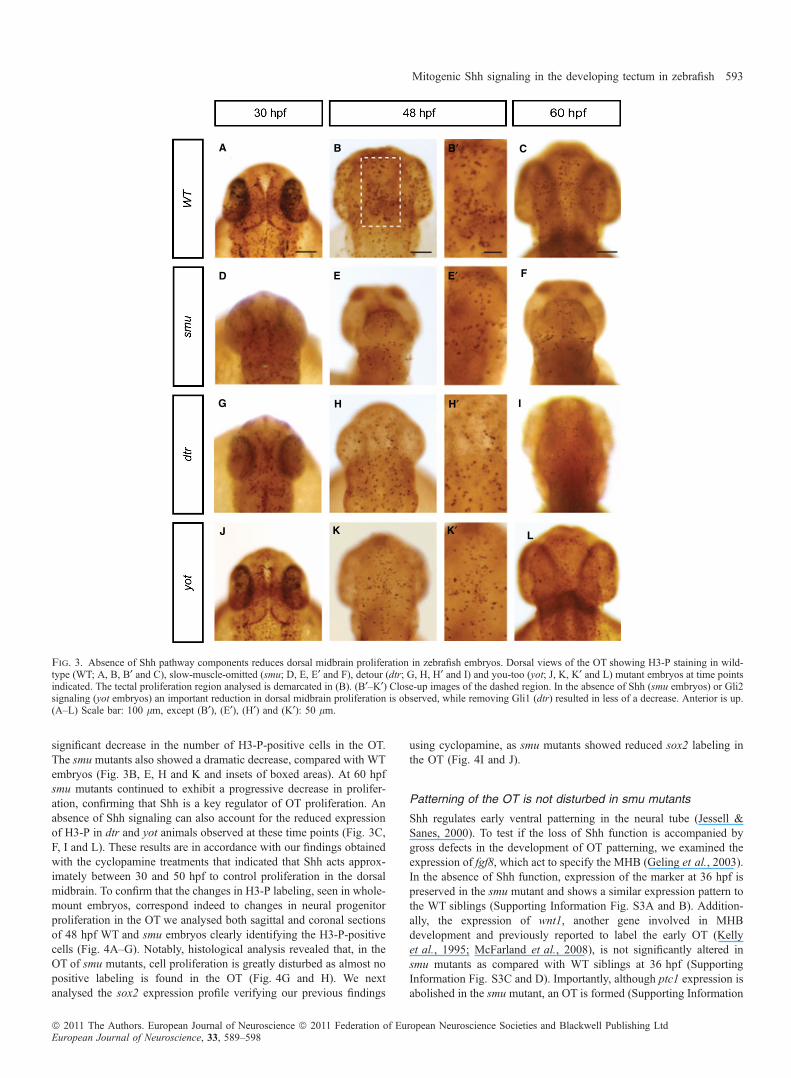
significant decrease in the number of H3-P-positive cells in the OT.The smu mutants also showed a dramatic decrease, compared with WTembryos (Fig. 3B, E, H and K and insets of boxed areas). At 60 hpfsmu mutants continued to exhibit a progressive decrease in prolifer-ation, confirming that Shh is a key regulator of OT proliferation. Anabsence of Shh signaling can also account for the reduced expressionof H3-P in dtr and yot animals observed at these time points (Fig. 3C,F, I and L). These results are in accordance with our findings obtainedwith the cyclopamine treatments that indicated that Shh acts approx-imately between 30 and 50 hpf to control proliferation in the dorsalmidbrain. To confirm that the changes in H3-P labeling, seen in whole-mount embryos, correspond indeed to changes in neural progenitorproliferation in the OT we analysed both sagittal and coronal sectionsof 48 hpf WT and smu embryos clearly identifying the H3-P-positivecells (Fig. 4A–G). Notably, histological analysis revealed that, in theOT of smu mutants, cell proliferation is greatly disturbed as almost nopositive labeling is found in the OT (Fig. 4G and H). We nextanalysed the sox2 expression profile verifying our previous findings
using cyclopamine, as smu mutants showed reduced sox2 labeling inthe OT (Fig. 4I and J).
Patterning of the OT is not disturbed in smu mutants
Shh regulates early ventral patterning in the neural tube (Jessell &Sanes, 2000). To test if the loss of Shh function is accompanied bygross defects in the development of OT patterning, we examined theexpression of fgf8, which act to specify the MHB (Geling et al., 2003).In the absence of Shh function, expression of the marker at 36 hpf ispreserved in the smu mutant and shows a similar expression pattern tothe WT siblings (Supporting Information Fig. S3A and B). Addition-ally, the expression of wnt1, another gene involved in MHBdevelopment and previously reported to label the early OT (Kellyet al., 1995; McFarland et al., 2008), is not significantly altered insmu mutants as compared with WT siblings at 36 hpf (SupportingInformation Fig. S3C and D). Importantly, although ptc1 expression isabolished in the smu mutant, an OT is formed (Supporting Information
A B C
D E F
LKJ
IHG
B′
E′
K′
H′
Fig. 3. Absence of Shh pathway components reduces dorsal midbrain proliferation in zebrafish embryos. Dorsal views of the OT showing H3-P staining in wild-type (WT; A, B, B¢ and C), slow-muscle-omitted (smu; D, E, E¢ and F), detour (dtr; G, H, H¢ and I) and you-too (yot; J, K, K¢ and L) mutant embryos at time pointsindicated. The tectal proliferation region analysed is demarcated in (B). (B¢–K¢) Close-up images of the dashed region. In the absence of Shh (smu embryos) or Gli2signaling (yot embryos) an important reduction in dorsal midbrain proliferation is observed, while removing Gli1 (dtr) resulted in less of a decrease. Anterior is up.(A–L) Scale bar: 100 lm, except (B¢), (E¢), (H¢) and (K¢): 50 lm.
Mitogenic Shh signaling in the developing tectum in zebrafish 593
ª 2011 The Authors. European Journal of Neuroscience ª 2011 Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 33, 589–598
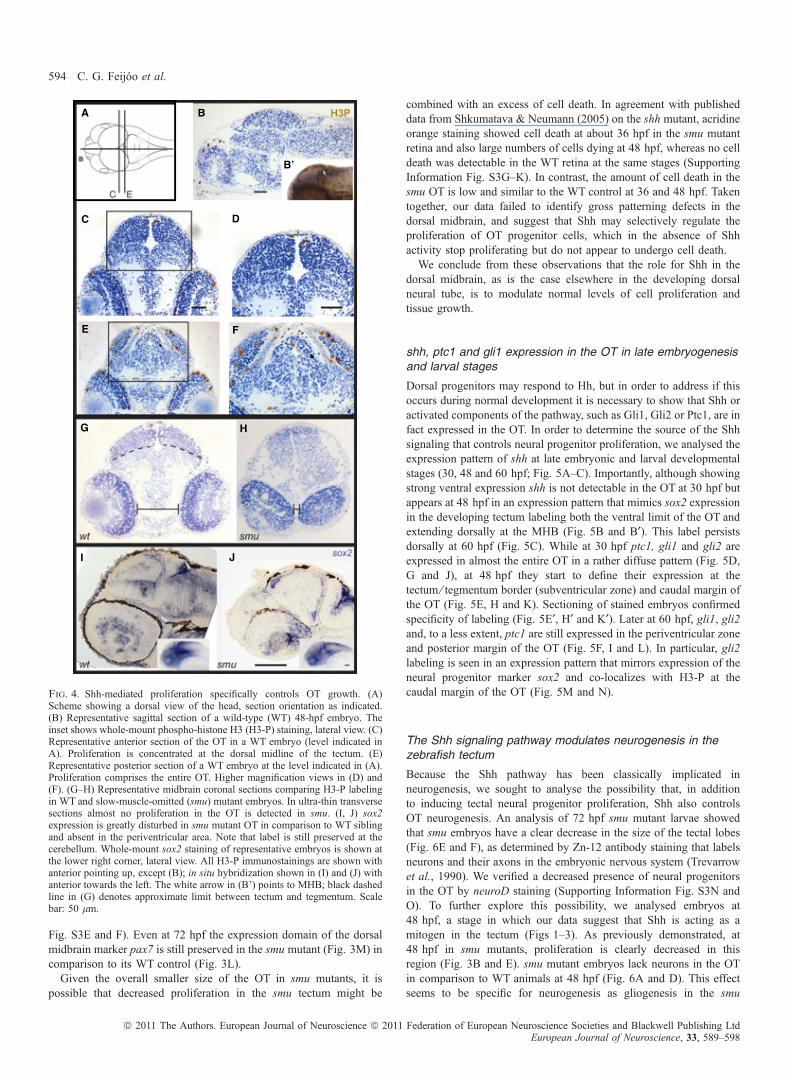
Fig. S3E and F). Even at 72 hpf the expression domain of the dorsalmidbrain marker pax7 is still preserved in the smu mutant (Fig. 3M) incomparison to its WT control (Fig. 3L).Given the overall smaller size of the OT in smu mutants, it is
possible that decreased proliferation in the smu tectum might be
combined with an excess of cell death. In agreement with publisheddata from Shkumatava & Neumann (2005) on the shh mutant, acridineorange staining showed cell death at about 36 hpf in the smu mutantretina and also large numbers of cells dying at 48 hpf, whereas no celldeath was detectable in the WT retina at the same stages (SupportingInformation Fig. S3G–K). In contrast, the amount of cell death in thesmu OT is low and similar to the WT control at 36 and 48 hpf. Takentogether, our data failed to identify gross patterning defects in thedorsal midbrain, and suggest that Shh may selectively regulate theproliferation of OT progenitor cells, which in the absence of Shhactivity stop proliferating but do not appear to undergo cell death.We conclude from these observations that the role for Shh in the
dorsal midbrain, as is the case elsewhere in the developing dorsalneural tube, is to modulate normal levels of cell proliferation andtissue growth.
shh, ptc1 and gli1 expression in the OT in late embryogenesisand larval stages
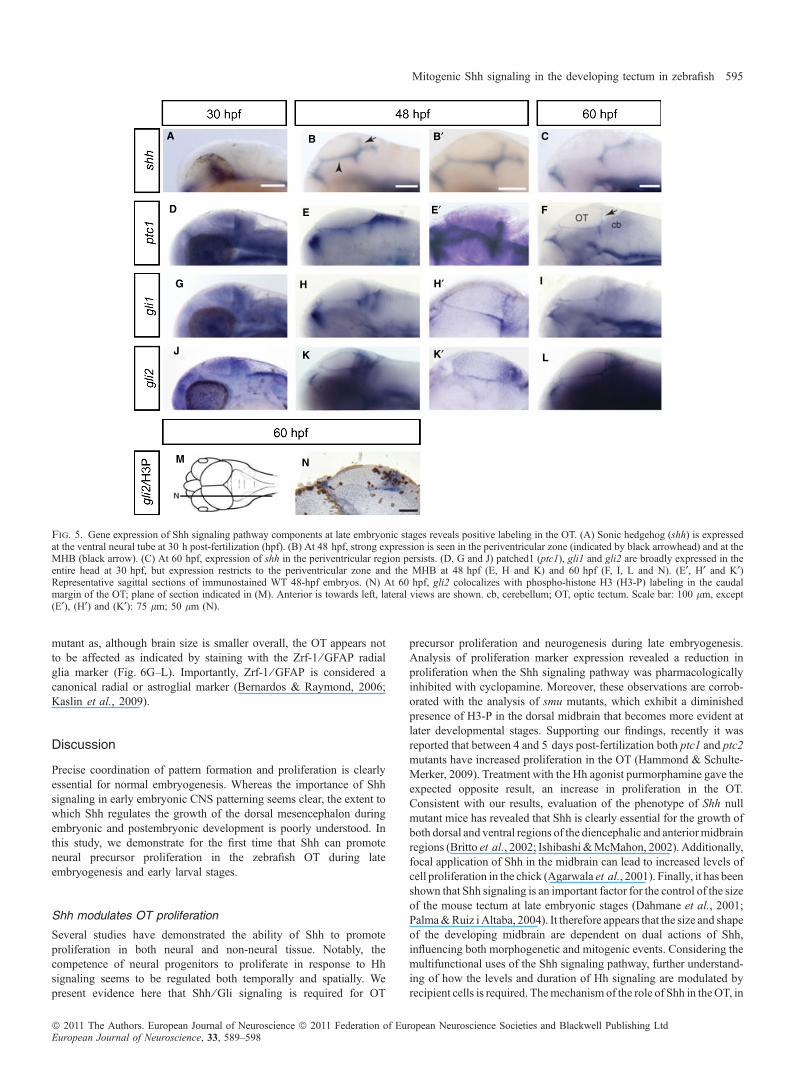
Dorsal progenitors may respond to Hh, but in order to address if thisoccurs during normal development it is necessary to show that Shh oractivated components of the pathway, such as Gli1, Gli2 or Ptc1, are infact expressed in the OT. In order to determine the source of the Shhsignaling that controls neural progenitor proliferation, we analysed theexpression pattern of shh at late embryonic and larval developmentalstages (30, 48 and 60 hpf; Fig. 5A–C). Importantly, although showingstrong ventral expression shh is not detectable in the OT at 30 hpf butappears at 48 hpf in an expression pattern that mimics sox2 expressionin the developing tectum labeling both the ventral limit of the OT andextending dorsally at the MHB (Fig. 5B and B¢). This label persistsdorsally at 60 hpf (Fig. 5C). While at 30 hpf ptc1, gli1 and gli2 areexpressed in almost the entire OT in a rather diffuse pattern (Fig. 5D,G and J), at 48 hpf they start to define their expression at thetectum ⁄ tegmentum border (subventricular zone) and caudal margin ofthe OT (Fig. 5E, H and K). Sectioning of stained embryos confirmedspecificity of labeling (Fig. 5E¢, H¢ and K¢). Later at 60 hpf, gli1, gli2and, to a less extent, ptc1 are still expressed in the periventricular zoneand posterior margin of the OT (Fig. 5F, I and L). In particular, gli2labeling is seen in an expression pattern that mirrors expression of theneural progenitor marker sox2 and co-localizes with H3-P at thecaudal margin of the OT (Fig. 5M and N).
The Shh signaling pathway modulates neurogenesis in thezebrafish tectum
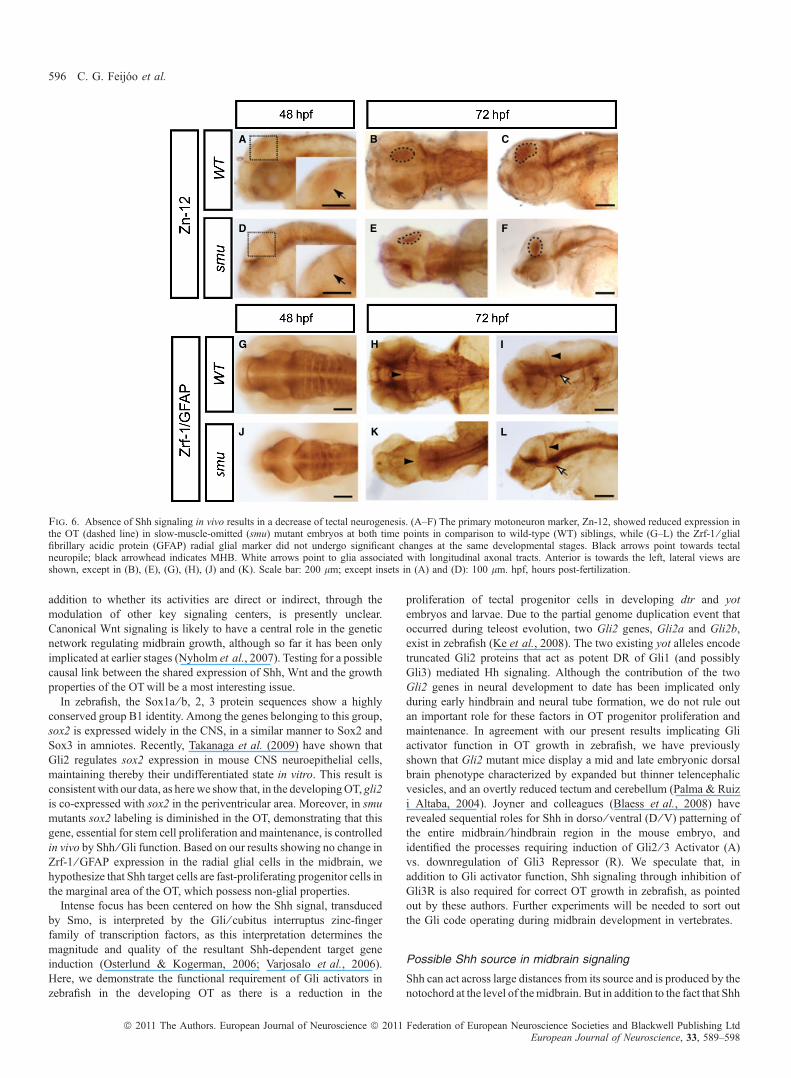
Because the Shh pathway has been classically implicated inneurogenesis, we sought to analyse the possibility that, in additionto inducing tectal neural progenitor proliferation, Shh also controlsOT neurogenesis. An analysis of 72 hpf smu mutant larvae showedthat smu embryos have a clear decrease in the size of the tectal lobes(Fig. 6E and F), as determined by Zn-12 antibody staining that labelsneurons and their axons in the embryonic nervous system (Trevarrowet al., 1990). We verified a decreased presence of neural progenitorsin the OT by neuroD staining (Supporting Information Fig. S3N andO). To further explore this possibility, we analysed embryos at48 hpf, a stage in which our data suggest that Shh is acting as amitogen in the tectum (Figs 1–3). As previously demonstrated, at48 hpf in smu mutants, proliferation is clearly decreased in thisregion (Fig. 3B and E). smu mutant embryos lack neurons in the OTin comparison to WT animals at 48 hpf (Fig. 6A and D). This effectseems to be specific for neurogenesis as gliogenesis in the smu
A B
B’
C D
E F
JI
HG
Fig. 4. Shh-mediated proliferation specifically controls OT growth. (A)Scheme showing a dorsal view of the head, section orientation as indicated.(B) Representative sagittal section of a wild-type (WT) 48-hpf embryo. Theinset shows whole-mount phospho-histone H3 (H3-P) staining, lateral view. (C)Representative anterior section of the OT in a WT embryo (level indicated inA). Proliferation is concentrated at the dorsal midline of the tectum. (E)Representative posterior section of a WT embryo at the level indicated in (A).Proliferation comprises the entire OT. Higher magnification views in (D) and(F). (G–H) Representative midbrain coronal sections comparing H3-P labelingin WT and slow-muscle-omitted (smu) mutant embryos. In ultra-thin transversesections almost no proliferation in the OT is detected in smu. (I, J) sox2expression is greatly disturbed in smu mutant OT in comparison to WT siblingand absent in the periventricular area. Note that label is still preserved at thecerebellum. Whole-mount sox2 staining of representative embryos is shown atthe lower right corner, lateral view. All H3-P immunostainings are shown withanterior pointing up, except (B); in situ hybridization shown in (I) and (J) withanterior towards the left. The white arrow in (B’) points to MHB; black dashedline in (G) denotes approximate limit between tectum and tegmentum. Scalebar: 50 lm.
594 C. G. Feijoo et al.
ª 2011 The Authors. European Journal of Neuroscience ª 2011 Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 33, 589–598
mutant as, although brain size is smaller overall, the OT appears notto be affected as indicated by staining with the Zrf-1 ⁄ GFAP radialglia marker (Fig. 6G–L). Importantly, Zrf-1 ⁄ GFAP is considered acanonical radial or astroglial marker (Bernardos & Raymond, 2006;Kaslin et al., 2009).
Discussion
Precise coordination of pattern formation and proliferation is clearlyessential for normal embryogenesis. Whereas the importance of Shhsignaling in early embryonic CNS patterning seems clear, the extent towhich Shh regulates the growth of the dorsal mesencephalon duringembryonic and postembryonic development is poorly understood. Inthis study, we demonstrate for the first time that Shh can promoteneural precursor proliferation in the zebrafish OT during lateembryogenesis and early larval stages.
Shh modulates OT proliferation
Several studies have demonstrated the ability of Shh to promoteproliferation in both neural and non-neural tissue. Notably, thecompetence of neural progenitors to proliferate in response to Hhsignaling seems to be regulated both temporally and spatially. Wepresent evidence here that Shh ⁄ Gli signaling is required for OT
precursor proliferation and neurogenesis during late embryogenesis.Analysis of proliferation marker expression revealed a reduction inproliferation when the Shh signaling pathway was pharmacologicallyinhibited with cyclopamine. Moreover, these observations are corrob-orated with the analysis of smu mutants, which exhibit a diminishedpresence of H3-P in the dorsal midbrain that becomes more evident atlater developmental stages. Supporting our findings, recently it wasreported that between 4 and 5 days post-fertilization both ptc1 and ptc2mutants have increased proliferation in the OT (Hammond & Schulte-Merker, 2009). Treatment with the Hh agonist purmorphamine gave theexpected opposite result, an increase in proliferation in the OT.Consistent with our results, evaluation of the phenotype of Shh nullmutant mice has revealed that Shh is clearly essential for the growth ofboth dorsal and ventral regions of the diencephalic and anteriormidbrainregions (Britto et al., 2002; Ishibashi &McMahon, 2002). Additionally,focal application of Shh in the midbrain can lead to increased levels ofcell proliferation in the chick (Agarwala et al., 2001). Finally, it has beenshown that Shh signaling is an important factor for the control of the sizeof the mouse tectum at late embryonic stages (Dahmane et al., 2001;Palma&Ruiz iAltaba, 2004). It therefore appears that the size and shapeof the developing midbrain are dependent on dual actions of Shh,influencing both morphogenetic and mitogenic events. Considering themultifunctional uses of the Shh signaling pathway, further understand-ing of how the levels and duration of Hh signaling are modulated byrecipient cells is required. Themechanism of the role of Shh in theOT, in
A B C
D E F
L
M N
KJ
IHG
B′
E′
K′
H′
Fig. 5. Gene expression of Shh signaling pathway components at late embryonic stages reveals positive labeling in the OT. (A) Sonic hedgehog (shh) is expressedat the ventral neural tube at 30 h post-fertilization (hpf). (B) At 48 hpf, strong expression is seen in the periventricular zone (indicated by black arrowhead) and at theMHB (black arrow). (C) At 60 hpf, expression of shh in the periventricular region persists. (D, G and J) patched1 (ptc1), gli1 and gli2 are broadly expressed in theentire head at 30 hpf, but expression restricts to the periventricular zone and the MHB at 48 hpf (E, H and K) and 60 hpf (F, I, L and N). (E¢, H¢ and K¢)Representative sagittal sections of immunostained WT 48-hpf embryos. (N) At 60 hpf, gli2 colocalizes with phospho-histone H3 (H3-P) labeling in the caudalmargin of the OT; plane of section indicated in (M). Anterior is towards left, lateral views are shown. cb, cerebellum; OT, optic tectum. Scale bar: 100 lm, except(E¢), (H¢) and (K¢): 75 lm; 50 lm (N).
Mitogenic Shh signaling in the developing tectum in zebrafish 595
ª 2011 The Authors. European Journal of Neuroscience ª 2011 Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 33, 589–598
addition to whether its activities are direct or indirect, through themodulation of other key signaling centers, is presently unclear.Canonical Wnt signaling is likely to have a central role in the geneticnetwork regulating midbrain growth, although so far it has been onlyimplicated at earlier stages (Nyholm et al., 2007). Testing for a possiblecausal link between the shared expression of Shh, Wnt and the growthproperties of the OTwill be a most interesting issue.In zebrafish, the Sox1a ⁄ b, 2, 3 protein sequences show a highly
conserved group B1 identity. Among the genes belonging to this group,sox2 is expressed widely in the CNS, in a similar manner to Sox2 andSox3 in amniotes. Recently, Takanaga et al. (2009) have shown thatGli2 regulates sox2 expression in mouse CNS neuroepithelial cells,maintaining thereby their undifferentiated state in vitro. This result isconsistent with our data, as herewe show that, in the developingOT, gli2is co-expressed with sox2 in the periventricular area. Moreover, in smumutants sox2 labeling is diminished in the OT, demonstrating that thisgene, essential for stem cell proliferation and maintenance, is controlledin vivo by Shh ⁄ Gli function. Based on our results showing no change inZrf-1 ⁄ GFAP expression in the radial glial cells in the midbrain, wehypothesize that Shh target cells are fast-proliferating progenitor cells inthe marginal area of the OT, which possess non-glial properties.Intense focus has been centered on how the Shh signal, transduced
by Smo, is interpreted by the Gli ⁄ cubitus interruptus zinc-fingerfamily of transcription factors, as this interpretation determines themagnitude and quality of the resultant Shh-dependent target geneinduction (Osterlund & Kogerman, 2006; Varjosalo et al., 2006).Here, we demonstrate the functional requirement of Gli activators inzebrafish in the developing OT as there is a reduction in the
proliferation of tectal progenitor cells in developing dtr and yotembryos and larvae. Due to the partial genome duplication event thatoccurred during teleost evolution, two Gli2 genes, Gli2a and Gli2b,exist in zebrafish (Ke et al., 2008). The two existing yot alleles encodetruncated Gli2 proteins that act as potent DR of Gli1 (and possiblyGli3) mediated Hh signaling. Although the contribution of the twoGli2 genes in neural development to date has been implicated onlyduring early hindbrain and neural tube formation, we do not rule outan important role for these factors in OT progenitor proliferation andmaintenance. In agreement with our present results implicating Gliactivator function in OT growth in zebrafish, we have previouslyshown that Gli2 mutant mice display a mid and late embryonic dorsalbrain phenotype characterized by expanded but thinner telencephalicvesicles, and an overtly reduced tectum and cerebellum (Palma & Ruizi Altaba, 2004). Joyner and colleagues (Blaess et al., 2008) haverevealed sequential roles for Shh in dorso ⁄ ventral (D ⁄ V) patterning ofthe entire midbrain ⁄ hindbrain region in the mouse embryo, andidentified the processes requiring induction of Gli2 ⁄ 3 Activator (A)vs. downregulation of Gli3 Repressor (R). We speculate that, inaddition to Gli activator function, Shh signaling through inhibition ofGli3R is also required for correct OT growth in zebrafish, as pointedout by these authors. Further experiments will be needed to sort outthe Gli code operating during midbrain development in vertebrates.
Possible Shh source in midbrain signaling
Shh can act across large distances from its source and is produced by thenotochord at the level of themidbrain. But in addition to the fact that Shh
A B C
D E F
G H I
J K L
Fig. 6. Absence of Shh signaling in vivo results in a decrease of tectal neurogenesis. (A–F) The primary motoneuron marker, Zn-12, showed reduced expression inthe OT (dashed line) in slow-muscle-omitted (smu) mutant embryos at both time points in comparison to wild-type (WT) siblings, while (G–L) the Zrf-1 ⁄ glialfibrillary acidic protein (GFAP) radial glial marker did not undergo significant changes at the same developmental stages. Black arrows point towards tectalneuropile; black arrowhead indicates MHB. White arrows point to glia associated with longitudinal axonal tracts. Anterior is towards the left, lateral views areshown, except in (B), (E), (G), (H), (J) and (K). Scale bar: 200 lm; except insets in (A) and (D): 100 lm. hpf, hours post-fertilization.
596 C. G. Feijoo et al.
ª 2011 The Authors. European Journal of Neuroscience ª 2011 Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 33, 589–598

itself diffuses a significant distance from the ventral midline, theremightbe a secondary relay of intermediate signals that propagates the originalShh signaling event through the tectum. Indeed, recent evidencesuggests that Shh can diffuse across the entire extent of the D ⁄ V axis,provided its movement is not significantly impeded by feedbackantagonists of Shh signaling such as Hhip1 (Jeong &McMahon, 2005).As the main Shh source, the notochord has most likely a key role inregulating the growth of the brain vesicle – in chick, transientdisplacement of the notochord from the midbrain results in a reducedexpansion of the brain vesicle. Implanting Shh-secreting cells restoredthe expansion (reviewed in Fleming et al., 2004). Of interest, recentreports have shown that several Hh-binding factors (MEGALIN, BOC)and Hh effectors (Smo, Gli1, 2, 3, DZIP) are expressed in the dorsalmidbrain and theMHB (Aglyamova&Agarwala, 2007). Here we reportdorsal expression of shh in 2-day-old zebrafish that correlates with asox2 positive labeling and high levels of proliferation in the OT.Interestingly, according to recent reportsHhpathway genes are excludedfrom the developing cerebellum at similar stages to the ones examined inour study (McFarland et al., 2008; Chaplin et al., 2010). It has thereforebeen proposed that Hh signaling is not involved in proliferation of thegranule cell precursor population in zebrafish (Chaplin et al., 2010).
Little is known about the differences between constitutivelyneurogenic and non-neurogenic post-embryonic brain regions inteleosts. Adult proliferation and neurogenesis seem to recapitulatesome of the cellular mechanisms used during embryonic development.In light of our results, a detailed study of shh expression should beperformed at later developmental stages.
Taken together, the findings in our study demonstrate that, in theabsence of Shh signaling, incomplete OT development results largelyfrom a deficiency in proliferation and differentiation of neuralprogenitor cells.
Supporting Information
Additional supporting information may be found in the online versionof this article:Fig. S1. Pharmacological treatments controls. Drug treatment wasinitiated at 24 hpf and embryos were fixed at 48 hpf (A–D) or startedat 30 hpf fixing embryos at 60 hpf (E–G). Cyclopamine effectivelyblocks the Shh pathway, as shown by the decreased expression of ptc1in the midbrain area in comparison to its vehicle (EtOH). In contrast,purmorphamine treatment showed increased expression of ptc1 incomparison to the vehicle (DMSO). (E and G) Controls weredeveloped at different times in order to match to their correspondingtreatment. Anterior is towards left, lateral views are showed.Arrowhead indicates MHB. Scale bar: 50 lm. cyc, cyclopamine.Fig. S2. Developing OT cells are sensitive to alterations of the Hhpathway. Sagittal confocal optical section analysis after pharmacolog-ical treatments at time frames as indicated. (A, D) Cyclopaminetreatment results in diminished H3-P-positive cell number in the OT. (C,D) The opposite effect is achieved by using purmorphamine where theproliferative cell number increases significantly. Means significantlydifferent in (C)P < 0.05. (E) Dorsal confocal sections show the locationof the active proliferative zone within the OT at 60 hpf. The level in thez-axis for each image is represented by a blue line. (F) The ventralregion, although showing strong POMC expression in the pituitaryanlage, only reveals a few proliferating cells. Anterior is towards theleft. Lateral views are shown from (A) to (C), and dorsal views in (E)and (F). Gray andwhite dashed lines represent the head limit and the eyeposition, respectively. e, eye; OT, optic tectum. Scale bar: 50 lm.Fig. S3. Patterning of midbrain and cell death in smu embryos. (A, B)The expression of the MHB marker, fgf8, is normal in smu embryos,
and (C, D) wnt1 demarcates the dorsal limit of the OT in a semi-circlesimilarly in WT and smu embryos at 36 hpf. (E, F) ptc1 expression insmu embryos is absent. At 72 hpf the expression of pax7 is preservedin smu (L, M). The proneural marker, neuroD, is reduced in the OT ofsmu mutants compared with WT (black arrowhead, in N vs O). (G–J)Whereas dead cells can be identified in the eye of smu mutant embryos(in both 36 hpf and 48 hpf, white arrows), there is no increased celldeath in the dorsal midbrain in smu embryos, as revealed by acridineorange staining (white arrowhead in H). (K) 48 hpf embryos treatedwith CuSO4 were shown as a positive control (Hernandez et al.,2007), indicating cell death of neuromasts located in the anteriorlateral line (small white arrowhead). Anterior is towards left, the entirepanel show lateral views. Scale bar: 100 lm.Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset by Wiley-Blackwell.Technical support issues arising from supporting information (otherthan missing files) should be addressed to the authors.
Acknowledgements
We would like to thank Catalina Lafourcade and former lab memberGustavo Cerda for technical assistance, and Dr Rolf Karlstrom for hiscritical advice and Hh pathway mutants lines. We also thank DrShlomo Melmed for the PomC::gfp line; Dr Miguel Allende for kindlyproviding zebrafish embryos and reagents and critical suggestions; DrKate Whitlock and members of the Palma lab for comments on themanuscript. Cyclopamine was kindly provided by Infinity Pharma-ceuticals. This work was supported by ICM P06-039F (V.P.), Fondecytgrant 1070248 (V.P.) and UNAB DI45-05 ⁄ R grant (C.G.F.).
Abbreviations
D ⁄ V, dorsal ⁄ ventral; DMSO, dimethylsulfoxide; DR, dominant repressor; dtr,detour; GFAP, glial fibrillary acidic protein; GFP, green fluorescent protein; H3-P, phospho-histone H3; Hh, hedgehog; hpf, hours post-fertilization; MHB,midbrain–hindbrain boundary; OT, optic tectum; Ptc1, Patched1; Shh, Sonichedgehog; Smo, Smoothened; smu, slow-muscle-omitted; WT, wild-type; yot,you-too.
References
Aanstad, P., Santos, N., Corbit, K.C., Scherz, P.J., Trinh, L.A., Salvenmoser,W., Huisken, J., Reiter, J.F. & Stainier, D.Y. (2009) The extracellular domainof Smoothened regulates ciliary localization and is required for high-level Hhsignaling. Curr. Biol., 19, 1034–1039.
Agarwala, S., Sanders, T.A. & Ragsdale, C.W. (2001) Sonic hedgehog controlof size and shape in midbrain pattern formation. Science, 291, 2147–2150.
Aglyamova, G.V. & Agarwala, S. (2007) Gene expression analysis of thehedgehog signaling cascade in the chick midbrain and spinal cord. Dev.Dyn., 236, 1363–1373.
Barresi, M.J., Stickney, H.L. & Devoto, S.H. (2000) The zebrafish slow-muscle-omitted gene product is required for Hedgehog signal transductionand the development of slow muscle identity. Development, 127, 2189–2199.
Bernardos, R.L. & Raymond, P.A. (2006) GFAP transgenic zebrafish. GeneExpr. Patterns, 6, 1007–1013.
Blaess, S., Stephen, D. & Joyner, A.L. (2008) Gli3 coordinates three-dimensional patterning and growth of the tectum and cerebellum byintegrating Shh and Fgf8 signaling. Development, 135, 2093–2103.
Briscoe, J. (2006) Agonizing hedgehog. Nat. Chem. Biol., 2, 10–11.Britto, J., Tannahill, D. & Keynes, R. (2002) A critical role for sonic hedgehog
signaling in the early expansion of the developing brain. Nat. Neurosci., 5,103–110.
Chaplin, N., Tendeng, C. & Wingate, R.J. (2010) Absence of an externalgerminal layer in zebrafish and shark reveals a distinct, anamniote groundplan of cerebellum development. J. Neurosci., 30, 3048–3057.
Mitogenic Shh signaling in the developing tectum in zebrafish 597
ª 2011 The Authors. European Journal of Neuroscience ª 2011 Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 33, 589–598

Concordet, J.P., Lewis, K.E., Moore, J.W., Goodrich, L.V., Johnson, R.L.,Scott, M.P. & Ingham, P.W. (1996) Spatial regulation of a zebrafishpatched homologue reflects the roles of sonic hedgehog and protein kinaseA in neural tube and somite patterning. Development, 122, 2835–2846.
Dahmane, N., Sanchez, P., Gitton, Y., Palma, V., Sun, T., Beyna, M., Weiner, H.& Ruiz i Altaba, A. (2001) The Sonic Hedgehog-Gli pathway regulatesdorsal brain growth and tumorigenesis. Development, 128, 5201–5212.
Fleming, A., Keynes, R. & Tannahill, D. (2004) A central role for thenotochord in vertebral patterning. Development, 131, 873–880.
Furthauer, M., Thisse, C. & Thisse, B. (1997) A role for FGF-8 in thedorsoventral patterning of the zebrafish gastrula. Development, 124, 4253–4264.
Geling, A., Itoh, M., Tallafuss, A., Chapouton, P., Tannhauser, B., Kuwada,J.Y., Chitnis, A.B. & Bally-Cuif, L. (2003) bHLH transcription factor Her5links patterning to regional inhibition of neurogenesis at the midbrain-hindbrain boundary. Development, 130, 1591–1604.
Graham, V., Khudyakov, J., Ellis, P. & Pevny, L. (2003) SOX2 functions tomaintain neural progenitor identity. Neuron, 39, 749–765.
Hammond, C.L. & Schulte-Merker, S. (2009) Two populations of endochondralosteoblasts with differential sensitivity to Hedgehog signaling. Development,136, 3991–4000.
Hammond, C.L., Hinits, Y., Osborn, D.P., Minchin, J.E., Tettamanti, G. &Hughes, S.M. (2007) Signals and myogenic regulatory factors restrict pax3and pax7 expression to dermomyotome-like tissue in zebrafish. Dev. Biol.,302, 504–521.
Hernandez, P.P., Olivari, F.A., Sarrazin, A.F., Sandoval, P.C. & Allende, M.L.(2007) Regeneration in zebrafish lateral line neuromasts: expression of theneural progenitor cell marker sox2 and proliferation-dependent and-independent mechanisms of hair cell renewal. Dev. Neurobiol., 67, 637–654.
Ishibashi, M. & McMahon, A.P. (2002) A sonic hedgehog-dependent signalingrelay regulates growth of diencephalic and mesencephalic primordia in theearly mouse embryo. Development, 129, 4807–4819.
Jeong, J. & McMahon, A.P. (2005) Growth and pattern of the mammalianneural tube are governed by partially overlapping feedback activities of thehedgehog antagonists patched 1 and Hhip1. Development, 132, 143–154.
Jessell, T.M. & Sanes, J.R. (2000) Development. The decade of the developingbrain. Curr. Opin. Neurobiol., 10, 599–611.
Jowett, T. & Lettice, L. (1994) Whole-mount in situ hybridizations on zebrafishembryos using a mixture of digoxigenin- and fluorescein-labelled probes.Trends Genet., 10, 73–74.
Karlstrom, R.O., Trowe, T., Klostermann, S., Baier, H., Brand, M., Crawford,A.D., Grunewald, B., Haffter, P., Hoffmann, H., Meyer, S.U., Muller, B.K.,Richter, S., van Eeden, F.J., Nusslein-Volhard, C. & Bonhoeffer, F. (1996)Zebrafish mutations affecting retinotectal axon pathfinding. Development,123, 427–438.
Karlstrom, R.O., Tyurina, O.V., Kawakami, A., Nishioka, N., Talbot, W.S.,Sasaki, H. & Schier, A.F. (2003) Genetic analysis of zebrafish gli1 and gli2reveals divergent requirements for gli genes in vertebrate development.Development, 130, 1549–1564.
Kaslin, J., Ganz, J., Geffarth, M., Grandel, H., Hans, S. & Brand, M. (2009)Stem cells in the adult zebrafish cerebellum: initiation and maintenance of anovel stem cell niche. J. Neurosci., 29, 6142–6153.
Kasper, M., Regl, G., Frischauf, A.M. & Aberger, F. (2006) GLI transcriptionfactors: mediators of oncogenic Hedgehog signalling. Eur. J. Cancer, 42,437–445.
Ke, Z., Kondrichin, I., Gong, Z. & Korzh, V. (2008) Combined activity of thetwo Gli2 genes of zebrafish play a major role in Hedgehog signaling duringzebrafish neurodevelopment. Mol. Cell. Neurosci., 37, 388–401.
Kelly, G.M., Erezyilmaz, D.F. & Moon, R.T. (1995) Induction of a secondaryembryonic axis in zebrafish occurs following the overexpression of beta-catenin. Mech. Dev., 53, 261–273.
Korzh, V., Sleptsova, I., Liao, J., He, J. & Gong, Z. (1998) Expression ofzebrafish bHLH genes ngn1 and nrd defines distinct stages of neuraldifferentiation. Dev. Dyn., 213, 92–104.
Krauss, S., Concordet, J.P. & Ingham, P.W. (1993) A functionally conservedhomolog of the Drosophila segment polarity gene hh is expressed in tissueswith polarizing activity in zebrafish embryos. Cell, 75, 1431–1444.
Liu, N., Huang, H., Yang, Z., Herzog, W., Hammerschmidt, M., Lin, S. &Melmed, S. (2003) Pituitary corticotroph ontogeny and regulation intransgenic zebrafish. Mol. Endocrinol., 17, 959–966.
Lum, L. & Beachy, P.A. (2004) The Hedgehog response network: sensors,switches, and routers. Science, 304, 1755–1759.
Lun, K. & Brand, M. (1998) A series of no isthmus (noi) alleles of the zebrafishpax2.1 gene reveals multiple signaling events in development of themidbrain-hindbrain boundary. Development, 125, 3049–3062.
Machold, R., Hayashi, S., Rutlin, M., Muzumdar, M.D., Nery, S., Corbin, J.G.,Gritli-Linde, A., Dellovade, T., Porter, J.A., Rubin, L.L., Dudek, H.,McMahon, A.P. & Fishell, G. (2003) Sonic hedgehog is required forprogenitor cell maintenance in telencephalic stem cell niches. Neuron, 39,937–950.
McFarland, K.A., Topczewska, J.M., Weidinger, G., Dorsky, R.I. & Appel, B.(2008) Hh and Wnt signaling regulate formation of olig2 + neurons in thezebrafish cerebellum. Dev. Biol., 318, 162–171.
Mueller, T. & Wullimann, M.F. (2003) Anatomy of neurogenesis in the earlyzebrafish brain. Dev. Brain Res., 140, 137–155.
Nguyen, V., Joly, J. & Bourrat, F. (2001a) An in situ screen for genescontrolling cell proliferation in the optic tectum of the medaka (Oryziaslatipes). Mech. Dev., 107, 55–67.
Nguyen, V., Candal Suarez, E.M., Sharif, A., Joly, J.S. & Bourrat, F. (2001b)Expression of OI-KIP, a cyclin-dependent kinase inhibitor, in embryonic andadult medaka (Oryzias latipes) central nervous system. Dev. Dyn., 222, 439–449.
Nyholm, M.K., Wu, S.F., Dorsky, R.I. & Grinblat, Y. (2007) The zebrafishzic2a-zic5 gene pair acts downstream of canonical Wnt signaling to controlcell proliferation in the developing tectum. Development, 134, 735–746.
Osterlund, T. & Kogerman, P. (2006) Hedgehog signalling: how to get fromSmo to Ci and Gli. Trends Cell Biol., 16, 176–180.
Palma, V. & Ruiz i Altaba, A. (2004) Hedgehog-GLI signaling regulates thebehavior of cells with stem cell properties in the developing neocortex.Development, 131, 337–345.
Palma, V., Lim, D.A., Dahmane, N., Sanchez, P., Brionne, T.C., Herzberg,C.D., Gitton, Y., Carleton, A., Alvarez-Buylla, A. & Ruiz i Altaba, A. (2005)Sonic hedgehog controls stem cell behavior in the postnatal and adult brain.Development, 132, 335–344.
Pevny, L.H. & Lovell-Badge, R. (1997) Sox genes find their feet. Curr. Opin.Genet. Dev., 7, 338–344.
Puelles, L. & Rubenstein, J.L. (1993) Expression patterns of homeobox andother putative regulatory genes in the embryonic mouse forebrain suggest aneuromeric organization. Trends Neurosci., 16, 472–479.
Sarrazin, A.F., Villablanca, E.J., Nunez, V.A., Sandoval, P.C., Ghysen, A. &Allende, M.L. (2006) Proneural gene requirement for hair cell differentiationin the zebrafish lateral line. Dev. Biol., 295, 534–545.
Schmatolla, E. (1972) Dependence of tectal neuron differentiation on opticinnervation in teleost fish. J. Embryol. Exp. Morphol., 27, 555–576.
Shkumatava, A. & Neumann, C.J. (2005) Shh directs cell-cycle exit byactivating p57Kip2 in the zebrafish retina. EMBO Rep., 6, 563–569.
Taipale, J., Chen, J.K., Cooper, M.K., Wang, B., Mann, R.K., Milenkovic, L.,Scott, M.P. & Beachy, P.A. (2000) Effects of oncogenic mutations inSmoothened and Patched can be reversed by cyclopamine. Nature, 406,1005–1009.
Takanaga, H., Tsuchida-Straeten, N., Nishide, K., Watanabe, A., Aburatani, H.& Kondo, T. (2009) Gli2 is a novel regulator of sox2 expression intelencephalic neuroepithelial cells. Stem Cells, 27, 165–174.
Trevarrow, B., Marks, D.L. & Kimmel, C.B. (1990) Organization of hindbrainsegments in the zebrafish embryo. Neuron, 4, 669–679.
Tyurina, O.V., Guner, B., Popova, E., Feng, J., Schier, A.F., Kohtz, J.D. &Karlstrom, R.O. (2005) Zebrafish Gli3 functions as both an activator and arepressor in Hedgehog signaling. Dev. Biol., 277, 537–556.
Ulloa, F. & Briscoe, J. (2007) Morphogens and the control of cell proliferationand patterning in the spinal cord. Cell Cycle, 6, 2640–2649.
Varga, Z.M., Amores, A., Lewis, K.E., Yan, Y.L., Postlethwait, J.H., Eisen, J.S.& Westerfield, M. (2001) Zebrafish smoothened functions in ventral neuraltube specification and axon tract formation. Development, 128, 3497–3509.
Varjosalo, M., Li, S.P. & Taipale, J. (2006) Divergence of hedgehog signaltransduction mechanism between Drosophila and mammals. Dev. Cell, 10,177–186.
van de Water, S., van de Wetering, M., Joore, J., Esseling, J., Bink, R., Clevers,H. & Zivkovic, D. (2001) Ectopic Wnt signal determines the eyelessphenotype of zebrafish masterblind mutant. Development, 128, 3877–3888.
Westerfield, M. (1993) The Zebrafish Book: A Guide for The Laboratory Use ofZebrafish (Brachydanio rerio). University of Oregon Press, Eugene, OR.
Williams, J.A., Barrios, A., Gatchalian, C., Rubin, L., Wilson, S.W. & Holder,N. (2000) Programmed cell death in zebrafish rohon beard neurons isinfluenced by TrkC1 ⁄ NT-3 signaling. Dev. Biol., 226, 220–230.
Wullimann, M.F. & Knipp, S. (2000) Proliferation pattern changes in thezebrafish brain from embryonic through early postembryonic stages. Anat.Embryol. (Berl), 202, 385–400.
Zhao, S., Nichols, J., Smith, A.G. & Li, M. (2004) SoxB transcription factorsspecify neuroectodermal lineage choice in ES cells. Mol. Cell. Neurosci., 27,332–342.
598 C. G. Feijoo et al.
ª 2011 The Authors. European Journal of Neuroscience ª 2011 Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 33, 589–598





























