Embed Size (px)
Citation preview

Sequencing, biochemical characterization, crystalstructure and molecular dynamics of cellobiohydrolaseCel7A from Geotrichum candidum 3CAnna S. Borisova1,2, Elena V. Eneyskaya2, Kirill S. Bobrov2, Suvamay Jana3, Anton Logachev4,Dmitrii E. Polev5, Alla L. Lapidus6, Farid M. Ibatullin2, Umair Saleem1, Mats Sandgren1,Christina M. Payne3, Anna A. Kulminskaya2,7 and Jerry St�ahlberg1
1 Department of Chemistry and Biotechnology, Swedish University of Agricultural Sciences, Uppsala, Sweden
2 National Research Centre «Kurchatov Institute», B.P. Konstantinov Petersburg Nuclear Physics Institute, Gatchina, Orlova roscha, Russia
3 Department of Chemical and Materials Engineering, University of Kentucky, Lexington, KY, USA
4 Theodosius Dobzhansky Center for Genome Bioinformatics, St. Petersburg State University, Russia
5 Research Resource Centre «Molecular and Cell Technologies», St. Petersburg State University, Russia
6 Centre for Algorithmic Biotechnology, St. Petersburg Academic University, Russia
7 Department of Medical Physics, Peter the Great St. Petersburg Polytechnic University, Russia
Keywords
biomass degradation; cellulase; Geotrichum
candidum; molecular dynamics; X-ray
structure
Correspondence
J. St�ahlberg, Department of Chemistry and
Biotechnology, Swedish University of
Agricultural Sciences, PO Box 7015, SE-750
07 Uppsala, Sweden
Tel: +46-18-673182
E-mail: [email protected]
C. M. Payne, Department of Chemical and
Materials Engineering, University of
Kentucky, 177 F. Paul Anderson Tower,
Lexington, KY 40506, USA
Fax: +1 859 323 1929
Tel: +1 859 257 2902
E-mail: [email protected]
A. A. Kulminskaya, National Research
Centre «Kurchatov Institute», B.P.
Konstantinov Petersburg Nuclear Physics
Institute, 188300, Gatchina, Orlova roscha,
Russia
Fax: +7 81371 32303
Tel: +7 813 7132014
E-mail: [email protected]
Present address
Umair Saleem, Birkedommervej 17, 3TH,
2400, København NV, Denmark
(Received 17 June 2015, revised 13 August
2015, accepted 4 September 2015)
The ascomycete Geotrichum candidum is a versatile and efficient decay
fungus that is involved, for example, in biodeterioration of compact
discs; notably, the 3C strain was previously shown to degrade filter
paper and cotton more efficiently than several industrial enzyme prepa-
rations. Glycoside hydrolase (GH) family 7 cellobiohydrolases (CBHs)
are the primary constituents of industrial cellulase cocktails employed in
biomass conversion, and feature tunnel-enclosed active sites that enable
processive hydrolytic cleavage of cellulose chains. Understanding the
structure–function relationships defining the activity and stability of
GH7 CBHs is thus of keen interest. Accordingly, we report the compre-
hensive characterization of the GH7 CBH secreted by G. candidum
(GcaCel7A). The bimodular cellulase consists of a family 1 cellulose-
binding module (CBM) and linker connected to a GH7 catalytic domain
that shares 64% sequence identity with the archetypal industrial GH7
CBH of Hypocrea jecorina (HjeCel7A). GcaCel7A shows activity on Avi-
cel cellulose similar to HjeCel7A, with less product inhibition, but has a
lower temperature optimum (50 °C versus 60–65 °C, respectively). Five
crystal structures, with and without bound thio-oligosaccharides, show
conformational diversity of tunnel-enclosing loops, including a form with
partial tunnel collapse at subsite –4 not reported previously in GH7.
Also, the first O-glycosylation site in a GH7 crystal structure is reported
– on a loop where the glycan probably influences loop contacts across
the active site and interactions with the cellulose surface. The GcaCel7A
structures indicate higher loop flexibility than HjeCel7A, in accordance
with sequence modifications. However, GcaCel7A retains small fluctua-
tions in molecular simulations, suggesting high processivity and low
endo-initiation probability, similar to HjeCel7A.
Database
Structural data are available in the Protein Data Bank under the accession numbers 5AMP,
4ZZV, 4ZZW, 4ZZT, and 4ZZU. The Geotrichum candidum GH family 7 cellobiohydrolase
nucleotide sequence is available in GenBank under accession number KJ958925.
1FEBS Journal (2015) ª 2015 FEBS

doi:10.1111/febs.13509 Enzymes
Glycoside hydrolase family 7 reducing end acting cellobiohydrolase
Introduction
Cellulose-degrading enzymes play a key role in global
carbon recycling, a process of considerable ecological
importance. As much as ~ 15% of all atmospheric car-
bon dioxide is fixed yearly by plants and incorporated
into vast amounts of biomass, which is eventually
degraded and recycled by enzymes from fungi and bac-
teria [1]. Lignocellulosic biomass is also by far the most
abundant renewable carbon source available to human-
ity in the transition from fossil-based to sustainable pro-
duction of fuels and chemicals. The potential of
enzymes for sustainable biomass utilization is attracting
growing attention and is expected to play a vital role in
the future. Already today, cellulases and associated
biomass-degrading enzymes constitute the third largest
group of industrially produced enzymes [2]; these
enzymes are commonly used in applications such as cot-
ton processing and paper recycling, as detergent
enzymes, in juice extraction, and as animal feed addi-
tives [3,4]. Given a viable worldwide biomass-to-biofuel
industry, cellulases will undoubtedly be the most highly
produced proteins in the world, by orders of magnitude.
Basic and applied research on cellulases for biofuel
production began in the 1970s [5,6]. The enzyme sys-
tem most extensively studied is that of the filamentous
ascomycete fungus Hypocrea jecorina (also known as
Trichoderma reesei). H. jecorina remains the predomi-
nant organism used for the production of cellulases,
thanks to the development of hyperproducing indus-
trial strains capable of secreting > 50 g�L�1 of protein
[7]. However, each component of the native
H. jecorina secretome is not necessarily primed for effi-
cient hydrolytic turnover and stability under industrial
conditions, prompting the search for alternative
enzymes among the vast array of cellulolytic microor-
ganisms that exist in nature.
The filamentous yeast-like fungus Geotrichum
candidum strain 3C was isolated from a rotting rope
[8] and found to have high cellulolytic and xylanolytic
activities [9,10]. As early as the 1970s, it was shown
that the G. candidum 3C cellulase complex was more
efficient than that of well-studied Trichoderma sp.
[8,11,12]. The cellulases of G. candidum 3C were ini-
tially characterized, but unfortunately, the enzymes
have been undeservedly forgotten for decades. Never-
theless, an enzyme preparation from this fungus, ‘Cel-
lokandin G10x’ [11], has been used in the pulp and
paper industry for waste paper utilization [13], and
applied research has been carried out on G. candidum
3C to improve the process of bleaching of softwood
and hardwood kraft pulp [9,14]. Interestingly,
G. candidum and related species have been found to be
able to degrade various natural and artificial materials,
and to be responsible for biodeterioration and destroy-
ing the information pits in compact discs [15]. From
an evolutionary point of view, G. candidum is rela-
tively distantly related to other ascomycete fungi with
characterized glycoside hydrolase (GH) family 7 cel-
lobiohydrolases (CBHs). The draft genome sequence
of G. candidum 3C has been published [16] and is
available in GenBank under the strain name Galacto-
myces candidus 3C. Also, the draft genome sequence of
another G. candidum strain CLIB 918 (ATCC 204307),
described as a dairy yeast within the Sachharomy-
cotina, has been published very recently [17].
The most abundant protein secreted by G. candidum
3C was previously isolated and partially characterized,
showing properties that are typical of GH7 CBHs
(EC 3.2.1.176) [8,10,11]. GH7 CBHs are commonly
the major components in the secretomes of potent cel-
lulolytic fungi and are also key rate-limiting factors in
commercial cellulase cocktails [18]. Approximately
Abbreviations
APO1, apo structure 1 of catalytic domain of Geotrichum candidum Cel7A; APO2, apo structure 2 of catalytic domain of Geotrichum
candidum Cel7A; BC, bacterial cellulose; CBH, cellobiohydrolase; CBM, cellulose-binding module; CMC, carboxymethyl cellulose; CNP-G2,
2-chloro-4-nitrophenyl b-cellobioside; G2, catalytic domain of Geotrichum candidum Cel7A ligand complex with cellobioside; G3, catalytic
domain of Geotrichum candidum Cel7A ligand complex with cellotrioside; G4, catalytic domain of Geotrichum candidum Cel7A ligand
complex with cellotetraoside; GcaCel7A_CD, catalytic domain of Geotrichum candidum Cel7A; GcaCel7A, Geotrichum candidum Cel7A; GH,
glycoside hydrolase; HirCel7A, Heterobasidion irregulare Cel7A; HjeCel7A, Hypocrea jecorina Cel7A; MD, molecular dynamics; PASC,
phosphoric acid-swollen cellulose; PchCel7D, Phanerochaete chrysosporium Cel7D; PDB, Protein Data Bank; PEG, poly(ethylene glycol);
pNP-G2, p-nitrophenyl β-cellobioside; pNP-Lac, p-nitrophenyl β-lactoside; RemCel7A, Rasamsonia emersonii Cel7A; RMSF, root-mean-square
fluctuation; XGO, xyloglucan oligosaccharide.
2 FEBS Journal (2015) ª 2015 FEBS
Characterization of G. candidum Cel7A A. S. Borisova et al.

one-third of the known GH7 members, including
Cel7A from G. candidum (GcaCel7A) and Cel7A from
H. jecorina (HjeCel7A), are bimodular proteins with a
family 1 cellulose-binding module (CBM) connected to
the GH7 catalytic module by a highly glycosylated
and flexible Ser/Thr-rich linker peptide.
Three-dimensional structures of the catalytic
domains of nine GH7 CBHs have been published pre-
viously, the first being HjeCel7A and the most recent
being Aspergillus fumigatus Cel7A [19–27]. The struc-
tures share a common b-jelly roll fold with a curved
b-sandwich constructed from two largely antiparallel
b-sheets packing face-to-face, forming an approxi-
mately 50-�A-long substrate-binding groove along the
GH7 catalytic module. A key structural feature of
GH7 CBHs is that long loops extend the edges of the
b-sandwich and effectively enclose the active site in a
tunnel. This enables the enzymes to act processively
along a cellulose chain and cleave off numerous cel-
lobiose units before detachment from the substrate,
which is believed to be key to their efficiency on highly
crystalline cellulose [18]. GH7 CBHs act preferentially
from the reducing towards the nonreducing end of cel-
lulose chains, in contrast with GH6 CBHs, which
work in the opposite direction.
The active site harbours 11 glucosyl-binding sub-
sites, numbered �7 to +4 from the nonreducing end of
the cellulose chain, and cleavage occurs between sub-
sites �1 and +1 [28]. Sequence identities are high
(> 50%) among the CBHs of family 7, and the cellu-
lose-binding active site is highly conserved, including
four Trp residues that serve as sugar-binding platforms
at subsites �7, �4, �2, and +1. Differences that may
relate to function occur primarily in the length and
sequence of loop regions, varying the accessibility of
the active site and the dynamics of loop movements; in
turn, these variations influence key enzyme properties,
such as processivity, product inhibition, endo-initiation
propensity, and the rate of release of non-productively
bound enzyme [23,29]. Computational investigation
offers complementary insights to those from X-ray
crystallography regarding protein dynamics and the
thermodynamics of protein–ligand interactions [30–33].Indeed, molecular dynamics (MD) simulations
revealed significant differences in loop dynamics
between HjeCel7A and Phanerochaete chrysosporium
Cel7D (PchCel7D), which have the most closed and
most open tunnels among known GH7 CBHs, respec-
tively. Heterobasidion irregulare Cel7A (HirCel7A)
showed intermediate properties [23]. Furthermore,
quantum mechanics/molecular mechanics MD simula-
tions of hydrolysis and cellulose chain threading in the
tunnel of HjeCel7A have enabled the calculation of
the free energy profile along the whole reaction coordi-
nate of the hydrolytic–processive cycle [33].
In this study, we report the identification, sequenc-
ing, biochemical characterization, and crystallization
of GcaCel7A, including five crystal structures of the
enzyme, both in its apo-form and in complex with
oligosaccharides. With the new structures, we also
conducted MD simulations to examine loop dynam-
ics and protein–substrate interactions with both
soluble and insoluble substrates for comparison with
GH7 homologues (HjeCel7A, HirCel7A, and
PchCel7D). Overall, our results highlight molecular-
level features that are important for understanding
this biologically and industrially relevant family of
GHs.
Results
Isolation and identification of GcaCel7A
From a culture of G. candidum 3C grown on filter paper
as the sole carbon source, the major cellulase enzyme
was purified to homogeneity with cellulose affinity, ion
exchange, and hydrophobic interaction chromatograpy.
The yield was 32 mg of purified protein per litre of cul-
ture, with a specific activity against crystalline cellulose
of 2.15 9 10�3 U/mg. SDS/PAGE analysis confirmed
that the enzyme was the most abundant protein in the
culture filtrate. Figure 1 shows the analysis of the puri-
fied protein after papain treatment to remove the linker-
CBM and isolate the catalytic domain (GcaCel7A_CD)
for protein crystallization. The proteolytic cleavage was
not complete. Both the full-length enzyme and the cat-
alytic domain were present, at 75 kDa and 46 kDa,
respectively, but as seen from the gel, they were well sep-
arated from each other. Trypsin digestion and peptide
mapping by MALDI-TOF mass spectrometry (data not
shown) identified the enzyme as a member of GH family
7.
Sequencing of the GcaCel7A-encoding gene
At the time of initial crystallization of GcaCel7A_CD,
neither the gene nor the protein had been sequenced.
Nevertheless, it was possible to solve the structure with
PchCel7D as a search model to build an initial struc-
ture model of GcaCel7A_CD. From this model, partial
amino acid sequence information could be derived for
the search for homologous genes in the NCBI and
MycoCosm databases. Alignment of the nucleotide
sequences of three close homologues (from Thermoascus
aurantiacus, Neosartorya fischeri, and Botryosphaeria
dothidea) revealed two conserved regions, near the
3FEBS Journal (2015) ª 2015 FEBS
A. S. Borisova et al. Characterization of G. candidum Cel7A

beginning and the end of the genes, suitable for primer
design. PCR against genomic DNA extracted from
G. candidum 3C yielded a PCR product of ~ 1200 bp
that was purified and sequenced. Subsequently, gen-
ome sequence data of G. candidum 3C became avail-
able [16], and the full-length gene could be retrieved.
The GcaCel7A gene consists of 1671 bp, including one
intron of 66 bp. The nucleotide sequence of the pro-
tein-encoding region, corresponding to mRNA, has
been deposited in GenBank (accession number
KJ958925), and was confirmed by sequencing of PCR-
amplified cDNA.
The encoded GcaCel7A preprotein consists of 535
amino acids, divided into a 17-residue signal peptide
followed by a GH7 catalytic module of ~ 436 residues,
a Ser/Thr-rich linker region of ~ 47 residues, and
finally a C-terminal CBM1 of ~ 35 residues (Fig. 2).
Three putative N-glycosylation sites and 56 potential
O-glycosylation sites were predicted in GcaCel7A_CD
by the NetNGlyc 1.0 (http://www.cbs.dtu.dk/services/
NetNGlyc/) and NetOGlyc 4.0 (http://www.cbs.dtu.
dk/services/NetOGlyc/ [35]) servers, respectively. The
crystal structures show attached N-glycans at two of
the predicted N-glycosylation sites (Asn57 and
Asn206) and also at Asn432 near the C terminus of
GcaCel7A_CD, whereas no attached sugar is visible at
the Asn98 site in any of the structures. Furthermore,
O-glycosylation at Ser196 is indicated by electron den-
sity for an a-linked sugar, presumably mannose, in
two of the structures. No glycosylation sites on the
catalytic domain are conserved throughout GH7 CBHs
(Fig. 3).
Temperature and pH dependence of GcaCel7A
activity and stability
Figure 4 shows the activity of GcaCel7A on Avicel
cellulose at different pH values and temperatures,
and the residual activity at 37 °C and pH 5.0, after
preincubation at the indicated temperatures (1 h, pH
5.0) and pH values (24 h, 37 °C). The enzyme
showed a broad pH optimum, with the highest activ-
ity at pH 5.0 and within > 85% from pH 4.0 to 6.5.
There was no loss of activity from pH 4.0 to pH 7.0,
and > 80% activity was retained after preincubation
at pH 3.5 and pH 7.5 (Fig. 4A). The temperature
optimum was at 50 °C (Fig. 4B). Loss of activity
after preincubation was detected from 50 °C and
upwards. After 1 h at 60 °C, the residual activity had
decreased to 45%.
GcaCel7A activity on polysaccharides
Specific activities of GcaCel7A against selected
polysaccharide substrates are shown in Table 1. On
the insoluble cellulose substrates, the enzyme showed
the highest activity on amorphous cellulose [phospho-
ric acid swollen cellulose (PASC)], followed by bacte-
rial cellulose (BC) and microcrystalline cellulose
(Avicel). The low activity on carboxymethyl cellulose
(CMC), although it is soluble, is in accordance with
poor accessibility of the restricted substrate-binding
tunnel of CBHs to substituted cellulose. Significant
activity was also detected on lichenan (b-1,3-1,4-glucan)but not on beechwood xylan (data not shown). The
isolated GcaCel7A_CD hydrolysed CMC at the same
rate as full-length GcaCel7A, whereas activity on
Avicel was reduced by half by cleaving the CBM
(1.02 9 103 U�mg�1). The latter finding is consistent
with previous reports indicating that the CBM
enhances hydrolysis of crystalline cellulose under rela-
tively low solid loading conditions [37]. Activity com-
parisons of GcaCel7A and HjeCel7A were performed
with Avicel as substrate, both with the CBH acting
alone and with the addition of a commercial cellulase
cocktail (Accellerase) from which HjeCel7A was selec-
tively removed. In both cases, GcaCel7A yielded simi-
lar amounts of soluble sugar as HjeCel7A, within the
standard error of the experiment (Table 2). Approxi-
mately 1.7–1.8-fold higher conversion was achieved
Fig. 1. SDS/PAGE analysis of fractions from the final size exclusion
chromatography purification step after papain cleavage of
GcaCel7A. Lanes 1–3, full-length GcaCel7A fractions. Lanes 4 and
5: GcaCel7A_CD fractions. Molecular masses in kDa are indicated
for the reference ladder proteins.
4 FEBS Journal (2015) ª 2015 FEBS
Characterization of G. candidum Cel7A A. S. Borisova et al.

with the enzyme cocktail than with either Cel7A acting
alone.
Enzyme kinetics and product inhibition
GcaCel7A_CD was used for the determination of enzyme
kinetic parameters, kcat and KM, on three chromogenic
disaccharide substrates, i.e. 2-chloro-4-nitrophenyl
b-cellobioside (CNP-G2), p-nitrophenyl b-cellobioside(pNP-G2), and p-nitrophenyl b-lactoside (pNP-Lac),
and studies of product inhibition by cellobiose. The
results are shown in Table 3 along with a comparison
of previously published data on pNP-Lac for
HjeCel7A and PchCel7D [38]. The kinetic parameters
N-glycosylation site (98...101)N-glycosylation site (57...60)
O-glycosylation site (196)N-glycosylation site (206...209)
Linker
CBM1 (484...518)Signal peptide (–17...–1) Catalytic region (1...436)
Fig. 2. Schematic representation of the amino acid sequence encoded by the GcaCel7A gene. Predicted N-glycosylation sites, two of which
were observed, and the observed O-glycosylation site are indicated by triangles. The picture was generated with the Vector NTI program by
using services of the SignalP 4.1 server (http://www.cbs.dtu.dk/services/SignalP/ [34]).
Fig. 3. Structure-based sequence alignment of the GH7 catalytic domains of GcaCel7A (PDB code 5AMP), RemCel7A (PDB code 1Q9H),
HjeCel7A (PDB code 1CEL), Trichoderma harzianum Cel7A (ThaCel7A; PDB code 2YOK), and HirCel7A (PDB code 2YG1). Secondary
structural elements of the GcaCel7A structure are indicated above the alignment (b-strand arrows and a-helices). Strictly identical residues
are marked in white letters on a black background. Regions of conserved, highly similar residues are framed in thin-lined boxes with bold
letters. Red frames indicate loop regions of interest, with loop nomenclature underneath. The green triangle indicates the O-glycosylated
Ser196, and blue triangles indicate the N-glycosylated Asn residues observed in GcaCel7A structures. The figure was prepared with the
ESPript web server with default parameters (http://espript.ibcp.fr [36]).
5FEBS Journal (2015) ª 2015 FEBS
A. S. Borisova et al. Characterization of G. candidum Cel7A

for GcaCel7A_CD on the cellobioside substrates are
practically identical. Thus, the 2-chloro substitution on
the nitrophenyl group of CNP-G2 is accommodated in
subsite +1 without influencing the activity. On pNP-
Lac, kcat is almost twice as high, and kcat/KM is higher,
than with CNP-G2 and pNP-G2. GcaCel7A_CD
shows an approximately two-fold higher kcat on pNP-
Lac than HjeCel7A and PchCel7D [38]. At the same
time, substrate binding is weaker (higher KM) and
catalytic efficiency (kcat/KM) is slightly lower for
GcaCel7A_CD than for HjeCel7A, whereas the oppo-
site is true relative to PchCel7D. Inhibition constants
indicate approximately two-fold weaker binding of cel-
lobiose to GcaCel7A_CD than to HjeCel7A.
GcaCel7A_CD crystal structures
The isolated GcaCel7A_CD was successfully crystal-
lized, and we report five crystal structures herein: two
apo structures (referred to as APO1 and APO2), and
three ligand complexes, with cellobioside (G2), cel-
lotrioside (G3), and cellotetraoside (G4) bound at the
active site. X-ray diffraction data and refinement
statistics are summarized in Table 4. The initial struc-
ture model of GcaCel7A was solved by molecular
replacement with the APO1 dataset and a structure of
PchCel7D as the search model [Protein Data Bank
(PDB) code 1GPI] [25]. All of the structures were
solved in the monoclinic space group P21 with one
protein chain per asymmetric unit, but two discrete
crystal packings differing by the length of the b-axis
were represented. To distinguish between those forms,
we use the terms ‘long’ and ‘short’ for the b-axis. The
APO1 and G4 structures have long b-axes (90.5 �A);
whereas, the APO2, G2 and G3 structures have short
b-axes (81–82 �A). APO1 was refined at 2.12 �A, and
the other structures at higher resolution (1.37–1.56 �A).
Figure 5 shows the protein backbones of all of the
GcaCel7A structures superimposed with HjeCel7A.
In both of the apo structures, all residues of the pro-
tein, 1–438, are visible in the electron density and are
Fig. 4. Effects of pH and temperature on the activity of GcaCel7A
on Avicel cellulose. (A) Temperature dependence. (B) pH
dependence. Filled circles show relative activities in terms of
released reducing sugar during 2 h of incubation of enzyme with
5 mg�mL�1 Avicel at the indicated temperatures (at pH 5.0) and pH
values (at 37 °C). Open circles show the residual activity measured
at pH 5.0 and 37 °C after preincubation at different temperatures
for 1 h at pH 5.0, and after preincubation at different pH values for
24 h at 37 °C. Error bars indicate standard errors of at least three
measurements.
Table 1. Specific activity of full-length GcaCel7A on polysaccharides. Released reducing sugar was assayed after incubation of 5 mg�mL�1
substrate with GcaCel7A at pH 5.0 and 37 °C. PASC and BC were incubated for 1 h with 0.075 mg�mL�1 and 0.5 mg�mL�1 enzyme,
respectively, and the other substrates were incubated for 20 h with 0.5 mg�mL�1 enzyme.
Substrate Avicel CMC BC PASC Lichenan
Enzyme activity (U�mg�1 9 103)a 2.15 � 0.13 1.86 � 0.11 40.0 � 2.4 275 � 17 1.91 � 0.11
a The specific activity, U�mg�1, is defined as the accumulated amount in lmol of glucose equivalents released per mg of enzyme divided by
the incubation time in minutes under the assay conditions.
6 FEBS Journal (2015) ª 2015 FEBS
Characterization of G. candidum Cel7A A. S. Borisova et al.
included in the structure models. The tip of loop B2 is
disordered in the G2 and G3 structures, and residues
199–201 have been excluded from the G2 structure
model, and residues 196–199 from the G3 structural
model, owing to insufficient electron density. In the
G4 structure, the density is weak and ambiguous for
Ser21, Gly22, and Gly23, indicating multiple confor-
mations. As compared with the other GcaCel7A_CD
structures and with Cel7 homologues, the APO1 struc-
ture seemed to fit best to the electron density in this
region of the G4 structure. Thus, the conformation of
these residues were taken from APO1 and were manu-
ally adjusted, refined, and included in the G4 structure
model. The terminal residues Gly437 and Thr438 are
not visible in the G4 structure; whereas, the other
structures clearly show Thr438 as the C-terminal resi-
due. In all structures, the N-terminal Gln is cyclized to
pyroglutamate, and all 16 Cys residues form disulfide
bonds. N-glycosylation is evident at three sites in all
five GcaCel7A_CD structures, with GlcNAc attached
to Asn57, Asn206, and Asn432. Interestingly, two con-
secutive GlcNAc residues are visible at Asn206, despite
deglycosylation treatment of the protein, which indi-
cates that endo-b-N-acetylglucosaminidase may be
unable to cleave the N-glycan at this site. Further-
more, there is convincing density for O-glycosylation
at Ser196 in the APO2 and G2 structures, where an a-linked mannose residue was refined. Positive density in
the difference map (Fo–Fc) of the G4 structure also
supports O-glycosylation at Ser196 but is not clear
enough for confident positioning of mannose. No
sugar is visible at this position in APO1 or in the G3
structure, where the entire Ser196 residue is missing.
As shown in Fig. 6, there is distinct electron density
for cello-oligosaccharides bound in the active site of
the GcaCel7A_CD ligand complexes. The G2 structure
was obtained by cocrystallization with laminaribiose,
and a disaccharide clearly occupies the product sub-
sites +1 and +2. Surprisingly, the density is not com-
patible with the b-1,3 linkage of laminaribiose, but
unambiguously shows that the bound sugar is cel-
lobiose (Fig. 6A). In the G3 complex, two thio-cel-
lotriose molecules are visible and included in the
structure model, in subsites �4/�3/�2 and �1/+1/+2,respectively (Fig. 6B). For the G4 structure, thio-cel-
lobiose was present in the cocrystallization and is visi-
ble at subsites +1 and +2. There is also contiguous
density for glucose residues all the way from subsite
�5 to subsite �1, indicating overlapping binding
modes (Fig. 6C). Consecutive glucose units connect
with proper thio-glycosidic bond geometry, which is
why we chose to refine two overlapping thio-cellote-
traose molecules at partial occupancy, spanning sub-
sites �5/�4/�3/�2 and �4/�3/�2/�1, respectively.
All glucosyl units are well defined except in subsite
�1, where the density is weaker and indicates the
presence of multiple conformations. The predominant
�1 conformer appears to be the a-anomer of a4C1 glucopyranosyl unit at partial occupancy. The
APO1 structure also shows a long stretch of contigu-
Table 2. Comparison of hydrolysis of 5 mg�mL�1 Avicel cellulose by GcaCel7A and HjeCel7A, acting alone (50 lg�mL�1) or added to a
Cel7A-depleted Hypocrea jecorina enzyme cocktail (25 lg�mL�1 Cel7A + 25 lg�mL�1 HjeCel7A-free Accellerase 1500), for 2 h at 40 °C and
pH 5.0. Excess b-glucosidase was subsequently added to convert all soluble sugars to glucose prior to the reducing sugar assay.
Enzyme
Cel7A alone Cel7A + HjeCel7A-free Accellerase
[Glc] (mg�mL�1)a Conversion (%) [Glc] (mg�mL�1)a Conversion (%)
GcaCel7A 0.39 � 0.021 6.9 0.65 � 0.072 11.7
HjeCel7A 0.42 � 0.025 7.6 0.75 � 0.039 13.5
a Average and standard deviation of five replicates.
Table 3. Enzyme kinetics parameters for GcaCel7A_CD on chromogenic disaccharide substrates and inhibition constants for cellobiose.
Enzyme kinetic parameters were derived by nonlinear regression from experiments performed at pH 5.0 and 30 °C with 0.01–5 mM
substrate. Previously published data for HjeCel7A and PchCel7D are provided here for comparison.
Enzyme Substrate kcat (s�1) KM (mM) kcat/KM (M�1 s�1) Ki (lM)
GcaCel7A_CD CNP-G2 0.11 � 0.02 0.84 � 0.12 131 –
GcaCel7A_CD pNP-G2 0.10 � 0.02 0.85 � 0.12 118 –
GcaCel7A_CD pNP-Lac 0.19 � 0.06 1.07 � 0.10 178 50 � 5
HjeCel7Aa pNP-Lac 0.093 0.41 226 24
PchCel7Da pNP-Lac 0.10 1.3 76 180
a Published data obtained at pH 5.0 and 30 °C [38]. Reported standard errors were 7–15% for KM and 3–6% for kcat.
7FEBS Journal (2015) ª 2015 FEBS
A. S. Borisova et al. Characterization of G. candidum Cel7A

ous density in the active site, probably representing
bound poly(ethylene glycol) (PEG) and/or glycerol.
The APO2 structure was obtained by crystal soaking
with xyloglucan oligosaccharide (XGO). Cocrystal-
lization with XGO was also performed in an attempt
to trap an ‘open-loop’ structure of the enzyme,
which would be required in order to accommodate
this branched and bulky sugar. However, no bound
sugars were seen in any structures from XGO-ex-
posed crystals.
Structure of GcaCel7A and comparison with
related enzymes
Overall, the GcaCel7A structures are very similar, as
reflected by low pairwise rmsd values (0.16–0.35 �A).
Notable differences occur at loop B2, which seems to
be flexible and shows elevated temperature factors,
and local disorder (the G2 and G3 structures), as
observed in other Cel7 CBHs [18,23]. In the APO2
structure, loop B2 adopts a new conformation not
previously seen in any Cel7 structure. The loop bends
further inwards and partially obstructs the cellulose-
binding path through the tunnel. As compared with
the APO1 structure, Ala199 CB at the tip of loop B2
has moved 10.7 �A into the tunnel, causing it to clash
with a glucosyl unit binding at subsite �4 (Fig. 7A).
This B2 loop conformation correlates with the short
unit cell b-axis and may be a crystallization artefact
caused by tight crystal packing in this region. The
crystal contacts also involve the O-glycosylation at
Ser196, which explains why the attached mannose
residue is most clearly revealed in the APO2 and G2
structures. A more relaxed B2 loop conformation,
Table 4. Diffraction data and refinement statistics for the five determined GcaCel7A_CD structures. The space group is P21 in all cases,
with one protein chain per asymmetric unit.
Structure Apo structure (APO1)
Apo structure
(APO2)
Cellobioside
complex (G2)
Cellotrioside
complex (G3)
Cellotetraoside
complex (G4)
PDB code 5AMP 4ZZV 4ZZW 4ZZT 4ZZU
b-axis Long Short Short Short Long
Unit cell, a, b,
c (�A); b-angle (°)
42.88, 90.56, 55.27;
109.86
42.42, 80.73,
54.91; 109.54
42.64, 81.82,
55.03; 109.63
42.47, 80.76,
55.06; 109.6
42.71, 90.52,
55.09; 109.62
Beamline I911-2, MAX-lab ID23-1, ESRF I911-3, MAX-lab ID23-1, ESRF I911-3, MAX-lab
Wavelength (�A) 1.0409 0.9918 1.0001 0.9918 0.9000
No. of observations 84 009 258 951 140 868 187 944 217 429
No. of unique reflections 21 970 68 772 54 088 48 793 70 198
Resolution range (�A) 30.12–2.12
(2.17–2.12)a43.57–1.37
(1.39–1.37)
24.9–1.50
(1.53–1.50)
51.87–1.56
(1.59–1.56)
27.62–1.44
(1.47–1.44)
Multiplicity 3.7 (2.6) 3.8 (3.8) 2.6 (2.6) 3.9 (4.0) 3.1 (2.5)
Completeness (%) 97.9 (75.1) 93.7 (92.4) 97.2 (95.7) 97.8 (97.8) 99.4 (97.9)
Rmerge (%)b 5.9 (16.9) 7.8 (37.6) 5.7 (36.8) 6.3 (22.6) 5.1 (22.4)
Mean [I/SD(I)] 15.3 (6.9) 7.3 (2.6) 7.1 (1.8) 10.5 (4.0) 13.7 (4.4)
CC1/2 99.6 (92.4) 99.5 (78.3) 99.2 (73.7) 97.7 (94.9) 99.7 (89.7)
Rwork/Rfree (%) 15.7/19.5 18.0/20.3 17.2/19.5 16.9/19.5 16.3/17.8
RMSD, bond lengths (�A) 0.0041 0.0045 0.0046 0.0047 0.0043
RMSD, bond angles (o) 0.97 1.06 1.09 1.14 1.12
Protein atoms: no.,
average B-factor (�A2)
3889, 14.0 4011, 14.0 3981, 13.0 3963, 14.0 4252, 11.0
Water molecules: no.,
average B-factor (�A2)
467, 21.96 530, 23.94 369, 24.0 394, 25.64 609, 21.20
Sugar ligands: no. of residues,
average B-factor (�A2)
0 0 2, 21.22 6, 29.93 6, 15.56
Glycosylation: no. of residues,
average B-factor (�A2)
4, 29.45 5, 22.73 5, 25.40 5, 26.54 4, 15.33
Other heteroatoms Glycerol 92 Mg2+ Glycerol 93; PEG 92;
Mg2+ 92
Mg2+ Glycerol
Ramachandran plot
outliersc, no. of residues
0 1 1 2 1
a Data within parentheses are for the outermost resolution shell.bRmerge = Σhkl Σi | Ii(hkl) � <I(hkl)> | Σhkl Σi Ii(hkl).c Calculated by use of a strict boundary Ramachandan plot [39].
8 FEBS Journal (2015) ª 2015 FEBS
Characterization of G. candidum Cel7A A. S. Borisova et al.

similar to that of HjeCel7A, is seen in the two
structures with long b-axes, i.e. APO1 and G4 (Figs 5
and 7).
The fold of GcaCel7A is very similar to those in
other Cel7 CBH structures (rmsd of < 0.8 �A), as
expected from high sequence identities [Rasamsonia
emersonii Cel7A (RemCel7A), 71%; HjeCel7A 64%;
Trichoderma harzianum Cel17A, 64%]. rmsd values
and sequence identities for further homologues are
provided in Table S1. The cellulose-binding active site
is highly conserved, including the four Trp platforms
at subsites �7, �4, �2 and +1 (Trp40, Trp38, Trp371,
and Trp380), and the catalytic triad Glu212 (nucle-
ophile), Asp214, and Glu217 (acid/base), having the
same residue numbers as in HjeCel7A. In GcaCel7A,
Leu213 replaces the Met that is conserved in the
nearest Cel7 homologues (Fig. 3).
At the B2/A3/B3 loop contact region, two Tyr resi-
dues at the tip of loop A3 play an important role in
tunnel-enclosing contacts with loops B2 and B3
across the active site (Tyr374 and Tyr375 in
GcaCel7A; Tyr370 and Tyr371 in HjeCel7A). In
GcaCel7A, the side chain of Tyr375 is flipped relative
to the predominant position of HjeCel7A Tyr371
above subsite �1, towards the +1 and +2 product
sites in a similar position as in the HjeCel7A struc-
tures 1CEL and 4P1J. There is also a shift outwards
of loop B3 and Tyr247 at its tip (Fig. 7). Loops A3
and B3 are practically identical in GcaCel7A and
HjeCel7A, whereas differences in loop B2 exist. At
the tip of loop B2, Asn198 in HjeCel7A, which inter-
acts with the �3 glucosyl unit, is replaced by the
smaller Ser198 in GcaCel7A, leading to the loss of
one direct protein–sugar interaction. Further size
reduction, of Ser195 and Thr201 in HjeCel7A to
Ala195 and Ser201 in GcaCel7A, reduces the contact
surface within the loop, with substrate, and with
the rest of the protein, suggesting that loop B2 would
be more flexible in GcaCel7A than in HjeCel7A.
The loop dynamics may also be influenced by the
O-glycosylation at Ser196 in GcaCel7A. To our
knowledge, there are no reports of O-glycosylation at
this position on any other Cel7 enzyme.
Superposition of the GcaCel7A ligand complexes
with the HjeCel7A Michaelis complex (4C4C; [40]),
shows very similar sugar binding at subsite �2, with a
gradual increase in deviation towards the entrance of
the tunnel (Fig. 8). At subsite �1, two binding modes
are observed. In the G4 complex, the �1 glucosyl is
connected to the �2 unit, and it binds in a similar
position as in HjeCel7A. However, the �1 glucosyl of
GcaCel7A is in the form of the a-anomer and adopts a4C1 chair conformation. In the G3 structure, on the
other hand, the �1 residue is connected to the +1 glu-
cosyl unit and is located further away from the cat-
alytic residues, illustrating that the �1 subsite is rather
spacious. At the product sites +1 and +2, the so-called
‘primed’ binding mode is observed in all of the
GcaCel7A complexes, in contrast with the ‘unprimed’
position in the HjeCel7A Michaelis complex. After
cleavage of the cellulose chain, the cellobiose product
can move away from the catalytic centre by pivoting
around OH6 of the +2 glucosyl unit [40,41]. In
GcaCel7A, the ‘primed’ mode is stabilized by Asp343
in loop B4, at hydrogen bonding distance to OH1 of
the reducing-end +2 glucose unit. Most GH7 CBHs
have an Asp at this position, but in H. jecorina
and Trichoderma species, it is missing owing to a one-
residue deletion in loop B4 [41].
MD
To investigate the significance of structural differences,
we conducted MD simulations of the GcaCel7A and
HjeCel7A catalytic domains in solution without a
bound ligand, in solution bound to a cellononaose
ligand, and complexed with the surface of a cellulose
microfibril (Fig. 9). Simulations of the enzymes in each
representative state enable us to directly examine the
effects of substrate on protein dynamics and to com-
pare relative endo-initiation abilities.
The relative degree of protein flexibility in response
to the environment over the course of MD simulation
provides a means to estimate similarity in dynamic
behaviour. Root-mean-square fluctuation (RMSF) of
Fig. 5. Overall structural alignment of the five GcaCel7A structures
with the HjeCel7A cellononaose complex (PDB code 4C4C [40]).
HjeCel7A protein and cellononaose ligand are coloured in light grey
with labelled loops in red. The GcaCel7A structures are coloured as
follows: APO1, orange; APO2, pale green; G2, yellow; G3, pink;
G4, cyan. Ligands in the GcaCel7A structures are not shown.
9FEBS Journal (2015) ª 2015 FEBS
A. S. Borisova et al. Characterization of G. candidum Cel7A

the protein backbone was calculated to evaluate the
flexibility of these two GH7s under the three different
substrate scenarios (Fig. 10A,C). The RMSF of
GcaCel7A and HjeCel7A without a ligand is visually
illustrated on the protein backbones in Fig. 10(B) and
Fig. 10(D), respectively.
Fig. 6. Electron density for the ligands in the GcaCel7A G2, G3, and G4 structures. (A) The G2 structure shows cellobiose bound in the
product-binding subsites +1 and +2. (B) The G3 complex shows two molecules of thio-cellotriose, at subsites �4/�3/�2 and �1/+1/+2,
respectively. (C) In the G4 complex, overlapping thio-cellotetraose molecules at partial occupancy were refined at subsites �5 to �2 and �4
to �1, respectively, and thio-cellobiose at subsites +1 and +2. At subsite �1, the density indicates the presence of multiple binding modes.
All glucosyl units adopt the 4C1 chair conformation. Sigma-averaged 2Fo�Fc electron density maps are contoured at 0.53 e/A3 in (A) and at
0.26 e/A3 in (B) and (C).
Fig. 7. Tunnel-enclosing loop contacts. (A) In the GcaCel7A APO2 structure (green), loop B2 bends into the tunnel, causing Ala199 at the tip
of the loop to clash with a glucosyl binding at subsite �4. (B) The B2/A3/B3 loop contacts in the GcaCel7A APO1, G3, and G4 structures.
(C) B2/A3/B3 loop contacts in HjeCel7A (G9 complex; PDB code 4C4C [40]). Two Tyr residues at the tip of loop A3 play an important role in
tunnel-enclosing contacts with loops B2 and B3 across the active site. In GcaCel7A, the Tyr375 side chain is flipped towards the +1 and +2
product sites, relative to the position of Tyr371 in HjeCel7A above subsite –1, accompanied by a slight shift outwards of loop B3 and Tyr247
at its tip. The colouring scheme is the same as in Fig. 5: HjeCel7A protein and cellononaose ligand, light grey with loops in red; GcaCel7A
APO1, orange; APO2, green; G2, yellow; G3, magenta; G4, cyan.
10 FEBS Journal (2015) ª 2015 FEBS
Characterization of G. candidum Cel7A A. S. Borisova et al.

As one might expect, the corresponding loop regions
in GcaCel7A and HjeCel7A show higher fluctuations
than the b-sandwich core of the proteins (Fig. 10A,C).
For HjeCel7A, the overall dynamics are dampened
with either bound cellulosic substrate as compared
with the ligand-free enzyme (Fig. 10C). The most sta-
bilizing effect is seen with the microfibril-bound
enzyme, in particular in the regions near the entrance
to the tunnel, including loops A1 and B1, which are in
direct contact with the microfibril surface. GcaCel7A
behaves differently, in the sense that there is little over-
all difference in fluctuations between the three scenar-
ios except for loops A2 and A4, which are more
flexible in the ligand-free enzyme than with substrate
bound in the active site (Fig. 10A). Surprisingly, there
is no significant stabilization of the microfibril-bound
GcaCel7A as compared with the cellononaose-bound
enzyme. One could reasonably expect that the presence
of a solid, crystalline substrate underneath the active
site loops would contribute additional stability, at the
very least on a localized basis. However, MD simula-
tions clearly indicate that this is not the case for
GcaCel7A; the RMSFs of the two substrate-bound
catalytic domains are almost indistinguishable
(Fig. 10A).
HjeCel7A shows a greater degree of flexibility in the
A4 and B4 loop regions at the product side of the
active site tunnel, as illustrated by multiple conforma-
tions and red regions in the rightmost loops in the
RMSF-coloured snapshot (Fig. 10D). The higher
stability of loop B4 in GcaCel7A (Fig. 10A) is attribu-
table to the insertion of Asp343, which forms a salt
bridge with Arg267. Also, an additional hydrogen
bond is provided by Lys338 in GcaCel7A (replacing
Glu335 in HjeCel7A). Both interactions will anchor
the loop more firmly to the rest of the protein. The
difference in behaviour of loop A4 between the two
enzymes is more difficult to rationalize. In GcaCel7A,
loop A4 is clearly stabilized by substrate binding
(Fig. 10A), as expected from the increase in local van
der Waals and hydrogen bond interaction opportuni-
ties when the product sites are occupied. However,
similar stabilization is not seen in HjeCel7A. The
sequences differ at 5 of 10 positions in loop A4, but
no obvious determinants of flexibility/stability can be
identified.
In a previous study, we investigated the structure
and dynamics of Cel7A from the tree pathogen Heter-
obasidion irregulare (HirCel7A [23]). Our investigation
revealed that the HirCel7A active site shows an ‘inter-
mediate’ degree of openness and flexibility relative to
PchCel7D and HjeCel7A (Fig. 11). We further suggest
that this degree of closure, resulting from active site
loop formation, impacts on processive ability, degree
Fig. 8. Oligosaccharide binding in the GcaCel7A complex structures in comparison with the HjeCel7A cellononaose complex (PDB code
4C4C; white). In (B), the view is rotated ~ 90 ° around the x-axis. The GcaCel7A G2 structure (yellow) contains cellobiose at subsites +1 and
+2, and the G3 complex (magenta) contains two 4,40-dithiocellotriose molecules at subsites �4/�3/�2 and �1/+1/+2, respectively. In the G4
structure (blue), 4-thiocellobiose was refined at subsites +1 and +2, and two overlapping molecules of 4,40,40 0 0-trithiocellotetraose in subsites
�5/�4/�3/�2 and �4/�3/�2/�1 at partial occupancy. Oxygen atoms are red, and sulfur atoms from thio-linkages are in yellow. Numbers
indicate the glucosyl-binding subsites.
11FEBS Journal (2015) ª 2015 FEBS
A. S. Borisova et al. Characterization of G. candidum Cel7A

of endo-initiation, and substrate dissociation. Compar-
ison of the flexibility of GcaCel7A with that of
HjeCel7A, HirCel7A, and PchCel7D indicates that
GcaCel7A is even more rigid than HjeCel7A. Nearly
all of the GcaCel7A primary active site loops fluctuate
less than those of the other three GH7s, which may
translate to higher ligand-binding free energy and a
higher degree of processive ability [42,43].
MD simulations also suggest that GcaCel7A and
HjeCel7A show a similarly low degree of endo-initi-
ated processive action in comparison with PchCel7D
and HirCel7A. In the literature, loop B3 has been ter-
med the exo-loop, as it is beneficial in facilitating pro-
cessive crystalline substrate degradation [38]. The
ability to conduct endo-initiated attack of crystalline
substrates is thought to be related to both the flexibil-
ity and the length of this loop, along with that of the
nearby loop B2. Both of these loops must open suffi-
ciently to allow the entry into the active site of an
internal part of a cellulose chain. PchCel7D shows the
shortest B2 and B3 loops of all four GH7s discussed
here and is known to conduct endo-initiated attack
Fig. 9. MD simulations of the behaviour of the GcaCel7A and HjeCel7A catalytic domains in solution without a ligand, bound to a
cellononaose ligand in solution, and bound to a portion of a cellulose Ib microfibril.
Fig. 10. Comparison of RMSFs of GcaCel7A and HjeCel7A. (A) The RMSF of each GcaCel7A residue over a 100-ns MD simulation, in
solution without a ligand, bound to a cellononaose ligand, and catalytically engaged with a cellulose Ib microfibril. Key active site loops are
labelled on the plot. (B) Twenty aligned snapshots of GcaCel7A from the ‘no ligand’ MD simulation are shown coloured by RMSF. Red
indicates regions of higher fluctuation, and blue indicates regions of lower fluctuation. White regions are intermediate. (C) The RMSF of
each HjeCel7A residue over the three 100-ns MD simulations. (D) Twenty aligned snapshots of the HjeCel7A ‘no ligand’ simulation shown
coloured by RMSF. The colour scales of (B) and (D) are identical from 0 to 3.3, the maximum value of the RMSF from the GcaCel7A ‘no
ligand’ simulation.
12 FEBS Journal (2015) ª 2015 FEBS
Characterization of G. candidum Cel7A A. S. Borisova et al.

more frequently than HjeCel7A [44]. Using this infor-
mation, we extrapolated substrate initiation modes for
GcaCel7A and the previously described HirCel7A [23],
based on evidence from MD simulation. We calculated
the minimum distances of nearby active site loops to
characterize the openness and movement of these
loops. The minimum distances were binned into his-
tograms to determine the probability that the loops
would be in a given position relative to each other.
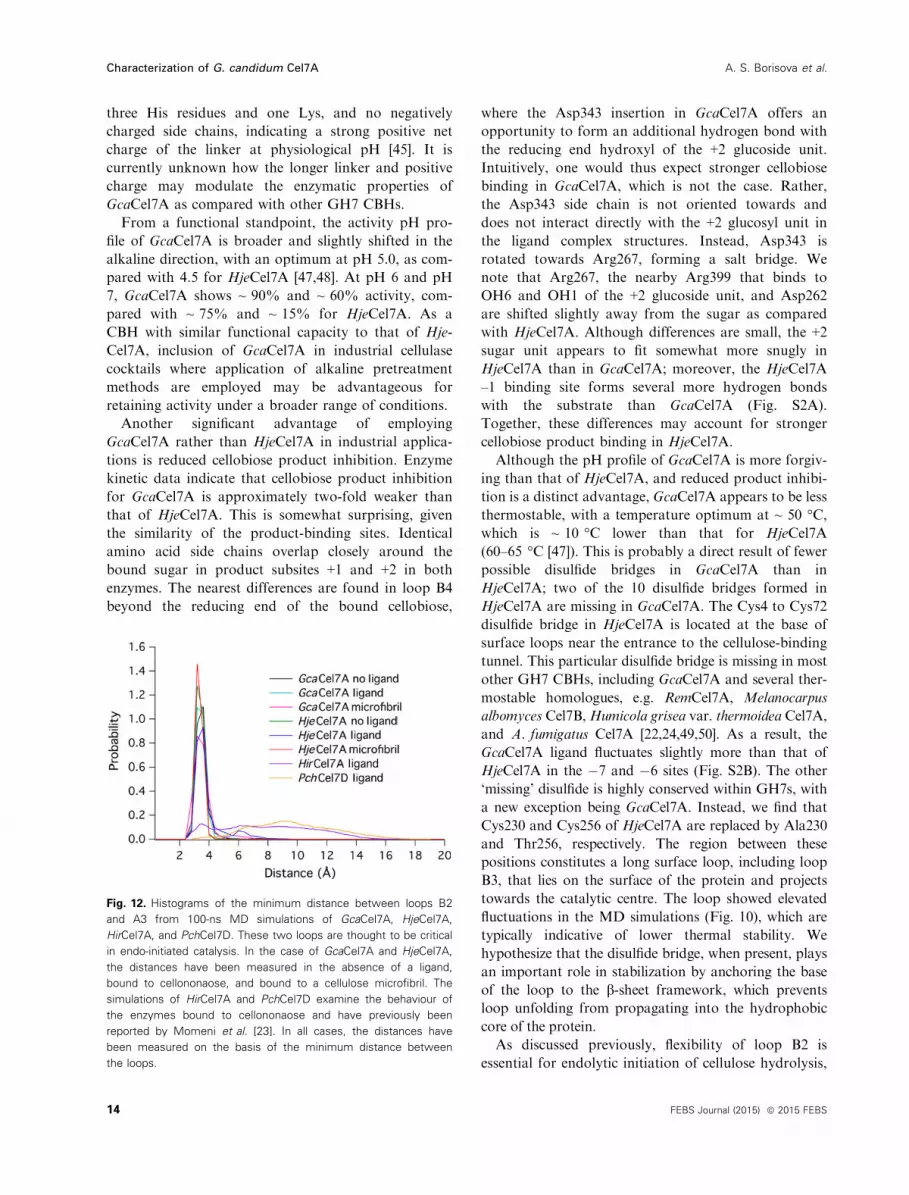
Examination of the minimum distance between loops
B2 and A3 indicate that GH7s performing primarily
endo-initiated attack have a broad range of conforma-
tions relative to each other (Fig. 12) and include
HirCel7A and PchCel7D [23]. Conversely, loops B2
and A3 of exo-initiating HjeCel7A seldom stray from
each other, maintaining a general minimum distance
of ~ 3.5 �A (Fig. 12). On the basis of these data, we
suggest that GcaCel7A also conducts exo-initiated
attack on crystalline cellulose substrates. As before,
this observed behaviour is the same regardless of the
bound cellulosic substrate. The minimum distances
between loops A1 and B2 (Fig. S3A) and between
loops B2 and B3 (Fig. S3B) in GcaCel7A and
HjeCel7A are greater than that between loops B2 and
A3, but demonstrate the same relative behaviour.
Discussion
We have found that GcaCel7A is biochemically and
structurally similar to the well-characterized HjeCel7A
and shows comparable activity on crystalline cellulose.
Slight differences between the two do exist, however.
In terms of modularity, the linker peptide connecting
the catalytic domain with the CBM in GcaCel7A is
significantly longer than that in HjeCel7A. In fact, the
47-residue linker of GcaCel7A is longer than nearly all
other fungal GH7 linker peptides. In 2012, Sammond
et al. reported the analyses of available sequence data
for linkers in bimodular GH6 and GH7 CBHs and
found an average length of 30 residues for GH7-
CBM1 linkers (42 residues for GH6-CBM1 linkers)
[45]. The GcaCel7A linker contains 18 Thr and 13 Ser
residues, which together make up more than two-
thirds of the amino acids in the linker. The Ser and
Thr residues are distributed along the entire length of
the linker region, suggesting that the whole peptide
may be O-glycosylated and able to extend the physical
distance between the CBM1 and the catalytic module
by ~ 50% as compared with the average GH7-CBM1
linker [46]. The GcaCel7A linker also shows an unusu-
ally large number of positively charged residues, i.e.
Fig. 11. Snapshots of GcaCel7A, HjeCel7A, HirCel7A, and
PchCel7D from MD simulations of the four enzymes bound to
cellononaose ligands. The aligned snapshots, 20 in each figure, are
shown coloured by RMSF calculated from the simulation. Red
regions indicate a high degree of fluctuation, and blue regions
indicate relatively little fluctuation. The colour scale in this figure is
from 0 to 6, where 6 is the highest RMSF observed for any
residue of the four enzymes during simulation [23]. The HirCel7A
and PchCel7D figures are based on data from previously published
simulations described here for comparison with GcaCel7A
behaviour [23].
13FEBS Journal (2015) ª 2015 FEBS
A. S. Borisova et al. Characterization of G. candidum Cel7A
three His residues and one Lys, and no negatively
charged side chains, indicating a strong positive net
charge of the linker at physiological pH [45]. It is
currently unknown how the longer linker and positive
charge may modulate the enzymatic properties of
GcaCel7A as compared with other GH7 CBHs.
From a functional standpoint, the activity pH pro-
file of GcaCel7A is broader and slightly shifted in the
alkaline direction, with an optimum at pH 5.0, as com-
pared with 4.5 for HjeCel7A [47,48]. At pH 6 and pH
7, GcaCel7A shows ~ 90% and ~ 60% activity, com-
pared with ~ 75% and ~ 15% for HjeCel7A. As a
CBH with similar functional capacity to that of Hje-
Cel7A, inclusion of GcaCel7A in industrial cellulase
cocktails where application of alkaline pretreatment
methods are employed may be advantageous for
retaining activity under a broader range of conditions.
Another significant advantage of employing
GcaCel7A rather than HjeCel7A in industrial applica-
tions is reduced cellobiose product inhibition. Enzyme
kinetic data indicate that cellobiose product inhibition
for GcaCel7A is approximately two-fold weaker than
that of HjeCel7A. This is somewhat surprising, given
the similarity of the product-binding sites. Identical
amino acid side chains overlap closely around the
bound sugar in product subsites +1 and +2 in both
enzymes. The nearest differences are found in loop B4
beyond the reducing end of the bound cellobiose,
where the Asp343 insertion in GcaCel7A offers an
opportunity to form an additional hydrogen bond with
the reducing end hydroxyl of the +2 glucoside unit.
Intuitively, one would thus expect stronger cellobiose
binding in GcaCel7A, which is not the case. Rather,
the Asp343 side chain is not oriented towards and
does not interact directly with the +2 glucosyl unit in
the ligand complex structures. Instead, Asp343 is
rotated towards Arg267, forming a salt bridge. We
note that Arg267, the nearby Arg399 that binds to
OH6 and OH1 of the +2 glucoside unit, and Asp262
are shifted slightly away from the sugar as compared
with HjeCel7A. Although differences are small, the +2sugar unit appears to fit somewhat more snugly in
HjeCel7A than in GcaCel7A; moreover, the HjeCel7A
–1 binding site forms several more hydrogen bonds
with the substrate than GcaCel7A (Fig. S2A).
Together, these differences may account for stronger
cellobiose product binding in HjeCel7A.
Although the pH profile of GcaCel7A is more forgiv-
ing than that of HjeCel7A, and reduced product inhibi-
tion is a distinct advantage, GcaCel7A appears to be less
thermostable, with a temperature optimum at ~ 50 °C,which is ~ 10 °C lower than that for HjeCel7A
(60–65 °C [47]). This is probably a direct result of fewer
possible disulfide bridges in GcaCel7A than in
HjeCel7A; two of the 10 disulfide bridges formed in
HjeCel7A are missing in GcaCel7A. The Cys4 to Cys72
disulfide bridge in HjeCel7A is located at the base of
surface loops near the entrance to the cellulose-binding
tunnel. This particular disulfide bridge is missing in most
other GH7 CBHs, including GcaCel7A and several ther-
mostable homologues, e.g. RemCel7A, Melanocarpus
albomyces Cel7B,Humicola grisea var. thermoidea Cel7A,
and A. fumigatus Cel7A [22,24,49,50]. As a result, the
GcaCel7A ligand fluctuates slightly more than that of
HjeCel7A in the �7 and �6 sites (Fig. S2B). The other
‘missing’ disulfide is highly conserved within GH7s, with
a new exception being GcaCel7A. Instead, we find that
Cys230 and Cys256 of HjeCel7A are replaced by Ala230
and Thr256, respectively. The region between these
positions constitutes a long surface loop, including loop
B3, that lies on the surface of the protein and projects
towards the catalytic centre. The loop showed elevated
fluctuations in the MD simulations (Fig. 10), which are
typically indicative of lower thermal stability. We
hypothesize that the disulfide bridge, when present, plays
an important role in stabilization by anchoring the base
of the loop to the b-sheet framework, which prevents
loop unfolding from propagating into the hydrophobic
core of the protein.
As discussed previously, flexibility of loop B2 is
essential for endolytic initiation of cellulose hydrolysis,
Fig. 12. Histograms of the minimum distance between loops B2
and A3 from 100-ns MD simulations of GcaCel7A, HjeCel7A,
HirCel7A, and PchCel7D. These two loops are thought to be critical
in endo-initiated catalysis. In the case of GcaCel7A and HjeCel7A,
the distances have been measured in the absence of a ligand,
bound to cellononaose, and bound to a cellulose microfibril. The
simulations of HirCel7A and PchCel7D examine the behaviour of
the enzymes bound to cellononaose and have previously been
reported by Momeni et al. [23]. In all cases, the distances have
been measured on the basis of the minimum distance between
the loops.
14 FEBS Journal (2015) ª 2015 FEBS
Characterization of G. candidum Cel7A A. S. Borisova et al.

and is also likely to affect processivity and enzyme dis-
sociation [23]. Loop B2 constitutes a 13–15-residueinsertion in CBHs relative to GH7 endoglucanases and
folds over the b-sandwich core to define the tunnel
around subsite –4. Residues at the tip of the loop
interact with the opposing loop A3 across the active
site, which effectively closes the ‘roof’ of the tunnel.
Thus, loop B2 must open to allow the enzyme to
access an internal unit of a cellulose chain. Several
indications from the crystal structure analysis suggest
that loop B2 is more flexible in GcaCel7A than in
HjeCel7A, such as elevated temperature factors, partial
disorder of the loop in some structures, and fewer con-
tacts with substrate and the opposing loop. However,
in the crystal structures, the loop is affected by crystal
contacts with neighbouring protein molecules; the MD
simulations do not show any increase in RMSFs or
the opening frequency of loop B2, but rather show
very similar dynamics in GcaCel7A and HjeCel7A.
Accordingly, we predict that the probability of endo-
mode initiation for GcaCel7A is low and similar to
what has been determined for HjeCel7A [29].
The GcaCel7A structure represents the first report
of O-glycosylation of a GH7 catalytic domain [18],
where a mannose residue is attached to Ser196 near
the tip of loop B2. We suggest that this is attributable
to the undercharacterization of GH7 glycosylation pat-
terns rather than being an outlier among the familial
members. To date, analysis of glycosylation has been
reported for only a very limited number of GH7
enzymes, and almost exclusively concerns N-glycosylation
[51–53]. The only GH7 CBH that has been extensively
characterized in terms of glycosylation is HjeCel7A,
where O-glycosylation was found on the CBM and at
numerous sites on the linker region but not in the
catalytic domain [54–58]. In previous GH7 crystal
structures, N-glycosylation has been observed, but no
O-glycosylation has been reported up to now. On the
other hand, carbohydrates attached at the surfaces of
proteins may be highly flexible and are often not
visible in the electron density, even if they are known
to be present. Thus, the question of the extent of
O-glycosylation on GH7 catalytic domains and the
frequency with which it occurs remains open. Ser is
conserved at this position in the enzymes aligned in
Fig. 3, but not across all known GH7 CBH sequences.
However, if the adjacent positions immediately before
and after Ser196 are also taken into consideration, Ser
or Thr seem to be ubiquitously present near the tip of
loop B2. A potential O-glycosylation site is missing
here in only one of 42 sequences covering phylogeneti-
cally diverse species in which GH7 CBHs have been
found, from ascomycete and basidiomycete fungi to
oomycetes, haptophytes, and parabasilids (Fig. S4).
The location of the O-glycan near the tip of loop B2
in GcaCel7A is interesting, as the mannose (or a
longer O-glycan) would significantly increase the sur-
face area of the loop contact across the active site. In
an earlier study, the introduction of an N-glycosylation
site at an adjacent position, Asn194, in Penicillium
funiculosum Cel7A, by the A196S mutation, yielded a
70% increase in cellulase activity [51]. The authors
point out that the site is located at the space between
the catalytic domain and the cellulose and speculate
that introduction of the N-glycan may deter nonspeci-
fic interactions with the substrate surface. Given
the similarities in circumstances, we suggest that the
O-glycan of loop B2 serves a similar functional role.
One could feasibly conceive that the addition of an
O-glycan in the same loop B2 region of HjeCel7A
would serve to further enhance the activity over the
wild-type; however, the requirements for inducing
O-glycosylation by the expression host are not well
understood, making engineering such a construct
difficult.
The implementation of GcaCel7A in whole cellulase
cocktails results in similar synergistic effects as
observed with HjeCel7A. Almost two-fold higher con-
version of Avicel cellulose was achieved when half of
the enzyme was replaced with a Cel7A-depleted
H. jecorina enzyme cocktail (Table 2). The overall
yields were slightly lower for GcaCel7A, although this
can potentially be attributed to coevolved synergistic
effects. In other words, GcaCel7A has coevolved with
the other enzymes in the G. candidum secretome and
may be fine-tuned for optimal synergism in that con-
text rather than in the context of the specific composi-
tion and properties of the H. jecorina cocktail. In fact,
earlier studies showed that naturally secreted cocktails
from G. candidum degraded filter paper and cotton
more efficiently than the industrial enzyme prepara-
tions to which they were compared, including Cellulase
Onozuka R-10 from Trichoderma viride (Japan), Meiji
cellulose from Acremonium cellulolyticus (Japan), and
rapidase from Aspergillus niger and Trichoderma
longibrachiatum (France) [8,11].
In conclusion, we have found that GcaCel7A shows
similar structural and functional characteristics to the
industrially relevant HjeCel7A. Nevertheless, notable
industrial advantages over HjeCel7A, including
reduced product inhibition and a broader optimal pH
range, are valuable in the development of cellulase
products, where the implemented process conditions
may vary significantly. For example, we envision that
replacing a portion of the HjeCel7A component with
GcaCel7A in lower-temperature, high-solid-loading
15FEBS Journal (2015) ª 2015 FEBS
A. S. Borisova et al. Characterization of G. candidum Cel7A

applications would result in improvements of
H. jecorina-based enzyme cocktails. Moreover, the minor
differences in the two structures, such as O-glycosylation
and modifications at loops B2 and B4, offer new direc-
tions in the pursuit of strategies for protein engineering
of GH7 CBHs. Finally, the performance of the G. can-
didum whole cellulase cocktail should be given serious
consideration in the development of new industrial
strains.
Experimental procedures
Fungal cultivation and protein preparation
The wild-type strain G. candidum 3C (a kind gift from
A. M. Bezborodov, Bach Institute of Biochemistry,
Moscow, Russia) was cultivated for 6 days at 28 °C in a
rotary shaker at 250 r.p.m. in 2-L flasks with 500 mL of
medium [1 g L�1 KH2PO4; 1.5 g�L�1 NaNO3; 1.5 g�L�1
(NH4)2SO4; 0.5 g�L�1 MgSO4.7H2O; 40 g�L�1 wheat bran;
and 10 g�L�1 filter paper]. The enzyme was purified accord-
ing to a protocol described previously [12]. Briefly, the fun-
gal biomass was removed by centrifugation, and the broth
supernatant was concentrated, desalted, and transferred to
0.1 M sodium acetate (pH 4.5) (buffer A), by ultrafiltration
(Vivaflow 200, 30-kDa cutoff; Sartorius, Germany). The
concentrated protein solution was loaded onto a glass col-
umn with Sigmacell Cellulose Type 50 (Sigma-Aldrich,
St. Louis, MO, USA). Buffer A was used to equilibrate the
column and to wash out unbound proteins; this was fol-
lowed by elution with distilled water. Fractions containing
GcaCel7A, as judged by cellulase activity measurements
and SDS/PAGE analysis, were pooled and transferred to
0.1 M Tris/HCl (pH 7.4) (buffer B) by dialysis. The protein
solution was fractionated on a BioSuite Q column (Waters
Co., Tokyo, Japan; 21.5 9 150 mm, flow rate of
2 mL�min�1) with a linear gradient of 0–0.5 M NaCl in buf-
fer B. Then, fractions with cellulase activity were loaded
onto a BioSuite Phenyl column (Waters Co.;
21.5 9 150 mm) in buffer B with 1.7 M (NH4)2SO4 fol-
lowed by a linear gradient of 1.7–0 M (NH4)2SO4 at a flow
rate 2 mL�min�1.
For structural studies, the CBM-linker portion of Gca-
Cel7A was removed proteolytically. Papain (26 lg) acti-
vated in 0.1 M sodium phosphate (pH 7.0), 2 mM DTT and
2 mM EDTA were added to 2 mg of GcaCel7A, and incu-
bated for 8 h at room temperature. The papain was then
inactivated by addition of iodoacetate to a concentration of
3.3 mM. Then, the protein was deglycosylated with 5 lg of
endo-b-N-acetylglucosaminidase (EC 3.2.1.96, from
Streptomyces plicatus) at room temperature overnight, and
this was followed by size exclusion chromatography on a
Hiload Superdex 200 16/60 column with 50 mM sodium
phosphate (pH 7.0) and 0.15 M NaCl as eluent. Fractions
containing GcaCel7A_CD were pooled, and the resulting
stock solution was concentrated and stored at �20 °C for
use in crystallization and biochemical experiments. Protein
purity was assessed by SDS/PAGE, and protein concentra-
tions were determined by measuring the absorbance at
280 nm and using extinction coefficients of
93 040 M�1�cm�1 and 78 810 M
�1�cm�1 for full-length
GcaCel7A and GcaCel7A_CD, respectively, which were
calculated by use of the ExPASy server [59].
Gene sequencing
For DNA and RNA isolation, mycelium of the strain
G. candidum 3C grown as described above was used. Cells
were filtered through gauze, washed with 15 mM Tris/HCl
and 150 mM NaCl (pH 7.0), and frozen with liquid nitro-
gen. Genomic DNA was extracted according to [60]. Total
RNA was isolated with the Fungal/Bacterial RNA Micro-
Prep kit (ZymoResearch, Irvine, CA, USA), according to
the manufacturer’s manual. The cDNA was synthesized on
the basis of total RNA with a RevertAid First Strand
cDNA Synthesis Kit (Thermo Scientific, Waltham, MA,
USA) and the Oligo(dT)18 primer.
The partial amino acid sequence of GcaCel7A, derived
from the initial structure, was used for a PBLAST homology
search against the NCBI and MycoCosm databases (http://
genome.jgi-psf.org/programs/fungi/index.jsf) and was found
to be most similar to a GH7 CBH from T. aurantiacus.
The MycoCosm database was then browsed with the geno-
mic nucleotide sequence of T. aurantiacus Cel7A. Two
other homologous Pezizomycotina gene sequences were
retrieved, from the species Neosartorya fischeri and
Botryosphaeria dothidea. Alignment of the three gene
sequences revealed two conserved regions of 21 bp and
17 bp near the ends of the genes, which served for the
design of four primers: GcCel7A_22, 50-CCGACCTT-
GATGTTGGAGTAGA-30; GcCel7A_22_cut21, 50-CCGA
CCTTGATGTTGGAGTAG-30; GcCel7A_22_cut17, 50-CCTTGATGTTGGAGTAG-30; and GcCel7A_17, 50-TACAC
CAACTGCTACAC-30. Gene amplification was performed
with an Eppendorf Mastercycler under the following condi-
tions: 2 min at 95 °C; 35 cycles of 30 s at 95 °C; 30 s at
52 °C; 45 s at 72 °C; and exposure at 72 °C for 5 min. The
reaction mixture contained 50 ng of template DNA, 5 lgof each primer, 0.2 mM each dNTP, and 2.5 U of Taq
polymerase (Evrogen, Moscow, Russia), in the appropriate
buffer. The resulting amplicons were sequenced at Evrogen
Co. The resulting PCR product, with a size of ~ 1200 bp,
was purified and sequenced, and the sequence was aligned
to a draft of the G. candidum whole genome sequence
(BioProject ID: PRJNA243259 [16]) to identify the com-
plete coding region (CDS) of the GcaCel7A gene and to
design primers for the CDS amplification. The complete
GcaCel7A CDS was PCR-amplified under the above condi-
tions, with primers CDS-F (50-ACCTTTGTCGTCCAT
CATGGC-30) and CDS-R (50-CCTTGCCTTGGATC
16 FEBS Journal (2015) ª 2015 FEBS
Characterization of G. candidum Cel7A A. S. Borisova et al.

TTAGTTTGG-30), and cDNA from G. candidum as the
PCR template. The resulting PCR product was sequenced
with a BigDye Terminator v3.1 Cycle Sequencing Kit (Life
Technologies, Carsbad, CA, USA) and a 3500xL Genetic
Analyzer (Life Technologies). The nucleotide sequence cod-
ing for GcaCel7A has been deposited in NCBI GenBank
under the accession number KJ958925.
Cellulase activity assays
Assessment of cellulase activity, e.g. during protein purifica-
tion, was routinely performed by incubating enzyme sam-
ples with 5 mg�mL�1 Avicel PH-101 (Fluka-Sigma-Aldrich)
microcrystalline cellulose as substrate in 100 lL of 0.1 M
sodium acetate (pH 5.0), at 37 °C for 120 min with vigor-
ous agitation every 20 min. The reaction was stopped by
addition of 250 lL of 1 M NaOH. After removal of the
remaining solids by centrifugation, the amount of released
reducing sugar was determined with p-hydroxybenzoic acid
hydrazide reagent as previously described [61], against glu-
cose standards.
Temperature and pH dependence of GcaCel7A
activity and stability
The pH dependence of GcaCel7A activity on Avicel was
determined as above, but with 0.1 M sodium/citrate/phos-
phate buffers ranging from pH 3.0 to pH 7.5. The pH depen-
dence of protein stability was assessed by preincubation of
GcaCel7A for 24 h at 37 °C in the sodium/citrate/phosphate
buffers, followed by measurement of the residual activity on
Avicel at pH 5.0 as above. The temperature dependence of
GcaCel7A activity was monitored by performing incubations
with Avicel at temperatures ranging from 30 °C to 60 °C (at
pH 5.0) and measuring the release of reducing sugar. Enzyme
thermal stability was assessed by preincubation of GcaCel7A
at different temperatures (20–60 °C) for 1 h, followed by
measurement of the residual activity on Avicel under the
standard assay conditions (pH 5.0, 37 °C, 2 h). The data
points are means of at least two or three independent experi-
ments, and errors were within 7–10%.
Substrate specificity assays
The specific activity of GcaCel7A on different polysaccha-
rides was assessed by incubation with 5 mg�mL�1 substrate
and determination of released reducing sugar as described
above (pH 5.0, 37 °C), except that enzyme concentrations
and incubation times were chosen to yield 3–5% substrate
conversion. The substrates were: Avicel, CMC (Sigma-
Aldrich, cat. no. 419311), xylan from beech wood (Sigma-
Aldrich; cat. no. X-4252), lichenan (b-1,3-1,4-glucan; MP
Biomedicals, Moscow, Russia, cat. no. 02155231), PASC
prepared from Avicel PH-101 as previously described
[62,63], and BC produced by Gluconacetobacter hansenii
B6756 (purchased from the Russian National Collection of
Industrial Microorganisms, Moscow, Russia) and prepared
as previously described [64]. PASC and BC were incubated
for 1 h with 0.075 mg�mL�1 and 0.5 mg�mL�1 enzyme,
respectively, and the other substrates were incubated for
20 h with 0.5 mg�mL�1 enzyme. The specific activity,
U�mg�1, is defined as the accumulated amount in lmol of
glucose equivalents released per mg of enzyme divided by
the incubation time in minutes under the assay conditions.
Comparison of GcaCel7A and HjeCel7A activity
The activities of GcaCel7A and HjeCel7A (prepared as previ-
ously described [65]) were compared on Avicel with Cel7A
enzyme acting alone and in cooperation with other cellulases.
For the latter experiment, an industrial H. jecorina enzyme
cocktail, Accellerase 1500 (Danisco-Genencor, Palo Alto,
CA, USA), was fractionated by anion exchange chromatog-
raphy, and Cel7A-containing fractions were removed. The
remaining fractions were pooled to yield a Cel7A-depleted
enzyme cocktail, referred to as Cel7A-free Accellerase.
Enzyme reactions contained 50 lg�mL�1 Cel7A enzyme
alone, or 25 lg�mL�1 Cel7A + 25 lg�mL�1 Cel7A-free
Accellerase, and 5 g�L�1 Avicel PH-101 in 300 lL of 0.1 M
sodium acetate (pH 5.0). Five replicates were incubated at
40 °C in closed Eppendorff tubes for 2 h with occasional
mixing (~ 15-min intervals). The reactions were stopped by
removal of the remaining substrate by filtration with 1-lmWhatman glass fibre 96-well plates in a vacuum filtration
unit (Porvair Sciences, Wrexham, Wales, UK). The filtrate
was incubated with 67 lg�mL�1 almond b-glucosidase(Sigma-Aldrich) for 1 h at 37 °C to hydrolyse cellobiose to
glucose, and this was followed by the p-hydroxybenzoic acid
hydrazide reducing sugar assay [61].
Enzyme kinetics and cellobiose inhibition on
chromogenic substrates
Enzyme kinetics experiments were performed in triplicate in
96-well microtitre plates with three chromogenic substrates:
CNP-G2, pNP-G2 and pNP-Lac (all from Sigma-Aldrich).
Reaction mixtures of 150 lL containing 0.01–5 mM sub-
strate and 0.64 lM GcaCel7A_CD in 50 mM sodium acetate
(pH 5.0) were incubated at 30 °C for 30 min, and the reac-
tions were stopped by the addition of 150 lL of 0.5 M
sodium carbonate. Absorbance was read at 404 nm with an
Eon Microplate Spectrophotometer (BioTek Instruments,
Winooski, VT, USA), and the concentration of released
chromophore was calculated by the use of extinction coeffi-
cients of 16 800 M�1�cm�1 for 2-chloro-4-nitrophenolate and
18 300 M�1�cm�1 for 4-nitrophenolate ion. Product inhibi-
tion experiments were carried out as above in duplicate, with
0.64 lM and 0.17 lM GcaCel7A_CD and pNP-Lac as sub-
17FEBS Journal (2015) ª 2015 FEBS
A. S. Borisova et al. Characterization of G. candidum Cel7A

strate, in the absence and presence of 20 lM and 80 lM cel-
lobiose. Appropriate controls were included to compensate
for any background absorbance by substrate solutions and/
or other compounds used. The Michaelis–Menten kinetics
parameters kcat and KM, and the competitive inhibition con-
stant, Ki, were derived by nonlinear regression with the pro-
gram ORIGIN 8.0 (OriginLab, Northampton, MA, USA).
X-ray crystallography
Crystallization experiments were carried out with deglyco-
sylated GcaCel7_CD. Screening for crystallization condi-
tions was performed in 96-well sitting drop trays with a
Mosquito crystallization robot (TTP Labtech, UK). The
most promising crystallization hits were obtained at room
temperature with Hampton PEG/Ion screen conditions,
with 0.2 M MgCl2 and 20% PEG 3350 as a precipitant.
The crystals used for data collection were grown by sitting
drop vapour diffusion under the same conditions after 1 : 1
mixing of precipitant with 6.5 mg�mL�1 GcaCel7A_CD in
0.15 M NaCl and 50 mM sodium phosphate (pH 7.0). No
ligand was added to the crystallization conditions for the
APO1 structure. The APO2 crystal was soaked for 15 min
with 5 mM XGO, but this ligand was not visible in the
structure. The G2, G3 and G4 structure complexes were
obtained from cocrystallization drops including lami-
naribiose (5 mM; Seikagaku Corporation, Tokyo, Japan),
4,40-dithio-cellotriose (5 mM), or 4-thio-cellobiose (10 mM),
respectively. Thio-cellooligosaccharides were synthesized by
sequential elongation of the thio-oligosaccaride chain from
the nonreducing to the reducing end, starting from
corresponding S-glycosyl isothiourea precursors and the
4-O-trifluoromethanesulfonyl derivative of galactose, with a
previously described approach [66,67]. A detailed descrip-
tion of the synthesis will be published elsewhere. NMR
spectra of all synthesized thiocellooligosaccharides were in
full agreement with previously published data [68]. Crystals
selected for X-ray data collection were briefly soaked in
mother liquor supplemented with 30% glycerol as cryopro-
tectant and then flash-frozen in liquid nitrogen. X-ray
diffraction data were collected at 100 K at the synchrotron
beamlines I911-2 and I911-3 (MAX-Lab, Lund, Sweden)
and ID23-1 (ESRF, Grenoble, France), as indicated in
Table 3. The data were integrated with XDS [69] and scaled
with SCALA or AIMLESS in the CCP4 suite [70]. The initial
GcaCel7A_CD structure model was solved by molecular
replacement with PHASER [71] and a structure of PchCel7D
as the search model (PDB code 1GPI [25]).
REFMAC5 [72] was used for structure model refinements,
and manual model rebuilding was performed with COOT
[73,74], with maximum likelihood A-weighted 2Fo–Fc
electron density maps [74]. For cross-validation of R and
Rfree calculations, 5% of the data were excluded from the
structure refinement [75]. Solvent molecules were automati-
cally added by use of the automatic water-picking function
in the ARP/WARP package [76]. Picked water molecules were
selected or discarded manually by visual inspection of
2Fo�Fc and Fo�Fc electron density maps. The coordinates
for the five final GcaCel7A_CD structure models and the
structure factors have been deposited in the PDB (http://
wwpdb.org/) with accession codes 5AMP, 4ZZV, 4ZZT,
4ZZW, 4ZZU, respectively.
MD simulations
Six total MD simulations were performed to examine the
protein dynamics of GcaCel7A relative to those of Hje-
Cel7A: GcaCel7A with no ligand, GcaCel7A bound to cel-
lononaose, GcaCel7A bound to a cellulose microfibril,
HjeCel7A with no ligand, HjeCel7A bound to cel-
lononaose, and HjeCel7A bound to a cellulose microfibril
(Fig. S1). In each case, only the catalytic domain of the
enzyme was considered. Additionally, the catalytic domains
were simulated in a deglycosylated state, as we have previ-
ously determined that catalytic domain glycosylation con-
tributes little to protein dynamics on the timescale of an
MD simulation [77,78]. Here, we briefly describe our
approach to construction, equilibration and execution of
the six MD simulations. Detailed simulation methodology
is provided in Doc. S1.
CHARMM was used to build and solvate the GcaCel7A
and HjeCel7A simulations from the reported GcaCel7A,
5AMP, and the 4C4C structure, respectively [41,79]. The
simulations of the catalytic domains with no bound ligands
were performed by simply removing the ligand from the
active site. Simulations of the bound catalytic domains in
solution included the cellononaose ligand from the 4C4C
structure of HjeCel7A, which was docked to GcaCel7A_CD
by alignment of the protein backbones. In the cellulose
microfibril simulations, the active site tunnels again bind
the 4C4C cellononaose ligand spanning the �7 to +2 bind-
ing sites, but the cellononaose ligand is also covalently
bound at the �7 pyranose to an edge chain of the cellulose
Ib hydrophobic surface [80,81]. The cellulose microfibril
represents the top half of a 36-chain cellulose Ib microfibril
with the corner chains removed [82]. The microfibril is 18
glucose residues in length and has a sufficient footprint to
allow the catalytic domain to fully interact. We have previ-
ously described similar system construction for the multido-
main HjeCel7A [83]. After construction, the systems were
explicitly solvated, and sodium ions were added for charge
neutrality. The catalytic domains in solution were (80 �A)3,
with ~ 52 000 atoms. The catalytic domains engaged with
cellulose microfibrils were solvated in 135 9 100 9 90-�A
boxes, resulting in systems of ~ 123 000 atoms.
The fully solvated systems were minimized and heated to
300 K and density-equilibrated in CHARMM. The 100-ns
production MD simulations were conducted in the canonical
ensemble at 300 K in NAMD with a 2-fs time step [84]. For all
simulations, the proteins were modelled by use of the CHAR-
18 FEBS Journal (2015) ª 2015 FEBS
Characterization of G. candidum Cel7A A. S. Borisova et al.

MM force field with CMAP correction [85,86]. The cellononaose
ligand and microfibrils were described by use of the CHARMM
C36 force field [87,88], and water was described by use of the
TIP3P model [89,90]. Analysis of the trajectories was per-
formed with CHARMM and VMD [79,91].
Acknowledgements
A. S. Borisova, M. Sandgren and J. St�ahlberg
acknowledge financial support from Formas (The
Swedish Research Council for Environment, Agricul-
tural Sciences and Spatial Planning) (grant no. 213-
2013-1607), and the Faculty of Natural Resources and
Agricultural Sciences, Swedish University of Agricul-
tural Sciences, through the programme ‘MicroDrivE’.
We thank Dr Majid Haddad Momeni for help with
activity measurements. C. M. Payne and S. Jana thank
the August T. Larsson Guest Researcher Programme
at the Swedish University of Agricultural Sciences for
funding and the opportunity to work alongside the
Sandgren and St�ahlberg research group. Computa-
tional time for this research was provided in part by
the National Science Foundation through Extreme
Science and Engineering Discovery Environment
(XSEDE), which is supported by National Science
Foundation grant number ACI-1053575
(TG-MCB090159). The research was also supported
by the Russian Foundation for Basic Research (project
#14-08-01041-a), the Russian Scientific Foundation
(grant #14-50-00069), and the Centre for Molecular
and Cell Technologies, Research Park, St Petersburg
State University. A. Logachev was supported by the St
Petersburg State University (grant #1.38.253.2015).
Author contributions
A. S. Borisova, E. V. Eneyskaya, K. S. Bobrov, S. Jana,
A. L. Lapidus, F. M. Ibatullin, U. Saleem, M. Sand-
gren, C. M. Payne, A. A. Kulminskaya and J. St�ahlberg
planned experiments. A. S. Borisova, E. V. Eneyskaya,
K. S. Bobrov, S. Jana, A. Logachev, D. E. Polev and
U. Saleem performed experiments. A. S. Borisova, E. V.
Eneyskaya, K. S. Bobrov, S. Jana, A. Logachev, D. E.
Polev, A. L. Lapidus, M. Sandgren, C. M. Payne
and J. St�ahlberg analysed data. F. M. Ibatullin
contributed essential material. A. S. Borisova, C. M.
Payne, A. A. Kulminskaya and J. St�ahlberg wrote the
paper.
References
1 Gottschalk G (1988) Cellulose degradation and the
carbon cycle. In Biochemistry and Genetics of Cellulose
Degradation (Aubert J-P, Beguin P & Millet J, eds), pp.
3–8. Academic Press, London.
2 Vuong TV & Wilson DB (2009) Processivity, synergism,
and substrate specificity of Thermobifida fusca Cel6B.
Appl Environ Microbiol 75, 6655–6661.3 Bhat MK (2000) Cellulases and related enzymes in
biotechnology. Biotechnol Adv 18, 355–383.4 Kirk O, Borchert TV & Fuglsang CC (2002) Industrial
enzyme applications. Curr Opin Biotechnol 13, 345–351.5 Mandels M (1976) Saccharification technology.
Biotechnol Bioeng Symp 6, 221–222.6 Reese ET & Mandels M (1971) Enzymatic Degradation.
Wiley-InterScience, New York.
7 Cherry JR & Fidantsef AL (2003) Directed evolution of
industrial enzymes: an update. Curr Opin Biotechnol 14,
438–443.8 Rodionova NA, Tiunova NA, Feniksova RV,
Kudriashova TI & Martinovich LI (1974) Cellulolytic
enzymes of Geotrichum candidum. Doklady Akademii
nauk SSSR 214, 1206–1209.9 Rodionova NA, Dubovaya NV, Eneyskaya EV,
Martinovich LI, Grasheva IM & Bezborodov AM
(2000) Purification and characterization of enda-1,4-
beta-xylanase from Geotrichum candidum 3C (in
russian). Prikl Biokhim Mikrobiol 36, 535–540.10 Tiunova NA, Rodionova NA, Martinovich LI &
Gogolev MN (1980) Preparation of cellulolytic enzymes
from Geotrichum candidum. Prikl Biokhim Mikrobiol 16,
185–190.11 Rodionova NA, Martinovich LI, Gukasyan GS &
Amelina DS (1988) Purification of cellulases from
Geotrichum candidum and Trichoderma longibrachiatum
by chromatography on microcrystalline cellulose. Prikl
Biokhim Mikrobiol 24, 370–379.12 Vasil’eva NV, Rodionova NA, Martinovich LI,
Tavobilov IM & Bezborodov AM (1989) Isolation of
endo-1,4-beta-glucanase II, bound to microcrystalline
cellulose, from a Geotrichum candidum 3C cellulase
preparation. Prikl Biokhim Mikrobiol 25, 322–332.13 Rodionova NA, Martinovich LI, Vasil’eva NV,
Tovobilov IM, Zagustina NA, Zacharov VI, De
MiloLE, Losyakova LS, Bochkareva NG, Belogortsev
YA et al. (1987) The production of glucose. In
Fermentation of Cellulose Containing Materials to the
Sugars and Biofuel. pp. 31–42. CBNTI, Minmedprom,
Moscow, 1987 (PRI GKNT USSR (21) 4262039/31-13
(22) 12.06.87 (46) 15.11.89, �A., ed) USSR.
14 Lapin VV, Rodionova NA, Zagustina NA, Kapanchan
AT, Dubovaya NV & Bezborodov AM (2002)
Application of enzyme preparartion cellokandin form
Geotrichum candidum 3C in waste paper utilization and
dehydration of glucose suspension. Prikl Biokhim
Mikrobiol 38, 452–454.15 Garcia-Guinea J, Cardenes V, Martinez AT &
Martinez MJ (2001) Fungal bioturbation
19FEBS Journal (2015) ª 2015 FEBS
A. S. Borisova et al. Characterization of G. candidum Cel7A

paths in a compact disk. Die Naturwissenschaften 88,
351–354.16 Polev DE, Bobrov KS, Eneyskaya EV & Kulminskaya
AA (2014) Draft genome sequence of geotrichum
candidum strain 3C. Genome Announc 2, e00956–14.17 Morel G, Sterck L, Swennen D, Marcet-Houben M,
Onesime D, Levasseur A, Jacques N, Mallet S,
Couloux A, Labadie K et al. (2015) Differential gene
retention as an evolutionary mechanism to generate
biodiversity and adaptation in yeasts. Sci Rep 5,
11571.
18 Payne CM, Knott BC, Mayes HB, Hansson H, Himmel
ME, Sandgren M, Stahlberg J & Beckham GT (2015)
Fungal cellulases. Chem Rev 115, 1308–1448.19 Divne C, Stahlberg J, Reinikainen T, Ruohonen L,
Pettersson G, Knowles JK, Teeri TT & Jones TA
(1994) The three-dimensional crystal structure of the
catalytic core of cellobiohydrolase I from Trichoderma
reesei. Science 265, 524–528.20 Grassick A, Murray PG, Thompson R, Collins CM,
Byrnes L, Birrane G, Higgins TM & Tuohy MG (2004)
Three-dimensional structure of a thermostable native
cellobiohydrolase, CBH IB, and molecular
characterization of the cel7 gene from the filamentous
fungus, Talaromyces emersonii. Eur J Biochem 271,
4495–4506.21 Kern M, McGeehan JE, Streeter SD, Martin RN,
Besser K, Elias L, Eborall W, Malyon GP, Payne CM,
Himmel ME et al. (2013) Structural characterization of
a unique marine animal family 7 cellobiohydrolase
suggests a mechanism of cellulase salt tolerance. Proc
Natl Acad Sci USA 110, 10189–10194.22 Momeni MH, Goedegebuur F, Hansson H,
Karkehabadi S, Askarieh G, Mitchinson C, Larenas
EA, Stahlberg J & Sandgren M (2014) Expression,
crystal structure and cellulase activity of the
thermostable cellobiohydrolase Cel7A from the fungus
Humicola grisea var. thermoidea. Acta Crystallogr D
Biol Crystallogr 70, 2356–2366.23 Momeni MH, Payne CM, Hansson H, Mikkelsen NE,
Svedberg J, Engstr€om �A, Sandgren M, Beckham GT &
St�ahlberg J (2013) Structural, biochemical, and
computational characterization of the glycoside
hydrolase family 7 cellobiohydrolase of the tree-killing
fungus Heterobasidion irregulare. J Biol Chem 288,
5861–5872.24 Moroz OV, Maranta M, Shaghasi T, Harris PV,
Wilson KS & Davies GJ (2015) The three-dimensional
structure of the cellobiohydrolase Cel7A from
Aspergillus fumigatus at 1.5 A resolution. Acta
Crystallogr F Struct Biol Commun 71, 114–120.25 Munoz IG, Ubhayasekera W, Henriksson H, Szabo I,
Pettersson G, Johansson G, Mowbray SL & Stahlberg
J (2001) Family 7 cellobiohydrolases from
Phanerochaete chrysosporium: crystal structure of the
catalytic module of Cel7D (CBH58) at 1.32 A
resolution and homology models of the isozymes.
J Mol Biol 314, 1097–1111.26 Parkkinen T, Koivula A, Vehmaanpera J & Rouvinen J
(2008) Crystal structures of Melanocarpus albomyces
cellobiohydrolase Cel7B in complex with cello-
oligomers show high flexibility in the substrate binding.
Protein Sci 17, 1383–1394.27 Textor LC, Colussi F, Silveira RL, Serpa V, de Mello
BL, Muniz JR, Squina FM, Pereira N Jr, Skaf MS &
Polikarpov I (2013) Joint X-ray crystallographic and
molecular dynamics study of cellobiohydrolase I from
Trichoderma harzianum: deciphering the structural
features of cellobiohydrolase catalytic activity. FEBS J
280, 56–69.28 Divne C, Stahlberg J, Teeri TT & Jones TA (1998)
High-resolution crystal structures reveal how a cellulose
chain is bound in the 50 A long tunnel of
cellobiohydrolase I from Trichoderma reesei. J Mol Biol
275, 309–325.29 Kurasin M & Valjamae P (2011) Processivity of
cellobiohydrolases is limited by the substrate. J Biol
Chem 286, 169–177.30 Beckham GT, Bomble YJ, Bayer EA, Himmel ME &
Crowley MF (2011) Applications of computational
science for understanding enzymatic deconstruction of
cellulose. Curr Opin Biotechnol 22, 231–238.31 Bu L, Nimlos MR, Shirts MR, Stahlberg J, Himmel
ME, Crowley MF & Beckham GT (2012) Product
binding varies dramatically between processive and
nonprocessive cellulase enzymes. J Biol Chem 287,
24807–24813.32 Chundawat SP, Beckham GT, Himmel ME & Dale BE
(2011) Deconstruction of lignocellulosic biomass to
fuels and chemicals. Annu Rev Chem Biomol Eng 2,
121–145.33 Knott BC, Crowley MF, Himmel ME, Stahlberg J &
Beckham GT (2014) Carbohydrate-protein interactions
that drive processive polysaccharide translocation in
enzymes revealed from a computational study of
cellobiohydrolase processivity. J Am Chem Soc 136,
8810–8819.34 Petersen TN, Brunak S, von Heijne G & Nielsen H
(2011) SignalP 4.0: discriminating signal peptides
from transmembrane regions. Nat Methods 8,
785–786.35 Steentoft C, Vakhrushev SY, Joshi HJ, Kong Y,
Vester-Christensen MB, Schjoldager KT, Lavrsen K,
Dabelsteen S, Pedersen NB, Marcos-Silva L et al.
(2013) Precision mapping of the human O-GalNAc
glycoproteome through SimpleCell technology. EMBO
J 32, 1478–1488.36 Robert X & Gouet P (2014) Deciphering key features
in protein structures with the new ENDscript server.
Nucleic Acids Res 42, W320–W324.
20 FEBS Journal (2015) ª 2015 FEBS
Characterization of G. candidum Cel7A A. S. Borisova et al.

37 Tomme P, Van Tilbeurgh H, Pettersson G, Van
Damme J, Vandekerckhove J, Knowles J, Teeri T &
Claeyssens M (1988) Studies of the cellulolytic system
of Trichoderma reesei QM 9414. Analysis of domain
function in two cellobiohydrolases by limited
proteolysis. Eur J Biochem 170, 575–581.38 von Ossowski I, St�ahlberg J, Koivula A, Piens K,
Becker D, Boer H, Harle R, Harris M, Divne C, Mahdi
S et al. (2003) Engineering the Exo-loop of Trichoderma
reesei Cellobiohydrolase, Cel7A. A comparison with
Phanerochaete chrysosporium Cel7D. J Mol Biol 333,
817–829.39 Kleywegt GJ & Jones TA (1996) Phi/psi-chology:
ramachandran revisited. Structure 4, 1395–1400.40 Knott BC, Haddad Momeni M, Crowley MF,
Mackenzie LF, G€otz AW, Sandgren M, Withers SG,
St�ahlberg J & Beckham GT (2013) The mechanism of
cellulose hydrolysis by a two-step, retaining
cellobiohydrolase elucidated by structural and
transition path sampling studies. J Am Chem Soc 136,
321–329.41 Ubhayasekera W, Munoz IG, Vasella A, Stahlberg J &
Mowbray SL (2005) Structures of Phanerochaete
chrysosporium Cel7D in complex with product and
inhibitors. FEBS J 272, 1952–1964.42 Payne CM, Baban J, Horn SJ, Backe PH, Arvai AS,
Dalhus B, Bjør�as M, Eijsink VGH, Sørlie M, Beckham
GT et al. (2012) Hallmarks of processivity in glycoside
hydrolases from Crystallographic and computational
studies of the Serratia marcescens chitinases. J Biol
Chem 287, 36322–36330.43 Payne CM, Jiang W, Shirts MR, Himmel ME, Crowley
MF & Beckham GT (2013) Glycoside hydrolase
processivity is directly related to oligosaccharide
binding free energy. J Am Chem Soc 135, 18831–18839.44 Kurasin M & Valjamae P (2010) Processivity of
cellobiohydrolases is limited by the substrate. J Biol
Chem 286, 169–177.45 Sammond DW, Payne CM, Brunecky R, Himmel ME,
Crowley MF & Beckham GT (2012) Cellulase linkers
are optimized based on domain type and function:
insights from sequence analysis, biophysical
measurements, and molecular simulation. PLoS One 7,
e48615.
46 Beckham GT, Bomble YJ, Matthews JF, Taylor CB,
Resch MG, Yarbrough JM, Decker SR, Bu L, Zhao X,
McCabe C et al. (2010) The O-glycosylated linker from
the Trichoderma reesei Family 7 cellulase is a flexible,
disordered protein. Biophys J 99, 3773–3781.47 Boer H & Koivula A (2003) The relationship between
thermal stability and pH optimum studied with wild-
type and mutant Trichoderma reesei cellobiohydrolase
Cel7A. Eur J Biochem 270, 841–848.48 Becker D, Braet C, Brumer H 3rd, Claeyssens M,
Divne C, Fagerstrom BR, Harris M, Jones TA,
Kleywegt GJ, Koivula A et al. (2001) Engineering of a
glycosidase Family 7 cellobiohydrolase to more alkaline
pH optimum: the pH behaviour of Trichoderma reesei
Cel7A and its E223S/A224H/L225V/T226A/D262G
mutant. Biochem J 356, 19–30.49 Tuohy MG, Walsh DJ, Murray PG, Claeyssens M,
Cuffe MM, Savage AV & Coughlan MP (2002) Kinetic
parameters and mode of action of the
cellobiohydrolases produced by Talaromyces emersonii.
Biochim Biophys Acta 1596, 366–380.50 Voutilainen SP, Boer H, Linder MB, Puranen T,
Rouvinen J, Vehmaanper€a J & Koivula A (2007)
Heterologous expression of Melanocarpus albomyces
cellobiohydrolase Cel7B, and random mutagenesis to
improve its thermostability. Enzyme Microb Technol 41,
234–243.51 Andey WS, Jeoh T, Beckham GT, Chou Y-C, Baker
JO, Michener W, Brunecky R & Himmel ME (2009)
Probing the role of N-linked glycans in the stability and
activity of fungal cellobiohydrolases by mutational
analysis. Cellulose 16, 669–709.52 Gao L, Gao F, Wang L, Geng C, Chi L, Zhao J & Qu
Y (2012) N-glycoform diversity of cellobiohydrolase I
from Penicillium decumbens and synergism of
nonhydrolytic glycoform in cellulose degradation.
J Biol Chem 287, 15906–15915.53 Garcia-Viloca M, Gao J, Karplus M & Truhlar DG
(2004) How enzymes work: analysis by modern rate
theory and computer simulations. Science 303, 186–195.54 Chen L, Drake MR, Resch MG, Greene ER, Himmel
ME, Chaffey PK, Beckham GT & Tan Z (2014)
Specificity of O-glycosylation in enhancing the stability
and cellulose binding affinity of Family 1 carbohydrate-
binding modules. Proc Natl Acad Sci USA 111,
7612–7617.55 Harrison MJ, Nouwens AS, Jardine DR, Zachara NE,
Gooley AA, Nevalainen H & Packer NH (1998)
Modified glycosylation of cellobiohydrolase I from a
high cellulase-producing mutant strain of Trichoderma
reesei. Eur J Biochem 256, 119–127.56 Jeoh T, Michener W, Himmel ME, Decker SR &
Adney WS (2008) Implications of cellobiohydrolase
glycosylation for use in biomass conversion. Biotechnol
Biofuels 1, 10.
57 Klarskov K, Piens K, Stahlberg J, Hoj PB, Beeumen
JV & Claeyssens M (1997) Cellobiohydrolase I from
Trichoderma reesei: identification of an active-site
nucleophile and additional information on sequence
including the glycosylation pattern of the core protein.
Carbohydr Res 304, 143–154.58 Taylor CB, Talib MF, McCabe C, Bu L, Adney WS,
Himmel ME, Crowley MF & Beckham GT (2012)
Computational investigation of glycosylation effects on
a family 1 carbohydrate-binding module. J Biol Chem
287, 3147–3155.
21FEBS Journal (2015) ª 2015 FEBS
A. S. Borisova et al. Characterization of G. candidum Cel7A

59 Gasteiger EHC, Gattiker A, Duvaud S, Wilkins MR,
Appel RD & Bairoch A (2005) Protein identification
and analysis tools on the ExPASy server. In The
Proteomics Protocols Handbook (Walker JM, ed) pp.
571–601. Humana Press Inc., Totowa, NJ.
60 Lebedeva L & Tvaruzek L (2006) Specialisation of
Rhynchosporium secalis (Oud.) J.J Davis infecting
barley and rye. Plant Protect Sci 42, 85–93.61 Hori C, Igarashi K, Katayama A & Samejima M
(2011) Effects of xylan and starch on secretome of the
basidiomycete Phanerochaete chrysosporium grown on
cellulose. FEMS Microbiol Lett 321, 14–23.62 Walseth CS (1952) Occurrence of cellulases in enzyme
preparations from microorganisms. Tappi J 35,
228–233.63 Wood TM (1988) Preparation of crystalline,
amorphous, and dyed cellulase substrates. Methods
Enzymol 160, 19–25.64 Tang W, Jia S, Jia Y & Yang H (2010) The influence of
fermentation conditions and post-treatment methods on
porosity of bacterial cellulose membrane. World J
Microbiol Biotechnol 26, 125–131.65 Stahlberg J, Divne C, Koivula A, Piens K, Claeyssens
M, Teeri TT & Jones TA (1996) Activity studies and
crystal structures of catalytically deficient mutants of
cellobiohydrolase I from Trichoderma reesei. J Mol Biol
264, 337–349.66 Ibatullin FM, Selivanov SI & Shavva AG (2001) A
general procedure for conversion of S-glycosyl
isothiourea derivatives into thioglycosides,
thiooligosaccharides and glycosyl thioesters Synthesis
2001, 419–422.67 Ibatullin FM, Shabalin KA, Janis JV & Selivanov SI
(2001) Stereoselective synthesis of
thioxylooligosaccharides from S-glycosyl isothiourea
precursors. Tetrahedron Lett 42, 4565–4567.68 Schou C, Rasmussen G, Schulein M, Henrissat B &
Driguez H (1993) 4-Thiocellooligosaccharides - their
synthesis and use as inhibitors of cellulases.
J Carbohydr Chem 12, 743–752.69 Kabsch W & Sander C (1983) Dictionary of protein
secondary structure: pattern recognition of hydrogen-
bonded and geometrical features. Biopolymers 22,
2577–2637.70 Evans PR (2011) An introduction to data reduction:
space-group determination, scaling and intensity
statistics. Acta Crystallogr D Biol Crystallogr 67,
282–292.71 McCoy AJ, Grosse-Kunstleve RW, Adams PD, Winn
MD, Storoni LC & Read RJ (2007) Phaser
crystallographic software. J Appl Crystallogr 40,
658–674.72 Murshudov GN, Skubak P, Lebedev AA, Pannu NS,
Steiner RA, Nicholls RA, Winn MD, Long F & Vagin
AA (2011) REFMAC5 for the refinement of
macromolecular crystal structures. Acta Crystallogr D
Biol Crystallogr 67, 355–367.73 Emsley P & Cowtan K (2004) Coot: model-building
tools for molecular graphics. Acta Crystallogr D Biol
Crystallogr 60, 2126–2132.74 Emsley P, Lohkamp B, Scott WG & Cowtan K (2010)
Features and development of Coot. Acta Crystallogr D
Biol Crystallogr 66, 486–501.75 Collaborative Computational Project, N (1994) The
CCP4 suite: programs for protein crystallography. Acta
Crystallogr D Biol Crystallogr 50, 760–763.76 Langer G, Cohen SX, Lamzin VS & Perrakis A (2008)
Automated macromolecular model building for X-ray
crystallography using ARP/wARP version 7. Nat
Protoc 3, 1171–1179.77 Payne CM, Bomble YJ, Taylor CB, McCabe C,
Himmel ME, Crowley MF & Beckham GT (2011)
Multiple functions of aromatic-carbohydrate
interactions in a processive cellulase examined with
molecular simulation. J Biol Chem 286, 41028–41035.78 Taylor CB, Payne CM, Himmel ME, Crowley MF,
McCabe C & Beckham GT (2013) Binding site
dynamics and aromatic-carbohydrate interactions in
processive and non-processive family 7 glycoside
hydrolases. J Phys Chem B 117, 4924–4933.79 Brooks BR, Brooks CL III, Mackerell AD Jr, Nilsson
L, Petrella RJ, Roux B, Won Y, Archontis G, Bartels
C, Boresch S et al. (2009) CHARMM: the biomolecular
simulation program. J Comput Chem 30, 1545–1614.80 Beckham GT, Matthews JF, Peters B, Bomble YJ,
Himmel ME & Crowley MF (2011) Molecular-Level
origins of biomass recalcitrance: decrystallization free
energies for four common cellulose polymorphs. J Phys
Chem B 115, 4118–4127.81 Lehtio J, Sugiyama J, Gustavsson M, Fransson L,
Linder M & Teeri TT (2003) The binding specificity
and affinity determinants of family 1 and family 3
cellulose binding modules. Proc Natl Acad Sci USA
100, 484–489.82 Nishiyama Y, Langan P & Chanzy H (2002) Crystal
structure and hydrogen-bonding system in cellulose Ibfrom synchrotron X-ray and neutron fiber diffraction.
J Am Chem Soc 124, 9074–9082.83 Payne CM, Resch MG, Chen L, Crowley MF, Himmel
ME, Taylor LE, Sandgren M, St�ahlberg J, Stals I, Tan
Z et al. (2013) Glycosylated linkers in multimodular
lignocellulose-degrading enzymes dynamically bind to
cellulose. Proc Natl Acad Sci 110, 14646–14651.84 Phillips JC, Braun R, Wang W, Gumbart J,
Tajkhorshid E, Villa E, Chipot C, Skeel RD, Kale L &
Schulten K (2005) Scalable molecular dynamics with
NAMD. J Comput Chem 26, 1781–1802.85 MacKerell AD, Bashford D, Bellott M, Dunbrack RL,
Evanseck JD, Field MJ, Fischer S, Gao J, Guo H, Ha
S et al. (1998) All-atom empirical potential for
22 FEBS Journal (2015) ª 2015 FEBS
Characterization of G. candidum Cel7A A. S. Borisova et al.

molecular modeling and dynamics studies of proteins.
J Phys Chem B 102, 3586–3616.86 MacKerell AD Jr, Feig M & Brooks CL III (2004)
Extending the treatment of backbone energetics in
protein force fields: limitations of gas-phase quantum
mechanics in reproducing protein conformational
distributions in molecular dynamics simulations.
J Comput Chem 25, 1400–1415.87 Guvench O, Greene SN, Kamath G, Brady JW,
Venable RM, Pastor RW & MacKerell AD Jr (2008)
Additive empirical force field for hexopyranose
monosaccharides. J Comput Chem 29, 2543–2564.88 Guvench O, Hatcher E, Venable RM, Pastor RW &
MacKerell AD Jr (2009) CHARMM additive all-atom
force field for glycosidic linkages between
hexopyranoses. J Chem Theory Comput 5, 2353–2370.89 Durell SR, Brooks BR & Bennaim A (1994) Solvent-
induced forces between 2 hydrophilic groups. J Phys
Chem-Us 98, 2198–2202.90 Jorgensen WL, Chandrasekhar J, Madura JD, Impey
RW & Klein ML (1983) Comparison of simple
potential functions for simulating liquid water. J Chem
Phys 79, 926–935.91 Humphrey W, Dalke A & Schulten K (1996) VMD:
visual molecular dynamics. J Mol Graph Model 14,
33–38.
Supporting information
Additional supporting information may be found in
the online version of this article at the publisher’s web
site:Table S1. Sequence identities and rmsd values between
GcaCel7A and other GH7 CBH structures.
Table S2. Simulation parameters for all six MD simu-
lations included in this study.
Fig. S1. Illustrations of the six MD simulations per-
formed as part of this study.
Fig. S2. MD simulation results comparing the active
site properties and dynamics of GcaCel7A and Hje-
Cel7A.
Fig. S3. Histograms of the minimum distances of (A)
loops A1 to B2 and (B) loops B2 to B3 for GcaCel7A
and HjeCel7A.
Fig. S4. Multiple sequence alignment at loop B2 of 42
GH7 CBH sequences from phylogenetically distant
species in the eukaryote tree of life suggests that
potential O-glycosylation sites in the form of Ser or
Thr are ubiquitously present near the tip of loop B2.
Doc. S1. Detailed MD protocol.
Doc. S2. Additional MD simulation results.
Doc. S3. References.
23FEBS Journal (2015) ª 2015 FEBS
A. S. Borisova et al. Characterization of G. candidum Cel7A























![9`T\Vj @]j 3c`_kV cV]ZVgVd % UVTRUVd `W ... - Daily Pioneer](https://img.dokumen.tips/doc/110x75/633e91f1b8d946e528072a9b/9tvj-j-3ckv-cvzvgvd-uvtruvd-w-daily-pioneer.jpg)





