Embed Size (px)
Citation preview

ARTICLE IN PRESS
0022-5193/$ - se
doi:10.1016/j.jtb
�CorrespondE-mail addr
Journal of Theoretical Biology 247 (2007) 733–742
www.elsevier.com/locate/yjtbi
Semi-discrete host–parasitoid models
Abhyudai Singha,�, Roger M. Nisbetb
aDepartment of Electrical and Computer Engineering, University of California at Santa Barbara, CA 93106-9610, USAbDepartment of Ecology, Evolution and Marine Biology, University of California at Santa Barbara, CA 93106-9610, USA
Received 1 December 2006; received in revised form 15 March 2007; accepted 4 April 2007
Available online 10 April 2007
Abstract
Arthropod host–parasitoid interactions constitute a very important class of consumer resource dynamics. Discrete-time models are a
tradition for such interactions and are characterized by an updating function, which relates the population densities at a fixed date in one
year to those at the same date in the previous year. Previous workers have investigated the effects of functional response and density
dependence on the stability of the host–parasitoid interaction by heuristically incorporating them in the updating function. Such an
approach ignores the effects of population changing continuously within a year due to different processes (for example intraspecific
competition, mortality from parasitism) that may act simultaneously. Their cumulative effect on the updating function is not obvious
and a more systematic methodology is needed. This paper uses a hybrid approach to formulate the updating function. This is done by
modeling the dynamics of various within-year processes in continuous-time, and reproduction as a discrete event. Using this formalism
we derive results connecting the stability of the host–parasitoid interaction with different forms of density dependence and the form of
the functional response. The latter results contradict previous conclusions from heuristically formulated models, and illustrate the need
for such a hybrid approach in discrete-time host–parasitoid theory.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Consumer–resource dynamics; Host–parasitoid models; Semi-discrete models
1. Introduction
One of the central themes in ecology is the interactionbetween populations of consumers (e.g. predators, para-sitoids) and resources (e.g. prey, host). There is a largebody of literature that studies such interactions using twowell-developed approaches: continuous-time models, whichare used to model populations with overlapping genera-tions and all year round reproduction, and discrete-timemodels, which are more suited for populations whichreproduce in a discrete pulse determined by season(Murdoch et al., 2003).
A large body of theory has focused on the dynamics ofarthropod host–parasitoid systems, dating back to theseminal work of Nicholson and Bailey (1935). Reviews canbe found in Hassell (2000) and Murdoch et al. (2003).
e front matter r 2007 Elsevier Ltd. All rights reserved.
i.2007.04.004
ing author. Tel.: +1805 403 2533.
esses: [email protected] (A. Singh),
csb.edu (R.M. Nisbet).
Discrete-time models are the traditional framework used, achoice that reflects the univoltine life histories of manytemperate-region insects. In such systems the host species isusually vulnerable to a particular parasitoid species at onlyone stage of its life cycle, commonly the larval stage. Onecommon life cycle involves adult hosts that emerge duringspring or summer, lay eggs and then die. The eggs matureinto larvae and for a short interval of time are vulnerable toattack by the parasitoids. Surviving host larvae pupate,overwinter in the pupal stage, and emerge as adult hoststhe following year. Adult parasitoids search for hostsduring the window of time that host larvae are present,then die. Parasitized hosts mature into a juvenile para-sitoid, pupate, overwinter, and emerge as adults thefollowing year. Synchronized life cycles, little overlap oflife stages, and no overlap of generations in both hosts andparasitoids suggest that discrete-time models are appro-priate for these systems.The updating function in a discrete-time, host–parasitoid
model relates the population densities at a fixed date in one

ARTICLE IN PRESS
Fig. 1. Life cycle of the host in year t.
A. Singh, R.M. Nisbet / Journal of Theoretical Biology 247 (2007) 733–742734
year to those at the same date in the previous year, anddescribes the cumulative effect of all the processes thathave occurred within the year. However, life processes arecontinuous, and the form taken by this cumulative effectmay be far from intuitive, especially where multipleprocesses operate simultaneously. A systematic way toformulate the updating function is presented in Box 4.1 ofMurdoch et al. (2003), and involves a hybrid approach,where the continuous processes involved in the within-yeardynamics are described by a continuous-time model, andreproduction is modeled as a discrete event. We followPachepsky et al. (2007), and call such models semi-discrete.
Semi-discrete models have previously been used toinvestigate interactions between adults and juveniles withina season with discrete between-season dynamics (Eskolaand Geritz, 2007), to model systems where a host orresource has discrete generations and is attacked by aconsumer with non-seasonal dynamics (e.g. Dugaw et al.,2004; Bonsall and Hassell, 1999; Briggs and Godfray,1996), and systems with discrete consumer generations andvery fast resource dynamics (Geritz and Kisdi, 2004). Semi-discrete models have also been used to address specificquestions relating to host–parasitoid dynamics; for exam-ple the stabilizing effects of within-season movement ofparasitoids (e.g. Rohani et al., 1994), and the consequencesof phenological asynchrony between parasitoids and theirhosts (Munster-Swendsen and Nachman, 1978; Godfrayet al., 1994). However, the main body of discrete-time,host–parasitoid theory does not take advantage of thisformalism.
In this paper we revisit discrete-time host–parasitoidtheory using semi-discrete models. We derive new resultsrelating stability to the form of the functional response,and to different forms of density dependence. Someprevious conclusions from heuristically formulated modelssurvive, but others are contradicted, notably those relatingthe form of the functional response to stability. Ourapproach can easily be extended to include more complexbiological interactions in contexts where a number ofprocesses can occur concurrently.
2. Model structure
A general model describing host–parasitoid dynamics indiscrete-time is given by
Htþ1 ¼ F ðHt;PtÞ, (1a)
Ptþ1 ¼ GðHt;PtÞ, (1b)
where Ht and Pt are the adult host and adult parasitoiddensities, respectively, at a fixed date near the start of year t
(where t is an integer). The host life cycle is illustrated inFig. 1 where Ht adult hosts give rise to eggs which matureinto RHt host larvae at the start of the vulnerable stage.Here R41 denotes the number of viable eggs produced byeach adult host. The time within the vulnerable larvae stageis denoted by t which varies from 0 to T corresponding to
the start and end of the vulnerable stage, respectively. Theupdate functions F, G are obtained by first writing acontinuous-time model that describes the various contin-uous-mortality sources that occur during the vulnerablestage of the host and is given by the following system ofordinary differential equations:
dLðt; tÞdt
¼ � g1½Lðt; tÞ;Pðt; tÞ; Iðt; tÞ�Pðt; tÞLðt; tÞ
� g2½Lðt; tÞ;Pðt; tÞ; Iðt; tÞ�Lðt; tÞ, ð2aÞ
dPðt; tÞdt
¼ �g3½Lðt; tÞ;Pðt; tÞ; Iðt; tÞ�Pðt; tÞ, (2b)
dIðt; tÞdt
¼ g1½Lðt; tÞ;Pðt; tÞ; Iðt; tÞ�Pðt; tÞLðt; tÞ
� g4½Lðt; tÞ;Pðt; tÞ; Iðt; tÞ�Iðt; tÞ, ð2cÞ
where Pðt; tÞ, Lðt; tÞ and Iðt; tÞ are the density ofparasitoids, un-parasitized and parasitized host larvae,respectively, at a time t within the vulnerable stage in yeart. For convenience of presentation, in the rest of the paperwe suppress the dependence on t and t and represent theabove differential equations by
dL
dt¼ �g1LP� g2L;
dP
dt¼ �g3P,
dI
dt¼ g1LP� g4I . (3)
The functions g2, g3 and g4 (units: time�1) represent(potentially density-dependent) host mortality due tocauses other than parasitism, parasitoid mortality, andmortality of the parasitized larvae, respectively. Thefunction g1 (units: time�1 parasitoid�1) is the (potentiallydensity-dependent) attack rate of the parasitoids. Theproduct g1L is frequently referred to as the functional
response and represents the instantaneous rate at whichhosts are attacked per parasitoid in the population. Theabove set of ordinary differential equations are thenintegrated starting from t ¼ 0 with initial conditions
Lð0; tÞ ¼ RHt; Pð0; tÞ ¼ Pt; Ið0; tÞ ¼ 0, (4)
up to t ¼ T . Assuming that each parasitized host larvaegives rise to k adult parasitoids in the next generation, theupdate functions can be obtained as
F ðHt;PtÞ ¼ LðT ; tÞ, (5a)
GðHt;PtÞ ¼ kIðT ; tÞ. (5b)

ARTICLE IN PRESSA. Singh, R.M. Nisbet / Journal of Theoretical Biology 247 (2007) 733–742 735
The stability analysis for the resulting discrete-time modelcan then be investigated using the standard Jury conditions(Elaydi, 1996).
If g2 ¼ g4 ¼ 0, i.e. host larvae mortality is solely throughparasitism and there is no parasitized host mortality, thenfrom (2) we have that Lðt; tÞ þ Iðt; tÞ is constant through-out the vulnerable period and is equal to RHt. In that casethe model (1) takes the more familiar form
Htþ1 ¼ RHtf ðHt;PtÞ, (6a)
Ptþ1 ¼ kRHt½1� f ðHt;PtÞ�, (6b)
where
f ðHt;PtÞ :¼F ðHt;PtÞ
RHt
(7)
denotes the fraction of host larvae that escape parasitismand is calculated from the solution of the system (2) ofdifferential equations.
3. Effects of functional responses
If the attack rate g1 is assumed to be a constant c,implying a linear functional response, and ifg2 ¼ g3 ¼ g4 ¼ 0, then integrating the corresponding con-tinuous-time model leads to the classic Nicholson–Baileymodel with
f ðHt;PtÞ ¼ expð�cTPtÞ, (8)
in the discrete-time model (6) (Box 4.1, Murdoch et al.,2003). A typical population time series of this model is asfollows: at low densities of hosts and parasitoids both thepopulations grow, when the population of hosts becomeslarge enough the parasitoid begins to overexploit the hostleading to a crash of the host population, followed by acrash of the parasitoids. Such cycles of over exploitationand crashes make this interaction unstable and both hostsand parasitoids show diverging oscillations. Variousauthors have investigated Type II (Rogers, 1972) and TypeIII functional responses (Hassell and Comins, 1978) withthe conclusion that they do not stabilize such discretegeneration host–parasitoid populations. They did this byassuming that the attack rate is a function of the initial hostlarvae density. For example, for a Type II functionalresponse, it was taken as
g1 ¼c
1þLð0; tÞ
LH
¼c
1þRHt
LH
(9)
for some constants c and LH . By contrast, our semi-discretemodel recognizes that in a real system, the number of hostslarvae declines throughout the vulnerable period due toattack by the parasitoids. Thus it is assumed that the attackrate is a function of Lðt; tÞ, the current host larvae densityinstead of Lð0; tÞ. We illustrate next, using a simple class offunctional responses which have the form cLmþ1, that thewithin season variation in host density can change thequalitative dynamics from instability to stability. These
accelerating functional responses are mathematically easyto analyze and provide insight on host–parasitoid interac-tions for the more complicated Type III functionalresponse which we investigate in Section 3.3. We beginby considering quadratic functional responses.
3.1. Quadratic functional responses
We consider the dynamical interaction
dL
dt¼ �cL2P;
dI
dt¼ cL2P;
dP
dt¼ 0, (10)
i.e. g1 ¼ cL, g2 ¼ 0, g3 ¼ 0 and g4 ¼ 0 which incorporatesa quadratic functional response. The solution to the abovecontinuous-time model is
Lðt; tÞ ¼RHt
1þ cRHtPtt; Pðt; tÞ ¼ Pt,
Iðt; tÞ ¼ RHt � Lðt; tÞ ð11Þ
which yields the following function f ðHt;PtÞ in thediscrete-time model (6)
f ðHt;RtÞ ¼LðT ; tÞ
RHt
¼1
1þ cTRHtPt
. (12)
Stability analysis of the model (see Appendix A) revealsthat the equilibrium point
H� ¼1ffiffiffiffiffiffiffiffiffiffiffiffi
kTRcp ; P� ¼ kðR� 1ÞH� (13)
is neutrally stable with a period of 2p= arctanðffiffiffiffiffiffiffiffiffiffiffiffiffiffiR2 � 1p
Þ forsmall amplitude oscillations. Thus, the cycle period is verylong when R is only slightly greater than one (the minimumvalue for viable populations), drops to six for R ¼ 2, andasymptotically approaches a four year period of oscillationwith increasing R.The neutral stability persists if we add a background
density-independent parasitoid mortality, i.e, g3 ¼ c1. Weobtain the same form for the function f as (12) with cT
being replaced by c½1� expð�c1TÞ�=c1. This does not affectthe above neutral stability result which holds independentof c.
3.2. Higher order functional responses
We now consider a generalization of the above model bytaking the functional response to be cLmþ1 with m40, i.e.g1 ¼ cLm. Using similar analysis to that above, we obtain
f ðHt;RtÞ ¼1
½1þ cTmðRHtÞmPt�
1=m, (14)
and the following equilibrium densities
H� ¼Rm � 1
cTkmRmðR� 1Þ
� �1=ðmþ1Þ
; P� ¼ kðR� 1ÞH�.
(15)

ARTICLE IN PRESS
Fig. 2. Plots of the fraction of host larvae escaping parasitism f as a
function of host density Ht, as given by (14) (dashed line, functional
response is a function of the current host larvae density) and (16) (solid
line, functional response is a function of the initial host larvae density) for
R ¼ 2;T ¼ 1; c ¼ :01;m ¼ 1 and Pt ¼ 1.
A. Singh, R.M. Nisbet / Journal of Theoretical Biology 247 (2007) 733–742736
In Appendix B we show that functional response of theform cLmþ1 can stabilize the host–parasitoid interaction form41.
It is instructive to contrast this result with previous work(Rogers, 1972; Hassell and Comins, 1978) where functionalresponses were incorporated by assuming that the attackrate g1 is a constant and equal to cLð0; tÞm ¼ cðRHtÞ
m, i.e. afunction of only the initial number of host larvae available.Such an assumption does not prevent the parasitoids fromoverexploiting the hosts as a large initial host density leadsto a high attack rate throughout the vulnerable stage inspite of the fact that the host population has been reducedto low levels towards the end of the stage. Not surprisingly,the equilibrium of the corresponding discrete-time modelgiven by
f ¼ exp½�cTðRHtÞmPt� (16)
is unstable for all mX0. The difference between thisfunction and the one obtained from a semi-discrete model(Eq. (14)) is illustrated in Fig. 2. Note the larger values of f
obtained from the semi-discrete approach, i.e. more hostlarvae escaping parasitism when Ht is large as compared to(16). This is simply because the semi-discrete approachtakes into account that the attack rate is not a constant butdecreases with the host larval densities throughout thevulnerable period. This leads to a larger host populationescaping parasitism and prevents overexploitation, whichcontributes to the stability of the semi-discrete model.
3.3. Type II and III functional responses
Changing the attack rate to
g1 ¼
cL
LH
� �m
1þL
LH
� �mþ1(17)
with mX1 incorporates a sigmoidal functional responsewith maximal value cLH . Analysis in Appendix C showsthat this yields the implicitly defined discrete-time model
Htþ1 �Lmþ1
H
mHmtþ1
¼ RHt �Lmþ1
H
mðRHtÞm � cTLHPt, (18a)
Ptþ1 ¼ kðRHt �Htþ1Þ (18b)
with the following equilibrium densities.
H� ¼ LH
Rm � 1
mRmðR� 1ÞðkcTLH � 1Þ
� �1=ðmþ1Þ
,
P� ¼ kðR� 1ÞH�; kcTLH41. ð19Þ
The product cTLH represents the number of hostsparasitized per parasitoid during the entire larval stagewhen the host larvae densities are large. Hence, the aboveequilibrium is only feasible if the population of parasitoidscan grow, i.e. their reproductive rate ðkcTLH Þ is larger thanone, when a large number of hosts are available.
The equilibrium in the above discrete-time model isstable for m�om where constant m� is the solution to
Rm�m�
Rm� � 1¼
kcTLHR� 1
ðkcTLH � 1ÞðR� 1Þ(20)
and has a value greater than one. If kcTLhb1 then m� � 1and if Rb1 then m� � kcTLH=ðkcTLH � 1Þ. From theabove analysis we conclude that a Type III functionalresponse can stabilize a host–parasitoid interaction as longas the initial portion of the functional response acceleratessufficiently strongly. Also, as m� is always greater than one,a Type II functional response, which would correspond tom ¼ 0, will lead to an unstable equilibrium.
4. Effects of density dependence
It is well known that density-dependent effects, ex-pressed through dependence of functions g1; g2; g3 and g4
on initial population density can stabilize the equilibria inthe Nicholson–Bailey model. For example, May et al.(1981) showed the stabilizing effect of density-dependentself-limitation in the host by introducing the discrete formof the logistic equation to the host equation and Hasselland Varley (1969) provided an analysis of the stabilizingeffects of density-dependent mutual interference betweensearching parasitoid adults. In this section we willinvestigate some density-dependent effects by incorporat-ing them in the continuous-time portion of our hybridmodel. A key advantage of using the hybrid approach here

ARTICLE IN PRESSA. Singh, R.M. Nisbet / Journal of Theoretical Biology 247 (2007) 733–742 737
is that these density-dependent effects can act concurrentlywith host mortality from parasitism. This contrasts withprevious studies where for ease of discrete-time modelformulation, it was explicitly or implicitly assumed that thedensity-dependent mortality acts at a stage before or afterthe stage where hosts are attacked by parasitoids.
4.1. Density dependent host mortality
We first consider the dynamical interaction
dL
dt¼ �cLP� c1L
2;dI
dt¼ cLP;
dP
dt¼ 0, (21)
i.e. g1 ¼ c; g2 ¼ c1L and g3 ¼ g4 ¼ 0. Here not only doboth the host mortalities occur concurrently but also thedensity-dependent mortality rate is a function of thecurrent density of hosts available. We assume thatparasitized larvae immediately becomes juvenile parasi-toids which do not face competition for resources with thehost larvae, and hence, have no density-dependencemortality acting on them, i.e. g4 ¼ 0. In the absence ofparasitoids, it is known that this system would lead toBeverton–Holt dynamics in the host (Gurney and Nisbet,1998, p. 125).
The solution to the above continuous-time modelis
Lðt; tÞ ¼RHt
expðcPttÞ þ c1RHt
expðcPttÞ � 1
cPt
, (22a)
Iðt; tÞ ¼cPt
c1ln 1þ c1RHt
1� expð�cPttÞcPt
� �, (22b)
Pðt; tÞ ¼ Pt. (22c)
which gives the discrete-time host–parasitoid model as
Htþ1 ¼ LðT ; tÞ ¼RHt expð�cTPtÞ
1þ c1RHt
1� expð�cTPtÞ
cPt
, (23a)
Ptþ1 ¼ IðT ; tÞ ¼kcPt
c1ln 1þ c1RHt
1� expð�cTPtÞ
cPt
� �.
(23b)
The above system has two non-trivial equilibrium points.The first is the no-parasitoid equilibrium that is set by thestrength of the host-density dependence
H�NC ¼R� 1
c1TR; P� ¼ 0. (24)
The Beverton–Holt-like host dynamics imply that the hostpopulation alone can never have overcompensation orexhibit cycles. Stability analysis using the Jury conditionsshow that the above equilibrium is stable in the full systemfor
lnRoc1
kc, (25)
a condition that precludes growth of a small parasitoidpopulation when the hosts are at carrying capacity. Thus,sufficiently large values of c1=ck, which can be interpretedas the strength of density-dependent mortality vs. parasit-ism, stabilize the no-parasitoid equilibrium. Similar stabi-lity analysis on Mathematica reveals that the secondequilibrium, where both host and parasitoid are presentand given by
H� ¼exp
c1
kc
� �� 1
1�exp
c1
kc
� �R
0BBBB@
1CCCCA
cP�
c1R; P� ¼
lnR�c1
kccT
, (26)
is stable for
z�oc1
cko lnR, (27)
where z� is the solution to
z� þ 1 ¼RðlnR� z�Þ
R� expðz�Þ. (28)
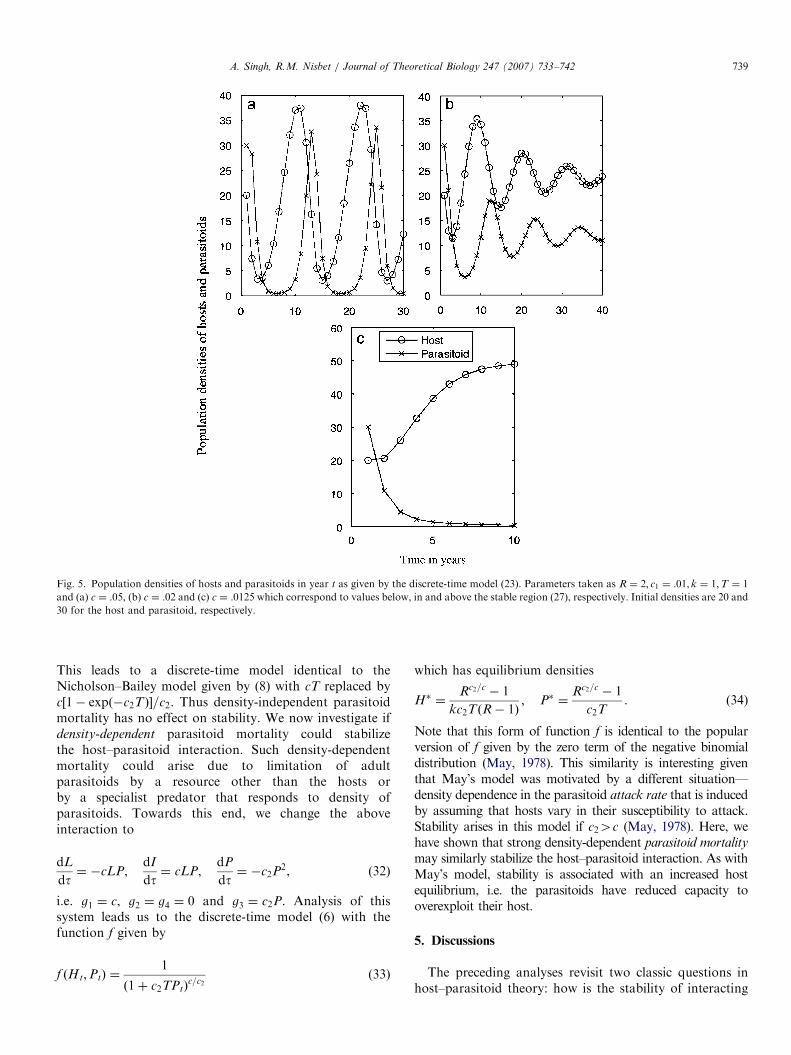
The constant z� is an increasing function of R, howeverincreasing not as fast as lnR. Thus the size of the stabilityregion specified in (27), increases with R as is illustrated inFig. 3. For c1=kc4 lnR the no-parasitoid equilibrium (24) isstable and as we will see, for c1=kcoz� one typically getspersistence of both host and parasitoid populations in thesense of bounded oscillations. Fig. 4 plots the aboveequilibrium densities and illustrates the different transitionsbetween stability and instability. Note that for each value ofthe parameters there is at most one stable equilibrium of thediscrete-time model (23). This is in contrast to the modelstudied by Neubert and Kot (1992) which reduces tothe Ricker model for the host in the absence of parasitoids.Due to the overcompensation in the Ricker model, theNeubert–Kot model may exhibit two locally stable attrac-tors, one where the consumer persists and other where it isextinct, for intermediate values of c1. Fig. 5 illustrates asimulation of the discrete-time model (23), with attack rate c
chosen such that c1=kc takes values below, in, and above thestable region (27). In summary, for a fixed parasitism rate,stability arises at intermediate levels of density-dependentmortality. At low levels of density-dependent mortality thehost population becomes large enough for the parasitoids tooverexploit it, which leads to familiar cycles of increase andcrashes. On the other hand, strong density-dependentmortality causes a very fast decrease of the hosts throughoutthe vulnerable stage and prevents the parasitoids fromexploiting the resource at all, which leads to their extinction.Stability occurs in the intermediate situation where para-sitoids can get enough recruits for the next generation butalso do not overexploit the hosts. The quantity
q ¼R exp
c1
kc� 1
� �lnR�
c1
kc
� �ðR� 1Þ R� exp
c1
kc
� � (29)
ARTICLE IN PRESS
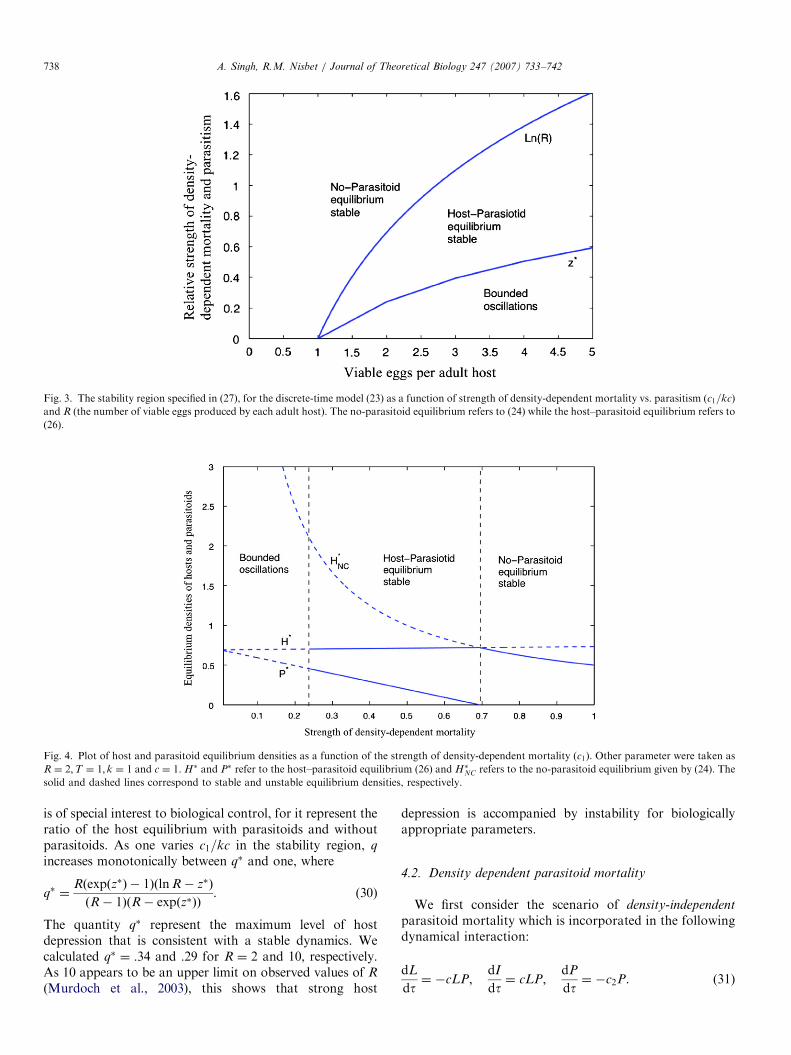
Fig. 4. Plot of host and parasitoid equilibrium densities as a function of the strength of density-dependent mortality (c1). Other parameter were taken as
R ¼ 2;T ¼ 1; k ¼ 1 and c ¼ 1. H� and P� refer to the host–parasitoid equilibrium (26) and H�NC refers to the no-parasitoid equilibrium given by (24). The
solid and dashed lines correspond to stable and unstable equilibrium densities, respectively.
Fig. 3. The stability region specified in (27), for the discrete-time model (23) as a function of strength of density-dependent mortality vs. parasitism (c1=kc)
and R (the number of viable eggs produced by each adult host). The no-parasitoid equilibrium refers to (24) while the host–parasitoid equilibrium refers to
(26).
A. Singh, R.M. Nisbet / Journal of Theoretical Biology 247 (2007) 733–742738
is of special interest to biological control, for it represent theratio of the host equilibrium with parasitoids and withoutparasitoids. As one varies c1=kc in the stability region, q
increases monotonically between q� and one, where
q� ¼Rðexpðz�Þ � 1ÞðlnR� z�Þ
ðR� 1ÞðR� expðz�ÞÞ. (30)
The quantity q� represent the maximum level of hostdepression that is consistent with a stable dynamics. Wecalculated q� ¼ :34 and :29 for R ¼ 2 and 10, respectively.As 10 appears to be an upper limit on observed values of R
(Murdoch et al., 2003), this shows that strong host
depression is accompanied by instability for biologicallyappropriate parameters.
4.2. Density dependent parasitoid mortality
We first consider the scenario of density-independent
parasitoid mortality which is incorporated in the followingdynamical interaction:
dL
dt¼ �cLP;
dI
dt¼ cLP;
dP
dt¼ �c2P. (31)
ARTICLE IN PRESS
Fig. 5. Population densities of hosts and parasitoids in year t as given by the discrete-time model (23). Parameters taken as R ¼ 2; c1 ¼ :01; k ¼ 1;T ¼ 1
and (a) c ¼ :05, (b) c ¼ :02 and (c) c ¼ :0125 which correspond to values below, in and above the stable region (27), respectively. Initial densities are 20 and
30 for the host and parasitoid, respectively.
A. Singh, R.M. Nisbet / Journal of Theoretical Biology 247 (2007) 733–742 739
This leads to a discrete-time model identical to theNicholson–Bailey model given by (8) with cT replaced byc½1� expð�c2TÞ�=c2. Thus density-independent parasitoidmortality has no effect on stability. We now investigate ifdensity-dependent parasitoid mortality could stabilizethe host–parasitoid interaction. Such density-dependentmortality could arise due to limitation of adultparasitoids by a resource other than the hosts orby a specialist predator that responds to density ofparasitoids. Towards this end, we change the aboveinteraction to
dL
dt¼ �cLP;
dI
dt¼ cLP;
dP
dt¼ �c2P
2, (32)
i.e. g1 ¼ c, g2 ¼ g4 ¼ 0 and g3 ¼ c2P. Analysis of thissystem leads us to the discrete-time model (6) with thefunction f given by
f ðHt;PtÞ ¼1
ð1þ c2TPtÞc=c2
(33)
which has equilibrium densities
H� ¼Rc2=c � 1
kc2TðR� 1Þ; P� ¼
Rc2=c � 1
c2T. (34)
Note that this form of function f is identical to the popularversion of f given by the zero term of the negative binomialdistribution (May, 1978). This similarity is interesting giventhat May’s model was motivated by a different situation—density dependence in the parasitoid attack rate that is inducedby assuming that hosts vary in their susceptibility to attack.Stability arises in this model if c24c (May, 1978). Here, wehave shown that strong density-dependent parasitoid mortality
may similarly stabilize the host–parasitoid interaction. As withMay’s model, stability is associated with an increased hostequilibrium, i.e. the parasitoids have reduced capacity tooverexploit their host.
5. Discussions
The preceding analyses revisit two classic questions inhost–parasitoid theory: how is the stability of interacting

ARTICLE IN PRESSA. Singh, R.M. Nisbet / Journal of Theoretical Biology 247 (2007) 733–742740
populations affected by the form of the functional responseand by density dependence. Our primary findings regardingdensity dependence are unsurprising, but the results on theeffects of the functional response contrast starkly withprevious work that used heuristically formulated models.
The key previous result on the effects of the functionalresponse was that of Hassell and Comins (1978) whoargued that accelerating functional response do notcompensate for the instability in the Nicholson–Baileymodel. This implies very different dynamics in discrete-time models than in their continuous-time counterpartswhere accelerating functional responses are stabilizing(Murdoch and Oaten, 1975). Fig. 2 shows the reason forthe different outcomes; essentially, our model recognizesthat parasitism slows down when the host populationbecomes small, thereby precluding the smallest hostpopulations. This leads to an increase in the equilibriumhost density and to stability. The ecological importance ofour findings on the stabilizing effects of acceleratingfunctional responses will depend on the mechanismresponsible for the acceleration. The model in this paperassumes that the parasitoid attack rate depends on theinstantaneous host density. This is obviously an idealiza-tion, as some time delay is inevitably involved. For type IIfunctional responses, there are mechanisms (e.g. thoseinvolving handling time) for which this delay is likely to beshort, but it is harder to defend this assumption withaccelerating functional responses caused by changes inparasitoid behavior. Murdoch and Bence (1987) arguedthat for aquatic predators, non-delayed, type III functionalresponses are seldom observed, and we suspect that asimilar situation holds for parasitoids. Yet the delaysinvolved in parasitoid response may plausibly be muchshorter than the duration of the vulnerable stage of thehost, and in such circumstances our model remains a morereasonable caricature of reality than its discrete-timecounterpart. The take-home message for applications isthe need for careful consideration of underlying mechan-isms and any associated time delays when constructingmodels of any particular system that includes acceleratingfunctional responses. Our conclusion that density depen-dence in either host or parasitoid is stabilizing is, by itself,unsurprising. But the dynamics of our model differs inimportant ways from previous work, not least in theabsence of multiple attractors (Neubert and Kot, 1992;Kon, 2006).
The work reported in this paper illustrates the limita-tions of models in the form of equations (6), namely
Htþ1 ¼ RHtf ðHt;PtÞ, (35a)
A ¼
1þ RH�qf ðHt;Pt
qHt
k R� 1� RH�qf ðHt;
qH
"266664
Ptþ1 ¼ kRHt½1� f ðHt;PtÞ�. (35b)
Much theory on the periods of population cycles inconsumer–resource systems assumes this form or general-izations that allow adult individuals to survive from oneyear to the next (Murdoch et al., 2001, 2003), with oneimportant conclusion being that in a broad range ofsituations, consumer–resource interactions lead to a lowerbound on cycle periods close to 6 time units. Shorterperiods typically require very large values of the geometricrate of increase, R, which is expected to be less than 10 innatural systems (Murdoch et al., 2003). By contrast, themodel in this paper with the quadratic functional responsehas a cycle period of 5 when R ¼ 3:23, well within theecologically feasible range. Yet the data analyzed byMurdoch et al. was broadly consistent with expectationsfrom the simpler model. Further work on cycle periodsusing the semi-discrete formalism is clearly called for.Finally, we note the generality of our approach. Here we
focused on two aspects of consumer–resource interactionsthat have for decades been considered important forstability of consumer–resource interactions. Many otherprocesses can be considered with semi-discrete models. Forexample, we have work in progress on the consequences ofhost-feeding where parasitoids face a choice between eatingor ovipositing in a host. Previous work (e.g. Yamamuraand Yano, 1988; Briggs et al., 1995; Kidd and Jervis, 1989)using continuous-time models has identified situationswhere host feeding has no effect on stability, and situationswhere it is stabilizing. With partially synchronized lifecycles, we lack intuition and systematic investigation isrequired.
Acknowledgments
We thank Cherie Briggs, Bill Murdoch and ElizavetaPachepsky for discussions. A. Singh would like to acknowl-edge support from J.P. Hespanha throughout this work.
Appendix A. Stability analysis for quadratic functional
responses
Denoting small fluctuation around the equilibrium (13)by htþ1 and ptþ1, one obtains after linearization thefollowing linear discrete system:
htþ1
ptþ1
" #¼ A
ht
pt
" #, (A.1)
with
ÞHt¼H�;Pt¼P�
RH�qf ðHt;PtÞ
qPt
Ht¼H� ;Pt¼P�
PtÞ
t
Ht¼H� ;Pt¼P�
#�kRH�
qf ðHt;PtÞ
qPt
Ht¼H�;Pt¼P�
377775. (A.2)

ARTICLE IN PRESSA. Singh, R.M. Nisbet / Journal of Theoretical Biology 247 (2007) 733–742 741
For stability of the above system we require the followingthree Jury conditions (Elaydi, 1996) to hold
1� trðAÞ þ detðAÞ40, (A.3a)
1þ trðAÞ þ detðAÞ40, (A.3b)
1� detðAÞ40. (A.3c)
Substituting the function
f ðHt;RtÞ ¼1
1þ cTRHtPt
(A.4)
and the equilibrium point
H� ¼1ffiffiffiffiffiffiffiffiffiffiffiffi
RcTkp ; P� ¼ kðR� 1ÞH� (A.5)
in (A.2) we have
trðAÞ ¼2
R; DetðAÞ ¼ 1 (A.6)
which corresponds to a neutrally stable equilibrium. Theroots of the characteristic equation, DetðA� lIÞ ¼ 0, ofthe matrix A are given by
l ¼ expð�yiÞ; y ¼ arctanðffiffiffiffiffiffiffiffiffiffiffiffiffiffiR2 � 1
pÞ, (A.7)
and hence, a period of oscillation of 2p= arctanðffiffiffiffiffiffiffiffiffiffiffiffiffiffiR2 � 1p
Þ.
Appendix B. Stability analysis for higher order functional
responses
Substituting the function
f ðHt;RtÞ ¼1
½1þ cTmðRHtÞmPt�
1=m(B.1)
and the equilibrium
H� ¼Rm � 1
cTkmRmðR� 1Þ
� �1=ðmþ1Þ
; P� ¼ kðR� 1ÞH�
(B.2)
in (A.2) we have
trðAÞ ¼1� R�m þ ðR� 1ÞmR�m
ðR� 1Þm,
DetðAÞ ¼Rð1� R�mÞ
ðR� 1Þm. ðB:3Þ
Using the fact that for all m41, R41 we have trðAÞ40,14DetðAÞ40 and
1� trðAÞ þ detðAÞ ¼ðmþ 1Þð1� R�mÞ
m40, (B.4)
one can see that the Jury conditions (A.3) hold, and hencethere is a stable equilibrium, for m41.
Appendix C. Stability analysis for Type II and III functional
responses
The solution for the corresponding continuous-timemodel
dLðt; tÞdt
¼ �
cLðt; tÞ
LH
� �m
1þLðt; tÞ
LH
� �mþ1
26664
37775Lðt; tÞPðt; tÞ, (C.1a)
Pðt; tÞ ¼ Pt, (C.1b)
RHt ¼ Iðt; tÞ þ Lðt; tÞ, (C.1c)
is
Lðt; tÞ �Lmþ1
H
mLmðt; tÞ¼ RHt �
Lmþ1H
mðRHtÞm � cLHPtt (C.2)
which gives us the implicitly defined discrete-time model
Htþ1 �Lmþ1
H
mHmtþ1
¼ RHt �Lmþ1
H
mðRHtÞm � cTLHPt, (C.3a)
Ptþ1 ¼ kðRHt �Htþ1Þ (C.3b)
and the following fixed point
H� ¼ LH
Rm � 1
mRmðR� 1ÞðkcTLH � 1Þ
� �1=ðmþ1Þ
,
P� ¼ kðR� 1ÞH�; kcTLH41. ðC:4Þ
Linearizing the above discrete-time model about thisequilibrium, we obtain the linear discrete system
htþ1
ptþ1
" #¼ A
ht
pt
" #, (C.5)
with
A ¼
qHtþ1
qHt
Ht¼H� ;Pt¼P�
qHtþ1
qPt
Ht¼H� ;Pt¼P�
qPtþ1
qHt
Ht¼H� ;Pt¼P�
qPtþ1
qPt
Ht¼H� ;Pt¼P�
266664
377775: ðC:6Þ
Using (C.3) and (C.4) we find that
trðAÞ ¼Rþ
gRm þ kcTLH
1þ g; DetðAÞ ¼
RkcTLH
1þ g,
g ¼ðkcTLH � 1ÞðR� 1ÞmRm
Rm � 1. ðC:7Þ
Substituting these in the Jury conditions (A.3), one findsthat inequalities (A.3a)–(A.3b) hold for all m40;kcTLH41;R41. The stability condition comes from

ARTICLE IN PRESSA. Singh, R.M. Nisbet / Journal of Theoretical Biology 247 (2007) 733–742742
inequality (A.3c) which holds for all m�om where constantm�41 is the solution to
m�
1�1
Rm�
� � ¼ kcTLHR� 1
ðkcTLH � 1ÞðR� 1Þ. (C.8)
References
Bonsall, M.B., Hassell, M.P., 1999. Parasitoid-mediated effects: apparent
competition and the persistence of host–parasitoid assemblages. Res.
Popul. Ecol. 41 (1), 59–68.
Briggs, C.J., Godfray, H.C.J., 1996. The dynamics of insect-pathogen
interactions in seasonal environments. Theor. Popul. Biol. 50, 149–177.
Briggs, C.J., Nisbet, R.M., Murdoch, W.W., Collier, T.R., Metz, J., 1995.
Dynamical effects of host-feeding. J. Anim. Ecol. 64, 403–416.
Dugaw, C.J., Hastings, A., Preisser, E.L., Strong, D.R., 2004. Parasitoid-
mediated effects: apparent competition and the persistence of
host–parasitoid assemblages. Bull. Math. Biol. 66 (3), 583–594.
Elaydi, S., 1996. An Introduction to Difference Equations. Springer, New
York.
Eskola, T.M., Geritz, S.A., 2007. On the derivation of discrete-time
population models by varying within-season patterns of reproduction
and aggression. Bull. Math. Biol. 69, 329–346.
Geritz, S.A., Kisdi, E., 2004. On the mechanistic underpinning of discrete-
time population models with complex dynamics. J. Theor. Biol. 228
(2), 261–269.
Godfray, H., Hassell, M., Holt, R., 1994. The population dynamic
consequences of phenological asynchrony between parasitoids and
their hosts. J. Anim. Ecol. 63 (1), 1–10.
Gurney, W.S.C., Nisbet, R.M., 1998. Ecological Dynamics. Oxford
University Press, Oxford.
Hassell, M.P., 2000. The Spatial and Temporal Dynamics of Host
Parasitoid Interactions. Oxford University Press, New York.
Hassell, M.P., Comins, H.N., 1978. Sigmoid functional responses and
population stability. Theor. Popul. Biol. 14, 62–66.
Hassell, M.P., Varley, G.C., 1969. New inductive population model for
insect parasites and its bearing on biological control. Nature 223,
1133–1137.
Kidd, N.A.C., Jervis, M.A., 1989. The effects of host-feeding behaviour
on the dynamics of parasitoid–host interactions, and the implications
for biological control. Res. Popul. Ecol. 31 (2), 235–274.
Kon, R., 2006. Multiple attractors in host–parasitoid interactions:
coexistence and extinction. Math. Biosci. 201, 172–183.
May, R.M., 1978. Host–parasitoids in patchy environments: a phenom-
enological model. Am. Nat. 47, 833–843.
May, R.M., Hassell, M.P., Anderson, R.M., Tonkyn, D.W., 1981. Density
dependence in host–parasitoid models. J. Anim. Ecol. 50, 855–865.
Munster-Swendsen, M., Nachman, G., 1978. Asynchrony in insect
host–parasite interaction and its effect on stability, studied by a
simulation model. J. Anim. Ecol. 47 (1), 159–171.
Murdoch, W.W., Bence, J.R., 1987. General predators and unstable prey
populations. In: Kerfoot, W.C., Sih, A. (Eds.), Predation in Aquatic
Communities Direct and Indirect Effects. New England, pp. 17–30.
Murdoch, W.W., Oaten, A., 1975. Predation and population stability.
Adv. Ecol. Res. 9, 1–131.
Murdoch, W.W., Kendall, B.E., Nisbet, R.M., Briggs, C.J., McCauley, E.,
Bolser, R., 2001. Single-species models for many-species food webs.
Nature 417, 541–543.
Murdoch, W.W., Briggs, C.J., Nisbet, R.M., 2003. Consumer–Resource
Dynamics. Princeton University Press, Princeton, NJ.
Neubert, M.G., Kot, M., 1992. The subcritical collapse of predator
populations in discrete-time predator–prey models. Math. Biosci. 110,
45–66.
Nicholson, A., Bailey, V.A., 1935. The balance of animal populations.
Part 1. Proc. Zool. Soc. London 3, 551–598.
Pachepsky, E., Nisbet, R.M., Murdoch, W.W., 2007. Between discrete and
continuous: Consumer-resource dynamics with synchronized repro-
duction. Ecology, in press.
Rogers, D.J., 1972. Random searching and incest population models.
J. Anim. Ecol. 41, 369–383.
Rohani, P., Godfray, H.C.J., Hassell, M.P., 1994. Aggregation and the
dynamics of host–parasitoid systems: a discrete-generation model with
within-generation redistribution. Am. Nat. 144 (3), 491–509.
Yamamura, N., Yano, E., 1988. A simple model of host–parasitoid
interaction with host-feeding. Res. Popul. Ecol. 30 (2), 353–369.





























