Embed Size (px)
Citation preview

Journal of Ecology 20201081101ndash1114 wileyonlinelibrarycomjournaljec emsp|emsp1101copy 2019 The Authors Journal of Ecology copy 2019 British Ecological Society
Received20June2019emsp |emsp Accepted2October2019DOI1011111365-274513306
R E S E A R C H A R T I C L E
Seagrass ecosystem metabolic carbon capture in response to green turtle grazing across Caribbean meadows
Robert A Johnson1 emsp| Alexandra G Gulick1 emsp| Nerine Constant1 emsp| Alan B Bolten1 emsp| Fee O H Smulders2 emsp| Marjolijn J A Christianen2 emsp| Mabel I Nava3emsp| Keith Kolasa4emsp| Karen A Bjorndal1
1ArchieCarrCenterforSeaTurtleResearchandDepartmentofBiologyUniversityofFloridaGainesvilleFLUSA2AquaticEcologyandWaterQualityManagementGroupWageningenUniversityampResearchWageningenTheNetherlands3SeaTurtleConservationBonaireKralendijkBonaireCaribbeanNetherlands4HernandoCountyPublicWorksHernandoBeachFLUSA
CorrespondenceRobertAJohnsonEmailjohnsonrobertufledu
Present addressRobertAJohnsonDepartmentofEcologyEvolutionandOrganismalBiologyIowaStateUniversityAmesIAUSA
Funding informationNationalScienceFoundationGrantAwardNumberDGE-1315138FitzsimmonsFund
HandlingEditorARandallHughes
Abstract1 Increasinggreenturtleabundancewillleadtoincreasedgrazingwithinseagrasshabitatsmdashecosystems that are important for carbon sequestration and storageHowever it is notwell understood how carbon dynamics in these ecosystemsrespondtograzingandwhetheraresponsediffersamongmeadowsorlocations
2 Wemeasuredseagrassecosystemmetabolism ingrazedandungrazedareasofThalassia testudinummeadowswithestablishedgreenturtleforagingareasacrosstheGreaterCaribbeanandGulfofMexicoWesampledmeadowsfromfivelo-cations thatdiffered in seagrassandenvironmental characteristicsEstablishedmeadowsoftheinvasiveseagrassHalophila stipulaceawerealsopresentattwooftheselocationsandwemeasuredecosystemmetabolisminthesemeadowsforcomparisontograzedandungrazedareasofthenativeT testudinum
3 Acrossallindividualsitesratesofnetecosystemproduction(NEP)rangedfrom56to96loweringrazedareasthanungrazedareasofT testudinummeadowsRatesofNEPwerealsostronglypositivelycorrelatedwithabove-groundseagrassbiomassacrosssitesWhilemetaboliccarboncapturerateswereloweringrazedareasheterotrophicrespirationwasnotstimulatedandgrazingthereforedidnotresultinsignificantmetabolicremineralizationofcarboninthesemeadowsNEPin H stipulaceameadowswassimilartoratesinT testudinummeadowsatallthreesitessuggestingthatmetaboliccarboncapturemayremainsimilarinCaribbeanmeadowswherethisinvasiveseagrassisreplacingnativespecies
4 SynthesisOurresultsshowthatthereisaconsistentresponseinmetaboliccarbondynamics to green turtle grazing inT testudinummeadows across theGreaterCaribbeanregionAnincreaseingrazingwillnotlikelystimulateremineralizationofcarbonastheseimportanthabitatsarereturnedtoanaturalgrazedstate
K E Y W O R D S
carbondynamicsgrazinggreenturtleHalophilastipulaceainvasivemetabolismplantndashherbivoreinteractionsThalassiatestudinum
1102emsp |emsp emspenspJournal of Ecology JOHNSON et al
1emsp |emspINTRODUC TION
Seagrasses form someof themost productive ecosystemson theplanet (DuarteampChiscano1999)High ratesofmetabolic carboncapture and subsequent biomass production is one of the mainprocessesbywhichseagrassescontributetocarbonsequestration(known as lsquoblue carbonrsquo in seagrass habitats Duarte et al 2010Kennedy et al 2010) Among seagrass meadows globally abouthalfofthetotalcarbonstoredinameadowonaveragemaybede-rivedfromseagrassbiomass(Kennedyetal2010)Meadowsalsoexport large amounts of biomass annually contributing to carbonstorage in peripheral habitats (DuarteampCebriaacuten 1996DuarteampKrause-Jensen2017)Coupledwiththeabilitytostorecarbonforcenturiestomillenniathroughthecreationofahypoxicsedimentaryenvironment(MateoRomeroPeacuterezLittlerampLittler1997SerranoMateoRenomampJuliagrave2012Terradosetal1999)highlyproduc-tiveseagrassmeadowsformefficientnaturalcarbonsinks(DuarteKennedyMarbagraveampHendriks2013)andtheirprotectionhasbeensuggestedasaclimatechangemitigationstrategy(Macreadieetal2017Murdiyarsoetal2015)
There is an estimated150000 km2 of seagrass habitat acrossthe Caribbean Sea Gulf of Mexico and The Bahamas (Green ampShort 2003 Jackson 1997 Wabnitz Andrefouet Torres-PullizaMuller-Karger amp Kramer 2008) Thesemeadows provide import-ant foraging habitat for numerous species (Hemminga amp Duarte2000 Ogden 1976 Scott et al 2018) including green turtles(Chelonia mydas) which were historically abundant in this regionpriortooverexploitationbyhumans(Jackson1997Jacksonetal2001)GreenturtleabundanceiscurrentlyincreasinginareassuchastheCaribbean (egnestnumbersroughlytripled1970ndash2005attheCaribbeanslargestrookery)asaresultofsuccessfulconserva-tionefforts (Chaloupkaetal2008MazarisSchofieldGkazinouAlmpanidouampHays 2017) Thiswill lead tomore seagrass areasbeing returned toanaturalgrazedstate (Figure1a)Green turtlesin theGreaterCaribbean regionnormally foragebycreatinggraz-ingplotswithinseagrassmeadowsinwhichtheycropthebladestoshortheightsabovethesedimentsurfaceandcontinuallyre-grazetheseareastoconsumenewtissuegrowth(Bjorndal1980Ogden1980)Thisgrazingstrategy leads toa reductionof thephotosyn-thetic biomass in a meadow At high densities green turtles arecapableofgrazinganentiremeadowandhavebeenrecordedcon-suming100ofnewseagrassgrowthdaily(Christianenetal2012)Giventhedesiretoconserveseagrassesfortheirabilitytosequesterandstorecarbon(Macreadieetal2017)andtheexpectationthatmoreseagrasswillreturntoanaturalgrazedstate inthefuture itisnecessarytounderstandhowmetaboliccarboncapture (carbonfixedthroughphotosynthesisormineralizedthroughrespiration)isaffectedbygrazingacrossseagrassmeadows
Much of the research on seagrassmetabolic carbon dynamicsto date has focused on differences among species (LindeboomampSandee1989MurrayampWetzel1987PollardampMoriarty1991)orbetween seagrasses and unvegetated sediments (Barroacuten DuarteFrankignoulleampBorges2006RheubanBergampMcGlathery2014
Stutes Cebrian Stutes Hunter amp Corcoran 2007) with little at-tentionbeinggiventograzedareasInaCaribbeanThalassia testu-dinummeadowgrazedbygreen turtles ratesofmetaboliccarboncapture were found to be lower compared to ungrazed seagrass(Johnson Gulick Bolten amp Bjorndal 2017) However long-term
F I G U R E 1 emspAThalassia testudinumseagrassmeadowwithanareagrazedbygreenturtles(panelaleftside)andanadjacentungrazedarea(panelarightside)inEleutheraTheBahamasTheinvasiveseagrassHalophila stipulaceainStCroixUSVirginIslands(b)AlightincubationchamberdeployedinagrazedareainBonaire(c)PhotosRJohnson[Colourfigurecanbeviewedatwileyonlinelibrarycom]
(a)
(b)
(c)
emspensp emsp | emsp1103Journal of EcologyJOHNSON et al
grazing(greaterthanayear)didnotleadtoaproportionalincreaseinbenthicrespirationandremineralizationofcarbonstored inthemeadow possibly due to lack of a significant heterotrophic com-munity These results are from a single Caribbean location (LittleCayman Cayman Islands) that has some of the highest recordedmetabolicratesamongseagrassmeadows(Johnsonetal2017)ItisnotknownifgrazinghassimilareffectsacrossseagrassmeadowsinotherlocationsorunderdifferentenvironmentalconditionsAbet-terunderstandingofthevariabilityamongmeadowsinresponsetograzingisneededtounderstandhowincreasinggreenturtleabun-danceandincreasedgrazingwillaffectseagrasscarboncapture intheGreaterCaribbeanregion
In addition to increased grazing seagrass meadows in theCaribbeanarealsobeinginvadedbytheseagrassHalophila stipula-cea(Figure1b)ThisseagrassspeciesmdashnativetotheIndianOceanmdashhas invaded themeadowsofmanyCaribbean islands following itsintroductiontotheregionin2002(RuizampBallantine2004Willetteetal2014)Initialresearchsuggeststhatgreenturtlesavoidgrazingthis invasiveseagrass infavourofnativespeciesandthatH stipu-laceamayspreadmorerapidlywithinareasofmeadowsgrazedbygreen turtles compared to areas left ungrazed (Christianen et al2018)Little isknownabouthowinvasionbyH stipulaceamayaf-fectcarbondynamicsorecosystemfunctioninginCaribbeanmead-owshowever(VianaSiriwardane-deZoysaWilletteampGillis2019)GiventhisinteractionbetweengrazingandH stipulaceaitiscriticaltounderstandhowcarbondynamicsareaffectedinmeadowswhereinvasionbythisspeciesisoccurringinadditiontograzing
Ourprimaryobjectiveinthisstudywastoevaluatethehypoth-esis that effectsof green turtle grazingonmetabolic carbon cap-tureratesareconsistentacrossseagrassmeadowsWealsoevaluatevariationinthestrengthoftheresponsetograzingamongmeadowsand how rates ofmetabolic carbon capture in invasiveH stipula-ceameadowscomparetonativemeadowswhereturtlesgrazeWecomparedratesofnetecosystemproduction(NEP)betweengrazedandungrazedareasofT testudinumseagrassmeadowsinfourloca-tionsaroundtheGreaterCaribbean(definitionfromPetuch2013Robertson amp Cramer 2014) and Gulf ofMexico with establishedgreen turtle grazing populations (in addition to those previouslysampled inLittleCayman)Thefivetotal locationsencompassedawidegeographicalareaandspannedarangeofenvironmentalandseagrassmeadowcharacteristicsrepresentativeofgreenturtlefor-agingareasinthisregionTheseagrassH stipulaceahadestablishedattwooftheselocationsandwecomparedratesofNEPinmead-owsofthis invasivespeciestothosemeasuredinbothgrazedandungrazedareasofnativemeadowsattheselocations
2emsp |emspMATERIAL S AND METHODS
21emsp|emspStudy sites
We sampled grazed and ungrazed areas ofT testudinum seagrassmeadowsatfivelocationsacrosstheGreaterCaribbeanandGulfofMexicowheregreenturtleshadestablishedforagingareas(Figure2)
OneoftheselocationsLittleCaymanCaymanIslandswassampledthreetimesduringJuneandJuly2016aspartofapreviousstudy(Johnsonetal2017)The remaining four locationsweresampledin2018StCroixUSVirginIslands(February)westcoastofFlorida(GulfofMexico side)USA (May)BonaireCaribbeanNetherlands(July)EleutheraTheBahamas(August)Weopportunisticallysam-pledtwoseparatesitesinbothBonaireandEleutheraandonesiteatallotherlocations(timeandorweatherconditionsdidnotallowsamplingadditionalsitesattheselocations)EstablishedmeadowsoftheseagrassH stipulaceawerepresentattwoofthelocationsmdashStCroixandBonairemdashandthesemeadowswerealsosampledforcom-parisontonearbygrazedandungrazedareasofnativeT testudinum meadows(oneofthesitesinBonaireonlyhadnearbygrazedT tes-tudinum for comparison) Coordinates and general environmentalcharacteristics (daylight hours water depth salinity temperatureandirradiance)forallsitesaregiveninTable1
Themeadowssampledinthisstudyvariedgreatlyinsizefromrelatively small (lt150m2) for the grazedT testudinum area at thenorthwestLacBaysiteinBonairetoverylarge(severalsquarekilo-metresofunbrokenseagrasshabitat)fortheungrazedT testudinum meadowattheNorthRacksiteinFlorida(westcoast)Meadowsatothersitesencompassedarangeofsizesbetweenthesetwosuchas~350m2 each for the grazed andungrazedT testudinum areasinLittleCaymanand~600m2forthegrazedareaandgt50000m2 for the ungrazed T testudinum meadow at the Arvida Bay site inEleuthera TheBahamasAt each site a grazed area and adjacentungrazedareawereselectedforsamplingthatwererepresentativeofthemeadowSamples(Sections22and23)werecollectedwithinseveralmetresoftheborderbetweenthegrazedandungrazedareastoavoidpossibleedgeeffectswhileensuringsimilarenvironmentalconditions
Allsampledgrazedareas(naturalgreenturtlegrazingplots)werenearly uniformly grazed The proportion of grazed T testudinum blades ranged from95 to 100 in all grazed areas Evidence ofgrazingbygreenturtleswasnotobservedinungrazedareasexcept
F I G U R E 2 emspMapofseagrassmeadowsamplinglocationsTwositesweresampledineachofBonaireandEleutheraandonesitewassampledateachoftheotherthreelocationsCircledsymbolsforStCroixandBonairedenotelocationswhereHalophila stipulaceameadowswerealsosampled
1104emsp |emsp emspenspJournal of Ecology JOHNSON et al
forinStCroixwhere~3ofT testudinumbladesshowedsignsofgrazing(croppedbladetips)Somefishesalsograzeonseagrassandevidenceoffishgrazing(egbitemarks)wasobservedinungrazedT testudinum meadows at some sites Biomass removal from fishgrazingwasminorcomparedtoturtlegrazinghoweverandwasnotquantifiedNoevidenceofgreenturtlegrazingwasobservedinanyH stipulaceameadows
WesampledsevenareasfromT testudinum-dominatedseagrassmeadowsthatdisplayedsignsofactivegreenturtlegrazing(hereaf-tergrazedareas)sixareasfromT testudinum-dominatedmeadowsthatwerenotgrazed(hereafterungrazedareas)andthreemeadowsthatweredominatedbytheinvasiveseagrassH stipulacea(hereaf-terH stipulaceameadows)Thesemeadowswere spreadacross awideareaoftheGreaterCaribbeanandGulfofMexicoregionsandvariedintheirbioticandabioticcharacteristicsSomeofthesampledseagrassmeadowssuchastheT testudinummeadows inBonairewerelargelymonospecificwhereasothersweredominatedbyonespecies(T testudinum or H stipulacea)withinterspersedSyringodium filiforme or Halodule wrightii seagrasses and various macroalgae(TableS1)Thesubstrateinallmeadowswasprimarilycalciumcar-bonate sand Additional meadow characteristics are provided inResults (Section31)Data from theLittleCayman sitehavebeenreportedpreviously(Johnsonetal2017)andarepresentedagainhereforcomparisontotheothersites
22emsp|emspSampling seagrass meadow characteristics
Seagrass meadow and environmental characteristics were sam-pledateachsiteatthetimemetabolicincubationswereconducted
(Section23)Environmentaltemperatureandirradianceweremeas-uredduringmetabolicincubationsatseagrasscanopyheight(5-minintervals) at each site with a HOBO Pendant data logger (OnsetComputer Corporation) A water sample was collected at canopyheight and salinity wasmeasured with a handheld AgTec SalinityRefractometer that was calibrated with freshwater each day be-fore sampling (Agriculture Solutions Strong ME no sample wascollectedattheFloridasite)Meadowdepthwasmeasuredateachlocationwitheitheratapemeasureorascubadiversdepthgauge(accuracyplusmn10cm)
Seagrassspeciescompositionshootdensityblademorphome-try(lengthwidthandsurfacearea)andabove-groundbiomassweredetermined in all sampled areas (grazed ungrazed H stipulacea)within1metreofwhere incubationchamberswereplacedShootdensitiesweremeasuredusing25times25cmquadrats(00625m2area)in T testudinummeadowsDuetothehighdensitiesofH stipulacea10times10cmquadrats(001m2area)wereusedtomeasureshootden-sitiesinmeadowsdominatedbythisspeciesAbove-groundbiomasssampleswerecollectedusing10times10cmquadrats(allmeadows)byclippingall bladesat the sediment surfacewith scissors Six repli-catequadrats (forbothshootdensityandbiomass)werecollectedfromallsampledareasateachsiteexceptfortheStCroix(n=5)andFlorida(n=3)sites(Table2)Bladelengthandwidthweremea-suredforthedominantspecies(T testudinum or H stipulacea)from30randomlyselectedseagrassbladesfromthesebiomasssamplesforeachareaBladesurfacearea(iephotosyntheticareaperblade)wascalculatedastwotimestheproductofbladelengthandwidth(for each species) Followingmeasurement all bladeswere gentlyscrapedcleanofsedimentsandepiphytes(lowepiphyteloadsatall
TA B L E 1 emspCoordinatesofeachsamplingsiteandenvironmentalparametersmeasuredintheseagrassmeadowsondaysofmetabolicincubations
Site
Latitude Longitude Daylight Depth Salinity Temperature Irradiance
Decimal degrees hours M permil degC Lux
Bonaire
LacCaiBeach 12104417 minus68223183 1275 10 35 300 324518
NWLacBay 12108546 minus68231562 1275 14 36 311 432217
StCroix
BIRNM 17784612 minus64624566 115 45 35 279 211904
LittleCayman
GrapeTreeBay 19696518 minus80059652 13 10 36 319 551840
19696518 minus80059652 13 10 37 317 465732
19696518 minus80059652 13 10 38 336 600715
Eleuthera
ArvidaBay 24722297 minus76190644 125 24 39 322 267648
HalfSound 24936641 minus76153420 125 06 36 344 717215
Florida
NorthRack 28564760 minus82779335 1375 20 mdash 292 409801
Note TemperatureandirradiancedataaremeanvaluesmeasuredduringincubationsTheLittleCaymansitewassampledthreetimesin2016Allothersitesweresampledoncein2018SamplingtimesaregiveninSection21AbbreviationBIRNMBuckIslandReefNationalMonumentsite
emspensp emsp | emsp1105Journal of EcologyJOHNSON et al
sites) rinsed infreshwateranddriedtoaconstantweightat60degCbeforeweighingfordrymass
23emsp|emspEcosystem metabolism measurements
Metaboliccarbondynamicsoftheseagrassecosystemsweremeas-uredusingbenthic incubationchambers (Figure1c)similartopre-viousstudies (BarroacutenMarbagraveTerradosKennedyampDuarte2004CallejaBarroacutenHaleFrazerampDuarte2006Johnsonetal2017OliveacuteSilvaCostaampSantos2015)ChamberswereconstructedbyinsertingaPVCcylinder(16cmdiameter002m2area)~75cmintothe sediment (attempting to avoid severing horizontal rhizomes)and attaching a flexible gas-tight polyethylene bagwith samplingporttothetop(HansenThamdrupampJoslashrgensen2000)Theuseofflexiblegas-tightbagsinchamberconstructionallowsthepropaga-tionofwave turbulence to the chambers interior tomore closelysimulateenvironmentalconditionsIncubationchambervolumewasmeasuredinthelaboratorytobe55ndash6LOnsamplingdayscham-berswere set up in themeadows between 1030 and 1130 andincubationswererunfor25ndash3hrThisincubationlengthwaschosenbecausesaturationeffectscanoccurwithinchambersduringlonger
incubationsandhavebeenshowntounderestimatemetabolicrates(Oliveacuteetal2015)Metabolicdynamicsweremeasuredthreetimesat2-weekintervalsinLittleCaymanaspartofapreviousstudyandshown to be relatively stable among sampling events (Johnson etal2017) Incubationswerethereforeconductedonceateachad-ditionalsiteinthepresentstudy
Wesetupthreelight(clear)andthreedark(opaque)chambersineacharea(grazedorungrazedT testudinumorH stipulacea)tomea-sureecosystemmetabolicrates(n=3forallareas)LightchamberswereusedtomeasureratesofNEPanddarkchamberswereusedtomeasure ratesofecosystemrespiration (RE)Metabolic rateswereestimatedfromchangesindissolvedoxygen(DO)concentrationin-side the chambersbetween thebeginningandendof the incuba-tionperiodThreewatersampleswerecollectedfromthechamberviathesamplingportin60-mlplasticsyringesatthebeginningandendoftheincubationUponcollectionsyringeswerecappedwitha silicon cap andbrought to the surfacewhereDOconcentrationwas measured directly in the syringe with an optical DO probe(Vadeboncoeur2011YSIProODOYellowSpringsOhio)TheDOprobewas calibrated at the beginning of each sampling daywithwater-saturatedair
TA B L E 2 emspSeagrasscharacteristicsofgrazedandungrazedareasofThalassia testudinummeadowsandHalophila stipulaceameadowsmeasuredateachsamplingsite
Site Meadow n
Seagrass density Canopy heightBlade surface area AG biomass
shootsm2 cm cm2 g DMm2
Bonaire
LacCaiBeach Grazed 6 7413plusmn1263 47plusmn22 63plusmn32 271plusmn76
Ungrazed 6 9547plusmn937 96plusmn39 214plusmn116 1309plusmn544
H stipulacea 6 38500plusmn8068 21plusmn06 19plusmn09 364plusmn133
NWLacBay Grazed 6 2373plusmn446 28plusmn14 28plusmn14 48plusmn28
H stipulacea 6 34833plusmn5492 20plusmn04 18plusmn06 171plusmn59
StCroix
BIRNM Grazed 5 19648plusmn2261 40plusmn23 44plusmn27 274plusmn102
Ungrazed 5 24064plusmn2501 81plusmn31 97plusmn45 516plusmn110
H stipulacea 5 37336plusmn12494 28plusmn05 32plusmn09 630plusmn240
LittleCayman
GrapeTreeBay Grazed 6 9573plusmn1485 20plusmn09 23plusmn12 123plusmn37
Ungrazed 6 10187plusmn2102 149plusmn85 307plusmn204 1832plusmn948
Eleuthera
ArvidaBay Grazed 6 16480plusmn3927 35plusmn19 42plusmn22 418plusmn139
Ungrazed 6 20533plusmn3299 152plusmn46 258plusmn88 2022plusmn348
HalfSound Grazed 6 16747plusmn3167 18plusmn07 14plusmn06 88plusmn20
Ungrazed 6 17840plusmn2847 94plusmn20 108plusmn31 965plusmn162
Florida
NorthRack Grazed 3 13227plusmn4441 25plusmn11 16plusmn08 93plusmn24
Ungrazed 3 13867plusmn1619 89plusmn27 78plusmn29 707plusmn233
Note nisnumberofreplicatesateachsiteValuesaremeansplusmnSDCanopyheightismeasuredfrommeanbladelengthBladesurfaceareaisperbladeofseagrassAbbreviationsAGabove-groundBIRNMBuckIslandReefNationalMonumentsiteDMdrymass
1106emsp |emsp emspenspJournal of Ecology JOHNSON et al
Tomeasureonlymetabolicratesoftheseagrassbenthiccompo-nentsofthesystem(thoseaffectingcarbonstorage)wecorrectedrates measured from incubation chambers for water column me-tabolismWemeasuredwater columnmetabolismusingbiologicaloxygendemand(BOD)bottlesatthesametimeasbenthicchamberincubationsThreeclear300-mlglassBODbottlesandthreeopaque300-mlglassBODbottleswereusedtomeasurewatercolumnpro-ductionandrespirationrespectivelyBottleswerefilledwithwaterat seagrasscanopyheightanchored to thebottomand incubatedunderinsituconditionsintheseagrassmeadowWatercolumnsam-pleswerecollectedatthebeginningofincubationsin60-mlsyringesatcanopyheightandDOconcentrationwasmeasuredinthesamemannerassamplesfromincubationchambersFollowingthe incu-bationperiodbottleswerecollectedand returned to thesurfaceonesamplewascollectedfromeachbottlewithasyringeandDOconcentrationwasmeasured
Hourlymetabolic rateswere calculated from thedifference inDOconcentrationwithinlight(NEP)anddark(RE)incubationcham-bersbetweenthebeginningandendoftheincubationperiodWatercolumn metabolic rates measured from BOD bottles were sub-tractedfromratesmeasuredinchambersHourlygrossprimarypro-duction(GPP)wasthencalculatedasthesumofhourlyNEPandRE DailyratesofGPPwerecalculatedbymultiplyinghourlyGPPbythelengthofthephotoperiodmultipliedby075(toaccountfor lowermetabolicratesatdawnandduskJohnsonetal2017Oliveacuteetal2015)anddailyratesofREwerecalculatedbymultiplyinghourlyRE by24hoursDailyNEPwascalculatedas thedifferencebetweendaily GPP and RE Therewere a few occasionswhen an increaseinDO concentrationwasmeasuredwithin a dark chamber or thedarkBODbottlesduringanincubationsuggestinganerrorThesemeasuredincreasesweresmallusuallywithintheprobesmarginoferror (01mgO2L)andweassumedrespirationtobezeroduringthese timesCorrecting for these respiration errors hadminor ef-fectsondailymetabolic rates in themeadows (TablesS2andS3)MeasuredDOconcentrationswereconvertedfrommgO2tommolO2andthentocarbonunits(mmolC)assumingphotosyntheticandrespiratoryquotientsofone(BarroacutenampDuarte2009)
24emsp|emspData analyses
Net ecosystem productionwas themetabolic variable of primaryinterestasitaccountsforbothcarboncaptureandlossandcanbeusedas an indicatorofwhether a system is currently ametaboliccarbonsinkorsourceThereforeNEPisthemetabolicvariableforwhichstatisticalresultsarepresented
Ouraimwastoevaluatewhethergreenturtlegrazinghasacon-sistenteffectonmetabolicratesacrossseagrassecosystemsmdashthatistoevaluatewhetheratanygivensiteatanygiventimeratesofNEPareloweringrazedareasthaninungrazedareasWeevaluateddifferences in ratesofNEPbetweenareasofgrazedandadjacentungrazedT testudinum from each sitewith paired t tests T testswereusedbecause thecomparisonof interestwas thedifferencebetween grazed and ungrazed areas (effect of grazing) within a
givensitenotdifferencesinNEPamongsitesForsiteswheretheinvasive seagrass H stipulacea was present we used one-wayANOVAstotestforsignificantdifferencesinratesofNEPbetweenH stipulaceameadowsandnearbygrazedandungrazedT testudinum areas(StCroixsiteandLacCaiBeachsiteBonaire)Forthenorth-west site in Lac Bay Bonaire where only a grazed area of T te-studinum and an invasive H stipulacea meadow were comparedanunpaired t testwasused IfANOVA resultswere significant aTukeyspost-hoctestwasusedtoidentifysignificantcomparisonsWetestedfordifferencesinseagrassmeadowparametersbetweengrazedandungrazedT testudinumareasandH stipulaceameadowsatsitesusingpairedttestsandANOVAsinthesamemannerasforNEPWeused linearregressiontotestrelationshipsbetweenNEPand explanatory variables (eg seagrass biomass environmentaltemperature)MeanvaluesforNEPandexplanatoryvariablesfromeachmeadowwereusedforlinearregressionstestssotherewasasingledatapointforeachgrazedandungrazedT testudinum area or H stipulaceameadowateachsamplingsite
AllanalyseswereperformedinRversion343(RCoreTeam2018)Datawere processed using the lsquodplyrrsquo package (Wickham FrancoisHenryampMuumlller2017)andpost-hocanalyseswereperformedusingthelsquoagricolaersquopackage(deMendiburu2017)Alldatamettheassump-tionsfortheseanalysesanddataarepresentedasmeanplusmnSD
3emsp |emspRESULTS
31emsp|emspSeagrass meadow characteristics
Total seagrass shoot densities varied widely among meadowsand locations (Table 2) though shoot densitieswere always high-est inH stipulaceameadows The lowest shoot density observed(2373plusmn446shootsm2)wasinagrazedarea(NWsiteinLacBayBonaire)buttherewasnocleartrendforhigherorlowerseagrassdensities between grazed and ungrazed areas across the sampledsitesThalassia testudinum density in LittleCaymanwasgreater inthe grazed area than ungrazed area (paired t test p = 02) whileT testudinumdensityattheLacCaiBeachsiteinBonairewasgreaterintheungrazedarea(pairedttestp=02)Thalassia testudinum den-sitydidnotdifferbetweengrazedandungrazedareasattheremain-ingsites(pairedttestspgt05)ShootdensityofH stipulaceawasnot significantly different between any of the threeH stipulacea meadowsmeasured(ANOVAF2=009p=92)
HalophilaseagrassestypicallygrowshortbladesCanopyheightoftheH stipulaceameadowwassimilartothatofthenearbygrazedarea inStCroix andwas shorter than thenearbygrazedareasatbothsitesinBonaire(Table2)Halophila stipulaceabladelengthwasgreaterinStCroixthanBonaire(ANOVAF2=2413plt01)AreasofT testudinum-dominatedseagrassmeadowsgrazedbygreentur-tleshad significantly shorterblades and therefore canopyheightthan adjacent ungrazed areas at all sampled sites (paired t testsplt05forallcomparisonsTable2)
Greenturtlesbycroppingseagrassbladesnearthesedimentsur-faceandreducingcanopyheightgreatlyreducetheabove-ground
emspensp emsp | emsp1107Journal of EcologyJOHNSON et al
seagrass biomass of meadows within their grazing areas Above-ground biomass ranged from 48 plusmn 28 to 418 plusmn 139 g DMm2 amonggrazedareas(Table2)andfrom516plusmn110to2022plusmn348gDMm2amongungrazedareasBiomasswasalwaysloweringrazedareas than adjacent ungrazed areas (p lt 05 for all comparisons)Biomass among the three sampledH stipulaceameadows rangedfrom171plusmn59to630plusmn240gDMm2TheH stipulacea meadow in St Croix had significantly greater above-ground biomass thaneither of theH stipulaceameadows sampled inBonaire (ANOVAF2=1202plt 01)Evidenceofgreenturtlegrazingwasonlyob-served in T testudinum-dominated meadows in this study notH stipulaceameadowsandgrazingresultedinagt79decreaseinbiomassingrazedcomparedtoungrazedareasatallsitesexceptStCroix(47Table3)
32emsp|emspEcosystem metabolic rates
Rates of NEP differed among meadow types (grazed ungrazedH stipulacea) across the studied locations (Figures 3 and 4 seeTable S2 for data) Across all sites NEP ranged from 42 plusmn 70 to517plusmn110mmolCmminus2 dayminus1amonggrazedareasandfrom523plusmn57to2253plusmn191mmolCmminus2 dayminus1amongungrazedareasThelargestandarddeviationsonsomeoftheungrazedareaNEPestimatesfromLittleCaymanwerearesultofoneincubationchamberproducingratesmuchdifferentfromtheothertwoAcrossallsitesNEPwasconsist-entlyloweringrazedareasthanungrazedareas(Figure3)DifferencesinNEPbetweengrazedandungrazedareasweresignificantat four
ofthesixsiteswhereungrazedT testudinumwaspresentStCroixLittleCayman (all threemeasurement times) theArvidaBay site inEleutheraandFlorida(pairedttestsplt05forallcomparisons)AttheLacCaiBeachsiteinBonaireandtheHalfSoundsiteinEleuthera
TA B L E 3 emspPercentdifferenceinmetabolicratesandabove-ground(AG)seagrassbiomassbetweengrazedandadjacentungrazedThalassia testudinumareas
Site
GPP RE NEP AG Biomass
difference
Bonaire
LacCaiBeach 818 909 656 793
StCroix
BIRNM 604 783 560 469
LittleCayman
GrapeTreeBay 922 955 891 933
899 796 960 933
885 870 900 933
Eleuthera
ArvidaBay 543 395 629 793
HalfSound 801 889 679 908
Florida
NorthRack 615 413 919 868
Note TheLittleCaymansitewassampledthreetimesin2016Allothersitesweresampledoncein2018SamplingtimesaregiveninSection21AbbreviationsBIRNMBuckIslandReefNationalMonumentsiteGPPgrossprimaryproductionNEPnetecosystemproductionREecosys-temrespiration
F I G U R E 3 emspRates(meanplusmnSD)ofnetecosystemproduction(a)grossprimaryproduction(b)andecosystemrespiration(c)fromgrazed(openpoints)andadjacentungrazed(solidpoints)areasofThalassia testudinumseagrassmeadowsRatesofNetecosystemproduction(NEP)weresignificantlyloweringrazedareasthanungrazedareasatallsitesexceptatLacCaiBeachBonaireandHalfSoundEleuthera(Section32)SitesorderedsouthtonorthBONBonaire(LCBLacCaiBeachsite)STXStCroixLCLittleCaymanELEleuthera(ABArvidaBaysiteHSHalfSoundsite)FLFloridaDashedlinein(a)denotesmetabolicbalance(NEP=0)Valuesabovelinerepresentnetmetaboliccarboncapture[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1108emsp |emsp emspenspJournal of Ecology JOHNSON et al
differencesinNEPbetweengrazedandungrazedareaswerenearlysignificant(pairedttestsp = 08 and p=09respectively)
Though NEP was significantly lower in grazed than ungrazedareas atmost sites rates ofNEPwere almost always positive asecosystem respiration was not stimulated in relation to primaryproductionevenwithconsiderablereductionofphotosyntheticsea-grassbiomassbygrazing(gt79biomassreductionatallsitesexceptStCroix Table3)NEP ranged from56 to96 lower in grazedareasthanungrazedareas(Table3)Asimilarrangeinthedifference
betweengrazedandungrazedareaswasmeasuredforgrossprimaryproduction (54ndash92 lower in grazed) and ecosystem respiration(40ndash96loweringrazed)
Halophila stipulacea meadows were present at three sites andratesofNEPintheseinvasiveseagrassmeadowswerecomparedtonearbynativeT testudinum-dominatedmeadowsNEPintheH stip-ulaceameadowswascomparedtonearbygrazedandungrazedareasofT testudinum-dominatedmeadowsattwoofthesites(StCroixandtheLacCaiBeachsiteinBonaire)WithineachsiteratesofNEPintheH stipulaceameadowwerebetweenthoseofthegrazedandungrazedareas (Figure 4) but the differences were not significant (ANOVApgt10forallpost-hoccomparisonswithineachsite)AnungrazedareawasnotavailableforcomparisonatthethirdsiteatwhichH stipula-ceawaspresent (NWLacBaysite inBonaire)asallT testudinumatthissitewasgrazedbygreenturtlesandratesofNEPdidnotdifferbetweenthe invasive-andnative-dominatedareasatthissiteeither(t test p = 56 Figure 4)Halophila stipulaceameadowNEP rangedfrom612plusmn226to989plusmn233mmolCmminus2 dayminus1(TableS2)anddidnotdiffersignificantlyamongthethreesitesatwhichitwaspresent(ANOVAF2=192p=23)eventhoughtheestablishmentofH stip-ulaceaseagrassmeadowsinStCroixwasrelativelyrecent(AGulickunpublisheddata)comparedtothoseinBonaire(Willetteetal2014)
33emsp|emspDrivers of metabolic rates
VariationinratesofseagrassecosystemmetabolismacrossgrazedandungrazedareasappearedtobedrivenbysomecharacteristicsoftheseagrassmeadowsbutnotbyenvironmentalfactorsMeadowNEPwasstronglypositivelyrelatedtoabove-groundseagrassbio-massacrosssites(linearregressionR2=82plt01Figure5a)NEP
F I G U R E 4 emspRates(meanplusmnSD)ofnetecosystemproductioninmeadowsdominatedbytheinvasiveseagrassHalophila stipulacea (triangles)comparedtonearbygrazed(opencircles)andungrazed(solidcircles)areasofThalassia testudinummeadowsBONBonaire(LCBLacCaiBeachsiteNWnorthwestLacBaysite)STXStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
F I G U R E 5 emspRelationshipbetweennetecosystemproductionandabove-groundseagrassbiomass(a)andtotalseagrassshootdensity(b)fromgrazed(openpoints)andungrazed(solidpoints)areasofThalassia testudinummeadowsSolidlinein(a)isthesignificantlinearregression(R2=82plt01)betweennetecosystemproduction(NEP)andbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows
emspensp emsp | emsp1109Journal of EcologyJOHNSON et al
wasalsopositivelycorrelatedwithseagrassmeadowcanopyheight(R2=84plt01)andbladesurfacearea(R2=92plt01)Above-groundbiomassislikelythemoreusefulpredictorofmeadowNEPhoweverasbiomassisinfluencedbybothcanopyheightandbladesurfaceareaandachange ineitherofthesevariablesresults inachangeinbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows(linearregressionR2lt01p=96Figure5b)norwerethererelationshipsbetweenNEPandenvironmentaltempera-ture(R2=01p=70)irradiance(R2lt01p=10)ormeadowdepth(R2lt01p=77)acrosssites(FigureS1)
4emsp |emspDISCUSSION
Through a comparative study of seagrass meadowsmdashgrazed andungrazedmdashacross a wide area encompassing sites in the GreaterCaribbean and Gulf of Mexico we found that green turtle graz-inghas a consistent effecton themetabolic carbon capture ratesofT testudinum-dominatedseagrassmeadowsRatesofmetaboliccarboncapturewerealwayslowerinareasgrazedbyturtlesthanun-grazedareasofmeadowsmdashupto96inLittleCaymanmdashbutratesofNEPwerealwaysnearorabovemetabolicbalanceEcosystemres-pirationwasnotstimulatedinrelationtoprimaryproductionwithinareas grazed by green turtles regardless of the amount of above-groundseagrassbiomassremovedthroughgrazingorotherdiffer-encesinbioticandabioticfactorsamongsitesThesefindingslendadditionalsupporttothehypothesisthatfutureratesofmetaboliccarboncapturemaybelowerinareasgrazedbyturtlesbutgrazingwillnot stimulateecosystemrespirationand result ina large rem-ineralizationof carbon currently stored in these seagrass habitats(Johnsonetal2017)
The similarity in results between St Croix (awintermeasure-ment)andtheotherfourlocationsinthepresentstudy(allsummermeasurements)strengthensourconclusionthatgrazingresultsinaconsistent response inmetaboliccarboncaptureacrossmeadowsHoweverwewerenotable toaddresspotential temporaldynam-icswithinsitesthatmayaffectnetcarboncaptureoveranannualcycle in these systems Future studies measuring carbon captureoverlongertemporalscales(ieseasonal)maybebeneficialtoourunderstanding of the effects of grazing on within-system carbondynamics
Thoughtheresponse inmetabolic rates tograzingwasconsis-tent acrossmeadows the strengthof this responsevariedamongindividual sites Some sites such as Little Cayman had consider-ablylowerratesofNEPingrazedareasthanungrazedareas(mean917)whereastheresponsetograzing(differenceinNEPbetweengrazedandadjacentungrazedareas)waslesspronouncedatothersites suchasStCroix (Table3) Seagrassmeadowcharacteristicssuch as shoot density canopy height and above-ground biomassare likely to bemore variable among ungrazed areas than grazedareasasgreenturtlegrazingresults insimilarcanopymorphome-tryacrossmeadowsintheGreaterCaribbeanregion(egBjorndal1980HernaacutendezampvanTussenbroek2014Williams1988)Given
the strong relationship between NEP and above-ground biomass(Figure 5a) high variability in NEPwould be expected across un-grazedmeadows(egFigure3a)IndeedtherangeinratesofNEPamongungrazedareaswasnearlythreetimesgreaterthantherangemeasuredacrossgrazedareasinthisstudyAssiteswerespecificallychosen that differed from each other in meadow characteristicsvariationinthestrengthofthemetabolicresponsetograzingwouldbeexpected
Variability in the strength of the metabolic response to graz-ing resulted in the lackofa significantdifferencebetweengrazedandungrazedareasattwositesmdashLacCaiBeachinBonaireandHalfSoundinEleuthera(Figure3a) It ispossiblethatotherfactorsnotaccounted for here such as local environmental factors or greenturtlegrazinghistory(egintensitylongevityofplotuse)mayhaveledtohighervariabilityinmetabolicratesatsomesitesandlackofastatisticaldifferenceInadditionmetabolicratesmayexhibitwithin-meadowvariation(patchiness)similartobelow-groundcarbonstor-age in meadows (OreskaMcglathery amp Porter 2017) leading tohighervariabilityinestimatesandhelpingtoexplainthelackofasig-nificantdifferenceinNEPatthesetwositesThiscouldbedriveninpartbywithin-meadowspatialheterogeneityinabove-groundbio-massgiventhestrongrelationshipbetweenbiomassandNEPHighwithin-meadow variability inmetabolic rates if presentmay alsohelpexplainthe largevariabilitymeasured intheungrazedarea inLittleCaymanFuturestudiesusingtheeddycorrelationtechnique(Bergetal2003) tomeasureecosystemmetabolismmaybeabletoovercomesomeofthiswithin-meadowvariabilityhoweverthegreatexpenseofthismethodmakesitprohibitiveformanystudies
Seagrass ecosystem metabolism is commonly estimated usingoxygendynamicsmeasuredwithbenthicincubationchambers(egBarroacutenetal2004Gaciaetal2005presentstudy)howeverthereare some limitations to thismethodBenthic chambersmay severbelow-ground rhizomesand release carbon that couldaffectmet-abolicestimatesIntactrhizomesontheotherhandcanaffectthediffusiveoxygenlossfromtheseagrasstotherhizosphere(BorumSand-JensenBinzerPedersenampGreve2006)andmayresultinanunderestimateofgrossprimaryproduction(andthereforeNEP)Thismaybeminimizedusingshorter incubation timesas in this studyIncubation length also strongly affects metabolic rates estimatedfromoxygendynamics (Oliveacute et al 2015)with longer incubationtimesbiasingestimatestowardsnetheterotrophyInadditiontoox-ygenmetabolismotherprocessessuchascarbonatedynamics(VanDamLopesOsburnampFourqurean2019)play important roles intotal ecosystem carbondynamicswithout affecting themeasuredproductivityofameadow
CaribbeanT testudinummeadows are highly productive (egJohnsonetal2017KochampMadden2001)andtherelationshipbetween biomass andNEP observed in this study suggests thatmeadows may maintain positive productivity as long as photo-synthetic biomass remains A threshold biomass below whichCaribbean meadows may become net heterotrophic cannot bedirectly addressed here but could be predicted to be near zeroThisisincontrasttoapreviouslysuggestedabove-groundbiomass
1110emsp |emsp emspenspJournal of Ecology JOHNSON et al
threshold(41gDMm2)belowwhichmeadowsmaybecomehet-erotrophic (Duarte et al 2010) However that threshold wascalculated for seagrasses globally and included less productivespecies Additional studies of the relationship between biomassandmeadowNEP from seagrass species and regions other thanthoseinthepresentstudywouldbebeneficialtofurthergeneral-izehowgrazingaffectsmetaboliccarboncaptureacrossseagrassecosystems
Environmental temperature and irradiance are strong driv-ers of seagrassmeadow productivity (Calleja et al 2006 Lee ampDunton 1997 Lee Park amp Kim 2007 Peacuterez amp Romero 1992)howeverpreviousstudiesexaminingcontrolsonseagrassmetabo-lismhavefocusedonungrazedseagrasses(egApostolakiHolmerMarbagraveampKarakassis2010Gaciaetal2005)Meadowmetabolicproductivity correlates stronglywith above-ground seagrass bio-mass(Johnsonetal2017Figure5a)thatcanbegreatlyaffectedby green turtle grazing (Bjorndal 1980CebriaacutenampDuarte 1998Christianenetal2012FourqureanManuelCoatesKenworthyampSmith2010Williams1988)Above-groundbiomassexplained82ofthevariationinNEPacrossgrazedandungrazedT testudi-nummeadowsinthisstudybutNEPwasnotcorrelatedwitheitherenvironmentaltemperatureorirradiance(FigureS1)WethereforeinvestigatedwhetherthelackofanobservedrelationshipbetweenNEPand temperatureand irradiancewasperhapsdue toa largereffectfromdifferencesinbiomasscausedbygrazingmaskinganyeffectoftheseenvironmentalvariablesWescaledmetabolicratestotheabove-groundbiomassineachgrazedandungrazedarea(unitmetabolismperunitbiomass)andcomparedthemtotemperatureandirradianceAccountingfordifferencesinabove-groundbiomassdidnot revealanunderlyingrelationshipbetweenNEPandtheseenvironmentalparametersacrossmeadows(grazedungrazedorallareascombinedFigureS2)ThelackofarelationshipbetweenNEPandenvironmentalparametersinthisstudymaybedueinparttothe relatively limited rangeof temperature and irradiance in sea-grassmeadowsgrazedbygreenturtles in theGreaterCaribbeanAllmeadowsinthisstudywerelocatedinrelativelyshallowwarmtropicalandsubtropicalenvironmentsreceivinghighlevelsofinci-dentsunlight
41emsp|emspEffects of Halophila stipulacea invasion on meadow carbon capture
In addition to the newmetabolic carbon capture estimates wecontribute from grazed areas in T testudinum meadows wealso present the first estimates of NEP from meadows domi-nated by H stipulacea seagrass Having been first reported inthe Caribbean in 2002 (Ruiz amp Ballantine 2004)H stipulacea has spread rapidly among islands and become invasive in thisregion (Christianenet al 2018Willette et al 2014)Halophila stipulaceamayalsospreadmorequicklywithinareasofseagrassgrazedbygreenturtlesthaninungrazedareas(Christianenetal2018) suggesting that grazing may inadvertently facilitate thespreadofthisinvasivespeciesonceithasbecomeestablishedin
anareaInLacBayBonairegreenturtleshavebeguntoexpandtheirgrazingplotsintoareasofpreviouslyungrazedT testudinum asH stipulacea slowly takes over the historically grazed areaswithin thebay (Christianenet al 2018 SmuldersVonk EngelampChristianen2017) If this relationshipbetweengreen turtlesnative seagrasses and invasive seagrass exists in other areas itcouldhaveimplicationsfortotalecosystemcarbondynamicsandcarbonstorageaswellasotherecosystemfunctionswithinthesemeadows
Our resultsdemonstrate that ratesofmetabolic carboncap-turemaybe similarbetweenH stipulaceameadowsand thena-tive T testudinum meadows that they are replacing (Figure 4)HowevergiventhelimitednumberofH stipulaceameadowswewereabletosample inthisstudy(n=3)cautioniswarrantedinextrapolating these findingswidely to other locations facing in-vasionbyH stipulacea seagrassWhile ratesofNEP inH stipu-laceameadowsweresimilar tobothgrazed (n=3)andungrazed(n=2)areasofT testudinum-dominatedmeadowsinStCroixandBonaire (within-site comparisons) multiple factors affect theseratesthatmaydifferandaffectthisrelationshipinotherlocations(egdominantspeciesofnativeseagrassdensityandbiomassofH stipulaceavsthenativespeciesbladeepiphytecoverage)Forexample this relationshipmay differ in areas such as Dominica(eastern Caribbean) where the meadows undergoing H stipu-lacea invasion are dominated byS filiforme seagrass (WilletteampAmbrose 2012) Measurements from additional locations willgreatlyaidinunderstandingthespatialextentofthesimilarityinmetaboliccarboncaptureratesbetweenH stipulaceaandnativeseagrassmeadowsseenhereHoweverthecomparablemetabolicratesbetweentheinvasiveandnativeseagrassesmeasuredinthisstudysuggestsomebenefitsofseagrasspresencemayberetainedinmeadowsfollowinginvasionbyH stipulaceasuchaslocalmit-igation of ocean acidification (through metabolic buffering ofpHCamp et al 2016Hendriks et al 2014Unsworth CollierHendersonampMcKenzie2012)
It is not yet known whether comparable rates of metaboliccarbon capture will translate to comparable carbon sequestra-tionor storage inmeadowsdominatedby invasiveH stipulacea UnlikeT testudinum(vanTussenbroeketal2006)seagrassspe-cieswithinthegenusHalophiladonotformadeepbelow-groundrhizome mat (Fonseca 1989) Robust rhizome mats formed bysomeseagrassspeciescontributetohigherbelow-groundcarbonstorage(Christianenetal2013)andthelackofasimilarrhizomematmayresult in lowersedimentcarbonstorageinH stipulacea meadowsHalophila tissues also decompose quickly in sediment(JosselynFonsecaNiesenampLarson1986)and thismayaffectseagrass-derivedcarboninputtothesedimentsininvadedmead-ows compared to native species with higher refractory organicmatter content (Trevathan-Tackett et al 2017) Further studieson carbondynamics inH stipulaceameadowswill beneeded toformabetterunderstandingofhowthecontinuedinvasionbythisspecieswill affect total ecosystem carbon dynamics in seagrassmeadowsintheCaribbean
emspensp emsp | emsp1111Journal of EcologyJOHNSON et al
42emsp|emspGreater global assessment of grazed meadows needed
UngrazedareasofT testudinum-dominatedseagrassmeadowsfromthisstudyexhibitedratesofNEP(median1040mmolCmminus2 dayminus1)neartheupperendofthosereportedintheliteratureforseagrassecosystemsandgrazedareasinthisstudyexhibitedrates(median381)nearthemedianvalueofNEPforallreportedungrazedsea-grassmeadows(median271Figure6)Measurementsofseagrassmetaboliccarboncapturearerelativelygeographicallylimitedhow-ever with the majority of measurementsmdashmostly from ungrazedT testudinum-dominated meadowsmdashcoming from the GreaterCaribbeanandGulfofMexicoregionsTheCaribbeanandGulfofMexicoareecologicallyimportantgrazingregionsbutonlyasingleestimateofseagrassmetabolismfromagrazedareaexistedpriortothisstudy(Johnsonetal2017)
Allmeadows in the present study exhibited a consistent re-sponseofmetaboliccarboncapturetogreenturtlegrazinghow-ever our studywas limited to the effects of grazing in tropicaland subtropicalT testudinum-dominatedmeadows It ispossiblethatmeadows inother regionsof theworldperhapsdominatedbydifferent seagrass speciesandunderdifferentenvironmentalconditionswillnotexhibitthesameresponsetograzingintermsofmetabolic carbon dynamicsWe are aware of only one otherstudy inwhichthiswas investigated Ina tropicalThalassia hem-prichiimeadow(Tanzania)NEPwasreducedasaresultofexper-imental clipping to simulategrazing (Dahletal2016)howeveronlyhourlyratesofdaytimeNEPweremeasuredWithoutcorre-spondingratesofrespirationwecannotevaluateifNEPremained
positive in thisT hemprichiimeadowoveradiel cycleand if theresponse toexperimental clipping inTanzaniawas similar to theresponse to grazing measured in our study Further studies oneffectsofgrazingonseagrassmeadowmetaboliccarbondynam-icswould be particularly beneficial in (a) additional areaswheregrazer abundance is increasing and (b) meadows dominated byspeciesotherthanT testudinum
5emsp |emspCONCLUSIONS
Given the importance of seagrass meadows as blue carbon eco-systems (Fourqureanetal2012)andtheir role incarbonseques-tration and potential climate changemitigation it is necessary tounderstand how meadow carbon dynamics are affected by graz-ing Our study adds critical information on the effects of grazingon seagrassmetabolic carbondynamics andhow theseprocessesmaybeaffectedwith increasinggreenturtleabundanceandgraz-ingWedemonstratethattheresponseinmetaboliccarboncapturetogreenturtlegrazing isconsistentacrossseagrassecosystems inthe Greater Caribbean and Gulf of Mexico regionsmdashan area thatsupportsvastexpansesofseagrass(GreenampShort2003Jackson1997Wabnitzetal2008)Thisisanecologicallyimportantgrazingregionforgreenturtlepopulationsandgivenincreasinggreenturtleabundanceacrosstheglobe(Chaloupkaetal2008Mazarisetal2017)itisexpectedthatmoreseagrassareawillreturntoanaturalgrazedstateOurresultssuggestthat increasedgrazingwill trans-latetolowerratesofmetaboliccarboncapturebutwillnotstimulaterespirationandthemetaboliclossofcarbonfromthesemeadows
F I G U R E 6 emspRates(meanplusmnSD)ofnetecosystemproductionfrommeadowsofvariousseagrassspeciescollectedfromtheliteratureOpencircleslsquocrossedrsquocirclesandopentrianglesaregrazedThalassia testudinum(n=7)ungrazedT testudinum(n=6)andHalophila stipulacea(n=3)meadowsrespectivelyfromthepresentstudyFilledcirclesarepreviouslypublishedestimatesofungrazedseagrassmeadownetecosystemproduction(NEP)andcomefromvariouslocationsaroundtheworldCriteriaforliteraturecollectionanddatainclusionaredescribedinJohnsonetal(2017)RatesofNEPfromungrazedT testudinum and H stipulaceameadowsfromthepresentstudyareneartheupperendofpublishedvaluesMedianNEPfromgrazedT testudinumareas(presentstudy)issimilartothemedianNEPofungrazedseagrassmeadowsglobally(Section42)FiguremodifiedfromJohnsonetal(2017)toincludeadditionalestimatesLettersabovepointsdenotesitesfromthepresentstudyndashBBonaireCLittleCaymanEEleutheraFFloridaSStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306

1102emsp |emsp emspenspJournal of Ecology JOHNSON et al
1emsp |emspINTRODUC TION
Seagrasses form someof themost productive ecosystemson theplanet (DuarteampChiscano1999)High ratesofmetabolic carboncapture and subsequent biomass production is one of the mainprocessesbywhichseagrassescontributetocarbonsequestration(known as lsquoblue carbonrsquo in seagrass habitats Duarte et al 2010Kennedy et al 2010) Among seagrass meadows globally abouthalfofthetotalcarbonstoredinameadowonaveragemaybede-rivedfromseagrassbiomass(Kennedyetal2010)Meadowsalsoexport large amounts of biomass annually contributing to carbonstorage in peripheral habitats (DuarteampCebriaacuten 1996DuarteampKrause-Jensen2017)Coupledwiththeabilitytostorecarbonforcenturiestomillenniathroughthecreationofahypoxicsedimentaryenvironment(MateoRomeroPeacuterezLittlerampLittler1997SerranoMateoRenomampJuliagrave2012Terradosetal1999)highlyproduc-tiveseagrassmeadowsformefficientnaturalcarbonsinks(DuarteKennedyMarbagraveampHendriks2013)andtheirprotectionhasbeensuggestedasaclimatechangemitigationstrategy(Macreadieetal2017Murdiyarsoetal2015)
There is an estimated150000 km2 of seagrass habitat acrossthe Caribbean Sea Gulf of Mexico and The Bahamas (Green ampShort 2003 Jackson 1997 Wabnitz Andrefouet Torres-PullizaMuller-Karger amp Kramer 2008) Thesemeadows provide import-ant foraging habitat for numerous species (Hemminga amp Duarte2000 Ogden 1976 Scott et al 2018) including green turtles(Chelonia mydas) which were historically abundant in this regionpriortooverexploitationbyhumans(Jackson1997Jacksonetal2001)GreenturtleabundanceiscurrentlyincreasinginareassuchastheCaribbean (egnestnumbersroughlytripled1970ndash2005attheCaribbeanslargestrookery)asaresultofsuccessfulconserva-tionefforts (Chaloupkaetal2008MazarisSchofieldGkazinouAlmpanidouampHays 2017) Thiswill lead tomore seagrass areasbeing returned toanaturalgrazedstate (Figure1a)Green turtlesin theGreaterCaribbean regionnormally foragebycreatinggraz-ingplotswithinseagrassmeadowsinwhichtheycropthebladestoshortheightsabovethesedimentsurfaceandcontinuallyre-grazetheseareastoconsumenewtissuegrowth(Bjorndal1980Ogden1980)Thisgrazingstrategy leads toa reductionof thephotosyn-thetic biomass in a meadow At high densities green turtles arecapableofgrazinganentiremeadowandhavebeenrecordedcon-suming100ofnewseagrassgrowthdaily(Christianenetal2012)Giventhedesiretoconserveseagrassesfortheirabilitytosequesterandstorecarbon(Macreadieetal2017)andtheexpectationthatmoreseagrasswillreturntoanaturalgrazedstate inthefuture itisnecessarytounderstandhowmetaboliccarboncapture (carbonfixedthroughphotosynthesisormineralizedthroughrespiration)isaffectedbygrazingacrossseagrassmeadows
Much of the research on seagrassmetabolic carbon dynamicsto date has focused on differences among species (LindeboomampSandee1989MurrayampWetzel1987PollardampMoriarty1991)orbetween seagrasses and unvegetated sediments (Barroacuten DuarteFrankignoulleampBorges2006RheubanBergampMcGlathery2014
Stutes Cebrian Stutes Hunter amp Corcoran 2007) with little at-tentionbeinggiventograzedareasInaCaribbeanThalassia testu-dinummeadowgrazedbygreen turtles ratesofmetaboliccarboncapture were found to be lower compared to ungrazed seagrass(Johnson Gulick Bolten amp Bjorndal 2017) However long-term
F I G U R E 1 emspAThalassia testudinumseagrassmeadowwithanareagrazedbygreenturtles(panelaleftside)andanadjacentungrazedarea(panelarightside)inEleutheraTheBahamasTheinvasiveseagrassHalophila stipulaceainStCroixUSVirginIslands(b)AlightincubationchamberdeployedinagrazedareainBonaire(c)PhotosRJohnson[Colourfigurecanbeviewedatwileyonlinelibrarycom]
(a)
(b)
(c)
emspensp emsp | emsp1103Journal of EcologyJOHNSON et al
grazing(greaterthanayear)didnotleadtoaproportionalincreaseinbenthicrespirationandremineralizationofcarbonstored inthemeadow possibly due to lack of a significant heterotrophic com-munity These results are from a single Caribbean location (LittleCayman Cayman Islands) that has some of the highest recordedmetabolicratesamongseagrassmeadows(Johnsonetal2017)ItisnotknownifgrazinghassimilareffectsacrossseagrassmeadowsinotherlocationsorunderdifferentenvironmentalconditionsAbet-terunderstandingofthevariabilityamongmeadowsinresponsetograzingisneededtounderstandhowincreasinggreenturtleabun-danceandincreasedgrazingwillaffectseagrasscarboncapture intheGreaterCaribbeanregion
In addition to increased grazing seagrass meadows in theCaribbeanarealsobeinginvadedbytheseagrassHalophila stipula-cea(Figure1b)ThisseagrassspeciesmdashnativetotheIndianOceanmdashhas invaded themeadowsofmanyCaribbean islands following itsintroductiontotheregionin2002(RuizampBallantine2004Willetteetal2014)Initialresearchsuggeststhatgreenturtlesavoidgrazingthis invasiveseagrass infavourofnativespeciesandthatH stipu-laceamayspreadmorerapidlywithinareasofmeadowsgrazedbygreen turtles compared to areas left ungrazed (Christianen et al2018)Little isknownabouthowinvasionbyH stipulaceamayaf-fectcarbondynamicsorecosystemfunctioninginCaribbeanmead-owshowever(VianaSiriwardane-deZoysaWilletteampGillis2019)GiventhisinteractionbetweengrazingandH stipulaceaitiscriticaltounderstandhowcarbondynamicsareaffectedinmeadowswhereinvasionbythisspeciesisoccurringinadditiontograzing
Ourprimaryobjectiveinthisstudywastoevaluatethehypoth-esis that effectsof green turtle grazingonmetabolic carbon cap-tureratesareconsistentacrossseagrassmeadowsWealsoevaluatevariationinthestrengthoftheresponsetograzingamongmeadowsand how rates ofmetabolic carbon capture in invasiveH stipula-ceameadowscomparetonativemeadowswhereturtlesgrazeWecomparedratesofnetecosystemproduction(NEP)betweengrazedandungrazedareasofT testudinumseagrassmeadowsinfourloca-tionsaroundtheGreaterCaribbean(definitionfromPetuch2013Robertson amp Cramer 2014) and Gulf ofMexico with establishedgreen turtle grazing populations (in addition to those previouslysampled inLittleCayman)Thefivetotal locationsencompassedawidegeographicalareaandspannedarangeofenvironmentalandseagrassmeadowcharacteristicsrepresentativeofgreenturtlefor-agingareasinthisregionTheseagrassH stipulaceahadestablishedattwooftheselocationsandwecomparedratesofNEPinmead-owsofthis invasivespeciestothosemeasuredinbothgrazedandungrazedareasofnativemeadowsattheselocations
2emsp |emspMATERIAL S AND METHODS
21emsp|emspStudy sites
We sampled grazed and ungrazed areas ofT testudinum seagrassmeadowsatfivelocationsacrosstheGreaterCaribbeanandGulfofMexicowheregreenturtleshadestablishedforagingareas(Figure2)
OneoftheselocationsLittleCaymanCaymanIslandswassampledthreetimesduringJuneandJuly2016aspartofapreviousstudy(Johnsonetal2017)The remaining four locationsweresampledin2018StCroixUSVirginIslands(February)westcoastofFlorida(GulfofMexico side)USA (May)BonaireCaribbeanNetherlands(July)EleutheraTheBahamas(August)Weopportunisticallysam-pledtwoseparatesitesinbothBonaireandEleutheraandonesiteatallotherlocations(timeandorweatherconditionsdidnotallowsamplingadditionalsitesattheselocations)EstablishedmeadowsoftheseagrassH stipulaceawerepresentattwoofthelocationsmdashStCroixandBonairemdashandthesemeadowswerealsosampledforcom-parisontonearbygrazedandungrazedareasofnativeT testudinum meadows(oneofthesitesinBonaireonlyhadnearbygrazedT tes-tudinum for comparison) Coordinates and general environmentalcharacteristics (daylight hours water depth salinity temperatureandirradiance)forallsitesaregiveninTable1
Themeadowssampledinthisstudyvariedgreatlyinsizefromrelatively small (lt150m2) for the grazedT testudinum area at thenorthwestLacBaysiteinBonairetoverylarge(severalsquarekilo-metresofunbrokenseagrasshabitat)fortheungrazedT testudinum meadowattheNorthRacksiteinFlorida(westcoast)Meadowsatothersitesencompassedarangeofsizesbetweenthesetwosuchas~350m2 each for the grazed andungrazedT testudinum areasinLittleCaymanand~600m2forthegrazedareaandgt50000m2 for the ungrazed T testudinum meadow at the Arvida Bay site inEleuthera TheBahamasAt each site a grazed area and adjacentungrazedareawereselectedforsamplingthatwererepresentativeofthemeadowSamples(Sections22and23)werecollectedwithinseveralmetresoftheborderbetweenthegrazedandungrazedareastoavoidpossibleedgeeffectswhileensuringsimilarenvironmentalconditions
Allsampledgrazedareas(naturalgreenturtlegrazingplots)werenearly uniformly grazed The proportion of grazed T testudinum blades ranged from95 to 100 in all grazed areas Evidence ofgrazingbygreenturtleswasnotobservedinungrazedareasexcept
F I G U R E 2 emspMapofseagrassmeadowsamplinglocationsTwositesweresampledineachofBonaireandEleutheraandonesitewassampledateachoftheotherthreelocationsCircledsymbolsforStCroixandBonairedenotelocationswhereHalophila stipulaceameadowswerealsosampled
1104emsp |emsp emspenspJournal of Ecology JOHNSON et al
forinStCroixwhere~3ofT testudinumbladesshowedsignsofgrazing(croppedbladetips)Somefishesalsograzeonseagrassandevidenceoffishgrazing(egbitemarks)wasobservedinungrazedT testudinum meadows at some sites Biomass removal from fishgrazingwasminorcomparedtoturtlegrazinghoweverandwasnotquantifiedNoevidenceofgreenturtlegrazingwasobservedinanyH stipulaceameadows
WesampledsevenareasfromT testudinum-dominatedseagrassmeadowsthatdisplayedsignsofactivegreenturtlegrazing(hereaf-tergrazedareas)sixareasfromT testudinum-dominatedmeadowsthatwerenotgrazed(hereafterungrazedareas)andthreemeadowsthatweredominatedbytheinvasiveseagrassH stipulacea(hereaf-terH stipulaceameadows)Thesemeadowswere spreadacross awideareaoftheGreaterCaribbeanandGulfofMexicoregionsandvariedintheirbioticandabioticcharacteristicsSomeofthesampledseagrassmeadowssuchastheT testudinummeadows inBonairewerelargelymonospecificwhereasothersweredominatedbyonespecies(T testudinum or H stipulacea)withinterspersedSyringodium filiforme or Halodule wrightii seagrasses and various macroalgae(TableS1)Thesubstrateinallmeadowswasprimarilycalciumcar-bonate sand Additional meadow characteristics are provided inResults (Section31)Data from theLittleCayman sitehavebeenreportedpreviously(Johnsonetal2017)andarepresentedagainhereforcomparisontotheothersites
22emsp|emspSampling seagrass meadow characteristics
Seagrass meadow and environmental characteristics were sam-pledateachsiteatthetimemetabolicincubationswereconducted
(Section23)Environmentaltemperatureandirradianceweremeas-uredduringmetabolicincubationsatseagrasscanopyheight(5-minintervals) at each site with a HOBO Pendant data logger (OnsetComputer Corporation) A water sample was collected at canopyheight and salinity wasmeasured with a handheld AgTec SalinityRefractometer that was calibrated with freshwater each day be-fore sampling (Agriculture Solutions Strong ME no sample wascollectedattheFloridasite)Meadowdepthwasmeasuredateachlocationwitheitheratapemeasureorascubadiversdepthgauge(accuracyplusmn10cm)
Seagrassspeciescompositionshootdensityblademorphome-try(lengthwidthandsurfacearea)andabove-groundbiomassweredetermined in all sampled areas (grazed ungrazed H stipulacea)within1metreofwhere incubationchamberswereplacedShootdensitiesweremeasuredusing25times25cmquadrats(00625m2area)in T testudinummeadowsDuetothehighdensitiesofH stipulacea10times10cmquadrats(001m2area)wereusedtomeasureshootden-sitiesinmeadowsdominatedbythisspeciesAbove-groundbiomasssampleswerecollectedusing10times10cmquadrats(allmeadows)byclippingall bladesat the sediment surfacewith scissors Six repli-catequadrats (forbothshootdensityandbiomass)werecollectedfromallsampledareasateachsiteexceptfortheStCroix(n=5)andFlorida(n=3)sites(Table2)Bladelengthandwidthweremea-suredforthedominantspecies(T testudinum or H stipulacea)from30randomlyselectedseagrassbladesfromthesebiomasssamplesforeachareaBladesurfacearea(iephotosyntheticareaperblade)wascalculatedastwotimestheproductofbladelengthandwidth(for each species) Followingmeasurement all bladeswere gentlyscrapedcleanofsedimentsandepiphytes(lowepiphyteloadsatall
TA B L E 1 emspCoordinatesofeachsamplingsiteandenvironmentalparametersmeasuredintheseagrassmeadowsondaysofmetabolicincubations
Site
Latitude Longitude Daylight Depth Salinity Temperature Irradiance
Decimal degrees hours M permil degC Lux
Bonaire
LacCaiBeach 12104417 minus68223183 1275 10 35 300 324518
NWLacBay 12108546 minus68231562 1275 14 36 311 432217
StCroix
BIRNM 17784612 minus64624566 115 45 35 279 211904
LittleCayman
GrapeTreeBay 19696518 minus80059652 13 10 36 319 551840
19696518 minus80059652 13 10 37 317 465732
19696518 minus80059652 13 10 38 336 600715
Eleuthera
ArvidaBay 24722297 minus76190644 125 24 39 322 267648
HalfSound 24936641 minus76153420 125 06 36 344 717215
Florida
NorthRack 28564760 minus82779335 1375 20 mdash 292 409801
Note TemperatureandirradiancedataaremeanvaluesmeasuredduringincubationsTheLittleCaymansitewassampledthreetimesin2016Allothersitesweresampledoncein2018SamplingtimesaregiveninSection21AbbreviationBIRNMBuckIslandReefNationalMonumentsite
emspensp emsp | emsp1105Journal of EcologyJOHNSON et al
sites) rinsed infreshwateranddriedtoaconstantweightat60degCbeforeweighingfordrymass
23emsp|emspEcosystem metabolism measurements
Metaboliccarbondynamicsoftheseagrassecosystemsweremeas-uredusingbenthic incubationchambers (Figure1c)similartopre-viousstudies (BarroacutenMarbagraveTerradosKennedyampDuarte2004CallejaBarroacutenHaleFrazerampDuarte2006Johnsonetal2017OliveacuteSilvaCostaampSantos2015)ChamberswereconstructedbyinsertingaPVCcylinder(16cmdiameter002m2area)~75cmintothe sediment (attempting to avoid severing horizontal rhizomes)and attaching a flexible gas-tight polyethylene bagwith samplingporttothetop(HansenThamdrupampJoslashrgensen2000)Theuseofflexiblegas-tightbagsinchamberconstructionallowsthepropaga-tionofwave turbulence to the chambers interior tomore closelysimulateenvironmentalconditionsIncubationchambervolumewasmeasuredinthelaboratorytobe55ndash6LOnsamplingdayscham-berswere set up in themeadows between 1030 and 1130 andincubationswererunfor25ndash3hrThisincubationlengthwaschosenbecausesaturationeffectscanoccurwithinchambersduringlonger
incubationsandhavebeenshowntounderestimatemetabolicrates(Oliveacuteetal2015)Metabolicdynamicsweremeasuredthreetimesat2-weekintervalsinLittleCaymanaspartofapreviousstudyandshown to be relatively stable among sampling events (Johnson etal2017) Incubationswerethereforeconductedonceateachad-ditionalsiteinthepresentstudy
Wesetupthreelight(clear)andthreedark(opaque)chambersineacharea(grazedorungrazedT testudinumorH stipulacea)tomea-sureecosystemmetabolicrates(n=3forallareas)LightchamberswereusedtomeasureratesofNEPanddarkchamberswereusedtomeasure ratesofecosystemrespiration (RE)Metabolic rateswereestimatedfromchangesindissolvedoxygen(DO)concentrationin-side the chambersbetween thebeginningandendof the incuba-tionperiodThreewatersampleswerecollectedfromthechamberviathesamplingportin60-mlplasticsyringesatthebeginningandendoftheincubationUponcollectionsyringeswerecappedwitha silicon cap andbrought to the surfacewhereDOconcentrationwas measured directly in the syringe with an optical DO probe(Vadeboncoeur2011YSIProODOYellowSpringsOhio)TheDOprobewas calibrated at the beginning of each sampling daywithwater-saturatedair
TA B L E 2 emspSeagrasscharacteristicsofgrazedandungrazedareasofThalassia testudinummeadowsandHalophila stipulaceameadowsmeasuredateachsamplingsite
Site Meadow n
Seagrass density Canopy heightBlade surface area AG biomass
shootsm2 cm cm2 g DMm2
Bonaire
LacCaiBeach Grazed 6 7413plusmn1263 47plusmn22 63plusmn32 271plusmn76
Ungrazed 6 9547plusmn937 96plusmn39 214plusmn116 1309plusmn544
H stipulacea 6 38500plusmn8068 21plusmn06 19plusmn09 364plusmn133
NWLacBay Grazed 6 2373plusmn446 28plusmn14 28plusmn14 48plusmn28
H stipulacea 6 34833plusmn5492 20plusmn04 18plusmn06 171plusmn59
StCroix
BIRNM Grazed 5 19648plusmn2261 40plusmn23 44plusmn27 274plusmn102
Ungrazed 5 24064plusmn2501 81plusmn31 97plusmn45 516plusmn110
H stipulacea 5 37336plusmn12494 28plusmn05 32plusmn09 630plusmn240
LittleCayman
GrapeTreeBay Grazed 6 9573plusmn1485 20plusmn09 23plusmn12 123plusmn37
Ungrazed 6 10187plusmn2102 149plusmn85 307plusmn204 1832plusmn948
Eleuthera
ArvidaBay Grazed 6 16480plusmn3927 35plusmn19 42plusmn22 418plusmn139
Ungrazed 6 20533plusmn3299 152plusmn46 258plusmn88 2022plusmn348
HalfSound Grazed 6 16747plusmn3167 18plusmn07 14plusmn06 88plusmn20
Ungrazed 6 17840plusmn2847 94plusmn20 108plusmn31 965plusmn162
Florida
NorthRack Grazed 3 13227plusmn4441 25plusmn11 16plusmn08 93plusmn24
Ungrazed 3 13867plusmn1619 89plusmn27 78plusmn29 707plusmn233
Note nisnumberofreplicatesateachsiteValuesaremeansplusmnSDCanopyheightismeasuredfrommeanbladelengthBladesurfaceareaisperbladeofseagrassAbbreviationsAGabove-groundBIRNMBuckIslandReefNationalMonumentsiteDMdrymass
1106emsp |emsp emspenspJournal of Ecology JOHNSON et al
Tomeasureonlymetabolicratesoftheseagrassbenthiccompo-nentsofthesystem(thoseaffectingcarbonstorage)wecorrectedrates measured from incubation chambers for water column me-tabolismWemeasuredwater columnmetabolismusingbiologicaloxygendemand(BOD)bottlesatthesametimeasbenthicchamberincubationsThreeclear300-mlglassBODbottlesandthreeopaque300-mlglassBODbottleswereusedtomeasurewatercolumnpro-ductionandrespirationrespectivelyBottleswerefilledwithwaterat seagrasscanopyheightanchored to thebottomand incubatedunderinsituconditionsintheseagrassmeadowWatercolumnsam-pleswerecollectedatthebeginningofincubationsin60-mlsyringesatcanopyheightandDOconcentrationwasmeasuredinthesamemannerassamplesfromincubationchambersFollowingthe incu-bationperiodbottleswerecollectedand returned to thesurfaceonesamplewascollectedfromeachbottlewithasyringeandDOconcentrationwasmeasured
Hourlymetabolic rateswere calculated from thedifference inDOconcentrationwithinlight(NEP)anddark(RE)incubationcham-bersbetweenthebeginningandendoftheincubationperiodWatercolumn metabolic rates measured from BOD bottles were sub-tractedfromratesmeasuredinchambersHourlygrossprimarypro-duction(GPP)wasthencalculatedasthesumofhourlyNEPandRE DailyratesofGPPwerecalculatedbymultiplyinghourlyGPPbythelengthofthephotoperiodmultipliedby075(toaccountfor lowermetabolicratesatdawnandduskJohnsonetal2017Oliveacuteetal2015)anddailyratesofREwerecalculatedbymultiplyinghourlyRE by24hoursDailyNEPwascalculatedas thedifferencebetweendaily GPP and RE Therewere a few occasionswhen an increaseinDO concentrationwasmeasuredwithin a dark chamber or thedarkBODbottlesduringanincubationsuggestinganerrorThesemeasuredincreasesweresmallusuallywithintheprobesmarginoferror (01mgO2L)andweassumedrespirationtobezeroduringthese timesCorrecting for these respiration errors hadminor ef-fectsondailymetabolic rates in themeadows (TablesS2andS3)MeasuredDOconcentrationswereconvertedfrommgO2tommolO2andthentocarbonunits(mmolC)assumingphotosyntheticandrespiratoryquotientsofone(BarroacutenampDuarte2009)
24emsp|emspData analyses
Net ecosystem productionwas themetabolic variable of primaryinterestasitaccountsforbothcarboncaptureandlossandcanbeusedas an indicatorofwhether a system is currently ametaboliccarbonsinkorsourceThereforeNEPisthemetabolicvariableforwhichstatisticalresultsarepresented
Ouraimwastoevaluatewhethergreenturtlegrazinghasacon-sistenteffectonmetabolicratesacrossseagrassecosystemsmdashthatistoevaluatewhetheratanygivensiteatanygiventimeratesofNEPareloweringrazedareasthaninungrazedareasWeevaluateddifferences in ratesofNEPbetweenareasofgrazedandadjacentungrazedT testudinum from each sitewith paired t tests T testswereusedbecause thecomparisonof interestwas thedifferencebetween grazed and ungrazed areas (effect of grazing) within a
givensitenotdifferencesinNEPamongsitesForsiteswheretheinvasive seagrass H stipulacea was present we used one-wayANOVAstotestforsignificantdifferencesinratesofNEPbetweenH stipulaceameadowsandnearbygrazedandungrazedT testudinum areas(StCroixsiteandLacCaiBeachsiteBonaire)Forthenorth-west site in Lac Bay Bonaire where only a grazed area of T te-studinum and an invasive H stipulacea meadow were comparedanunpaired t testwasused IfANOVA resultswere significant aTukeyspost-hoctestwasusedtoidentifysignificantcomparisonsWetestedfordifferencesinseagrassmeadowparametersbetweengrazedandungrazedT testudinumareasandH stipulaceameadowsatsitesusingpairedttestsandANOVAsinthesamemannerasforNEPWeused linearregressiontotestrelationshipsbetweenNEPand explanatory variables (eg seagrass biomass environmentaltemperature)MeanvaluesforNEPandexplanatoryvariablesfromeachmeadowwereusedforlinearregressionstestssotherewasasingledatapointforeachgrazedandungrazedT testudinum area or H stipulaceameadowateachsamplingsite
AllanalyseswereperformedinRversion343(RCoreTeam2018)Datawere processed using the lsquodplyrrsquo package (Wickham FrancoisHenryampMuumlller2017)andpost-hocanalyseswereperformedusingthelsquoagricolaersquopackage(deMendiburu2017)Alldatamettheassump-tionsfortheseanalysesanddataarepresentedasmeanplusmnSD
3emsp |emspRESULTS
31emsp|emspSeagrass meadow characteristics
Total seagrass shoot densities varied widely among meadowsand locations (Table 2) though shoot densitieswere always high-est inH stipulaceameadows The lowest shoot density observed(2373plusmn446shootsm2)wasinagrazedarea(NWsiteinLacBayBonaire)buttherewasnocleartrendforhigherorlowerseagrassdensities between grazed and ungrazed areas across the sampledsitesThalassia testudinum density in LittleCaymanwasgreater inthe grazed area than ungrazed area (paired t test p = 02) whileT testudinumdensityattheLacCaiBeachsiteinBonairewasgreaterintheungrazedarea(pairedttestp=02)Thalassia testudinum den-sitydidnotdifferbetweengrazedandungrazedareasattheremain-ingsites(pairedttestspgt05)ShootdensityofH stipulaceawasnot significantly different between any of the threeH stipulacea meadowsmeasured(ANOVAF2=009p=92)
HalophilaseagrassestypicallygrowshortbladesCanopyheightoftheH stipulaceameadowwassimilartothatofthenearbygrazedarea inStCroix andwas shorter than thenearbygrazedareasatbothsitesinBonaire(Table2)Halophila stipulaceabladelengthwasgreaterinStCroixthanBonaire(ANOVAF2=2413plt01)AreasofT testudinum-dominatedseagrassmeadowsgrazedbygreentur-tleshad significantly shorterblades and therefore canopyheightthan adjacent ungrazed areas at all sampled sites (paired t testsplt05forallcomparisonsTable2)
Greenturtlesbycroppingseagrassbladesnearthesedimentsur-faceandreducingcanopyheightgreatlyreducetheabove-ground
emspensp emsp | emsp1107Journal of EcologyJOHNSON et al
seagrass biomass of meadows within their grazing areas Above-ground biomass ranged from 48 plusmn 28 to 418 plusmn 139 g DMm2 amonggrazedareas(Table2)andfrom516plusmn110to2022plusmn348gDMm2amongungrazedareasBiomasswasalwaysloweringrazedareas than adjacent ungrazed areas (p lt 05 for all comparisons)Biomass among the three sampledH stipulaceameadows rangedfrom171plusmn59to630plusmn240gDMm2TheH stipulacea meadow in St Croix had significantly greater above-ground biomass thaneither of theH stipulaceameadows sampled inBonaire (ANOVAF2=1202plt 01)Evidenceofgreenturtlegrazingwasonlyob-served in T testudinum-dominated meadows in this study notH stipulaceameadowsandgrazingresultedinagt79decreaseinbiomassingrazedcomparedtoungrazedareasatallsitesexceptStCroix(47Table3)
32emsp|emspEcosystem metabolic rates
Rates of NEP differed among meadow types (grazed ungrazedH stipulacea) across the studied locations (Figures 3 and 4 seeTable S2 for data) Across all sites NEP ranged from 42 plusmn 70 to517plusmn110mmolCmminus2 dayminus1amonggrazedareasandfrom523plusmn57to2253plusmn191mmolCmminus2 dayminus1amongungrazedareasThelargestandarddeviationsonsomeoftheungrazedareaNEPestimatesfromLittleCaymanwerearesultofoneincubationchamberproducingratesmuchdifferentfromtheothertwoAcrossallsitesNEPwasconsist-entlyloweringrazedareasthanungrazedareas(Figure3)DifferencesinNEPbetweengrazedandungrazedareasweresignificantat four
ofthesixsiteswhereungrazedT testudinumwaspresentStCroixLittleCayman (all threemeasurement times) theArvidaBay site inEleutheraandFlorida(pairedttestsplt05forallcomparisons)AttheLacCaiBeachsiteinBonaireandtheHalfSoundsiteinEleuthera
TA B L E 3 emspPercentdifferenceinmetabolicratesandabove-ground(AG)seagrassbiomassbetweengrazedandadjacentungrazedThalassia testudinumareas
Site
GPP RE NEP AG Biomass
difference
Bonaire
LacCaiBeach 818 909 656 793
StCroix
BIRNM 604 783 560 469
LittleCayman
GrapeTreeBay 922 955 891 933
899 796 960 933
885 870 900 933
Eleuthera
ArvidaBay 543 395 629 793
HalfSound 801 889 679 908
Florida
NorthRack 615 413 919 868
Note TheLittleCaymansitewassampledthreetimesin2016Allothersitesweresampledoncein2018SamplingtimesaregiveninSection21AbbreviationsBIRNMBuckIslandReefNationalMonumentsiteGPPgrossprimaryproductionNEPnetecosystemproductionREecosys-temrespiration
F I G U R E 3 emspRates(meanplusmnSD)ofnetecosystemproduction(a)grossprimaryproduction(b)andecosystemrespiration(c)fromgrazed(openpoints)andadjacentungrazed(solidpoints)areasofThalassia testudinumseagrassmeadowsRatesofNetecosystemproduction(NEP)weresignificantlyloweringrazedareasthanungrazedareasatallsitesexceptatLacCaiBeachBonaireandHalfSoundEleuthera(Section32)SitesorderedsouthtonorthBONBonaire(LCBLacCaiBeachsite)STXStCroixLCLittleCaymanELEleuthera(ABArvidaBaysiteHSHalfSoundsite)FLFloridaDashedlinein(a)denotesmetabolicbalance(NEP=0)Valuesabovelinerepresentnetmetaboliccarboncapture[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1108emsp |emsp emspenspJournal of Ecology JOHNSON et al
differencesinNEPbetweengrazedandungrazedareaswerenearlysignificant(pairedttestsp = 08 and p=09respectively)
Though NEP was significantly lower in grazed than ungrazedareas atmost sites rates ofNEPwere almost always positive asecosystem respiration was not stimulated in relation to primaryproductionevenwithconsiderablereductionofphotosyntheticsea-grassbiomassbygrazing(gt79biomassreductionatallsitesexceptStCroix Table3)NEP ranged from56 to96 lower in grazedareasthanungrazedareas(Table3)Asimilarrangeinthedifference
betweengrazedandungrazedareaswasmeasuredforgrossprimaryproduction (54ndash92 lower in grazed) and ecosystem respiration(40ndash96loweringrazed)
Halophila stipulacea meadows were present at three sites andratesofNEPintheseinvasiveseagrassmeadowswerecomparedtonearbynativeT testudinum-dominatedmeadowsNEPintheH stip-ulaceameadowswascomparedtonearbygrazedandungrazedareasofT testudinum-dominatedmeadowsattwoofthesites(StCroixandtheLacCaiBeachsiteinBonaire)WithineachsiteratesofNEPintheH stipulaceameadowwerebetweenthoseofthegrazedandungrazedareas (Figure 4) but the differences were not significant (ANOVApgt10forallpost-hoccomparisonswithineachsite)AnungrazedareawasnotavailableforcomparisonatthethirdsiteatwhichH stipula-ceawaspresent (NWLacBaysite inBonaire)asallT testudinumatthissitewasgrazedbygreenturtlesandratesofNEPdidnotdifferbetweenthe invasive-andnative-dominatedareasatthissiteeither(t test p = 56 Figure 4)Halophila stipulaceameadowNEP rangedfrom612plusmn226to989plusmn233mmolCmminus2 dayminus1(TableS2)anddidnotdiffersignificantlyamongthethreesitesatwhichitwaspresent(ANOVAF2=192p=23)eventhoughtheestablishmentofH stip-ulaceaseagrassmeadowsinStCroixwasrelativelyrecent(AGulickunpublisheddata)comparedtothoseinBonaire(Willetteetal2014)
33emsp|emspDrivers of metabolic rates
VariationinratesofseagrassecosystemmetabolismacrossgrazedandungrazedareasappearedtobedrivenbysomecharacteristicsoftheseagrassmeadowsbutnotbyenvironmentalfactorsMeadowNEPwasstronglypositivelyrelatedtoabove-groundseagrassbio-massacrosssites(linearregressionR2=82plt01Figure5a)NEP
F I G U R E 4 emspRates(meanplusmnSD)ofnetecosystemproductioninmeadowsdominatedbytheinvasiveseagrassHalophila stipulacea (triangles)comparedtonearbygrazed(opencircles)andungrazed(solidcircles)areasofThalassia testudinummeadowsBONBonaire(LCBLacCaiBeachsiteNWnorthwestLacBaysite)STXStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
F I G U R E 5 emspRelationshipbetweennetecosystemproductionandabove-groundseagrassbiomass(a)andtotalseagrassshootdensity(b)fromgrazed(openpoints)andungrazed(solidpoints)areasofThalassia testudinummeadowsSolidlinein(a)isthesignificantlinearregression(R2=82plt01)betweennetecosystemproduction(NEP)andbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows
emspensp emsp | emsp1109Journal of EcologyJOHNSON et al
wasalsopositivelycorrelatedwithseagrassmeadowcanopyheight(R2=84plt01)andbladesurfacearea(R2=92plt01)Above-groundbiomassislikelythemoreusefulpredictorofmeadowNEPhoweverasbiomassisinfluencedbybothcanopyheightandbladesurfaceareaandachange ineitherofthesevariablesresults inachangeinbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows(linearregressionR2lt01p=96Figure5b)norwerethererelationshipsbetweenNEPandenvironmentaltempera-ture(R2=01p=70)irradiance(R2lt01p=10)ormeadowdepth(R2lt01p=77)acrosssites(FigureS1)
4emsp |emspDISCUSSION
Through a comparative study of seagrass meadowsmdashgrazed andungrazedmdashacross a wide area encompassing sites in the GreaterCaribbean and Gulf of Mexico we found that green turtle graz-inghas a consistent effecton themetabolic carbon capture ratesofT testudinum-dominatedseagrassmeadowsRatesofmetaboliccarboncapturewerealwayslowerinareasgrazedbyturtlesthanun-grazedareasofmeadowsmdashupto96inLittleCaymanmdashbutratesofNEPwerealwaysnearorabovemetabolicbalanceEcosystemres-pirationwasnotstimulatedinrelationtoprimaryproductionwithinareas grazed by green turtles regardless of the amount of above-groundseagrassbiomassremovedthroughgrazingorotherdiffer-encesinbioticandabioticfactorsamongsitesThesefindingslendadditionalsupporttothehypothesisthatfutureratesofmetaboliccarboncapturemaybelowerinareasgrazedbyturtlesbutgrazingwillnot stimulateecosystemrespirationand result ina large rem-ineralizationof carbon currently stored in these seagrass habitats(Johnsonetal2017)
The similarity in results between St Croix (awintermeasure-ment)andtheotherfourlocationsinthepresentstudy(allsummermeasurements)strengthensourconclusionthatgrazingresultsinaconsistent response inmetaboliccarboncaptureacrossmeadowsHoweverwewerenotable toaddresspotential temporaldynam-icswithinsitesthatmayaffectnetcarboncaptureoveranannualcycle in these systems Future studies measuring carbon captureoverlongertemporalscales(ieseasonal)maybebeneficialtoourunderstanding of the effects of grazing on within-system carbondynamics
Thoughtheresponse inmetabolic rates tograzingwasconsis-tent acrossmeadows the strengthof this responsevariedamongindividual sites Some sites such as Little Cayman had consider-ablylowerratesofNEPingrazedareasthanungrazedareas(mean917)whereastheresponsetograzing(differenceinNEPbetweengrazedandadjacentungrazedareas)waslesspronouncedatothersites suchasStCroix (Table3) Seagrassmeadowcharacteristicssuch as shoot density canopy height and above-ground biomassare likely to bemore variable among ungrazed areas than grazedareasasgreenturtlegrazingresults insimilarcanopymorphome-tryacrossmeadowsintheGreaterCaribbeanregion(egBjorndal1980HernaacutendezampvanTussenbroek2014Williams1988)Given
the strong relationship between NEP and above-ground biomass(Figure 5a) high variability in NEPwould be expected across un-grazedmeadows(egFigure3a)IndeedtherangeinratesofNEPamongungrazedareaswasnearlythreetimesgreaterthantherangemeasuredacrossgrazedareasinthisstudyAssiteswerespecificallychosen that differed from each other in meadow characteristicsvariationinthestrengthofthemetabolicresponsetograzingwouldbeexpected
Variability in the strength of the metabolic response to graz-ing resulted in the lackofa significantdifferencebetweengrazedandungrazedareasattwositesmdashLacCaiBeachinBonaireandHalfSoundinEleuthera(Figure3a) It ispossiblethatotherfactorsnotaccounted for here such as local environmental factors or greenturtlegrazinghistory(egintensitylongevityofplotuse)mayhaveledtohighervariabilityinmetabolicratesatsomesitesandlackofastatisticaldifferenceInadditionmetabolicratesmayexhibitwithin-meadowvariation(patchiness)similartobelow-groundcarbonstor-age in meadows (OreskaMcglathery amp Porter 2017) leading tohighervariabilityinestimatesandhelpingtoexplainthelackofasig-nificantdifferenceinNEPatthesetwositesThiscouldbedriveninpartbywithin-meadowspatialheterogeneityinabove-groundbio-massgiventhestrongrelationshipbetweenbiomassandNEPHighwithin-meadow variability inmetabolic rates if presentmay alsohelpexplainthe largevariabilitymeasured intheungrazedarea inLittleCaymanFuturestudiesusingtheeddycorrelationtechnique(Bergetal2003) tomeasureecosystemmetabolismmaybeabletoovercomesomeofthiswithin-meadowvariabilityhoweverthegreatexpenseofthismethodmakesitprohibitiveformanystudies
Seagrass ecosystem metabolism is commonly estimated usingoxygendynamicsmeasuredwithbenthicincubationchambers(egBarroacutenetal2004Gaciaetal2005presentstudy)howeverthereare some limitations to thismethodBenthic chambersmay severbelow-ground rhizomesand release carbon that couldaffectmet-abolicestimatesIntactrhizomesontheotherhandcanaffectthediffusiveoxygenlossfromtheseagrasstotherhizosphere(BorumSand-JensenBinzerPedersenampGreve2006)andmayresultinanunderestimateofgrossprimaryproduction(andthereforeNEP)Thismaybeminimizedusingshorter incubation timesas in this studyIncubation length also strongly affects metabolic rates estimatedfromoxygendynamics (Oliveacute et al 2015)with longer incubationtimesbiasingestimatestowardsnetheterotrophyInadditiontoox-ygenmetabolismotherprocessessuchascarbonatedynamics(VanDamLopesOsburnampFourqurean2019)play important roles intotal ecosystem carbondynamicswithout affecting themeasuredproductivityofameadow
CaribbeanT testudinummeadows are highly productive (egJohnsonetal2017KochampMadden2001)andtherelationshipbetween biomass andNEP observed in this study suggests thatmeadows may maintain positive productivity as long as photo-synthetic biomass remains A threshold biomass below whichCaribbean meadows may become net heterotrophic cannot bedirectly addressed here but could be predicted to be near zeroThisisincontrasttoapreviouslysuggestedabove-groundbiomass
1110emsp |emsp emspenspJournal of Ecology JOHNSON et al
threshold(41gDMm2)belowwhichmeadowsmaybecomehet-erotrophic (Duarte et al 2010) However that threshold wascalculated for seagrasses globally and included less productivespecies Additional studies of the relationship between biomassandmeadowNEP from seagrass species and regions other thanthoseinthepresentstudywouldbebeneficialtofurthergeneral-izehowgrazingaffectsmetaboliccarboncaptureacrossseagrassecosystems
Environmental temperature and irradiance are strong driv-ers of seagrassmeadow productivity (Calleja et al 2006 Lee ampDunton 1997 Lee Park amp Kim 2007 Peacuterez amp Romero 1992)howeverpreviousstudiesexaminingcontrolsonseagrassmetabo-lismhavefocusedonungrazedseagrasses(egApostolakiHolmerMarbagraveampKarakassis2010Gaciaetal2005)Meadowmetabolicproductivity correlates stronglywith above-ground seagrass bio-mass(Johnsonetal2017Figure5a)thatcanbegreatlyaffectedby green turtle grazing (Bjorndal 1980CebriaacutenampDuarte 1998Christianenetal2012FourqureanManuelCoatesKenworthyampSmith2010Williams1988)Above-groundbiomassexplained82ofthevariationinNEPacrossgrazedandungrazedT testudi-nummeadowsinthisstudybutNEPwasnotcorrelatedwitheitherenvironmentaltemperatureorirradiance(FigureS1)WethereforeinvestigatedwhetherthelackofanobservedrelationshipbetweenNEPand temperatureand irradiancewasperhapsdue toa largereffectfromdifferencesinbiomasscausedbygrazingmaskinganyeffectoftheseenvironmentalvariablesWescaledmetabolicratestotheabove-groundbiomassineachgrazedandungrazedarea(unitmetabolismperunitbiomass)andcomparedthemtotemperatureandirradianceAccountingfordifferencesinabove-groundbiomassdidnot revealanunderlyingrelationshipbetweenNEPandtheseenvironmentalparametersacrossmeadows(grazedungrazedorallareascombinedFigureS2)ThelackofarelationshipbetweenNEPandenvironmentalparametersinthisstudymaybedueinparttothe relatively limited rangeof temperature and irradiance in sea-grassmeadowsgrazedbygreenturtles in theGreaterCaribbeanAllmeadowsinthisstudywerelocatedinrelativelyshallowwarmtropicalandsubtropicalenvironmentsreceivinghighlevelsofinci-dentsunlight
41emsp|emspEffects of Halophila stipulacea invasion on meadow carbon capture
In addition to the newmetabolic carbon capture estimates wecontribute from grazed areas in T testudinum meadows wealso present the first estimates of NEP from meadows domi-nated by H stipulacea seagrass Having been first reported inthe Caribbean in 2002 (Ruiz amp Ballantine 2004)H stipulacea has spread rapidly among islands and become invasive in thisregion (Christianenet al 2018Willette et al 2014)Halophila stipulaceamayalsospreadmorequicklywithinareasofseagrassgrazedbygreenturtlesthaninungrazedareas(Christianenetal2018) suggesting that grazing may inadvertently facilitate thespreadofthisinvasivespeciesonceithasbecomeestablishedin
anareaInLacBayBonairegreenturtleshavebeguntoexpandtheirgrazingplotsintoareasofpreviouslyungrazedT testudinum asH stipulacea slowly takes over the historically grazed areaswithin thebay (Christianenet al 2018 SmuldersVonk EngelampChristianen2017) If this relationshipbetweengreen turtlesnative seagrasses and invasive seagrass exists in other areas itcouldhaveimplicationsfortotalecosystemcarbondynamicsandcarbonstorageaswellasotherecosystemfunctionswithinthesemeadows
Our resultsdemonstrate that ratesofmetabolic carboncap-turemaybe similarbetweenH stipulaceameadowsand thena-tive T testudinum meadows that they are replacing (Figure 4)HowevergiventhelimitednumberofH stipulaceameadowswewereabletosample inthisstudy(n=3)cautioniswarrantedinextrapolating these findingswidely to other locations facing in-vasionbyH stipulacea seagrassWhile ratesofNEP inH stipu-laceameadowsweresimilar tobothgrazed (n=3)andungrazed(n=2)areasofT testudinum-dominatedmeadowsinStCroixandBonaire (within-site comparisons) multiple factors affect theseratesthatmaydifferandaffectthisrelationshipinotherlocations(egdominantspeciesofnativeseagrassdensityandbiomassofH stipulaceavsthenativespeciesbladeepiphytecoverage)Forexample this relationshipmay differ in areas such as Dominica(eastern Caribbean) where the meadows undergoing H stipu-lacea invasion are dominated byS filiforme seagrass (WilletteampAmbrose 2012) Measurements from additional locations willgreatlyaidinunderstandingthespatialextentofthesimilarityinmetaboliccarboncaptureratesbetweenH stipulaceaandnativeseagrassmeadowsseenhereHoweverthecomparablemetabolicratesbetweentheinvasiveandnativeseagrassesmeasuredinthisstudysuggestsomebenefitsofseagrasspresencemayberetainedinmeadowsfollowinginvasionbyH stipulaceasuchaslocalmit-igation of ocean acidification (through metabolic buffering ofpHCamp et al 2016Hendriks et al 2014Unsworth CollierHendersonampMcKenzie2012)
It is not yet known whether comparable rates of metaboliccarbon capture will translate to comparable carbon sequestra-tionor storage inmeadowsdominatedby invasiveH stipulacea UnlikeT testudinum(vanTussenbroeketal2006)seagrassspe-cieswithinthegenusHalophiladonotformadeepbelow-groundrhizome mat (Fonseca 1989) Robust rhizome mats formed bysomeseagrassspeciescontributetohigherbelow-groundcarbonstorage(Christianenetal2013)andthelackofasimilarrhizomematmayresult in lowersedimentcarbonstorageinH stipulacea meadowsHalophila tissues also decompose quickly in sediment(JosselynFonsecaNiesenampLarson1986)and thismayaffectseagrass-derivedcarboninputtothesedimentsininvadedmead-ows compared to native species with higher refractory organicmatter content (Trevathan-Tackett et al 2017) Further studieson carbondynamics inH stipulaceameadowswill beneeded toformabetterunderstandingofhowthecontinuedinvasionbythisspecieswill affect total ecosystem carbon dynamics in seagrassmeadowsintheCaribbean
emspensp emsp | emsp1111Journal of EcologyJOHNSON et al
42emsp|emspGreater global assessment of grazed meadows needed
UngrazedareasofT testudinum-dominatedseagrassmeadowsfromthisstudyexhibitedratesofNEP(median1040mmolCmminus2 dayminus1)neartheupperendofthosereportedintheliteratureforseagrassecosystemsandgrazedareasinthisstudyexhibitedrates(median381)nearthemedianvalueofNEPforallreportedungrazedsea-grassmeadows(median271Figure6)Measurementsofseagrassmetaboliccarboncapturearerelativelygeographicallylimitedhow-ever with the majority of measurementsmdashmostly from ungrazedT testudinum-dominated meadowsmdashcoming from the GreaterCaribbeanandGulfofMexicoregionsTheCaribbeanandGulfofMexicoareecologicallyimportantgrazingregionsbutonlyasingleestimateofseagrassmetabolismfromagrazedareaexistedpriortothisstudy(Johnsonetal2017)
Allmeadows in the present study exhibited a consistent re-sponseofmetaboliccarboncapturetogreenturtlegrazinghow-ever our studywas limited to the effects of grazing in tropicaland subtropicalT testudinum-dominatedmeadows It ispossiblethatmeadows inother regionsof theworldperhapsdominatedbydifferent seagrass speciesandunderdifferentenvironmentalconditionswillnotexhibitthesameresponsetograzingintermsofmetabolic carbon dynamicsWe are aware of only one otherstudy inwhichthiswas investigated Ina tropicalThalassia hem-prichiimeadow(Tanzania)NEPwasreducedasaresultofexper-imental clipping to simulategrazing (Dahletal2016)howeveronlyhourlyratesofdaytimeNEPweremeasuredWithoutcorre-spondingratesofrespirationwecannotevaluateifNEPremained
positive in thisT hemprichiimeadowoveradiel cycleand if theresponse toexperimental clipping inTanzaniawas similar to theresponse to grazing measured in our study Further studies oneffectsofgrazingonseagrassmeadowmetaboliccarbondynam-icswould be particularly beneficial in (a) additional areaswheregrazer abundance is increasing and (b) meadows dominated byspeciesotherthanT testudinum
5emsp |emspCONCLUSIONS
Given the importance of seagrass meadows as blue carbon eco-systems (Fourqureanetal2012)andtheir role incarbonseques-tration and potential climate changemitigation it is necessary tounderstand how meadow carbon dynamics are affected by graz-ing Our study adds critical information on the effects of grazingon seagrassmetabolic carbondynamics andhow theseprocessesmaybeaffectedwith increasinggreenturtleabundanceandgraz-ingWedemonstratethattheresponseinmetaboliccarboncapturetogreenturtlegrazing isconsistentacrossseagrassecosystems inthe Greater Caribbean and Gulf of Mexico regionsmdashan area thatsupportsvastexpansesofseagrass(GreenampShort2003Jackson1997Wabnitzetal2008)Thisisanecologicallyimportantgrazingregionforgreenturtlepopulationsandgivenincreasinggreenturtleabundanceacrosstheglobe(Chaloupkaetal2008Mazarisetal2017)itisexpectedthatmoreseagrassareawillreturntoanaturalgrazedstateOurresultssuggestthat increasedgrazingwill trans-latetolowerratesofmetaboliccarboncapturebutwillnotstimulaterespirationandthemetaboliclossofcarbonfromthesemeadows
F I G U R E 6 emspRates(meanplusmnSD)ofnetecosystemproductionfrommeadowsofvariousseagrassspeciescollectedfromtheliteratureOpencircleslsquocrossedrsquocirclesandopentrianglesaregrazedThalassia testudinum(n=7)ungrazedT testudinum(n=6)andHalophila stipulacea(n=3)meadowsrespectivelyfromthepresentstudyFilledcirclesarepreviouslypublishedestimatesofungrazedseagrassmeadownetecosystemproduction(NEP)andcomefromvariouslocationsaroundtheworldCriteriaforliteraturecollectionanddatainclusionaredescribedinJohnsonetal(2017)RatesofNEPfromungrazedT testudinum and H stipulaceameadowsfromthepresentstudyareneartheupperendofpublishedvaluesMedianNEPfromgrazedT testudinumareas(presentstudy)issimilartothemedianNEPofungrazedseagrassmeadowsglobally(Section42)FiguremodifiedfromJohnsonetal(2017)toincludeadditionalestimatesLettersabovepointsdenotesitesfromthepresentstudyndashBBonaireCLittleCaymanEEleutheraFFloridaSStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306

emspensp emsp | emsp1103Journal of EcologyJOHNSON et al
grazing(greaterthanayear)didnotleadtoaproportionalincreaseinbenthicrespirationandremineralizationofcarbonstored inthemeadow possibly due to lack of a significant heterotrophic com-munity These results are from a single Caribbean location (LittleCayman Cayman Islands) that has some of the highest recordedmetabolicratesamongseagrassmeadows(Johnsonetal2017)ItisnotknownifgrazinghassimilareffectsacrossseagrassmeadowsinotherlocationsorunderdifferentenvironmentalconditionsAbet-terunderstandingofthevariabilityamongmeadowsinresponsetograzingisneededtounderstandhowincreasinggreenturtleabun-danceandincreasedgrazingwillaffectseagrasscarboncapture intheGreaterCaribbeanregion
In addition to increased grazing seagrass meadows in theCaribbeanarealsobeinginvadedbytheseagrassHalophila stipula-cea(Figure1b)ThisseagrassspeciesmdashnativetotheIndianOceanmdashhas invaded themeadowsofmanyCaribbean islands following itsintroductiontotheregionin2002(RuizampBallantine2004Willetteetal2014)Initialresearchsuggeststhatgreenturtlesavoidgrazingthis invasiveseagrass infavourofnativespeciesandthatH stipu-laceamayspreadmorerapidlywithinareasofmeadowsgrazedbygreen turtles compared to areas left ungrazed (Christianen et al2018)Little isknownabouthowinvasionbyH stipulaceamayaf-fectcarbondynamicsorecosystemfunctioninginCaribbeanmead-owshowever(VianaSiriwardane-deZoysaWilletteampGillis2019)GiventhisinteractionbetweengrazingandH stipulaceaitiscriticaltounderstandhowcarbondynamicsareaffectedinmeadowswhereinvasionbythisspeciesisoccurringinadditiontograzing
Ourprimaryobjectiveinthisstudywastoevaluatethehypoth-esis that effectsof green turtle grazingonmetabolic carbon cap-tureratesareconsistentacrossseagrassmeadowsWealsoevaluatevariationinthestrengthoftheresponsetograzingamongmeadowsand how rates ofmetabolic carbon capture in invasiveH stipula-ceameadowscomparetonativemeadowswhereturtlesgrazeWecomparedratesofnetecosystemproduction(NEP)betweengrazedandungrazedareasofT testudinumseagrassmeadowsinfourloca-tionsaroundtheGreaterCaribbean(definitionfromPetuch2013Robertson amp Cramer 2014) and Gulf ofMexico with establishedgreen turtle grazing populations (in addition to those previouslysampled inLittleCayman)Thefivetotal locationsencompassedawidegeographicalareaandspannedarangeofenvironmentalandseagrassmeadowcharacteristicsrepresentativeofgreenturtlefor-agingareasinthisregionTheseagrassH stipulaceahadestablishedattwooftheselocationsandwecomparedratesofNEPinmead-owsofthis invasivespeciestothosemeasuredinbothgrazedandungrazedareasofnativemeadowsattheselocations
2emsp |emspMATERIAL S AND METHODS
21emsp|emspStudy sites
We sampled grazed and ungrazed areas ofT testudinum seagrassmeadowsatfivelocationsacrosstheGreaterCaribbeanandGulfofMexicowheregreenturtleshadestablishedforagingareas(Figure2)
OneoftheselocationsLittleCaymanCaymanIslandswassampledthreetimesduringJuneandJuly2016aspartofapreviousstudy(Johnsonetal2017)The remaining four locationsweresampledin2018StCroixUSVirginIslands(February)westcoastofFlorida(GulfofMexico side)USA (May)BonaireCaribbeanNetherlands(July)EleutheraTheBahamas(August)Weopportunisticallysam-pledtwoseparatesitesinbothBonaireandEleutheraandonesiteatallotherlocations(timeandorweatherconditionsdidnotallowsamplingadditionalsitesattheselocations)EstablishedmeadowsoftheseagrassH stipulaceawerepresentattwoofthelocationsmdashStCroixandBonairemdashandthesemeadowswerealsosampledforcom-parisontonearbygrazedandungrazedareasofnativeT testudinum meadows(oneofthesitesinBonaireonlyhadnearbygrazedT tes-tudinum for comparison) Coordinates and general environmentalcharacteristics (daylight hours water depth salinity temperatureandirradiance)forallsitesaregiveninTable1
Themeadowssampledinthisstudyvariedgreatlyinsizefromrelatively small (lt150m2) for the grazedT testudinum area at thenorthwestLacBaysiteinBonairetoverylarge(severalsquarekilo-metresofunbrokenseagrasshabitat)fortheungrazedT testudinum meadowattheNorthRacksiteinFlorida(westcoast)Meadowsatothersitesencompassedarangeofsizesbetweenthesetwosuchas~350m2 each for the grazed andungrazedT testudinum areasinLittleCaymanand~600m2forthegrazedareaandgt50000m2 for the ungrazed T testudinum meadow at the Arvida Bay site inEleuthera TheBahamasAt each site a grazed area and adjacentungrazedareawereselectedforsamplingthatwererepresentativeofthemeadowSamples(Sections22and23)werecollectedwithinseveralmetresoftheborderbetweenthegrazedandungrazedareastoavoidpossibleedgeeffectswhileensuringsimilarenvironmentalconditions
Allsampledgrazedareas(naturalgreenturtlegrazingplots)werenearly uniformly grazed The proportion of grazed T testudinum blades ranged from95 to 100 in all grazed areas Evidence ofgrazingbygreenturtleswasnotobservedinungrazedareasexcept
F I G U R E 2 emspMapofseagrassmeadowsamplinglocationsTwositesweresampledineachofBonaireandEleutheraandonesitewassampledateachoftheotherthreelocationsCircledsymbolsforStCroixandBonairedenotelocationswhereHalophila stipulaceameadowswerealsosampled
1104emsp |emsp emspenspJournal of Ecology JOHNSON et al
forinStCroixwhere~3ofT testudinumbladesshowedsignsofgrazing(croppedbladetips)Somefishesalsograzeonseagrassandevidenceoffishgrazing(egbitemarks)wasobservedinungrazedT testudinum meadows at some sites Biomass removal from fishgrazingwasminorcomparedtoturtlegrazinghoweverandwasnotquantifiedNoevidenceofgreenturtlegrazingwasobservedinanyH stipulaceameadows
WesampledsevenareasfromT testudinum-dominatedseagrassmeadowsthatdisplayedsignsofactivegreenturtlegrazing(hereaf-tergrazedareas)sixareasfromT testudinum-dominatedmeadowsthatwerenotgrazed(hereafterungrazedareas)andthreemeadowsthatweredominatedbytheinvasiveseagrassH stipulacea(hereaf-terH stipulaceameadows)Thesemeadowswere spreadacross awideareaoftheGreaterCaribbeanandGulfofMexicoregionsandvariedintheirbioticandabioticcharacteristicsSomeofthesampledseagrassmeadowssuchastheT testudinummeadows inBonairewerelargelymonospecificwhereasothersweredominatedbyonespecies(T testudinum or H stipulacea)withinterspersedSyringodium filiforme or Halodule wrightii seagrasses and various macroalgae(TableS1)Thesubstrateinallmeadowswasprimarilycalciumcar-bonate sand Additional meadow characteristics are provided inResults (Section31)Data from theLittleCayman sitehavebeenreportedpreviously(Johnsonetal2017)andarepresentedagainhereforcomparisontotheothersites
22emsp|emspSampling seagrass meadow characteristics
Seagrass meadow and environmental characteristics were sam-pledateachsiteatthetimemetabolicincubationswereconducted
(Section23)Environmentaltemperatureandirradianceweremeas-uredduringmetabolicincubationsatseagrasscanopyheight(5-minintervals) at each site with a HOBO Pendant data logger (OnsetComputer Corporation) A water sample was collected at canopyheight and salinity wasmeasured with a handheld AgTec SalinityRefractometer that was calibrated with freshwater each day be-fore sampling (Agriculture Solutions Strong ME no sample wascollectedattheFloridasite)Meadowdepthwasmeasuredateachlocationwitheitheratapemeasureorascubadiversdepthgauge(accuracyplusmn10cm)
Seagrassspeciescompositionshootdensityblademorphome-try(lengthwidthandsurfacearea)andabove-groundbiomassweredetermined in all sampled areas (grazed ungrazed H stipulacea)within1metreofwhere incubationchamberswereplacedShootdensitiesweremeasuredusing25times25cmquadrats(00625m2area)in T testudinummeadowsDuetothehighdensitiesofH stipulacea10times10cmquadrats(001m2area)wereusedtomeasureshootden-sitiesinmeadowsdominatedbythisspeciesAbove-groundbiomasssampleswerecollectedusing10times10cmquadrats(allmeadows)byclippingall bladesat the sediment surfacewith scissors Six repli-catequadrats (forbothshootdensityandbiomass)werecollectedfromallsampledareasateachsiteexceptfortheStCroix(n=5)andFlorida(n=3)sites(Table2)Bladelengthandwidthweremea-suredforthedominantspecies(T testudinum or H stipulacea)from30randomlyselectedseagrassbladesfromthesebiomasssamplesforeachareaBladesurfacearea(iephotosyntheticareaperblade)wascalculatedastwotimestheproductofbladelengthandwidth(for each species) Followingmeasurement all bladeswere gentlyscrapedcleanofsedimentsandepiphytes(lowepiphyteloadsatall
TA B L E 1 emspCoordinatesofeachsamplingsiteandenvironmentalparametersmeasuredintheseagrassmeadowsondaysofmetabolicincubations
Site
Latitude Longitude Daylight Depth Salinity Temperature Irradiance
Decimal degrees hours M permil degC Lux
Bonaire
LacCaiBeach 12104417 minus68223183 1275 10 35 300 324518
NWLacBay 12108546 minus68231562 1275 14 36 311 432217
StCroix
BIRNM 17784612 minus64624566 115 45 35 279 211904
LittleCayman
GrapeTreeBay 19696518 minus80059652 13 10 36 319 551840
19696518 minus80059652 13 10 37 317 465732
19696518 minus80059652 13 10 38 336 600715
Eleuthera
ArvidaBay 24722297 minus76190644 125 24 39 322 267648
HalfSound 24936641 minus76153420 125 06 36 344 717215
Florida
NorthRack 28564760 minus82779335 1375 20 mdash 292 409801
Note TemperatureandirradiancedataaremeanvaluesmeasuredduringincubationsTheLittleCaymansitewassampledthreetimesin2016Allothersitesweresampledoncein2018SamplingtimesaregiveninSection21AbbreviationBIRNMBuckIslandReefNationalMonumentsite
emspensp emsp | emsp1105Journal of EcologyJOHNSON et al
sites) rinsed infreshwateranddriedtoaconstantweightat60degCbeforeweighingfordrymass
23emsp|emspEcosystem metabolism measurements
Metaboliccarbondynamicsoftheseagrassecosystemsweremeas-uredusingbenthic incubationchambers (Figure1c)similartopre-viousstudies (BarroacutenMarbagraveTerradosKennedyampDuarte2004CallejaBarroacutenHaleFrazerampDuarte2006Johnsonetal2017OliveacuteSilvaCostaampSantos2015)ChamberswereconstructedbyinsertingaPVCcylinder(16cmdiameter002m2area)~75cmintothe sediment (attempting to avoid severing horizontal rhizomes)and attaching a flexible gas-tight polyethylene bagwith samplingporttothetop(HansenThamdrupampJoslashrgensen2000)Theuseofflexiblegas-tightbagsinchamberconstructionallowsthepropaga-tionofwave turbulence to the chambers interior tomore closelysimulateenvironmentalconditionsIncubationchambervolumewasmeasuredinthelaboratorytobe55ndash6LOnsamplingdayscham-berswere set up in themeadows between 1030 and 1130 andincubationswererunfor25ndash3hrThisincubationlengthwaschosenbecausesaturationeffectscanoccurwithinchambersduringlonger
incubationsandhavebeenshowntounderestimatemetabolicrates(Oliveacuteetal2015)Metabolicdynamicsweremeasuredthreetimesat2-weekintervalsinLittleCaymanaspartofapreviousstudyandshown to be relatively stable among sampling events (Johnson etal2017) Incubationswerethereforeconductedonceateachad-ditionalsiteinthepresentstudy
Wesetupthreelight(clear)andthreedark(opaque)chambersineacharea(grazedorungrazedT testudinumorH stipulacea)tomea-sureecosystemmetabolicrates(n=3forallareas)LightchamberswereusedtomeasureratesofNEPanddarkchamberswereusedtomeasure ratesofecosystemrespiration (RE)Metabolic rateswereestimatedfromchangesindissolvedoxygen(DO)concentrationin-side the chambersbetween thebeginningandendof the incuba-tionperiodThreewatersampleswerecollectedfromthechamberviathesamplingportin60-mlplasticsyringesatthebeginningandendoftheincubationUponcollectionsyringeswerecappedwitha silicon cap andbrought to the surfacewhereDOconcentrationwas measured directly in the syringe with an optical DO probe(Vadeboncoeur2011YSIProODOYellowSpringsOhio)TheDOprobewas calibrated at the beginning of each sampling daywithwater-saturatedair
TA B L E 2 emspSeagrasscharacteristicsofgrazedandungrazedareasofThalassia testudinummeadowsandHalophila stipulaceameadowsmeasuredateachsamplingsite
Site Meadow n
Seagrass density Canopy heightBlade surface area AG biomass
shootsm2 cm cm2 g DMm2
Bonaire
LacCaiBeach Grazed 6 7413plusmn1263 47plusmn22 63plusmn32 271plusmn76
Ungrazed 6 9547plusmn937 96plusmn39 214plusmn116 1309plusmn544
H stipulacea 6 38500plusmn8068 21plusmn06 19plusmn09 364plusmn133
NWLacBay Grazed 6 2373plusmn446 28plusmn14 28plusmn14 48plusmn28
H stipulacea 6 34833plusmn5492 20plusmn04 18plusmn06 171plusmn59
StCroix
BIRNM Grazed 5 19648plusmn2261 40plusmn23 44plusmn27 274plusmn102
Ungrazed 5 24064plusmn2501 81plusmn31 97plusmn45 516plusmn110
H stipulacea 5 37336plusmn12494 28plusmn05 32plusmn09 630plusmn240
LittleCayman
GrapeTreeBay Grazed 6 9573plusmn1485 20plusmn09 23plusmn12 123plusmn37
Ungrazed 6 10187plusmn2102 149plusmn85 307plusmn204 1832plusmn948
Eleuthera
ArvidaBay Grazed 6 16480plusmn3927 35plusmn19 42plusmn22 418plusmn139
Ungrazed 6 20533plusmn3299 152plusmn46 258plusmn88 2022plusmn348
HalfSound Grazed 6 16747plusmn3167 18plusmn07 14plusmn06 88plusmn20
Ungrazed 6 17840plusmn2847 94plusmn20 108plusmn31 965plusmn162
Florida
NorthRack Grazed 3 13227plusmn4441 25plusmn11 16plusmn08 93plusmn24
Ungrazed 3 13867plusmn1619 89plusmn27 78plusmn29 707plusmn233
Note nisnumberofreplicatesateachsiteValuesaremeansplusmnSDCanopyheightismeasuredfrommeanbladelengthBladesurfaceareaisperbladeofseagrassAbbreviationsAGabove-groundBIRNMBuckIslandReefNationalMonumentsiteDMdrymass
1106emsp |emsp emspenspJournal of Ecology JOHNSON et al
Tomeasureonlymetabolicratesoftheseagrassbenthiccompo-nentsofthesystem(thoseaffectingcarbonstorage)wecorrectedrates measured from incubation chambers for water column me-tabolismWemeasuredwater columnmetabolismusingbiologicaloxygendemand(BOD)bottlesatthesametimeasbenthicchamberincubationsThreeclear300-mlglassBODbottlesandthreeopaque300-mlglassBODbottleswereusedtomeasurewatercolumnpro-ductionandrespirationrespectivelyBottleswerefilledwithwaterat seagrasscanopyheightanchored to thebottomand incubatedunderinsituconditionsintheseagrassmeadowWatercolumnsam-pleswerecollectedatthebeginningofincubationsin60-mlsyringesatcanopyheightandDOconcentrationwasmeasuredinthesamemannerassamplesfromincubationchambersFollowingthe incu-bationperiodbottleswerecollectedand returned to thesurfaceonesamplewascollectedfromeachbottlewithasyringeandDOconcentrationwasmeasured
Hourlymetabolic rateswere calculated from thedifference inDOconcentrationwithinlight(NEP)anddark(RE)incubationcham-bersbetweenthebeginningandendoftheincubationperiodWatercolumn metabolic rates measured from BOD bottles were sub-tractedfromratesmeasuredinchambersHourlygrossprimarypro-duction(GPP)wasthencalculatedasthesumofhourlyNEPandRE DailyratesofGPPwerecalculatedbymultiplyinghourlyGPPbythelengthofthephotoperiodmultipliedby075(toaccountfor lowermetabolicratesatdawnandduskJohnsonetal2017Oliveacuteetal2015)anddailyratesofREwerecalculatedbymultiplyinghourlyRE by24hoursDailyNEPwascalculatedas thedifferencebetweendaily GPP and RE Therewere a few occasionswhen an increaseinDO concentrationwasmeasuredwithin a dark chamber or thedarkBODbottlesduringanincubationsuggestinganerrorThesemeasuredincreasesweresmallusuallywithintheprobesmarginoferror (01mgO2L)andweassumedrespirationtobezeroduringthese timesCorrecting for these respiration errors hadminor ef-fectsondailymetabolic rates in themeadows (TablesS2andS3)MeasuredDOconcentrationswereconvertedfrommgO2tommolO2andthentocarbonunits(mmolC)assumingphotosyntheticandrespiratoryquotientsofone(BarroacutenampDuarte2009)
24emsp|emspData analyses
Net ecosystem productionwas themetabolic variable of primaryinterestasitaccountsforbothcarboncaptureandlossandcanbeusedas an indicatorofwhether a system is currently ametaboliccarbonsinkorsourceThereforeNEPisthemetabolicvariableforwhichstatisticalresultsarepresented
Ouraimwastoevaluatewhethergreenturtlegrazinghasacon-sistenteffectonmetabolicratesacrossseagrassecosystemsmdashthatistoevaluatewhetheratanygivensiteatanygiventimeratesofNEPareloweringrazedareasthaninungrazedareasWeevaluateddifferences in ratesofNEPbetweenareasofgrazedandadjacentungrazedT testudinum from each sitewith paired t tests T testswereusedbecause thecomparisonof interestwas thedifferencebetween grazed and ungrazed areas (effect of grazing) within a
givensitenotdifferencesinNEPamongsitesForsiteswheretheinvasive seagrass H stipulacea was present we used one-wayANOVAstotestforsignificantdifferencesinratesofNEPbetweenH stipulaceameadowsandnearbygrazedandungrazedT testudinum areas(StCroixsiteandLacCaiBeachsiteBonaire)Forthenorth-west site in Lac Bay Bonaire where only a grazed area of T te-studinum and an invasive H stipulacea meadow were comparedanunpaired t testwasused IfANOVA resultswere significant aTukeyspost-hoctestwasusedtoidentifysignificantcomparisonsWetestedfordifferencesinseagrassmeadowparametersbetweengrazedandungrazedT testudinumareasandH stipulaceameadowsatsitesusingpairedttestsandANOVAsinthesamemannerasforNEPWeused linearregressiontotestrelationshipsbetweenNEPand explanatory variables (eg seagrass biomass environmentaltemperature)MeanvaluesforNEPandexplanatoryvariablesfromeachmeadowwereusedforlinearregressionstestssotherewasasingledatapointforeachgrazedandungrazedT testudinum area or H stipulaceameadowateachsamplingsite
AllanalyseswereperformedinRversion343(RCoreTeam2018)Datawere processed using the lsquodplyrrsquo package (Wickham FrancoisHenryampMuumlller2017)andpost-hocanalyseswereperformedusingthelsquoagricolaersquopackage(deMendiburu2017)Alldatamettheassump-tionsfortheseanalysesanddataarepresentedasmeanplusmnSD
3emsp |emspRESULTS
31emsp|emspSeagrass meadow characteristics
Total seagrass shoot densities varied widely among meadowsand locations (Table 2) though shoot densitieswere always high-est inH stipulaceameadows The lowest shoot density observed(2373plusmn446shootsm2)wasinagrazedarea(NWsiteinLacBayBonaire)buttherewasnocleartrendforhigherorlowerseagrassdensities between grazed and ungrazed areas across the sampledsitesThalassia testudinum density in LittleCaymanwasgreater inthe grazed area than ungrazed area (paired t test p = 02) whileT testudinumdensityattheLacCaiBeachsiteinBonairewasgreaterintheungrazedarea(pairedttestp=02)Thalassia testudinum den-sitydidnotdifferbetweengrazedandungrazedareasattheremain-ingsites(pairedttestspgt05)ShootdensityofH stipulaceawasnot significantly different between any of the threeH stipulacea meadowsmeasured(ANOVAF2=009p=92)
HalophilaseagrassestypicallygrowshortbladesCanopyheightoftheH stipulaceameadowwassimilartothatofthenearbygrazedarea inStCroix andwas shorter than thenearbygrazedareasatbothsitesinBonaire(Table2)Halophila stipulaceabladelengthwasgreaterinStCroixthanBonaire(ANOVAF2=2413plt01)AreasofT testudinum-dominatedseagrassmeadowsgrazedbygreentur-tleshad significantly shorterblades and therefore canopyheightthan adjacent ungrazed areas at all sampled sites (paired t testsplt05forallcomparisonsTable2)
Greenturtlesbycroppingseagrassbladesnearthesedimentsur-faceandreducingcanopyheightgreatlyreducetheabove-ground
emspensp emsp | emsp1107Journal of EcologyJOHNSON et al
seagrass biomass of meadows within their grazing areas Above-ground biomass ranged from 48 plusmn 28 to 418 plusmn 139 g DMm2 amonggrazedareas(Table2)andfrom516plusmn110to2022plusmn348gDMm2amongungrazedareasBiomasswasalwaysloweringrazedareas than adjacent ungrazed areas (p lt 05 for all comparisons)Biomass among the three sampledH stipulaceameadows rangedfrom171plusmn59to630plusmn240gDMm2TheH stipulacea meadow in St Croix had significantly greater above-ground biomass thaneither of theH stipulaceameadows sampled inBonaire (ANOVAF2=1202plt 01)Evidenceofgreenturtlegrazingwasonlyob-served in T testudinum-dominated meadows in this study notH stipulaceameadowsandgrazingresultedinagt79decreaseinbiomassingrazedcomparedtoungrazedareasatallsitesexceptStCroix(47Table3)
32emsp|emspEcosystem metabolic rates
Rates of NEP differed among meadow types (grazed ungrazedH stipulacea) across the studied locations (Figures 3 and 4 seeTable S2 for data) Across all sites NEP ranged from 42 plusmn 70 to517plusmn110mmolCmminus2 dayminus1amonggrazedareasandfrom523plusmn57to2253plusmn191mmolCmminus2 dayminus1amongungrazedareasThelargestandarddeviationsonsomeoftheungrazedareaNEPestimatesfromLittleCaymanwerearesultofoneincubationchamberproducingratesmuchdifferentfromtheothertwoAcrossallsitesNEPwasconsist-entlyloweringrazedareasthanungrazedareas(Figure3)DifferencesinNEPbetweengrazedandungrazedareasweresignificantat four
ofthesixsiteswhereungrazedT testudinumwaspresentStCroixLittleCayman (all threemeasurement times) theArvidaBay site inEleutheraandFlorida(pairedttestsplt05forallcomparisons)AttheLacCaiBeachsiteinBonaireandtheHalfSoundsiteinEleuthera
TA B L E 3 emspPercentdifferenceinmetabolicratesandabove-ground(AG)seagrassbiomassbetweengrazedandadjacentungrazedThalassia testudinumareas
Site
GPP RE NEP AG Biomass
difference
Bonaire
LacCaiBeach 818 909 656 793
StCroix
BIRNM 604 783 560 469
LittleCayman
GrapeTreeBay 922 955 891 933
899 796 960 933
885 870 900 933
Eleuthera
ArvidaBay 543 395 629 793
HalfSound 801 889 679 908
Florida
NorthRack 615 413 919 868
Note TheLittleCaymansitewassampledthreetimesin2016Allothersitesweresampledoncein2018SamplingtimesaregiveninSection21AbbreviationsBIRNMBuckIslandReefNationalMonumentsiteGPPgrossprimaryproductionNEPnetecosystemproductionREecosys-temrespiration
F I G U R E 3 emspRates(meanplusmnSD)ofnetecosystemproduction(a)grossprimaryproduction(b)andecosystemrespiration(c)fromgrazed(openpoints)andadjacentungrazed(solidpoints)areasofThalassia testudinumseagrassmeadowsRatesofNetecosystemproduction(NEP)weresignificantlyloweringrazedareasthanungrazedareasatallsitesexceptatLacCaiBeachBonaireandHalfSoundEleuthera(Section32)SitesorderedsouthtonorthBONBonaire(LCBLacCaiBeachsite)STXStCroixLCLittleCaymanELEleuthera(ABArvidaBaysiteHSHalfSoundsite)FLFloridaDashedlinein(a)denotesmetabolicbalance(NEP=0)Valuesabovelinerepresentnetmetaboliccarboncapture[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1108emsp |emsp emspenspJournal of Ecology JOHNSON et al
differencesinNEPbetweengrazedandungrazedareaswerenearlysignificant(pairedttestsp = 08 and p=09respectively)
Though NEP was significantly lower in grazed than ungrazedareas atmost sites rates ofNEPwere almost always positive asecosystem respiration was not stimulated in relation to primaryproductionevenwithconsiderablereductionofphotosyntheticsea-grassbiomassbygrazing(gt79biomassreductionatallsitesexceptStCroix Table3)NEP ranged from56 to96 lower in grazedareasthanungrazedareas(Table3)Asimilarrangeinthedifference
betweengrazedandungrazedareaswasmeasuredforgrossprimaryproduction (54ndash92 lower in grazed) and ecosystem respiration(40ndash96loweringrazed)
Halophila stipulacea meadows were present at three sites andratesofNEPintheseinvasiveseagrassmeadowswerecomparedtonearbynativeT testudinum-dominatedmeadowsNEPintheH stip-ulaceameadowswascomparedtonearbygrazedandungrazedareasofT testudinum-dominatedmeadowsattwoofthesites(StCroixandtheLacCaiBeachsiteinBonaire)WithineachsiteratesofNEPintheH stipulaceameadowwerebetweenthoseofthegrazedandungrazedareas (Figure 4) but the differences were not significant (ANOVApgt10forallpost-hoccomparisonswithineachsite)AnungrazedareawasnotavailableforcomparisonatthethirdsiteatwhichH stipula-ceawaspresent (NWLacBaysite inBonaire)asallT testudinumatthissitewasgrazedbygreenturtlesandratesofNEPdidnotdifferbetweenthe invasive-andnative-dominatedareasatthissiteeither(t test p = 56 Figure 4)Halophila stipulaceameadowNEP rangedfrom612plusmn226to989plusmn233mmolCmminus2 dayminus1(TableS2)anddidnotdiffersignificantlyamongthethreesitesatwhichitwaspresent(ANOVAF2=192p=23)eventhoughtheestablishmentofH stip-ulaceaseagrassmeadowsinStCroixwasrelativelyrecent(AGulickunpublisheddata)comparedtothoseinBonaire(Willetteetal2014)
33emsp|emspDrivers of metabolic rates
VariationinratesofseagrassecosystemmetabolismacrossgrazedandungrazedareasappearedtobedrivenbysomecharacteristicsoftheseagrassmeadowsbutnotbyenvironmentalfactorsMeadowNEPwasstronglypositivelyrelatedtoabove-groundseagrassbio-massacrosssites(linearregressionR2=82plt01Figure5a)NEP
F I G U R E 4 emspRates(meanplusmnSD)ofnetecosystemproductioninmeadowsdominatedbytheinvasiveseagrassHalophila stipulacea (triangles)comparedtonearbygrazed(opencircles)andungrazed(solidcircles)areasofThalassia testudinummeadowsBONBonaire(LCBLacCaiBeachsiteNWnorthwestLacBaysite)STXStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
F I G U R E 5 emspRelationshipbetweennetecosystemproductionandabove-groundseagrassbiomass(a)andtotalseagrassshootdensity(b)fromgrazed(openpoints)andungrazed(solidpoints)areasofThalassia testudinummeadowsSolidlinein(a)isthesignificantlinearregression(R2=82plt01)betweennetecosystemproduction(NEP)andbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows
emspensp emsp | emsp1109Journal of EcologyJOHNSON et al
wasalsopositivelycorrelatedwithseagrassmeadowcanopyheight(R2=84plt01)andbladesurfacearea(R2=92plt01)Above-groundbiomassislikelythemoreusefulpredictorofmeadowNEPhoweverasbiomassisinfluencedbybothcanopyheightandbladesurfaceareaandachange ineitherofthesevariablesresults inachangeinbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows(linearregressionR2lt01p=96Figure5b)norwerethererelationshipsbetweenNEPandenvironmentaltempera-ture(R2=01p=70)irradiance(R2lt01p=10)ormeadowdepth(R2lt01p=77)acrosssites(FigureS1)
4emsp |emspDISCUSSION
Through a comparative study of seagrass meadowsmdashgrazed andungrazedmdashacross a wide area encompassing sites in the GreaterCaribbean and Gulf of Mexico we found that green turtle graz-inghas a consistent effecton themetabolic carbon capture ratesofT testudinum-dominatedseagrassmeadowsRatesofmetaboliccarboncapturewerealwayslowerinareasgrazedbyturtlesthanun-grazedareasofmeadowsmdashupto96inLittleCaymanmdashbutratesofNEPwerealwaysnearorabovemetabolicbalanceEcosystemres-pirationwasnotstimulatedinrelationtoprimaryproductionwithinareas grazed by green turtles regardless of the amount of above-groundseagrassbiomassremovedthroughgrazingorotherdiffer-encesinbioticandabioticfactorsamongsitesThesefindingslendadditionalsupporttothehypothesisthatfutureratesofmetaboliccarboncapturemaybelowerinareasgrazedbyturtlesbutgrazingwillnot stimulateecosystemrespirationand result ina large rem-ineralizationof carbon currently stored in these seagrass habitats(Johnsonetal2017)
The similarity in results between St Croix (awintermeasure-ment)andtheotherfourlocationsinthepresentstudy(allsummermeasurements)strengthensourconclusionthatgrazingresultsinaconsistent response inmetaboliccarboncaptureacrossmeadowsHoweverwewerenotable toaddresspotential temporaldynam-icswithinsitesthatmayaffectnetcarboncaptureoveranannualcycle in these systems Future studies measuring carbon captureoverlongertemporalscales(ieseasonal)maybebeneficialtoourunderstanding of the effects of grazing on within-system carbondynamics
Thoughtheresponse inmetabolic rates tograzingwasconsis-tent acrossmeadows the strengthof this responsevariedamongindividual sites Some sites such as Little Cayman had consider-ablylowerratesofNEPingrazedareasthanungrazedareas(mean917)whereastheresponsetograzing(differenceinNEPbetweengrazedandadjacentungrazedareas)waslesspronouncedatothersites suchasStCroix (Table3) Seagrassmeadowcharacteristicssuch as shoot density canopy height and above-ground biomassare likely to bemore variable among ungrazed areas than grazedareasasgreenturtlegrazingresults insimilarcanopymorphome-tryacrossmeadowsintheGreaterCaribbeanregion(egBjorndal1980HernaacutendezampvanTussenbroek2014Williams1988)Given
the strong relationship between NEP and above-ground biomass(Figure 5a) high variability in NEPwould be expected across un-grazedmeadows(egFigure3a)IndeedtherangeinratesofNEPamongungrazedareaswasnearlythreetimesgreaterthantherangemeasuredacrossgrazedareasinthisstudyAssiteswerespecificallychosen that differed from each other in meadow characteristicsvariationinthestrengthofthemetabolicresponsetograzingwouldbeexpected
Variability in the strength of the metabolic response to graz-ing resulted in the lackofa significantdifferencebetweengrazedandungrazedareasattwositesmdashLacCaiBeachinBonaireandHalfSoundinEleuthera(Figure3a) It ispossiblethatotherfactorsnotaccounted for here such as local environmental factors or greenturtlegrazinghistory(egintensitylongevityofplotuse)mayhaveledtohighervariabilityinmetabolicratesatsomesitesandlackofastatisticaldifferenceInadditionmetabolicratesmayexhibitwithin-meadowvariation(patchiness)similartobelow-groundcarbonstor-age in meadows (OreskaMcglathery amp Porter 2017) leading tohighervariabilityinestimatesandhelpingtoexplainthelackofasig-nificantdifferenceinNEPatthesetwositesThiscouldbedriveninpartbywithin-meadowspatialheterogeneityinabove-groundbio-massgiventhestrongrelationshipbetweenbiomassandNEPHighwithin-meadow variability inmetabolic rates if presentmay alsohelpexplainthe largevariabilitymeasured intheungrazedarea inLittleCaymanFuturestudiesusingtheeddycorrelationtechnique(Bergetal2003) tomeasureecosystemmetabolismmaybeabletoovercomesomeofthiswithin-meadowvariabilityhoweverthegreatexpenseofthismethodmakesitprohibitiveformanystudies
Seagrass ecosystem metabolism is commonly estimated usingoxygendynamicsmeasuredwithbenthicincubationchambers(egBarroacutenetal2004Gaciaetal2005presentstudy)howeverthereare some limitations to thismethodBenthic chambersmay severbelow-ground rhizomesand release carbon that couldaffectmet-abolicestimatesIntactrhizomesontheotherhandcanaffectthediffusiveoxygenlossfromtheseagrasstotherhizosphere(BorumSand-JensenBinzerPedersenampGreve2006)andmayresultinanunderestimateofgrossprimaryproduction(andthereforeNEP)Thismaybeminimizedusingshorter incubation timesas in this studyIncubation length also strongly affects metabolic rates estimatedfromoxygendynamics (Oliveacute et al 2015)with longer incubationtimesbiasingestimatestowardsnetheterotrophyInadditiontoox-ygenmetabolismotherprocessessuchascarbonatedynamics(VanDamLopesOsburnampFourqurean2019)play important roles intotal ecosystem carbondynamicswithout affecting themeasuredproductivityofameadow
CaribbeanT testudinummeadows are highly productive (egJohnsonetal2017KochampMadden2001)andtherelationshipbetween biomass andNEP observed in this study suggests thatmeadows may maintain positive productivity as long as photo-synthetic biomass remains A threshold biomass below whichCaribbean meadows may become net heterotrophic cannot bedirectly addressed here but could be predicted to be near zeroThisisincontrasttoapreviouslysuggestedabove-groundbiomass
1110emsp |emsp emspenspJournal of Ecology JOHNSON et al
threshold(41gDMm2)belowwhichmeadowsmaybecomehet-erotrophic (Duarte et al 2010) However that threshold wascalculated for seagrasses globally and included less productivespecies Additional studies of the relationship between biomassandmeadowNEP from seagrass species and regions other thanthoseinthepresentstudywouldbebeneficialtofurthergeneral-izehowgrazingaffectsmetaboliccarboncaptureacrossseagrassecosystems
Environmental temperature and irradiance are strong driv-ers of seagrassmeadow productivity (Calleja et al 2006 Lee ampDunton 1997 Lee Park amp Kim 2007 Peacuterez amp Romero 1992)howeverpreviousstudiesexaminingcontrolsonseagrassmetabo-lismhavefocusedonungrazedseagrasses(egApostolakiHolmerMarbagraveampKarakassis2010Gaciaetal2005)Meadowmetabolicproductivity correlates stronglywith above-ground seagrass bio-mass(Johnsonetal2017Figure5a)thatcanbegreatlyaffectedby green turtle grazing (Bjorndal 1980CebriaacutenampDuarte 1998Christianenetal2012FourqureanManuelCoatesKenworthyampSmith2010Williams1988)Above-groundbiomassexplained82ofthevariationinNEPacrossgrazedandungrazedT testudi-nummeadowsinthisstudybutNEPwasnotcorrelatedwitheitherenvironmentaltemperatureorirradiance(FigureS1)WethereforeinvestigatedwhetherthelackofanobservedrelationshipbetweenNEPand temperatureand irradiancewasperhapsdue toa largereffectfromdifferencesinbiomasscausedbygrazingmaskinganyeffectoftheseenvironmentalvariablesWescaledmetabolicratestotheabove-groundbiomassineachgrazedandungrazedarea(unitmetabolismperunitbiomass)andcomparedthemtotemperatureandirradianceAccountingfordifferencesinabove-groundbiomassdidnot revealanunderlyingrelationshipbetweenNEPandtheseenvironmentalparametersacrossmeadows(grazedungrazedorallareascombinedFigureS2)ThelackofarelationshipbetweenNEPandenvironmentalparametersinthisstudymaybedueinparttothe relatively limited rangeof temperature and irradiance in sea-grassmeadowsgrazedbygreenturtles in theGreaterCaribbeanAllmeadowsinthisstudywerelocatedinrelativelyshallowwarmtropicalandsubtropicalenvironmentsreceivinghighlevelsofinci-dentsunlight
41emsp|emspEffects of Halophila stipulacea invasion on meadow carbon capture
In addition to the newmetabolic carbon capture estimates wecontribute from grazed areas in T testudinum meadows wealso present the first estimates of NEP from meadows domi-nated by H stipulacea seagrass Having been first reported inthe Caribbean in 2002 (Ruiz amp Ballantine 2004)H stipulacea has spread rapidly among islands and become invasive in thisregion (Christianenet al 2018Willette et al 2014)Halophila stipulaceamayalsospreadmorequicklywithinareasofseagrassgrazedbygreenturtlesthaninungrazedareas(Christianenetal2018) suggesting that grazing may inadvertently facilitate thespreadofthisinvasivespeciesonceithasbecomeestablishedin
anareaInLacBayBonairegreenturtleshavebeguntoexpandtheirgrazingplotsintoareasofpreviouslyungrazedT testudinum asH stipulacea slowly takes over the historically grazed areaswithin thebay (Christianenet al 2018 SmuldersVonk EngelampChristianen2017) If this relationshipbetweengreen turtlesnative seagrasses and invasive seagrass exists in other areas itcouldhaveimplicationsfortotalecosystemcarbondynamicsandcarbonstorageaswellasotherecosystemfunctionswithinthesemeadows
Our resultsdemonstrate that ratesofmetabolic carboncap-turemaybe similarbetweenH stipulaceameadowsand thena-tive T testudinum meadows that they are replacing (Figure 4)HowevergiventhelimitednumberofH stipulaceameadowswewereabletosample inthisstudy(n=3)cautioniswarrantedinextrapolating these findingswidely to other locations facing in-vasionbyH stipulacea seagrassWhile ratesofNEP inH stipu-laceameadowsweresimilar tobothgrazed (n=3)andungrazed(n=2)areasofT testudinum-dominatedmeadowsinStCroixandBonaire (within-site comparisons) multiple factors affect theseratesthatmaydifferandaffectthisrelationshipinotherlocations(egdominantspeciesofnativeseagrassdensityandbiomassofH stipulaceavsthenativespeciesbladeepiphytecoverage)Forexample this relationshipmay differ in areas such as Dominica(eastern Caribbean) where the meadows undergoing H stipu-lacea invasion are dominated byS filiforme seagrass (WilletteampAmbrose 2012) Measurements from additional locations willgreatlyaidinunderstandingthespatialextentofthesimilarityinmetaboliccarboncaptureratesbetweenH stipulaceaandnativeseagrassmeadowsseenhereHoweverthecomparablemetabolicratesbetweentheinvasiveandnativeseagrassesmeasuredinthisstudysuggestsomebenefitsofseagrasspresencemayberetainedinmeadowsfollowinginvasionbyH stipulaceasuchaslocalmit-igation of ocean acidification (through metabolic buffering ofpHCamp et al 2016Hendriks et al 2014Unsworth CollierHendersonampMcKenzie2012)
It is not yet known whether comparable rates of metaboliccarbon capture will translate to comparable carbon sequestra-tionor storage inmeadowsdominatedby invasiveH stipulacea UnlikeT testudinum(vanTussenbroeketal2006)seagrassspe-cieswithinthegenusHalophiladonotformadeepbelow-groundrhizome mat (Fonseca 1989) Robust rhizome mats formed bysomeseagrassspeciescontributetohigherbelow-groundcarbonstorage(Christianenetal2013)andthelackofasimilarrhizomematmayresult in lowersedimentcarbonstorageinH stipulacea meadowsHalophila tissues also decompose quickly in sediment(JosselynFonsecaNiesenampLarson1986)and thismayaffectseagrass-derivedcarboninputtothesedimentsininvadedmead-ows compared to native species with higher refractory organicmatter content (Trevathan-Tackett et al 2017) Further studieson carbondynamics inH stipulaceameadowswill beneeded toformabetterunderstandingofhowthecontinuedinvasionbythisspecieswill affect total ecosystem carbon dynamics in seagrassmeadowsintheCaribbean
emspensp emsp | emsp1111Journal of EcologyJOHNSON et al
42emsp|emspGreater global assessment of grazed meadows needed
UngrazedareasofT testudinum-dominatedseagrassmeadowsfromthisstudyexhibitedratesofNEP(median1040mmolCmminus2 dayminus1)neartheupperendofthosereportedintheliteratureforseagrassecosystemsandgrazedareasinthisstudyexhibitedrates(median381)nearthemedianvalueofNEPforallreportedungrazedsea-grassmeadows(median271Figure6)Measurementsofseagrassmetaboliccarboncapturearerelativelygeographicallylimitedhow-ever with the majority of measurementsmdashmostly from ungrazedT testudinum-dominated meadowsmdashcoming from the GreaterCaribbeanandGulfofMexicoregionsTheCaribbeanandGulfofMexicoareecologicallyimportantgrazingregionsbutonlyasingleestimateofseagrassmetabolismfromagrazedareaexistedpriortothisstudy(Johnsonetal2017)
Allmeadows in the present study exhibited a consistent re-sponseofmetaboliccarboncapturetogreenturtlegrazinghow-ever our studywas limited to the effects of grazing in tropicaland subtropicalT testudinum-dominatedmeadows It ispossiblethatmeadows inother regionsof theworldperhapsdominatedbydifferent seagrass speciesandunderdifferentenvironmentalconditionswillnotexhibitthesameresponsetograzingintermsofmetabolic carbon dynamicsWe are aware of only one otherstudy inwhichthiswas investigated Ina tropicalThalassia hem-prichiimeadow(Tanzania)NEPwasreducedasaresultofexper-imental clipping to simulategrazing (Dahletal2016)howeveronlyhourlyratesofdaytimeNEPweremeasuredWithoutcorre-spondingratesofrespirationwecannotevaluateifNEPremained
positive in thisT hemprichiimeadowoveradiel cycleand if theresponse toexperimental clipping inTanzaniawas similar to theresponse to grazing measured in our study Further studies oneffectsofgrazingonseagrassmeadowmetaboliccarbondynam-icswould be particularly beneficial in (a) additional areaswheregrazer abundance is increasing and (b) meadows dominated byspeciesotherthanT testudinum
5emsp |emspCONCLUSIONS
Given the importance of seagrass meadows as blue carbon eco-systems (Fourqureanetal2012)andtheir role incarbonseques-tration and potential climate changemitigation it is necessary tounderstand how meadow carbon dynamics are affected by graz-ing Our study adds critical information on the effects of grazingon seagrassmetabolic carbondynamics andhow theseprocessesmaybeaffectedwith increasinggreenturtleabundanceandgraz-ingWedemonstratethattheresponseinmetaboliccarboncapturetogreenturtlegrazing isconsistentacrossseagrassecosystems inthe Greater Caribbean and Gulf of Mexico regionsmdashan area thatsupportsvastexpansesofseagrass(GreenampShort2003Jackson1997Wabnitzetal2008)Thisisanecologicallyimportantgrazingregionforgreenturtlepopulationsandgivenincreasinggreenturtleabundanceacrosstheglobe(Chaloupkaetal2008Mazarisetal2017)itisexpectedthatmoreseagrassareawillreturntoanaturalgrazedstateOurresultssuggestthat increasedgrazingwill trans-latetolowerratesofmetaboliccarboncapturebutwillnotstimulaterespirationandthemetaboliclossofcarbonfromthesemeadows
F I G U R E 6 emspRates(meanplusmnSD)ofnetecosystemproductionfrommeadowsofvariousseagrassspeciescollectedfromtheliteratureOpencircleslsquocrossedrsquocirclesandopentrianglesaregrazedThalassia testudinum(n=7)ungrazedT testudinum(n=6)andHalophila stipulacea(n=3)meadowsrespectivelyfromthepresentstudyFilledcirclesarepreviouslypublishedestimatesofungrazedseagrassmeadownetecosystemproduction(NEP)andcomefromvariouslocationsaroundtheworldCriteriaforliteraturecollectionanddatainclusionaredescribedinJohnsonetal(2017)RatesofNEPfromungrazedT testudinum and H stipulaceameadowsfromthepresentstudyareneartheupperendofpublishedvaluesMedianNEPfromgrazedT testudinumareas(presentstudy)issimilartothemedianNEPofungrazedseagrassmeadowsglobally(Section42)FiguremodifiedfromJohnsonetal(2017)toincludeadditionalestimatesLettersabovepointsdenotesitesfromthepresentstudyndashBBonaireCLittleCaymanEEleutheraFFloridaSStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306
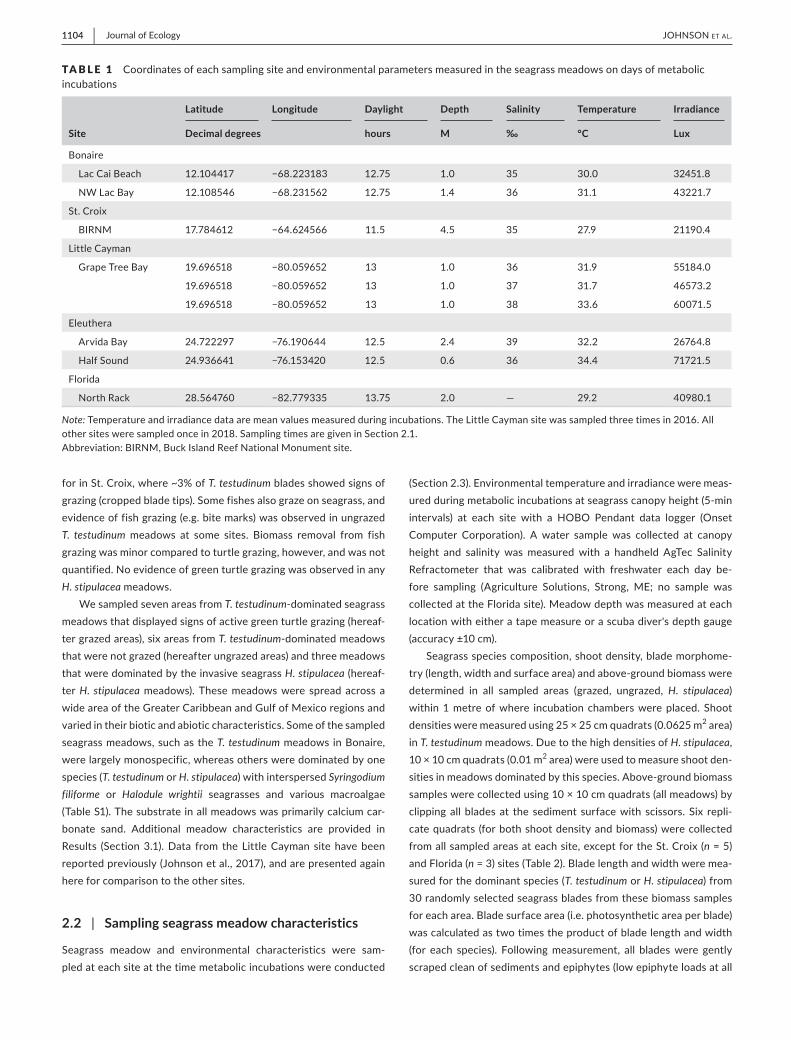
1104emsp |emsp emspenspJournal of Ecology JOHNSON et al
forinStCroixwhere~3ofT testudinumbladesshowedsignsofgrazing(croppedbladetips)Somefishesalsograzeonseagrassandevidenceoffishgrazing(egbitemarks)wasobservedinungrazedT testudinum meadows at some sites Biomass removal from fishgrazingwasminorcomparedtoturtlegrazinghoweverandwasnotquantifiedNoevidenceofgreenturtlegrazingwasobservedinanyH stipulaceameadows
WesampledsevenareasfromT testudinum-dominatedseagrassmeadowsthatdisplayedsignsofactivegreenturtlegrazing(hereaf-tergrazedareas)sixareasfromT testudinum-dominatedmeadowsthatwerenotgrazed(hereafterungrazedareas)andthreemeadowsthatweredominatedbytheinvasiveseagrassH stipulacea(hereaf-terH stipulaceameadows)Thesemeadowswere spreadacross awideareaoftheGreaterCaribbeanandGulfofMexicoregionsandvariedintheirbioticandabioticcharacteristicsSomeofthesampledseagrassmeadowssuchastheT testudinummeadows inBonairewerelargelymonospecificwhereasothersweredominatedbyonespecies(T testudinum or H stipulacea)withinterspersedSyringodium filiforme or Halodule wrightii seagrasses and various macroalgae(TableS1)Thesubstrateinallmeadowswasprimarilycalciumcar-bonate sand Additional meadow characteristics are provided inResults (Section31)Data from theLittleCayman sitehavebeenreportedpreviously(Johnsonetal2017)andarepresentedagainhereforcomparisontotheothersites
22emsp|emspSampling seagrass meadow characteristics
Seagrass meadow and environmental characteristics were sam-pledateachsiteatthetimemetabolicincubationswereconducted
(Section23)Environmentaltemperatureandirradianceweremeas-uredduringmetabolicincubationsatseagrasscanopyheight(5-minintervals) at each site with a HOBO Pendant data logger (OnsetComputer Corporation) A water sample was collected at canopyheight and salinity wasmeasured with a handheld AgTec SalinityRefractometer that was calibrated with freshwater each day be-fore sampling (Agriculture Solutions Strong ME no sample wascollectedattheFloridasite)Meadowdepthwasmeasuredateachlocationwitheitheratapemeasureorascubadiversdepthgauge(accuracyplusmn10cm)
Seagrassspeciescompositionshootdensityblademorphome-try(lengthwidthandsurfacearea)andabove-groundbiomassweredetermined in all sampled areas (grazed ungrazed H stipulacea)within1metreofwhere incubationchamberswereplacedShootdensitiesweremeasuredusing25times25cmquadrats(00625m2area)in T testudinummeadowsDuetothehighdensitiesofH stipulacea10times10cmquadrats(001m2area)wereusedtomeasureshootden-sitiesinmeadowsdominatedbythisspeciesAbove-groundbiomasssampleswerecollectedusing10times10cmquadrats(allmeadows)byclippingall bladesat the sediment surfacewith scissors Six repli-catequadrats (forbothshootdensityandbiomass)werecollectedfromallsampledareasateachsiteexceptfortheStCroix(n=5)andFlorida(n=3)sites(Table2)Bladelengthandwidthweremea-suredforthedominantspecies(T testudinum or H stipulacea)from30randomlyselectedseagrassbladesfromthesebiomasssamplesforeachareaBladesurfacearea(iephotosyntheticareaperblade)wascalculatedastwotimestheproductofbladelengthandwidth(for each species) Followingmeasurement all bladeswere gentlyscrapedcleanofsedimentsandepiphytes(lowepiphyteloadsatall
TA B L E 1 emspCoordinatesofeachsamplingsiteandenvironmentalparametersmeasuredintheseagrassmeadowsondaysofmetabolicincubations
Site
Latitude Longitude Daylight Depth Salinity Temperature Irradiance
Decimal degrees hours M permil degC Lux
Bonaire
LacCaiBeach 12104417 minus68223183 1275 10 35 300 324518
NWLacBay 12108546 minus68231562 1275 14 36 311 432217
StCroix
BIRNM 17784612 minus64624566 115 45 35 279 211904
LittleCayman
GrapeTreeBay 19696518 minus80059652 13 10 36 319 551840
19696518 minus80059652 13 10 37 317 465732
19696518 minus80059652 13 10 38 336 600715
Eleuthera
ArvidaBay 24722297 minus76190644 125 24 39 322 267648
HalfSound 24936641 minus76153420 125 06 36 344 717215
Florida
NorthRack 28564760 minus82779335 1375 20 mdash 292 409801
Note TemperatureandirradiancedataaremeanvaluesmeasuredduringincubationsTheLittleCaymansitewassampledthreetimesin2016Allothersitesweresampledoncein2018SamplingtimesaregiveninSection21AbbreviationBIRNMBuckIslandReefNationalMonumentsite
emspensp emsp | emsp1105Journal of EcologyJOHNSON et al
sites) rinsed infreshwateranddriedtoaconstantweightat60degCbeforeweighingfordrymass
23emsp|emspEcosystem metabolism measurements
Metaboliccarbondynamicsoftheseagrassecosystemsweremeas-uredusingbenthic incubationchambers (Figure1c)similartopre-viousstudies (BarroacutenMarbagraveTerradosKennedyampDuarte2004CallejaBarroacutenHaleFrazerampDuarte2006Johnsonetal2017OliveacuteSilvaCostaampSantos2015)ChamberswereconstructedbyinsertingaPVCcylinder(16cmdiameter002m2area)~75cmintothe sediment (attempting to avoid severing horizontal rhizomes)and attaching a flexible gas-tight polyethylene bagwith samplingporttothetop(HansenThamdrupampJoslashrgensen2000)Theuseofflexiblegas-tightbagsinchamberconstructionallowsthepropaga-tionofwave turbulence to the chambers interior tomore closelysimulateenvironmentalconditionsIncubationchambervolumewasmeasuredinthelaboratorytobe55ndash6LOnsamplingdayscham-berswere set up in themeadows between 1030 and 1130 andincubationswererunfor25ndash3hrThisincubationlengthwaschosenbecausesaturationeffectscanoccurwithinchambersduringlonger
incubationsandhavebeenshowntounderestimatemetabolicrates(Oliveacuteetal2015)Metabolicdynamicsweremeasuredthreetimesat2-weekintervalsinLittleCaymanaspartofapreviousstudyandshown to be relatively stable among sampling events (Johnson etal2017) Incubationswerethereforeconductedonceateachad-ditionalsiteinthepresentstudy
Wesetupthreelight(clear)andthreedark(opaque)chambersineacharea(grazedorungrazedT testudinumorH stipulacea)tomea-sureecosystemmetabolicrates(n=3forallareas)LightchamberswereusedtomeasureratesofNEPanddarkchamberswereusedtomeasure ratesofecosystemrespiration (RE)Metabolic rateswereestimatedfromchangesindissolvedoxygen(DO)concentrationin-side the chambersbetween thebeginningandendof the incuba-tionperiodThreewatersampleswerecollectedfromthechamberviathesamplingportin60-mlplasticsyringesatthebeginningandendoftheincubationUponcollectionsyringeswerecappedwitha silicon cap andbrought to the surfacewhereDOconcentrationwas measured directly in the syringe with an optical DO probe(Vadeboncoeur2011YSIProODOYellowSpringsOhio)TheDOprobewas calibrated at the beginning of each sampling daywithwater-saturatedair
TA B L E 2 emspSeagrasscharacteristicsofgrazedandungrazedareasofThalassia testudinummeadowsandHalophila stipulaceameadowsmeasuredateachsamplingsite
Site Meadow n
Seagrass density Canopy heightBlade surface area AG biomass
shootsm2 cm cm2 g DMm2
Bonaire
LacCaiBeach Grazed 6 7413plusmn1263 47plusmn22 63plusmn32 271plusmn76
Ungrazed 6 9547plusmn937 96plusmn39 214plusmn116 1309plusmn544
H stipulacea 6 38500plusmn8068 21plusmn06 19plusmn09 364plusmn133
NWLacBay Grazed 6 2373plusmn446 28plusmn14 28plusmn14 48plusmn28
H stipulacea 6 34833plusmn5492 20plusmn04 18plusmn06 171plusmn59
StCroix
BIRNM Grazed 5 19648plusmn2261 40plusmn23 44plusmn27 274plusmn102
Ungrazed 5 24064plusmn2501 81plusmn31 97plusmn45 516plusmn110
H stipulacea 5 37336plusmn12494 28plusmn05 32plusmn09 630plusmn240
LittleCayman
GrapeTreeBay Grazed 6 9573plusmn1485 20plusmn09 23plusmn12 123plusmn37
Ungrazed 6 10187plusmn2102 149plusmn85 307plusmn204 1832plusmn948
Eleuthera
ArvidaBay Grazed 6 16480plusmn3927 35plusmn19 42plusmn22 418plusmn139
Ungrazed 6 20533plusmn3299 152plusmn46 258plusmn88 2022plusmn348
HalfSound Grazed 6 16747plusmn3167 18plusmn07 14plusmn06 88plusmn20
Ungrazed 6 17840plusmn2847 94plusmn20 108plusmn31 965plusmn162
Florida
NorthRack Grazed 3 13227plusmn4441 25plusmn11 16plusmn08 93plusmn24
Ungrazed 3 13867plusmn1619 89plusmn27 78plusmn29 707plusmn233
Note nisnumberofreplicatesateachsiteValuesaremeansplusmnSDCanopyheightismeasuredfrommeanbladelengthBladesurfaceareaisperbladeofseagrassAbbreviationsAGabove-groundBIRNMBuckIslandReefNationalMonumentsiteDMdrymass
1106emsp |emsp emspenspJournal of Ecology JOHNSON et al
Tomeasureonlymetabolicratesoftheseagrassbenthiccompo-nentsofthesystem(thoseaffectingcarbonstorage)wecorrectedrates measured from incubation chambers for water column me-tabolismWemeasuredwater columnmetabolismusingbiologicaloxygendemand(BOD)bottlesatthesametimeasbenthicchamberincubationsThreeclear300-mlglassBODbottlesandthreeopaque300-mlglassBODbottleswereusedtomeasurewatercolumnpro-ductionandrespirationrespectivelyBottleswerefilledwithwaterat seagrasscanopyheightanchored to thebottomand incubatedunderinsituconditionsintheseagrassmeadowWatercolumnsam-pleswerecollectedatthebeginningofincubationsin60-mlsyringesatcanopyheightandDOconcentrationwasmeasuredinthesamemannerassamplesfromincubationchambersFollowingthe incu-bationperiodbottleswerecollectedand returned to thesurfaceonesamplewascollectedfromeachbottlewithasyringeandDOconcentrationwasmeasured
Hourlymetabolic rateswere calculated from thedifference inDOconcentrationwithinlight(NEP)anddark(RE)incubationcham-bersbetweenthebeginningandendoftheincubationperiodWatercolumn metabolic rates measured from BOD bottles were sub-tractedfromratesmeasuredinchambersHourlygrossprimarypro-duction(GPP)wasthencalculatedasthesumofhourlyNEPandRE DailyratesofGPPwerecalculatedbymultiplyinghourlyGPPbythelengthofthephotoperiodmultipliedby075(toaccountfor lowermetabolicratesatdawnandduskJohnsonetal2017Oliveacuteetal2015)anddailyratesofREwerecalculatedbymultiplyinghourlyRE by24hoursDailyNEPwascalculatedas thedifferencebetweendaily GPP and RE Therewere a few occasionswhen an increaseinDO concentrationwasmeasuredwithin a dark chamber or thedarkBODbottlesduringanincubationsuggestinganerrorThesemeasuredincreasesweresmallusuallywithintheprobesmarginoferror (01mgO2L)andweassumedrespirationtobezeroduringthese timesCorrecting for these respiration errors hadminor ef-fectsondailymetabolic rates in themeadows (TablesS2andS3)MeasuredDOconcentrationswereconvertedfrommgO2tommolO2andthentocarbonunits(mmolC)assumingphotosyntheticandrespiratoryquotientsofone(BarroacutenampDuarte2009)
24emsp|emspData analyses
Net ecosystem productionwas themetabolic variable of primaryinterestasitaccountsforbothcarboncaptureandlossandcanbeusedas an indicatorofwhether a system is currently ametaboliccarbonsinkorsourceThereforeNEPisthemetabolicvariableforwhichstatisticalresultsarepresented
Ouraimwastoevaluatewhethergreenturtlegrazinghasacon-sistenteffectonmetabolicratesacrossseagrassecosystemsmdashthatistoevaluatewhetheratanygivensiteatanygiventimeratesofNEPareloweringrazedareasthaninungrazedareasWeevaluateddifferences in ratesofNEPbetweenareasofgrazedandadjacentungrazedT testudinum from each sitewith paired t tests T testswereusedbecause thecomparisonof interestwas thedifferencebetween grazed and ungrazed areas (effect of grazing) within a
givensitenotdifferencesinNEPamongsitesForsiteswheretheinvasive seagrass H stipulacea was present we used one-wayANOVAstotestforsignificantdifferencesinratesofNEPbetweenH stipulaceameadowsandnearbygrazedandungrazedT testudinum areas(StCroixsiteandLacCaiBeachsiteBonaire)Forthenorth-west site in Lac Bay Bonaire where only a grazed area of T te-studinum and an invasive H stipulacea meadow were comparedanunpaired t testwasused IfANOVA resultswere significant aTukeyspost-hoctestwasusedtoidentifysignificantcomparisonsWetestedfordifferencesinseagrassmeadowparametersbetweengrazedandungrazedT testudinumareasandH stipulaceameadowsatsitesusingpairedttestsandANOVAsinthesamemannerasforNEPWeused linearregressiontotestrelationshipsbetweenNEPand explanatory variables (eg seagrass biomass environmentaltemperature)MeanvaluesforNEPandexplanatoryvariablesfromeachmeadowwereusedforlinearregressionstestssotherewasasingledatapointforeachgrazedandungrazedT testudinum area or H stipulaceameadowateachsamplingsite
AllanalyseswereperformedinRversion343(RCoreTeam2018)Datawere processed using the lsquodplyrrsquo package (Wickham FrancoisHenryampMuumlller2017)andpost-hocanalyseswereperformedusingthelsquoagricolaersquopackage(deMendiburu2017)Alldatamettheassump-tionsfortheseanalysesanddataarepresentedasmeanplusmnSD
3emsp |emspRESULTS
31emsp|emspSeagrass meadow characteristics
Total seagrass shoot densities varied widely among meadowsand locations (Table 2) though shoot densitieswere always high-est inH stipulaceameadows The lowest shoot density observed(2373plusmn446shootsm2)wasinagrazedarea(NWsiteinLacBayBonaire)buttherewasnocleartrendforhigherorlowerseagrassdensities between grazed and ungrazed areas across the sampledsitesThalassia testudinum density in LittleCaymanwasgreater inthe grazed area than ungrazed area (paired t test p = 02) whileT testudinumdensityattheLacCaiBeachsiteinBonairewasgreaterintheungrazedarea(pairedttestp=02)Thalassia testudinum den-sitydidnotdifferbetweengrazedandungrazedareasattheremain-ingsites(pairedttestspgt05)ShootdensityofH stipulaceawasnot significantly different between any of the threeH stipulacea meadowsmeasured(ANOVAF2=009p=92)
HalophilaseagrassestypicallygrowshortbladesCanopyheightoftheH stipulaceameadowwassimilartothatofthenearbygrazedarea inStCroix andwas shorter than thenearbygrazedareasatbothsitesinBonaire(Table2)Halophila stipulaceabladelengthwasgreaterinStCroixthanBonaire(ANOVAF2=2413plt01)AreasofT testudinum-dominatedseagrassmeadowsgrazedbygreentur-tleshad significantly shorterblades and therefore canopyheightthan adjacent ungrazed areas at all sampled sites (paired t testsplt05forallcomparisonsTable2)
Greenturtlesbycroppingseagrassbladesnearthesedimentsur-faceandreducingcanopyheightgreatlyreducetheabove-ground
emspensp emsp | emsp1107Journal of EcologyJOHNSON et al
seagrass biomass of meadows within their grazing areas Above-ground biomass ranged from 48 plusmn 28 to 418 plusmn 139 g DMm2 amonggrazedareas(Table2)andfrom516plusmn110to2022plusmn348gDMm2amongungrazedareasBiomasswasalwaysloweringrazedareas than adjacent ungrazed areas (p lt 05 for all comparisons)Biomass among the three sampledH stipulaceameadows rangedfrom171plusmn59to630plusmn240gDMm2TheH stipulacea meadow in St Croix had significantly greater above-ground biomass thaneither of theH stipulaceameadows sampled inBonaire (ANOVAF2=1202plt 01)Evidenceofgreenturtlegrazingwasonlyob-served in T testudinum-dominated meadows in this study notH stipulaceameadowsandgrazingresultedinagt79decreaseinbiomassingrazedcomparedtoungrazedareasatallsitesexceptStCroix(47Table3)
32emsp|emspEcosystem metabolic rates
Rates of NEP differed among meadow types (grazed ungrazedH stipulacea) across the studied locations (Figures 3 and 4 seeTable S2 for data) Across all sites NEP ranged from 42 plusmn 70 to517plusmn110mmolCmminus2 dayminus1amonggrazedareasandfrom523plusmn57to2253plusmn191mmolCmminus2 dayminus1amongungrazedareasThelargestandarddeviationsonsomeoftheungrazedareaNEPestimatesfromLittleCaymanwerearesultofoneincubationchamberproducingratesmuchdifferentfromtheothertwoAcrossallsitesNEPwasconsist-entlyloweringrazedareasthanungrazedareas(Figure3)DifferencesinNEPbetweengrazedandungrazedareasweresignificantat four
ofthesixsiteswhereungrazedT testudinumwaspresentStCroixLittleCayman (all threemeasurement times) theArvidaBay site inEleutheraandFlorida(pairedttestsplt05forallcomparisons)AttheLacCaiBeachsiteinBonaireandtheHalfSoundsiteinEleuthera
TA B L E 3 emspPercentdifferenceinmetabolicratesandabove-ground(AG)seagrassbiomassbetweengrazedandadjacentungrazedThalassia testudinumareas
Site
GPP RE NEP AG Biomass
difference
Bonaire
LacCaiBeach 818 909 656 793
StCroix
BIRNM 604 783 560 469
LittleCayman
GrapeTreeBay 922 955 891 933
899 796 960 933
885 870 900 933
Eleuthera
ArvidaBay 543 395 629 793
HalfSound 801 889 679 908
Florida
NorthRack 615 413 919 868
Note TheLittleCaymansitewassampledthreetimesin2016Allothersitesweresampledoncein2018SamplingtimesaregiveninSection21AbbreviationsBIRNMBuckIslandReefNationalMonumentsiteGPPgrossprimaryproductionNEPnetecosystemproductionREecosys-temrespiration
F I G U R E 3 emspRates(meanplusmnSD)ofnetecosystemproduction(a)grossprimaryproduction(b)andecosystemrespiration(c)fromgrazed(openpoints)andadjacentungrazed(solidpoints)areasofThalassia testudinumseagrassmeadowsRatesofNetecosystemproduction(NEP)weresignificantlyloweringrazedareasthanungrazedareasatallsitesexceptatLacCaiBeachBonaireandHalfSoundEleuthera(Section32)SitesorderedsouthtonorthBONBonaire(LCBLacCaiBeachsite)STXStCroixLCLittleCaymanELEleuthera(ABArvidaBaysiteHSHalfSoundsite)FLFloridaDashedlinein(a)denotesmetabolicbalance(NEP=0)Valuesabovelinerepresentnetmetaboliccarboncapture[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1108emsp |emsp emspenspJournal of Ecology JOHNSON et al
differencesinNEPbetweengrazedandungrazedareaswerenearlysignificant(pairedttestsp = 08 and p=09respectively)
Though NEP was significantly lower in grazed than ungrazedareas atmost sites rates ofNEPwere almost always positive asecosystem respiration was not stimulated in relation to primaryproductionevenwithconsiderablereductionofphotosyntheticsea-grassbiomassbygrazing(gt79biomassreductionatallsitesexceptStCroix Table3)NEP ranged from56 to96 lower in grazedareasthanungrazedareas(Table3)Asimilarrangeinthedifference
betweengrazedandungrazedareaswasmeasuredforgrossprimaryproduction (54ndash92 lower in grazed) and ecosystem respiration(40ndash96loweringrazed)
Halophila stipulacea meadows were present at three sites andratesofNEPintheseinvasiveseagrassmeadowswerecomparedtonearbynativeT testudinum-dominatedmeadowsNEPintheH stip-ulaceameadowswascomparedtonearbygrazedandungrazedareasofT testudinum-dominatedmeadowsattwoofthesites(StCroixandtheLacCaiBeachsiteinBonaire)WithineachsiteratesofNEPintheH stipulaceameadowwerebetweenthoseofthegrazedandungrazedareas (Figure 4) but the differences were not significant (ANOVApgt10forallpost-hoccomparisonswithineachsite)AnungrazedareawasnotavailableforcomparisonatthethirdsiteatwhichH stipula-ceawaspresent (NWLacBaysite inBonaire)asallT testudinumatthissitewasgrazedbygreenturtlesandratesofNEPdidnotdifferbetweenthe invasive-andnative-dominatedareasatthissiteeither(t test p = 56 Figure 4)Halophila stipulaceameadowNEP rangedfrom612plusmn226to989plusmn233mmolCmminus2 dayminus1(TableS2)anddidnotdiffersignificantlyamongthethreesitesatwhichitwaspresent(ANOVAF2=192p=23)eventhoughtheestablishmentofH stip-ulaceaseagrassmeadowsinStCroixwasrelativelyrecent(AGulickunpublisheddata)comparedtothoseinBonaire(Willetteetal2014)
33emsp|emspDrivers of metabolic rates
VariationinratesofseagrassecosystemmetabolismacrossgrazedandungrazedareasappearedtobedrivenbysomecharacteristicsoftheseagrassmeadowsbutnotbyenvironmentalfactorsMeadowNEPwasstronglypositivelyrelatedtoabove-groundseagrassbio-massacrosssites(linearregressionR2=82plt01Figure5a)NEP
F I G U R E 4 emspRates(meanplusmnSD)ofnetecosystemproductioninmeadowsdominatedbytheinvasiveseagrassHalophila stipulacea (triangles)comparedtonearbygrazed(opencircles)andungrazed(solidcircles)areasofThalassia testudinummeadowsBONBonaire(LCBLacCaiBeachsiteNWnorthwestLacBaysite)STXStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
F I G U R E 5 emspRelationshipbetweennetecosystemproductionandabove-groundseagrassbiomass(a)andtotalseagrassshootdensity(b)fromgrazed(openpoints)andungrazed(solidpoints)areasofThalassia testudinummeadowsSolidlinein(a)isthesignificantlinearregression(R2=82plt01)betweennetecosystemproduction(NEP)andbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows
emspensp emsp | emsp1109Journal of EcologyJOHNSON et al
wasalsopositivelycorrelatedwithseagrassmeadowcanopyheight(R2=84plt01)andbladesurfacearea(R2=92plt01)Above-groundbiomassislikelythemoreusefulpredictorofmeadowNEPhoweverasbiomassisinfluencedbybothcanopyheightandbladesurfaceareaandachange ineitherofthesevariablesresults inachangeinbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows(linearregressionR2lt01p=96Figure5b)norwerethererelationshipsbetweenNEPandenvironmentaltempera-ture(R2=01p=70)irradiance(R2lt01p=10)ormeadowdepth(R2lt01p=77)acrosssites(FigureS1)
4emsp |emspDISCUSSION
Through a comparative study of seagrass meadowsmdashgrazed andungrazedmdashacross a wide area encompassing sites in the GreaterCaribbean and Gulf of Mexico we found that green turtle graz-inghas a consistent effecton themetabolic carbon capture ratesofT testudinum-dominatedseagrassmeadowsRatesofmetaboliccarboncapturewerealwayslowerinareasgrazedbyturtlesthanun-grazedareasofmeadowsmdashupto96inLittleCaymanmdashbutratesofNEPwerealwaysnearorabovemetabolicbalanceEcosystemres-pirationwasnotstimulatedinrelationtoprimaryproductionwithinareas grazed by green turtles regardless of the amount of above-groundseagrassbiomassremovedthroughgrazingorotherdiffer-encesinbioticandabioticfactorsamongsitesThesefindingslendadditionalsupporttothehypothesisthatfutureratesofmetaboliccarboncapturemaybelowerinareasgrazedbyturtlesbutgrazingwillnot stimulateecosystemrespirationand result ina large rem-ineralizationof carbon currently stored in these seagrass habitats(Johnsonetal2017)
The similarity in results between St Croix (awintermeasure-ment)andtheotherfourlocationsinthepresentstudy(allsummermeasurements)strengthensourconclusionthatgrazingresultsinaconsistent response inmetaboliccarboncaptureacrossmeadowsHoweverwewerenotable toaddresspotential temporaldynam-icswithinsitesthatmayaffectnetcarboncaptureoveranannualcycle in these systems Future studies measuring carbon captureoverlongertemporalscales(ieseasonal)maybebeneficialtoourunderstanding of the effects of grazing on within-system carbondynamics
Thoughtheresponse inmetabolic rates tograzingwasconsis-tent acrossmeadows the strengthof this responsevariedamongindividual sites Some sites such as Little Cayman had consider-ablylowerratesofNEPingrazedareasthanungrazedareas(mean917)whereastheresponsetograzing(differenceinNEPbetweengrazedandadjacentungrazedareas)waslesspronouncedatothersites suchasStCroix (Table3) Seagrassmeadowcharacteristicssuch as shoot density canopy height and above-ground biomassare likely to bemore variable among ungrazed areas than grazedareasasgreenturtlegrazingresults insimilarcanopymorphome-tryacrossmeadowsintheGreaterCaribbeanregion(egBjorndal1980HernaacutendezampvanTussenbroek2014Williams1988)Given
the strong relationship between NEP and above-ground biomass(Figure 5a) high variability in NEPwould be expected across un-grazedmeadows(egFigure3a)IndeedtherangeinratesofNEPamongungrazedareaswasnearlythreetimesgreaterthantherangemeasuredacrossgrazedareasinthisstudyAssiteswerespecificallychosen that differed from each other in meadow characteristicsvariationinthestrengthofthemetabolicresponsetograzingwouldbeexpected
Variability in the strength of the metabolic response to graz-ing resulted in the lackofa significantdifferencebetweengrazedandungrazedareasattwositesmdashLacCaiBeachinBonaireandHalfSoundinEleuthera(Figure3a) It ispossiblethatotherfactorsnotaccounted for here such as local environmental factors or greenturtlegrazinghistory(egintensitylongevityofplotuse)mayhaveledtohighervariabilityinmetabolicratesatsomesitesandlackofastatisticaldifferenceInadditionmetabolicratesmayexhibitwithin-meadowvariation(patchiness)similartobelow-groundcarbonstor-age in meadows (OreskaMcglathery amp Porter 2017) leading tohighervariabilityinestimatesandhelpingtoexplainthelackofasig-nificantdifferenceinNEPatthesetwositesThiscouldbedriveninpartbywithin-meadowspatialheterogeneityinabove-groundbio-massgiventhestrongrelationshipbetweenbiomassandNEPHighwithin-meadow variability inmetabolic rates if presentmay alsohelpexplainthe largevariabilitymeasured intheungrazedarea inLittleCaymanFuturestudiesusingtheeddycorrelationtechnique(Bergetal2003) tomeasureecosystemmetabolismmaybeabletoovercomesomeofthiswithin-meadowvariabilityhoweverthegreatexpenseofthismethodmakesitprohibitiveformanystudies
Seagrass ecosystem metabolism is commonly estimated usingoxygendynamicsmeasuredwithbenthicincubationchambers(egBarroacutenetal2004Gaciaetal2005presentstudy)howeverthereare some limitations to thismethodBenthic chambersmay severbelow-ground rhizomesand release carbon that couldaffectmet-abolicestimatesIntactrhizomesontheotherhandcanaffectthediffusiveoxygenlossfromtheseagrasstotherhizosphere(BorumSand-JensenBinzerPedersenampGreve2006)andmayresultinanunderestimateofgrossprimaryproduction(andthereforeNEP)Thismaybeminimizedusingshorter incubation timesas in this studyIncubation length also strongly affects metabolic rates estimatedfromoxygendynamics (Oliveacute et al 2015)with longer incubationtimesbiasingestimatestowardsnetheterotrophyInadditiontoox-ygenmetabolismotherprocessessuchascarbonatedynamics(VanDamLopesOsburnampFourqurean2019)play important roles intotal ecosystem carbondynamicswithout affecting themeasuredproductivityofameadow
CaribbeanT testudinummeadows are highly productive (egJohnsonetal2017KochampMadden2001)andtherelationshipbetween biomass andNEP observed in this study suggests thatmeadows may maintain positive productivity as long as photo-synthetic biomass remains A threshold biomass below whichCaribbean meadows may become net heterotrophic cannot bedirectly addressed here but could be predicted to be near zeroThisisincontrasttoapreviouslysuggestedabove-groundbiomass
1110emsp |emsp emspenspJournal of Ecology JOHNSON et al
threshold(41gDMm2)belowwhichmeadowsmaybecomehet-erotrophic (Duarte et al 2010) However that threshold wascalculated for seagrasses globally and included less productivespecies Additional studies of the relationship between biomassandmeadowNEP from seagrass species and regions other thanthoseinthepresentstudywouldbebeneficialtofurthergeneral-izehowgrazingaffectsmetaboliccarboncaptureacrossseagrassecosystems
Environmental temperature and irradiance are strong driv-ers of seagrassmeadow productivity (Calleja et al 2006 Lee ampDunton 1997 Lee Park amp Kim 2007 Peacuterez amp Romero 1992)howeverpreviousstudiesexaminingcontrolsonseagrassmetabo-lismhavefocusedonungrazedseagrasses(egApostolakiHolmerMarbagraveampKarakassis2010Gaciaetal2005)Meadowmetabolicproductivity correlates stronglywith above-ground seagrass bio-mass(Johnsonetal2017Figure5a)thatcanbegreatlyaffectedby green turtle grazing (Bjorndal 1980CebriaacutenampDuarte 1998Christianenetal2012FourqureanManuelCoatesKenworthyampSmith2010Williams1988)Above-groundbiomassexplained82ofthevariationinNEPacrossgrazedandungrazedT testudi-nummeadowsinthisstudybutNEPwasnotcorrelatedwitheitherenvironmentaltemperatureorirradiance(FigureS1)WethereforeinvestigatedwhetherthelackofanobservedrelationshipbetweenNEPand temperatureand irradiancewasperhapsdue toa largereffectfromdifferencesinbiomasscausedbygrazingmaskinganyeffectoftheseenvironmentalvariablesWescaledmetabolicratestotheabove-groundbiomassineachgrazedandungrazedarea(unitmetabolismperunitbiomass)andcomparedthemtotemperatureandirradianceAccountingfordifferencesinabove-groundbiomassdidnot revealanunderlyingrelationshipbetweenNEPandtheseenvironmentalparametersacrossmeadows(grazedungrazedorallareascombinedFigureS2)ThelackofarelationshipbetweenNEPandenvironmentalparametersinthisstudymaybedueinparttothe relatively limited rangeof temperature and irradiance in sea-grassmeadowsgrazedbygreenturtles in theGreaterCaribbeanAllmeadowsinthisstudywerelocatedinrelativelyshallowwarmtropicalandsubtropicalenvironmentsreceivinghighlevelsofinci-dentsunlight
41emsp|emspEffects of Halophila stipulacea invasion on meadow carbon capture
In addition to the newmetabolic carbon capture estimates wecontribute from grazed areas in T testudinum meadows wealso present the first estimates of NEP from meadows domi-nated by H stipulacea seagrass Having been first reported inthe Caribbean in 2002 (Ruiz amp Ballantine 2004)H stipulacea has spread rapidly among islands and become invasive in thisregion (Christianenet al 2018Willette et al 2014)Halophila stipulaceamayalsospreadmorequicklywithinareasofseagrassgrazedbygreenturtlesthaninungrazedareas(Christianenetal2018) suggesting that grazing may inadvertently facilitate thespreadofthisinvasivespeciesonceithasbecomeestablishedin
anareaInLacBayBonairegreenturtleshavebeguntoexpandtheirgrazingplotsintoareasofpreviouslyungrazedT testudinum asH stipulacea slowly takes over the historically grazed areaswithin thebay (Christianenet al 2018 SmuldersVonk EngelampChristianen2017) If this relationshipbetweengreen turtlesnative seagrasses and invasive seagrass exists in other areas itcouldhaveimplicationsfortotalecosystemcarbondynamicsandcarbonstorageaswellasotherecosystemfunctionswithinthesemeadows
Our resultsdemonstrate that ratesofmetabolic carboncap-turemaybe similarbetweenH stipulaceameadowsand thena-tive T testudinum meadows that they are replacing (Figure 4)HowevergiventhelimitednumberofH stipulaceameadowswewereabletosample inthisstudy(n=3)cautioniswarrantedinextrapolating these findingswidely to other locations facing in-vasionbyH stipulacea seagrassWhile ratesofNEP inH stipu-laceameadowsweresimilar tobothgrazed (n=3)andungrazed(n=2)areasofT testudinum-dominatedmeadowsinStCroixandBonaire (within-site comparisons) multiple factors affect theseratesthatmaydifferandaffectthisrelationshipinotherlocations(egdominantspeciesofnativeseagrassdensityandbiomassofH stipulaceavsthenativespeciesbladeepiphytecoverage)Forexample this relationshipmay differ in areas such as Dominica(eastern Caribbean) where the meadows undergoing H stipu-lacea invasion are dominated byS filiforme seagrass (WilletteampAmbrose 2012) Measurements from additional locations willgreatlyaidinunderstandingthespatialextentofthesimilarityinmetaboliccarboncaptureratesbetweenH stipulaceaandnativeseagrassmeadowsseenhereHoweverthecomparablemetabolicratesbetweentheinvasiveandnativeseagrassesmeasuredinthisstudysuggestsomebenefitsofseagrasspresencemayberetainedinmeadowsfollowinginvasionbyH stipulaceasuchaslocalmit-igation of ocean acidification (through metabolic buffering ofpHCamp et al 2016Hendriks et al 2014Unsworth CollierHendersonampMcKenzie2012)
It is not yet known whether comparable rates of metaboliccarbon capture will translate to comparable carbon sequestra-tionor storage inmeadowsdominatedby invasiveH stipulacea UnlikeT testudinum(vanTussenbroeketal2006)seagrassspe-cieswithinthegenusHalophiladonotformadeepbelow-groundrhizome mat (Fonseca 1989) Robust rhizome mats formed bysomeseagrassspeciescontributetohigherbelow-groundcarbonstorage(Christianenetal2013)andthelackofasimilarrhizomematmayresult in lowersedimentcarbonstorageinH stipulacea meadowsHalophila tissues also decompose quickly in sediment(JosselynFonsecaNiesenampLarson1986)and thismayaffectseagrass-derivedcarboninputtothesedimentsininvadedmead-ows compared to native species with higher refractory organicmatter content (Trevathan-Tackett et al 2017) Further studieson carbondynamics inH stipulaceameadowswill beneeded toformabetterunderstandingofhowthecontinuedinvasionbythisspecieswill affect total ecosystem carbon dynamics in seagrassmeadowsintheCaribbean
emspensp emsp | emsp1111Journal of EcologyJOHNSON et al
42emsp|emspGreater global assessment of grazed meadows needed
UngrazedareasofT testudinum-dominatedseagrassmeadowsfromthisstudyexhibitedratesofNEP(median1040mmolCmminus2 dayminus1)neartheupperendofthosereportedintheliteratureforseagrassecosystemsandgrazedareasinthisstudyexhibitedrates(median381)nearthemedianvalueofNEPforallreportedungrazedsea-grassmeadows(median271Figure6)Measurementsofseagrassmetaboliccarboncapturearerelativelygeographicallylimitedhow-ever with the majority of measurementsmdashmostly from ungrazedT testudinum-dominated meadowsmdashcoming from the GreaterCaribbeanandGulfofMexicoregionsTheCaribbeanandGulfofMexicoareecologicallyimportantgrazingregionsbutonlyasingleestimateofseagrassmetabolismfromagrazedareaexistedpriortothisstudy(Johnsonetal2017)
Allmeadows in the present study exhibited a consistent re-sponseofmetaboliccarboncapturetogreenturtlegrazinghow-ever our studywas limited to the effects of grazing in tropicaland subtropicalT testudinum-dominatedmeadows It ispossiblethatmeadows inother regionsof theworldperhapsdominatedbydifferent seagrass speciesandunderdifferentenvironmentalconditionswillnotexhibitthesameresponsetograzingintermsofmetabolic carbon dynamicsWe are aware of only one otherstudy inwhichthiswas investigated Ina tropicalThalassia hem-prichiimeadow(Tanzania)NEPwasreducedasaresultofexper-imental clipping to simulategrazing (Dahletal2016)howeveronlyhourlyratesofdaytimeNEPweremeasuredWithoutcorre-spondingratesofrespirationwecannotevaluateifNEPremained
positive in thisT hemprichiimeadowoveradiel cycleand if theresponse toexperimental clipping inTanzaniawas similar to theresponse to grazing measured in our study Further studies oneffectsofgrazingonseagrassmeadowmetaboliccarbondynam-icswould be particularly beneficial in (a) additional areaswheregrazer abundance is increasing and (b) meadows dominated byspeciesotherthanT testudinum
5emsp |emspCONCLUSIONS
Given the importance of seagrass meadows as blue carbon eco-systems (Fourqureanetal2012)andtheir role incarbonseques-tration and potential climate changemitigation it is necessary tounderstand how meadow carbon dynamics are affected by graz-ing Our study adds critical information on the effects of grazingon seagrassmetabolic carbondynamics andhow theseprocessesmaybeaffectedwith increasinggreenturtleabundanceandgraz-ingWedemonstratethattheresponseinmetaboliccarboncapturetogreenturtlegrazing isconsistentacrossseagrassecosystems inthe Greater Caribbean and Gulf of Mexico regionsmdashan area thatsupportsvastexpansesofseagrass(GreenampShort2003Jackson1997Wabnitzetal2008)Thisisanecologicallyimportantgrazingregionforgreenturtlepopulationsandgivenincreasinggreenturtleabundanceacrosstheglobe(Chaloupkaetal2008Mazarisetal2017)itisexpectedthatmoreseagrassareawillreturntoanaturalgrazedstateOurresultssuggestthat increasedgrazingwill trans-latetolowerratesofmetaboliccarboncapturebutwillnotstimulaterespirationandthemetaboliclossofcarbonfromthesemeadows
F I G U R E 6 emspRates(meanplusmnSD)ofnetecosystemproductionfrommeadowsofvariousseagrassspeciescollectedfromtheliteratureOpencircleslsquocrossedrsquocirclesandopentrianglesaregrazedThalassia testudinum(n=7)ungrazedT testudinum(n=6)andHalophila stipulacea(n=3)meadowsrespectivelyfromthepresentstudyFilledcirclesarepreviouslypublishedestimatesofungrazedseagrassmeadownetecosystemproduction(NEP)andcomefromvariouslocationsaroundtheworldCriteriaforliteraturecollectionanddatainclusionaredescribedinJohnsonetal(2017)RatesofNEPfromungrazedT testudinum and H stipulaceameadowsfromthepresentstudyareneartheupperendofpublishedvaluesMedianNEPfromgrazedT testudinumareas(presentstudy)issimilartothemedianNEPofungrazedseagrassmeadowsglobally(Section42)FiguremodifiedfromJohnsonetal(2017)toincludeadditionalestimatesLettersabovepointsdenotesitesfromthepresentstudyndashBBonaireCLittleCaymanEEleutheraFFloridaSStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306
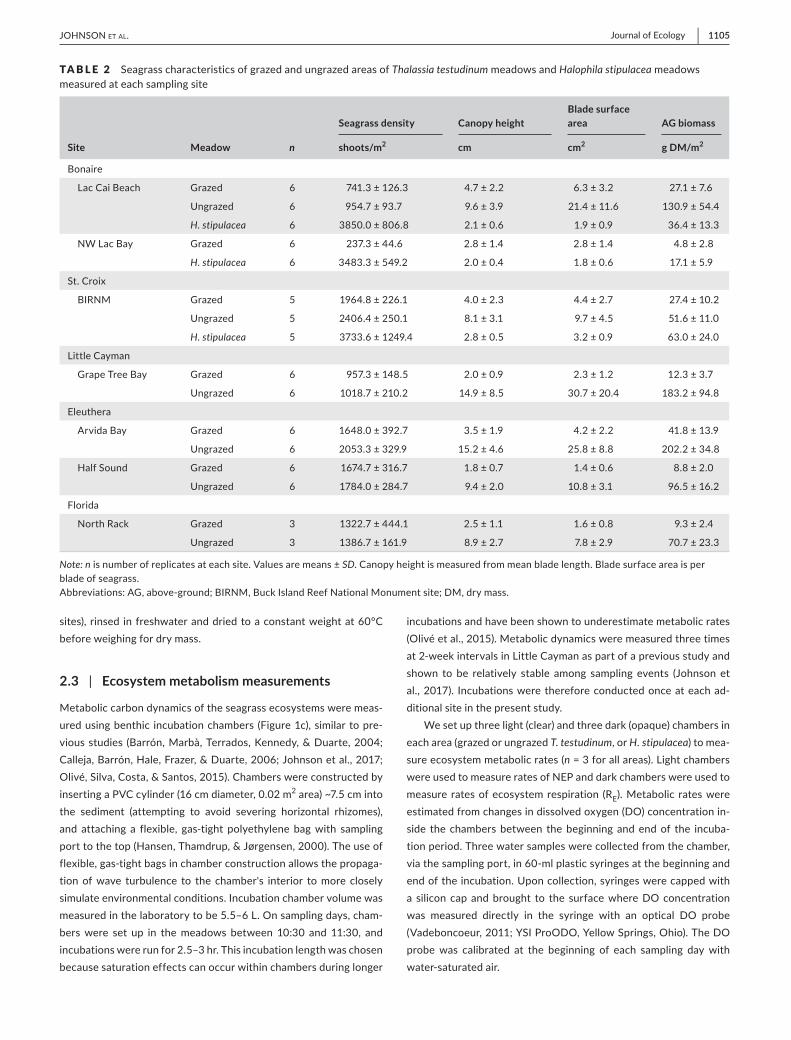
emspensp emsp | emsp1105Journal of EcologyJOHNSON et al
sites) rinsed infreshwateranddriedtoaconstantweightat60degCbeforeweighingfordrymass
23emsp|emspEcosystem metabolism measurements
Metaboliccarbondynamicsoftheseagrassecosystemsweremeas-uredusingbenthic incubationchambers (Figure1c)similartopre-viousstudies (BarroacutenMarbagraveTerradosKennedyampDuarte2004CallejaBarroacutenHaleFrazerampDuarte2006Johnsonetal2017OliveacuteSilvaCostaampSantos2015)ChamberswereconstructedbyinsertingaPVCcylinder(16cmdiameter002m2area)~75cmintothe sediment (attempting to avoid severing horizontal rhizomes)and attaching a flexible gas-tight polyethylene bagwith samplingporttothetop(HansenThamdrupampJoslashrgensen2000)Theuseofflexiblegas-tightbagsinchamberconstructionallowsthepropaga-tionofwave turbulence to the chambers interior tomore closelysimulateenvironmentalconditionsIncubationchambervolumewasmeasuredinthelaboratorytobe55ndash6LOnsamplingdayscham-berswere set up in themeadows between 1030 and 1130 andincubationswererunfor25ndash3hrThisincubationlengthwaschosenbecausesaturationeffectscanoccurwithinchambersduringlonger
incubationsandhavebeenshowntounderestimatemetabolicrates(Oliveacuteetal2015)Metabolicdynamicsweremeasuredthreetimesat2-weekintervalsinLittleCaymanaspartofapreviousstudyandshown to be relatively stable among sampling events (Johnson etal2017) Incubationswerethereforeconductedonceateachad-ditionalsiteinthepresentstudy
Wesetupthreelight(clear)andthreedark(opaque)chambersineacharea(grazedorungrazedT testudinumorH stipulacea)tomea-sureecosystemmetabolicrates(n=3forallareas)LightchamberswereusedtomeasureratesofNEPanddarkchamberswereusedtomeasure ratesofecosystemrespiration (RE)Metabolic rateswereestimatedfromchangesindissolvedoxygen(DO)concentrationin-side the chambersbetween thebeginningandendof the incuba-tionperiodThreewatersampleswerecollectedfromthechamberviathesamplingportin60-mlplasticsyringesatthebeginningandendoftheincubationUponcollectionsyringeswerecappedwitha silicon cap andbrought to the surfacewhereDOconcentrationwas measured directly in the syringe with an optical DO probe(Vadeboncoeur2011YSIProODOYellowSpringsOhio)TheDOprobewas calibrated at the beginning of each sampling daywithwater-saturatedair
TA B L E 2 emspSeagrasscharacteristicsofgrazedandungrazedareasofThalassia testudinummeadowsandHalophila stipulaceameadowsmeasuredateachsamplingsite
Site Meadow n
Seagrass density Canopy heightBlade surface area AG biomass
shootsm2 cm cm2 g DMm2
Bonaire
LacCaiBeach Grazed 6 7413plusmn1263 47plusmn22 63plusmn32 271plusmn76
Ungrazed 6 9547plusmn937 96plusmn39 214plusmn116 1309plusmn544
H stipulacea 6 38500plusmn8068 21plusmn06 19plusmn09 364plusmn133
NWLacBay Grazed 6 2373plusmn446 28plusmn14 28plusmn14 48plusmn28
H stipulacea 6 34833plusmn5492 20plusmn04 18plusmn06 171plusmn59
StCroix
BIRNM Grazed 5 19648plusmn2261 40plusmn23 44plusmn27 274plusmn102
Ungrazed 5 24064plusmn2501 81plusmn31 97plusmn45 516plusmn110
H stipulacea 5 37336plusmn12494 28plusmn05 32plusmn09 630plusmn240
LittleCayman
GrapeTreeBay Grazed 6 9573plusmn1485 20plusmn09 23plusmn12 123plusmn37
Ungrazed 6 10187plusmn2102 149plusmn85 307plusmn204 1832plusmn948
Eleuthera
ArvidaBay Grazed 6 16480plusmn3927 35plusmn19 42plusmn22 418plusmn139
Ungrazed 6 20533plusmn3299 152plusmn46 258plusmn88 2022plusmn348
HalfSound Grazed 6 16747plusmn3167 18plusmn07 14plusmn06 88plusmn20
Ungrazed 6 17840plusmn2847 94plusmn20 108plusmn31 965plusmn162
Florida
NorthRack Grazed 3 13227plusmn4441 25plusmn11 16plusmn08 93plusmn24
Ungrazed 3 13867plusmn1619 89plusmn27 78plusmn29 707plusmn233
Note nisnumberofreplicatesateachsiteValuesaremeansplusmnSDCanopyheightismeasuredfrommeanbladelengthBladesurfaceareaisperbladeofseagrassAbbreviationsAGabove-groundBIRNMBuckIslandReefNationalMonumentsiteDMdrymass
1106emsp |emsp emspenspJournal of Ecology JOHNSON et al
Tomeasureonlymetabolicratesoftheseagrassbenthiccompo-nentsofthesystem(thoseaffectingcarbonstorage)wecorrectedrates measured from incubation chambers for water column me-tabolismWemeasuredwater columnmetabolismusingbiologicaloxygendemand(BOD)bottlesatthesametimeasbenthicchamberincubationsThreeclear300-mlglassBODbottlesandthreeopaque300-mlglassBODbottleswereusedtomeasurewatercolumnpro-ductionandrespirationrespectivelyBottleswerefilledwithwaterat seagrasscanopyheightanchored to thebottomand incubatedunderinsituconditionsintheseagrassmeadowWatercolumnsam-pleswerecollectedatthebeginningofincubationsin60-mlsyringesatcanopyheightandDOconcentrationwasmeasuredinthesamemannerassamplesfromincubationchambersFollowingthe incu-bationperiodbottleswerecollectedand returned to thesurfaceonesamplewascollectedfromeachbottlewithasyringeandDOconcentrationwasmeasured
Hourlymetabolic rateswere calculated from thedifference inDOconcentrationwithinlight(NEP)anddark(RE)incubationcham-bersbetweenthebeginningandendoftheincubationperiodWatercolumn metabolic rates measured from BOD bottles were sub-tractedfromratesmeasuredinchambersHourlygrossprimarypro-duction(GPP)wasthencalculatedasthesumofhourlyNEPandRE DailyratesofGPPwerecalculatedbymultiplyinghourlyGPPbythelengthofthephotoperiodmultipliedby075(toaccountfor lowermetabolicratesatdawnandduskJohnsonetal2017Oliveacuteetal2015)anddailyratesofREwerecalculatedbymultiplyinghourlyRE by24hoursDailyNEPwascalculatedas thedifferencebetweendaily GPP and RE Therewere a few occasionswhen an increaseinDO concentrationwasmeasuredwithin a dark chamber or thedarkBODbottlesduringanincubationsuggestinganerrorThesemeasuredincreasesweresmallusuallywithintheprobesmarginoferror (01mgO2L)andweassumedrespirationtobezeroduringthese timesCorrecting for these respiration errors hadminor ef-fectsondailymetabolic rates in themeadows (TablesS2andS3)MeasuredDOconcentrationswereconvertedfrommgO2tommolO2andthentocarbonunits(mmolC)assumingphotosyntheticandrespiratoryquotientsofone(BarroacutenampDuarte2009)
24emsp|emspData analyses
Net ecosystem productionwas themetabolic variable of primaryinterestasitaccountsforbothcarboncaptureandlossandcanbeusedas an indicatorofwhether a system is currently ametaboliccarbonsinkorsourceThereforeNEPisthemetabolicvariableforwhichstatisticalresultsarepresented
Ouraimwastoevaluatewhethergreenturtlegrazinghasacon-sistenteffectonmetabolicratesacrossseagrassecosystemsmdashthatistoevaluatewhetheratanygivensiteatanygiventimeratesofNEPareloweringrazedareasthaninungrazedareasWeevaluateddifferences in ratesofNEPbetweenareasofgrazedandadjacentungrazedT testudinum from each sitewith paired t tests T testswereusedbecause thecomparisonof interestwas thedifferencebetween grazed and ungrazed areas (effect of grazing) within a
givensitenotdifferencesinNEPamongsitesForsiteswheretheinvasive seagrass H stipulacea was present we used one-wayANOVAstotestforsignificantdifferencesinratesofNEPbetweenH stipulaceameadowsandnearbygrazedandungrazedT testudinum areas(StCroixsiteandLacCaiBeachsiteBonaire)Forthenorth-west site in Lac Bay Bonaire where only a grazed area of T te-studinum and an invasive H stipulacea meadow were comparedanunpaired t testwasused IfANOVA resultswere significant aTukeyspost-hoctestwasusedtoidentifysignificantcomparisonsWetestedfordifferencesinseagrassmeadowparametersbetweengrazedandungrazedT testudinumareasandH stipulaceameadowsatsitesusingpairedttestsandANOVAsinthesamemannerasforNEPWeused linearregressiontotestrelationshipsbetweenNEPand explanatory variables (eg seagrass biomass environmentaltemperature)MeanvaluesforNEPandexplanatoryvariablesfromeachmeadowwereusedforlinearregressionstestssotherewasasingledatapointforeachgrazedandungrazedT testudinum area or H stipulaceameadowateachsamplingsite
AllanalyseswereperformedinRversion343(RCoreTeam2018)Datawere processed using the lsquodplyrrsquo package (Wickham FrancoisHenryampMuumlller2017)andpost-hocanalyseswereperformedusingthelsquoagricolaersquopackage(deMendiburu2017)Alldatamettheassump-tionsfortheseanalysesanddataarepresentedasmeanplusmnSD
3emsp |emspRESULTS
31emsp|emspSeagrass meadow characteristics
Total seagrass shoot densities varied widely among meadowsand locations (Table 2) though shoot densitieswere always high-est inH stipulaceameadows The lowest shoot density observed(2373plusmn446shootsm2)wasinagrazedarea(NWsiteinLacBayBonaire)buttherewasnocleartrendforhigherorlowerseagrassdensities between grazed and ungrazed areas across the sampledsitesThalassia testudinum density in LittleCaymanwasgreater inthe grazed area than ungrazed area (paired t test p = 02) whileT testudinumdensityattheLacCaiBeachsiteinBonairewasgreaterintheungrazedarea(pairedttestp=02)Thalassia testudinum den-sitydidnotdifferbetweengrazedandungrazedareasattheremain-ingsites(pairedttestspgt05)ShootdensityofH stipulaceawasnot significantly different between any of the threeH stipulacea meadowsmeasured(ANOVAF2=009p=92)
HalophilaseagrassestypicallygrowshortbladesCanopyheightoftheH stipulaceameadowwassimilartothatofthenearbygrazedarea inStCroix andwas shorter than thenearbygrazedareasatbothsitesinBonaire(Table2)Halophila stipulaceabladelengthwasgreaterinStCroixthanBonaire(ANOVAF2=2413plt01)AreasofT testudinum-dominatedseagrassmeadowsgrazedbygreentur-tleshad significantly shorterblades and therefore canopyheightthan adjacent ungrazed areas at all sampled sites (paired t testsplt05forallcomparisonsTable2)
Greenturtlesbycroppingseagrassbladesnearthesedimentsur-faceandreducingcanopyheightgreatlyreducetheabove-ground
emspensp emsp | emsp1107Journal of EcologyJOHNSON et al
seagrass biomass of meadows within their grazing areas Above-ground biomass ranged from 48 plusmn 28 to 418 plusmn 139 g DMm2 amonggrazedareas(Table2)andfrom516plusmn110to2022plusmn348gDMm2amongungrazedareasBiomasswasalwaysloweringrazedareas than adjacent ungrazed areas (p lt 05 for all comparisons)Biomass among the three sampledH stipulaceameadows rangedfrom171plusmn59to630plusmn240gDMm2TheH stipulacea meadow in St Croix had significantly greater above-ground biomass thaneither of theH stipulaceameadows sampled inBonaire (ANOVAF2=1202plt 01)Evidenceofgreenturtlegrazingwasonlyob-served in T testudinum-dominated meadows in this study notH stipulaceameadowsandgrazingresultedinagt79decreaseinbiomassingrazedcomparedtoungrazedareasatallsitesexceptStCroix(47Table3)
32emsp|emspEcosystem metabolic rates
Rates of NEP differed among meadow types (grazed ungrazedH stipulacea) across the studied locations (Figures 3 and 4 seeTable S2 for data) Across all sites NEP ranged from 42 plusmn 70 to517plusmn110mmolCmminus2 dayminus1amonggrazedareasandfrom523plusmn57to2253plusmn191mmolCmminus2 dayminus1amongungrazedareasThelargestandarddeviationsonsomeoftheungrazedareaNEPestimatesfromLittleCaymanwerearesultofoneincubationchamberproducingratesmuchdifferentfromtheothertwoAcrossallsitesNEPwasconsist-entlyloweringrazedareasthanungrazedareas(Figure3)DifferencesinNEPbetweengrazedandungrazedareasweresignificantat four
ofthesixsiteswhereungrazedT testudinumwaspresentStCroixLittleCayman (all threemeasurement times) theArvidaBay site inEleutheraandFlorida(pairedttestsplt05forallcomparisons)AttheLacCaiBeachsiteinBonaireandtheHalfSoundsiteinEleuthera
TA B L E 3 emspPercentdifferenceinmetabolicratesandabove-ground(AG)seagrassbiomassbetweengrazedandadjacentungrazedThalassia testudinumareas
Site
GPP RE NEP AG Biomass
difference
Bonaire
LacCaiBeach 818 909 656 793
StCroix
BIRNM 604 783 560 469
LittleCayman
GrapeTreeBay 922 955 891 933
899 796 960 933
885 870 900 933
Eleuthera
ArvidaBay 543 395 629 793
HalfSound 801 889 679 908
Florida
NorthRack 615 413 919 868
Note TheLittleCaymansitewassampledthreetimesin2016Allothersitesweresampledoncein2018SamplingtimesaregiveninSection21AbbreviationsBIRNMBuckIslandReefNationalMonumentsiteGPPgrossprimaryproductionNEPnetecosystemproductionREecosys-temrespiration
F I G U R E 3 emspRates(meanplusmnSD)ofnetecosystemproduction(a)grossprimaryproduction(b)andecosystemrespiration(c)fromgrazed(openpoints)andadjacentungrazed(solidpoints)areasofThalassia testudinumseagrassmeadowsRatesofNetecosystemproduction(NEP)weresignificantlyloweringrazedareasthanungrazedareasatallsitesexceptatLacCaiBeachBonaireandHalfSoundEleuthera(Section32)SitesorderedsouthtonorthBONBonaire(LCBLacCaiBeachsite)STXStCroixLCLittleCaymanELEleuthera(ABArvidaBaysiteHSHalfSoundsite)FLFloridaDashedlinein(a)denotesmetabolicbalance(NEP=0)Valuesabovelinerepresentnetmetaboliccarboncapture[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1108emsp |emsp emspenspJournal of Ecology JOHNSON et al
differencesinNEPbetweengrazedandungrazedareaswerenearlysignificant(pairedttestsp = 08 and p=09respectively)
Though NEP was significantly lower in grazed than ungrazedareas atmost sites rates ofNEPwere almost always positive asecosystem respiration was not stimulated in relation to primaryproductionevenwithconsiderablereductionofphotosyntheticsea-grassbiomassbygrazing(gt79biomassreductionatallsitesexceptStCroix Table3)NEP ranged from56 to96 lower in grazedareasthanungrazedareas(Table3)Asimilarrangeinthedifference
betweengrazedandungrazedareaswasmeasuredforgrossprimaryproduction (54ndash92 lower in grazed) and ecosystem respiration(40ndash96loweringrazed)
Halophila stipulacea meadows were present at three sites andratesofNEPintheseinvasiveseagrassmeadowswerecomparedtonearbynativeT testudinum-dominatedmeadowsNEPintheH stip-ulaceameadowswascomparedtonearbygrazedandungrazedareasofT testudinum-dominatedmeadowsattwoofthesites(StCroixandtheLacCaiBeachsiteinBonaire)WithineachsiteratesofNEPintheH stipulaceameadowwerebetweenthoseofthegrazedandungrazedareas (Figure 4) but the differences were not significant (ANOVApgt10forallpost-hoccomparisonswithineachsite)AnungrazedareawasnotavailableforcomparisonatthethirdsiteatwhichH stipula-ceawaspresent (NWLacBaysite inBonaire)asallT testudinumatthissitewasgrazedbygreenturtlesandratesofNEPdidnotdifferbetweenthe invasive-andnative-dominatedareasatthissiteeither(t test p = 56 Figure 4)Halophila stipulaceameadowNEP rangedfrom612plusmn226to989plusmn233mmolCmminus2 dayminus1(TableS2)anddidnotdiffersignificantlyamongthethreesitesatwhichitwaspresent(ANOVAF2=192p=23)eventhoughtheestablishmentofH stip-ulaceaseagrassmeadowsinStCroixwasrelativelyrecent(AGulickunpublisheddata)comparedtothoseinBonaire(Willetteetal2014)
33emsp|emspDrivers of metabolic rates
VariationinratesofseagrassecosystemmetabolismacrossgrazedandungrazedareasappearedtobedrivenbysomecharacteristicsoftheseagrassmeadowsbutnotbyenvironmentalfactorsMeadowNEPwasstronglypositivelyrelatedtoabove-groundseagrassbio-massacrosssites(linearregressionR2=82plt01Figure5a)NEP
F I G U R E 4 emspRates(meanplusmnSD)ofnetecosystemproductioninmeadowsdominatedbytheinvasiveseagrassHalophila stipulacea (triangles)comparedtonearbygrazed(opencircles)andungrazed(solidcircles)areasofThalassia testudinummeadowsBONBonaire(LCBLacCaiBeachsiteNWnorthwestLacBaysite)STXStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
F I G U R E 5 emspRelationshipbetweennetecosystemproductionandabove-groundseagrassbiomass(a)andtotalseagrassshootdensity(b)fromgrazed(openpoints)andungrazed(solidpoints)areasofThalassia testudinummeadowsSolidlinein(a)isthesignificantlinearregression(R2=82plt01)betweennetecosystemproduction(NEP)andbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows
emspensp emsp | emsp1109Journal of EcologyJOHNSON et al
wasalsopositivelycorrelatedwithseagrassmeadowcanopyheight(R2=84plt01)andbladesurfacearea(R2=92plt01)Above-groundbiomassislikelythemoreusefulpredictorofmeadowNEPhoweverasbiomassisinfluencedbybothcanopyheightandbladesurfaceareaandachange ineitherofthesevariablesresults inachangeinbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows(linearregressionR2lt01p=96Figure5b)norwerethererelationshipsbetweenNEPandenvironmentaltempera-ture(R2=01p=70)irradiance(R2lt01p=10)ormeadowdepth(R2lt01p=77)acrosssites(FigureS1)
4emsp |emspDISCUSSION
Through a comparative study of seagrass meadowsmdashgrazed andungrazedmdashacross a wide area encompassing sites in the GreaterCaribbean and Gulf of Mexico we found that green turtle graz-inghas a consistent effecton themetabolic carbon capture ratesofT testudinum-dominatedseagrassmeadowsRatesofmetaboliccarboncapturewerealwayslowerinareasgrazedbyturtlesthanun-grazedareasofmeadowsmdashupto96inLittleCaymanmdashbutratesofNEPwerealwaysnearorabovemetabolicbalanceEcosystemres-pirationwasnotstimulatedinrelationtoprimaryproductionwithinareas grazed by green turtles regardless of the amount of above-groundseagrassbiomassremovedthroughgrazingorotherdiffer-encesinbioticandabioticfactorsamongsitesThesefindingslendadditionalsupporttothehypothesisthatfutureratesofmetaboliccarboncapturemaybelowerinareasgrazedbyturtlesbutgrazingwillnot stimulateecosystemrespirationand result ina large rem-ineralizationof carbon currently stored in these seagrass habitats(Johnsonetal2017)
The similarity in results between St Croix (awintermeasure-ment)andtheotherfourlocationsinthepresentstudy(allsummermeasurements)strengthensourconclusionthatgrazingresultsinaconsistent response inmetaboliccarboncaptureacrossmeadowsHoweverwewerenotable toaddresspotential temporaldynam-icswithinsitesthatmayaffectnetcarboncaptureoveranannualcycle in these systems Future studies measuring carbon captureoverlongertemporalscales(ieseasonal)maybebeneficialtoourunderstanding of the effects of grazing on within-system carbondynamics
Thoughtheresponse inmetabolic rates tograzingwasconsis-tent acrossmeadows the strengthof this responsevariedamongindividual sites Some sites such as Little Cayman had consider-ablylowerratesofNEPingrazedareasthanungrazedareas(mean917)whereastheresponsetograzing(differenceinNEPbetweengrazedandadjacentungrazedareas)waslesspronouncedatothersites suchasStCroix (Table3) Seagrassmeadowcharacteristicssuch as shoot density canopy height and above-ground biomassare likely to bemore variable among ungrazed areas than grazedareasasgreenturtlegrazingresults insimilarcanopymorphome-tryacrossmeadowsintheGreaterCaribbeanregion(egBjorndal1980HernaacutendezampvanTussenbroek2014Williams1988)Given
the strong relationship between NEP and above-ground biomass(Figure 5a) high variability in NEPwould be expected across un-grazedmeadows(egFigure3a)IndeedtherangeinratesofNEPamongungrazedareaswasnearlythreetimesgreaterthantherangemeasuredacrossgrazedareasinthisstudyAssiteswerespecificallychosen that differed from each other in meadow characteristicsvariationinthestrengthofthemetabolicresponsetograzingwouldbeexpected
Variability in the strength of the metabolic response to graz-ing resulted in the lackofa significantdifferencebetweengrazedandungrazedareasattwositesmdashLacCaiBeachinBonaireandHalfSoundinEleuthera(Figure3a) It ispossiblethatotherfactorsnotaccounted for here such as local environmental factors or greenturtlegrazinghistory(egintensitylongevityofplotuse)mayhaveledtohighervariabilityinmetabolicratesatsomesitesandlackofastatisticaldifferenceInadditionmetabolicratesmayexhibitwithin-meadowvariation(patchiness)similartobelow-groundcarbonstor-age in meadows (OreskaMcglathery amp Porter 2017) leading tohighervariabilityinestimatesandhelpingtoexplainthelackofasig-nificantdifferenceinNEPatthesetwositesThiscouldbedriveninpartbywithin-meadowspatialheterogeneityinabove-groundbio-massgiventhestrongrelationshipbetweenbiomassandNEPHighwithin-meadow variability inmetabolic rates if presentmay alsohelpexplainthe largevariabilitymeasured intheungrazedarea inLittleCaymanFuturestudiesusingtheeddycorrelationtechnique(Bergetal2003) tomeasureecosystemmetabolismmaybeabletoovercomesomeofthiswithin-meadowvariabilityhoweverthegreatexpenseofthismethodmakesitprohibitiveformanystudies
Seagrass ecosystem metabolism is commonly estimated usingoxygendynamicsmeasuredwithbenthicincubationchambers(egBarroacutenetal2004Gaciaetal2005presentstudy)howeverthereare some limitations to thismethodBenthic chambersmay severbelow-ground rhizomesand release carbon that couldaffectmet-abolicestimatesIntactrhizomesontheotherhandcanaffectthediffusiveoxygenlossfromtheseagrasstotherhizosphere(BorumSand-JensenBinzerPedersenampGreve2006)andmayresultinanunderestimateofgrossprimaryproduction(andthereforeNEP)Thismaybeminimizedusingshorter incubation timesas in this studyIncubation length also strongly affects metabolic rates estimatedfromoxygendynamics (Oliveacute et al 2015)with longer incubationtimesbiasingestimatestowardsnetheterotrophyInadditiontoox-ygenmetabolismotherprocessessuchascarbonatedynamics(VanDamLopesOsburnampFourqurean2019)play important roles intotal ecosystem carbondynamicswithout affecting themeasuredproductivityofameadow
CaribbeanT testudinummeadows are highly productive (egJohnsonetal2017KochampMadden2001)andtherelationshipbetween biomass andNEP observed in this study suggests thatmeadows may maintain positive productivity as long as photo-synthetic biomass remains A threshold biomass below whichCaribbean meadows may become net heterotrophic cannot bedirectly addressed here but could be predicted to be near zeroThisisincontrasttoapreviouslysuggestedabove-groundbiomass
1110emsp |emsp emspenspJournal of Ecology JOHNSON et al
threshold(41gDMm2)belowwhichmeadowsmaybecomehet-erotrophic (Duarte et al 2010) However that threshold wascalculated for seagrasses globally and included less productivespecies Additional studies of the relationship between biomassandmeadowNEP from seagrass species and regions other thanthoseinthepresentstudywouldbebeneficialtofurthergeneral-izehowgrazingaffectsmetaboliccarboncaptureacrossseagrassecosystems
Environmental temperature and irradiance are strong driv-ers of seagrassmeadow productivity (Calleja et al 2006 Lee ampDunton 1997 Lee Park amp Kim 2007 Peacuterez amp Romero 1992)howeverpreviousstudiesexaminingcontrolsonseagrassmetabo-lismhavefocusedonungrazedseagrasses(egApostolakiHolmerMarbagraveampKarakassis2010Gaciaetal2005)Meadowmetabolicproductivity correlates stronglywith above-ground seagrass bio-mass(Johnsonetal2017Figure5a)thatcanbegreatlyaffectedby green turtle grazing (Bjorndal 1980CebriaacutenampDuarte 1998Christianenetal2012FourqureanManuelCoatesKenworthyampSmith2010Williams1988)Above-groundbiomassexplained82ofthevariationinNEPacrossgrazedandungrazedT testudi-nummeadowsinthisstudybutNEPwasnotcorrelatedwitheitherenvironmentaltemperatureorirradiance(FigureS1)WethereforeinvestigatedwhetherthelackofanobservedrelationshipbetweenNEPand temperatureand irradiancewasperhapsdue toa largereffectfromdifferencesinbiomasscausedbygrazingmaskinganyeffectoftheseenvironmentalvariablesWescaledmetabolicratestotheabove-groundbiomassineachgrazedandungrazedarea(unitmetabolismperunitbiomass)andcomparedthemtotemperatureandirradianceAccountingfordifferencesinabove-groundbiomassdidnot revealanunderlyingrelationshipbetweenNEPandtheseenvironmentalparametersacrossmeadows(grazedungrazedorallareascombinedFigureS2)ThelackofarelationshipbetweenNEPandenvironmentalparametersinthisstudymaybedueinparttothe relatively limited rangeof temperature and irradiance in sea-grassmeadowsgrazedbygreenturtles in theGreaterCaribbeanAllmeadowsinthisstudywerelocatedinrelativelyshallowwarmtropicalandsubtropicalenvironmentsreceivinghighlevelsofinci-dentsunlight
41emsp|emspEffects of Halophila stipulacea invasion on meadow carbon capture
In addition to the newmetabolic carbon capture estimates wecontribute from grazed areas in T testudinum meadows wealso present the first estimates of NEP from meadows domi-nated by H stipulacea seagrass Having been first reported inthe Caribbean in 2002 (Ruiz amp Ballantine 2004)H stipulacea has spread rapidly among islands and become invasive in thisregion (Christianenet al 2018Willette et al 2014)Halophila stipulaceamayalsospreadmorequicklywithinareasofseagrassgrazedbygreenturtlesthaninungrazedareas(Christianenetal2018) suggesting that grazing may inadvertently facilitate thespreadofthisinvasivespeciesonceithasbecomeestablishedin
anareaInLacBayBonairegreenturtleshavebeguntoexpandtheirgrazingplotsintoareasofpreviouslyungrazedT testudinum asH stipulacea slowly takes over the historically grazed areaswithin thebay (Christianenet al 2018 SmuldersVonk EngelampChristianen2017) If this relationshipbetweengreen turtlesnative seagrasses and invasive seagrass exists in other areas itcouldhaveimplicationsfortotalecosystemcarbondynamicsandcarbonstorageaswellasotherecosystemfunctionswithinthesemeadows
Our resultsdemonstrate that ratesofmetabolic carboncap-turemaybe similarbetweenH stipulaceameadowsand thena-tive T testudinum meadows that they are replacing (Figure 4)HowevergiventhelimitednumberofH stipulaceameadowswewereabletosample inthisstudy(n=3)cautioniswarrantedinextrapolating these findingswidely to other locations facing in-vasionbyH stipulacea seagrassWhile ratesofNEP inH stipu-laceameadowsweresimilar tobothgrazed (n=3)andungrazed(n=2)areasofT testudinum-dominatedmeadowsinStCroixandBonaire (within-site comparisons) multiple factors affect theseratesthatmaydifferandaffectthisrelationshipinotherlocations(egdominantspeciesofnativeseagrassdensityandbiomassofH stipulaceavsthenativespeciesbladeepiphytecoverage)Forexample this relationshipmay differ in areas such as Dominica(eastern Caribbean) where the meadows undergoing H stipu-lacea invasion are dominated byS filiforme seagrass (WilletteampAmbrose 2012) Measurements from additional locations willgreatlyaidinunderstandingthespatialextentofthesimilarityinmetaboliccarboncaptureratesbetweenH stipulaceaandnativeseagrassmeadowsseenhereHoweverthecomparablemetabolicratesbetweentheinvasiveandnativeseagrassesmeasuredinthisstudysuggestsomebenefitsofseagrasspresencemayberetainedinmeadowsfollowinginvasionbyH stipulaceasuchaslocalmit-igation of ocean acidification (through metabolic buffering ofpHCamp et al 2016Hendriks et al 2014Unsworth CollierHendersonampMcKenzie2012)
It is not yet known whether comparable rates of metaboliccarbon capture will translate to comparable carbon sequestra-tionor storage inmeadowsdominatedby invasiveH stipulacea UnlikeT testudinum(vanTussenbroeketal2006)seagrassspe-cieswithinthegenusHalophiladonotformadeepbelow-groundrhizome mat (Fonseca 1989) Robust rhizome mats formed bysomeseagrassspeciescontributetohigherbelow-groundcarbonstorage(Christianenetal2013)andthelackofasimilarrhizomematmayresult in lowersedimentcarbonstorageinH stipulacea meadowsHalophila tissues also decompose quickly in sediment(JosselynFonsecaNiesenampLarson1986)and thismayaffectseagrass-derivedcarboninputtothesedimentsininvadedmead-ows compared to native species with higher refractory organicmatter content (Trevathan-Tackett et al 2017) Further studieson carbondynamics inH stipulaceameadowswill beneeded toformabetterunderstandingofhowthecontinuedinvasionbythisspecieswill affect total ecosystem carbon dynamics in seagrassmeadowsintheCaribbean
emspensp emsp | emsp1111Journal of EcologyJOHNSON et al
42emsp|emspGreater global assessment of grazed meadows needed
UngrazedareasofT testudinum-dominatedseagrassmeadowsfromthisstudyexhibitedratesofNEP(median1040mmolCmminus2 dayminus1)neartheupperendofthosereportedintheliteratureforseagrassecosystemsandgrazedareasinthisstudyexhibitedrates(median381)nearthemedianvalueofNEPforallreportedungrazedsea-grassmeadows(median271Figure6)Measurementsofseagrassmetaboliccarboncapturearerelativelygeographicallylimitedhow-ever with the majority of measurementsmdashmostly from ungrazedT testudinum-dominated meadowsmdashcoming from the GreaterCaribbeanandGulfofMexicoregionsTheCaribbeanandGulfofMexicoareecologicallyimportantgrazingregionsbutonlyasingleestimateofseagrassmetabolismfromagrazedareaexistedpriortothisstudy(Johnsonetal2017)
Allmeadows in the present study exhibited a consistent re-sponseofmetaboliccarboncapturetogreenturtlegrazinghow-ever our studywas limited to the effects of grazing in tropicaland subtropicalT testudinum-dominatedmeadows It ispossiblethatmeadows inother regionsof theworldperhapsdominatedbydifferent seagrass speciesandunderdifferentenvironmentalconditionswillnotexhibitthesameresponsetograzingintermsofmetabolic carbon dynamicsWe are aware of only one otherstudy inwhichthiswas investigated Ina tropicalThalassia hem-prichiimeadow(Tanzania)NEPwasreducedasaresultofexper-imental clipping to simulategrazing (Dahletal2016)howeveronlyhourlyratesofdaytimeNEPweremeasuredWithoutcorre-spondingratesofrespirationwecannotevaluateifNEPremained
positive in thisT hemprichiimeadowoveradiel cycleand if theresponse toexperimental clipping inTanzaniawas similar to theresponse to grazing measured in our study Further studies oneffectsofgrazingonseagrassmeadowmetaboliccarbondynam-icswould be particularly beneficial in (a) additional areaswheregrazer abundance is increasing and (b) meadows dominated byspeciesotherthanT testudinum
5emsp |emspCONCLUSIONS
Given the importance of seagrass meadows as blue carbon eco-systems (Fourqureanetal2012)andtheir role incarbonseques-tration and potential climate changemitigation it is necessary tounderstand how meadow carbon dynamics are affected by graz-ing Our study adds critical information on the effects of grazingon seagrassmetabolic carbondynamics andhow theseprocessesmaybeaffectedwith increasinggreenturtleabundanceandgraz-ingWedemonstratethattheresponseinmetaboliccarboncapturetogreenturtlegrazing isconsistentacrossseagrassecosystems inthe Greater Caribbean and Gulf of Mexico regionsmdashan area thatsupportsvastexpansesofseagrass(GreenampShort2003Jackson1997Wabnitzetal2008)Thisisanecologicallyimportantgrazingregionforgreenturtlepopulationsandgivenincreasinggreenturtleabundanceacrosstheglobe(Chaloupkaetal2008Mazarisetal2017)itisexpectedthatmoreseagrassareawillreturntoanaturalgrazedstateOurresultssuggestthat increasedgrazingwill trans-latetolowerratesofmetaboliccarboncapturebutwillnotstimulaterespirationandthemetaboliclossofcarbonfromthesemeadows
F I G U R E 6 emspRates(meanplusmnSD)ofnetecosystemproductionfrommeadowsofvariousseagrassspeciescollectedfromtheliteratureOpencircleslsquocrossedrsquocirclesandopentrianglesaregrazedThalassia testudinum(n=7)ungrazedT testudinum(n=6)andHalophila stipulacea(n=3)meadowsrespectivelyfromthepresentstudyFilledcirclesarepreviouslypublishedestimatesofungrazedseagrassmeadownetecosystemproduction(NEP)andcomefromvariouslocationsaroundtheworldCriteriaforliteraturecollectionanddatainclusionaredescribedinJohnsonetal(2017)RatesofNEPfromungrazedT testudinum and H stipulaceameadowsfromthepresentstudyareneartheupperendofpublishedvaluesMedianNEPfromgrazedT testudinumareas(presentstudy)issimilartothemedianNEPofungrazedseagrassmeadowsglobally(Section42)FiguremodifiedfromJohnsonetal(2017)toincludeadditionalestimatesLettersabovepointsdenotesitesfromthepresentstudyndashBBonaireCLittleCaymanEEleutheraFFloridaSStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306

1106emsp |emsp emspenspJournal of Ecology JOHNSON et al
Tomeasureonlymetabolicratesoftheseagrassbenthiccompo-nentsofthesystem(thoseaffectingcarbonstorage)wecorrectedrates measured from incubation chambers for water column me-tabolismWemeasuredwater columnmetabolismusingbiologicaloxygendemand(BOD)bottlesatthesametimeasbenthicchamberincubationsThreeclear300-mlglassBODbottlesandthreeopaque300-mlglassBODbottleswereusedtomeasurewatercolumnpro-ductionandrespirationrespectivelyBottleswerefilledwithwaterat seagrasscanopyheightanchored to thebottomand incubatedunderinsituconditionsintheseagrassmeadowWatercolumnsam-pleswerecollectedatthebeginningofincubationsin60-mlsyringesatcanopyheightandDOconcentrationwasmeasuredinthesamemannerassamplesfromincubationchambersFollowingthe incu-bationperiodbottleswerecollectedand returned to thesurfaceonesamplewascollectedfromeachbottlewithasyringeandDOconcentrationwasmeasured
Hourlymetabolic rateswere calculated from thedifference inDOconcentrationwithinlight(NEP)anddark(RE)incubationcham-bersbetweenthebeginningandendoftheincubationperiodWatercolumn metabolic rates measured from BOD bottles were sub-tractedfromratesmeasuredinchambersHourlygrossprimarypro-duction(GPP)wasthencalculatedasthesumofhourlyNEPandRE DailyratesofGPPwerecalculatedbymultiplyinghourlyGPPbythelengthofthephotoperiodmultipliedby075(toaccountfor lowermetabolicratesatdawnandduskJohnsonetal2017Oliveacuteetal2015)anddailyratesofREwerecalculatedbymultiplyinghourlyRE by24hoursDailyNEPwascalculatedas thedifferencebetweendaily GPP and RE Therewere a few occasionswhen an increaseinDO concentrationwasmeasuredwithin a dark chamber or thedarkBODbottlesduringanincubationsuggestinganerrorThesemeasuredincreasesweresmallusuallywithintheprobesmarginoferror (01mgO2L)andweassumedrespirationtobezeroduringthese timesCorrecting for these respiration errors hadminor ef-fectsondailymetabolic rates in themeadows (TablesS2andS3)MeasuredDOconcentrationswereconvertedfrommgO2tommolO2andthentocarbonunits(mmolC)assumingphotosyntheticandrespiratoryquotientsofone(BarroacutenampDuarte2009)
24emsp|emspData analyses
Net ecosystem productionwas themetabolic variable of primaryinterestasitaccountsforbothcarboncaptureandlossandcanbeusedas an indicatorofwhether a system is currently ametaboliccarbonsinkorsourceThereforeNEPisthemetabolicvariableforwhichstatisticalresultsarepresented
Ouraimwastoevaluatewhethergreenturtlegrazinghasacon-sistenteffectonmetabolicratesacrossseagrassecosystemsmdashthatistoevaluatewhetheratanygivensiteatanygiventimeratesofNEPareloweringrazedareasthaninungrazedareasWeevaluateddifferences in ratesofNEPbetweenareasofgrazedandadjacentungrazedT testudinum from each sitewith paired t tests T testswereusedbecause thecomparisonof interestwas thedifferencebetween grazed and ungrazed areas (effect of grazing) within a
givensitenotdifferencesinNEPamongsitesForsiteswheretheinvasive seagrass H stipulacea was present we used one-wayANOVAstotestforsignificantdifferencesinratesofNEPbetweenH stipulaceameadowsandnearbygrazedandungrazedT testudinum areas(StCroixsiteandLacCaiBeachsiteBonaire)Forthenorth-west site in Lac Bay Bonaire where only a grazed area of T te-studinum and an invasive H stipulacea meadow were comparedanunpaired t testwasused IfANOVA resultswere significant aTukeyspost-hoctestwasusedtoidentifysignificantcomparisonsWetestedfordifferencesinseagrassmeadowparametersbetweengrazedandungrazedT testudinumareasandH stipulaceameadowsatsitesusingpairedttestsandANOVAsinthesamemannerasforNEPWeused linearregressiontotestrelationshipsbetweenNEPand explanatory variables (eg seagrass biomass environmentaltemperature)MeanvaluesforNEPandexplanatoryvariablesfromeachmeadowwereusedforlinearregressionstestssotherewasasingledatapointforeachgrazedandungrazedT testudinum area or H stipulaceameadowateachsamplingsite
AllanalyseswereperformedinRversion343(RCoreTeam2018)Datawere processed using the lsquodplyrrsquo package (Wickham FrancoisHenryampMuumlller2017)andpost-hocanalyseswereperformedusingthelsquoagricolaersquopackage(deMendiburu2017)Alldatamettheassump-tionsfortheseanalysesanddataarepresentedasmeanplusmnSD
3emsp |emspRESULTS
31emsp|emspSeagrass meadow characteristics
Total seagrass shoot densities varied widely among meadowsand locations (Table 2) though shoot densitieswere always high-est inH stipulaceameadows The lowest shoot density observed(2373plusmn446shootsm2)wasinagrazedarea(NWsiteinLacBayBonaire)buttherewasnocleartrendforhigherorlowerseagrassdensities between grazed and ungrazed areas across the sampledsitesThalassia testudinum density in LittleCaymanwasgreater inthe grazed area than ungrazed area (paired t test p = 02) whileT testudinumdensityattheLacCaiBeachsiteinBonairewasgreaterintheungrazedarea(pairedttestp=02)Thalassia testudinum den-sitydidnotdifferbetweengrazedandungrazedareasattheremain-ingsites(pairedttestspgt05)ShootdensityofH stipulaceawasnot significantly different between any of the threeH stipulacea meadowsmeasured(ANOVAF2=009p=92)
HalophilaseagrassestypicallygrowshortbladesCanopyheightoftheH stipulaceameadowwassimilartothatofthenearbygrazedarea inStCroix andwas shorter than thenearbygrazedareasatbothsitesinBonaire(Table2)Halophila stipulaceabladelengthwasgreaterinStCroixthanBonaire(ANOVAF2=2413plt01)AreasofT testudinum-dominatedseagrassmeadowsgrazedbygreentur-tleshad significantly shorterblades and therefore canopyheightthan adjacent ungrazed areas at all sampled sites (paired t testsplt05forallcomparisonsTable2)
Greenturtlesbycroppingseagrassbladesnearthesedimentsur-faceandreducingcanopyheightgreatlyreducetheabove-ground
emspensp emsp | emsp1107Journal of EcologyJOHNSON et al
seagrass biomass of meadows within their grazing areas Above-ground biomass ranged from 48 plusmn 28 to 418 plusmn 139 g DMm2 amonggrazedareas(Table2)andfrom516plusmn110to2022plusmn348gDMm2amongungrazedareasBiomasswasalwaysloweringrazedareas than adjacent ungrazed areas (p lt 05 for all comparisons)Biomass among the three sampledH stipulaceameadows rangedfrom171plusmn59to630plusmn240gDMm2TheH stipulacea meadow in St Croix had significantly greater above-ground biomass thaneither of theH stipulaceameadows sampled inBonaire (ANOVAF2=1202plt 01)Evidenceofgreenturtlegrazingwasonlyob-served in T testudinum-dominated meadows in this study notH stipulaceameadowsandgrazingresultedinagt79decreaseinbiomassingrazedcomparedtoungrazedareasatallsitesexceptStCroix(47Table3)
32emsp|emspEcosystem metabolic rates
Rates of NEP differed among meadow types (grazed ungrazedH stipulacea) across the studied locations (Figures 3 and 4 seeTable S2 for data) Across all sites NEP ranged from 42 plusmn 70 to517plusmn110mmolCmminus2 dayminus1amonggrazedareasandfrom523plusmn57to2253plusmn191mmolCmminus2 dayminus1amongungrazedareasThelargestandarddeviationsonsomeoftheungrazedareaNEPestimatesfromLittleCaymanwerearesultofoneincubationchamberproducingratesmuchdifferentfromtheothertwoAcrossallsitesNEPwasconsist-entlyloweringrazedareasthanungrazedareas(Figure3)DifferencesinNEPbetweengrazedandungrazedareasweresignificantat four
ofthesixsiteswhereungrazedT testudinumwaspresentStCroixLittleCayman (all threemeasurement times) theArvidaBay site inEleutheraandFlorida(pairedttestsplt05forallcomparisons)AttheLacCaiBeachsiteinBonaireandtheHalfSoundsiteinEleuthera
TA B L E 3 emspPercentdifferenceinmetabolicratesandabove-ground(AG)seagrassbiomassbetweengrazedandadjacentungrazedThalassia testudinumareas
Site
GPP RE NEP AG Biomass
difference
Bonaire
LacCaiBeach 818 909 656 793
StCroix
BIRNM 604 783 560 469
LittleCayman
GrapeTreeBay 922 955 891 933
899 796 960 933
885 870 900 933
Eleuthera
ArvidaBay 543 395 629 793
HalfSound 801 889 679 908
Florida
NorthRack 615 413 919 868
Note TheLittleCaymansitewassampledthreetimesin2016Allothersitesweresampledoncein2018SamplingtimesaregiveninSection21AbbreviationsBIRNMBuckIslandReefNationalMonumentsiteGPPgrossprimaryproductionNEPnetecosystemproductionREecosys-temrespiration
F I G U R E 3 emspRates(meanplusmnSD)ofnetecosystemproduction(a)grossprimaryproduction(b)andecosystemrespiration(c)fromgrazed(openpoints)andadjacentungrazed(solidpoints)areasofThalassia testudinumseagrassmeadowsRatesofNetecosystemproduction(NEP)weresignificantlyloweringrazedareasthanungrazedareasatallsitesexceptatLacCaiBeachBonaireandHalfSoundEleuthera(Section32)SitesorderedsouthtonorthBONBonaire(LCBLacCaiBeachsite)STXStCroixLCLittleCaymanELEleuthera(ABArvidaBaysiteHSHalfSoundsite)FLFloridaDashedlinein(a)denotesmetabolicbalance(NEP=0)Valuesabovelinerepresentnetmetaboliccarboncapture[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1108emsp |emsp emspenspJournal of Ecology JOHNSON et al
differencesinNEPbetweengrazedandungrazedareaswerenearlysignificant(pairedttestsp = 08 and p=09respectively)
Though NEP was significantly lower in grazed than ungrazedareas atmost sites rates ofNEPwere almost always positive asecosystem respiration was not stimulated in relation to primaryproductionevenwithconsiderablereductionofphotosyntheticsea-grassbiomassbygrazing(gt79biomassreductionatallsitesexceptStCroix Table3)NEP ranged from56 to96 lower in grazedareasthanungrazedareas(Table3)Asimilarrangeinthedifference
betweengrazedandungrazedareaswasmeasuredforgrossprimaryproduction (54ndash92 lower in grazed) and ecosystem respiration(40ndash96loweringrazed)
Halophila stipulacea meadows were present at three sites andratesofNEPintheseinvasiveseagrassmeadowswerecomparedtonearbynativeT testudinum-dominatedmeadowsNEPintheH stip-ulaceameadowswascomparedtonearbygrazedandungrazedareasofT testudinum-dominatedmeadowsattwoofthesites(StCroixandtheLacCaiBeachsiteinBonaire)WithineachsiteratesofNEPintheH stipulaceameadowwerebetweenthoseofthegrazedandungrazedareas (Figure 4) but the differences were not significant (ANOVApgt10forallpost-hoccomparisonswithineachsite)AnungrazedareawasnotavailableforcomparisonatthethirdsiteatwhichH stipula-ceawaspresent (NWLacBaysite inBonaire)asallT testudinumatthissitewasgrazedbygreenturtlesandratesofNEPdidnotdifferbetweenthe invasive-andnative-dominatedareasatthissiteeither(t test p = 56 Figure 4)Halophila stipulaceameadowNEP rangedfrom612plusmn226to989plusmn233mmolCmminus2 dayminus1(TableS2)anddidnotdiffersignificantlyamongthethreesitesatwhichitwaspresent(ANOVAF2=192p=23)eventhoughtheestablishmentofH stip-ulaceaseagrassmeadowsinStCroixwasrelativelyrecent(AGulickunpublisheddata)comparedtothoseinBonaire(Willetteetal2014)
33emsp|emspDrivers of metabolic rates
VariationinratesofseagrassecosystemmetabolismacrossgrazedandungrazedareasappearedtobedrivenbysomecharacteristicsoftheseagrassmeadowsbutnotbyenvironmentalfactorsMeadowNEPwasstronglypositivelyrelatedtoabove-groundseagrassbio-massacrosssites(linearregressionR2=82plt01Figure5a)NEP
F I G U R E 4 emspRates(meanplusmnSD)ofnetecosystemproductioninmeadowsdominatedbytheinvasiveseagrassHalophila stipulacea (triangles)comparedtonearbygrazed(opencircles)andungrazed(solidcircles)areasofThalassia testudinummeadowsBONBonaire(LCBLacCaiBeachsiteNWnorthwestLacBaysite)STXStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
F I G U R E 5 emspRelationshipbetweennetecosystemproductionandabove-groundseagrassbiomass(a)andtotalseagrassshootdensity(b)fromgrazed(openpoints)andungrazed(solidpoints)areasofThalassia testudinummeadowsSolidlinein(a)isthesignificantlinearregression(R2=82plt01)betweennetecosystemproduction(NEP)andbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows
emspensp emsp | emsp1109Journal of EcologyJOHNSON et al
wasalsopositivelycorrelatedwithseagrassmeadowcanopyheight(R2=84plt01)andbladesurfacearea(R2=92plt01)Above-groundbiomassislikelythemoreusefulpredictorofmeadowNEPhoweverasbiomassisinfluencedbybothcanopyheightandbladesurfaceareaandachange ineitherofthesevariablesresults inachangeinbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows(linearregressionR2lt01p=96Figure5b)norwerethererelationshipsbetweenNEPandenvironmentaltempera-ture(R2=01p=70)irradiance(R2lt01p=10)ormeadowdepth(R2lt01p=77)acrosssites(FigureS1)
4emsp |emspDISCUSSION
Through a comparative study of seagrass meadowsmdashgrazed andungrazedmdashacross a wide area encompassing sites in the GreaterCaribbean and Gulf of Mexico we found that green turtle graz-inghas a consistent effecton themetabolic carbon capture ratesofT testudinum-dominatedseagrassmeadowsRatesofmetaboliccarboncapturewerealwayslowerinareasgrazedbyturtlesthanun-grazedareasofmeadowsmdashupto96inLittleCaymanmdashbutratesofNEPwerealwaysnearorabovemetabolicbalanceEcosystemres-pirationwasnotstimulatedinrelationtoprimaryproductionwithinareas grazed by green turtles regardless of the amount of above-groundseagrassbiomassremovedthroughgrazingorotherdiffer-encesinbioticandabioticfactorsamongsitesThesefindingslendadditionalsupporttothehypothesisthatfutureratesofmetaboliccarboncapturemaybelowerinareasgrazedbyturtlesbutgrazingwillnot stimulateecosystemrespirationand result ina large rem-ineralizationof carbon currently stored in these seagrass habitats(Johnsonetal2017)
The similarity in results between St Croix (awintermeasure-ment)andtheotherfourlocationsinthepresentstudy(allsummermeasurements)strengthensourconclusionthatgrazingresultsinaconsistent response inmetaboliccarboncaptureacrossmeadowsHoweverwewerenotable toaddresspotential temporaldynam-icswithinsitesthatmayaffectnetcarboncaptureoveranannualcycle in these systems Future studies measuring carbon captureoverlongertemporalscales(ieseasonal)maybebeneficialtoourunderstanding of the effects of grazing on within-system carbondynamics
Thoughtheresponse inmetabolic rates tograzingwasconsis-tent acrossmeadows the strengthof this responsevariedamongindividual sites Some sites such as Little Cayman had consider-ablylowerratesofNEPingrazedareasthanungrazedareas(mean917)whereastheresponsetograzing(differenceinNEPbetweengrazedandadjacentungrazedareas)waslesspronouncedatothersites suchasStCroix (Table3) Seagrassmeadowcharacteristicssuch as shoot density canopy height and above-ground biomassare likely to bemore variable among ungrazed areas than grazedareasasgreenturtlegrazingresults insimilarcanopymorphome-tryacrossmeadowsintheGreaterCaribbeanregion(egBjorndal1980HernaacutendezampvanTussenbroek2014Williams1988)Given
the strong relationship between NEP and above-ground biomass(Figure 5a) high variability in NEPwould be expected across un-grazedmeadows(egFigure3a)IndeedtherangeinratesofNEPamongungrazedareaswasnearlythreetimesgreaterthantherangemeasuredacrossgrazedareasinthisstudyAssiteswerespecificallychosen that differed from each other in meadow characteristicsvariationinthestrengthofthemetabolicresponsetograzingwouldbeexpected
Variability in the strength of the metabolic response to graz-ing resulted in the lackofa significantdifferencebetweengrazedandungrazedareasattwositesmdashLacCaiBeachinBonaireandHalfSoundinEleuthera(Figure3a) It ispossiblethatotherfactorsnotaccounted for here such as local environmental factors or greenturtlegrazinghistory(egintensitylongevityofplotuse)mayhaveledtohighervariabilityinmetabolicratesatsomesitesandlackofastatisticaldifferenceInadditionmetabolicratesmayexhibitwithin-meadowvariation(patchiness)similartobelow-groundcarbonstor-age in meadows (OreskaMcglathery amp Porter 2017) leading tohighervariabilityinestimatesandhelpingtoexplainthelackofasig-nificantdifferenceinNEPatthesetwositesThiscouldbedriveninpartbywithin-meadowspatialheterogeneityinabove-groundbio-massgiventhestrongrelationshipbetweenbiomassandNEPHighwithin-meadow variability inmetabolic rates if presentmay alsohelpexplainthe largevariabilitymeasured intheungrazedarea inLittleCaymanFuturestudiesusingtheeddycorrelationtechnique(Bergetal2003) tomeasureecosystemmetabolismmaybeabletoovercomesomeofthiswithin-meadowvariabilityhoweverthegreatexpenseofthismethodmakesitprohibitiveformanystudies
Seagrass ecosystem metabolism is commonly estimated usingoxygendynamicsmeasuredwithbenthicincubationchambers(egBarroacutenetal2004Gaciaetal2005presentstudy)howeverthereare some limitations to thismethodBenthic chambersmay severbelow-ground rhizomesand release carbon that couldaffectmet-abolicestimatesIntactrhizomesontheotherhandcanaffectthediffusiveoxygenlossfromtheseagrasstotherhizosphere(BorumSand-JensenBinzerPedersenampGreve2006)andmayresultinanunderestimateofgrossprimaryproduction(andthereforeNEP)Thismaybeminimizedusingshorter incubation timesas in this studyIncubation length also strongly affects metabolic rates estimatedfromoxygendynamics (Oliveacute et al 2015)with longer incubationtimesbiasingestimatestowardsnetheterotrophyInadditiontoox-ygenmetabolismotherprocessessuchascarbonatedynamics(VanDamLopesOsburnampFourqurean2019)play important roles intotal ecosystem carbondynamicswithout affecting themeasuredproductivityofameadow
CaribbeanT testudinummeadows are highly productive (egJohnsonetal2017KochampMadden2001)andtherelationshipbetween biomass andNEP observed in this study suggests thatmeadows may maintain positive productivity as long as photo-synthetic biomass remains A threshold biomass below whichCaribbean meadows may become net heterotrophic cannot bedirectly addressed here but could be predicted to be near zeroThisisincontrasttoapreviouslysuggestedabove-groundbiomass
1110emsp |emsp emspenspJournal of Ecology JOHNSON et al
threshold(41gDMm2)belowwhichmeadowsmaybecomehet-erotrophic (Duarte et al 2010) However that threshold wascalculated for seagrasses globally and included less productivespecies Additional studies of the relationship between biomassandmeadowNEP from seagrass species and regions other thanthoseinthepresentstudywouldbebeneficialtofurthergeneral-izehowgrazingaffectsmetaboliccarboncaptureacrossseagrassecosystems
Environmental temperature and irradiance are strong driv-ers of seagrassmeadow productivity (Calleja et al 2006 Lee ampDunton 1997 Lee Park amp Kim 2007 Peacuterez amp Romero 1992)howeverpreviousstudiesexaminingcontrolsonseagrassmetabo-lismhavefocusedonungrazedseagrasses(egApostolakiHolmerMarbagraveampKarakassis2010Gaciaetal2005)Meadowmetabolicproductivity correlates stronglywith above-ground seagrass bio-mass(Johnsonetal2017Figure5a)thatcanbegreatlyaffectedby green turtle grazing (Bjorndal 1980CebriaacutenampDuarte 1998Christianenetal2012FourqureanManuelCoatesKenworthyampSmith2010Williams1988)Above-groundbiomassexplained82ofthevariationinNEPacrossgrazedandungrazedT testudi-nummeadowsinthisstudybutNEPwasnotcorrelatedwitheitherenvironmentaltemperatureorirradiance(FigureS1)WethereforeinvestigatedwhetherthelackofanobservedrelationshipbetweenNEPand temperatureand irradiancewasperhapsdue toa largereffectfromdifferencesinbiomasscausedbygrazingmaskinganyeffectoftheseenvironmentalvariablesWescaledmetabolicratestotheabove-groundbiomassineachgrazedandungrazedarea(unitmetabolismperunitbiomass)andcomparedthemtotemperatureandirradianceAccountingfordifferencesinabove-groundbiomassdidnot revealanunderlyingrelationshipbetweenNEPandtheseenvironmentalparametersacrossmeadows(grazedungrazedorallareascombinedFigureS2)ThelackofarelationshipbetweenNEPandenvironmentalparametersinthisstudymaybedueinparttothe relatively limited rangeof temperature and irradiance in sea-grassmeadowsgrazedbygreenturtles in theGreaterCaribbeanAllmeadowsinthisstudywerelocatedinrelativelyshallowwarmtropicalandsubtropicalenvironmentsreceivinghighlevelsofinci-dentsunlight
41emsp|emspEffects of Halophila stipulacea invasion on meadow carbon capture
In addition to the newmetabolic carbon capture estimates wecontribute from grazed areas in T testudinum meadows wealso present the first estimates of NEP from meadows domi-nated by H stipulacea seagrass Having been first reported inthe Caribbean in 2002 (Ruiz amp Ballantine 2004)H stipulacea has spread rapidly among islands and become invasive in thisregion (Christianenet al 2018Willette et al 2014)Halophila stipulaceamayalsospreadmorequicklywithinareasofseagrassgrazedbygreenturtlesthaninungrazedareas(Christianenetal2018) suggesting that grazing may inadvertently facilitate thespreadofthisinvasivespeciesonceithasbecomeestablishedin
anareaInLacBayBonairegreenturtleshavebeguntoexpandtheirgrazingplotsintoareasofpreviouslyungrazedT testudinum asH stipulacea slowly takes over the historically grazed areaswithin thebay (Christianenet al 2018 SmuldersVonk EngelampChristianen2017) If this relationshipbetweengreen turtlesnative seagrasses and invasive seagrass exists in other areas itcouldhaveimplicationsfortotalecosystemcarbondynamicsandcarbonstorageaswellasotherecosystemfunctionswithinthesemeadows
Our resultsdemonstrate that ratesofmetabolic carboncap-turemaybe similarbetweenH stipulaceameadowsand thena-tive T testudinum meadows that they are replacing (Figure 4)HowevergiventhelimitednumberofH stipulaceameadowswewereabletosample inthisstudy(n=3)cautioniswarrantedinextrapolating these findingswidely to other locations facing in-vasionbyH stipulacea seagrassWhile ratesofNEP inH stipu-laceameadowsweresimilar tobothgrazed (n=3)andungrazed(n=2)areasofT testudinum-dominatedmeadowsinStCroixandBonaire (within-site comparisons) multiple factors affect theseratesthatmaydifferandaffectthisrelationshipinotherlocations(egdominantspeciesofnativeseagrassdensityandbiomassofH stipulaceavsthenativespeciesbladeepiphytecoverage)Forexample this relationshipmay differ in areas such as Dominica(eastern Caribbean) where the meadows undergoing H stipu-lacea invasion are dominated byS filiforme seagrass (WilletteampAmbrose 2012) Measurements from additional locations willgreatlyaidinunderstandingthespatialextentofthesimilarityinmetaboliccarboncaptureratesbetweenH stipulaceaandnativeseagrassmeadowsseenhereHoweverthecomparablemetabolicratesbetweentheinvasiveandnativeseagrassesmeasuredinthisstudysuggestsomebenefitsofseagrasspresencemayberetainedinmeadowsfollowinginvasionbyH stipulaceasuchaslocalmit-igation of ocean acidification (through metabolic buffering ofpHCamp et al 2016Hendriks et al 2014Unsworth CollierHendersonampMcKenzie2012)
It is not yet known whether comparable rates of metaboliccarbon capture will translate to comparable carbon sequestra-tionor storage inmeadowsdominatedby invasiveH stipulacea UnlikeT testudinum(vanTussenbroeketal2006)seagrassspe-cieswithinthegenusHalophiladonotformadeepbelow-groundrhizome mat (Fonseca 1989) Robust rhizome mats formed bysomeseagrassspeciescontributetohigherbelow-groundcarbonstorage(Christianenetal2013)andthelackofasimilarrhizomematmayresult in lowersedimentcarbonstorageinH stipulacea meadowsHalophila tissues also decompose quickly in sediment(JosselynFonsecaNiesenampLarson1986)and thismayaffectseagrass-derivedcarboninputtothesedimentsininvadedmead-ows compared to native species with higher refractory organicmatter content (Trevathan-Tackett et al 2017) Further studieson carbondynamics inH stipulaceameadowswill beneeded toformabetterunderstandingofhowthecontinuedinvasionbythisspecieswill affect total ecosystem carbon dynamics in seagrassmeadowsintheCaribbean
emspensp emsp | emsp1111Journal of EcologyJOHNSON et al
42emsp|emspGreater global assessment of grazed meadows needed
UngrazedareasofT testudinum-dominatedseagrassmeadowsfromthisstudyexhibitedratesofNEP(median1040mmolCmminus2 dayminus1)neartheupperendofthosereportedintheliteratureforseagrassecosystemsandgrazedareasinthisstudyexhibitedrates(median381)nearthemedianvalueofNEPforallreportedungrazedsea-grassmeadows(median271Figure6)Measurementsofseagrassmetaboliccarboncapturearerelativelygeographicallylimitedhow-ever with the majority of measurementsmdashmostly from ungrazedT testudinum-dominated meadowsmdashcoming from the GreaterCaribbeanandGulfofMexicoregionsTheCaribbeanandGulfofMexicoareecologicallyimportantgrazingregionsbutonlyasingleestimateofseagrassmetabolismfromagrazedareaexistedpriortothisstudy(Johnsonetal2017)
Allmeadows in the present study exhibited a consistent re-sponseofmetaboliccarboncapturetogreenturtlegrazinghow-ever our studywas limited to the effects of grazing in tropicaland subtropicalT testudinum-dominatedmeadows It ispossiblethatmeadows inother regionsof theworldperhapsdominatedbydifferent seagrass speciesandunderdifferentenvironmentalconditionswillnotexhibitthesameresponsetograzingintermsofmetabolic carbon dynamicsWe are aware of only one otherstudy inwhichthiswas investigated Ina tropicalThalassia hem-prichiimeadow(Tanzania)NEPwasreducedasaresultofexper-imental clipping to simulategrazing (Dahletal2016)howeveronlyhourlyratesofdaytimeNEPweremeasuredWithoutcorre-spondingratesofrespirationwecannotevaluateifNEPremained
positive in thisT hemprichiimeadowoveradiel cycleand if theresponse toexperimental clipping inTanzaniawas similar to theresponse to grazing measured in our study Further studies oneffectsofgrazingonseagrassmeadowmetaboliccarbondynam-icswould be particularly beneficial in (a) additional areaswheregrazer abundance is increasing and (b) meadows dominated byspeciesotherthanT testudinum
5emsp |emspCONCLUSIONS
Given the importance of seagrass meadows as blue carbon eco-systems (Fourqureanetal2012)andtheir role incarbonseques-tration and potential climate changemitigation it is necessary tounderstand how meadow carbon dynamics are affected by graz-ing Our study adds critical information on the effects of grazingon seagrassmetabolic carbondynamics andhow theseprocessesmaybeaffectedwith increasinggreenturtleabundanceandgraz-ingWedemonstratethattheresponseinmetaboliccarboncapturetogreenturtlegrazing isconsistentacrossseagrassecosystems inthe Greater Caribbean and Gulf of Mexico regionsmdashan area thatsupportsvastexpansesofseagrass(GreenampShort2003Jackson1997Wabnitzetal2008)Thisisanecologicallyimportantgrazingregionforgreenturtlepopulationsandgivenincreasinggreenturtleabundanceacrosstheglobe(Chaloupkaetal2008Mazarisetal2017)itisexpectedthatmoreseagrassareawillreturntoanaturalgrazedstateOurresultssuggestthat increasedgrazingwill trans-latetolowerratesofmetaboliccarboncapturebutwillnotstimulaterespirationandthemetaboliclossofcarbonfromthesemeadows
F I G U R E 6 emspRates(meanplusmnSD)ofnetecosystemproductionfrommeadowsofvariousseagrassspeciescollectedfromtheliteratureOpencircleslsquocrossedrsquocirclesandopentrianglesaregrazedThalassia testudinum(n=7)ungrazedT testudinum(n=6)andHalophila stipulacea(n=3)meadowsrespectivelyfromthepresentstudyFilledcirclesarepreviouslypublishedestimatesofungrazedseagrassmeadownetecosystemproduction(NEP)andcomefromvariouslocationsaroundtheworldCriteriaforliteraturecollectionanddatainclusionaredescribedinJohnsonetal(2017)RatesofNEPfromungrazedT testudinum and H stipulaceameadowsfromthepresentstudyareneartheupperendofpublishedvaluesMedianNEPfromgrazedT testudinumareas(presentstudy)issimilartothemedianNEPofungrazedseagrassmeadowsglobally(Section42)FiguremodifiedfromJohnsonetal(2017)toincludeadditionalestimatesLettersabovepointsdenotesitesfromthepresentstudyndashBBonaireCLittleCaymanEEleutheraFFloridaSStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306
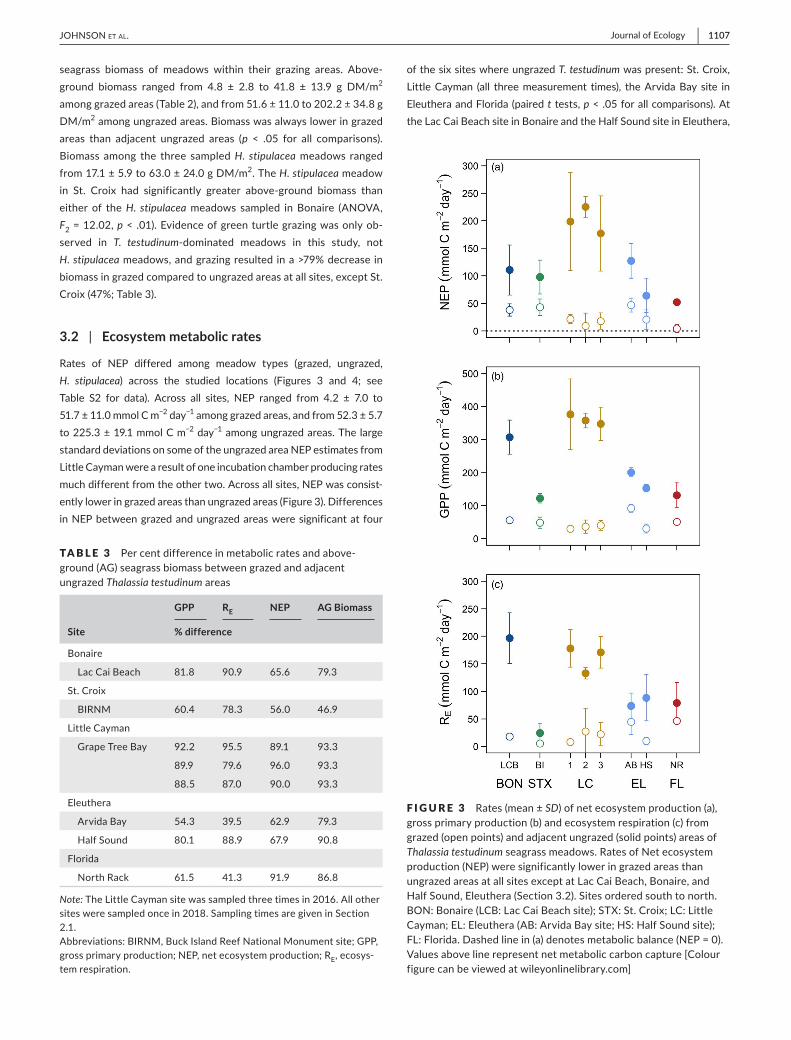
emspensp emsp | emsp1107Journal of EcologyJOHNSON et al
seagrass biomass of meadows within their grazing areas Above-ground biomass ranged from 48 plusmn 28 to 418 plusmn 139 g DMm2 amonggrazedareas(Table2)andfrom516plusmn110to2022plusmn348gDMm2amongungrazedareasBiomasswasalwaysloweringrazedareas than adjacent ungrazed areas (p lt 05 for all comparisons)Biomass among the three sampledH stipulaceameadows rangedfrom171plusmn59to630plusmn240gDMm2TheH stipulacea meadow in St Croix had significantly greater above-ground biomass thaneither of theH stipulaceameadows sampled inBonaire (ANOVAF2=1202plt 01)Evidenceofgreenturtlegrazingwasonlyob-served in T testudinum-dominated meadows in this study notH stipulaceameadowsandgrazingresultedinagt79decreaseinbiomassingrazedcomparedtoungrazedareasatallsitesexceptStCroix(47Table3)
32emsp|emspEcosystem metabolic rates
Rates of NEP differed among meadow types (grazed ungrazedH stipulacea) across the studied locations (Figures 3 and 4 seeTable S2 for data) Across all sites NEP ranged from 42 plusmn 70 to517plusmn110mmolCmminus2 dayminus1amonggrazedareasandfrom523plusmn57to2253plusmn191mmolCmminus2 dayminus1amongungrazedareasThelargestandarddeviationsonsomeoftheungrazedareaNEPestimatesfromLittleCaymanwerearesultofoneincubationchamberproducingratesmuchdifferentfromtheothertwoAcrossallsitesNEPwasconsist-entlyloweringrazedareasthanungrazedareas(Figure3)DifferencesinNEPbetweengrazedandungrazedareasweresignificantat four
ofthesixsiteswhereungrazedT testudinumwaspresentStCroixLittleCayman (all threemeasurement times) theArvidaBay site inEleutheraandFlorida(pairedttestsplt05forallcomparisons)AttheLacCaiBeachsiteinBonaireandtheHalfSoundsiteinEleuthera
TA B L E 3 emspPercentdifferenceinmetabolicratesandabove-ground(AG)seagrassbiomassbetweengrazedandadjacentungrazedThalassia testudinumareas
Site
GPP RE NEP AG Biomass
difference
Bonaire
LacCaiBeach 818 909 656 793
StCroix
BIRNM 604 783 560 469
LittleCayman
GrapeTreeBay 922 955 891 933
899 796 960 933
885 870 900 933
Eleuthera
ArvidaBay 543 395 629 793
HalfSound 801 889 679 908
Florida
NorthRack 615 413 919 868
Note TheLittleCaymansitewassampledthreetimesin2016Allothersitesweresampledoncein2018SamplingtimesaregiveninSection21AbbreviationsBIRNMBuckIslandReefNationalMonumentsiteGPPgrossprimaryproductionNEPnetecosystemproductionREecosys-temrespiration
F I G U R E 3 emspRates(meanplusmnSD)ofnetecosystemproduction(a)grossprimaryproduction(b)andecosystemrespiration(c)fromgrazed(openpoints)andadjacentungrazed(solidpoints)areasofThalassia testudinumseagrassmeadowsRatesofNetecosystemproduction(NEP)weresignificantlyloweringrazedareasthanungrazedareasatallsitesexceptatLacCaiBeachBonaireandHalfSoundEleuthera(Section32)SitesorderedsouthtonorthBONBonaire(LCBLacCaiBeachsite)STXStCroixLCLittleCaymanELEleuthera(ABArvidaBaysiteHSHalfSoundsite)FLFloridaDashedlinein(a)denotesmetabolicbalance(NEP=0)Valuesabovelinerepresentnetmetaboliccarboncapture[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1108emsp |emsp emspenspJournal of Ecology JOHNSON et al
differencesinNEPbetweengrazedandungrazedareaswerenearlysignificant(pairedttestsp = 08 and p=09respectively)
Though NEP was significantly lower in grazed than ungrazedareas atmost sites rates ofNEPwere almost always positive asecosystem respiration was not stimulated in relation to primaryproductionevenwithconsiderablereductionofphotosyntheticsea-grassbiomassbygrazing(gt79biomassreductionatallsitesexceptStCroix Table3)NEP ranged from56 to96 lower in grazedareasthanungrazedareas(Table3)Asimilarrangeinthedifference
betweengrazedandungrazedareaswasmeasuredforgrossprimaryproduction (54ndash92 lower in grazed) and ecosystem respiration(40ndash96loweringrazed)
Halophila stipulacea meadows were present at three sites andratesofNEPintheseinvasiveseagrassmeadowswerecomparedtonearbynativeT testudinum-dominatedmeadowsNEPintheH stip-ulaceameadowswascomparedtonearbygrazedandungrazedareasofT testudinum-dominatedmeadowsattwoofthesites(StCroixandtheLacCaiBeachsiteinBonaire)WithineachsiteratesofNEPintheH stipulaceameadowwerebetweenthoseofthegrazedandungrazedareas (Figure 4) but the differences were not significant (ANOVApgt10forallpost-hoccomparisonswithineachsite)AnungrazedareawasnotavailableforcomparisonatthethirdsiteatwhichH stipula-ceawaspresent (NWLacBaysite inBonaire)asallT testudinumatthissitewasgrazedbygreenturtlesandratesofNEPdidnotdifferbetweenthe invasive-andnative-dominatedareasatthissiteeither(t test p = 56 Figure 4)Halophila stipulaceameadowNEP rangedfrom612plusmn226to989plusmn233mmolCmminus2 dayminus1(TableS2)anddidnotdiffersignificantlyamongthethreesitesatwhichitwaspresent(ANOVAF2=192p=23)eventhoughtheestablishmentofH stip-ulaceaseagrassmeadowsinStCroixwasrelativelyrecent(AGulickunpublisheddata)comparedtothoseinBonaire(Willetteetal2014)
33emsp|emspDrivers of metabolic rates
VariationinratesofseagrassecosystemmetabolismacrossgrazedandungrazedareasappearedtobedrivenbysomecharacteristicsoftheseagrassmeadowsbutnotbyenvironmentalfactorsMeadowNEPwasstronglypositivelyrelatedtoabove-groundseagrassbio-massacrosssites(linearregressionR2=82plt01Figure5a)NEP
F I G U R E 4 emspRates(meanplusmnSD)ofnetecosystemproductioninmeadowsdominatedbytheinvasiveseagrassHalophila stipulacea (triangles)comparedtonearbygrazed(opencircles)andungrazed(solidcircles)areasofThalassia testudinummeadowsBONBonaire(LCBLacCaiBeachsiteNWnorthwestLacBaysite)STXStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
F I G U R E 5 emspRelationshipbetweennetecosystemproductionandabove-groundseagrassbiomass(a)andtotalseagrassshootdensity(b)fromgrazed(openpoints)andungrazed(solidpoints)areasofThalassia testudinummeadowsSolidlinein(a)isthesignificantlinearregression(R2=82plt01)betweennetecosystemproduction(NEP)andbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows
emspensp emsp | emsp1109Journal of EcologyJOHNSON et al
wasalsopositivelycorrelatedwithseagrassmeadowcanopyheight(R2=84plt01)andbladesurfacearea(R2=92plt01)Above-groundbiomassislikelythemoreusefulpredictorofmeadowNEPhoweverasbiomassisinfluencedbybothcanopyheightandbladesurfaceareaandachange ineitherofthesevariablesresults inachangeinbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows(linearregressionR2lt01p=96Figure5b)norwerethererelationshipsbetweenNEPandenvironmentaltempera-ture(R2=01p=70)irradiance(R2lt01p=10)ormeadowdepth(R2lt01p=77)acrosssites(FigureS1)
4emsp |emspDISCUSSION
Through a comparative study of seagrass meadowsmdashgrazed andungrazedmdashacross a wide area encompassing sites in the GreaterCaribbean and Gulf of Mexico we found that green turtle graz-inghas a consistent effecton themetabolic carbon capture ratesofT testudinum-dominatedseagrassmeadowsRatesofmetaboliccarboncapturewerealwayslowerinareasgrazedbyturtlesthanun-grazedareasofmeadowsmdashupto96inLittleCaymanmdashbutratesofNEPwerealwaysnearorabovemetabolicbalanceEcosystemres-pirationwasnotstimulatedinrelationtoprimaryproductionwithinareas grazed by green turtles regardless of the amount of above-groundseagrassbiomassremovedthroughgrazingorotherdiffer-encesinbioticandabioticfactorsamongsitesThesefindingslendadditionalsupporttothehypothesisthatfutureratesofmetaboliccarboncapturemaybelowerinareasgrazedbyturtlesbutgrazingwillnot stimulateecosystemrespirationand result ina large rem-ineralizationof carbon currently stored in these seagrass habitats(Johnsonetal2017)
The similarity in results between St Croix (awintermeasure-ment)andtheotherfourlocationsinthepresentstudy(allsummermeasurements)strengthensourconclusionthatgrazingresultsinaconsistent response inmetaboliccarboncaptureacrossmeadowsHoweverwewerenotable toaddresspotential temporaldynam-icswithinsitesthatmayaffectnetcarboncaptureoveranannualcycle in these systems Future studies measuring carbon captureoverlongertemporalscales(ieseasonal)maybebeneficialtoourunderstanding of the effects of grazing on within-system carbondynamics
Thoughtheresponse inmetabolic rates tograzingwasconsis-tent acrossmeadows the strengthof this responsevariedamongindividual sites Some sites such as Little Cayman had consider-ablylowerratesofNEPingrazedareasthanungrazedareas(mean917)whereastheresponsetograzing(differenceinNEPbetweengrazedandadjacentungrazedareas)waslesspronouncedatothersites suchasStCroix (Table3) Seagrassmeadowcharacteristicssuch as shoot density canopy height and above-ground biomassare likely to bemore variable among ungrazed areas than grazedareasasgreenturtlegrazingresults insimilarcanopymorphome-tryacrossmeadowsintheGreaterCaribbeanregion(egBjorndal1980HernaacutendezampvanTussenbroek2014Williams1988)Given
the strong relationship between NEP and above-ground biomass(Figure 5a) high variability in NEPwould be expected across un-grazedmeadows(egFigure3a)IndeedtherangeinratesofNEPamongungrazedareaswasnearlythreetimesgreaterthantherangemeasuredacrossgrazedareasinthisstudyAssiteswerespecificallychosen that differed from each other in meadow characteristicsvariationinthestrengthofthemetabolicresponsetograzingwouldbeexpected
Variability in the strength of the metabolic response to graz-ing resulted in the lackofa significantdifferencebetweengrazedandungrazedareasattwositesmdashLacCaiBeachinBonaireandHalfSoundinEleuthera(Figure3a) It ispossiblethatotherfactorsnotaccounted for here such as local environmental factors or greenturtlegrazinghistory(egintensitylongevityofplotuse)mayhaveledtohighervariabilityinmetabolicratesatsomesitesandlackofastatisticaldifferenceInadditionmetabolicratesmayexhibitwithin-meadowvariation(patchiness)similartobelow-groundcarbonstor-age in meadows (OreskaMcglathery amp Porter 2017) leading tohighervariabilityinestimatesandhelpingtoexplainthelackofasig-nificantdifferenceinNEPatthesetwositesThiscouldbedriveninpartbywithin-meadowspatialheterogeneityinabove-groundbio-massgiventhestrongrelationshipbetweenbiomassandNEPHighwithin-meadow variability inmetabolic rates if presentmay alsohelpexplainthe largevariabilitymeasured intheungrazedarea inLittleCaymanFuturestudiesusingtheeddycorrelationtechnique(Bergetal2003) tomeasureecosystemmetabolismmaybeabletoovercomesomeofthiswithin-meadowvariabilityhoweverthegreatexpenseofthismethodmakesitprohibitiveformanystudies
Seagrass ecosystem metabolism is commonly estimated usingoxygendynamicsmeasuredwithbenthicincubationchambers(egBarroacutenetal2004Gaciaetal2005presentstudy)howeverthereare some limitations to thismethodBenthic chambersmay severbelow-ground rhizomesand release carbon that couldaffectmet-abolicestimatesIntactrhizomesontheotherhandcanaffectthediffusiveoxygenlossfromtheseagrasstotherhizosphere(BorumSand-JensenBinzerPedersenampGreve2006)andmayresultinanunderestimateofgrossprimaryproduction(andthereforeNEP)Thismaybeminimizedusingshorter incubation timesas in this studyIncubation length also strongly affects metabolic rates estimatedfromoxygendynamics (Oliveacute et al 2015)with longer incubationtimesbiasingestimatestowardsnetheterotrophyInadditiontoox-ygenmetabolismotherprocessessuchascarbonatedynamics(VanDamLopesOsburnampFourqurean2019)play important roles intotal ecosystem carbondynamicswithout affecting themeasuredproductivityofameadow
CaribbeanT testudinummeadows are highly productive (egJohnsonetal2017KochampMadden2001)andtherelationshipbetween biomass andNEP observed in this study suggests thatmeadows may maintain positive productivity as long as photo-synthetic biomass remains A threshold biomass below whichCaribbean meadows may become net heterotrophic cannot bedirectly addressed here but could be predicted to be near zeroThisisincontrasttoapreviouslysuggestedabove-groundbiomass
1110emsp |emsp emspenspJournal of Ecology JOHNSON et al
threshold(41gDMm2)belowwhichmeadowsmaybecomehet-erotrophic (Duarte et al 2010) However that threshold wascalculated for seagrasses globally and included less productivespecies Additional studies of the relationship between biomassandmeadowNEP from seagrass species and regions other thanthoseinthepresentstudywouldbebeneficialtofurthergeneral-izehowgrazingaffectsmetaboliccarboncaptureacrossseagrassecosystems
Environmental temperature and irradiance are strong driv-ers of seagrassmeadow productivity (Calleja et al 2006 Lee ampDunton 1997 Lee Park amp Kim 2007 Peacuterez amp Romero 1992)howeverpreviousstudiesexaminingcontrolsonseagrassmetabo-lismhavefocusedonungrazedseagrasses(egApostolakiHolmerMarbagraveampKarakassis2010Gaciaetal2005)Meadowmetabolicproductivity correlates stronglywith above-ground seagrass bio-mass(Johnsonetal2017Figure5a)thatcanbegreatlyaffectedby green turtle grazing (Bjorndal 1980CebriaacutenampDuarte 1998Christianenetal2012FourqureanManuelCoatesKenworthyampSmith2010Williams1988)Above-groundbiomassexplained82ofthevariationinNEPacrossgrazedandungrazedT testudi-nummeadowsinthisstudybutNEPwasnotcorrelatedwitheitherenvironmentaltemperatureorirradiance(FigureS1)WethereforeinvestigatedwhetherthelackofanobservedrelationshipbetweenNEPand temperatureand irradiancewasperhapsdue toa largereffectfromdifferencesinbiomasscausedbygrazingmaskinganyeffectoftheseenvironmentalvariablesWescaledmetabolicratestotheabove-groundbiomassineachgrazedandungrazedarea(unitmetabolismperunitbiomass)andcomparedthemtotemperatureandirradianceAccountingfordifferencesinabove-groundbiomassdidnot revealanunderlyingrelationshipbetweenNEPandtheseenvironmentalparametersacrossmeadows(grazedungrazedorallareascombinedFigureS2)ThelackofarelationshipbetweenNEPandenvironmentalparametersinthisstudymaybedueinparttothe relatively limited rangeof temperature and irradiance in sea-grassmeadowsgrazedbygreenturtles in theGreaterCaribbeanAllmeadowsinthisstudywerelocatedinrelativelyshallowwarmtropicalandsubtropicalenvironmentsreceivinghighlevelsofinci-dentsunlight
41emsp|emspEffects of Halophila stipulacea invasion on meadow carbon capture
In addition to the newmetabolic carbon capture estimates wecontribute from grazed areas in T testudinum meadows wealso present the first estimates of NEP from meadows domi-nated by H stipulacea seagrass Having been first reported inthe Caribbean in 2002 (Ruiz amp Ballantine 2004)H stipulacea has spread rapidly among islands and become invasive in thisregion (Christianenet al 2018Willette et al 2014)Halophila stipulaceamayalsospreadmorequicklywithinareasofseagrassgrazedbygreenturtlesthaninungrazedareas(Christianenetal2018) suggesting that grazing may inadvertently facilitate thespreadofthisinvasivespeciesonceithasbecomeestablishedin
anareaInLacBayBonairegreenturtleshavebeguntoexpandtheirgrazingplotsintoareasofpreviouslyungrazedT testudinum asH stipulacea slowly takes over the historically grazed areaswithin thebay (Christianenet al 2018 SmuldersVonk EngelampChristianen2017) If this relationshipbetweengreen turtlesnative seagrasses and invasive seagrass exists in other areas itcouldhaveimplicationsfortotalecosystemcarbondynamicsandcarbonstorageaswellasotherecosystemfunctionswithinthesemeadows
Our resultsdemonstrate that ratesofmetabolic carboncap-turemaybe similarbetweenH stipulaceameadowsand thena-tive T testudinum meadows that they are replacing (Figure 4)HowevergiventhelimitednumberofH stipulaceameadowswewereabletosample inthisstudy(n=3)cautioniswarrantedinextrapolating these findingswidely to other locations facing in-vasionbyH stipulacea seagrassWhile ratesofNEP inH stipu-laceameadowsweresimilar tobothgrazed (n=3)andungrazed(n=2)areasofT testudinum-dominatedmeadowsinStCroixandBonaire (within-site comparisons) multiple factors affect theseratesthatmaydifferandaffectthisrelationshipinotherlocations(egdominantspeciesofnativeseagrassdensityandbiomassofH stipulaceavsthenativespeciesbladeepiphytecoverage)Forexample this relationshipmay differ in areas such as Dominica(eastern Caribbean) where the meadows undergoing H stipu-lacea invasion are dominated byS filiforme seagrass (WilletteampAmbrose 2012) Measurements from additional locations willgreatlyaidinunderstandingthespatialextentofthesimilarityinmetaboliccarboncaptureratesbetweenH stipulaceaandnativeseagrassmeadowsseenhereHoweverthecomparablemetabolicratesbetweentheinvasiveandnativeseagrassesmeasuredinthisstudysuggestsomebenefitsofseagrasspresencemayberetainedinmeadowsfollowinginvasionbyH stipulaceasuchaslocalmit-igation of ocean acidification (through metabolic buffering ofpHCamp et al 2016Hendriks et al 2014Unsworth CollierHendersonampMcKenzie2012)
It is not yet known whether comparable rates of metaboliccarbon capture will translate to comparable carbon sequestra-tionor storage inmeadowsdominatedby invasiveH stipulacea UnlikeT testudinum(vanTussenbroeketal2006)seagrassspe-cieswithinthegenusHalophiladonotformadeepbelow-groundrhizome mat (Fonseca 1989) Robust rhizome mats formed bysomeseagrassspeciescontributetohigherbelow-groundcarbonstorage(Christianenetal2013)andthelackofasimilarrhizomematmayresult in lowersedimentcarbonstorageinH stipulacea meadowsHalophila tissues also decompose quickly in sediment(JosselynFonsecaNiesenampLarson1986)and thismayaffectseagrass-derivedcarboninputtothesedimentsininvadedmead-ows compared to native species with higher refractory organicmatter content (Trevathan-Tackett et al 2017) Further studieson carbondynamics inH stipulaceameadowswill beneeded toformabetterunderstandingofhowthecontinuedinvasionbythisspecieswill affect total ecosystem carbon dynamics in seagrassmeadowsintheCaribbean
emspensp emsp | emsp1111Journal of EcologyJOHNSON et al
42emsp|emspGreater global assessment of grazed meadows needed
UngrazedareasofT testudinum-dominatedseagrassmeadowsfromthisstudyexhibitedratesofNEP(median1040mmolCmminus2 dayminus1)neartheupperendofthosereportedintheliteratureforseagrassecosystemsandgrazedareasinthisstudyexhibitedrates(median381)nearthemedianvalueofNEPforallreportedungrazedsea-grassmeadows(median271Figure6)Measurementsofseagrassmetaboliccarboncapturearerelativelygeographicallylimitedhow-ever with the majority of measurementsmdashmostly from ungrazedT testudinum-dominated meadowsmdashcoming from the GreaterCaribbeanandGulfofMexicoregionsTheCaribbeanandGulfofMexicoareecologicallyimportantgrazingregionsbutonlyasingleestimateofseagrassmetabolismfromagrazedareaexistedpriortothisstudy(Johnsonetal2017)
Allmeadows in the present study exhibited a consistent re-sponseofmetaboliccarboncapturetogreenturtlegrazinghow-ever our studywas limited to the effects of grazing in tropicaland subtropicalT testudinum-dominatedmeadows It ispossiblethatmeadows inother regionsof theworldperhapsdominatedbydifferent seagrass speciesandunderdifferentenvironmentalconditionswillnotexhibitthesameresponsetograzingintermsofmetabolic carbon dynamicsWe are aware of only one otherstudy inwhichthiswas investigated Ina tropicalThalassia hem-prichiimeadow(Tanzania)NEPwasreducedasaresultofexper-imental clipping to simulategrazing (Dahletal2016)howeveronlyhourlyratesofdaytimeNEPweremeasuredWithoutcorre-spondingratesofrespirationwecannotevaluateifNEPremained
positive in thisT hemprichiimeadowoveradiel cycleand if theresponse toexperimental clipping inTanzaniawas similar to theresponse to grazing measured in our study Further studies oneffectsofgrazingonseagrassmeadowmetaboliccarbondynam-icswould be particularly beneficial in (a) additional areaswheregrazer abundance is increasing and (b) meadows dominated byspeciesotherthanT testudinum
5emsp |emspCONCLUSIONS
Given the importance of seagrass meadows as blue carbon eco-systems (Fourqureanetal2012)andtheir role incarbonseques-tration and potential climate changemitigation it is necessary tounderstand how meadow carbon dynamics are affected by graz-ing Our study adds critical information on the effects of grazingon seagrassmetabolic carbondynamics andhow theseprocessesmaybeaffectedwith increasinggreenturtleabundanceandgraz-ingWedemonstratethattheresponseinmetaboliccarboncapturetogreenturtlegrazing isconsistentacrossseagrassecosystems inthe Greater Caribbean and Gulf of Mexico regionsmdashan area thatsupportsvastexpansesofseagrass(GreenampShort2003Jackson1997Wabnitzetal2008)Thisisanecologicallyimportantgrazingregionforgreenturtlepopulationsandgivenincreasinggreenturtleabundanceacrosstheglobe(Chaloupkaetal2008Mazarisetal2017)itisexpectedthatmoreseagrassareawillreturntoanaturalgrazedstateOurresultssuggestthat increasedgrazingwill trans-latetolowerratesofmetaboliccarboncapturebutwillnotstimulaterespirationandthemetaboliclossofcarbonfromthesemeadows
F I G U R E 6 emspRates(meanplusmnSD)ofnetecosystemproductionfrommeadowsofvariousseagrassspeciescollectedfromtheliteratureOpencircleslsquocrossedrsquocirclesandopentrianglesaregrazedThalassia testudinum(n=7)ungrazedT testudinum(n=6)andHalophila stipulacea(n=3)meadowsrespectivelyfromthepresentstudyFilledcirclesarepreviouslypublishedestimatesofungrazedseagrassmeadownetecosystemproduction(NEP)andcomefromvariouslocationsaroundtheworldCriteriaforliteraturecollectionanddatainclusionaredescribedinJohnsonetal(2017)RatesofNEPfromungrazedT testudinum and H stipulaceameadowsfromthepresentstudyareneartheupperendofpublishedvaluesMedianNEPfromgrazedT testudinumareas(presentstudy)issimilartothemedianNEPofungrazedseagrassmeadowsglobally(Section42)FiguremodifiedfromJohnsonetal(2017)toincludeadditionalestimatesLettersabovepointsdenotesitesfromthepresentstudyndashBBonaireCLittleCaymanEEleutheraFFloridaSStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306

1108emsp |emsp emspenspJournal of Ecology JOHNSON et al
differencesinNEPbetweengrazedandungrazedareaswerenearlysignificant(pairedttestsp = 08 and p=09respectively)
Though NEP was significantly lower in grazed than ungrazedareas atmost sites rates ofNEPwere almost always positive asecosystem respiration was not stimulated in relation to primaryproductionevenwithconsiderablereductionofphotosyntheticsea-grassbiomassbygrazing(gt79biomassreductionatallsitesexceptStCroix Table3)NEP ranged from56 to96 lower in grazedareasthanungrazedareas(Table3)Asimilarrangeinthedifference
betweengrazedandungrazedareaswasmeasuredforgrossprimaryproduction (54ndash92 lower in grazed) and ecosystem respiration(40ndash96loweringrazed)
Halophila stipulacea meadows were present at three sites andratesofNEPintheseinvasiveseagrassmeadowswerecomparedtonearbynativeT testudinum-dominatedmeadowsNEPintheH stip-ulaceameadowswascomparedtonearbygrazedandungrazedareasofT testudinum-dominatedmeadowsattwoofthesites(StCroixandtheLacCaiBeachsiteinBonaire)WithineachsiteratesofNEPintheH stipulaceameadowwerebetweenthoseofthegrazedandungrazedareas (Figure 4) but the differences were not significant (ANOVApgt10forallpost-hoccomparisonswithineachsite)AnungrazedareawasnotavailableforcomparisonatthethirdsiteatwhichH stipula-ceawaspresent (NWLacBaysite inBonaire)asallT testudinumatthissitewasgrazedbygreenturtlesandratesofNEPdidnotdifferbetweenthe invasive-andnative-dominatedareasatthissiteeither(t test p = 56 Figure 4)Halophila stipulaceameadowNEP rangedfrom612plusmn226to989plusmn233mmolCmminus2 dayminus1(TableS2)anddidnotdiffersignificantlyamongthethreesitesatwhichitwaspresent(ANOVAF2=192p=23)eventhoughtheestablishmentofH stip-ulaceaseagrassmeadowsinStCroixwasrelativelyrecent(AGulickunpublisheddata)comparedtothoseinBonaire(Willetteetal2014)
33emsp|emspDrivers of metabolic rates
VariationinratesofseagrassecosystemmetabolismacrossgrazedandungrazedareasappearedtobedrivenbysomecharacteristicsoftheseagrassmeadowsbutnotbyenvironmentalfactorsMeadowNEPwasstronglypositivelyrelatedtoabove-groundseagrassbio-massacrosssites(linearregressionR2=82plt01Figure5a)NEP
F I G U R E 4 emspRates(meanplusmnSD)ofnetecosystemproductioninmeadowsdominatedbytheinvasiveseagrassHalophila stipulacea (triangles)comparedtonearbygrazed(opencircles)andungrazed(solidcircles)areasofThalassia testudinummeadowsBONBonaire(LCBLacCaiBeachsiteNWnorthwestLacBaysite)STXStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
F I G U R E 5 emspRelationshipbetweennetecosystemproductionandabove-groundseagrassbiomass(a)andtotalseagrassshootdensity(b)fromgrazed(openpoints)andungrazed(solidpoints)areasofThalassia testudinummeadowsSolidlinein(a)isthesignificantlinearregression(R2=82plt01)betweennetecosystemproduction(NEP)andbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows
emspensp emsp | emsp1109Journal of EcologyJOHNSON et al
wasalsopositivelycorrelatedwithseagrassmeadowcanopyheight(R2=84plt01)andbladesurfacearea(R2=92plt01)Above-groundbiomassislikelythemoreusefulpredictorofmeadowNEPhoweverasbiomassisinfluencedbybothcanopyheightandbladesurfaceareaandachange ineitherofthesevariablesresults inachangeinbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows(linearregressionR2lt01p=96Figure5b)norwerethererelationshipsbetweenNEPandenvironmentaltempera-ture(R2=01p=70)irradiance(R2lt01p=10)ormeadowdepth(R2lt01p=77)acrosssites(FigureS1)
4emsp |emspDISCUSSION
Through a comparative study of seagrass meadowsmdashgrazed andungrazedmdashacross a wide area encompassing sites in the GreaterCaribbean and Gulf of Mexico we found that green turtle graz-inghas a consistent effecton themetabolic carbon capture ratesofT testudinum-dominatedseagrassmeadowsRatesofmetaboliccarboncapturewerealwayslowerinareasgrazedbyturtlesthanun-grazedareasofmeadowsmdashupto96inLittleCaymanmdashbutratesofNEPwerealwaysnearorabovemetabolicbalanceEcosystemres-pirationwasnotstimulatedinrelationtoprimaryproductionwithinareas grazed by green turtles regardless of the amount of above-groundseagrassbiomassremovedthroughgrazingorotherdiffer-encesinbioticandabioticfactorsamongsitesThesefindingslendadditionalsupporttothehypothesisthatfutureratesofmetaboliccarboncapturemaybelowerinareasgrazedbyturtlesbutgrazingwillnot stimulateecosystemrespirationand result ina large rem-ineralizationof carbon currently stored in these seagrass habitats(Johnsonetal2017)
The similarity in results between St Croix (awintermeasure-ment)andtheotherfourlocationsinthepresentstudy(allsummermeasurements)strengthensourconclusionthatgrazingresultsinaconsistent response inmetaboliccarboncaptureacrossmeadowsHoweverwewerenotable toaddresspotential temporaldynam-icswithinsitesthatmayaffectnetcarboncaptureoveranannualcycle in these systems Future studies measuring carbon captureoverlongertemporalscales(ieseasonal)maybebeneficialtoourunderstanding of the effects of grazing on within-system carbondynamics
Thoughtheresponse inmetabolic rates tograzingwasconsis-tent acrossmeadows the strengthof this responsevariedamongindividual sites Some sites such as Little Cayman had consider-ablylowerratesofNEPingrazedareasthanungrazedareas(mean917)whereastheresponsetograzing(differenceinNEPbetweengrazedandadjacentungrazedareas)waslesspronouncedatothersites suchasStCroix (Table3) Seagrassmeadowcharacteristicssuch as shoot density canopy height and above-ground biomassare likely to bemore variable among ungrazed areas than grazedareasasgreenturtlegrazingresults insimilarcanopymorphome-tryacrossmeadowsintheGreaterCaribbeanregion(egBjorndal1980HernaacutendezampvanTussenbroek2014Williams1988)Given
the strong relationship between NEP and above-ground biomass(Figure 5a) high variability in NEPwould be expected across un-grazedmeadows(egFigure3a)IndeedtherangeinratesofNEPamongungrazedareaswasnearlythreetimesgreaterthantherangemeasuredacrossgrazedareasinthisstudyAssiteswerespecificallychosen that differed from each other in meadow characteristicsvariationinthestrengthofthemetabolicresponsetograzingwouldbeexpected
Variability in the strength of the metabolic response to graz-ing resulted in the lackofa significantdifferencebetweengrazedandungrazedareasattwositesmdashLacCaiBeachinBonaireandHalfSoundinEleuthera(Figure3a) It ispossiblethatotherfactorsnotaccounted for here such as local environmental factors or greenturtlegrazinghistory(egintensitylongevityofplotuse)mayhaveledtohighervariabilityinmetabolicratesatsomesitesandlackofastatisticaldifferenceInadditionmetabolicratesmayexhibitwithin-meadowvariation(patchiness)similartobelow-groundcarbonstor-age in meadows (OreskaMcglathery amp Porter 2017) leading tohighervariabilityinestimatesandhelpingtoexplainthelackofasig-nificantdifferenceinNEPatthesetwositesThiscouldbedriveninpartbywithin-meadowspatialheterogeneityinabove-groundbio-massgiventhestrongrelationshipbetweenbiomassandNEPHighwithin-meadow variability inmetabolic rates if presentmay alsohelpexplainthe largevariabilitymeasured intheungrazedarea inLittleCaymanFuturestudiesusingtheeddycorrelationtechnique(Bergetal2003) tomeasureecosystemmetabolismmaybeabletoovercomesomeofthiswithin-meadowvariabilityhoweverthegreatexpenseofthismethodmakesitprohibitiveformanystudies
Seagrass ecosystem metabolism is commonly estimated usingoxygendynamicsmeasuredwithbenthicincubationchambers(egBarroacutenetal2004Gaciaetal2005presentstudy)howeverthereare some limitations to thismethodBenthic chambersmay severbelow-ground rhizomesand release carbon that couldaffectmet-abolicestimatesIntactrhizomesontheotherhandcanaffectthediffusiveoxygenlossfromtheseagrasstotherhizosphere(BorumSand-JensenBinzerPedersenampGreve2006)andmayresultinanunderestimateofgrossprimaryproduction(andthereforeNEP)Thismaybeminimizedusingshorter incubation timesas in this studyIncubation length also strongly affects metabolic rates estimatedfromoxygendynamics (Oliveacute et al 2015)with longer incubationtimesbiasingestimatestowardsnetheterotrophyInadditiontoox-ygenmetabolismotherprocessessuchascarbonatedynamics(VanDamLopesOsburnampFourqurean2019)play important roles intotal ecosystem carbondynamicswithout affecting themeasuredproductivityofameadow
CaribbeanT testudinummeadows are highly productive (egJohnsonetal2017KochampMadden2001)andtherelationshipbetween biomass andNEP observed in this study suggests thatmeadows may maintain positive productivity as long as photo-synthetic biomass remains A threshold biomass below whichCaribbean meadows may become net heterotrophic cannot bedirectly addressed here but could be predicted to be near zeroThisisincontrasttoapreviouslysuggestedabove-groundbiomass
1110emsp |emsp emspenspJournal of Ecology JOHNSON et al
threshold(41gDMm2)belowwhichmeadowsmaybecomehet-erotrophic (Duarte et al 2010) However that threshold wascalculated for seagrasses globally and included less productivespecies Additional studies of the relationship between biomassandmeadowNEP from seagrass species and regions other thanthoseinthepresentstudywouldbebeneficialtofurthergeneral-izehowgrazingaffectsmetaboliccarboncaptureacrossseagrassecosystems
Environmental temperature and irradiance are strong driv-ers of seagrassmeadow productivity (Calleja et al 2006 Lee ampDunton 1997 Lee Park amp Kim 2007 Peacuterez amp Romero 1992)howeverpreviousstudiesexaminingcontrolsonseagrassmetabo-lismhavefocusedonungrazedseagrasses(egApostolakiHolmerMarbagraveampKarakassis2010Gaciaetal2005)Meadowmetabolicproductivity correlates stronglywith above-ground seagrass bio-mass(Johnsonetal2017Figure5a)thatcanbegreatlyaffectedby green turtle grazing (Bjorndal 1980CebriaacutenampDuarte 1998Christianenetal2012FourqureanManuelCoatesKenworthyampSmith2010Williams1988)Above-groundbiomassexplained82ofthevariationinNEPacrossgrazedandungrazedT testudi-nummeadowsinthisstudybutNEPwasnotcorrelatedwitheitherenvironmentaltemperatureorirradiance(FigureS1)WethereforeinvestigatedwhetherthelackofanobservedrelationshipbetweenNEPand temperatureand irradiancewasperhapsdue toa largereffectfromdifferencesinbiomasscausedbygrazingmaskinganyeffectoftheseenvironmentalvariablesWescaledmetabolicratestotheabove-groundbiomassineachgrazedandungrazedarea(unitmetabolismperunitbiomass)andcomparedthemtotemperatureandirradianceAccountingfordifferencesinabove-groundbiomassdidnot revealanunderlyingrelationshipbetweenNEPandtheseenvironmentalparametersacrossmeadows(grazedungrazedorallareascombinedFigureS2)ThelackofarelationshipbetweenNEPandenvironmentalparametersinthisstudymaybedueinparttothe relatively limited rangeof temperature and irradiance in sea-grassmeadowsgrazedbygreenturtles in theGreaterCaribbeanAllmeadowsinthisstudywerelocatedinrelativelyshallowwarmtropicalandsubtropicalenvironmentsreceivinghighlevelsofinci-dentsunlight
41emsp|emspEffects of Halophila stipulacea invasion on meadow carbon capture
In addition to the newmetabolic carbon capture estimates wecontribute from grazed areas in T testudinum meadows wealso present the first estimates of NEP from meadows domi-nated by H stipulacea seagrass Having been first reported inthe Caribbean in 2002 (Ruiz amp Ballantine 2004)H stipulacea has spread rapidly among islands and become invasive in thisregion (Christianenet al 2018Willette et al 2014)Halophila stipulaceamayalsospreadmorequicklywithinareasofseagrassgrazedbygreenturtlesthaninungrazedareas(Christianenetal2018) suggesting that grazing may inadvertently facilitate thespreadofthisinvasivespeciesonceithasbecomeestablishedin
anareaInLacBayBonairegreenturtleshavebeguntoexpandtheirgrazingplotsintoareasofpreviouslyungrazedT testudinum asH stipulacea slowly takes over the historically grazed areaswithin thebay (Christianenet al 2018 SmuldersVonk EngelampChristianen2017) If this relationshipbetweengreen turtlesnative seagrasses and invasive seagrass exists in other areas itcouldhaveimplicationsfortotalecosystemcarbondynamicsandcarbonstorageaswellasotherecosystemfunctionswithinthesemeadows
Our resultsdemonstrate that ratesofmetabolic carboncap-turemaybe similarbetweenH stipulaceameadowsand thena-tive T testudinum meadows that they are replacing (Figure 4)HowevergiventhelimitednumberofH stipulaceameadowswewereabletosample inthisstudy(n=3)cautioniswarrantedinextrapolating these findingswidely to other locations facing in-vasionbyH stipulacea seagrassWhile ratesofNEP inH stipu-laceameadowsweresimilar tobothgrazed (n=3)andungrazed(n=2)areasofT testudinum-dominatedmeadowsinStCroixandBonaire (within-site comparisons) multiple factors affect theseratesthatmaydifferandaffectthisrelationshipinotherlocations(egdominantspeciesofnativeseagrassdensityandbiomassofH stipulaceavsthenativespeciesbladeepiphytecoverage)Forexample this relationshipmay differ in areas such as Dominica(eastern Caribbean) where the meadows undergoing H stipu-lacea invasion are dominated byS filiforme seagrass (WilletteampAmbrose 2012) Measurements from additional locations willgreatlyaidinunderstandingthespatialextentofthesimilarityinmetaboliccarboncaptureratesbetweenH stipulaceaandnativeseagrassmeadowsseenhereHoweverthecomparablemetabolicratesbetweentheinvasiveandnativeseagrassesmeasuredinthisstudysuggestsomebenefitsofseagrasspresencemayberetainedinmeadowsfollowinginvasionbyH stipulaceasuchaslocalmit-igation of ocean acidification (through metabolic buffering ofpHCamp et al 2016Hendriks et al 2014Unsworth CollierHendersonampMcKenzie2012)
It is not yet known whether comparable rates of metaboliccarbon capture will translate to comparable carbon sequestra-tionor storage inmeadowsdominatedby invasiveH stipulacea UnlikeT testudinum(vanTussenbroeketal2006)seagrassspe-cieswithinthegenusHalophiladonotformadeepbelow-groundrhizome mat (Fonseca 1989) Robust rhizome mats formed bysomeseagrassspeciescontributetohigherbelow-groundcarbonstorage(Christianenetal2013)andthelackofasimilarrhizomematmayresult in lowersedimentcarbonstorageinH stipulacea meadowsHalophila tissues also decompose quickly in sediment(JosselynFonsecaNiesenampLarson1986)and thismayaffectseagrass-derivedcarboninputtothesedimentsininvadedmead-ows compared to native species with higher refractory organicmatter content (Trevathan-Tackett et al 2017) Further studieson carbondynamics inH stipulaceameadowswill beneeded toformabetterunderstandingofhowthecontinuedinvasionbythisspecieswill affect total ecosystem carbon dynamics in seagrassmeadowsintheCaribbean
emspensp emsp | emsp1111Journal of EcologyJOHNSON et al
42emsp|emspGreater global assessment of grazed meadows needed
UngrazedareasofT testudinum-dominatedseagrassmeadowsfromthisstudyexhibitedratesofNEP(median1040mmolCmminus2 dayminus1)neartheupperendofthosereportedintheliteratureforseagrassecosystemsandgrazedareasinthisstudyexhibitedrates(median381)nearthemedianvalueofNEPforallreportedungrazedsea-grassmeadows(median271Figure6)Measurementsofseagrassmetaboliccarboncapturearerelativelygeographicallylimitedhow-ever with the majority of measurementsmdashmostly from ungrazedT testudinum-dominated meadowsmdashcoming from the GreaterCaribbeanandGulfofMexicoregionsTheCaribbeanandGulfofMexicoareecologicallyimportantgrazingregionsbutonlyasingleestimateofseagrassmetabolismfromagrazedareaexistedpriortothisstudy(Johnsonetal2017)
Allmeadows in the present study exhibited a consistent re-sponseofmetaboliccarboncapturetogreenturtlegrazinghow-ever our studywas limited to the effects of grazing in tropicaland subtropicalT testudinum-dominatedmeadows It ispossiblethatmeadows inother regionsof theworldperhapsdominatedbydifferent seagrass speciesandunderdifferentenvironmentalconditionswillnotexhibitthesameresponsetograzingintermsofmetabolic carbon dynamicsWe are aware of only one otherstudy inwhichthiswas investigated Ina tropicalThalassia hem-prichiimeadow(Tanzania)NEPwasreducedasaresultofexper-imental clipping to simulategrazing (Dahletal2016)howeveronlyhourlyratesofdaytimeNEPweremeasuredWithoutcorre-spondingratesofrespirationwecannotevaluateifNEPremained
positive in thisT hemprichiimeadowoveradiel cycleand if theresponse toexperimental clipping inTanzaniawas similar to theresponse to grazing measured in our study Further studies oneffectsofgrazingonseagrassmeadowmetaboliccarbondynam-icswould be particularly beneficial in (a) additional areaswheregrazer abundance is increasing and (b) meadows dominated byspeciesotherthanT testudinum
5emsp |emspCONCLUSIONS
Given the importance of seagrass meadows as blue carbon eco-systems (Fourqureanetal2012)andtheir role incarbonseques-tration and potential climate changemitigation it is necessary tounderstand how meadow carbon dynamics are affected by graz-ing Our study adds critical information on the effects of grazingon seagrassmetabolic carbondynamics andhow theseprocessesmaybeaffectedwith increasinggreenturtleabundanceandgraz-ingWedemonstratethattheresponseinmetaboliccarboncapturetogreenturtlegrazing isconsistentacrossseagrassecosystems inthe Greater Caribbean and Gulf of Mexico regionsmdashan area thatsupportsvastexpansesofseagrass(GreenampShort2003Jackson1997Wabnitzetal2008)Thisisanecologicallyimportantgrazingregionforgreenturtlepopulationsandgivenincreasinggreenturtleabundanceacrosstheglobe(Chaloupkaetal2008Mazarisetal2017)itisexpectedthatmoreseagrassareawillreturntoanaturalgrazedstateOurresultssuggestthat increasedgrazingwill trans-latetolowerratesofmetaboliccarboncapturebutwillnotstimulaterespirationandthemetaboliclossofcarbonfromthesemeadows
F I G U R E 6 emspRates(meanplusmnSD)ofnetecosystemproductionfrommeadowsofvariousseagrassspeciescollectedfromtheliteratureOpencircleslsquocrossedrsquocirclesandopentrianglesaregrazedThalassia testudinum(n=7)ungrazedT testudinum(n=6)andHalophila stipulacea(n=3)meadowsrespectivelyfromthepresentstudyFilledcirclesarepreviouslypublishedestimatesofungrazedseagrassmeadownetecosystemproduction(NEP)andcomefromvariouslocationsaroundtheworldCriteriaforliteraturecollectionanddatainclusionaredescribedinJohnsonetal(2017)RatesofNEPfromungrazedT testudinum and H stipulaceameadowsfromthepresentstudyareneartheupperendofpublishedvaluesMedianNEPfromgrazedT testudinumareas(presentstudy)issimilartothemedianNEPofungrazedseagrassmeadowsglobally(Section42)FiguremodifiedfromJohnsonetal(2017)toincludeadditionalestimatesLettersabovepointsdenotesitesfromthepresentstudyndashBBonaireCLittleCaymanEEleutheraFFloridaSStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306

emspensp emsp | emsp1109Journal of EcologyJOHNSON et al
wasalsopositivelycorrelatedwithseagrassmeadowcanopyheight(R2=84plt01)andbladesurfacearea(R2=92plt01)Above-groundbiomassislikelythemoreusefulpredictorofmeadowNEPhoweverasbiomassisinfluencedbybothcanopyheightandbladesurfaceareaandachange ineitherofthesevariablesresults inachangeinbiomassNEPwasnotrelatedtoseagrassshootdensityacrossmeadows(linearregressionR2lt01p=96Figure5b)norwerethererelationshipsbetweenNEPandenvironmentaltempera-ture(R2=01p=70)irradiance(R2lt01p=10)ormeadowdepth(R2lt01p=77)acrosssites(FigureS1)
4emsp |emspDISCUSSION
Through a comparative study of seagrass meadowsmdashgrazed andungrazedmdashacross a wide area encompassing sites in the GreaterCaribbean and Gulf of Mexico we found that green turtle graz-inghas a consistent effecton themetabolic carbon capture ratesofT testudinum-dominatedseagrassmeadowsRatesofmetaboliccarboncapturewerealwayslowerinareasgrazedbyturtlesthanun-grazedareasofmeadowsmdashupto96inLittleCaymanmdashbutratesofNEPwerealwaysnearorabovemetabolicbalanceEcosystemres-pirationwasnotstimulatedinrelationtoprimaryproductionwithinareas grazed by green turtles regardless of the amount of above-groundseagrassbiomassremovedthroughgrazingorotherdiffer-encesinbioticandabioticfactorsamongsitesThesefindingslendadditionalsupporttothehypothesisthatfutureratesofmetaboliccarboncapturemaybelowerinareasgrazedbyturtlesbutgrazingwillnot stimulateecosystemrespirationand result ina large rem-ineralizationof carbon currently stored in these seagrass habitats(Johnsonetal2017)
The similarity in results between St Croix (awintermeasure-ment)andtheotherfourlocationsinthepresentstudy(allsummermeasurements)strengthensourconclusionthatgrazingresultsinaconsistent response inmetaboliccarboncaptureacrossmeadowsHoweverwewerenotable toaddresspotential temporaldynam-icswithinsitesthatmayaffectnetcarboncaptureoveranannualcycle in these systems Future studies measuring carbon captureoverlongertemporalscales(ieseasonal)maybebeneficialtoourunderstanding of the effects of grazing on within-system carbondynamics
Thoughtheresponse inmetabolic rates tograzingwasconsis-tent acrossmeadows the strengthof this responsevariedamongindividual sites Some sites such as Little Cayman had consider-ablylowerratesofNEPingrazedareasthanungrazedareas(mean917)whereastheresponsetograzing(differenceinNEPbetweengrazedandadjacentungrazedareas)waslesspronouncedatothersites suchasStCroix (Table3) Seagrassmeadowcharacteristicssuch as shoot density canopy height and above-ground biomassare likely to bemore variable among ungrazed areas than grazedareasasgreenturtlegrazingresults insimilarcanopymorphome-tryacrossmeadowsintheGreaterCaribbeanregion(egBjorndal1980HernaacutendezampvanTussenbroek2014Williams1988)Given
the strong relationship between NEP and above-ground biomass(Figure 5a) high variability in NEPwould be expected across un-grazedmeadows(egFigure3a)IndeedtherangeinratesofNEPamongungrazedareaswasnearlythreetimesgreaterthantherangemeasuredacrossgrazedareasinthisstudyAssiteswerespecificallychosen that differed from each other in meadow characteristicsvariationinthestrengthofthemetabolicresponsetograzingwouldbeexpected
Variability in the strength of the metabolic response to graz-ing resulted in the lackofa significantdifferencebetweengrazedandungrazedareasattwositesmdashLacCaiBeachinBonaireandHalfSoundinEleuthera(Figure3a) It ispossiblethatotherfactorsnotaccounted for here such as local environmental factors or greenturtlegrazinghistory(egintensitylongevityofplotuse)mayhaveledtohighervariabilityinmetabolicratesatsomesitesandlackofastatisticaldifferenceInadditionmetabolicratesmayexhibitwithin-meadowvariation(patchiness)similartobelow-groundcarbonstor-age in meadows (OreskaMcglathery amp Porter 2017) leading tohighervariabilityinestimatesandhelpingtoexplainthelackofasig-nificantdifferenceinNEPatthesetwositesThiscouldbedriveninpartbywithin-meadowspatialheterogeneityinabove-groundbio-massgiventhestrongrelationshipbetweenbiomassandNEPHighwithin-meadow variability inmetabolic rates if presentmay alsohelpexplainthe largevariabilitymeasured intheungrazedarea inLittleCaymanFuturestudiesusingtheeddycorrelationtechnique(Bergetal2003) tomeasureecosystemmetabolismmaybeabletoovercomesomeofthiswithin-meadowvariabilityhoweverthegreatexpenseofthismethodmakesitprohibitiveformanystudies
Seagrass ecosystem metabolism is commonly estimated usingoxygendynamicsmeasuredwithbenthicincubationchambers(egBarroacutenetal2004Gaciaetal2005presentstudy)howeverthereare some limitations to thismethodBenthic chambersmay severbelow-ground rhizomesand release carbon that couldaffectmet-abolicestimatesIntactrhizomesontheotherhandcanaffectthediffusiveoxygenlossfromtheseagrasstotherhizosphere(BorumSand-JensenBinzerPedersenampGreve2006)andmayresultinanunderestimateofgrossprimaryproduction(andthereforeNEP)Thismaybeminimizedusingshorter incubation timesas in this studyIncubation length also strongly affects metabolic rates estimatedfromoxygendynamics (Oliveacute et al 2015)with longer incubationtimesbiasingestimatestowardsnetheterotrophyInadditiontoox-ygenmetabolismotherprocessessuchascarbonatedynamics(VanDamLopesOsburnampFourqurean2019)play important roles intotal ecosystem carbondynamicswithout affecting themeasuredproductivityofameadow
CaribbeanT testudinummeadows are highly productive (egJohnsonetal2017KochampMadden2001)andtherelationshipbetween biomass andNEP observed in this study suggests thatmeadows may maintain positive productivity as long as photo-synthetic biomass remains A threshold biomass below whichCaribbean meadows may become net heterotrophic cannot bedirectly addressed here but could be predicted to be near zeroThisisincontrasttoapreviouslysuggestedabove-groundbiomass
1110emsp |emsp emspenspJournal of Ecology JOHNSON et al
threshold(41gDMm2)belowwhichmeadowsmaybecomehet-erotrophic (Duarte et al 2010) However that threshold wascalculated for seagrasses globally and included less productivespecies Additional studies of the relationship between biomassandmeadowNEP from seagrass species and regions other thanthoseinthepresentstudywouldbebeneficialtofurthergeneral-izehowgrazingaffectsmetaboliccarboncaptureacrossseagrassecosystems
Environmental temperature and irradiance are strong driv-ers of seagrassmeadow productivity (Calleja et al 2006 Lee ampDunton 1997 Lee Park amp Kim 2007 Peacuterez amp Romero 1992)howeverpreviousstudiesexaminingcontrolsonseagrassmetabo-lismhavefocusedonungrazedseagrasses(egApostolakiHolmerMarbagraveampKarakassis2010Gaciaetal2005)Meadowmetabolicproductivity correlates stronglywith above-ground seagrass bio-mass(Johnsonetal2017Figure5a)thatcanbegreatlyaffectedby green turtle grazing (Bjorndal 1980CebriaacutenampDuarte 1998Christianenetal2012FourqureanManuelCoatesKenworthyampSmith2010Williams1988)Above-groundbiomassexplained82ofthevariationinNEPacrossgrazedandungrazedT testudi-nummeadowsinthisstudybutNEPwasnotcorrelatedwitheitherenvironmentaltemperatureorirradiance(FigureS1)WethereforeinvestigatedwhetherthelackofanobservedrelationshipbetweenNEPand temperatureand irradiancewasperhapsdue toa largereffectfromdifferencesinbiomasscausedbygrazingmaskinganyeffectoftheseenvironmentalvariablesWescaledmetabolicratestotheabove-groundbiomassineachgrazedandungrazedarea(unitmetabolismperunitbiomass)andcomparedthemtotemperatureandirradianceAccountingfordifferencesinabove-groundbiomassdidnot revealanunderlyingrelationshipbetweenNEPandtheseenvironmentalparametersacrossmeadows(grazedungrazedorallareascombinedFigureS2)ThelackofarelationshipbetweenNEPandenvironmentalparametersinthisstudymaybedueinparttothe relatively limited rangeof temperature and irradiance in sea-grassmeadowsgrazedbygreenturtles in theGreaterCaribbeanAllmeadowsinthisstudywerelocatedinrelativelyshallowwarmtropicalandsubtropicalenvironmentsreceivinghighlevelsofinci-dentsunlight
41emsp|emspEffects of Halophila stipulacea invasion on meadow carbon capture
In addition to the newmetabolic carbon capture estimates wecontribute from grazed areas in T testudinum meadows wealso present the first estimates of NEP from meadows domi-nated by H stipulacea seagrass Having been first reported inthe Caribbean in 2002 (Ruiz amp Ballantine 2004)H stipulacea has spread rapidly among islands and become invasive in thisregion (Christianenet al 2018Willette et al 2014)Halophila stipulaceamayalsospreadmorequicklywithinareasofseagrassgrazedbygreenturtlesthaninungrazedareas(Christianenetal2018) suggesting that grazing may inadvertently facilitate thespreadofthisinvasivespeciesonceithasbecomeestablishedin
anareaInLacBayBonairegreenturtleshavebeguntoexpandtheirgrazingplotsintoareasofpreviouslyungrazedT testudinum asH stipulacea slowly takes over the historically grazed areaswithin thebay (Christianenet al 2018 SmuldersVonk EngelampChristianen2017) If this relationshipbetweengreen turtlesnative seagrasses and invasive seagrass exists in other areas itcouldhaveimplicationsfortotalecosystemcarbondynamicsandcarbonstorageaswellasotherecosystemfunctionswithinthesemeadows
Our resultsdemonstrate that ratesofmetabolic carboncap-turemaybe similarbetweenH stipulaceameadowsand thena-tive T testudinum meadows that they are replacing (Figure 4)HowevergiventhelimitednumberofH stipulaceameadowswewereabletosample inthisstudy(n=3)cautioniswarrantedinextrapolating these findingswidely to other locations facing in-vasionbyH stipulacea seagrassWhile ratesofNEP inH stipu-laceameadowsweresimilar tobothgrazed (n=3)andungrazed(n=2)areasofT testudinum-dominatedmeadowsinStCroixandBonaire (within-site comparisons) multiple factors affect theseratesthatmaydifferandaffectthisrelationshipinotherlocations(egdominantspeciesofnativeseagrassdensityandbiomassofH stipulaceavsthenativespeciesbladeepiphytecoverage)Forexample this relationshipmay differ in areas such as Dominica(eastern Caribbean) where the meadows undergoing H stipu-lacea invasion are dominated byS filiforme seagrass (WilletteampAmbrose 2012) Measurements from additional locations willgreatlyaidinunderstandingthespatialextentofthesimilarityinmetaboliccarboncaptureratesbetweenH stipulaceaandnativeseagrassmeadowsseenhereHoweverthecomparablemetabolicratesbetweentheinvasiveandnativeseagrassesmeasuredinthisstudysuggestsomebenefitsofseagrasspresencemayberetainedinmeadowsfollowinginvasionbyH stipulaceasuchaslocalmit-igation of ocean acidification (through metabolic buffering ofpHCamp et al 2016Hendriks et al 2014Unsworth CollierHendersonampMcKenzie2012)
It is not yet known whether comparable rates of metaboliccarbon capture will translate to comparable carbon sequestra-tionor storage inmeadowsdominatedby invasiveH stipulacea UnlikeT testudinum(vanTussenbroeketal2006)seagrassspe-cieswithinthegenusHalophiladonotformadeepbelow-groundrhizome mat (Fonseca 1989) Robust rhizome mats formed bysomeseagrassspeciescontributetohigherbelow-groundcarbonstorage(Christianenetal2013)andthelackofasimilarrhizomematmayresult in lowersedimentcarbonstorageinH stipulacea meadowsHalophila tissues also decompose quickly in sediment(JosselynFonsecaNiesenampLarson1986)and thismayaffectseagrass-derivedcarboninputtothesedimentsininvadedmead-ows compared to native species with higher refractory organicmatter content (Trevathan-Tackett et al 2017) Further studieson carbondynamics inH stipulaceameadowswill beneeded toformabetterunderstandingofhowthecontinuedinvasionbythisspecieswill affect total ecosystem carbon dynamics in seagrassmeadowsintheCaribbean
emspensp emsp | emsp1111Journal of EcologyJOHNSON et al
42emsp|emspGreater global assessment of grazed meadows needed
UngrazedareasofT testudinum-dominatedseagrassmeadowsfromthisstudyexhibitedratesofNEP(median1040mmolCmminus2 dayminus1)neartheupperendofthosereportedintheliteratureforseagrassecosystemsandgrazedareasinthisstudyexhibitedrates(median381)nearthemedianvalueofNEPforallreportedungrazedsea-grassmeadows(median271Figure6)Measurementsofseagrassmetaboliccarboncapturearerelativelygeographicallylimitedhow-ever with the majority of measurementsmdashmostly from ungrazedT testudinum-dominated meadowsmdashcoming from the GreaterCaribbeanandGulfofMexicoregionsTheCaribbeanandGulfofMexicoareecologicallyimportantgrazingregionsbutonlyasingleestimateofseagrassmetabolismfromagrazedareaexistedpriortothisstudy(Johnsonetal2017)
Allmeadows in the present study exhibited a consistent re-sponseofmetaboliccarboncapturetogreenturtlegrazinghow-ever our studywas limited to the effects of grazing in tropicaland subtropicalT testudinum-dominatedmeadows It ispossiblethatmeadows inother regionsof theworldperhapsdominatedbydifferent seagrass speciesandunderdifferentenvironmentalconditionswillnotexhibitthesameresponsetograzingintermsofmetabolic carbon dynamicsWe are aware of only one otherstudy inwhichthiswas investigated Ina tropicalThalassia hem-prichiimeadow(Tanzania)NEPwasreducedasaresultofexper-imental clipping to simulategrazing (Dahletal2016)howeveronlyhourlyratesofdaytimeNEPweremeasuredWithoutcorre-spondingratesofrespirationwecannotevaluateifNEPremained
positive in thisT hemprichiimeadowoveradiel cycleand if theresponse toexperimental clipping inTanzaniawas similar to theresponse to grazing measured in our study Further studies oneffectsofgrazingonseagrassmeadowmetaboliccarbondynam-icswould be particularly beneficial in (a) additional areaswheregrazer abundance is increasing and (b) meadows dominated byspeciesotherthanT testudinum
5emsp |emspCONCLUSIONS
Given the importance of seagrass meadows as blue carbon eco-systems (Fourqureanetal2012)andtheir role incarbonseques-tration and potential climate changemitigation it is necessary tounderstand how meadow carbon dynamics are affected by graz-ing Our study adds critical information on the effects of grazingon seagrassmetabolic carbondynamics andhow theseprocessesmaybeaffectedwith increasinggreenturtleabundanceandgraz-ingWedemonstratethattheresponseinmetaboliccarboncapturetogreenturtlegrazing isconsistentacrossseagrassecosystems inthe Greater Caribbean and Gulf of Mexico regionsmdashan area thatsupportsvastexpansesofseagrass(GreenampShort2003Jackson1997Wabnitzetal2008)Thisisanecologicallyimportantgrazingregionforgreenturtlepopulationsandgivenincreasinggreenturtleabundanceacrosstheglobe(Chaloupkaetal2008Mazarisetal2017)itisexpectedthatmoreseagrassareawillreturntoanaturalgrazedstateOurresultssuggestthat increasedgrazingwill trans-latetolowerratesofmetaboliccarboncapturebutwillnotstimulaterespirationandthemetaboliclossofcarbonfromthesemeadows
F I G U R E 6 emspRates(meanplusmnSD)ofnetecosystemproductionfrommeadowsofvariousseagrassspeciescollectedfromtheliteratureOpencircleslsquocrossedrsquocirclesandopentrianglesaregrazedThalassia testudinum(n=7)ungrazedT testudinum(n=6)andHalophila stipulacea(n=3)meadowsrespectivelyfromthepresentstudyFilledcirclesarepreviouslypublishedestimatesofungrazedseagrassmeadownetecosystemproduction(NEP)andcomefromvariouslocationsaroundtheworldCriteriaforliteraturecollectionanddatainclusionaredescribedinJohnsonetal(2017)RatesofNEPfromungrazedT testudinum and H stipulaceameadowsfromthepresentstudyareneartheupperendofpublishedvaluesMedianNEPfromgrazedT testudinumareas(presentstudy)issimilartothemedianNEPofungrazedseagrassmeadowsglobally(Section42)FiguremodifiedfromJohnsonetal(2017)toincludeadditionalestimatesLettersabovepointsdenotesitesfromthepresentstudyndashBBonaireCLittleCaymanEEleutheraFFloridaSStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306

1110emsp |emsp emspenspJournal of Ecology JOHNSON et al
threshold(41gDMm2)belowwhichmeadowsmaybecomehet-erotrophic (Duarte et al 2010) However that threshold wascalculated for seagrasses globally and included less productivespecies Additional studies of the relationship between biomassandmeadowNEP from seagrass species and regions other thanthoseinthepresentstudywouldbebeneficialtofurthergeneral-izehowgrazingaffectsmetaboliccarboncaptureacrossseagrassecosystems
Environmental temperature and irradiance are strong driv-ers of seagrassmeadow productivity (Calleja et al 2006 Lee ampDunton 1997 Lee Park amp Kim 2007 Peacuterez amp Romero 1992)howeverpreviousstudiesexaminingcontrolsonseagrassmetabo-lismhavefocusedonungrazedseagrasses(egApostolakiHolmerMarbagraveampKarakassis2010Gaciaetal2005)Meadowmetabolicproductivity correlates stronglywith above-ground seagrass bio-mass(Johnsonetal2017Figure5a)thatcanbegreatlyaffectedby green turtle grazing (Bjorndal 1980CebriaacutenampDuarte 1998Christianenetal2012FourqureanManuelCoatesKenworthyampSmith2010Williams1988)Above-groundbiomassexplained82ofthevariationinNEPacrossgrazedandungrazedT testudi-nummeadowsinthisstudybutNEPwasnotcorrelatedwitheitherenvironmentaltemperatureorirradiance(FigureS1)WethereforeinvestigatedwhetherthelackofanobservedrelationshipbetweenNEPand temperatureand irradiancewasperhapsdue toa largereffectfromdifferencesinbiomasscausedbygrazingmaskinganyeffectoftheseenvironmentalvariablesWescaledmetabolicratestotheabove-groundbiomassineachgrazedandungrazedarea(unitmetabolismperunitbiomass)andcomparedthemtotemperatureandirradianceAccountingfordifferencesinabove-groundbiomassdidnot revealanunderlyingrelationshipbetweenNEPandtheseenvironmentalparametersacrossmeadows(grazedungrazedorallareascombinedFigureS2)ThelackofarelationshipbetweenNEPandenvironmentalparametersinthisstudymaybedueinparttothe relatively limited rangeof temperature and irradiance in sea-grassmeadowsgrazedbygreenturtles in theGreaterCaribbeanAllmeadowsinthisstudywerelocatedinrelativelyshallowwarmtropicalandsubtropicalenvironmentsreceivinghighlevelsofinci-dentsunlight
41emsp|emspEffects of Halophila stipulacea invasion on meadow carbon capture
In addition to the newmetabolic carbon capture estimates wecontribute from grazed areas in T testudinum meadows wealso present the first estimates of NEP from meadows domi-nated by H stipulacea seagrass Having been first reported inthe Caribbean in 2002 (Ruiz amp Ballantine 2004)H stipulacea has spread rapidly among islands and become invasive in thisregion (Christianenet al 2018Willette et al 2014)Halophila stipulaceamayalsospreadmorequicklywithinareasofseagrassgrazedbygreenturtlesthaninungrazedareas(Christianenetal2018) suggesting that grazing may inadvertently facilitate thespreadofthisinvasivespeciesonceithasbecomeestablishedin
anareaInLacBayBonairegreenturtleshavebeguntoexpandtheirgrazingplotsintoareasofpreviouslyungrazedT testudinum asH stipulacea slowly takes over the historically grazed areaswithin thebay (Christianenet al 2018 SmuldersVonk EngelampChristianen2017) If this relationshipbetweengreen turtlesnative seagrasses and invasive seagrass exists in other areas itcouldhaveimplicationsfortotalecosystemcarbondynamicsandcarbonstorageaswellasotherecosystemfunctionswithinthesemeadows
Our resultsdemonstrate that ratesofmetabolic carboncap-turemaybe similarbetweenH stipulaceameadowsand thena-tive T testudinum meadows that they are replacing (Figure 4)HowevergiventhelimitednumberofH stipulaceameadowswewereabletosample inthisstudy(n=3)cautioniswarrantedinextrapolating these findingswidely to other locations facing in-vasionbyH stipulacea seagrassWhile ratesofNEP inH stipu-laceameadowsweresimilar tobothgrazed (n=3)andungrazed(n=2)areasofT testudinum-dominatedmeadowsinStCroixandBonaire (within-site comparisons) multiple factors affect theseratesthatmaydifferandaffectthisrelationshipinotherlocations(egdominantspeciesofnativeseagrassdensityandbiomassofH stipulaceavsthenativespeciesbladeepiphytecoverage)Forexample this relationshipmay differ in areas such as Dominica(eastern Caribbean) where the meadows undergoing H stipu-lacea invasion are dominated byS filiforme seagrass (WilletteampAmbrose 2012) Measurements from additional locations willgreatlyaidinunderstandingthespatialextentofthesimilarityinmetaboliccarboncaptureratesbetweenH stipulaceaandnativeseagrassmeadowsseenhereHoweverthecomparablemetabolicratesbetweentheinvasiveandnativeseagrassesmeasuredinthisstudysuggestsomebenefitsofseagrasspresencemayberetainedinmeadowsfollowinginvasionbyH stipulaceasuchaslocalmit-igation of ocean acidification (through metabolic buffering ofpHCamp et al 2016Hendriks et al 2014Unsworth CollierHendersonampMcKenzie2012)
It is not yet known whether comparable rates of metaboliccarbon capture will translate to comparable carbon sequestra-tionor storage inmeadowsdominatedby invasiveH stipulacea UnlikeT testudinum(vanTussenbroeketal2006)seagrassspe-cieswithinthegenusHalophiladonotformadeepbelow-groundrhizome mat (Fonseca 1989) Robust rhizome mats formed bysomeseagrassspeciescontributetohigherbelow-groundcarbonstorage(Christianenetal2013)andthelackofasimilarrhizomematmayresult in lowersedimentcarbonstorageinH stipulacea meadowsHalophila tissues also decompose quickly in sediment(JosselynFonsecaNiesenampLarson1986)and thismayaffectseagrass-derivedcarboninputtothesedimentsininvadedmead-ows compared to native species with higher refractory organicmatter content (Trevathan-Tackett et al 2017) Further studieson carbondynamics inH stipulaceameadowswill beneeded toformabetterunderstandingofhowthecontinuedinvasionbythisspecieswill affect total ecosystem carbon dynamics in seagrassmeadowsintheCaribbean
emspensp emsp | emsp1111Journal of EcologyJOHNSON et al
42emsp|emspGreater global assessment of grazed meadows needed
UngrazedareasofT testudinum-dominatedseagrassmeadowsfromthisstudyexhibitedratesofNEP(median1040mmolCmminus2 dayminus1)neartheupperendofthosereportedintheliteratureforseagrassecosystemsandgrazedareasinthisstudyexhibitedrates(median381)nearthemedianvalueofNEPforallreportedungrazedsea-grassmeadows(median271Figure6)Measurementsofseagrassmetaboliccarboncapturearerelativelygeographicallylimitedhow-ever with the majority of measurementsmdashmostly from ungrazedT testudinum-dominated meadowsmdashcoming from the GreaterCaribbeanandGulfofMexicoregionsTheCaribbeanandGulfofMexicoareecologicallyimportantgrazingregionsbutonlyasingleestimateofseagrassmetabolismfromagrazedareaexistedpriortothisstudy(Johnsonetal2017)
Allmeadows in the present study exhibited a consistent re-sponseofmetaboliccarboncapturetogreenturtlegrazinghow-ever our studywas limited to the effects of grazing in tropicaland subtropicalT testudinum-dominatedmeadows It ispossiblethatmeadows inother regionsof theworldperhapsdominatedbydifferent seagrass speciesandunderdifferentenvironmentalconditionswillnotexhibitthesameresponsetograzingintermsofmetabolic carbon dynamicsWe are aware of only one otherstudy inwhichthiswas investigated Ina tropicalThalassia hem-prichiimeadow(Tanzania)NEPwasreducedasaresultofexper-imental clipping to simulategrazing (Dahletal2016)howeveronlyhourlyratesofdaytimeNEPweremeasuredWithoutcorre-spondingratesofrespirationwecannotevaluateifNEPremained
positive in thisT hemprichiimeadowoveradiel cycleand if theresponse toexperimental clipping inTanzaniawas similar to theresponse to grazing measured in our study Further studies oneffectsofgrazingonseagrassmeadowmetaboliccarbondynam-icswould be particularly beneficial in (a) additional areaswheregrazer abundance is increasing and (b) meadows dominated byspeciesotherthanT testudinum
5emsp |emspCONCLUSIONS
Given the importance of seagrass meadows as blue carbon eco-systems (Fourqureanetal2012)andtheir role incarbonseques-tration and potential climate changemitigation it is necessary tounderstand how meadow carbon dynamics are affected by graz-ing Our study adds critical information on the effects of grazingon seagrassmetabolic carbondynamics andhow theseprocessesmaybeaffectedwith increasinggreenturtleabundanceandgraz-ingWedemonstratethattheresponseinmetaboliccarboncapturetogreenturtlegrazing isconsistentacrossseagrassecosystems inthe Greater Caribbean and Gulf of Mexico regionsmdashan area thatsupportsvastexpansesofseagrass(GreenampShort2003Jackson1997Wabnitzetal2008)Thisisanecologicallyimportantgrazingregionforgreenturtlepopulationsandgivenincreasinggreenturtleabundanceacrosstheglobe(Chaloupkaetal2008Mazarisetal2017)itisexpectedthatmoreseagrassareawillreturntoanaturalgrazedstateOurresultssuggestthat increasedgrazingwill trans-latetolowerratesofmetaboliccarboncapturebutwillnotstimulaterespirationandthemetaboliclossofcarbonfromthesemeadows
F I G U R E 6 emspRates(meanplusmnSD)ofnetecosystemproductionfrommeadowsofvariousseagrassspeciescollectedfromtheliteratureOpencircleslsquocrossedrsquocirclesandopentrianglesaregrazedThalassia testudinum(n=7)ungrazedT testudinum(n=6)andHalophila stipulacea(n=3)meadowsrespectivelyfromthepresentstudyFilledcirclesarepreviouslypublishedestimatesofungrazedseagrassmeadownetecosystemproduction(NEP)andcomefromvariouslocationsaroundtheworldCriteriaforliteraturecollectionanddatainclusionaredescribedinJohnsonetal(2017)RatesofNEPfromungrazedT testudinum and H stipulaceameadowsfromthepresentstudyareneartheupperendofpublishedvaluesMedianNEPfromgrazedT testudinumareas(presentstudy)issimilartothemedianNEPofungrazedseagrassmeadowsglobally(Section42)FiguremodifiedfromJohnsonetal(2017)toincludeadditionalestimatesLettersabovepointsdenotesitesfromthepresentstudyndashBBonaireCLittleCaymanEEleutheraFFloridaSStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306
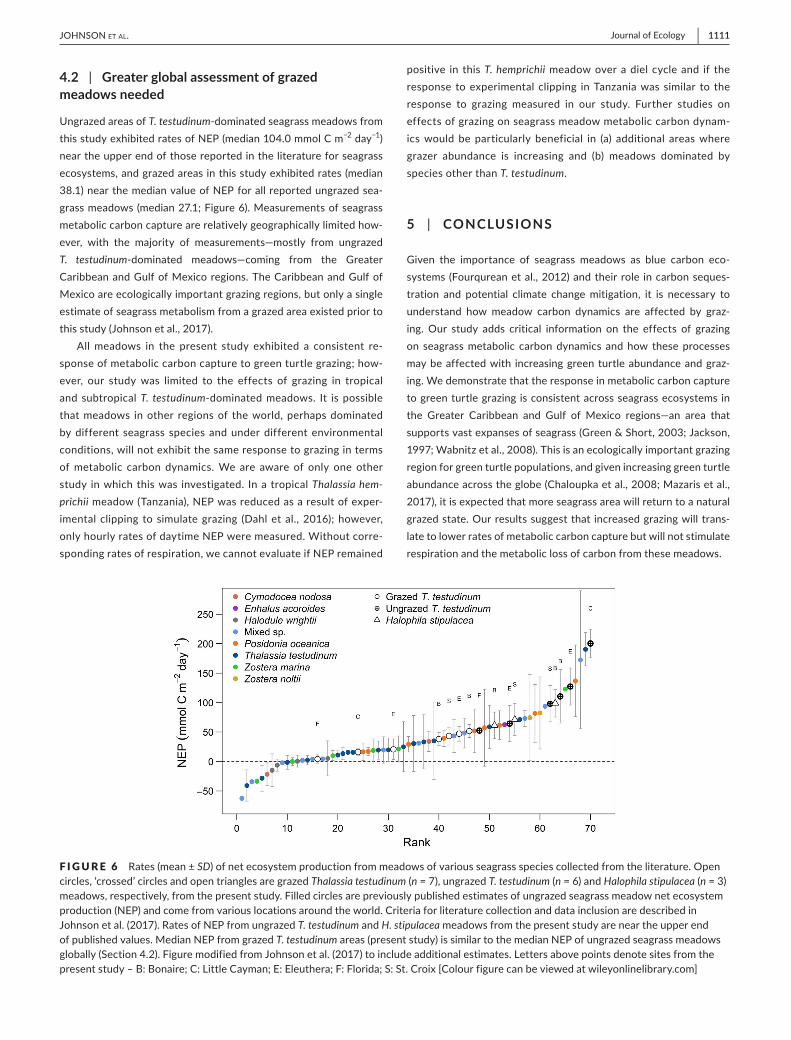
emspensp emsp | emsp1111Journal of EcologyJOHNSON et al
42emsp|emspGreater global assessment of grazed meadows needed
UngrazedareasofT testudinum-dominatedseagrassmeadowsfromthisstudyexhibitedratesofNEP(median1040mmolCmminus2 dayminus1)neartheupperendofthosereportedintheliteratureforseagrassecosystemsandgrazedareasinthisstudyexhibitedrates(median381)nearthemedianvalueofNEPforallreportedungrazedsea-grassmeadows(median271Figure6)Measurementsofseagrassmetaboliccarboncapturearerelativelygeographicallylimitedhow-ever with the majority of measurementsmdashmostly from ungrazedT testudinum-dominated meadowsmdashcoming from the GreaterCaribbeanandGulfofMexicoregionsTheCaribbeanandGulfofMexicoareecologicallyimportantgrazingregionsbutonlyasingleestimateofseagrassmetabolismfromagrazedareaexistedpriortothisstudy(Johnsonetal2017)
Allmeadows in the present study exhibited a consistent re-sponseofmetaboliccarboncapturetogreenturtlegrazinghow-ever our studywas limited to the effects of grazing in tropicaland subtropicalT testudinum-dominatedmeadows It ispossiblethatmeadows inother regionsof theworldperhapsdominatedbydifferent seagrass speciesandunderdifferentenvironmentalconditionswillnotexhibitthesameresponsetograzingintermsofmetabolic carbon dynamicsWe are aware of only one otherstudy inwhichthiswas investigated Ina tropicalThalassia hem-prichiimeadow(Tanzania)NEPwasreducedasaresultofexper-imental clipping to simulategrazing (Dahletal2016)howeveronlyhourlyratesofdaytimeNEPweremeasuredWithoutcorre-spondingratesofrespirationwecannotevaluateifNEPremained
positive in thisT hemprichiimeadowoveradiel cycleand if theresponse toexperimental clipping inTanzaniawas similar to theresponse to grazing measured in our study Further studies oneffectsofgrazingonseagrassmeadowmetaboliccarbondynam-icswould be particularly beneficial in (a) additional areaswheregrazer abundance is increasing and (b) meadows dominated byspeciesotherthanT testudinum
5emsp |emspCONCLUSIONS
Given the importance of seagrass meadows as blue carbon eco-systems (Fourqureanetal2012)andtheir role incarbonseques-tration and potential climate changemitigation it is necessary tounderstand how meadow carbon dynamics are affected by graz-ing Our study adds critical information on the effects of grazingon seagrassmetabolic carbondynamics andhow theseprocessesmaybeaffectedwith increasinggreenturtleabundanceandgraz-ingWedemonstratethattheresponseinmetaboliccarboncapturetogreenturtlegrazing isconsistentacrossseagrassecosystems inthe Greater Caribbean and Gulf of Mexico regionsmdashan area thatsupportsvastexpansesofseagrass(GreenampShort2003Jackson1997Wabnitzetal2008)Thisisanecologicallyimportantgrazingregionforgreenturtlepopulationsandgivenincreasinggreenturtleabundanceacrosstheglobe(Chaloupkaetal2008Mazarisetal2017)itisexpectedthatmoreseagrassareawillreturntoanaturalgrazedstateOurresultssuggestthat increasedgrazingwill trans-latetolowerratesofmetaboliccarboncapturebutwillnotstimulaterespirationandthemetaboliclossofcarbonfromthesemeadows
F I G U R E 6 emspRates(meanplusmnSD)ofnetecosystemproductionfrommeadowsofvariousseagrassspeciescollectedfromtheliteratureOpencircleslsquocrossedrsquocirclesandopentrianglesaregrazedThalassia testudinum(n=7)ungrazedT testudinum(n=6)andHalophila stipulacea(n=3)meadowsrespectivelyfromthepresentstudyFilledcirclesarepreviouslypublishedestimatesofungrazedseagrassmeadownetecosystemproduction(NEP)andcomefromvariouslocationsaroundtheworldCriteriaforliteraturecollectionanddatainclusionaredescribedinJohnsonetal(2017)RatesofNEPfromungrazedT testudinum and H stipulaceameadowsfromthepresentstudyareneartheupperendofpublishedvaluesMedianNEPfromgrazedT testudinumareas(presentstudy)issimilartothemedianNEPofungrazedseagrassmeadowsglobally(Section42)FiguremodifiedfromJohnsonetal(2017)toincludeadditionalestimatesLettersabovepointsdenotesitesfromthepresentstudyndashBBonaireCLittleCaymanEEleutheraFFloridaSStCroix[Colourfigurecanbeviewedatwileyonlinelibrarycom]
1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306

1112emsp |emsp emspenspJournal of Ecology JOHNSON et al
Therearemanydifferentfluxesinadditiontometabolismthatcontribute to totalecosystemcarbondynamics inseagrassmead-ows (Mateo CebriaacutenDuntonampMutchler 2006) To fully under-standtheeffectsofgreenturtlegrazingontotalcarboncaptureandstorage inseagrassecosystemsabetterunderstanding isneededonhowothercarbonfluxesandstocksalsorespondtograzingInaddition to reducing above-ground biomass green turtle grazingalsoledtoreducedbelow-groundrhizomecarbohydratereservesinameadowinBermuda(Fourqureanetal2010)Howeverabetterunderstandingof temporaldynamicsofcarbonstored inseagrasstissues is needed among naturally grazed areas The majority ofcarbon inseagrassmeadows isstored inbelow-groundsediments(Fourqureanetal2012)andthetrappingoforganicparticlesisanimportant process bywhich seagrasses accumulate sediment car-bon (GaciaGranataampDuarte1999)Recent researchhasshownthatgreenturtlegrazingmaynotdirectlyleadtoincreasederosionofsedimentsfromcoastalmeadowsinthisregion(JohnsonGulickBoltenampBjorndal2019)butfurtherresearchisneededtounder-standhowlong-term(egdecadal)grazingmayaffectsedimentcar-bonstocksFuturestudiesonadditionalcarbonfluxesandstockswill bebeneficial and complementary toour results onmetaboliccarbondynamicswhichshowthatanincreaseingreenturtleabun-danceandgrazingpressureisnotexpectedtoresultinalargerem-ineralizationofcarbonfromtheseimportantecosystems
ACKNOWLEDG EMENTS
WethankLauraPalmaforassistancewithfieldsamplinginStCroixandOlivierKramerforassistanceinBonaireWealsothankthefol-lowingpersonsandentitiesforassistanceandlogisticalsupportinplanningthesamplingatthevarioussitesinthisstudystaffoftheCentralCaribbeanMarineInstitute(LittleCayman)ClaytonPollockZandy Hillis-Starr Rich Berey and the National Park Service atBuckIslandReefNationalMonument (StCroix)SabineEngelandSTINAPA(Bonaire)NathanRobinsonAnnabelleBrooksandstaffof theCape Eleuthera Institute (Eleuthera) Thismaterial is baseduponworksupportedbytheNationalScienceFoundationGraduateResearch Fellowship under Grant No DGE-1315138 Additionalfundingwasprovidedby the Jeff andMonetteFitzsimmonsFund(ACCSTR)Theauthorsdeclarenoconflictsofinterest
AUTHORSrsquo CONTRIBUTIONS
RAJABBandKABconceivedofanddesignedthestudyAllauthorscontributedtoplanningfieldsamplingatthevarious loca-tionsRAJAGGandNCcollectedthedataRAJanalysedthedataAllauthorscontributedtothewritingofthemanuscript
DATA AVAIL ABILIT Y S TATEMENT
Data are archived in theEnvironmentalData Initiative repositoryhttpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc(JohnsonGulickConstantetal2019)
ORCID
Robert A Johnson httpsorcidorg0000-0003-4072-5623
Alexandra G Gulick httpsorcidorg0000-0002-9082-3136
Nerine Constant httpsorcidorg0000-0003-2561-719X
Alan B Bolten httpsorcidorg0000-0002-4834-2742
Fee O H Smulders httpsorcidorg0000-0003-4124-8355
Marjolijn J A Christianen httpsorcidorg0000-0001-5839-2981
Karen A Bjorndal httpsorcidorg0000-0002-6286-1901
R E FE R E N C E S
ApostolakiETHolmerMMarbagraveNampKarakassisI(2010)Metabolicimbalance in coastal vegetated (Posidonia oceanica) and unvege-tated benthic ecosystemsEcosystems13(3) 459ndash471 httpsdoiorg101007s10021-010-9330-9
BarroacutenCampDuarteCM(2009)Dissolvedorganicmatterreleaseina Posidonia oceanica meadow Marine Ecology Progress Series 37475ndash84httpsdoiorg103354meps07715
Barroacuten C Duarte C M Frankignoulle M amp Borges A V (2006)Organic carbon metabolism and carbonate dynamics in aMediterraneanseagrass (Posidonia oceanica)meadowEstuaries and Coasts29(3)417ndash426httpsdoiorg101007BF02784990
Barroacuten C Marbagrave N Terrados J Kennedy H amp Duarte C M(2004)Communitymetabolismandcarbonbudgetsalongagra-dient of seagrass (Cymodocea nodosa) colonization Limnology and Oceanography 49(5) 1642ndash1651 httpsdoiorg104319lo20044951642
BergPRoslashyHJanssenFMeyerVJoslashrgensenBBHuettelMampDeBeerD(2003)Oxygenuptakebyaquaticsedimentsmeasuredwithanovelnon-invasiveeddy-correlationtechniqueMarine Ecology Progress Series26175ndash83httpsdoiorg103354meps261075
Bjorndal K A (1980) Nutrition and grazing behavior of the greenturtle Chelonia mydas Marine Biology 56(2) 147ndash154 httpsdoiorg101007BF00397131
BorumJSand-JensenKBinzerTPedersenOampGreveTM(2006)InAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp255ndash270)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_10
CallejaMLBarroacutenCHaleJAFrazerTKampDuarteCM(2006)Lightregulationofbenthicsulfatereductionratesmediatedbysea-grass(Thalassia testudinum)metabolismEstuaries and Coasts29(6)1255ndash1264httpsdoiorg101007BF02781825
Camp E F Suggett D J Gendron G Jompa J Manfrino C ampSmithDJ(2016)MangroveandseagrassbedsprovidedifferentbiogeochemicalservicesforcoralsthreatenedbyclimatechangeFrontiers in Marine Science 3 1ndash16 httpsdoiorg103389fmars201600052
Cebriaacuten JampDuarteCM (1998) Patterns in leaf herbivoryon sea-grasses Aquatic Botany 60(1) 67ndash82 httpsdoiorg101016S0304-3770(97)00070-3
Chaloupka M Bjorndal K A Balazs G H Bolten A B EhrhartL M Limpus C J hellip Yamaguchi M (2008) Encouraging out-look for recovery of a once severely exploitedmarinemegaherbi-vore Global Ecology and Biogeography17(2) 297ndash304 httpsdoiorg101111j1466-8238200700367x
ChristianenM JAGoversLLBoumaT JKiswaraWRoelofsJ G M Lamers L P M amp van Katwijk M M (2012) Marinemegaherbivore grazing may increase seagrass tolerance to highnutrient loads Journal of Ecology 100(2) 546ndash560 httpsdoiorg101111j1365-2745201101900x
emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306

emspensp emsp | emsp1113Journal of EcologyJOHNSON et al
ChristianenMJASmuldersFOHEngelMSNavaMIWillisSDebrotAOhellipBeckingLE(2018)Megaherbivoresmayimpactexpansionof invasiveseagrass intheCaribbeanJournal of Ecology10745ndash57httpsdoiorg1011111365-274513021
ChristianenMJAvanBelzenJHermanPMJvanKatwijkMMLamersLPMvanLeentPJMampBoumaTJ(2013)Low-canopyseagrassbedsstillprovideimportantcoastalprotectionservicesPLoS ONE8(5)e62413httpsdoiorg101371journalpone0062413
DahlMDeyanovaDLyimoLDNaumlslundJSamuelssonGSMtoleraMSPhellipGullstroumlmM(2016)Effectsofshadingandsimulatedgraz-ingoncarbonsequestrationinatropicalseagrassmeadowJournal of Ecology104(3)654ndash664httpsdoiorg1011111365-274512564
deMendiburuF(2017)agricolaeStatisticalProceduresforAgriculturalResearch R package version 12-6 Retrieved from httpscranr-projectorgpackage=agricolae
DuarteCMampCebriaacutenJ(1996)Thefateofmarineautotrophicpro-ductionLimnology and Oceanography41(8)1758ndash1766httpsdoiorg104319lo19964181758
DuarteCMampChiscanoCL (1999)Seagrassbiomassandproduc-tionAreassessmentAquatic Botany65(1ndash4)159ndash174httpsdoiorg101016S0304-3770(99)00038-8
DuarteCMKennedyHMarbagraveNampHendriksI(2013)AssessingthecapacityofseagrassmeadowsforcarbonburialCurrentlimita-tionsandfuturestrategiesOcean amp Coastal Management8332ndash38httpsdoiorg101016jocecoaman201109001
DuarteCMampKrause-JensenD(2017)Exportfromseagrassmead-owscontributestomarinecarbonsequestrationFrontiers in Marine Science4(13)1ndash7httpsdoiorg103389fmars201700013
DuarteCMMarbagraveNGaciaEFourqureanJWBegginsJBarroacutenCampApostolakiET(2010)SeagrasscommunitymetabolismAssessingthecarbonsinkcapacityofseagrassmeadowsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003793
FonsecaM S (1989) Sediment stabilization byHalophila decipiens in comparisontootherseagrassesEstuarine Coastal and Shelf Science29(5)501ndash507httpsdoiorg1010160272-7714(89)90083-8
Fourqurean JWDuarteCMKennedyHMarbagraveNHolmerMMateoMAhellipSerranoO(2012)SeagrassecosystemsasagloballysignificantcarbonstockNature Geoscience5(7)505ndash509httpsdoiorg101038ngeo1477
FourqureanJWManuelSCoatesKAKenworthyWJampSmithSR (2010)Effectsofexcludingsea turtleherbivores fromasea-grassbedOvergrazingmayhaveledtolossofseagrassmeadowsinBermudaMarine Ecology Progress Series419223ndash232httpsdoiorg103354meps08853
GaciaEGranataTCampDuarteCM (1999)Anapproachtomea-surement of particle flux and sediment retention within seagrass(Posidonia oceanica) meadows Aquatic Botany 65(1ndash4) 255ndash268httpsdoiorg101016S0304-3770(99)00044-3
Gacia E Kennedy H Duarte C M Terrados J Marbagrave NPapadimitriouSampFortesM(2005)Light-dependenceofthemet-abolicbalanceofahighlyproductivePhilippineseagrasscommunityJournal of Experimental Marine Biology and Ecology 316(1) 55ndash67httpsdoiorg101016jjembe200410008
GreenEPampShortFT(2003)World Atlas of SeagrassesBerkeleyCAUniversityofCaliforniaPress
Hansen JW Thamdrup B amp Joslashrgensen B B (2000) Anoxic incu-bationof sediment in gas-tightplasticbagsAmethod forbiogeo-chemicalprocessstudiesMarine Ecology Progress Series208(1989)273ndash282httpsdoiorg103354meps208273
HemmingaMAampDuarteCM(2000)Seagrass ecologyCambridgeUKCambridgeUniversityPress
HendriksIEOlsenYSRamajoLBassoLSteckbauerAMooreTShellipDuarteCM(2014)Photosyntheticactivitybuffersoceanacidification in seagrassmeadowsBiogeosciences11(2) 333ndash346httpsdoiorg105194bg-11-333-2014
HernaacutendezA LMamp vanTussenbroekB I (2014) Patch dynamicsandspeciesshiftsinseagrasscommunitiesundermoderateandhighgrazingpressurebygreenseaturtlesMarine Ecology Progress Series517143ndash157httpsdoiorg103354meps11068
JacksonJBC(1997)ReefssinceColumbusCoral Reefs16S23ndashS32httpsdoiorg101007s003380050238
JacksonJBCKirbyMXBergerWHBjorndalKABotsfordLWBourqueBJhellipWarnerRR(2001)Historicaloverfishingandthe recent collapseof coastal ecosystemsScience293(629)629ndash637httpsdoiorg101126science1059199
Johnson R A Gulick A G Bolten A B amp Bjorndal K A (2017)Bluecarbonstoresintropicalseagrassmeadowsmaintainedundergreen turtle grazing Scientific Reports 7(1) 13545 httpsdoiorg101038s41598-017-13142-4
JohnsonRAGulickAGBoltenABampBjorndalKA(2019)Ratesof sediment resuspension and erosion following green turtle graz-inginashallowCaribbeanThalassia testudinum meadow Ecosystems httpsdoiorg101007s10021-019-00372-y
JohnsonRAGulickAGConstantNBoltenABSmuldersFOHChristianenMJAhellipBjorndalKA(2019)DatafromSeagrassecosystem metabolic carbon capture in response to green turtlegrazingacrossCaribbeanmeadows2016ndash2018Environmental Data Initiative httpsdoiorg106073pastac2ba534cb03a7d1ef203a3af63bfe6bc
Josselyn M Fonseca M Niesen T amp Larson R (1986) BiomassproductionanddecompositionofadeepwaterseagrassHalophiladecipiens ostenf Aquatic Botany 25(C) 47ndash61 httpsdoiorg1010160304-3770(86)90039-2
KennedyHBegginsJDuarteCMFourqureanJWHolmerMMarbaacuteNampMiddelburgJJ(2010)SeagrasssedimentsasaglobalcarbonsinkIsotopicconstraintsGlobal Biogeochemical Cycles24(4)1ndash8httpsdoiorg1010292010GB003848
KochMSampMaddenCJ(2001)Patternsofprimaryproductionandnutrient availability in aBahamas lagoonwith fringingmangrovesMarine Ecology Progress Series 219(1998) 109ndash119 httpsdoiorg103354meps219109
LeeKSampDuntonKH (1997)Effectsof insitu light reductiononthe maintenance growth and partitioning of carbon resourcesin Thalassia testudinum Banks ex Konig Journal of Experimental Marine Biology and Ecology210(1)53ndash73httpsdoiorg101016S0022-0981(96)02720-7
Lee K S Park S R ampKim Y K (2007) Effects of irradiance tem-peratureandnutrientsongrowthdynamicsofseagrassesAreviewJournal of Experimental Marine Biology and Ecology 350(1ndash2) 144ndash175httpsdoiorg101016jjembe200706016
LindeboomHJampSandeeAJJ(1989)Productionandconsumptionof tropical seagrass fields inEastern IndonesiameasuredwithbelljarsandmicroelectrodesNetherlands Journal of Sea Research23(2)181ndash190httpsdoiorg1010160077-7579(89)90012-4
MacreadieP INielsenDAKellewayJJAtwoodTBSeymourJRPetrouKhellipRalphPJ (2017)Canwemanagecoastaleco-systemstosequestermorebluecarbonFrontiers in Ecology and the Environment15(4)206ndash213httpsdoiorg101002fee1484
Mateo M A Cebriaacuten J Dunton K amp Mutchler T (2006) CarbonfluxinseagrassecosystemsInAWDLarkumRJOrthampCMDuarte(Eds)Seagrasses Biology ecology and conservation(pp159ndash192)DordrechtTheNetherlandsSpringer
MateoMARomeroJPeacuterezMLittlerMMampLittlerDS(1997)Dynamics ofmillenary organic deposits resulting from the growthof the Mediterranean seagrass Posidonia oceanica Estuarine Coastal and Shelf Science44(1) 103ndash110 httpsdoiorg101006ecss19960116
MazarisADSchofieldGGkazinouCAlmpanidouVampHaysGC(2017)GlobalseaturtleconservationsuccessesScience Advances3(9)e1600730httpsdoiorg101126sciadv1600730
1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306

1114emsp |emsp emspenspJournal of Ecology JOHNSON et al
MurdiyarsoDPurbopuspitoJKauffmanJBWarrenMWSasmitoSDDonatoDChellipKurniantoS(2015)ThepotentialofIndonesianmangroveforestsforglobalclimatechangemitigationNature Climate Change5(12)1089ndash1092httpsdoiorg101038nclimate2734
MurrayLampWetzelR(1987)Oxygenproductionandconsumptionas-sociatedwiththemajorautotrophiccomponents intwotemperateseagrass communitiesMarine Ecology Progress Series38 231ndash239httpsdoiorg103354meps038231
Ogden J C (1976) Some aspects of herbivore-plant relationships onCaribbean reefs and seagrassbedsAquatic Botany2(C) 103ndash116httpsdoiorg1010160304-3770(76)90013-9
OgdenJC(1980)FaunalrelationshipsinCaribbeanseagrassbedsInRCPhillipsampCPMcRoy(Eds)Handbook of seagrass biology An eco-system perspective(pp173ndash198)NewYorkNYGarlandSTPMPress
Oliveacute I Silva J Costa M M amp Santos R (2015) Estimating sea-grass community metabolism using benthic chambers The effectofincubationtimeEstuaries and Coasts39(1)138ndash144httpsdoiorg101007s12237-015-9973-z
OreskaMPJMcglatheryKJampPorterJH(2017)Seagrassbluecarbonspatialpatternsatthemeadow-scalePLoS ONE12(4)1ndash18httpsdoiorg101371journalpone0176630
Peacuterez M amp Romero J (1992) Photosynthetic response to light andtemperature of the seagrass Cymodocea nodosa and the predic-tion of its seasonality Aquatic Botany 43(1) 51ndash62 httpsdoiorg1010160304-3770(92)90013-9
Petuch E (2013) Biogeography and biodiversity of western Atlantic Mollusks(1sted)httpsdoiorg101201b14798
PollardPCampMoriartyDJW(1991)Organiccarbondecompositionpri-maryandbacterialproductivityandsulphatereductionintropicalsea-grassbedsoftheGulfofCarpentariaAustraliaMarine Ecology Progress Series69(1ndash2)149ndash159httpsdoiorg103354meps069149
RCoreTeam (2018)R A language and environment for statistical com-puting R foundation for statistical computingRetrieved fromhttpwwwr-projectorg
RheubanJEBergPampMcGlatheryKJ(2014)Ecosystemmetabo-lismalongacolonizationgradientofeelgrass (Zostera marina)mea-suredbyeddycorrelationLimnology and Oceanography59(4)1376ndash1387httpsdoiorg104319lo20145941376
RobertsonDRampCramerKL(2014)DefininganddividingtheGreaterCaribbeanInsightsfromthebiogeographyofshorefishesPLoS ONE9(7)e102918httpsdoiorg101371journalpone0102918
RuizHampBallantineDL(2004)OccurrenceoftheseagrassHalophila stipulacea in the tropical west Atlantic Bulletin of Marine Science75(1)131ndash135
ScottALYorkPHDuncanCMacreadiePIConnollyRMEllisM ThellipRasheedMA (2018) The role of herbivory in structur-ing tropical seagrass ecosystem service delivery Frontiers in Plant Science91ndash10httpsdoiorg103389fpls201800127
SerranoOMateoMARenomPampJuliagraveR(2012)CharacterizationofsoilsbeneathaPosidonia oceanica meadow Geoderma185ndash18626ndash36httpsdoiorg101016jgeoderma201203020
SmuldersFOHVonkJAEngelMSampChristianenMJA(2017)Expansion and fragment settlement of the non-native seagrassHalophila stipulacea in a Caribbean bay Marine Biology Research13(9)967ndash974httpsdoiorg1010801745100020171333620
StutesJCebrianJStutesALHunterAampCorcoranAA(2007)Benthic metabolism across a gradient of anthropogenic impact inthreeshallowcoastallagoonsinNWFloridaMarine Ecology Progress Series34855ndash70httpsdoiorg103354meps07036
TerradosJDuarteCMKamp-NielsenLAgawinNGaciaELacapDhellipGreveT(1999)AreseagrassgrowthandsurvivalconstrainedbythereducingconditionsofthesedimentAquatic Botany65(1ndash4)175ndash197httpsdoiorg101016S0304-3770(99)00039-X
Trevathan-Tackett S M Macreadie P I Sanderman J BaldockJHowes JMampRalph P J (2017) A global assessment of the
chemicalrecalcitranceofseagrasstissuesImplicationsforlong-termcarbonsequestrationFrontiers in Plant Science81ndash18httpsdoiorg103389fpls201700925
Unsworth R K F Collier C JHendersonGMampMcKenzie L J(2012)Tropical seagrassmeadowsmodifyseawatercarbonchem-istry Implications for coral reefs impacted by ocean acidifica-tion Environmental Research Letters 7(024026) 9 httpsdoiorg1010881748-932672024026
Vadeboncoeur Y (2011) Fish diversity nutrient recycling and pe-riphyton productivity in the littoral zone of Lake Tanganyika YSI Environmental httpswwwysicomFile20LibraryDocumentsApplication20NotesA591-Lake-Tanganyika-Productivityand- Nutrient-Uptake-ProODOpdf
VanDamBRLopesCOsburnCLampFourqureanJW(2019)NetheterotrophyandcarbonatedissolutionintwosubtropicalseagrassmeadowsBiogeosciences Discussions1ndash26httpsdoiorg105194bg-2019-191
van Tussenbroek B I Vonk J A Stapel J Erftemeijer P L AMiddelburg J JampZieman JC (2006) InAWDLarkumR JOrthampCMDuarte(Eds)Seagrasses Biology ecology and conser-vation(pp409ndash439)DordrechtTheNetherlandsSpringerhttpsdoiorg101007978-1-4020-2983-7_18
VianaIGSiriwardane-deZoysaRWilletteDAampGillisLG(2019)Exploringhownon-nativeseagrass speciescouldprovideessentialecosystemsservicesAperspectiveonthehighlyinvasiveseagrassHalophilastipulaceaintheCaribbeanSeaBiological Invasions21(5)1461ndash1472httpsdoiorg101007s10530-019-01924-y
Wabnitz C C Andrefouet S Torres-Pulliza DMuller-Karger F EampKramerPA (2008)Regional-scaleseagrasshabitatmapping intheWiderCaribbeanregionusingLandsatsensorsApplicationstoconservation and ecology Remote Sensing of Environment 112(8)3455ndash3467httpsdoiorg101016jrse200801020
WickhamHFrancoisRHenryLampMuumlllerK(2017)dplyr A grammar of data manipulation R package version 072Retrievedfromhttpscranrndashprojectorgpackage=dplyr
Willette D A amp Ambrose R F (2012) Effects of the invasive sea-grassHalophilastipulaceaonthenativeseagrassSyringodiumfili-formeandassociatedfishandepibiotacommunitiesintheEasternCaribbean Aquatic Botany 103 74ndash82 httpsdoiorg101016jaquabot201206007
Willette D A Chalifour J Debrot A O D Engel M S Miller JOxenford H A hellip Veacutedie F (2014) Continued expansion of thetrans-Atlantic invasive marine angiosperm Halophila stipulacea in the Eastern Caribbean Aquatic Botany 112 98ndash102 httpsdoiorg101016jaquabot201310001
WilliamsSL (1988)Thalassia testudinumproductivityandgrazingbygreenturtlesinahighlydisturbedseagrassbedMarine Biology98(3)447ndash455httpsdoiorg101007BF00391121
SUPPORTING INFORMATION
Additional supporting information may be found online in theSupportingInformationsection
How to cite this articleJohnsonRAGulickAGConstantNetalSeagrassecosystemmetaboliccarboncaptureinresponsetogreenturtlegrazingacrossCaribbeanmeadowsJ Ecol 20201081101ndash1114 httpsdoiorg1011111365-274513306





























