Embed Size (px)
Citation preview

Screening and typing of Patagonian wine yeasts forglycosidase activities
M.E. Rodrıguez1*, C.A. Lopes1*, M. van Broock2*, S. Valles3, D. Ramon3,4 and A.C. Caballero1
1Laboratorio de Microbiologıa y Biotecnologıa, Departamento de Quımica, Facultad de Ingenierıa, Universidad Nacional del Comahue,
Neuquen, Argentina, 2Laboratorio de Microbiologıa y Biotecnologıa, Departamento de Biologıa, Centro Regional Universitario Bariloche,
Universidad Nacional del Comahue, Bariloche, Rıo Negro, Argentina, 3Departamento de Biotecnologıa, Instituto de Agroquımica y
Tecnologıa de Alimentos (C.S.I.C.), Paterna, Valencia, Spain, and 4Departamento de Medicina Preventiva y Salud Publica,
Bromatologıa, Toxicologıa y Medicina Legal, Universitat de Valencia, Valencia, Spain
2003/0457: received 2 June 2003, revised 7 August 2003 and accepted 9 August 2003
ABSTRACT
M.E . RODR IGUEZ, C .A . LOPES, M. VAN BROOCK, S . VALLES, D. RAMON AND A.C . CABALLERO. 2003.
Aims: The purpose of this study was to select autochthonous glycosidase producer yeasts with potential use in
industrial production of Patagonian red wines.
Methods and Results: The study was carried out in oenological autochthonous yeasts from Comahue region
(Argentinean North Patagonia). A set of screenable yeast phenotypic characteristics indicative of their potential
usefulness in more aromatic red wine production was defined and tested in both, Saccharomyces and non-
Saccharomyces populations. Twelve isolates showing six different glycosidase phenotypes were selected and they
were characterized at species and strain levels using molecular methods. A close correlation between molecular and
phenotypic characteristics was observed. Five strains belonging to Candida guilliermondii, C. pulcherrima and
Kloeckera apiculata with highest constitutive b-glucosidase activity levels without anthocyanase activity were
discriminated. Some of them also showed constitutive b-xylosidase and inductive a-rhamnosidase activities.
Conclusions: The extension of the selection of oenological yeast to non-Saccharomyces species provided strains
possessing novel and interesting oenological characteristics which could have significant implications in the
production of more aromatic young red wine.
Significance and Impact of the Study: As these non-Saccharomyces are indigenous to wine, they can be used in
mixed starters at the beginning or as pure cultures at the end fermentation to contribute in enhancing the wine
nuance that is typical of this specific area.
Keywords: anthocyanase activity, glycosidase activities, killer behaviour, non-Saccharomyces diversity, Patagonian
oenological yeast.
INTRODUCTION
Flavour (aroma and taste) is the most important distin-
guishing characteristic of wine (Lambrechts and Pretorius
2000). Wine flavour is classified according to the sources of
the different compounds contributing to it, this includes
varietal, prefermentative, fermentative and postfermentative
flavours (Schreier 1979; Rapp 1998). It is well known that
several secondary metabolites accumulated in grapes are
responsible for providing the basis of the �varietal character�of the wine (Rapp and Versini 1991). These compounds can
be found in grapes and musts as free, volatile and odorous
forms as well as in flavourless, nonvolatile forms b-glyco-
sidically bound to disaccharide molecules such as 6-O-a,
*Consejo Nacional de Investigaciones Cientıficas y Tecnicas de la Republica
Argentina (CONICET).
Correspondence to: Adriana C. Caballero, Laboratorio de Microbiologıa y
Biotecnologıa, Departamento de Quımica, Facultad de Ingenierıa, Universidad
Nacional del Comahue, Neuquen, Argentina (e-mail: ecastro@neunet.
com.ar).
ª 2003 The Society for Applied Microbiology
Journal of Applied Microbiology 2004, 96, 84–95 doi:10.1046/j.1365-2672.2003.02032.x

LL-arabinofuranosyl-, 6-O-a, LL-rhamnopyranosyl- and 6-O-
b-DD-apiofuranosil-b-DD-glucose (Vasserot et al. 1995; Win-
terhalter and Skouroumounis 1997). These flavourless
glycosidic complexes are generally more abundant than free
odorous forms and they represent an important potential
source of the fragrant compounds in wine requiring
enzymatic or acidic hydrolysis for the liberation of their
fragrances (Gunata et al. 1985; Williams et al. 1989). The
enzymatic hydrolysis of these glycosidic complexes is a two-
step sequential reaction: first, an a-LL-arabinofuranosidase,
an a-LL-rhamnosidase or a b-DD-apiofuranosidase activity
cleaves the (1 fi 6) osidic linkage, and secondly, the flavour
compounds are liberated from the monoglucosides by the
action of a b-DD-glucosidase activity (Gunata et al. 1988).
Unlike acidic hydrolysis, enzymatic hydrolysis does not
result in modifications of the aromatic character (Gunata
et al. 1990a). Because of their characteristics (glucose
inhibition, poor stability at the low pH), grape glycosidase
activities have a minimal effect on enhancing varietal aroma
during winemaking (Canal-Llauberes 1993). As an alternat-
ive to the inefficient grape glycosidases, the presence of these
activities has been investigated in several fungal and yeast
species and it has been found that some of them possess
interesting properties for practical uses (Gunata et al. 1985,
1990a,b; Aryan et al. 1987; Vasserot et al. 1991; Rosi et al.
1994; Rogerson et al. 1995; Riou et al. 1998; Riccio et al.
1999; Spagna et al. 2002).
In this work, we investigated the presence of several
glycosidase activities in oenological indigenous yeasts from
Comahue region (north-western Patagonia, Argentina) in
order to use these yeasts themselves, or their enzymes, to
increase the varietal character of the regional red wines. To
do this, a large number of Saccharomyces and non-
Saccharomyces yeasts isolated from grape surfaces and
fermentation musts were screened twice to determine the
presence of b-glucosidase, b-xylosidase, a-LL-arabinofura-
nosidase and a-LL-rhamnosidase activities and their antho-
cyanase capabilities. The taxonomic identity of 12
indigenous yeast isolates with highest b-glucosidase activity
was confirmed by molecular methods at species and strain
levels, and their killer phenotype was characterized. On the
basis of the relationships between their phenotypic and
genotypic characteristics, these indigenous yeast isolates
were clustered in six different strains, some of which showed
interesting characteristics for winemaking.
MATERIALS AND METHODS
Yeasts
A total of 180 indigenous yeast isolates belonging to the
genera Candida (74 isolates), Torulaspora (six isolates),
Kloeckera (25 isolates) and Saccharomyces (75 isolates) were
analysed. The yeast isolates were obtained in our laboratory
from Merlot, Malbec and Trousseau type grapes (van
Broock et al. 1996; Lopes 1999) and from three different
fermentation stages of their musts spontaneously fermented
at laboratory and industrial scales (Rodrıguez 1999). Yeasts
have already been identified at genus and species level
according to conventional physiological and morphological
tests and keys proposed by Kreger-van Rij (1984) and
Kurtzman and Fell (1998) (van Broock et al. 1996; Lopes
1999; Rodrıguez 1999).
Glycosidase screening methods
Screening for detecting b-glucosidase activity was carried
out on agar plates with arbutin as substrate. The medium,
containing 6Æ7 g yeast nitrogen base (YNB; Difco), 5 g
arbutin, 20 g agar per litre and pH 5Æ0, was autoclaved at
120�C for 15 min. Immediately after sterilization 2 ml of a
sterile 1% (w/v) ferric ammonium citrate solution was
added to 100 ml melted medium (Rosi et al. 1994). Each
plate was inoculated in radial streaks with 24-h-old yeast
cultures, incubated at 25�C and examined daily during a
week. An uninoculated plate served as the control. Yeast
isolates with b-glucosidase activity hydrolyse the substrate
and a dark brown halo develops in the agar medium.
Screening for detecting all b-glucosidase, b-xylosidase,
a-rhamnosidase and a-arabinofuranosidase activities was
carried out on agar plates containing 1Æ7 g YNB (without
amino acids and ammonium sulphate; Difco), 5 g ammo-
nium sulphate, 5 g source carbon and 20 g agar per litre.
The pH was adjusted to 5Æ5. The carbon sources assayed
were either arabinose, cellobiose, glucose, rhamnose and
xylose (Sigma). Glycosidase activities were determined by
using the appropriate 4-methylumbelliferyl glycoside (Sig-
ma) as substrate, as described by Manzanares et al. (1999).
The presence of the enzymatic activity was visualized as a
fluorescent halo surrounding yeast growth by plate expo-
sition to u.v. light.
b-Glucosidase activity
Liquid culture conditions. A medium containing 1Æ7 g
YNB, 5Æ0 g (NH4)2SO4, 5Æ0 g yeast extract, 5Æ0 g peptone,
5Æ0 g glucose per litre and pH 5Æ0 was used to examine
yeasts showing b-glucosidase activity and to localize the
enzymatic activity. Inocula were prepared by growing a
loopful of cells from GPY plate (5Æ0 g yeast extract, 5Æ0 g
peptone, 40Æ0 g dextrose, 20 g agar per litre; pH 5Æ0) in
10 ml of liquid medium. After 24 h, a 2% volume of the
inoculum was added to 100 ml Erlenmeyer flasks filled to
20% of their volume and shaken at 180 rev min)1 in a
Vicking shaker at 25�C for 24 h (aerobic conditions). In
studies on the influence of anaerobic growth conditions, the
GLYCOSIDASE ACTIVIT IES IN PATAGONIAN YEASTS 85
ª 2003 The Society for Applied Microbiology, Journal of Applied Microbiology, 96, 84–95, doi:10.1046/j.1365-2672.2003.02032.x

yeasts were grown in 20 ml screw-capped tubes totally filled
and incubated without shaking at 25�C for 3 days. Growth
control was checked by monitoring the culture absorbance at
600 nm and tubes or Erlenmeyer flasks containing uninoc-
ulated media were used as the blank. Once the assays were
finished, cultures were centrifuged (5000 g, 10 min, 4�C)
and the cells and culture supernatant were assayed for
enzymatic activity and the dry weight was determined.
Whole cell preparation. Cells harvested from 1 ml of
culture as indicated above, were washed twice with cold
sterile distilled water, centrifuged and the pellet was
resuspended in 0Æ5 ml of 100 mmol l)1 citrate–phosphate
buffer, pH 5Æ0, and used for enzymatic assay.
Permeabilized cell preparation. The procedure of
Salmon (1984) with some modifications was used: 5 ml of
liquid culture were centrifuged and washed with 5 ml of cold
sterile distilled water. The pellet was resuspended in 1 ml of
75 mmol l)1 imidazole buffer pH 7Æ5, and added quickly to
50 ll of 0Æ3 mmol l)1 glutathion, 10 ll of 10% Triton X-100
and 50 ll of toluene/ethanol (1 : 4 v/v). The suspension was
shaken vigorously for 5 min and then centrifuged. The yeast
pellet was suspended in 5 ml cold sterile water: 1 ml of
this suspension was centrifuged, washed with cold sterile
distilled water and the pellet was resuspended in 0Æ5 ml
citrate–phosphate buffer and used for enzymatic assay.
Enzyme assay. b-Glucosidase activity was assayed by
measuring the amount of p-nitrophenol (pNP) released from
p-nitrophenyl-b-DD-glucoside (pNPG) used as substrate.
Enzyme solution (0Æ1 ml) was mixed with 0Æ4 ml of a
2 mmol l)1 solution of pNPG in 100 mmol l)1 citrate–
phosphate buffer, pH 5Æ0. The reaction mixture was incu-
bated at 30�C for 5–15 min and subsequently, 2Æ0 ml of
250 mmol l)1 Na2CO3 was added to stop the reaction. The
released pNP in this mixture was measured spectrophoto-
metrically at 405 nm in a Shimadzu UV–V spectrophoto-
meter (Shimadzu Corporation, Kyoto, Japan). The pNP molar
extinction coefficient used was � ¼ 18 300 mol)1 l cm)1.
One unit (U) of enzyme activity was defined as the quantity
of enzyme that released 1 lmol of pNP per minute under the
above experimental conditions. The values of intracellular
activity were obtained subtracting the whole cell enzymatic
activity values from the permeabilized cell values. All assays
were performed in duplicate.
Dry weight. Ten millilitres of culture samples were
centrifuged at 10 000 g 5 min. The pellets were washed
twice with 5 ml of cold sterile distilled water and resus-
pended in sterile distilled water. These cell suspensions were
placed on preweighed dishes and dried at 105�C until
constant weight.
PCR–RFLP analysis of rDNA
The 5Æ8S-ITS region was amplified in a Progene thermo-
cycler (Techne, Cambridge, UK) using ITS1 (5¢-TCCGT
AGGTGAACCTGCGG-3¢) and ITS4 (5¢-TCCTCCGCT
TATTGATATGC-3¢) primers already described (White
et al. 1990). PCR conditions were identical to those
described by Esteve-Zarzoso et al. (1999). The amplified
DNAs (0Æ5–10 lg) were digested without further purifica-
tion with the restriction endonucleases CfoI, HaeIII, HinfI
and DdeI (Roche Molecular Biochemicals, Mannheim,
Germany) according to the supplier’s instructions. PCR
products and their restriction fragments were separated on
1Æ4% (w/v)and 3% (w/v) agarose gels, respectively, with 1x
TAE buffer (40 mmol l)1 Tris–acetate, 1 mmol l)1 EDTA,
pH 8). After electrophoresis, gels were stained with ethi-
dium bromide (5 lg ml)1) and visualized under u.v. light. A
100-bp DNA ladder marker (Gibco BRL, Gaithersburg,
MD, USA) served as size standard.
The taxonomic identity of indigenous S. cerevisiae isolates
tested in this study has already been confirmed by us using
the same molecular method (Lopes et al. 2002).
Karyotype analysis
DNA was prepared in agarose plugs as described by Carle
and Olson (1985). Chromosomal profiles were determined
using the CHEF technique with a CHEF-DRIII (Bio-Rad,
Richmond CA, USA). The yeast chromosomes were
separated on 1% (w/v) chromosomal grade agarose gels
(Bio-Rad) under the following running conditions: electric
field, 6 V cm)1; angle, 120�; temperature, 14�C; block 1,
60 s pulse time for 15 h; block 2, 120 s pulse time for 13 h;
buffer 0Æ5 · TBE (45 mmol l)1 Tris–borate pH 7Æ5,
1 mmol l)1 EDTA pH 8Æ0). After electrophoresis, gels were
stained with ethidium bromide (5 lg ml)1) and visualized
under u.v. light.
mtDNA-RFLP analysis
Total DNA extraction and mtDNA restriction analysis were
performed by the method of Querol et al. (1992) modified
by Lopez et al. (2001). Yeast DNA was digested with HinfI
restriction enzyme (Roche Molecular Biochemicals, Mann-
heim, Germany) and the fragments were separated in TAE
buffer containing 1% (w/v) agarose gels (Fernandez-
Espinar et al. 2000).
Killer behaviour
Killer behaviour was assayed using the seeded agar-plate
technique as described by Sangorrın et al. (2002). The killer
sensitivity of the indigenous yeasts were determined
86 M.E. RODRIGUEZ ET AL.
ª 2003 The Society for Applied Microbiology, Journal of Applied Microbiology, 96, 84–95, doi:10.1046/j.1365-2672.2003.02032.x

contrasting them to reference panel of the 10 collection well-
known killer strains and their killer character was evaluated
against two collection sensitive strains. The collection killer
strains used were S. cerevisiae YAT 679 (K1 type), S.
cerevisiae NCYC 738 (K2 type), S. capensis NCYC 671 (K3
type), C. glabrata NCYC 388 (K4 type), Hansenula anomala
NCYC 434 (K5 type), Kluyveromyces fragilis NCYC 587 (K6
type), C. valida NCYC 327 (K7 type), H. anomala NCYC
435 (K8 type) and H. mrakii NCYC 500 (K9 type), K.
drosophilarum NCYC 575 (K10 type), provided by Professor
Isato Kono (Industrial Technology Center of Okayama
Prefecture, Japan), and S. cerevisiae P352 (K1 type) from
PROIMI yeast collection from Tucuman (Argentina). The
collection-sensitive strains used were S. cerevisiae P351
(PROIMI yeast collection) and C. glabrata NCYC 388 above
cited.
Anthocyanase activity
Anthocyanase activity was measured using red wine as
substrate. Aerobically grown YNB-cellobiose or rhamnose
cells of 1.5 ml was collected by centrifugation and resus-
pended in 1Æ5 ml of red wine and anaerobically incubated at
4�C during a week. Wine decolourization was monitored
measuring the absorbance at 520 nm in 2 mm light path
cuvettes. Glycosidase activity was also assayed using the
appropriate pNP glycoside (Sigma) as substrate.
Chemicals
Arbutin, pNPG, pNP, MUG, MUX, MUA, MUR,
glutathion and Triton X-100 were supplied by Sigma, and
culture medium constituents by Difco. All other chemicals
used were of reagent grade.
Data analysis
Significant differences between experimental figures were
estimated using Student’s test. Dendrograms were gener-
ated using an unweighed pair group with arithmetic average
(UPGMA) algorithm and the concordance between indi-
viduals was (1-SM) matching.
RESULTS
Glycosidase screenings and characterization
Patagonian Saccharomyces and non-Saccharomyces yeast
isolates, all of oenological origin, were screened for
b-glucosidase activity on agar plates containing arbutin as
substrate. Forty-five of 180 analysed isolates showed enzy-
matic activity and they were characterized as strong (11
isolates), moderate (eight isolates) or weak (26 isolates)
according to the colour intensity (dark, fair or light brown)
of the halo (Table 1). Among these producer isolates, 44
belonged to C. guilliermondii, C. pulcherrima and Kloeckeraapiculata species and only one belonged to S. cerevisiae; this
isolate came from grape berry surface and it was a weak
producer (Table 1).
All isolates characterized as enzyme producers on plate
assay were grown in liquid media under aerobic conditions
and they were tested for b-glucosidase activity using
p-nitrophenyl-b-DD-glucoside as substrate (in vitro assays).
In these assays, whole cells and culture media were used as
enzyme source. Under assay conditions, none of the yeast
isolates showed any capacity to excrete the enzyme into the
medium. Analysis of whole cell activity data, using the box-
and-whiskers statistic method, revealed that within
C. guilliermondii, C. pulcherrima and K. apiculata producer
species, the activity levels found in must isolates (median
values of 13Æ00, 1Æ00 and 0Æ07 U g)1 dry weight, respect-
ively) were greater than in grape isolates (median values of
2Æ1, 0Æ40 and 0Æ05 U g)1 dry weight, respectively). These
results also evidence that C. guilliermondii isolates from must
(M229, 231 and 425) were the greatest b-glucosidase
producers. However, some single grape isolates of this
species (A135, M102 and M119) as well as some single
isolates of C. pulcherrima (T152 and M227) and K. apiculata
(M321, 325 and 328) also showed very high enzyme activity
levels (outlier values), between four to 40 times higher than
the medians of their populations. These 11 best b-glucosi-
dase producers, characterized as strong on plate assay, were
selected to continue the study. The moderate to weak
producer K. apiculata M326 was also included in order to
continue checking the screening and selection method and
the 12 isolates were renamed as V1–V12 (see Table 2).
Because yeast strains may have exocellular, parietal (cell
wall bound) and intracellular b-glycosidase activity, this
activity was assayed on the culture supernatant medium
(extracellular), on whole cells (parietal) and on permeabi-
lized cells; intracellular activity was estimated from the latter
(see �Methods�). Taking into account that winemaking is an
anaerobic process, yeast cells were grown under aerobic and
partially anaerobic conditions, and total b-glucosidase
activity was estimated as the sum of the three activities.
As a whole, the results from these assays showed two
different behaviour patterns among indigenous isolates.
C. pulcherrima isolates and four of the six C. guilliermondii
isolates (V3, V5, V7 and V8) yielded the highest levels of
enzyme activity and neither b-glucosidase production nor
cellular location were significantly affected by culture
aeration conditions (Table 2). Additionally, whereas in the
first isolates almost all the activity was detected in perme-
abilized cells, indicating the presence of an intracellular
enzyme, the last isolates showed important levels in whole
cells and permeabilized cells, indicating the presence of
GLYCOSIDASE ACTIVIT IES IN PATAGONIAN YEASTS 87
ª 2003 The Society for Applied Microbiology, Journal of Applied Microbiology, 96, 84–95, doi:10.1046/j.1365-2672.2003.02032.x

both parietal and intracellular enzymes (Table 2). However,
under anaerobic conditions, the other two C. guilliermondii
isolates (V1 and V2) and K. apiculata isolates showed the
highest levels of activity on whole cells (‡89Æ5 and ‡96Æ9% of
the total activity, respectively), evidencing the presence of
parietal enzyme. Aerobic growth conditions greatly stimu-
lated b-glycosidase activity in these isolates (Table 2).
Candida guilliermondii V2 shows fivefold more enzymatic
activity under this condition than under anaerobic condi-
tions, with a significant increase of both intracellular and
parietal activities. Although their activity values are lower,
the same trend may be observed in C. guilliermondii V1 and
all K. apiculata isolates. Additionally, certain activity was
observed in supernatant media, exocellular enzyme, but its
level was small and, except for V8 isolate, it was only
observed under anaerobic conditions (Table 2).
A second plate screening for detecting other glycosidase
activities were realized on the yeast isolates. In this assay,
yeast cells were grown on agar plates containing different
carbon source and b-DD-xylosidase, a-LL-rhamnosidase and
a-LL-arabinofuranosidase as well as b-DD-glucosidase were
detected using the appropriate 4-methylumbelliferyl-
glycoside as substrate. The results showed in Table 3 reveal
that all C. guilliermondii and C. pulcherrima isolates were
capable to produce both b-glucosidase and b-xylosidase
activities on all carbon source where they can grow,
indicating that the enzymes could be synthesized constitu-
tively, whilst K. apiculata isolates only produced b-glucosi-
dase. Candida guilliermondii V2 also showed capability to
hydrolyse MUR (a-rhamnosidase activity), but this enzy-
matic activity was only detected using rhamnose as substrate
(inducible activity) (Table 3). None of the 12 yeast strains
produced a-arabinofuranosidase activity.
Killer behaviour
Under assay conditions, neither C. pulcherrima, K. apiculatanor C. guilliermondii isolates were capable of producing killer
toxin. However, whereas the isolates belonging to the two
former species were immune against all assayed killer toxins
Table 1 Indigenous yeast isolates tested for
b-glucosidase activity
Species Source
Tested
isolate
number Activity* Isolate identity�
Saccharomyces cerevisiae Grape 1 +(1) A102w
Must 72 – –
Saccharomyces kluyveri Must 2 –
Kloeckera apiculata Grape 4 +(3) A117w and 119w; T124m
Must 21 +(15) M201m, 204m, 313w, 315w, 321s,
325s, 326m, 327s; 402w, 407w, 411w,
412w and 414w T202w and 206w
Torulaspora delbrueckii Must 4 – –
Torulaspora pretoriensis Must 2 – –
Candida colliculosa Must 7 – –
Candida dattila Must 16 – –
Candida guilliermondii Grape 13 +(13) M102s and 119s; A108w,
109w, 110w, 111w,
114w, 131w, 132m, 135s;
142w, 146w and 150w
Must 3 +(3) M229s, 231s and 425s
Candida parapsilosis Grape 2 – –
Candida pulcherrima Grape 10 +(6) T140w, 149w and 152s A123w,
128w and 162m
Must 17 +(4) M227s T205m; 218m and 220w
Candida stellata Must 6 – –
Total 180 +(45)
*Activity is expressed as hydrolysis of arbutin. The figures between brackets indicate the number
of yeast isolates with b-glucosidase activity.
�The capital letters indicate the grape variety: A, Merlot; M, Malbec; T, Trousseau. The first
digit indicates the source: 1, grape; 2, must completely processed in laboratory; 3, must partially
processed in laboratory; 4, must completely processed in winery and the last two digits represent
the isolate number within each series. The superscript letters s, m and w indicate strong, moderate
or weak b-glucosidase production, respectively.
88 M.E. RODRIGUEZ ET AL.
ª 2003 The Society for Applied Microbiology, Journal of Applied Microbiology, 96, 84–95, doi:10.1046/j.1365-2672.2003.02032.x
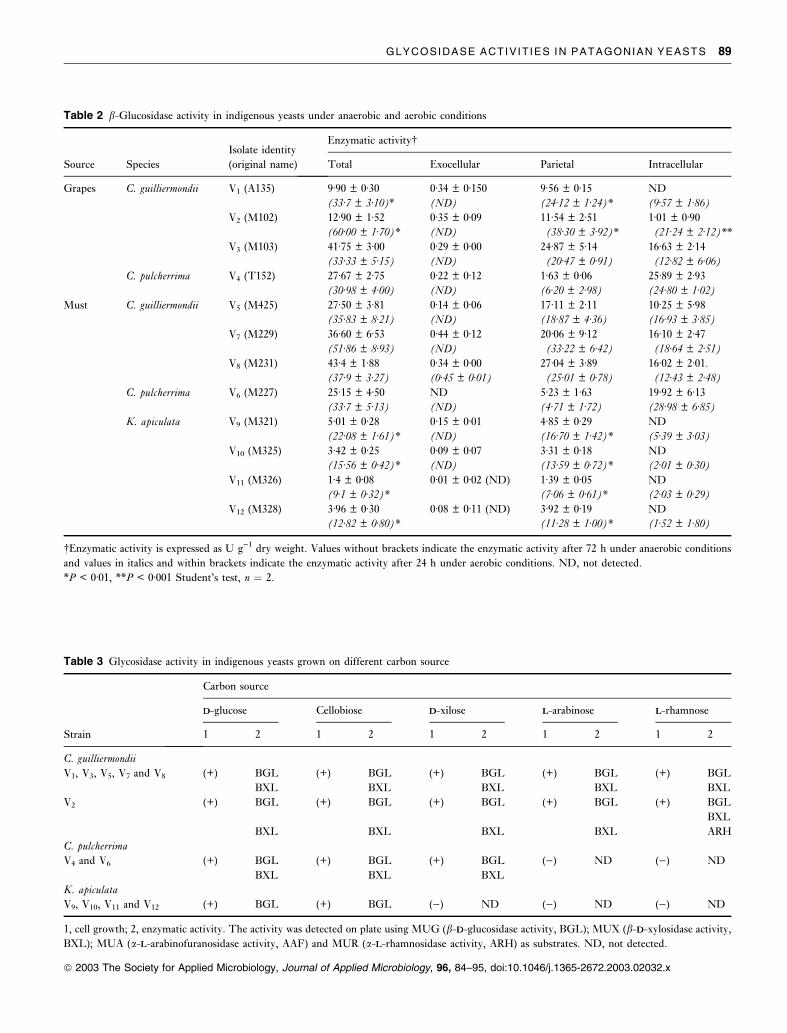
Table 3 Glycosidase activity in indigenous yeasts grown on different carbon source
Strain
Carbon source
DD-glucose Cellobiose DD-xilose LL-arabinose LL-rhamnose
1 2 1 2 1 2 1 2 1 2
C. guilliermondii
V1, V3, V5, V7 and V8 (+) BGL (+) BGL (+) BGL (+) BGL (+) BGL
BXL BXL BXL BXL BXL
V2 (+) BGL (+) BGL (+) BGL (+) BGL (+) BGL
BXL
BXL BXL BXL BXL ARH
C. pulcherrima
V4 and V6 (+) BGL (+) BGL (+) BGL ()) ND ()) ND
BXL BXL BXL
K. apiculata
V9, V10, V11 and V12 (+) BGL (+) BGL ()) ND ()) ND ()) ND
1, cell growth; 2, enzymatic activity. The activity was detected on plate using MUG (b-DD-glucosidase activity, BGL); MUX (b-DD-xylosidase activity,
BXL); MUA (a-LL-arabinofuranosidase activity, AAF) and MUR (a-LL-rhamnosidase activity, ARH) as substrates. ND, not detected.
Table 2 b-Glucosidase activity in indigenous yeasts under anaerobic and aerobic conditions
Source Species
Isolate identity
(original name)
Enzymatic activity�
Total Exocellular Parietal Intracellular
Grapes C. guilliermondii V1 (A135) 9Æ90 ± 0Æ30
(33Æ7 ± 3Æ10)*
0Æ34 ± 0Æ150
(ND)
9Æ56 ± 0Æ15
(24Æ12 ± 1Æ24)*
ND
(9Æ57 ± 1Æ86)
V2 (M102) 12Æ90 ± 1Æ52
(60Æ00 ± 1Æ70)*
0Æ35 ± 0Æ09
(ND)
11Æ54 ± 2Æ51
(38Æ30 ± 3Æ92)*
1Æ01 ± 0Æ90
(21Æ24 ± 2Æ12)**
V3 (M103) 41Æ75 ± 3Æ00
(33Æ33 ± 5Æ15)
0Æ29 ± 0Æ00
(ND)
24Æ87 ± 5Æ14
(20Æ47 ± 0Æ91)
16Æ63 ± 2Æ14
(12Æ82 ± 6Æ06)
C. pulcherrima V4 (T152) 27Æ67 ± 2Æ75
(30Æ98 ± 4Æ00)
0Æ22 ± 0Æ12
(ND)
1Æ63 ± 0Æ06
(6Æ20 ± 2Æ98)
25Æ89 ± 2Æ93
(24Æ80 ± 1Æ02)
Must C. guilliermondii V5 (M425) 27Æ50 ± 3Æ81
(35Æ83 ± 8Æ21)
0Æ14 ± 0Æ06
(ND)
17Æ11 ± 2Æ11
(18Æ87 ± 4Æ36)
10Æ25 ± 5Æ98
(16Æ93 ± 3Æ85)
V7 (M229) 36Æ60 ± 6Æ53
(51Æ86 ± 8Æ93)
0Æ44 ± 0Æ12
(ND)
20Æ06 ± 9Æ12
(33Æ22 ± 6Æ42)
16Æ10 ± 2Æ47
(18Æ64 ± 2Æ51)
V8 (M231) 43Æ4 ± 1Æ88
(37Æ9 ± 3Æ27)
0Æ34 ± 0Æ00
(0Æ45 ± 0Æ01)
27Æ04 ± 3Æ89
(25Æ01 ± 0Æ78)
16Æ02 ± 2Æ01.
(12Æ43 ± 2Æ48)
C. pulcherrima V6 (M227) 25Æ15 ± 4Æ50
(33Æ7 ± 5Æ13)
ND
(ND)
5Æ23 ± 1Æ63
(4Æ71 ± 1Æ72)
19Æ92 ± 6Æ13
(28Æ98 ± 6Æ85)
K. apiculata V9 (M321) 5Æ01 ± 0Æ28
(22Æ08 ± 1Æ61)*
0Æ15 ± 0Æ01
(ND)
4Æ85 ± 0Æ29
(16Æ70 ± 1Æ42)*
ND
(5Æ39 ± 3Æ03)
V10 (M325) 3Æ42 ± 0Æ25
(15Æ56 ± 0Æ42)*
0Æ09 ± 0Æ07
(ND)
3Æ31 ± 0Æ18
(13Æ59 ± 0Æ72)*
ND
(2Æ01 ± 0Æ30)
V11 (M326) 1Æ4 ± 0Æ08
(9Æ1 ± 0Æ32)*
0Æ01 ± 0Æ02 (ND) 1Æ39 ± 0Æ05
(7Æ06 ± 0Æ61)*
ND
(2Æ03 ± 0Æ29)
V12 (M328) 3Æ96 ± 0Æ30
(12Æ82 ± 0Æ80)*
0Æ08 ± 0Æ11 (ND) 3Æ92 ± 0Æ19
(11Æ28 ± 1Æ00)*
ND
(1Æ52 ± 1Æ80)
�Enzymatic activity is expressed as U g)1 dry weight. Values without brackets indicate the enzymatic activity after 72 h under anaerobic conditions
and values in italics and within brackets indicate the enzymatic activity after 24 h under aerobic conditions. ND, not detected.
*P < 0Æ01, **P < 0Æ001 Student’s test, n ¼ 2.
GLYCOSIDASE ACTIVIT IES IN PATAGONIAN YEASTS 89
ª 2003 The Society for Applied Microbiology, Journal of Applied Microbiology, 96, 84–95, doi:10.1046/j.1365-2672.2003.02032.x
displaying a neutral phenotype (S)K)), C. guilliermondii
isolates showed a sensitive phenotype to K8, K9 and K10
isolates (S+K)).
Antocyanase activity
In this assay yeast cells grown on cellobiose (cell bound b-
glucosidase activities ‡10Æ00 ± 1Æ50 U g)1 dry weight) or
rhamnose (cell bound V2a-rhamnosidase activity ¼5Æ84 U g)1 dry weight) were used as enzyme source and
the progress of decolourization of a commercial red wine was
studied during a week. No significant differences in colour
figures were observed between control (uninoculated red
wine, A520 ¼ 0Æ731 ± 0Æ010, n ¼ 2) and experimental sam-
ples (V2, V5, V6, V9-cellobiose and V2-rhamnose inoculated
red wine, A520 ¼ 0Æ669± 0Æ039, 0Æ671 ± 0Æ051, 0Æ780 ±
0Æ021, 0Æ702 ± 0Æ060 and 0Æ739 ± 0Æ014, respectively;
n ¼ 2). However, low but significant residual b-glycosidase
(‡100 ± 9 mU g)1 dry weight) and a-rhamnosidase activ-
ities (700 ± 23 mU g)1 dry weight) were observed at the
end of the assay.
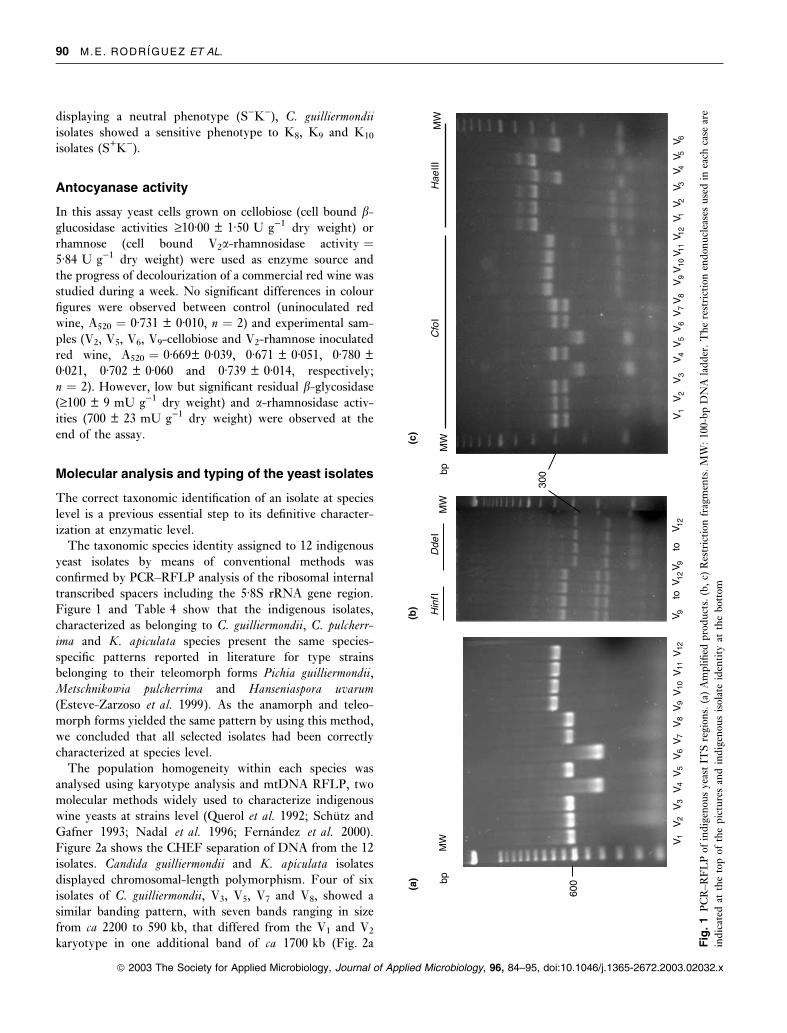
Molecular analysis and typing of the yeast isolates
The correct taxonomic identification of an isolate at species
level is a previous essential step to its definitive character-
ization at enzymatic level.
The taxonomic species identity assigned to 12 indigenous
yeast isolates by means of conventional methods was
confirmed by PCR–RFLP analysis of the ribosomal internal
transcribed spacers including the 5Æ8S rRNA gene region.
Figure 1 and Table 4 show that the indigenous isolates,
characterized as belonging to C. guilliermondii, C. pulcherr-
ima and K. apiculata species present the same species-
specific patterns reported in literature for type strains
belonging to their teleomorph forms Pichia guilliermondii,
Metschnikowia pulcherrima and Hanseniaspora uvarum
(Esteve-Zarzoso et al. 1999). As the anamorph and teleo-
morph forms yielded the same pattern by using this method,
we concluded that all selected isolates had been correctly
characterized at species level.
The population homogeneity within each species was
analysed using karyotype analysis and mtDNA RFLP, two
molecular methods widely used to characterize indigenous
wine yeasts at strains level (Querol et al. 1992; Schutz and
Gafner 1993; Nadal et al. 1996; Fernandez et al. 2000).
Figure 2a shows the CHEF separation of DNA from the 12
isolates. Candida guilliermondii and K. apiculata isolates
displayed chromosomal-length polymorphism. Four of six
isolates of C. guilliermondii, V3, V5, V7 and V8, showed a
similar banding pattern, with seven bands ranging in size
from ca 2200 to 590 kb, that differed from the V1 and V2
karyotype in one additional band of ca 1700 kb (Fig. 2a
300
Cfo
IH
aeIII
MW
MW
MW
600
MW
(a)
(b)
(c)
bpbp
Hin
fID
deI
V1
V2
V3
V4
V5
V6
V7
V8
V9
V10
V11
V12
V 9to
V 12V 9
toV 1
2V
1V
2V
3V
4V
5V
6V
7V
8V
9V
10V 1
1V 1
2V 1
V 2V 3
V 4V 5
V 6
Fig
.1
PC
R–
RF
LP
ofin
dig
enou
sye
ast
ITS
regi
ons.
(a)
Am
pli
fied
pro
du
cts.
(b,
c)R
estr
icti
onfr
agm
ents
.M
W:
100-
bp
DN
Ala
dd
er.
Th
ere
stri
ctio
nen
don
ucl
ease
su
sed
inea
chca
sear
e
ind
icat
edat
the
top
ofth
ep
ictu
res
and
ind
igen
ous
isol
ate
iden
tity
atth
eb
otto
m
90 M.E. RODRIGUEZ ET AL.
ª 2003 The Society for Applied Microbiology, Journal of Applied Microbiology, 96, 84–95, doi:10.1046/j.1365-2672.2003.02032.x
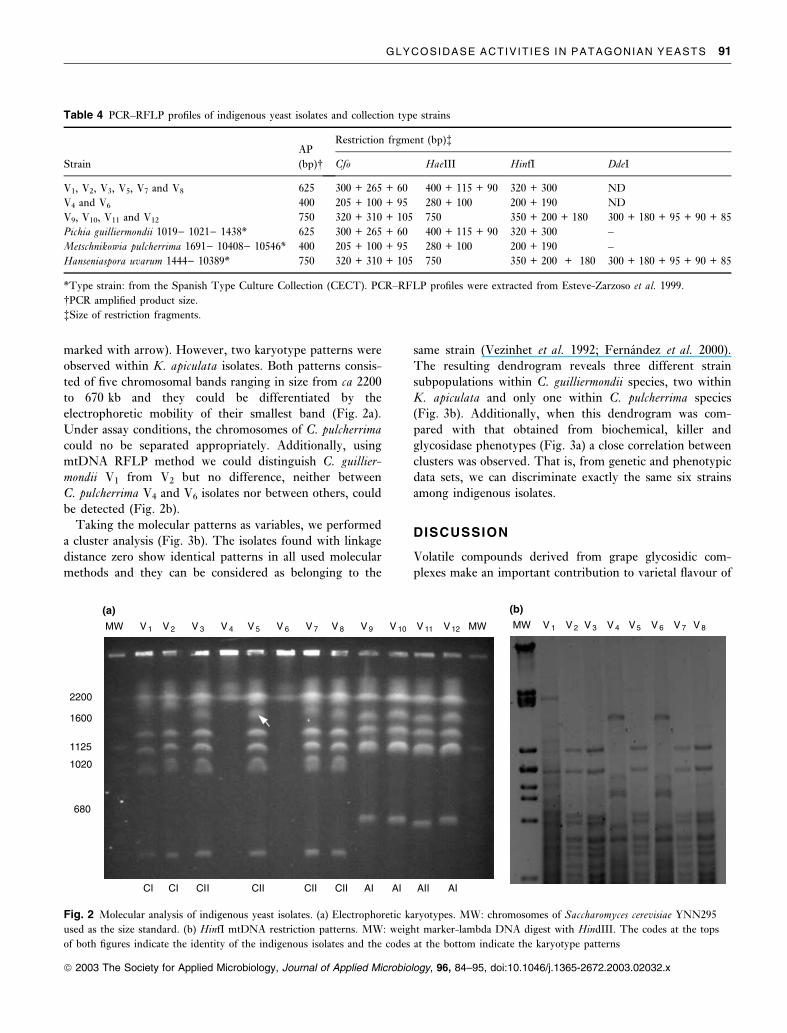
marked with arrow). However, two karyotype patterns were
observed within K. apiculata isolates. Both patterns consis-
ted of five chromosomal bands ranging in size from ca 2200
to 670 kb and they could be differentiated by the
electrophoretic mobility of their smallest band (Fig. 2a).
Under assay conditions, the chromosomes of C. pulcherrima
could no be separated appropriately. Additionally, using
mtDNA RFLP method we could distinguish C. guillier-mondii V1 from V2 but no difference, neither between
C. pulcherrima V4 and V6 isolates nor between others, could
be detected (Fig. 2b).
Taking the molecular patterns as variables, we performed
a cluster analysis (Fig. 3b). The isolates found with linkage
distance zero show identical patterns in all used molecular
methods and they can be considered as belonging to the
same strain (Vezinhet et al. 1992; Fernandez et al. 2000).
The resulting dendrogram reveals three different strain
subpopulations within C. guilliermondii species, two within
K. apiculata and only one within C. pulcherrima species
(Fig. 3b). Additionally, when this dendrogram was com-
pared with that obtained from biochemical, killer and
glycosidase phenotypes (Fig. 3a) a close correlation between
clusters was observed. That is, from genetic and phenotypic
data sets, we can discriminate exactly the same six strains
among indigenous isolates.
DISCUSSION
Volatile compounds derived from grape glycosidic com-
plexes make an important contribution to varietal flavour of
Table 4 PCR–RFLP profiles of indigenous yeast isolates and collection type strains
Strain
AP
(bp)�
Restriction frgment (bp)�
Cfo HaeIII HinfI DdeI
V1, V2, V3, V5, V7 and V8 625 300 + 265 + 60 400 + 115 + 90 320 + 300 ND
V4 and V6 400 205 + 100 + 95 280 + 100 200 + 190 ND
V9, V10, V11 and V12 750 320 + 310 + 105 750 350 + 200 + 180 300 + 180 + 95 + 90 + 85
Pichia guilliermondii 1019) 1021) 1438* 625 300 + 265 + 60 400 + 115 + 90 320 + 300 –
Metschnikowia pulcherrima 1691) 10408) 10546* 400 205 + 100 + 95 280 + 100 200 + 190 –
Hanseniaspora uvarum 1444) 10389* 750 320 + 310 + 105 750 350 + 200 + 180 300 + 180 + 95 + 90 + 85
*Type strain: from the Spanish Type Culture Collection (CECT). PCR–RFLP profiles were extracted from Esteve-Zarzoso et al. 1999.
�PCR amplified product size.
�Size of restriction fragments.
1020
2200
1125
1600
MW MWV 1 V 2 V 3 V 4 V 5 V 6 V 7 V 8 V 9 V 10 V 11 V 12
CI CI CII CII CII CII AI AI AII AI
(a)V 1 V 2 V 3 V 4 V5 V 6 V 7 V 8MW
(b)
680
Fig. 2 Molecular analysis of indigenous yeast isolates. (a) Electrophoretic karyotypes. MW: chromosomes of Saccharomyces cerevisiae YNN295
used as the size standard. (b) HinfI mtDNA restriction patterns. MW: weight marker-lambda DNA digest with HindIII. The codes at the tops
of both figures indicate the identity of the indigenous isolates and the codes at the bottom indicate the karyotype patterns
GLYCOSIDASE ACTIVIT IES IN PATAGONIAN YEASTS 91
ª 2003 The Society for Applied Microbiology, Journal of Applied Microbiology, 96, 84–95, doi:10.1046/j.1365-2672.2003.02032.x

the wine (Francis et al. 1992, 1999; Williams and Francis
1996; Gueguen et al. 1997). b-Glucosidase plays a key role
in this aroma enhancing process and this activity has been
extensively researched in wine yeast. Although some
activity in different strains of S. cerevisiae has been
described (Delcroix et al. 1994; Mateo and Di Stefano
1997; Hernandez et al. 2002), most studies demonstrate
that higher b-glucosidase producers are non-Saccharomycesspecies (Rosi et al. 1994; Miklosy and Polos 1995;
Charoenchai et al. 1997; Fernandez et al. 2000; Mendes
Ferreira et al. 2001; Strauss et al. 2001). Our screening
and preliminary in vitro assay results confirm these find-
ings. The best b-glucosidase activity producer were all
non-Saccharomyces yeasts belonging to C. guilliermondii,
C. pulcherrima and K. apiculata species (Table 1). This
yeast isolates were preselected and they were characterized
according to a set of selectable phenotypic characteristics
indicative of their potential usefulness in more aromatic
red wine production.
The first criterion to discriminate among strains included
the levels of specific b-glucosidase activity, its cellular
location and the effect of aeration conditions on them.
b-Glucosidase, by releasing aglicon from monoglycosidic
complexes, enhances the varietal aroma of wines. The
second criterion was the presence of the other glycosidases
activities involved in the first step of glycosidic complex
Linkage distance
V11
V12
V10
V9
V6
V4
V8
V7
V5
V3
V2
V1
0·00 0·05 0·10 0·15 0·20 0·25 0·30 0·35 0·40 0.45 0.50
C. guilliermondii
C. guilliermondii
C. guilliermondii
C. guilliermondii
C. guilliermondii
C. guilliermondii
C. pulcherrima
C. pulcherrima
K. apiculata
K. apiculata
K. apiculata
K. apiculata
(a)
Linkage distance
V11
V12
V10
V9
V6
V4
V8
V7
V5
V3
V2
V1
0·0 0·2 0·4 0·6 0·8 1·0 1·2
C. guilliermondii
C. guilliermondii
C. guilliermondii
C. guilliermondii
C. guilliermondii
C. guilliermondii
C. pulcherrima
C. pulcherrima
K. apiculata
K. apiculata
K. apiculata
K. apiculata
StrainI
StrainII
Strains III
Strains IV
Strains V
Strain VI
(b)
––
–
Fig. 3 UPGMA cluster analysis of selected yeast isolates based on phenotype (a) and molecular (b) characteristics
92 M.E. RODRIGUEZ ET AL.
ª 2003 The Society for Applied Microbiology, Journal of Applied Microbiology, 96, 84–95, doi:10.1046/j.1365-2672.2003.02032.x

sequential hydrolysis as well as b-xylosidase activity. There
are no data available about the potential of this activity to
improve aroma and flavour properties of wine. However,
b-xylosidase contribution to the formation of flavour and
colour in other foods obtained from fermentation industries
has been reported (Ohta et al. 1991; Kimura et al. 1999),
so we decided to include it. The third criterion was a
technological property, the killer behaviour of the isolates.
Non-Saccharomyces yeasts can be employed at the begin of
winemaking as mixed starter cultures in conjunction with
more ethanol-tolerant S. cerevisiae strains (Rainieri and
Pretorius 2000). To determine the killer behaviour is very
important because different killer interactions between
starters and between starters and wild yeast strains could
be established during fermentation affecting its normal
evolution. These interactions become relevant in areas
where killer yeasts are widespread such as the north
Patagonian region (Sangorrın et al. 2001). The fourth and
last, unfavourable criterion, was the presence of anthocy-
anase activity. Anthocyanins are phenolic molecules com-
posed of a glycosilated flavylium ion and they are
responsible for wine colour. Some b-glucosidase from
fungis and yeasts break the linkage between the glucose
and the anthocyanidin moieties inducing loss of wine
colour (Wightman et al. 1997; Sanchez-Torres et al. 1998;
Manzanares et al. 2000).
On the basis of these results, we characterized six
phenotypically different subpopulation among the selected
isolates including the weak isolate used as control
(Fig. 3a). Then, using molecular methods we confirmed
their taxonomical identity at species level (Fig. 1 and
Table 4) and their intraspecific variability (Figs 2 and 3b).
From the ecological point of view, these results showed a
great diversity of Patagonian indigenous glycosidase
producer strains (Tables 2 and 3), with significantly
different b-glucosidase profiles. These different glycosi-
dase profiles seem to be more closely related to strains
than to species. However, as far as is known, our study is
the first one that shows a correlation between molecular
polymorphism and specific phenotypic traits in wild
indigenous strains. In order to simplify the selection
process, the use of molecular methods before the appli-
cation of oenological criteria have been proposed to
discriminate among strains (Esteve-Zarzoso et al. 2000).
However, more than one molecular method could be used
so as not to miss any strain. With these considerations,
molecular polymorphism analysis may be very useful to
identify strains with specific phenotypic properties in
selection protocols.
From the technological point of view, some characteristics
of these Patagonian indigenous strains could justify their
utilization in industrial production of more aromatic red
wine. One of them is the high b-glucosidase activity levels
showed by these strains under anaerobic conditions, much
higher than those reported by other authors for yeast strains
belonging to these same species (Rosi et al. 1994; Fernandez
et al. 2000; Manzanares et al. 2000; Mendes Ferreira et al.
2001). Another characteristic is the lack of killer character
and their immunity to S. cerevisiae killer toxins. Although
S. cerevisiae killer toxins are only active against a few yeast
species including S. cerevisiae and C. glabrata, non-Sac-
charomyces killer toxins have a broader anti-yeast spectrum
including S. cerevisiae species. We have observed that
indigenous non-Saccharomyces killer yeasts resident in
Patagonian winery surfaces and initial musts could hinder
the implantation of commercial S. cerevisiae starters in
guided fermentation, inducing stuck fermentation
(I. Zajonskovsky, T.L. Lavalle, M.E. Rodrıguez, Ch. Lopes,
M. Sangorrın and A.C. Caballero 2002, personal commu-
nication). The last remarkable trait of these strains is the
presence of b-glucosidase activities lacking anthocyanase
activity and maintaining low but significant residual levels
after exposition to finished wine conditions during a
significant period of time.
Because some non-Saccharomyces yeasts produce undes-
irable concentrations of acetic acid and ethyl acetate from
sugars, they had always been regarded as unsuitable for
winemaking. The potential application of non-Saccharomy-
ces yeast strains in this process has been explored only fairly
recently. Due to their sensitivity to ethanol, non-Saccharo-myces yeast could be employed at the beginning of
winemaking as mixed cultures together with Saccharomyces
strains or in sequential non-Saccharomyces–Saccharomyces
inoculation protocols (Rainieri and Pretorius 2000; Soden
et al. 2000). However, the results obtained in this study
allow us to think in an additional and more advantageous
way of using these non-Saccharomyces strains towards the
end of fermentation, on the young wine.
ACKNOWLEDGEMENTS
This work was supported by the Comahue University Grant
(B091). The authors wish to thank S. Genoves and E. Ibanez
for help during the work in IATA and Ph.D. S. Bramardi
(U.N. Comahue) for his statistical assistance.
REFERENCES
Aryan, A.P., Wilson, B., Strauss, C.R. and Williams, P.J. (1987) The
properties of glycosidases of Vitis vinifera and comparison of their
b-glucosidase activity with that of exogenous enzymes. An assess-
ment of possible applications in enology. American Journal of Enology
and Viticulture 38, 182–188.
van Broock, M.R., Zajonskovsky, I.E., Assadourian, M.I., Lavalle,
T.L. and Caballero de Castro, A.C. (1996) Wine yeasts associated to
Merlot type grapes from north Patagonian region. An ecological
GLYCOSIDASE ACTIVIT IES IN PATAGONIAN YEASTS 93
ª 2003 The Society for Applied Microbiology, Journal of Applied Microbiology, 96, 84–95, doi:10.1046/j.1365-2672.2003.02032.x

study. In 10th International Biotechnology Symposium and 9th
International Symposium on Yeasts. pp. 13–15. Sydney, Australia.
Canal-Llauberes, R.M. (1993) Enzymes in winemaking. In Wine
Microbiology and Biotechnology ed. Fleet, G.H. pp. 477–506.
Switzerland: Harwood Academic Publishers.
Carle, G. F. and Olson M. V. (1985) An electrophoretic karyotype of
yeast. Proceedings of the National Academy of Sciences USA 82, 3756–
3760.
Charoenchai, C., Fleet, G.H., Henschke, P.A. and Todd, B.E.N.
(1997) Screening of non-Saccharomyces yeasts for the presence of
extracellular hydrolytic enzymes. Australian Journal of Grape and
Wine Research 3, 2–8.
Delcroix, A., Gunata Z., Sapis, J.C., Salmon, J.M. and Bayonove, C.
(1994) Glycosidase activities of the three enological yeast strains
during winemaking: effect on the terpenols content of Muscat wine.
American Journal of Enology and Viticulture 45, 291–296.
Esteve-Zarzoso, B., Belloch, C., Uruburu, F. and Querol, A. (1999)
Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and
two ribosomal internal transcribed spacers. International Journal of
Systematic Bacteriology 49, 329–337.
Esteve-Zarzoso, B., Gostıncar, A., Bobet, R., Uruburu, F. and Querol,
A. (2000) Selection and molecular characterization of wine yeasts
isolated from the ��El Penedes�� area (Spain). Food Microbiology 17,
553–562.
Fernandez, M., Ubeda, J.F. and Briones, A.I. (2000) Typing of
non-Saccharomyces yeasts with enzymatic activities of interest
in wine-making. International Journal of Food Microbiology 59,
29–36.
Fernandez-Espinar, M., Esteve-Zarzoso, B., Querol, A. and Barrio, E.
(2000) RFLP analysis of the ribosomal internal transcribed spacers
and the 5.8S rRNA gene region of the genus Saccharomyces: a fast
method for species identification and the differentiation of flor
yeasts. Antonie van Leeuwenhoeck 78, 87–97.
Francis, I.L., Sefton, M.A. and Williams, P.J. (1992) Sensory
descriptive analysis of hydrolyzed precursor fractions from Semillon,
Chardonnay, and Semillon Blanc grape juices. Journal of the Science
of Food and Agriculture 59, 511–520.
Francis, I.L., Kassara, S., Noble, A.C. and Williams, P.J. (1999) The
contribution of glycoside precursors to Cabernet Sauvignon and
Merlot aroma. Sensory and compositional studies. In Chemistry of
Wine Flavour, ACS Symposium Series 714, Chapter 12. ed.
Waterhouse A.L. and Ebeler S.E. pp. 13–30. Washington DC:
American Chemical Society.
Gueguen, Y., Chemardin, S.P., Arnaud, A. and Galzy, P. (1997)
Enhancement of aromatic quality of Muscat wine by the use of
immobilized b-glucosidase. Journal of Biotechnology 55, 151–156.
Gunata, Y.Z., Bayonove, C., Baumes, R. and Codornier, R.E. (1985)
The aroma of grapes. Extraction and determination of free and
glycosidically bound fractions of some grape aroma components.
Journal of Chromatography 331, 83–90.
Gunata, Z., Bitteur, S., Brillouet, J. Bayonove, C. and Cordonnier, R.
(1988) Sequential enzymatic hydrolysis of potentially aromatic
glycosides from grape. Carbohydrate Research 184, 139–149.
Gunata, Y.Z., Bayonove, C., Tapiero, C. and Cordonnier, R.E. (1990a)
Hydrolysis of grape monoterpenyl b-D-glucosides by various
b-glucosidases. Journal of Agricultural and Food Chemistry 38,
1232–1236.
Gunata, Y.Z., Bayonove, C., Arnaud, A. and Galzy, P. (1990b)
Hydrolysis of grape monoterpenyl glycosides by Candida molischiana
and Candida wickerhamii b-glucosidases. Journal of the Science of
Food and Agriculture 50, 499–506.
Hernandez, L.F., Espinoza, J.C., Fernandez-Gonzalez, M. and
Briones, A. (2002) b-Glucosidase activity in a Saccharomyces
cerevisiae wine strain. International Journal of Food Microbiology 80,
171–176.
Kimura, I., Yoshioka, N. and Tajima, S. (1999) Purification and
characterization of a b-glucosidase with b-xylosidase activity from
Aspergillus sajae. Journal of Bioscience and Bioengineering 87, 538–541.
Kreger-van Rij, N.J.W. (1984) The Yeasts, A Taxonomic Study, 3rd
edn. Amsterdam: Elsevier Science Publications.
Kurtzman, C.P. and Fell, J.W. (1998) The Yeast, a Taxonomic Study.
Amsterdam: Elseiver Science Publications.
Lambrechts, M.G. and Pretorius, I.S. (2000) Yeast and its importance
to wine aroma – a review. South African Journal of Enology and
Viticulture 21, 97–129.
Lopes, Ch. A. (1999) Aislamiento, cuantificacion y caracterizacion de la
flora de levaduras asociada a uvas regionales. Graduate thesis,
Universidad Nacional del Comahue, Argentina.
Lopes, C.A., van Broock, M., Querol, A. and Caballero, A.C. (2002)
Saccharomyces cerevisiae wine yeast populations in a cold region in
Argentinean Patagonia. A study at different fermentation scales.
Journal of Applied Microbiology 93, 608–615.
Lopez, V., Querol, A., Ramon, D. and Fernandez-Espinar, M.T.
(2001) A simplified procedure to analyse mitochondrial DNA from
industrial yeasts. International Journal of Food Microbiology 68,
75–81.
Manzanares, P., Ramon, D. and Querol, A. (1999) Screening of non-
Saccharomyces wine yeasts for the production of b-D-xylosidase
activity. International Journal of Food Microbiology 46, 105–112.
Manzanares, P., Rojas, V., Genoves, S. and Valles, S. (2000)
A preliminary search for anthocyanin-b-D-glucosidase activity in
non-Saccharomyces wine yeasts. International Journal of Food Science
and Technology 35, 95–103.
Mateo, J.J. and Di Stefano, R. (1997) Description of the b-glucosidase
activity of wine yeasts. Food Microbiology 14, 583–591.
Mendes Ferreira, A., Clımaco, M.C. and Mendes Faia, A. (2001) The
role of non-Saccharomyces species in releasing glycosidic bound
fraction of grape aroma components-a preliminary study. Journal of
Applied Microbiology 91, 67–71.
Miklosy, E. and Polos, V. (1995) Yeasts with b-glucosidase activity:
properties and possible application in winemaking processes. Acta
alimentaria 24, 167–180.
Nadal, D., Colomer, B. and Pina, B. (1996) Molecular polymorphism
distribution in phenotypically distinct populations of wine yeast
strains. Applied Environmental and Microbiology 62, 1944–1950.
Ohta, T., Omori, T., Shimoji, H., Hashimoto, K., Samuta, T. and
Ohba, T. (1991) Identification of monoterpenes alcohol b-glucoside
in sweet potatoes and purification of a Shiro-Koji b-glucosidase.
Agricultural Biological Chemistry 55, 1811–1816.
Querol, A., Barrio, E., Huerta, T. and Ramon, D. (1992) Molecular
monitoring of wine fermentations conducted by active dry yeast
strains. Applied and Environmental Microbiology 58, 2948–2953.
Rainieri, S. and Pretorius, I.S. (2000) Selection and improvement of
wine yeasts. Annals of Microbiology 50, 15–31.
94 M.E. RODRIGUEZ ET AL.
ª 2003 The Society for Applied Microbiology, Journal of Applied Microbiology, 96, 84–95, doi:10.1046/j.1365-2672.2003.02032.x

Rapp, A. (1998) Volatile flavour of wine. Correlation between
instrumental analysis and sensory perception. Nahrung 42, 351–363.
Rapp, A. and Versini, G. (1991) Influence of nitrogen compounds in
grapes on aroma compounds of wine. In Proceedings of the International
Symposium on Nitrogen in Grapes and Wine ed. Rantz, J.M. pp. 156–
164. Davis, CA: American Society for Enology and Viticulture.
Riccio, P., Rossano, R., Vinella, M., Domizio, P., Zito, F., Sanseverino,
F., D’Elia, A. and Rossi, I. (1999) Extraction and immobilization in
one sep of two b-glucosidases released from a yeast strain of
Debaryomyces hansenii. Enzyme and Microbial Technology 24, 123–129.
Riou, Ch., Salmon, J.M., Vallier, M.J., Gunata, Z. and Barre, P. (1998)
Purification, characterization, and substrate specificity of a novel
highly glucose-tolerant b-glucosidase from Aspergillus oryzae.
Applied and Environmental Microbiology 64, 3607–3614.
Rodrıguez, M.E. (1999) Flora de levaduras y caracterısticas enologicas
de mostos fermentados espontaneamente en ambientes de bodega y
de laboratorio. Graduate thesis, Universidad Nacional del Comahue,
Argentina.
Rogerson, F.S., Grande, H.J. and Silva, M.C.M. (1995) Enzymatic
enhancement of the free monoterpenol content of Portuguese wine
from a single, native grape variety, ��Trajadura��. Biotechnology Letters
17, 35–40.
Rosi, I., Vinella, M. and Domizio, P. (1994) Characterization of b-
glucosidase activity in yeasts of oenological origin. Journal of Appllied
Bacteriology 77, 519–527.
Salmon, J.M. (1984) Application of the technique of cellular perme-
abilization to the study of the enzymatic activities of Saccharomyces
cerevisiae in continuous alcoholic fermentation. Biotechnology Letters
6, 43–48.
Sanchez-Torres, P., Gonzalez-Candelas, L. and Ramon, D. (1998)
Heterologous expression of a Candida molischiana anthocyanin-b-
glucosidase in a wine yeast strain. Journal of Agricultural and Food
Chemistry 46, 354–360.
Sangorrın, M., Zajonskovsky, I., Lopes, Ch., Rodrıguez, M., van
Broock, M. and Caballero A. (2001) Killer behaviour in wild wine
yeasts associated with Merlot and Malbec type musts spontaneously
fermented from Northwestern Patagonia (Argentina). Journal of
Basic Microbiology 41, 105–113.
Sangorrın, M., Zajonskovsky, I., van Broock, M. and Caballero, A.
(2002) The use of killer biotyping in an ecological survey of yeast in
an old Patagonian winery. World Journal Microbiology and Biotech-
nology 18, 115–120.
Schreier, P. (1979) Flavour composition of wines: a review. CRC
Critical Review in Food Science and Nutrition 12, 59–111.
Schutz, M. and Gafner, J. (1993) Analysis of yeast diversity during
spontaneous and induced alcoholic fermentations. Journal of Applied
Bacteriology 75, 551–558.
Soden, A., Francis, I.L., Oakey, H. and Henschke, P.A. (2000) Effects
of co-fermentation with Candida stellata and Saccharomyces cerevisiae
on aroma and composition of Chardonnay wine. Australian Journal of
Grape and Wine Research 6, 21–30.
Spagna, G., Barbagallo, R.N., Palmeri, R., Restuccia, C. and Giudici,
P. (2002) Properties of endogenous b-glucosidase of a Pichia anomala
strain isolated from Sicilian musts and wines. Enzyme and Microbial
Technology 31, 1036–1041.
Strauss, M.L., Jolly, N.P., Lambrechts, M.G. and van Rensburg, P.
(2001) Screening for the production of extracellular hydrolytic
enzymes by non-Saccharomyces wine yeasts. Applied Microbiology 91,
182–190.
Vasserot, Y., Chemardin, P., Arnaud, A. and Galzy, P. (1991)
Purification and properties of Candida molischiana able to work at
low pH values: possible use in the liberation of bound terpenols.
Journal Basic Microbiology 31, 301–312.
Vasserot, Y., Arnaud, A. and Galzy, P. (1995) Monoterpenyl glycosides
in plants and their biotechnological transformation. Acta Biotechno-
logy 15, 77–95.
Vezinhet, F., Hallet, J.N., Valade, M. and Poulard, A. (1992)
Ecological survey of wine yeast strains by molecular methods of
identification. American Journal of Enology and Viticulture 43, 83–86.
White, T.J., Bruns, T., Lee, S. and Taylor, J. (1990). PCR protocols. A
guide to methods and applications. In Amplification and Direct
Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics ed.
Innis, M.A., Gelfand, D.H., Sninsky, J.J. and With, T.J. pp. 315–
322. San Diego: Academic Press.
Wightman, J.D., Price, S.F., Watson, B.T. and Wrolstad, R.E. (1997)
Some effects of processing enzymes on anthocyanino and phenolics
in Pinot noir and Cabernet Sauvignon wines. American Journal of
Enology and Viticulture 48, 39–48.
Williams, P.J. and Francis, L. (1996) Sensory analysis and quantitative
determination of grape glycosides. The contribution of these data to
winemaking and viticulture. In Biotechnology for Improved Foods and
Flavors, ACS Symposium Series 637, ed. Takcoka, G.R., Teranishi,
R., Williams, P.I. and Kobayashi, A. Washington DC: American
Chemical Society.
Williams, P.J., Sefton, M.A. and Wilson, B. (1989) Nonvolatile
conjugates of secondary metabolites as precursors of varietal grape
flavor components. In Flavor Chemistry: Trends and Developments ed.
Teranishi R., Buttery R. and Shahidi R. pp. 35–48. Washington DC:
American Chemical Society.
Winterhalter, P. and Skouroumounis, G. (1997) Glycoconjugated
aroma compounds: occurrence, role and biotechnological transfor-
mation. In Advances in Biochemical Engineering and Biotechnology,
vol. 55, ed. Scheper, T. pp. 73–105. Berlin, Heidelberg, Germany:
Springer-Verlag.
GLYCOSIDASE ACTIVIT IES IN PATAGONIAN YEASTS 95
ª 2003 The Society for Applied Microbiology, Journal of Applied Microbiology, 96, 84–95, doi:10.1046/j.1365-2672.2003.02032.x





























