Embed Size (px)
Citation preview

May 27, 2009 19:38 WSPC/178-JTCC 00487
Journal of Theoretical and Computational ChemistryVol. 8, No. 3 (2009) 529–539c© World Scientific Publishing Company
ROLE OF ADENINE AND GUANINE SITES INHOLE HOPPING IN DNA NANOWIRE
INDERPREET KAUR∗,§, GIRISH S. KULKARNI∗, RAM AJORE∗,RICHA BHARADWAJ∗, BHANU PRAKASH KOTAMARTHI†,
NIMAL SINGH‡ and LALIT M. BHARADWAJ∗,¶
∗Biomolecular Electronics and Nanotechnology Division (BEND),Central Scientific Instruments Organization (CSIO),
Sector-30C, Chandigarh, India
†Indian Institute of Chemical Technology, Hyderabad, India
‡Department of Physics, Panjab University,Sector-14, Chandigarh, India¶[email protected]
Received 24 June 2008Accepted 29 August 2008
Transfer integrals for oligos with different bases have been calculated using INDO/Koopman’s approximation to unveil the charge transport mechanism in DNA. Thesequences, G(A)nG, n = 1, 2, . . . , 10; G(A)xG(A)yG, x + y = 9; and G(A)xG(A)y
G(A)zG, x + y + z = 8, were employed to interpret the Guanine (G) and Adenine(A)hopping. Adenine hopping is found to be faster in G(A)nG sequences with longer Ade-nine bridges (n ≥ 3). Inserting G-bases in between G(A)10G led to a decrease in thevalue of transfer integrals. Close analysis has revealed that bridge closer to 3′-end formsa hopping bottleneck; however, the presence of bridge at 5′-end enhances the chargetransfer through A-hopping. Further insertion of single G sites in G(A)xG(A)yG (wherex + y = 9) reduces the transfer integrals, thus explaining the hampering of A-hopping.Hence, sequences of the type G(A)nG, n > 3, are better suited for their application asmolecular wire. Finally, studies on the effect of flipping of bases, i.e. flipping G:C to C:Gon transfer integrals, have revealed that helical distortions and conformational changesdue to sequence variations lead to changes in coupling, which is highly unpredictable.
Keywords: DNA nanowire; coupling parameters; molecular electronics; HOMO–LUMO;population analysis.
1. Introduction
The ideal structure and molecular recognition properties of DNA make it a suit-able contender for fulfiling the goal of self-assembling nanodevices with a defi-nite molecular architecture.1–3 Sequence variability,4 nonperiodicity, and structuraldynamicity5–10 of DNA have given experimentalists very controversial observations,e.g. researchers have reported conductive11–17 and others reported insulating18–21
529

May 27, 2009 19:38 WSPC/178-JTCC 00487
530 I. Kaur et al.
behavior on charge transport that made theoreticians to propose number of trans-port mechanisms,22–27 which are still not complete to explain the charge transportin DNA. DNA base sequence, conformational dynamics, and local flexibility allcontribute to coupling within the base pair π-stacked array and to the efficiencyof the DNA-mediated charge transport reaction. Therefore, theoretical approacheswere employed so that base-by-base feasibility of charge transport in DNA could beexplained. The aim of the present work is to reveal the transfer integrals, which aredirectly related to coupling parameters, to understand effect of sequence variationand helical changes due to base pair arrangements on the charge transport in DNAbases.
Previous theoretical studies on DNA base pair coupling analysis to pre-dict sequence variations are mostly on double and triple stacks28,29 and theirconformational distortions like slide “S,” twist “T,” and roll “R”.30–32 Here, tak-ing longer sequences, up to 12 bases, assuming that π-stacking is playing a dom-inant role in charge transfer (CT) in DNA33,34 and hopping mechanism, eitherdomain35–37 or multistep,38 is a long-range transport mechanism. Therefore, back-bone is being neglected to study aromatic interactions in π-stacks of base pairs.Since G-hopping38,39 and A-hopping40 are reported many times in literature; hence,sequences with intrastrand G and A combination were taken. Although enhancedefficiency of Thymine (T) relative to A as mediator between guanine in G+XY · · ·G (X, Y = T and A) has been reported by Voityuk et al.41 but this particularly fea-tures “super exchange induced hole hopping” mechanism, which is not the interestof present work.
Theoretical modeling of large molecules particularly π-stacking molecules suchas DNA is an extremely difficult task. Voityuk42 has done a significant analysisof quantum mechanical methods predicting that the semi-empirical method/INDOwith Koopman’s approximation is most reliable and computationally very efficientfor relative/comparative study of electronic coupling. Among all the semi-empiricalmethods/approximations, e.g. MNDO/3, INDO, AM1, PM3 and Koopman/INDOprovide good estimates of the CT parameters. While the MNDO, AM1, and PM3methods give reasonable CT energies, they strongly underestimate the couplingsmatrix elements, and therefore, cannot be applied to model CT in π-stacks. There-fore, INDO method with Koopman’s approximation was used to calculate couplingparameters for sequences of the type G(A)nG, n = 1, 2, . . . , 10; G(A)xG(A)yG,x + y = 9; and G(A)xG(A)yG(A)zG, x + y + z = 8. Analysis on G(A)nG,n = 1, 2, . . . , 10 confirms that A is also participating in long-range charge transport.The present study is also in accordance with the experimental observation reportedby Giese et al.43–45 It is further elaborated by taking G(A)10G sequence by addingone guanine site making it G(A)xG(A)yG; x+ y = 9 and two guanine sites formingsequence G(A)xG(A)yG(A)zG; x + y + z = 8 that with the increasing number ofG sites electronic coupling decreases.46 While studying G(A)xG(A)yG, x + y = 9sequences, another very interesting observation is made that A:T bridge lengths at5′-end and 3′-end may also affect transfer integrals.

May 27, 2009 19:38 WSPC/178-JTCC 00487
Role of Adenine and Guanine Sites in Hole Hopping in DNA Nanowire 531
Furthermore, a systematic study suggested that for different sequences of samelength, DNA helix is not the same, which can be attributed to the differentpotentials of base pairs. Even a flip of base pair, i.e. G:C flipped to C:G inthe same sequence, can show an effective change in transport. These helicalchanges/distortions are analyzed by flipping in the sequences G(A)xG(A)yG andG(A)xG(A)yG(A)zG where G:C is flipped to C:G. This revealed that flipping some-times lead to drastic changes; on the other hand, it may lead to almost negligiblevariation in the transfer integrals. These flips may increase or decrease couplingdepending on whether the helical distortions caused by the flips tend to favor orhamper charge migration. The distortions in terms of transfer integrals have alsobeen studied by Voityuk et al.,32 by taking AA stack with all possible conforma-tional combinations by twisting, sliding, and rolling the base pairs they suggestedthat electronic coupling matrix elements may change considerably thereby makinghole donors and acceptors in resonance or out of resonance for CT.47
2. Methodology
To examine the various combinations of neutral base pairs to understand thesequence dependence of CT in DNA, fragments of stacked pairs of nucleobasesare considered as mediators of CT; however, the sugar–phosphate backbone isneglected. This approximation seems to be quite adequate since π-stacking interac-tions of nucleobases play the dominant role for the hole transfer in DNA.33,34 Hereabbreviated notations have been used; for instance, GA denotes a fragment thatconsists of two Watson crick pairs (GC) and (AT) in the standard order 5′ → 3′.All these structures are constructed with the program HYPERCHEM using thestep parameters of regular B-DNA (rise, 3.38 A; twist, 36◦). To study the aromaticinteractions (transfer integrals) between the adjacent base pairs, semi-empiricalintermediate neglect of differential overlap (INDO) Hamiltonian calculations wereperformed. Transfer integrals, t, are calculated using Koopman’s approximation,and estimated splitting t = E2 −E1 of the cation radical states as the difference ofthe one-electron energies of the (HOMO and HOMO−1) orbitals calculated for thecorresponding neutral system:
t = EHOMO − EHOMO − 1 for hole transfer .
INDO calculations were done using GAUSSIAN 03W.
3. Results and Discussion
3.1. Effect of increasing the number of A:T pairs intervening
G:C pairs
Giese et al.4 initially proposed that charge transport occurs in DNA by hoppingthrough guanine bases and tunneling through intervening A:T pairs. It was alsoreported that the increasing number of A:T base pairs led to decrease in cou-pling between the guanine bases.4 The experimental study of Boon and Barton48
May 27, 2009 19:38 WSPC/178-JTCC 00487
532 I. Kaur et al.
Table 1. Transfer integrals for G(A)nG, where n = 1, 2, . . . , 10.
Sequence HOMO−1 HOMO HOMO−(HOMO−1)
GAG −8.539 −8.488 0.051G(A)2G −8.564 −8.465 0.099G(A)3G −8.572 −8.455 0.117G(A)4G −8.575 −8.451 0.125G(A)5G −8.577 −8.448 0.128G(A)6G −8.577 −8.446 0.131G(A)7G −8.577 −8.445 0.133G(A)8G −8.577 −8.443 0.133G(A)9G −8.576 −8.442 0.134G(A)10G −8.576 −8.441 0.135G(A)12G −8.577 −8.441 0.136
0 2 4 6 8 100.04
0.06
0.08
0.10
0.12
0.14
Tra
nsf
er
Inte
gra
l [H
-(H
-1)]
eV
n[G:C(A:T)
nG:C]
Hole Transport [HOMO-(HOMO-1)]
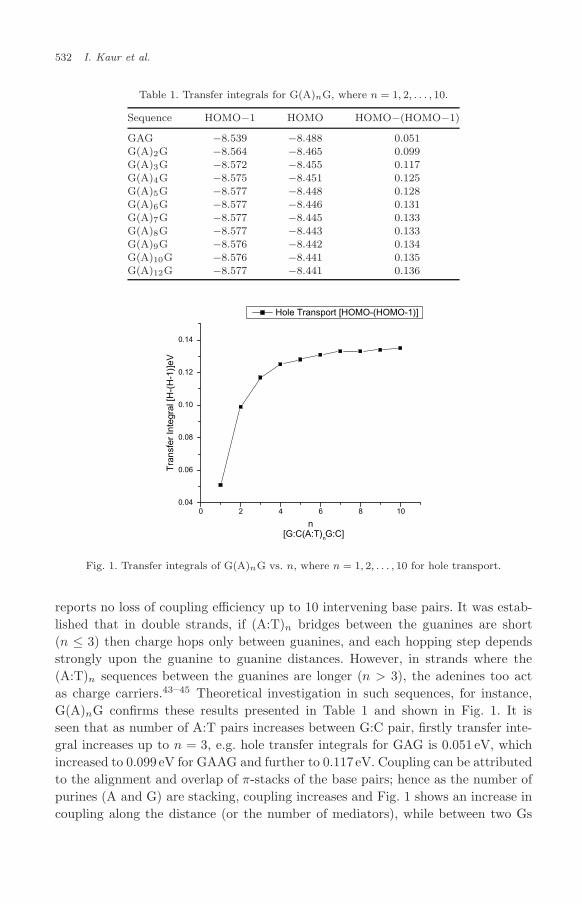
Fig. 1. Transfer integrals of G(A)nG vs. n, where n = 1, 2, . . . , 10 for hole transport.
reports no loss of coupling efficiency up to 10 intervening base pairs. It was estab-lished that in double strands, if (A:T)n bridges between the guanines are short(n ≤ 3) then charge hops only between guanines, and each hopping step dependsstrongly upon the guanine to guanine distances. However, in strands where the(A:T)n sequences between the guanines are longer (n > 3), the adenines too actas charge carriers.43–45 Theoretical investigation in such sequences, for instance,G(A)nG confirms these results presented in Table 1 and shown in Fig. 1. It isseen that as number of A:T pairs increases between G:C pair, firstly transfer inte-gral increases up to n = 3, e.g. hole transfer integrals for GAG is 0.051 eV, whichincreased to 0.099 eV for GAAG and further to 0.117 eV. Coupling can be attributedto the alignment and overlap of π-stacks of the base pairs; hence as the number ofpurines (A and G) are stacking, coupling increases and Fig. 1 shows an increase incoupling along the distance (or the number of mediators), while between two Gs

May 27, 2009 19:38 WSPC/178-JTCC 00487
Role of Adenine and Guanine Sites in Hole Hopping in DNA Nanowire 533
the coupling decreases exponentially henceforth leading to decrease in hole trans-port for G(A)nG where n = 1,2,3 thereby emphasizing the hole hopping is boundto take place on G-sites only. On adding one more Adenine in between Gs aftern > 3 change is not appreciable (Fig. 1), hence showing that A can play a role ofintermediate hole carriers.46 It was further reported that the oxidation of A by G·+
is endothermic.46 With respect to the low efficiency of this hole-hopping step, it wassuggested that once A·+ has been generated, A-hopping proceeds faster. In fact,the rate of A-hopping49kHT has been determined to be 1010 s−1, while the rate fora single hopping50 step from G to GG kHOP was reported to be 106–108 s−1.51 Incomparison to G-hopping, A-hopping proceeds faster, more efficiently, and almostdistance independently. Recent calculations suggest that hole transport throughstacked A-T base pairs may be most favorable.52
This long-range hopping was further explained by taking orbital plots of HOMOand HOMO−1, since two adiabatic states that are involved in hole transfer canbe represented by a linear combination of HOMO and HOMO−1 orbitals of thecorresponding neutral system (Koopman’s approximation). Therefore, one canrationalize the coupling between two bases in terms of the overlap between thecorresponding orbitals. In Figs. 2 and 3, HOMO−1 is lying on left G, i.e. 3′-end,
Fig. 2. Orbital plots for HOMO − 1 and HOMO for G(A)4G.
Fig. 3. Orbital plots for HOMO − 1 and HOMO for G(A)10G.

May 27, 2009 19:38 WSPC/178-JTCC 00487
534 I. Kaur et al.
and HOMO is lying on right G, i.e. 5′-end, indicating that hole is moved from leftG (3′-end) to right G (5′-end), thus long-range CT is possible in A-bridges. Negli-gible delocalization of HOMO and HOMO−1 also supports that hole hops throughspace. To strengthen this fact, the transfer integral for G(A)12G was also calculatedand found an approximate value to G(A)10G.
3.2. Introducing G sites in between G(A)10G
(a) When one G-site is introduced: Next, A single guanine site between the inter-vening A:T pairs was introduced in order to study its effect on CT integrals.Keeping the intervening base pairs to a limit of 10, we obtained the resultsshown in Table 2 for all possible combinations. It is clear that transfer integralvalues decrease gradually from G(A)10G = 0.135 to 0 eV for the combinationG(A)1G(A)8G. Further, results revealed that charge transport efficiency is morewhen the A:T bridge is closer to 5′-end than the 3′-end.
These results are in accordance with earlier reported data,53 which showthat HOMO has a tendency to concentrate on 5′-end.53 Table 2 also reflectsthat as the A:T bridge gets longer toward 3′-end it dampens up the CT, posinga hopping bottleneck,38 for example, 5′-G(A)5G(A)4G-3′ has almost doublecoupling than 5′-G(A)4G(A)5G-3′. Similarly when bridge of (A)8 comes at 3′-end in G(A)1G(A)8G it decreases the transfer integral to zero while reversingthe bridge, for instance, G(A)8G(A)1G, coupling increases sharply to 0.101 eV.Similar trends are observed in all other possible combinations (Table 2).
(b) Introducing two G-sites: A number of combinations were tried for introduc-ing second G-site to the above sequences. The addition of another guaninesite would lead to further reduction in coupling thereby reducing A-hopping.Such A-hopping can occur if G is not present within the sequential context.46
Although the coupling decreases, symmetrical insertion of G in the sequencesenhances the coupling value, for example, results for combination X :Y :Z =3:3:2 has higher coupling than 4:2:2, 2:4:2, and 4:3:1. Such sequences supportG-hopping. Second, bridge length, on the other hand, longer than that of threeA:T base pairs towards 5′-end can enhance A-hopping. It can increase the valuefor transfer integrals, provided the central region, which may form a bottle neck,should not be large. Hence, it can be considered that G should not be present
Table 2. Transfer integrals for G(A)xG(A)yG, where x + y = 9.
5′-G(A)xG(A)yG-3′ 5′-G(A)xG(A)yG-3′x + y = 9; x > 5 HOMO−(HOMO−1) x + y = 9; y > 5 HOMO−(HOMO−1)
G(A)8G(A)1G 0.101 G(A)1G(A)8G 0.000G(A)7G(A)2G 0.063 G(A)2G(A)7G 0.022G(A)6G(A)3G 0.079 G(A)3G(A)6G 0.061G(A)5G(A)4G 0.071 G(A)4G(A)5G 0.037

May 27, 2009 19:38 WSPC/178-JTCC 00487
Role of Adenine and Guanine Sites in Hole Hopping in DNA Nanowire 535
in sequence in sequential context to get long-range A-hopping. Otherwise, G-hopping dominates over A-hopping and sometimes could lead to delocalizationof charge on the sequence.
3.3. Flipping the bases
Sequence-dependent study was done, wherein intermediate bases were flipped ofsome of the combinations studied in Tables 2 and 3.
The transfer integrals do not show regular pattern that flipping the bases insome cases increases the transfer integral and in some cases decreases it. Theseflippings of bases can be related to helical distortions and hence to relative transferintegral variation among stacks of bases in DNA. When G:C flipped to C:G in thesame sequence (Table 4). It can be attributed to realignment of helical structure andnearing of G bases due to helical distortions in DNA, which can promote or demoteconductance. Structural distortions in DNA are of many types and very complex.Here it is assumed that flipping of bases can cause helical distortions in terms ofrotational or translational changes, which may bring hole donors and acceptors intoresonance or out of resonance32 hence allowing the hole to adopt the zigzag pathwaymechanism or inter-strand hopping of hole. Second, such flips can enhance or reduce“through space” hopping such as G-hopping by causing the hopping sites to comecloser or moving them away. However, if hopping is considered through bond then
Table 3. Transfer integrals for G(A)xG(A)yG(A)zG , where x + y + z = 8.
5′-G(A)xG(A)y G(A)zG-3′;x + y + z = 8 HOMO−1 HOMO H−(H − 1)
G(A)5G(A)2G(A)1G −8.392 −8.368 0.024G(A)5G(A)1G(A)2G −8.405 −8.362 0.043G(A)2G(A)1G(A)5G −8.424 −8.381 0.042G(A)4G(A)3G(A)1G −8.402 −8.363 0.039G(A)4G(A)1G(A)3G −8.414 −8.366 0.048G(A)4G(A)2G(A)2G −8.406 −8.400 0.007G(A)2G(A)4G(A)2G −8.428 −8.409 0.019G(A)3G(A)3G(A)2G −8.407 −8.360 0.047
Table 4. Transfer integrals for DNA sequences after flipping middle andend bases.
Sequence HOMO−1 HOMO H − (H − 1)
G(A)5C(A)4G −8.439 −8.401 0.038G(A)5C(A)4C −8.510 −8.400 0.110G(A)3C(A)3G(A)2G −8.435 −8.353 0.081G(A)3G(A)3C(A)2G −8.426 −8.367 0.059G(A)3C(A)3C(A)2G −8.435 −8.401 0.034G(A)5G(A)4C −8.503 −8.361 0.142
May 27, 2009 19:38 WSPC/178-JTCC 00487
536 I. Kaur et al.
increase in transfer integrals in some cases such as G(A)5C(A)4C has almost doublevalue than G(A)4C(A)5C, which can be due to break in electron channel that isformed by purines in intra-strand. Hence, flip will reduce the intrastrand π-stackingentrapment region, thereby reducing the hole entrapment probability. This type ofhelical distortion can be related to polaron like species — a local distortion of theDNA structure where a long π-stacking of purines is delocalized along the lengthand has a tendency to entrap a hole.10
3.4. Alternate sequences
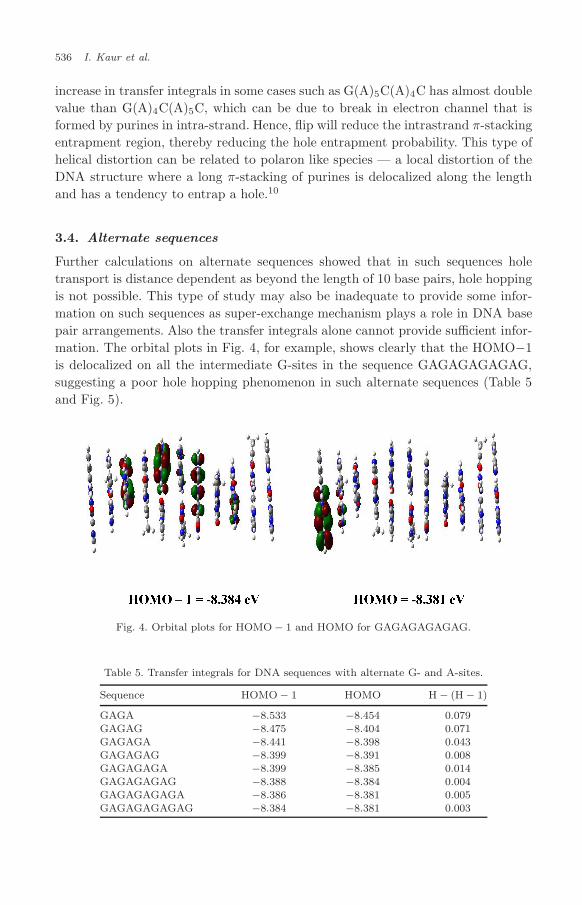
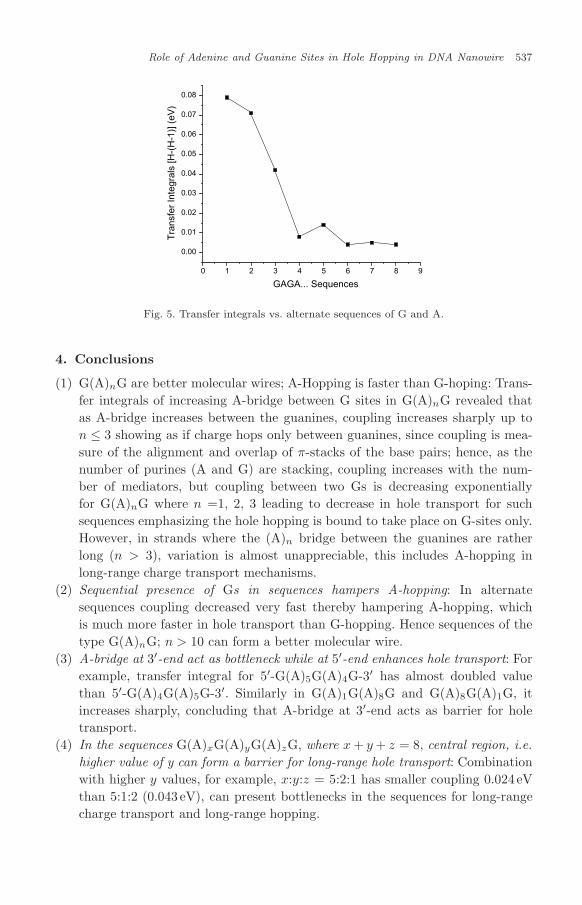
Further calculations on alternate sequences showed that in such sequences holetransport is distance dependent as beyond the length of 10 base pairs, hole hoppingis not possible. This type of study may also be inadequate to provide some infor-mation on such sequences as super-exchange mechanism plays a role in DNA basepair arrangements. Also the transfer integrals alone cannot provide sufficient infor-mation. The orbital plots in Fig. 4, for example, shows clearly that the HOMO−1is delocalized on all the intermediate G-sites in the sequence GAGAGAGAGAG,suggesting a poor hole hopping phenomenon in such alternate sequences (Table 5and Fig. 5).
Fig. 4. Orbital plots for HOMO − 1 and HOMO for GAGAGAGAGAG.
Table 5. Transfer integrals for DNA sequences with alternate G- and A-sites.
Sequence HOMO − 1 HOMO H − (H − 1)
GAGA −8.533 −8.454 0.079GAGAG −8.475 −8.404 0.071GAGAGA −8.441 −8.398 0.043GAGAGAG −8.399 −8.391 0.008GAGAGAGA −8.399 −8.385 0.014GAGAGAGAG −8.388 −8.384 0.004GAGAGAGAGA −8.386 −8.381 0.005GAGAGAGAGAG −8.384 −8.381 0.003
May 27, 2009 19:38 WSPC/178-JTCC 00487
Role of Adenine and Guanine Sites in Hole Hopping in DNA Nanowire 537
0 1 2 3 4 5 6 7 8 9
0.00
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
Tra
nsf
er
Inte
gra
ls [H
-(H
-1)]
(eV
)
GAGA... Sequences
Fig. 5. Transfer integrals vs. alternate sequences of G and A.
4. Conclusions
(1) G(A)nG are better molecular wires; A-Hopping is faster than G-hoping: Trans-fer integrals of increasing A-bridge between G sites in G(A)nG revealed thatas A-bridge increases between the guanines, coupling increases sharply up ton ≤ 3 showing as if charge hops only between guanines, since coupling is mea-sure of the alignment and overlap of π-stacks of the base pairs; hence, as thenumber of purines (A and G) are stacking, coupling increases with the num-ber of mediators, but coupling between two Gs is decreasing exponentiallyfor G(A)nG where n =1, 2, 3 leading to decrease in hole transport for suchsequences emphasizing the hole hopping is bound to take place on G-sites only.However, in strands where the (A)n bridge between the guanines are ratherlong (n > 3), variation is almost unappreciable, this includes A-hopping inlong-range charge transport mechanisms.
(2) Sequential presence of Gs in sequences hampers A-hopping: In alternatesequences coupling decreased very fast thereby hampering A-hopping, whichis much more faster in hole transport than G-hopping. Hence sequences of thetype G(A)nG; n > 10 can form a better molecular wire.
(3) A-bridge at 3′-end act as bottleneck while at 5′-end enhances hole transport: Forexample, transfer integral for 5′-G(A)5G(A)4G-3′ has almost doubled valuethan 5′-G(A)4G(A)5G-3′. Similarly in G(A)1G(A)8G and G(A)8G(A)1G, itincreases sharply, concluding that A-bridge at 3′-end acts as barrier for holetransport.
(4) In the sequences G(A)xG(A)yG(A)zG, where x + y + z = 8, central region, i.e.higher value of y can form a barrier for long-range hole transport: Combinationwith higher y values, for example, x:y:z = 5:2:1 has smaller coupling 0.024 eVthan 5:1:2 (0.043 eV), can present bottlenecks in the sequences for long-rangecharge transport and long-range hopping.

May 27, 2009 19:38 WSPC/178-JTCC 00487
538 I. Kaur et al.
(5) By flipping the base pairs, if possible, DNA conductivity can be improved: Thiscan be attributed to realignment of helical structure and nearing of G bases dueto helical distortions in DNA, which can either promote or demote conductance;either allow the hole to adopt the zigzag pathway mechanism or inter-strandhopping of hole; or can reduce a local distortion of the DNA structure where along π-stacking of purines is delocalized along the length and have a tendencyto entrap a hole.
Acknowledgments
This research is supported by Department of Bio Technology (DBT), India andDepartment of Science and Technology (DST), India. One of the authors, Inder-preet, acknowledges the support of the Council of Scientific and Industrial Research(CSIR), India for Doctoral education on CSIR-Junior Research Fellowship. Theauthors also acknowledge other group members at CSIO, for their valuable sugges-tions and inputs.
References
1. Mao C et al., Nature (London) 407:493, 2000.2. Yurke B et al., Nature 406:605, 2000.3. Winfree E et al., Nature 394:539, 1998.4. Meggers E, Michel-Beyerle ME, Giese B, J Am Chem Soc 120:12950, 1998.5. Brauns EB, Murphy CJ, Berg MA, J Am Chem Soc 120:2449, 1998.6. Borer PN, Pante SR, Kumar A, Zanatta N, Martin A, Hakkinen A, Levy GC, Biochem
33:2441, 1994.7. Kojima C, Ono A, Kainosho M, James TL, J Magn Reson 135:1333, 1998.8. Georghiou S, Bradrick TD, Philippetis A, Beechem JM, Biopys J 70:1909, 1996.9. Sriram M, Wang AHJ, in Hecht SM (ed.), Oxford University Press, New York, 1996,
pp. 105–143.10. Ly D, Sanii L, Schuster GB, J Am Chem Soc 121:9400, 1999.11. Kasumov Y, Kociak M, Gueron S, Reulet B, Volkov VT, Klinov DV, Bouchiat H,
Science 291:240, 2001.12. Porath D, Cuniberti G, Di Felice R, Top Curr Chem 237:183, 2004.13. Porath D, Bezryadin A, de Vries S, Dekker C, Nature 403:635, 2000.14. Hwang JS, Lee GS, Ahn D, Hwang SW, Appl Phys Lett 81:1134, 2002.15. Shigematsu T, Shimotani K, Manabe C, Watanabe H, Shimizu M, J Chem Phys
118:4245, 2003.16. Fink HW, Schonenberger C, Nature 398:407, 1999.17. Watanabe H, Manabe C, Shigematsu T, Shimotani K, Shimizu M, Appl Phys Lett
79:2462, 2001.18. Braun E, Eichen Y, Sivan U, Ben-Yoseph G, Nature 391:775, 1998.19. de Pablo PJ, Moreno-Herrero F, Colchero J, Gomez Herrero J, Herrero P, Baro AM,
Ordejon P, Soler JM, Artacho E, Phys Rev Lett 85:4992, 2000.20. Storm AJ, van Noort J, de Vries S, Dekker C, Appl Phys Lett 79:3881, 2001.21. Zhang Y, Austin RH, Kraeft J, Cox EC, Ong NP, Phys Rev Lett 89:198102, 2002.22. Murphy CJ, Arkin MA, Jenkins Y, Ghatlia ND, Bossman S, Turro NJ, Barton JK,
Science 262:1025, 1993.

May 27, 2009 19:38 WSPC/178-JTCC 00487
Role of Adenine and Guanine Sites in Hole Hopping in DNA Nanowire 539
23. Bixon M, Giese B, Wessely S, Langenbacher T, Michel-Beyerle ME, Jortner J, ProcNat Acad Sci USA 96:11713, 1999.
24. Conwell EM, Rakhmanova SV, Proc Nat Acad Sci USA 97:4556, 2000.25. Bixon M, Jortner J, J Am Chem Soc 123:12556, 2001.26. Jortner J, Bixon M, Langenbacher T, Michel-Beyerle ME, Proc Nat Acad Sci USA
95:12759, 1998.27. Bixon M, Jortner J, J Phys Chem B 104:3906, 2000.28. Voityuk AA, Rosch N, Bixon M, Jortner J, J Phys Chem B 104: 9740, 2000.29. Nunez ME, Hall DB, Barton JK, Chem Biol 6:85, 1999.30. Troisi A, Orlandi G, J Phys Chem B 106:2093, 2002.31. Sadowska-Aleksiejew A, Rak J, Voityuk AA, Chem Phys Lett 429:546, 2006.32. Voityuk AA, Siriwong K, Rosch N, Phys Chem Chem Phys 3:5421, 2001.33. Liu T, Barton JK, J Am Chem Soc 127:10160, 2005.34. O Neill MA, Barton JK, Top Curr Chem 236:67, 2004.35. Delaney S, Barton JK, J Org Chem 68:6475, 2003.36. Schuster G, Acc Chem Res 33:253, 2000.37. Henderson PT, Jones D, Hampikian G, Kan Y, Schuster G, Proc Nat Acad Sci USA
96:8353, 1999.38. Giese B, Acc Chem Res 33:631, 2000.39. Grozema FC, Berlin YA, Siebbeles LDA, J Am Chem Soc 122:10903, 2000.40. Giese B, Biland A, Chem Commun 667, 2002.41. Voityuk AA, Siriwong K, Rosch N, Phys Chem Chem Phys 3:5421, 2001.42. Voityuk AA, Chem Phys Lett 427:177, 2006.43. Giese B, Spitchy M, Wessely S, Pure Appl Chem 73:449, 2001.44. Giese B, Spichty M, Chem Phys Chem 1:195, 2000.45. Giese B, Amaudrut J, Kohler A-K, Spormann M, Wessely S, Nature 412:318, 2001.46. Wagenknecht H-A, Charge Transfer in DNA, Wiley InterScience, 2005.47. Ly D, Sanii L, Schuster GB, J Am Chem Soc 121:9400, 1999.48. Boon EM, Barton JK, Curr Opin Struct Biol 12:320, 2002.49. Takada T, Kawai K, Cai X, Sugimoto A, Fujitsuka M, Majima T, J Am Chem Soc
126:1125, 2004.50. Lewis FD, Liu X, Liu J, Miller SE, Hayes RT, Wasielewski MR, Nature 406:51, 2000.51. Giese B, Kendrick T, Chem Commun 2016, 2002.52. Li X, Cai Z, Sevilla MD, J Phys Chem A 106:9345, 2002.53. Prat F, Houk KN, Foote CS, J Am Chem Soc 120:845, 1998.




























