Embed Size (px)
Citation preview

REPORT
S. R. Scheffers Æ G. Nieuwland Æ R. P. M. Bak
F. C. van Duyl
Removal of bacteria and nutrient dynamics within the coralreef framework of Curacao (Netherlands Antilles)
Received: 5 May 2003 / Accepted: 3 November 2003 / Published online: 9 July 2004� Springer-Verlag 2004
Abstract The authors studied removal rates of bacteriaand the regeneration of inorganic nutrients in coral reefcavities in the reef slope of Curacao, NetherlandsAntilles. We found that in cavities the hard substratumsurface area (=ca 68% of cavity surface area) is 65%covered with sessile filter feeders. The cryptic cavitysurface area exceeds the projected surface area of thereef by 1.5–8 times. Consequently, the organisms livingin these cryptic habitats have potentially a large impacton pico- and nano-plankton densities and are importantin reef water nutrient dynamics. We closed cavities(±70 l volume, 15 m depth) in seven experiments tostudy changes in bacterial densities and dissolved inor-ganic nutrients (DIN, DIP, and silicate) over time.Water samples were taken from the middle of the cavityat 5-min intervals, for 30 min, and analyzed for het-erotrophic bacterial abundance and nutrient concentra-tions. After closure, bacterial abundance droppedrapidly. Of the initial bacterial concentration in thecavities, 50–60% had disappeared after 30 min, anaverage disappearance rate of 1.43·104 bacteria ml)1
min)1 (0.62 mg C l)1 d)1; or 30.1 mg C m)2 cavitysurface area d)1). NOx concentrations increased signifi-cantly during the time of closure. Efflux rates variedbetween 1.02–9.77 mmol m)2 cavity surface area d)1.NH4
+ and PO43) concentrations were variable and
did not show a consistent change over time inthe experiments. Comparison of bacterial organic
nitrogen disappearance rates and DIN (NOx+NH4+)
release rates suggests that on average only 30–40%of additional sources of N besides bacteria were requiredto balance the nitrogen budget. This highlights theimportance of heterotrophic bacterioplankton as foodfor cryptic filter feeders on coral reefs. Silicate concen-trations significantly decreased after closure with0.50 mmol m)2 cavity surface area d)1, suggesting thenet deposition of SiO4
2) in spicules of cryptic filterfeeding sponges. We conclude that coral reef cavities area major sink for heterotrophic bacteria, a sink for dis-solved silicon (DSi), and a source for NOx. That reefcavities are a source for NOx suggests strong reminer-alization and nitrification in cavities with a potential rolefor sponge-symbiotic microbial nitrification.
Keywords Coral reef Æ Bacteria Æ Cryptic sessile fauna ÆNutrient regeneration Æ Cavities Æ Silicate
Introduction
Cryptic habitats inside the reef framework are a largeand yet unexplored component of the coral reef system.Cavities present the largest cryptic habitat on reefs. Theyoccur under coral colony overhangs and can be found asvoids in the coral reef framework. Volumes of individualcavities on reefs range from several to hundreds of liters.Cavities encompass 30 to 75% of the bulk volume ofcorals reefs, offering a large surface (75% of the totalsurface on reefs) for the growth and exploitation ofcryptic sessile organisms (Ginsburg 1983). Cavity sur-face area can exceed that of the projected reef by a factor1.5–8.0, depending of reef zone and three-dimensionalcomplexity of the reef (Richter et al. 2001; Scheffers et al.2003). The biological activity in cavities is stronglyassociated with surface area.
Bottom cavities are semi-closed systems, with reducedexchange with ambient water. Water quality character-istics often differ between cavity and ambient water.
Communicated by K.S. Sealey
S. R. Scheffers (&) Æ G. Nieuwland Æ R. P. M. BakF. C. van DuylRoyal Netherlands Institute for Sea Research (NIOZ),1790 AB Den Burg, P.O. Box 59, Texel, The NetherlandsE-mail: [email protected]
S. R. ScheffersCARMABI Foundation, P.O. Box 2090,Willemstad, Curacao, Netherlands Antilles
R. P. M. BakUniversity of Amsterdam, Mauritskade 57,Amsterdam, The Netherlands
Coral Reefs (2004) 23: 413–422DOI 10.1007/s00338-004-0400-3

Depletion of chlorophyll a in cavity water relative toambient water has been reported for framework cavitiesin the Mediterranean and in the Red Sea (Gili et al.1986; Richter and Wunsch 1999). On the reefs alongCuracao, Gast et al. (1998) found that bacterial densitieswere usually lower inside cavities than outside and thatthe chemical composition of water in cavities often dif-fers from that in overlying water (Van Duyl and Gast2001). These differences in water quality characteristicshave been ascribed to activities by cryptofauna or coe-lobites living on the walls of cavities (Richter et al.2001). The majority of cryptofauna species are suspen-sion feeders dominated in cover by encrusting sponges(Kobluk and Van Soest 1989; Wunsch et al. 2002) thatfilter nano- and pico-plankton from the water column.Many tropical sponges utilize heterotrophic bacteria astheir main food source (Reiswig 1971; Pile 1996; Ribesand Gili 1999; Kotter and Pernthaler 2002). Particles inthe size class of <2 lm are preferably removed from theambient water by sponges (Pile et al. 1996).
Inorganic nutrient concentrations are often enhancednear the reef framework and in coral cavities (Gast et al.1998; Rasheed et al. 2002). The regenerative spaces inthe reef are apparently a net source of inorganic N and P(Hatcher 1997). The cavity fauna decomposes organicmatter and regenerates inorganic nutrients (Richter et al.2001). Open water sponges excrete excess N in the formof nitrate into the water column (Corredor et al. 1988;Diaz and Ward 1997). Biotic activity with respect tonutrient regeneration is not limited to cavity wallinhabitants. It also occurs in cavity bottom sediments(Capone et al. 1992) and within coral skeletons byendolithic communities (Ferrer and Szmant 1988). Free-living residents of cavities, such as fishes and motileinvertebrates, may also influence nutrient concentrationsnear the reef bottom. In view of such effects of fauna onthe water quality, it must be expected that the biologicaland chemical characteristics of water within cavitiesdiffer from ambient reef water.
The aim is to quantify these water characteristics interms of fluxes of bacteria and nutrients. We assesseduptake rates of bacteria and inorganic nutrient cycling incoral cavities and established the role of these processesin coral reef trophodynamics. In order to do so wemeasured changes in bacterial densities and inorganicnutrient concentrations over time intervals after cavitieswere closed off. Closure eliminates exchange of thecavity water with overlying reef water and this allowedus to estimate the bacterial removal and mineralizationrates by the cavity biota.
Materials and methods
We focused our studies on two cavities at two sites(WF and B1, Fig. 1) on the fringing reef along theleeward coast of Curacao (Netherlands Antilles) (Bak1977; Van Duyl 1985). The cavities were situated at15-m depth on the reef slope. We measured cavity
geometry (cavity volume and cavity surface area) withthe cave-profiler, an instrument to measure the 3-Dstructure of reef cavities (Scheffers et al. 2003)
Cryptofauna cover in the cavities was estimated with aSony 900E digital CaveCam adapted from Wunsch andRichter (1998). Per cave, 48 random, non-overlappingpictures (4·4 cm) were taken and analyzed with AdobePhotoShop 4.0. Random points ( n=100) were projectedover the picture and substratum types lying directlybeneath these points were identified. We distinguishedcover of suspension feeders, cover of sponges as share ofthe suspension feeders, cover of calcareous algae, andbaresubstratum. To study in situ removal rate of bacteria andnutrient dynamics, we stopped water exchange betweencavity and reef overlying water by closing each cavitywitha tightly woven cloth. The separation of cavity and reefwater was tested, releasing fluorescin in the closed offcavity.We did not observe any leakage of the dye from theclosed cavities within 30 min, which indicates the effec-tiveness of the closure. Surface wave-action had an effecteven at 15 m and made the cloth, and subsequently thecavity water, move gently back and forth.
Experimentswere conducted ondifferent days between27 September and 13December 2001.Water samples weretaken from the center of the cavity through a fixed siliconetube. Before the start of the experiment, duplicate watersamples ( n =2) were taken from cavity and overlyingwater (in front of the cavity entrance 1 m distance fromsubstrate). After closing the cavity, water samples ( n=2)were taken at 0, 5, 10, 15, 20, and 30 min. After sampling,the syringes were brought to the surface and kept on ice inthe dark, while being shipped to the laboratory. For eachtime interval, we analyzed DSi (dissolved silica), NO3,
NO2, NH4, PO4, and heterotrophic bacterial abundance.Duplicate samples for nutrient analyses were filtered overa disposable 0.22-lm sterile polycarbonate filter andsubsequently frozen and stored at )20 �C (N and P) andstored at +4 �C (Si) in 6-ml Packard pony vials. Allnutrient concentrations were analyzed with a Traacs 800auto-analyzer (Technicon).
Duplicate 10-ml samples for bacterial counts werestained with acridine orange and filtered over blackNuclepore 0.22-lm polycarbonate filters (Hobbie et al.1977). Bacteria were counted with a Zeiss epifluores-cence microscope with magnification 1250x. Conversionfactors used for bacterial carbon and nitrogen contentwere 30 fg and 5.8 fg per bacteria, respectively (Fukudaet al. 1998). We kept at least a 1-day interval betweeneach experiment in order to allow pre-experiment con-ditions in the cavities to be restored. We did fourexperiments with the cavity at WF, and three with thecavity at B1. In seven experiments, the removal of bac-teria was assessed, and in five experiments, nutrientconcentration changes were followed.
Bacterial removal rates were calculated assumingexponential clearance of prey in closed cavities:
NðtÞ ¼ Nð0Þ e�at ð1Þ
414

N =number of bacteria, t =time, and a =ratecoefficient.
Nutrient mineralization rates were calculatedassuming a linear increase in inorganic nutrients inclosed cavities:
CðtÞ ¼ aCð0Þ þ b ð2Þ
C =nutrient concentration, t =time, a =rate coef-ficient, and b=constant.
An indication of relationship between the processesof bacterial removal and nutrient production/disap-pearance is found when the rate coefficients of theseprocesses are similar.
The fluxes based on concentration changes in time(mgC.l)1.d)1) in closed-off cavities were recalculated tounit cavity surface area. This was accomplished bymultiplying these initial fluxes by cavity volume andthen dividing this number by cavity surface area (thetotal surface area and the surface area covered byall suspension feeders together, sponges alone, respec-tively). Cavity surface areas and volumes wereobtained with the cave-profiler (Scheffers et al. 2003)and cover of suspension feeders was obtained with theCaveCAM.
Results
Cavity geometry and cryptic organisms
The cavities for this study are typical of such structuresin this habitat (Scheffers et al. 2003). They are charac-
terized by hard substratum roofs and walls and havesandy bottoms. Cavities had frontal openings of,respectively, 0.31 and 0.27 m2, which faced off the reefslope. The total inner surface area of the cavities amountto 1.41 m2 (B1) and 1.52 m2 (WF), of which ca. 2/3 ishard substratum and ca. 1/3 is sand on the bottom(Table 1). The hard substratum was covered for morethan 93% with cryptic organisms. Encrusting calcareousalgae dominated cover in the entrance part, and filterfeeders in the deeper part. Sponges, polychaetes, bry-ozoans, ascidians, and bivalves represent the filter/suspension feeding community. The cover of this com-munity is dominated by encrusting sponges (Demo-spongiae and Calcarea). Morphological and bioticcharacteristics were roughly similar for the two cavities.We found that fluxes were also comparable betweencavities. Therefore fluxes are not discussed separately fordifferent cavities.
Fig. 1 Locations of sites alongthe Caribbean island ofCuracao (Netherlands Antilles),where crevices in the reef slopeat 15 m of depth were sampled.WF water factory, B1 buoy 1.The inset shows the position ofCuracao in the Caribbean Sea,north of Venezuela
Table 1 Volume and absolute cover of solid (hard) and sand (soft)substrata and the absolute coverage of solid substratum by differ-ent cryptic organisms are given for the cavities WF and B1. Filterfeeders include sponges, tunicates, bryozoans, polychaetes, andbivalves. Between brackets, the filter feeder cover by sponges isgiven. The cover of calcareous algae includes 10–17% cover byunspecified organisms
Cavity Volume(m3)
Hardsubstratum(m2)
Sand(m2)
Filter feeders(sponges)(m2)
Calcareousalgae(m2)
WF 0.073 1.01 0.51 0.68 (0.40) 0.33B1 0.070 0.98 0.44 0.61 (0.28) 0.30
415

Heterotrophic bacteria
Bacterial concentrations were always higher outsidethan inside the cavity. Initial densities inside cavitiesdiffered substantially between experimental days. Bac-
terial densities inside of the cavities decreased on averagewith 1.43·104±4.90·103 ml)1 min)1 after closure (cal-culated with Eq. (1), and see Fig. 2, Table 2). Convert-ing this in mg C disappearing m)2 d)1 of Total SurfaceArea (TSA=Hard substratum+Soft sediment within a
Fig. 2 Significant decrease inbacterial abundance 30 minafter closure of cavities. Resultsof all experiments in cavity WFand B1 are shown. Standarddeviations are based onduplicate samples. The line is anexponential decay regression fit.Each panel contains a productmoment correlation coefficient(R) and the significance level (p)
416

cavity), gives an average removal of 30.1±10.6 mg Cm)2 d)1. If the filter feeder community (FF) in thecavities is responsible for total removal of bacteria, theaverage uptake is 68.0±23.4 mg C m)2 FF d)1. Spongeshave the highest percentage cover in the FF (Table 1).Assuming sponges take up all bacteria, it appears that,on average 134.0±63.3 mg C is removed m)2 sponged)1. After 30 min, bacterial densities had declined, onaverage, 50%. The average bacterial N removal rate was0.44±0.16 mmol N m)2 TSA d)1. The rate coefficientsfor bacterial uptake were for all experiments and forboth cavities within the same range (Table 3), in spite ofdifferent values in initial densities. On the other hand,bacterial removal rates increased significantly (R=0.87,p <0.01) with increasing initial bacterial concentrations(Fig. 3).
Nutrients
Initial nitrate concentrations were always higher insidethan outside of the cavities, with average concentrationsof 0.78±0.48 lmol l)1 inside, and 0.65±0.42 lmoll)1outside the cavities. Initial nitrite concentrations werein all cases lower inside than outside of the cavities withaverage concentrations of 0.07±0.03 lmol l)1 inside and0.18±0.19 lmol l)1 outside cavities. Nitrate and nitritefluxes were different between experiments, but concen-trations usually showed a net increase over time. Theaverage NO3
- efflux was 1.41±1.45 mmol m)2 TSA d)1,and the average NO2
- efflux was 0.03±0.05 mmol m2
TSA d)1. The average NOx (NO3)+NO2
)) efflux was1.42±1.45 mmol m)2 TSA d)1. The rate coefficients for
NOx fluxes varied between 0.006–0.056 (Table 3), withthe steepest concentration increase at experiment WF-1(Fig. 4).
Initial ammonium concentrations were always higheroutside than inside of the cavities, with average con-centrations of 1.86±1.98 lmol l)1 inside and 0.72±0.63lmol l)1 outside cavities. We found a net NH4
+ influx inthree experiments (0.25–2.00 mmol m)2 TSA d)1) and anet efflux in two experiments (0.03 and 0.34 mmol m)2
TSA d)1). The average DIN (NO3)+NO2
)+NH4+)
efflux was 0.67±1.22 mmol m)2 TSA d)1. One experi-ment showed an influx of 0.30 mmol m)2 TSA d)1
(Table 4).On average, initial DIP concentrations were higher
outside (0.37±0.55 lmol l)1) than inside (0.12±0.06lmol l)1) of the cavity. DIP fluxes were highly variable.We found a significant net increase in concentration inthree experiments (0.076–0.540 mmol m)2 TSA d)1) anda net decrease in two experiments (0.11–6.49 mmol m)2
TSA d)1).Initial silicate (measured as SiO4
2)) concentrationswere on average higher outside (2.29±0.64 lmol l)1) thecavity than inside (1.81±0.38 lmol l)1). During theexperiments, we found a consistent net decrease in sili-cate concentrations (Fig. 5). The average rate of silicateremoval was 0.50±0.21 mmol m)2 TSA d)1 (Table 4).All inorganic nutrient fluxes are calculated with Eq. 2.
Discussion
Fluxes in coral reef cavities are best understood whenrelated to the cavity wall/bottom biota. Due to the thinand encrusting growth forms, area will be related tobiomass in these cryptic organisms. Actual measurementof cryptofauna biomass is difficult, without destructionof the habitat. Here we use surface area of the cavitiesand area covered by cryptic biota as the relevant vari-ables to understand fluxes of matter in coral reef cavities.
Bacterial fluxes
Bacterial disappearance starts immediately after closureof a cavity. There are several processes that can beresponsible for this instantaneous decrease. A number of
Table 2 Removal rates ofbacteria, bacterial C of sevenexperiments (four in cavity WFand three in B1) on differentdays in September to mid-December. Rates are expressedas disappearance of bacterialcells and of bacterial carbon pervolume cavity water (ml and l)and per surface area (totalcavity surface area and surfacearea covered with filter feeders),per unit time (minute and day)
Cavity Date Removal of bacteria Removal of carbon
Cavity water Cavity water Total cavitysurface area
Filter feedersurface area
x1000 ml)1min)1 mgC l)1d)1 mgC TSA m)2 d)1 mgC FF m)2 d)1
WF-1 01-Oct 14.30 0.62 29.72 68.15WF-2 03-Oct 14.14 0.61 29.38 67.38WF-3 05-Oct 14.37 0.62 29.86 68.46WF-4 27-Sep 9.57 0.41 19.89 45.61B1–1 08-Oct 23.06 1.00 49.34 109.91B1–2 09-Oct 16.51 0.71 35.33 78.71B1–3 13-Dec 7.97 0.34 17.06 38.00Mean (SD) 14.27 (4.90) 0.62 (0.21) 30.08 (10.59) 68.03 (23.36)
Table 3 Rate coefficients of removal of bacteria and NOx and DINeffluxes across the cavity substratum–cavity water interface of fiveexperiments (two in cavity WF and three in B1). NOx=NO3+NO2, DIN=dissolved inorganic nitrogen
Cavity Removalof bacteria
NOx efflux DIN efflux
WF-1 0.029 0.056 0.040WF-2 0.026 0.012 0.006B1–1 0.031 0.006 0.006B1–2 0.033 0.006 0.003B1–3 0.028 0.022 )0.006
417
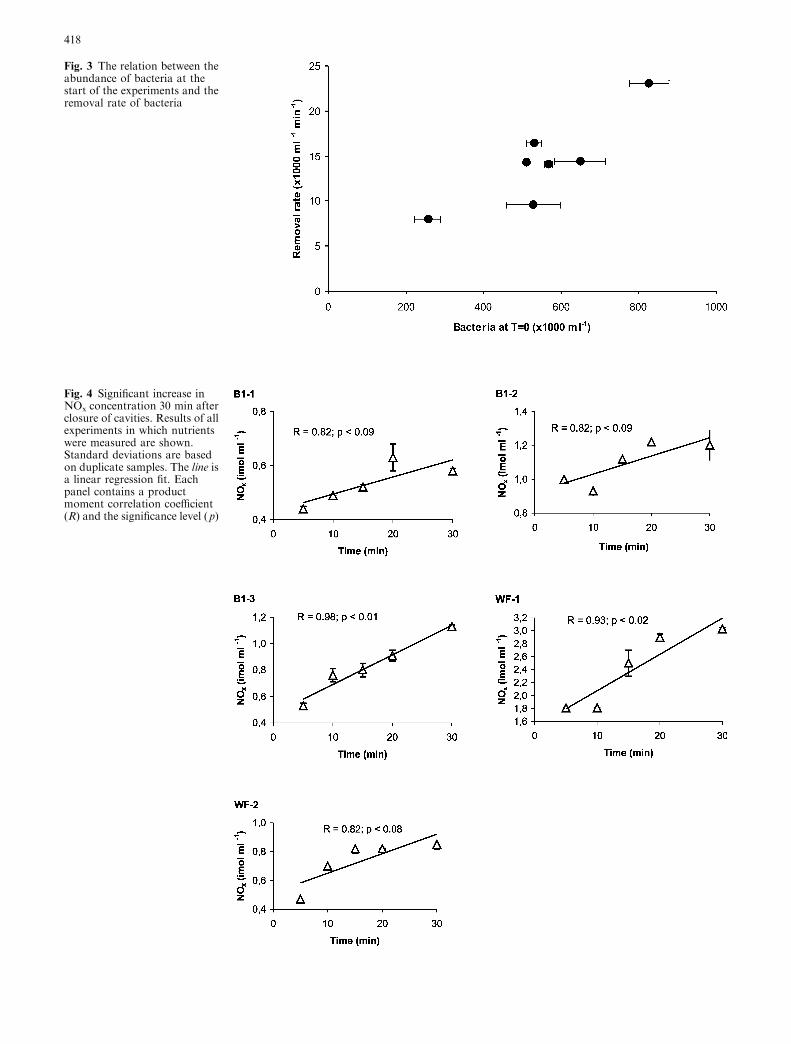
Fig. 3 The relation between theabundance of bacteria at thestart of the experiments and theremoval rate of bacteria
Fig. 4 Significant increase inNOx concentration 30 min afterclosure of cavities. Results of allexperiments in which nutrientswere measured are shown.Standard deviations are basedon duplicate samples. The line isa linear regression fit. Eachpanel contains a productmoment correlation coefficient(R) and the significance level (p)
418

these processes are, however, unlikely to be important,either because the process effects are comparable insideand outside of cavities, such as in microbial grazing(Gast et al. 1999) and viral infections (Noble et al. 1999),
or because process rates are known to be very slow, suchas in adsorption and sedimentation of bacteria. Con-finement effects, such as increase of bacterial productionand increased viral bacteriophagy, common in smallclosed volumes, are not expected in these large volumes(Noble et al. 1999). The most likely factor explaining thedisappearance of bacteria is bacterivory by sessile sus-pension feeding cryptofauna.
Bacterial removal rates were on average 68 mg C m)2
filter feeding area d)1. This rate is in the same range as
rates given in the literature for bacterial feeding in a
Table 4 Overview of the silicon influxes and inorganic N effluxes across the cavity substratum–cavity water interface of five experiments(two in cavity WF and three in B1). For comparison, the influx of bacterial N is given in the last column
Cavity DSi influx(mmol TSA m)2d)1)
NOx efflux(mmol TSA m)2d)1)
DIN efflux(mmol TSA m)2d)1)
Bacterial N influx(mmol TSA m)2d)1)
WF-1 0.72 3.88 2.77 0.41WF-2 0.26 0.80 0.42 0.41B1–1 0.50 0.43 0.31 0.68B1–2 0.32 0.43 0.14 0.49B1–3 0.68 1.57 )0.30 0.24Mean (SD) 0.50 (0.21) 1.42 (1.45) 0.67 (1.21) 0.44 (0.16)
Fig. 5 Consistent decrease in DSi concentration 30 min afterclosure of cavities. Results of all experiments in which DSiconcentrations were measured are shown. Error bars refer to the95% confidence limits of concentrations as established during themeasuring period. The line is a linear regression fit. Each panelcontains a product moment correlation coefficient (R) and thesignificance level (p)
419

range of suspension and filter feeders. Bak et al. (1998)gives an estimate of 0.75–1.07·109 bacteria 100 cm)2 h)1
for the ascidian Trididemnum solidum, and the sclerac-tinian coral Madracis mirabilis (Curacao). This is equalto 54–77 mg C m)2 d)1 (calculated with a conversionfactor of 30 fg C per heterotrophic bacterial cell),assuming that uptake is taking place at the same rateduring 24 h d)1. Pile et al. (1996) calculated the grazingcapacity of the boreal sponge Mycale lyngua on het-erotrophic bacteria to be 29 mg C m)2 d)1. Kotter andPernthaler (2002) found for the cryptic Caribbeansponges Ulosa ruetzleri, Halisarca caerula , Clathriararaechelae , Merlia normani , Desmanthus incrustans ,and Diplastrella megastellata, after recalculation andconversion with 30 fg C per heterotrophic bacterium,respectively, 101, 729, 162, 163, 9, and 58 mg C m)2 d)1.Their removal rates are in agreement with the rate of(heterotrophic) bacterial carbon removal by crypticsponges in our study (133 mg C sponge m)2 d)1).
Grazing estimates that include a larger size range ofplanktonic organisms may be higher. Depletion ofphytoplankton in reef overlying waters and in reef cav-ities has been reported (Ayukai 1995; Yahel et al. 1998;Richter et al. 2001; Van Duyl et al. 2002). Richter andWunsch (1999) found phytoplankton removal rates inthe range of 900 mg C m)2 d)1 for coelobite communi-ties in the Red Sea on basis of chlorophyll a. Conse-quently, their total nano-and pico-plankton removalincluding heterotrophic bacterial biomass may be atleast twice as high. Our data are definitely more con-servative. We measured only the uptake of heterotrophicbacteria and therefore we may underestimate the totalflux of particulate organic matter into cavities.
Bacterioplankton production in reef waters is low.Production under normal trophic circumstances is onaverage 7 lg C l)1 d)1 in reef overlying water alongCuracao (Gast et al. 1999). When we compare ourbacterial removal rates with bacterioplankton produc-tion, even with extremely high productivity such as un-der the eutrophic reef conditions off the harbor ofCuracao (Gast et al. 1999), it is evident that our removalrates of bacteria in cavities exceed bacterial productionby far (bacterial production of 7 versus removal of620 lg C.l)1.d)1, respectively). This implies that coralcavities depend on water exchange with the passingoceanic water mass for their food supply.
Initial concentrations of bacteria inside cavities var-ied widely between experiments. Gast et al. (1999)showed temporal variations in bacterial concentrationsin the reef water column. From day to day, and evenduring the day, concentrations of bacteria in the reefwater may vary depending on the water exchange be-tween ocean, reef boundary water, and coral cavitywater. This could explain differences between initialconcentrations of bacteria within crevices. Initial con-centrations appear to influence uptake rates m)2 d)1.In combination with the consistent rate coefficients,we argue that average pumping rate of the crypticsuspension feeders is relatively constant (Ribes et al.
1999). Filter feeders may not alter their pumping/filter-ing rates under experimental conditions, and bacterialuptake rates appear to increase with increasing initialconcentrations of bacteria in the different experiments.
Nutrient fluxes
The organic nitrogen uptake by cryptofauna in theform of bacteria (0.44±0.16 mmol TSA m)2 d)1) is inthe same range as the average efflux of DIN(0.67±1.22 mmol TSA m)2 d)1), considering the largestandard deviations. The N efflux is on average only30–40% higher than the bacterial N removal rate. Itsuggests that bacteria may well be the main food sourcefor the heterotrophic biota in cavities. This is surprisingsince DIN release does not merely depend on minerali-zation of the heterotrophic bacterial biomass. DIN isalso released by the mineralization of other sources oforganic matter in the cavity, such as picophytoplankton,detrital material in the bottom of the cavity, anddead-cavity-dwelling organisms. It is possible that thiscontribution to the DIN efflux is negligible. An overes-timation of bacteria N content would mean lower influxrates. Another explanation could be that cavities are netsinks of N and that the biomass or organic matter (OM)content in cavities gradually increases.
The inorganic N-species composition, and concen-tration, in cavity water is determined by the supply, andthe N transformations that take place in cavities. Theobservation that ammonia concentrations did not sig-nificantly vary during the experiments and that NOx
concentrations significantly increased in closed off cavi-ties, suggest nitrification, the oxidation of ammonia tonitrite and subsequently to nitrate, to be an importantpathway in the N remineralization in cavities. Theregeneration of ammonia and the ammonia supply fromthe sediment and from outside the cavity may be therate-limiting step in the nitrification in cavities. Anargument to support this is that the NOx efflux ratesexceed the efflux of DIN, implying that the initial dif-ference in ammonia concentrations with higher concen-trations outside than inside the cavity and the occasionaldecrease of ammonia during experiments may be due tonitrification. This suggests that ammonia imported fromoutside into the cavity is rapidly nitrified. Cavities areapparently nitrification hot spots with nitrificationassociated with substrates/interfaces in the cavity. NOx
efflux rates of 0.4 to 3.9 mmol N.m)2 TSA.d)1 are highcompared to, for instance, those in aphotic North Seasediments with rates of 0.12–0.48 mmol N.m)2.d)1,which are comparable to rates in oligotrophic deep-seasediment (Lohse et al. 1993). Accounting for the factthat the cryptic surface on the reef slope exceeds theprojected reef surface by up to 8 times, our rates can bemultiplied by 8, yielding rates which fall in the samerange as estimates reported for coral reefs in the Red Sea(Rasheed et al. 2002), with efflux rates from the reefframework of 8–9 mmol NOx. m
)2 projected reef.d)1. It
420

is evident that the enhanced NOx concentrations com-monly found in reef overlying waters, compared toconcentrations in adjacent oceanic waters, originate atthe reef bottom.
Nitrifying bacteria play an important role in reefs.Both corals and sponges have been reported to releasenitrate (Corredor et al. 1988; Diaz and Ward 1997;Wafar et al. 1990). These authors ascribe the nitraterelease to nitrifying bacteria, which live associated withthe host. Diaz and Ward (1997) demonstrated thepresence of ammonia oxidizing bacteria in the Carib-bean reef sponge ?Pseudaxinella zeai, which releases5.8–10.9 mmol nitrate m)2 d)1. Diaz (1997) and Hent-schel et al. (2002) established the common occurrence ofnitrite oxidizers in sponges. Considering these findings,it is highly likely that cryptic sponges may also harbornitrifying bacteria. This has not been confirmed yet.
The NOx flux from cavities may also come fromsediments in the cavity. When NOx efflux from cavities iscompletely ascribed to the sediment in cavities, we ob-tain rates of 1.82±2.99 mmol m)2 sand.d)1. Such ratesare in the same range as mineralization rates for reefsediments as reported by Capone et al. (1992) of1.68 mmol N m)2 d)1.
Inorganic phosphorous fluxes showed an irregularpattern in our experiments. Initial DIP concentrationswere lower inside than outside cavities, suggesting a netinflux of DIP. However, we found three times a signifi-cant net efflux. The DIP efflux is consistent withenhanced DIP concentrations in coral cavities comparedto ambient water (Andrews and Muller 1983; Van Duyland Gast 2001; Rasheed et al. 2002). Pile (1996) foundreef sponges to release DIP.
Silicate influxes (0.50 mmol m)2 TSA d)1) were veryhigh, considering that uptake of silicate is restricted toorganisms with a silicate skeleton, such as the frustule ofdiatoms and the spiculae in sponges. Sponges are wellrepresented on cavity walls. Confining the DSi uptake totheir surface area yields uptake rates of 2.30 mmol m)2
sponge.d)1. This is extremely high for sponges and notequaled in the literature. The highest rates we could findamounted to 0.2 lmol m)2 d)1 for the sponge Hali-chondria panacea (Frohlich and Barthel 1997; Reinckeand Barthel 1997). DSi concentrations in the cavitywater were already low at the start of the experiments.With these low concentrations, an extremely high uptakeaffinity for DSi is required. It is unknown whether thecryptofaunal organisms with a silicon skeleton have suchhigh affinities.
To assess the impact of cryptic biota on the tropho-dynamics of coral reefs, we extrapolate the bacterialremoval rate (as measured in cavities) to the total crypticsurfaces below 1 m2 projected reef surface and comparethis with the average benthic primary production m)2
reef. The cryptic surface area in the reef is 1.5 to 8 timeslarger than that of the projected reef with the largestsurface enlargement occurring at 15-m depth (Richteret al. 2001). We calculated the bacterial removal for 1 m2
of projected reef surface at 15-m depth. We assumed
that the overall cover of suspension feeders in cryptichabitats is comparable to the area coverage in our studycavities. The calculation shows that the cryptic surfacesremove 240 mg bacterial C m)2 projected reef d)1at15-m depth (8*30 mg C m)2 d)1, Table 2). Picophyto-plankton (dominated by photoautotrophic bacteria) alsodisappears from the reef overlying waters at Curacao(Van Duyl et al. 2002). This suggests that the removalrates of plankton may well exceed 240 mg C m)2 d)1 at15-m depth and may even double, considering thecomparable biomasses of heterotrophic and photoau-totrophic bacteria in oligotrophic waters (Campbellet al. 1994) and the comparable retention efficiency ofsuch small particles by tropical coral reef sponges (Pile1996; Kotter and Pernthaler 2002). Doubling 240 resultsin a removal of C of 480 mgC.m)2.d)1 at 15 m. Grossproduction of a coral reef is about 3,000 mgC.m)2.d)1,of which 97% is respired (Hatcher 1997). Organic matterrelease by benthic primary producers on the reef (17–275 mg C m)2 d)1, e.g., Richman et al. 1975; Crossland1987) may indirectly (via bacterioplankton secondaryproduction in reef water, Van Duyl and Gast 2001) fuelthe cryptic biota, which depends on the matter advectedto them by the water movement. Considering therespiratory losses in trophic transfers (uptake of dis-solved organic matter by bacteria), as well as the loss ofthis organic matter to the ocean, it is unlikely that thecoral reef benthos can fully provide the energy demandof cryptic biota. This may imply that the cryptic biota oncoral reefs along Curacao are predominantly dependenton organic matter supply from the adjacent water mas-ses, and as such, more dependent on allochthonous thanautochthonous organic matter. Inorganic nutrients re-leased by mineralization of this predominantly allo-chthonous organic matter are available for reef benthicprimary producers. It is evident that cryptic biota mayplay an important role in reef trophodynamics byincorporating ocean-derived organic matter in the reefbenthos and supply the reef with extra inorganic nutri-ents.
Conclusion
The experiments show that coral reef cavities are a majorsink for heterotrophic bacteria and a source for DIN inthe form of NOx. The first points to strong heterotrophicactivity of the cryptic suspension feeders in cavities, andthe latter suggests that cavities are key regenerativespaces and hot spots of nitrification. Our silicateremoval rates in cavities are the fastest ever measured inthe coral reef environment. Observed fluxes can only bemaintained through rapid exchange with ambient reefand oceanic water.
Acknowledgements We wish to thank the staff of the CARMABIFoundation (Curacao, Netherlands Antilles) and especially ‘‘Don’’Carlos and Brian Leysner for their invaluable cooperation. Wethank Jan van Ooyen, Evaline van Weerlee, and Karel Bakker
421

(NIOZ nutrient lab) for the nutrient analyses. Very special thanksgo to Talitha Miedema for her assistance in the field and Rob vanSoest and Cees Veth for discussions on experimental procedure.This study is financed by the WOTRO grant nr. W84–439.
References
Andrews JC, Muller H (1983) Space-time variability of nutrients ina lagoonal patch reef. Limnol Oceanogr 28:215–227
Ayukai T (1995) Retention of phytoplankton and planktonic mi-crobes on coral reefs within the Great Barrier Reef, Australia.Coral Reefs 14:141–147
Bak RPM (1977) Coral reefs and their zonation in the NetherlandsAntilles. Stud Geol 4:3–16
Bak RPM, Joenje M, De Jong I, Lambrechts DYM, Nieuwland G(1998) Bacterial suspension feeding by coral reef benthicorganisms. Mar Ecol Prog Ser 175:285–288
Campbell L, Nolla HA, Vaulot D (1994) The importance of Pro-chlorococcus to community structure in the Central North Pa-cific Ocean. Limnol Oceanogr 39:954–961
Capone DG, Dunham SE, Horrigan SG, Duguay LE (1992)Microbial nitrogen transformations in unconsolidated coralreef sediments. Mar Ecol Prog Ser 80:75–88
Corredor JE, Wilkinson CR, Vicente VP, Morell JM, Otero E(1988) Nitrate release by Caribbean reef sponges. LimnolOceanogr 33:114–120
Crossland CJ (1987) In situ release of mucus and DOC-lipid fromthe corals Acropora variabilis and Stylophora pistillata in dif-ferent light regimes. Coral Reefs 6:35–42
Diaz MC (1997) Molecular detection and characterization of spe-cific bacterial groups associated with tropical sponges. Proc 8thInt Coral Reef Symp, Panama, pp 1399–1402
Diaz MC, Ward BB (1997) Sponge mediated nitrification in trop-ical benthic communities. Mar Ecol Prog Ser 156:97–107
Ferrer LM, Szmant AM (1988) Nutrient regeneration by theendolithic community in coral skeletons. Proc 6th Int CoralReef Symp, Townsville, Australia, 8th 12th August 3
Frohlich H, Barthel D (1997) Silica uptake of the marine spongeHalichondria panicea in Kiel Bight. Mar Biol 126:115–125
Fukuda R, Ogawa H, Nakata T, Koike I (1998) Direct determi-nation of carbon and nitrogen contents of natural bacterialassemblages in marine environments. Appl Environ Microb64:3352–3358
Gast GJ, Jonkers PJ, Van Duyl FC, Bak RPM (1999) Bacteria,flagellates and nutrients in island fringing coral reef waters:influence of the ocean, the reef and eutrophication. Bull Mar Sci65:523–538
Gast GJ, Wiegman S, Wieringa E, Van Duyl FC, Bak RPM (1998)Bacteria in coral reef water types: removal of cells, stimulationof growth and mineralization. Mar Ecol Prog Ser 167:37–45
Gili J-M, Riera T, Zabala M (1986) Physical and biological gra-dients in a submarine cave on the Western Mediterranean coast(north-east Spain). Mar Biol 90:291–297
Ginsburg RN (1983) Geological and biological roles of cavities incoral reefs. In: Barnes DJ (ed) Perspectives on coral reefs.Australian Institute of Marine Science, Townsville, Australia(pp 148–153)
Hatcher B (1997) Organic production and decomposition. In:Birkeland C (ed) Life and death of coral reefs. Chapman andHall, New York, pp 140–174
Hentschel U, Hopke J, Horn M, Friedrich AB, Wagner M, HackerJ, Moore BS (2002) Molecular evidence for a uniform microbialcommunity in sponges from different oceans. Appl EnvironMicrob 68:4431–4440
Hobbie JE, Daley RJ, Jasper S (1977) Use of nuclepore filters forcounting bacteria by fluorescence microscopy. Appl EnvironMicrobiol 33:1225–1228
Kobluk DR, Van Soest RWM (1989) Cavity-dwelling sponges in asouthern Caribbean coral reef and their paleontological impli-cations. Bull Mar Sci 44:1207–1235
Kotter I, Pernthaler J (2002) In situ feeding rates of obligate andfacultative coelobite (cavity-dwelling) sponges in a Caribbeancoral reef. Proc 9th Int Coral Reef Symp, Bali, Indonesia. vol.1:347–352
Lohse L, Malschaert JFP, Slomp CP, Helder W, Van RaaphorstW (1993) Nitrogen cycling in North Sea sediments: interac-tion of denitrification and nitrification in offshore and coastalareas. Mar Ecol Prog Ser 101:283–296
Noble RT, Middelboe M, Fuhrman JA (1999) Effects of viralenrichment on the mortality and growth of heterotrophic bac-terioplankton. Aq Microb Ecol 18:1–13
Pile AJ (1996) The role of microbial food webs in benthic-pelagiccoupling in freshwater and marine ecosystems. PhD Thesis,College William and Mary, Virginia, pp 1–166
Pile AJ, Patterson MR, Witman JD (1996) In situ grazing onplankton <10 um by the boreal sponge Mycale lingua. MarEcol Prog Ser 141:95–102
Rasheed M, Badran MI, Richter C, Huettel M (2002) Effect of reefframework and bottom sediment on nutrient enrichment in acoral reef of the Gulf of Aqaba, Red Sea. Mar Ecol Prog Ser239:277–285
Reincke T, Barthel (1997) Silica uptake kinetics of Halichondriapanicea in Kiel Bight. Mar Biol 129:591–593
Reiswig H (1971) Particle feeding in natural populations of threemarine demosponges. Biol Bull 141:568–591
Ribes M, Gili J-M (1999) Seasonal variation of particulate or-ganic carbon, dissolved organic carbon and the contributionof microbial communities to the live particulate organic car-bon in a shallow near-bottom ecosystem at the northwesternMediterranean Sea. J Plankton Res 21:1077–1100
Richman S, Loya Y, Slobodkin LR (1975) The rate of mucusproduction by corals and its assimilation by the coral reefcopepod Acartia negligens. Limnol Oceanogr 20:918–923
Richter C, Wunsch M (1999) Cavity-dwelling suspension feeders incoral reefs-a new link in reef trophodynamics. Mar Ecol ProgSer 188:105–116
Richter C, Wunsch M, Rasheed M, Kotter I, Badran MI (2001)Endoscopic exploration of Red Sea coral reefs reveals densepopulations of cavity-dwelling sponges. Nature 413:726–730
Scheffers SR, de Goeij J, Van Duyl FC, Bak RPM (2003) The cave-profiler: a simple tool to describe the 3D structure of inacces-sible coral reef cavities. Coral Reefs 22:49–53
Van Duyl FC (1985) Atlas of the living reefs of Curacao andBonaire (Netherlands Antilles). Foundation for ScientificResearch in Surinam and the Netherlands Antilles, Utrecht,pp 1–63
Van Duyl FC, Gast GJ (2001) Linkage of small-scale spatialvariations in DOC, inorganic nutrients, and between bacterio-plankton growth with different coral reef water types. AquatMicrob Ecol 24:17–26
Van Duyl FC, Gast GJ, Steinhoff W, Kloff S, Veldhuis MJW, BakRPM (2002) Factors influencing the short-term variation inphytoplankton composition and biomass in coral reef waters.Coral Reefs 21:293–306
Wafar M, Wafar S, David JD (1990) Nitrification in coral reefs.Limnol Oceanogr 35:725–730
Wunsch M, Al-Moghrabi SM, Kotter I (2002) Communities ofcoral reef cavities in Jordan, Gulf of Aqaba (Red Sea). Proc 9thInt Coral Reef Symp, Bali, Indonesia. vol 1, pp 595–600
Wunsch M, Richter C (1998) The CaveCam—an endoscopicunderwater video system for the exploration of cryptic habitats.Mar Ecol Prog Ser 169:277–282
Yahel G, Post AF, Fabricius K, Marie D, Vaulot D, Genin A(1998) Phytoplankton distribution and grazing near coral reefs.Limnol Oceanogr 43:551–563
422





























