Embed Size (px)
Citation preview

MOLECULAR AND CELLULAR NEUROSCIENCES 3,487-496 (19%)
Rat Anterior Pitu itary Estrogen Receptor mRNA Levels a fter Estrogen and Progesterone Treatment
L. CHORICH, M. ROSSER, E. HOWARD, P. ZAMORANO, AND V. B. MAHESH~
Department of Physiology and Endocrinology, Medical College of Georgia, Augusta, Georgia 30912-3000
Received for publication April 9, 1992
Previous work from our laboratory has demonstrated that progesterone can induce a rapid decrease in estrogen receptor (ER) binding in the anterior pituitary of the rat. This effect of progesterone on ER correlates well with the loss of estrogen action. The objectives of this study were to determine whether the rapid decrease in ER binding in the rat pituitary after a single injection of es- tradiol and the decrease in ER binding induced by pro- gesterone in the estrogen-pr imed rat could be due to a suppression of mRNA levels for the estrogen receptor. Ovariectomy for 14 days decreased ER mRNA levels and injection of 2 Bg of estradiol in ethanol:saline for 3 days increased ER mRNA levels. A single injection of 1 rg of estradiol brought about no change in ER mRNA levels in the pituitary of ovariectomized rats within 1 h of ad- ministration. Five micrograms of estradiol increased the ER mRNA levels, whereas 10 pg of estradiol brought about a decrease in ER mRNA levels. Thus, the reported decrease in pituitary ER binding after 1 and 5 pg of es- tradiol was not accompanied by a similar change in ER mRNA levels, suggest ing that the decrease in ER binding may be due to accelerated processing of ER or to de- creased translation of ER mRNA. When 2 pg of estradiol was injected into estrogen-pr imed ovariectomized rats, the recovery of ER binding at 18 h was preceded by an increase in ER mRNA levels at 12 h, suggest ing new ER synthesis during the recovery process. Progesterone (0.8 mg/kg body wt), in the absence of estradiol-priming, in- creased ER mRNA levels in the pituitary at 1,6, and 12 h and decreased them at 18 h. In estrogen-pr imed ani- mals, the changes in ER mRNA levels brought about by progesterone combined with estradiol were similar to those observed with estradiol alone, with the except ion that ER mRNA levels were elevated in the progesterone- treated group at 1 h, a time when ER binding was further suppressed. Thus the progesterone- induced decrease in ER binding was not due to a suppression of ER mRNA levels. In estrogen-pr imed rats 0.8 and 4.0 mg/kg body wt doses of progesterone increased ER mRNA levels, whereas the 2.0 mg/kg body wt dose did not. These results
1 To whom correspondence should be addressed.
indicate that estradiol a lone has a time- and dose-depen- dent effect on ER mRNA levels in the anterior pituitary. Progesterone by itself and in the presence of estrogen priming has a dose-related effect on anterior pituitary ER mRNA levels. Furthermore, the effect of progesterone on decreasing ER binding appears to be due either to a post-transcriptional effect and/or to accelerated receptor processing. 0 1992 Academic Press, Inc.
INTRODUCTION
Tissue responsiveness to estradiol and progesterone is believed to be dependent upon the presence of high affinity intracellular receptors that are specific for these hormones (l-4). W ithin the first hour after the injection of estradiol, there is a rapid depletion of cytosolic or loosely bound unoccupied nuclear receptors with a concurrent accu- mulation of estrogen bound nuclear receptors (l-4). This receptor redistribution is not stoichiometric and there is a net loss of total estrogen receptors (ER) (5). Once the ER are depleted, they remain low for 5 to 6 h. Replenish- ment of the ER starts around 5 h and ER return to base- line levels around 15-18 h (4). The initial decrease in ER binding (ER content as measured by specific high-affinity binding of estradiol) following an injection of estradiol could be due to a suppression of new ER synthesis or due to alterations in post-transcriptional events such as de- creased translation or increased receptor processing (re- ceptor degradation and hormone-receptor complex uti- lization) or to a combination of the above-mentioned pro- cesses. The relative contribution of each of these processes to the decrease in ER binding is currently unknown. Therefore, the purpose of this study was to investigate the changes in ER mRNA levels during estradiol-induced ER binding depletion and replenishment in the anterior pituitary.
Recent studies in our laboratory have demonstrated that in addition to estradiol, progesterone is also a potent regulator of ER binding in the anterior pituitary (6-8). In the estrogen-primed rat, the administration of proges- terone 1 h before the final estradiol injection brought
487 1044-7431/92 $5.00 Copyright 0 1992 by Academic Press, Inc.
All rights of reproduction in any form reserved.

488 CHORICH ET AL
about a decrease in occupied nuclear estrogen receptors (6-B). Progesterone was also shown to exert a multiphasic effect on gonadotropin secretion depending upon the dose used (9, 10). A similar multiphasic effect of progesterone was also observed in the extent of occupied nuclear pi- tuitary ER binding after various doses of progesterone (8). The progesterone-induced decrease in ER binding in the anterior pituitary appeared to be biologically signifi- cant because it correlated with the loss of estrogen action in progesterone receptor synthesis (7) and induction of prolactin release (11). Similar to the effects of estradiol on ER binding, the progesterone-induced nuclear ER de- pletion in the anterior pituitary observed in our previous studies could be due to either accelerated processing of the ER or inhibition of ER synthesis or both. Therefore, a second objective of our study was to determine changes in anterior pituitary ER mRNA during progesterone-in- duced depletion of ER binding. Furthermore, because progesterone exerts a dose-related multiphasic decrease in occupied nuclear ER binding in the anterior pituitary (B), the effects of various doses of progesterone on anterior pituitary ER mRNA were also examined.
MATERIALS AND METHODS
Animals
Mature, female, virus-free, Holtzman rats (Madison, WI) were obtained at 63 days of age. They were main- tained in an air-conditioned, light-controlled room with a 14-h light:lO-h dark cycle (lights on 0500 h; off 1900 h EST) and were given water and rat chow (Wayne) ad libitum. Bilateral ovariectomy, where indicated, was car- ried out under light ether anesthesia at 65 days (Day 0) of age. The various experimental designs used are sum- marized in Fig. 1 and are briefly described below. All ve- hicle and steroid injections were ip.
Experiment 1. The objective of the first study was to determine if the treatment of ovariectomized rats with estradiol or progesterone or a combination of the two brought about a significant change in P-actin mRNA lev- els. This was necessary because we intended to use p- actin mRNA levels to standardize ER mRNA levels. Two groups of 11 ovariectomized rats received the steroid ve- hicle (0.3 ml, 20% ethanol/BO% saline) for 3 days starting on Day 11 after ovariectomy, while two other groups re- ceived 2 pug of estradiol ip. On Day 14 after ovariectomy, the vehicle-treated rats received either vehicle or 0.8 mg/ kg body wt of progesterone. Both groups received vehicle 1 h later and were killed 1 h after the vehicle injection. The two estrogen-treated groups received either vehicle or 0.8 mg/kg body wt of progesterone on Day 14 after ovariectomy and both groups received 2 pg of estradiol 1 h later. They were killed 1 h after the last estradiol in- jection. @Actin mRNA levels were determined in all four groups.
Experiment 2. The objective of this study was to de- termine the effect of ovariectomy and estrogen treatment on ER mRNA levels. One group of 11 intact adult female rats and two groups of 11 adult ovariectomized rats were used. The intact rats and one group of ovariectomized rats received the estrogen vehicle (0.3 ml, 20% ethanol/ 80% saline) for 3 days starting on Day 76 of age on Day 11 after ovariectomy, while the second group of ovariec- tomized rats received 2 pg of estradiol for 3 days starting on Day 11 after ovariectomy. The animals were killed on Day 79 of age (Day 14 after ovariectomy) and the anterior pituitaries were collected for ER mRNA determination.
Experiment 3. To determine the effect of various doses of estradiol on anterior pituitary ER mRNA levels in ovariectomized rats, groups of 11 ovariectomized rats were injected with vehicle or 1, 5, or 10 /*g of estradiol on Day 14 after ovariectomy. One hour after the injections, the rats were killed by decapitation and the anterior pitui- taries collected for ER mRNA.
Experiment 4. To study the effect of progesterone on ER mRNA in ovariectomized rats, groups of rats were injected with vehicle on Days 11, 12, and 13 and either vehicle or 0.8 mg/kg body wt of progesterone on Day 14 after ovariectomy. After receiving a final vehicle injection 1 h after the vehicle or progesterone injection, the animals were killed 1, 6, 12, or 18 h later and the anterior pitui- taries collected for ER mRNA determination.
Experiment 5. To study the effect of progesterone on acute ER depletion and the effect of estradiol and pro- gesterone on ER replenishment, groups of ovariectomized rats received 2 pg of estradiol for 3 days starting on Day 11 after ovariectomy. On Day 14 after ovariectomy, the rats received either 0.8 mg/kg body wt of progesterone or vehicle. One hour later, all rats received 2 pg of estradiol. All animals were killed 1, 6, 12, or 18 h later and the anterior pituitaries collected for ER mRNA determina- tion. The doses of steroids used and the injection paradigm employed were adopted from Fuentes et al. (B), who mea- sured ER binding under identical conditions 1 h after the last estradiol injection.
Experiment 6. In a final study, three doses of proges- terone 0.8, 2.0, and 4.0 mg/kg body wt or vehicle were injected on Day 14 into ovariectomized rats that were primed for 3 days with 2 pg of estradiol starting on Day 11 after ovariectomy. One hour after the progesterone injection, all groups of rats were injected with 2 pg of estradiol and killed 1 h later. ‘Anterior pituitaries were collected for ER mRNA determination.
Measurement of ER mRNA
Anterior pituitaries were removed immediately after decapitation. Total RNA was prepared by homogenizing anterior pituitaries from 11 rats in RNAzol (100 mg tis- sue/2 ml RNAzol) (Biotecx Laboratories), followed by extraction with chloroform, precipitation with isopropa-

ESTROGEN RECEPTOR mRNA LEVELS IN RAT PITUITARY 489
EXPERIMENTAL PROTOCOLS
(Day 11) (Day 12) (Day 13) (Day 14)
Veh
<
rbVeh 5 Smxiflce
<
Veh -Veh -Veh lh
P4 OVX.
-Veh b sacririce Veh
<
?- E2 - sacrifice E2 - E2 -E2
P4L E2 ” Srxriflce
Intact Veh -Veh -Veh Sacrifice
<
Veh -Veh -Veh Sacrifice
ovx.
E2 - E2 -E2 Sacrifice
Ih E2 lpg - Sacrifice
lh 122 spg - Sacrifice
Ih E2 1Org - SPerlnce
ovx. Veh -Veh -Veh Sacrifice 1,6,12&1Sh
-Veh -
Exot. ovx.
Veh ? E2 _ E2 - E2- E2 -cP
Sacrifice Ih 1,6,12&1Sh
4- E2 -
Exot.
ovx.
Veh lh E2 Ih Sacrifice
0.8 P4 EE2 lh - Sacrifice E2 - E2 - E2 2.0 P4 Ih E2 Ih Sacrifice
4.0 P4 ‘h E2 rh Sacrifice
FIG. 1. Summary of the six protocols used in this study. With the exception of one group in Experiment 2, adult female rats were ovariectomized (OVX on Day 0) on Day 65 of age and treated with vehicle (veh; 0.3 ml, 20% ethanol/80% saline) or estradiol (E2; 2 pg) ip on Days 11, 12, and 13 as shown. On Day 14 they were killed either without further treatment (Experiment 2) or 1 h after various doses of estradiol (Experiment 3) or various combinations of vehicle, progesterone (P,; 0.8 mg/kg body wt), or estradiol (Ex; 2 pg) treatment as shown (Experiments 4 and 5). In Experiment 6, different doses of progesterone were used.
nol, and ethanol washes, according to the manufacturer’s every 1.5 h, and transferred by capillary action to a nylon protocol (12). Initial studies utilized Northern blotting membrane (Gene Screen Plus, New England Nuclear Re- for size confirmation of the rat and human ER mRNA. search Products) with 10X SSC (1X SSC: 0.15 M sodium Total RNA from T47D human breast cancer cells was chloride (NaCl) , 15 n&f sodium citrate). Slot blotting was used as a positive ER mRNA control in the Northern subsequently chosen for message quantification. For slot blots. Total RNA (20 pg/sample) was denatured by blots, samples of total RNA (20 pg) were denatured by glyoxalation, subjected to electrophoresis in a 1.5% aga- glyoxalation and blotted to nylon membranes. Slot blots rose gel with low ionic strength buffer which was changed were sequentially hybridized with cDNA probes for the

490 CHORICH ET AL.
rat ER and P-actin. Slot blots were stripped between hybridizations with an initial wash of 10 mM Tris(hydroxymethyl)aminomethane hydrochloride (Tris- HCl) pH 7.5,l mM disodium ethylenediaminetetraacetate (EDTA), 1% (wt/vol) sodium dodecyl sulfate (SDS) at 100°C followed by a second wash with 0.1X SSC con- taining 0.1% (wt/vol) SDS at lOO”C, allowing each wash to cool to 30°C with agitation. This was followed by a final wash of 2X SSPE (20X SSPE: 3.6 M NaCl, 0.2 M sodium phosphate dibasic, pH 6.8, 0.02 M EDTA) for 30 min with agitation. The plasmid harboring the 2.1-kb rat ER cDNA, pSVBER, was generously provided by M. Sakai (University of Tokyo, Tokyo, Japan) (13). P. Chambon (National Institutes of Medical Research, Strasbourg, France) (14) kindly provided pKCR2-ER, containing the hER cDNA probe of 1.8kb size. N. Shimizu (University of Chicago, Chicago, IL) (15) donated the pp-actin plasmid hosting the 1.15-kb P-actin cDNA which was used to con- trol for RNA levels. Each cDNA insert was excised from its plasmid construct by digestion with EcoRI, EcoRI, and PstI, respectively. Electrophoresis of the digests was car- ried out in 1.5% low-melting-point agarose in TEA buffer (40 mM Tris base, 20 mM sodium acetate, 2 mM EDTA, final pH 7.5). The 1.8kb human ER cDNA fragment, the 2.1-kb rat ER cDNA fragment, and the 1.15-kb P-actin fragment were excised and labeled in agarose with random primers (Pharmacia, Oligolabelling Kit) and [a-32P]dCTP (ICN, 3000 Ci/mmol). Probe specific activities were l-4 X lo8 Cerenkov cpm/pg DNA. Hybridization was carried out in 50% (vol/vol) formamide, 10% (wt/vol) Dextran sulfate, 50 mM Tris, pH 7.5, 1% (wt/vol) SDS, 1 M NaCl, and 100 pg/ml sheared, denatured salmon sperm DNA for 18 h at 42°C. Subsequently, membranes were washed three times with agitation in 0.2X SSC, 0.1% (wt/vol) SDS, the first at room temperature and the second and third at 60°C. The membrane was then washed in 2~ SSPE at room temperature and subjected to autoradi- ography. Hybridized band intensities were determined by scanning the autoradiograph with a Schimadzu densi- tometer. Levels of specific mRNA were expressed in ar- bitrary densitometric units and normalized to the level of hybridized ,&actin mRNA.
Measurement of ER Binding
Estradiol receptor binding was performed according to Zamorano et al. (16). Briefly, anterior pituitaries were placed in cold homogenization buffer (TEDSP; 10 mM Tris-HCl, 1.5 mM EDTA, 1.0 mM dithiothreitol (DTT), 250 mM sucrose, and 1 mMphenylmethylsulfony1 fluoride (PMSF), pH 7.4; 0.8 ml/per pituitary) and homogenized manually on ice with 10 strokes of a Teflon/glass ho- mogenizer (clearance, 180 PM). Nuclear pellets were ob- tained by centrifuging the homogenate at 8OOg for 10 min. The supernatants were then centrifuged at 105,OOOg for 1 h to obtain the cytosolic fraction. Nuclear pellets were washed three times and resuspended in nuclear buffer
(TDSPM; Tris-HCl 10 mM, 1.0 mM DTT, 250 mM su- crose, 1 mM PMSF, and 2.5 mM magnesium chloride (MgC12), pH 7.4; 0.4 m per pituitary) with a Dounce ho- mogenizer (pestle B) prior to the binding assays. Receptor binding assays were performed by incubating 250 ~1 of cytosol or nuclear suspension with five different concen- trations of [3H]estradiol (0.1-2.5 nM) in a final volume of 500 ~1. Nonspecific binding was measured in the pres- ence of lOO-fold molar excess of diethylstilbestrol. Total nuclear receptor determination was carried out at 30°C for 1 h, followed by 1 h at O”C, and the total cytosolic receptor binding assay was incubated at 22’C for 18 h, followed by 1 h at 0°C (17). Nuclear incubation was stopped with 1 ml of cold nuclear buffer and the unbound hormone was removed by three washes of 1 ml TDSPM buffer. Bound hormone was extracted by incubating with 1 ml ethanol at 30°C for 30 min. After centrifugation at 1OOOg for 10 min, the ethanol supernatants were added to 10 ml scintillation cocktail. Cytosolic incubation was terminated by incubating with 0.5 ml dextran-coated charcoal (0.5% Norit A, 0.5% dextran) for 10 min at 0°C. The dextran-coated charcoal was removed by centrifu- gation at 1OOOg for 10 min. The supernatant was added to 10 ml scintillation cocktail and 1 ml ethanol and counted for 3H content using a Beckman LS-5801 spec- trometer with 45% efficiency for tritium. Results were normalized per milligram of DNA or protein content of nuclei and cytosol, respectively. The DNA content was determined by the method of Burton (18) and the protein content by the method of Lowry et al. (19). The ER bind- ing activity was calculated by using Scatchard analysis (20). Final values were calculated on a per pituitary basis. This was done because it represents the total receptor content of the cytosol and the nucleus and therefore is a good index of the total change in the tissue. Because the ER binding patterns after an injection of estradiol have been reported extensively in literature (l-8), a single de- termination using a 5-point Scatchard plot was carried out to document the results at 1, 6, 12, and 18 h under our experimental conditions.
Statistical Analysis
The results given in the text are expressed as the mean + SE. The differences between experimental groups were analyzed using the Student t test when two groups were compared. Analysis of variance followed by the Fisher PLSD test was used when more than two groups were compared. Differences were considered to be significant at P < 0.05.
RESULTS
Establishment of Techniques for Measuring ER mRNA
Northern blots of total RNA extracted from uterine tissue were prepared and hybridized with 32P-labeled rat

ESTROGEN RECEPTOR mRNA LEVELS IN RAT PITUITARY 491
and human ER cDNA. A prominent 6.4-kb ER species TABLE 1 was identified along with a faint band at 5.5 kb (Fig. 2). In all groups the 5.5kb band was too faint to obtain a reliable densitometric measurement. The slot blotting method was used for the auantitation of ER mRNA levels -
Effect of Estradiol, Progesterone, and Estradiol plus Progesterone on fi-Actin Message Levels
in Anterior Pituitary RNA
after standardizing the conditions of hybridization. Groups
Appropriateness of the Use of /3-Actin mRNA Levels to Standardize ER mRNA Content
Before @-actin mRNA levels were used to standardize ER mRNA content, it was considered appropriate to es- tablish that they did not change in the anterior pituitary after estradiol and progesterone treatment. The results in Table 1 show that @-actin mRNA levels after estradiol, progesterone, and a combination of estradiol and proges- terone treatment, expressed as the percentage of vehicle controls, did not show significant differences from each other. Therefore, the results of ER mRNA content in an- terior pituitary tissue were normalized to P-actin mRNA levels.
Effect of Ovariectomy and 3-Day E&radio1 Priming on Anterior Pituitary ER mRNA Levels
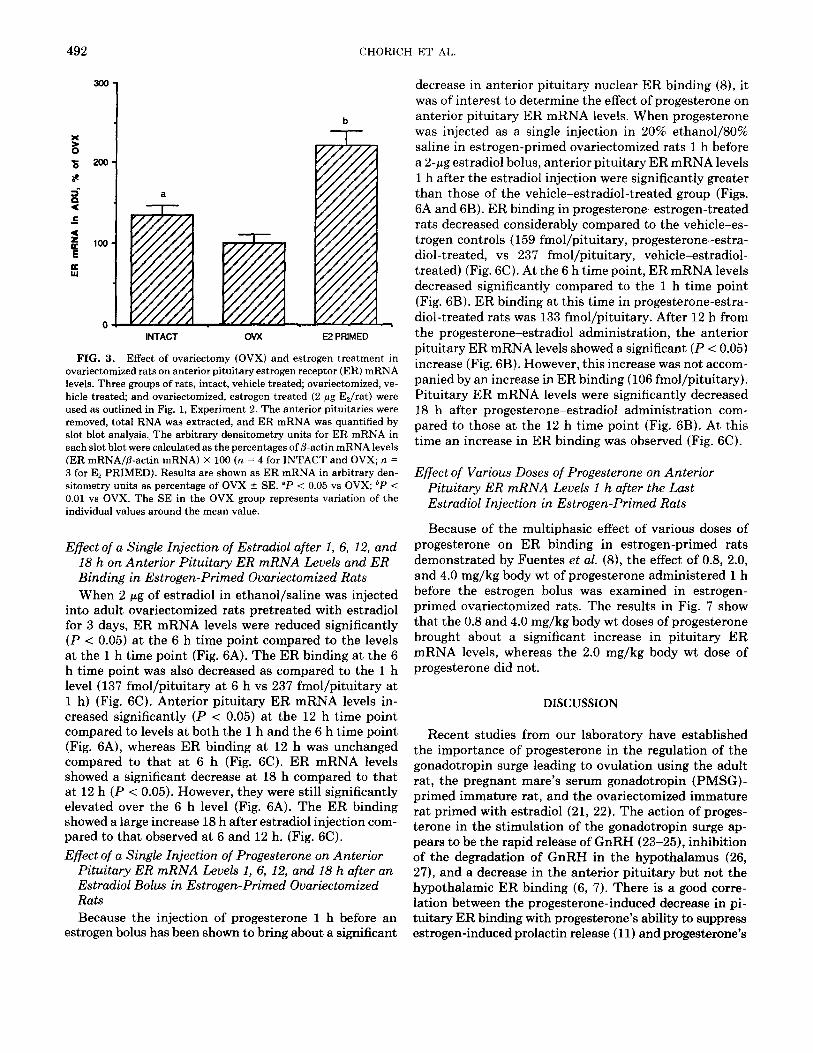
To determine whether endogenous estrogens regulate ER mRNA levels of the anterior pituitary, ER mRNA levels were measured in randomly cycling adult female rats, rats ovariectomized for 14 days, and rats ovariec- tomized for 14 days that received 2 pg/day of estradiol during the last 3 days. As shown in Fig. 3, ovariectomy for 14 days resulted in a significant decrease in anterior pituitary ER mRNA levels compared to intact controls.
kb
6.4
5.5
FIG. 2. Northern blots from two anterior pituitary RNA extracts hybridized with labeled 2.1-kb rat estrogen receptor (ER) cDNA frag- ment. Note the prominent 6.4-kb ER band and the barely visible 5.5- kb band.
Veh E2 p4 E2 plus P4 Veh Kz veh Veh
fi-Actin 1.00 + 0.28 1.13 z!c 0.10 1.26 f 0.09 1.29 * 0.12 n=7 n=6 n=4 n=6
Note. Total anterior pituitary RNA was extracted from ovariectomized rats treated with vehicle, vehicle and 0.8 mg/kg body weight progesterone (P4), estradiol (Ez, 2 pg/rat), or Ez plus P, as outlined in Fig. 1, Exper- iment 1. Slot blots of this mRNA were hybridized to a j3-actin probe. fl-Actin values for each hormone treatment were divided by the mean vehicle /3-actin value. Results are expressed as the mean + SE. No sig- nificant difference was found between the various groups. The SE of the Veh/Veh group represents the variation of individual values around the mean value.
Treatment with estradiol for 3 days brought about a sig- nificant increase in anterior pituitary ER mRNA levels.
Acute Effects of Various Doses of Estradiol on Anterior Pituitary ER mRNA Levels in Ovariectomized Adult Rats
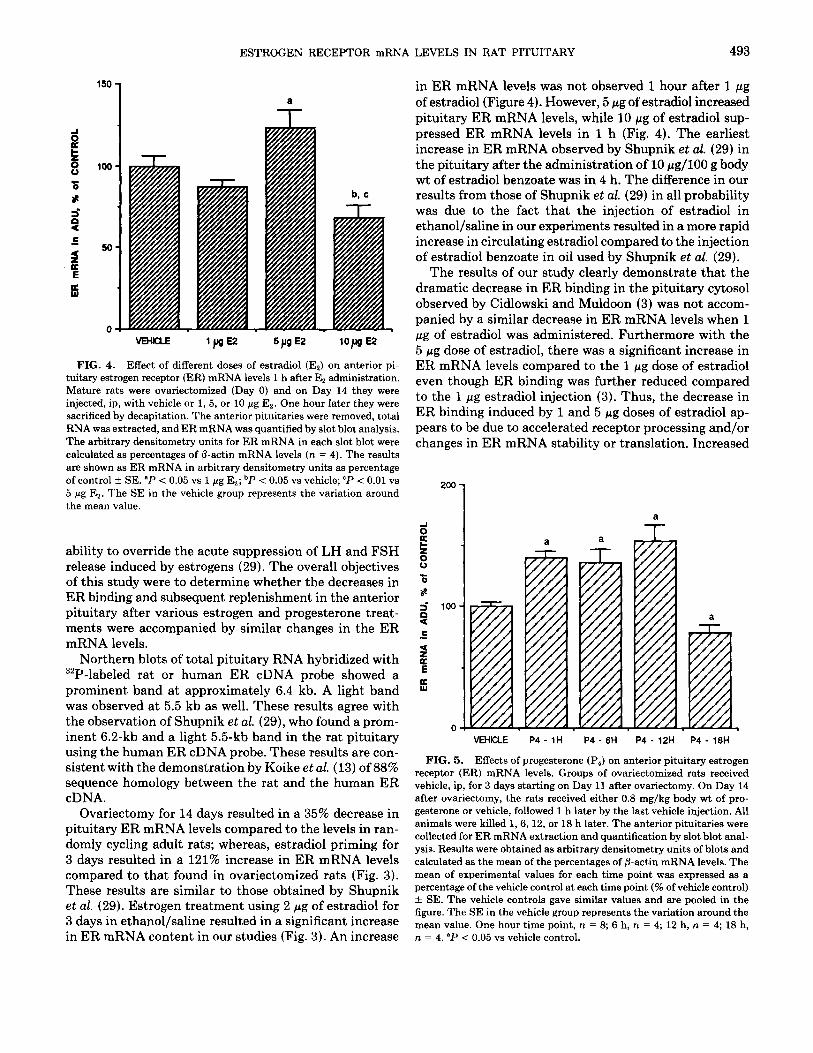
Based on the ER binding studies of Cidlowski and Muldoon (3) that showed an acute decrease in ER binding in the anterior pituitary cytosol after 1 h of estradiol ad- ministration, three doses of estradiol (1, 5, and 10 pg) were injected into adult ovariectomized rats and anterior pituitary ER mRNA levels were determined 1 h later. The administration of 1 wg of estradiol did not bring about any changes in anterior pituitary ER mRNA levels at the 1 h time point (Fig. 4). The administration of 5 pg of estradiol resulted in a significant (P < 0.05) increase in anterior pituitary ER mRNA levels compared to the 1 pg dose of estradiol, while the 10 pg dose brought about a significant (P < 0.05) reduction of ER mRNA levels after 1 h (Fig. 4).
Effect of Progesterone on Anterior Pituitary ER mRNA Levels in the Nonestrogen-Treated Ovariectomized Rat
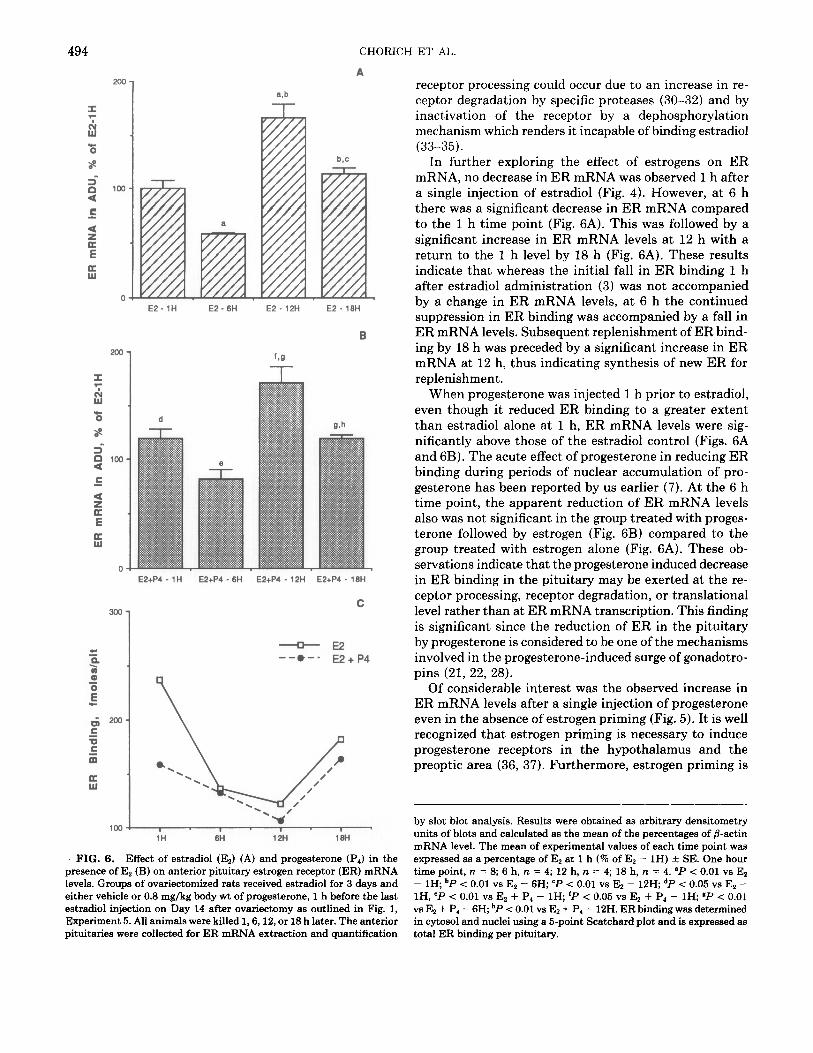
In the ovariectomized rat, the administration of 0.8 mg/ kg body wt of progesterone brought about a stimulatory effect on ER mRNA levels of the anterior pituitary at the 1, 6, and 12 h time points (P < 0.05) (Fig. 5). The mean ER mRNA content was 140% of the control at 1 h, 136% of the control at 6 h, and 153% of the control at 12 h. At the 18 h time point, progesterone significantly (P < 0.05) reduced the ER mRNA content in the anterior pituitary, the levels being 78% of the control (Fig. 5).
492 CHORICH ET AL
300
200
a
b
INTACT OVX E2 PRIMED
FIG. 3. Effect of ovariectomy (OVX) and estrogen treatment in ovariectomized rats on anterior pituitary estrogen receptor (ER) mRNA levels. Three groups of rats, intact, vehicle treated; ovariectomized, ve- hicle treated; and ovariectomized, estrogen treated (2 pg E&at) were used as outlined in Fig. 1, Experiment 2. The anterior pituitaries were removed, total RNA was extracted, and ER mRNA was quantified by slot blot analysis. The arbitrary densitometry units for ER mRNA in each slot blot were calculated as the percentages of p-actin mRNA levels (ER mRNA/P-actin mRNA) X 100 (n = 4 for INTACT and OVX; n = 3 for E2 PRIMED). Results are shown as ER mRNA in arbitrary den- sitometry units as percentage of OVX + SE. “P < 0.05 vs OVX; bP < 0.01 vs OVX. The SE in the OVX group represents variation of the individual values around the mean value.
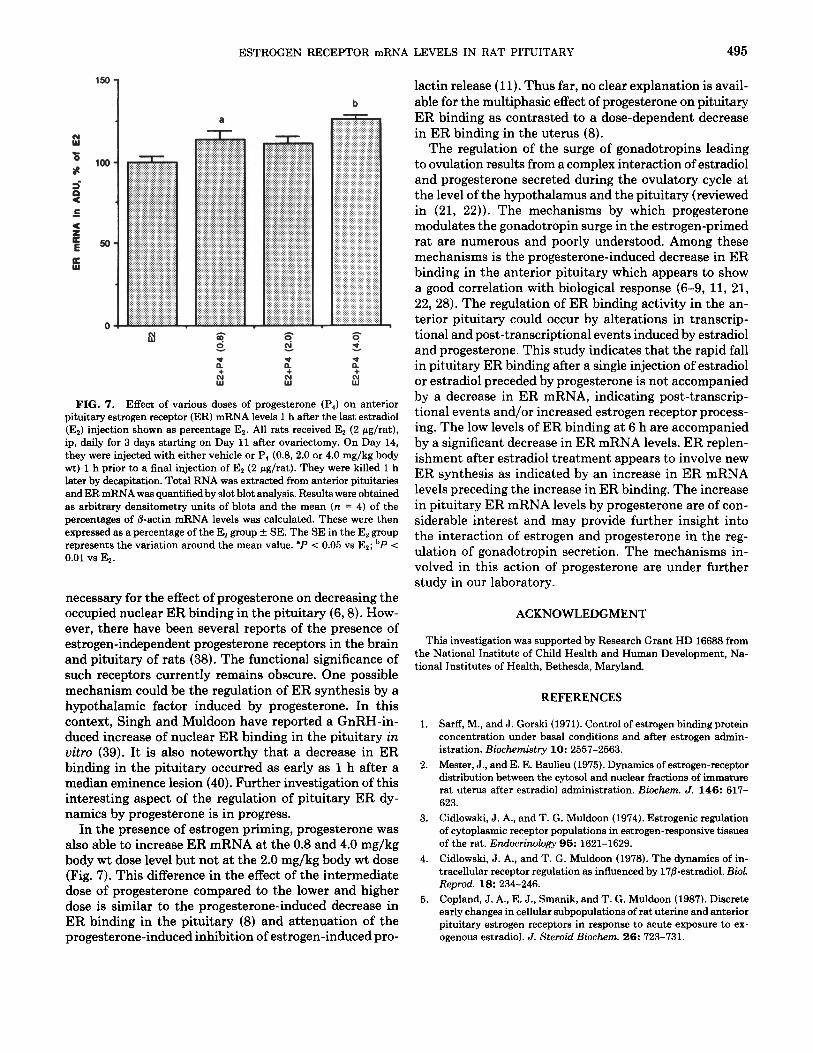
Effect of a Single Injection of Estradiol after 1, 6, 12, and 18 h on Anterior Pituitary ER mRNA Levels and ER Binding in Estrogen-Primed Ovariectomized Rats When 2 pg of e&radio1 in ethanol/saline was injected
into adult ovariectomized rats pretreated with e&radio1 for 3 days, ER mRNA levels were reduced significantly (P < 0.05) at the 6 h time point compared to the levels at the 1 h time point (Fig. 6A). The ER binding at the 6 h time point was also decreased as compared to the 1 h level (137 fmol/pituitary at 6 h vs 237 fmol/pituitary at 1 h) (Fig. 6C). Anterior pituitary ER mRNA levels in- creased significantly (P < 0.05) at the 12 h time point compared to levels at both the 1 h and the 6 h time point (Fig. 6A), whereas ER binding at 12 h was unchanged compared to that at 6 h (Fig. 6C). ER mRNA levels showed a significant decrease at 18 h compared to that at 12 h (P < 0.05). However, they were still significantly elevated over the 6 h level (Fig. 6A). The ER binding showed a large increase 18 h after estradiol injection com- pared to that observed at 6 and 12 h. (Fig. 6C). Effect of a Single Injection of Progesterone on Anterior
Pituitary ER mRNA Levels 1, 6, 12, and 18 h after an Estradiol Bolus in Estrogen-Primed Ovariectomized Rats Because the injection of progesterone 1 h before an
estrogen bolus has been shown to bring about a significant
decrease in anterior pituitary nuclear ER binding (8), it was of interest to determine the effect of progesterone on anterior pituitary ER mRNA levels. When progesterone was injected as a single injection in 20% ethanol/80% saline in estrogen-primed ovariectomized rats 1 h before a 2-pg e&radio1 bolus, anterior pituitary ER mRNA levels 1 h after the e&radio1 injection were significantly greater than those of the vehicle-estradiol-treated group (Figs. 6A and 6B). ER binding in progesterone-estrogen-treated rats decreased considerably compared to the vehicle-es- trogen controls (159 fmol/pituitary, progesterone-estra- diol-treated, vs 237 fmol/pituitary, vehicle-estradiol- treated) (Fig. 6C). At the 6 h time point, ER mRNA levels decreased significantly compared to the 1 h time point (Fig. 6B). ER binding at this time in progesterone-estra- diol-treated rats was 133 fmol/pituitary. After 12 h from the progesterone-estradiol administration, the anterior pituitary ER mRNA levels showed a significant (P < 0.05) increase (Fig. 6B). However, this increase was not accom- panied by an increase in ER binding (106 fmol/pituitary). Pituitary ER mRNA levels were significantly decreased 18 h after progesterone-estradiol administration com- pared to those at the 12 h time point (Fig. 6B). At this time an increase in ER binding was observed (Fig. 6C).
Effect of Various Doses of Progesterone on Anterior Pituitary ER mRNA Levels 1 h after the Last Estradiol Injection in Estrogen-Primed Rats
Because of the multiphasic effect of various doses of progesterone on ER binding in estrogen-primed rats demonstrated by Fuentes et al. (8), the effect of 0.8, 2.0, and 4.0 mg/kg body wt of progesterone administered 1 h before the estrogen bolus was examined in estrogen- primed ovariectomized rats. The results in Fig. 7 show that the 0.8 and 4.0 mg/kg body wt doses of progesterone brought about a significant increase in pituitary ER mRNA levels, whereas the 2.0 mg/kg body wt dose of progesterone did not.
DISCUSSION
Recent studies from our laboratory have established the importance of progesterone in the regulation of the gonadotropin surge leading to ovulation using the adult rat, the pregnant mare’s serum gonadotropin (PMSG)- primed immature rat, and the ovariectomized immature rat primed with estradiol (21, 22). The action of proges- terone in the stimulation of the gonadotropin surge ap- pears to be the rapid release of GnRH (23-25), inhibition of the degradation of GnRH in the hypothalamus (26, 27), and a decrease in the anterior pituitary but not the hypothalamic ER binding (6, 7). There is a good corre- lation between the progesterone-induced decrease in pi- tuitary ER binding with progesterone’s ability to suppress estrogen-induced prolactin release (11) and progesterone’s
ESTROGEN RECEPTOR mRNA LEVELS IN RAT PITUITARY 493
v -v- -r
VEHCLE ‘Pm 5pgE2 lop E2
FIG. 4. Effect of different doses of estradiol (Es) on anterior pi- tuitary estrogen receptor (ER) mRNA levels 1 h after E, administration. Mature rats were ovariectomized (Day 0) and on Day 14 they were injected, ip, with vehicle or 1, 5, or 10 pg Es. One hour later they were sacrificed by decapitation. The anterior pituitaries were removed, total RNA was extracted, and ER mRNA was quantified by slot blot analysis. The arbitrary densitometry units for ER mRNA in each slot blot were calculated as percentages of fl-actin mRNA levels (n = 4). The results are shown as ER mRNA in arbitrary densitometry units as percentage of control + SE. “P < 0.05 vs 1 pg Es; bP i 0.05 vs vehicle; "P < 0.01 vs 5 pg Er. The SE in the vehicle group represents the variation around the mean value.
ability to override the acute suppression of LH and FSH release induced by estrogens (29). The overall objectives of this study were to determine whether the decreases in ER binding and subsequent replenishment in the anterior pituitary after various estrogen and progesterone treat- ments were accompanied by similar changes in the ER mRNA levels.
Northern blots of total pituitary RNA hybridized with 32P-labeled rat or human ER cDNA probe showed a prominent band at approximately 6.4 kb. A light band was observed at 5.5 kb as well. These results agree with the observation of Shupnik et al. (29), who found a prom- inent 6.2-kb and a light 5.5kb band in the rat pituitary using the human ER cDNA probe. These results are con- sistent with the demonstration by Koike et al. (13) of 88% sequence homology between the rat and the human ER cDNA.
Ovariectomy for 14 days resulted in a 35% decrease in pituitary ER mRNA levels compared to the levels in ran- domly cycling adult rats; whereas, estradiol priming for 3 days resulted in a 121% increase in ER mRNA levels compared to that found in ovariectomized rats (Fig. 3). These results are similar to those obtained by Shupnik et al. (29). Estrogen treatment using 2 pg of estradiol for 3 days in ethanol/saline resulted in a significant increase in ER mRNA content in our studies (Fig. 3). An increase
in ER mRNA levels was not observed 1 hour after 1 pg of estradiol (Figure 4). However, 5 pg of estradiol increased pituitary ER mRNA levels, while 10 pg of estradiol sup- pressed ER mRNA levels in 1 h (Fig. 4). The earliest increase in ER mRNA observed by Shupnik et al. (29) in the pituitary after the administration of 10 @g/100 g body wt of estradiol benzoate was in 4 h. The difference in our results from those of Shupnik et al. (29) in all probability was due to the fact that the injection of estradiol in ethanol/saline in our experiments resulted in a more rapid increase in circulating estradiol compared to the injection of estradiol benzoate in oil used by Shupnik et al. (29).
The results of our study clearly demonstrate that the dramatic decrease in ER binding in the pituitary cytosol observed by Cidlowski and Muldoon (3) was not accom- panied by a similar decrease in ER mRNA levels when 1 pg of estradiol was administered. Furthermore with the 5 pg dose of estradiol, there was a significant increase in ER mRNA levels compared to the 1 pg dose of estradiol even though ER binding was further reduced compared to the 1 pg estradiol injection (3). Thus, the decrease in ER binding induced by 1 and 5 pg doses of estradiol ap- pears to be due to accelerated receptor processing and/or changes in ER mRNA stability or translation. Increased
d
ii! E
6
a l-
VEHKLE P4- IH P4 - 6H P4 - 12H P4 - 18H
FIG. 5. Effects of progesterone (P,) on anterior pituitary estrogen receptor (ER) mRNA levels. Groups of ovariectomized rats received vehicle, ip, for 3 days starting on Day 11 after ovariectomy. On Day 14 after ovariectomy, the rats received either 0.8 mg/kg body wt of pro- gesterone or vehicle, followed 1 h later by the last vehicle injection. All animals were killed 1, 6, 12, or 18 h later. The anterior pituitaries were collected for ER mRNA extraction and quantification by slot blot anal- ysis. Results were obtained as arbitrary densitometry units of blots and calculated as the mean of the percentages of fl-actin mRNA levels. The mean of experimental values for each time point was expressed as a percentage of the vehicle control at each time point (% of vehicle control) + SE. The vehicle controls gave similar values and are pooled in the figure. The SE in the vehicle group represents the variation around the mean value. One hour time point, n = 8; 6 h, n = 4; 12 h, n = 4; 18 h, n = 4. “P < 0.05 vs vehicle control.
494 CHORICH ET AL
EZ-1H E2 - BH E2 - 12H E2-18H
B
E2+P4 - 1H E2+P4 - 6H E2tP4 - 12H E2+P4 - 18H
C
-E2 -me-. E2 + P4
FIG. 6. Effect of e&radio1 (E,) (A) and progesterone (PI) in the presence of Er (B) on anterior pituitary estrogen receptor (ER) mRNA levels. Groups of ovariectomized rats received e&radio1 for 3 days and either vehicle or 0.8 mg/kg body wt of progesterone, 1 h before the last estradiol injection on Day 14 after ovariectomy as outlined in Fig. 1, Experiment 5. All animals were killed 1,6,12, or 18 h later. The anterior pituitaries were collected for ER mRNA extraction and quantification
receptor processing could occur due to an increase in re- ceptor degradation by specific proteases (30-32) and by inactivation of the receptor by a dephosphorylation mechanism which renders it incapable of binding estradiol (33-35).
In further exploring the effect of estrogens on ER mRNA, no decrease in ER mRNA was observed 1 h after a single injection of estradiol (Fig. 4). However, at 6 h there was a significant decrease in ER mRNA compared to the 1 h time point (Fig. 6A). This was followed by a significant increase in ER mRNA levels at 12 h with a return to the 1 h level by 18 h (Fig. 6A). These results indicate that whereas the initial fall in ER binding 1 h after estradiol administration (3) was not accompanied by a change in ER mRNA levels, at 6 h the continued suppression in ER binding was accompanied by a fall in ER mRNA levels. Subsequent replenishment of ER bind- ing by 18 h was preceded by a significant increase in ER mRNA at 12 h, thus indicating synthesis of new ER for replenishment.
When progesterone was injected 1 h prior to estradiol, even though it reduced ER binding to a greater extent than estradiol alone at 1 h, ER mRNA levels were sig- nificantly above those of the estradiol control (Figs. 6A and 6B). The acute effect of progesterone in reducing ER binding during periods of nuclear accumulation of pro- gesterone has been reported by us earlier (7). At the 6 h time point, the apparent reduction of ER mRNA levels also was not significant in the group treated with proges- terone followed by estrogen (Fig. 6B) compared to the group treated with estrogen alone (Fig. 6A). These ob- servations indicate that the progesterone induced decrease in ER binding in the pituitary may be exerted at the re- ceptor processing, receptor degradation, or translational level rather than at ER mRNA transcription. This finding is significant since the reduction of ER in the pituitary by progesterone is considered to be one of the mechanisms involved in the progesterone-induced surge of gonadotro- pins (21, 22, 28).
Of considerable interest was the observed increase in ER mRNA levels after a single injection of progesterone even in the absence of estrogen priming (Fig. 5). It is well recognized that estrogen priming is necessary to induce progesterone receptors in the hypothalamus and the preoptic area (36, 37). Furthermore, estrogen priming is
by slot blot analysis. Results were obtained as arbitrary densitometry units of blots and calculated as the mean of the percentages of @actin mRNA level. The mean of experimental values of each time point was expressed as a percentage of Es at 1 h (% of Es - 1H) f SE. One hour time point, n = 8; 6 h, n = 4; 12 h, n = 4; 18 h, n = 4. ‘P < 0.01 vs Er - 1H; bP < 0.01 vs Es - 6H; 'P < 0.01 vs Ez - 12H; dP < 0.05 vs El - lH, 'P < 0.01 vs Es + P, - 1H; 'P < 0.05 vs Ez + Pd - 1H; gP < 0.01 vs Ez + P, - 6H; hP < 0.01 vs Es + P, - 12H. ER binding was determined in cytosol and nuclei using a B-point Scatchard plot and is expressed as total ER binding per pituitary.
ESTROGEN RECEPTOR mRNA
i L
lOO-
!s-
OS
a 7
2 x 2 + + + D z D FIG. 7. Effect of various doses of progesterone (P4) on anterior
pituitary estrogen receptor (ER) mRNA levels 1 h after the last estradiol (Er) injection shown as percentage Ex. All rats received E2 (2 pg/rat), ip, daily for 3 days starting on Day 11 after ovariectomy. On Day 14, they were injected with either vehicle or P1 (0.8, 2.0 or 4.0 mg/kg body wt) 1 h prior to a final injection of Er (2 pg/rat). They were killed 1 h later by decapitation. Total RNA was extracted from anterior pituitaries and ER mRNA was quantified by slot blot analysis. Results were obtained as arbitrary densitometry units of blots and the mean (n = 4) of the percentages of &actin mRNA levels was calculated. These were then expressed as a percentage of the Ex group + SE. The SE in the E2 group represents the variation around the mean value. “P < 0.05 vs E,; bP < 0.01 vs Es.
necessary for the effect of progesterone on decreasing the occupied nuclear ER binding in the pituitary (6,8). How- ever, there have been several reports of the presence of estrogen-independent progesterone receptors in the brain and pituitary of rats (38). The functional significance of such receptors currently remains obscure. One possible mechanism could be the regulation of ER synthesis by a hypothalamic factor induced by progesterone. In this context, Singh and Muldoon have reported a GnRH-in- duced increase of nuclear ER binding in the pituitary in vitro (39). It is also noteworthy that a decrease in ER binding in the pituitary occurred as early as 1 h after a median eminence lesion (40). Further investigation of this interesting aspect of the regulation of pituitary ER dy- namics by progesterone is in progress.
In the presence of estrogen priming, progesterone was also able to increase ER mRNA at the 0.8 and 4.0 mg/kg body wt dose level but not at the 2.0 mg/kg body wt dose (Fig. 7). This difference in the effect of the intermediate dose of progesterone compared to the lower and higher dose is similar to the progesterone-induced decrease in ER binding in the pituitary (8) and attenuation of the progesterone-induced inhibition of estrogen-induced pro-
LEVELS IN RAT PITUITARY 495
lactin release (11). Thus far, no clear explanation is avail- able for the multiphasic effect of progesterone on pituitary ER binding as contrasted to a dose-dependent decrease in ER binding in the uterus (8).
The regulation of the surge of gonadotropins leading to ovulation results from a complex interaction of estradiol and progesterone secreted during the ovulatory cycle at the level of the hypothalamus and the pituitary (reviewed in (21, 22)). The mechanisms by which progesterone modulates the gonadotropin surge in the estrogen-primed rat are numerous and poorly understood. Among these mechanisms is the progesterone-induced decrease in ER binding in the anterior pituitary which appears to show a good correlation with biological response (6-9, 11, 21, 22, 28). The regulation of ER binding activity in the an- terior pituitary could occur by alterations in transcrip- tional and post-transcriptional events induced by estradiol and progesterone. This study indicates that the rapid fall in pituitary ER binding after a single injection of estradiol or estradiol preceded by progesterone is not accompanied by a decrease in ER mRNA, indicating post-transcrip- tional events and/or increased estrogen receptor process- ing. The low levels of ER binding at 6 h are accompanied by a significant decrease in ER mRNA levels. ER replen- ishment after estradiol treatment appears to involve new ER synthesis as indicated by an increase in ER mRNA levels preceding the increase in ER binding. The increase in pituitary ER mRNA levels by progesterone are of con- siderable interest and may provide further insight into the interaction of estrogen and progesterone in the reg- ulation of gonadotropin secretion. The mechanisms in- volved in this action of progesterone are under further study in our laboratory.
ACKNOWLEDGMENT
This investigation was supported by Research Grant HD 16666 from the National Institute of Child Health and Human Development, Na- tional Institutes of Health, Bethesda, Maryland.
1.
2.
3.
4.
5.
REFERENCES
Sarff, M., and J. Gorski (1971). Control of estrogen binding protein concentration under basal conditions and after estrogen admin- istration. Biochemistry 10: 2557-2563. Me&r, J., and E. E. Baulieu (1975). Dynamics of estrogen-receptor distribution between the cytosol and nuclear fractions of immature rat uterus after estradiol administration. Biochem. J. 146: 617- 623. Cidlowski, J. A., and T. G. Muldoon (1974). Estrogenic regulation of cytoplasmic receptor populations in estrogen-responsive tissues of the rat. Endocrinology 95: 1621-1629. Cidlowski, J. A., and T. G. Muldoon (1978). The dynamics of in- tracellular receptor regulation as influenced by 17fl-estradiol. Biol. Reprod. l&234-246. Copland, J. A., E. J., Smanik, and T. G. Muldoon (1987). Discrete early changes in cellular subpopulations of rat uterine and anterior pituitary estrogen receptors in response to acute exposure to ex- ogenous estradiol. J. Steroid Biochem. 26: 723-731.

496 CHORICH ET AL.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
Smanik, E., K. Young, T. G. Muldoon, and V. B. Mahesh (1983). Analysis of the effect of progesterone in vivo on estrogen receptor distribution in the rat anterior pituitary and hypothalamus. En- docrinology 113: 15-22. Calderon, J. J., T. G. Muldoon, and V. B. Mahesh (1987). Receptor- mediated interrelationships between progesterone and estradiol action on the anterior pituitary-hypothalamic axis of the ovari- ectomized rat. Endocrinology 120: 2428-2435. Fuentes, M. A., T. G. Muldoon, and V. B. Mahesh (1990). Inhibitory effect of progesterone on occupied estrogen receptors of anterior pituitary and uterus in adult rats. J. Neuroendocrinol. 2: 517-522. McPherson, J., A. Costoff, J. Eldridge, and V. B. Mahesh (1974). Effects of various progestational preparations on gonadotropin se- cretion in ovariectomized immature female rats. Fertil. Steril. 25: 1063-1070. McPherson, J. C., and V. B. Mahesh (1979). Dose-related effect of a single injection of progesterone on gonadotropin secretion and pituitary sensitivity to LHRH in estrogen-primed castrated female rats. Biol. Reprod. 20: 763-772. Brann, D. W., C. D. Putnam, and V. B. Mahesh (1990). Dose related effects of progesterone and 5-a-dihydroprogesterone upon estrogen- induced prolactin release. J. Neuroendocrinol. 2: 341-345. Chomczynski, P., and N. Sacchi (1987). Single-step methodof RNA isolation by acid guanidium thiocyanate-phenolchloroform ex- traction. Anal. Biochem. 162: 156-159. Koike, S., M. Sakai, and M. Muramatou (1987). Molecular cloning and characterization of rat estrogen receptor cDNA. Nucleic Acids Res. 15: 2499-2513. Green, S., P. Walker, V. Kumar, A. Krust, J. M. Bornert, P. Argos, and P. Chambon (1986). Oestrogen receptor cDNA: Sequence, expression and homology to v-erb-A. Nature 320: 134-139. Shimizu, N., S. Kamel-Reid, and R. Zak (1988). Expression of actin mRNAs in denervated chicken skeletal muscle. Den Biol. 128:435-440. Zamorano, P., J. Steinsapir, and V. B. Mahesb (1992). Effect of 5o-dihydrotestosterone and dexamethasone on estrogen receptors of anterior pituitary and uterus. Steroids 57: 18-26. Smanik, E. J., J. A. Copland, and T. G. Muldoon (1984). Cryptic estrogen binding protein complicates analysis of estrogen receptor distribution. Biochem. Biophys. Res. Commun. 125: 577-583. Burton, K. (1956). A study of the conditions and mechanism of the diphenylamine reaction for the calorimetric estimation of de- oxyribonucleic acid. Biochem. J. 62: 315-323. Lowry, 0. H., N. J. Rosengrough, A. L. Furr, and R. J. Randall (1951). Protein measurement with the Folin phenol reagent. J. Biol.Chem. 193:265-275. Scatchard, G. (1949). The attraction of proteins for small molecules and ions. Ann. N. Y. Acad. Sci. 51: 660-672. Brann, D. W., and V. B. Mahesh (1991). Regulation of gonadotropin secretion by steroid hormones. Front. Neuroendocrinol. 12: 165- 207. Mahesh, V. B., and D. W. Brann (1992). Interaction between ovar- ian and adrenal steroids in the regulation of gonadotropin secretion. J. Steroid Biochem. Mol. Biol. 41: 495-513. Peduto, J. C., and V. B. Mahesh (1985). Effects of progesterone on hypothalamic and plasma LHRH. Neuroendocrinology 40: 238- 245. Kim, K., and V. D. Ramirez (1982). In vitro progesterone stimulates the release of luteinizing hormone-releasing hormone from super- fused hypothalamic tissue from ovariectomized estradiol-primed prepubertal rats. Endocrinology 111: 750-757.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
Levine, J. E., and V. D. Ramirez (1980). In vivo release of luteinizing hormone-releasing hormone estimated with push-pull cannulae from the mediobasal hypothalamus of ovariectomized steroid primed rats. Endocrinology 107: 1782-1790. O’Conner, J. L., and V. B. Mahesh (1988). A possible role for pro- gesterone in the preovulatory gonadotropin surge through modu- lation of LHRH degrading activity. J. Steroid Biochem. 29: 257- 263. Advis, J. P., J. E. Krause, and J. F. McKelvy (1982). Luteinizing hormone-releasing hormone peptidase activities in discrete hy- pothalamic regions and anterior pituitary of the rat: Apparent reg- ulation during the prepubertal period and first estrous cycle of puberty. Endocrinology 110: 1238-1245. Brann, D. W., C. D. Putnam, and V. B. Mahesh (1991). Validation of the mechanisms proposed for the stimulatory and inhibitory effects of progesterone and gonadotropin secretion in the estrogen- primed rat: A possible role for adrenal steroids. Steroids 56: 103- 111. Shupnik, M. A., M. S. Gordon, and W. W. Chin (1989). Tissue- specific regulation of rat estrogen receptor mRNAs. Mol. Endo- crinol. 3: 660-665. Kassis, J. A., and J. Gorski (1983). On the mechanism of estrogen receptor replenishment: recycling, resynthesis and/or processing. Mol. Cell. Biochem. 52: 27-36. Horwitz, K. B., and W. L. McGuire (1978). Estrogen control of progesterone receptor in human breast cancer: Correlation with nuclear process of estrogen receptor. J. Biol. Chem. 253: 2223- 2228.
Horwitz, K. B., and W. L. McGuire (1986). Nuclear mechanisms of estrogen action: Effects of estradiol and antiestrogens on estrogen receptors and nuclear processing. J. Biol. Chem. 253: 8185-8195.
Auricchio, F., A. Migliaccio, and A. Rotondi (1981). Inactivations of estrogen receptor in vitro by nuclear dephosphorylation. Biochem. J. 194:569-574. Auricchio, F., A. Migliaccio, and G. Castoria (1981). Dephosphor- ylation of estrogen receptor in uitro. Biochem. J. 198: 699-702.
Auricchio, F., A. Migliaccio, G. Castoria, S. Lastoria, and A. Ron- tondi (1982). Evidence that in uiuo estradiol receptor translocated into nuclei is dephosphorylated and released into cytoplasm. Biochem. Biophys. Res. Commun. 106: 149-157.
MacLusky, N., and B. McEwen (1980). Progestin receptors in rat brain: Distribution and properties of cytoplasmic progestin-binding sites. Endocrinology 106: 192-197.
Thornton, J. E., B. Neck, B. S. McEwen, and H. H. Feder (1986). Estrogen induction of progestin receptors in microdissected hy- pothalamic and limbic nuclei of female guinea pigs. Neuroendo- crinology 43: 182-188.
Karavolas, H. J., D. Hodges, and D. O’Brien (1976). Uptake of [3H]progesterone and [aH]So-dihydroprogesterone in viuo and analysis of accumulative radioactivity: Accumulation of 5a-dihy- droprogesterone by pituitary and hypothalamic tissues. Endocri- nology 98: 164-175.
Singh, P., and T. G. Muldoon (1983). Specific estrogen-sensitive alterations in anterior pituitary cytoplasmic and nuclear estrogen receptors actuated by LHRH. Neuroendocrinology 37: 98-105.
Weisenberg, L. S., A. F. DeNicola, M. C. Arakelian, and C. Libertun (1979). Effect of median eminence lesions on [sHIestradio binding in the anterior pituitary and hypothalamus. Endocrinology 105: 1152-1157.





























