Embed Size (px)
Citation preview

Anim Cogn (2009) 12:97–105
DOI 10.1007/s10071-008-0174-zORIGINAL PAPER
Quantity judgments of sequentially presented food items by capuchin monkeys (Cebus apella)
Theodore A. Evans · Michael J. Beran · Emily H. Harris · Daniel F. Rice
Received: 7 April 2008 / Revised: 11 June 2008 / Accepted: 19 July 2008 / Published online: 1 August 2008© Springer-Verlag 2008
Abstract Recent assessments have shown that capuchinmonkeys, like chimpanzees and other Old World primatespecies, are sensitive to quantitative diVerences betweensets of visible stimuli. In the present study, we examinedcapuchins’ performance in a more sophisticated quantityjudgment task that required the ability to form representa-tions of food quantities while viewing the quantities onlyone piece at a time. In three experiments, we presentedmonkeys with the choice between two sets of discretehomogeneous food items and allowed the monkeys to con-sume the set of their choice. In Experiments 1 and 2, mon-keys compared an entirely visible food set to a second set,presented item-by-item into an opaque container. All mon-keys exhibited high accuracy in choosing the larger set,even when the entirely visible set was presented last, pre-venting the use of one-to-one item correspondence to com-pare quantities. In Experiment 3, monkeys compared twosets that were each presented item-by-item into opaquecontainers, but at diVerent rates to control for temporalcues. Some monkeys performed well in this experiment,though others exhibited near-chance performance, suggest-ing that this species’ ability to form representations of foodquantities may be limited compared to previously testedspecies such as chimpanzees. Overall, these Wndings sup-port the analog magnitude model of quantity representationas an explanation for capuchin monkeys’ quantiWcation ofsequentially presented food items.
Keywords Capuchin monkey · Cebus · Quantity judgment · Sequential presentation · Analog magnitude estimation
Introduction
Although many animal species have demonstrated basicnumerical abilities (see Brannon and Roitman 2003), thechimpanzee (Pan troglodytes) and rhesus macaque(Macaca mulatta) have dominated the comparative psy-chology literature as nonhuman primate models of numeri-cal cognition. More recently another nonhuman primatespecies, the capuchin monkey (Cebus apella), has provenuseful as a model of numerical cognition. This species is aNew World primate and is valuable to comparative studiesbecause it shares an older common ancestor with humansthan do Old World primates such as rhesus macaques andchimpanzees. To date, researchers have shown that capu-chins exhibit a similar degree of competency to rhesusmacaques and chimpanzees with regard to ordinal judg-ments of computerized item arrays (Brannon and Terrace2000; Judge et al. 2005), ordinal judgments of Arabicnumerals and tokens (Addessi et al. 2007; Beran et al.2008a; Tomonaga and Matsuzawa 2000; Washburn andRumbaugh 1991), and discrete quantity conservation ofdigital dot arrays (Beran 2007a, 2008; Muncer 1983; Sudaand Call 2005).
Researchers also have investigated this species’ abilityto compare discrete quantities of naturally relevant objectsand arrays. Addessi et al. (2008) demonstrated that capu-chins reliably select the larger of two visible quantities of1–5 food items. Beran et al. (2008b) demonstrated thatcapuchins could judge the relative quantity of non-visiblesets of 1–5 food items shown brieXy one whole set at a
T. A. Evans (&) · M. J. Beran · E. H. Harris · D. F. RiceLanguage Research Center, Georgia State University, Atlanta, USAe-mail: [email protected]
123

98 Anim Cogn (2009) 12:97–105
time, as well as make accurate judgments between food setsafter extra items were added to one of the sets. Further, vanMarle et al. (2006) showed that capuchins could makeaccurate judgments between sets of 1–4 food items pre-sented one item at a time (as opposed to one whole set at atime). Success in that task suggested that these monkeys,like chimpanzees (Beran 2001; Beran and Beran 2004) andrhesus macaques (Hauser et al. 2000), could form mentalrepresentations of food sets after viewing only individualpieces of each set for a brief duration. However, in the vanMarle et al. (2006) study, all food items were presented at asimilar rate, so set quantity was confounded with presenta-tion duration. Therefore, it was unclear whether capuchinmonkeys were judging the relative quantity of food items orjudging the relative length of presentation time betweencomparison sets.
The test in which food sets are presented one item at atime is an important assessment of quantity discriminationbecause it has implications for our understanding of theevolution of enumerative processes. Sequentially presenteditems evoke an enumerative process, or at least a summa-tive process. However, most tests of food quantity discrimi-nation may not actually be number discriminations becauseitem number often co-varies with overall food amount (i.e.,mass, volume, or surface area). This is because investiga-tors often present sets containing food items that are verysimilar to one another in size and appearance. For example,if asking an animal to discriminate 2 M&M® candies from5 M&M® candies, it is not possible to say whether the ani-mal decided that the larger set of candies was more numer-ous than the smaller set or if the larger set of candies wasmore massive than the smaller set.
While the process of comparing two sets of food itemsmay not require any recognition of the number of items inthose sets, it does require repeated updates of a represen-tational system used for retaining quantity informationlong enough to support a judgment. Therefore, compari-sons between multiple sets of sequentially-presenteditems or comparisons between sets of sequentially-pre-sented items and simultaneously-presented items requirecomplicated interactions between the enumerative/sum-mative process, working memory, and the decision-mak-ing component of the system that must discriminatebetween those representations. Although there are dis-agreements about the mechanisms used to form these rep-resentations (e.g., Beran 2004, 2007b; Hanus and Call2007; Hauser et al. 2000), it is clear that Old World pri-mates, such as great apes and rhesus monkeys can succeedin this task. The data from van Marle et al. (2006) sug-gested that capuchins also succeed with this task, but theconfounding of quantity with duration requires furtherinvestigation as temporal cues have been ruled out as
important features for Old World primates (e.g., Beranand Beran 2004).
Our primary goal was to clarify whether capuchin mon-keys, like rhesus monkeys and chimpanzees, can performthe comparison of nonvisible food sets presented one itemat a time. We particularly were interested in removing tem-poral presentation duration as a possible useful cue in thistask so that we could more accurately compare the enu-merative/summative skills of these monkeys to the skillshown by the Old World primates. Success at this taskwould indicate that capuchins can form mental representa-tions of food quantities while viewing only individual ele-ments of those quantities. In Experiment 1 and Experiment2, we provided the monkeys with an intermediate versionof the test by providing a comparison between one entirelyvisible food set and one set in which items were presentedone at a time. In Experiment 1, we always presented theentirely visible set Wrst, in the interest of providing themonkeys with the best chance of maintaining an accuratequantity representation. We anticipated that the monkeysmight have diYculty in keeping the sequentially-presentedquantity in working memory following the presentation ofthe static set because of the prepotent nature of the entirelyvisible set of food items. However, this presentation orderalso allowed the monkeys two mechanisms to solve thetask. Monkeys could use one-to-one correspondencebetween items of the already visible static set and itemsobserved one at a time in the second set, or they couldform a mental representation of the sequentially presentedset for comparison with the entirely visible set. However,in Experiment 2, only the latter strategy could beemployed, because we always presented the entirely visi-ble set last. We hypothesized that the monkeys would formmental representations of sets presented item-by-item, andthus, would succeed in choosing the larger set of itemsreliably in both the experiments. Importantly, monkeyscould not use any timing cues in these tasks as one ofthe comparison sets was always presented in its entirety,and as such oVered no temporal dimension in relation toquantity.
In the Wnal experiment, we provided the monkeys with achoice between two food sets, both presented in an item-by-item manner. This method was more comparable towhat was used in earlier studies with chimpanzees and rhe-sus monkeys. It was also similar to the method used by vanMarle et al. (2006) to test capuchins, except that weincluded a control for temporal cuing. We hypothesizedthat the capuchins would persist in selecting food quantitiescontaining the greater amount/number of items in thisexperiment, despite the temporal control, and thereforeexhibit similar food quantity representation capacity tochimpanzees and rhesus monkeys.
123

Anim Cogn (2009) 12:97–105 99
Experiment 1
Methods
Participants
We observed six capuchin monkeys: Liam (male, 3 yearsold), Wren (female, 4 years old), Nala (female, 4 years old),Gabe (male, 9 years old), GriYn (male, 9 years old), andLily (female, 10 years old). All monkeys participated in oneor more previous studies involving quantity judgments. Intwo of these studies, they ordered completely visible digitalobject arrays (Beran 2008) or completely visible digitalnumeral arrays (Beran et al. 2008a) while working at auto-mated computer test stations. In a third study, these mon-keys chose between two sets of food items that were eithervisible at choice time or were entirely visible to monkeys atsome time during the trial (Beran et al. 2008b). Thus, priorto the present study, these monkeys had never been evalu-ated for the ability to quantify sets of food items visibleonly one piece at a time, a task which requires greater enu-merative and working memory capacities than the previ-ously tested judgments.
Design and procedure
We separated participants into individual enclosures fortesting. Monkeys observed the presentation of food setsthrough a Lexan® (clear plastic) panel that was attached tothe side of the enclosure facing the experimenters. Thepanel had two holes large enough for the monkeys to reachthrough, in order to respond to stimuli. These holes forcedthe monkeys to choose only one of the two sets of fooditems, and the monkeys all had previous experience learn-ing that they could only reach through one hole at a time, ora response would not be rewarded (Beran et al. 2008b).
Two experimenters participated in test sessions. At thebeginning of a trial, Experimenter 1 (the Wrst author) heldtwo containers in front of the monkey. One container was ashallow lidless bowl; the other was a deeper opaque boxwith a lid. Experimenter 1 closed his eyes while Experi-menter 2 (the second, third or fourth author) placed one setof Cheerios® cereal pieces in the shallow bowl all at onetime. Experimenter 2 then dropped another set of cerealpieces into the opaque box, one piece at a time (at a rate ofapproximately 1 piece per second), and then closed the lid.The side and bottom surfaces of the opaque box were linedwith thick and soft fabric that prevented the cereal piecesfrom producing sound or vibration when they contactedthese surfaces. Thus, neither the monkey nor Experimenter1 could detect any auditory or tactile cues regarding thequantity of items being dropped into the box. If the monkeyturned away from the experimenter during the presentation
of the second set, we aborted the trial and returned to it atthe end of the session. In all other trials, Experimenter 1reopened his eyes and moved the containers close enoughto the Lexan® panel for the monkey to choose while Exper-imenter 2 looked away. Thus, we eliminated any possibilityfor cuing of the monkeys, as only Experimenter 2 knew thecorrect choice and only Experimenter 1 directly interactedwith the monkeys. The monkeys directly retrieved the fooditems from their choice container while the experimentersimultaneously pulled the other container out of reach. Insummary, the monkeys could see the Wrst set in its entiretyfor the duration of the trial, but they could only see the sec-ond set one piece at a time. Monkeys completed six 10-trialsessions over the course of 3 weeks. We presented everycombination of 1–5 food items, six times throughout theexperiment in a random order. We also randomized the left/right positioning of quantities across trials. Each quantitywas presented all-at-once and item-by-item an equal num-ber of times.
Results
All monkeys chose the larger quantity over the smallerquantity (hereafter, “performed”) signiWcantly better thanexpected by chance, as assessed with two-tailed sign tests(Table 1). An exploratory analysis of the data suggestedthat the monkeys performed better when either the mini-mum or maximum quantity for the experiment (i.e., 1 or 5)was visible during a trial. We conWrmed this pattern using aWilcoxon Matched-Pairs Signed-Ranks test to compareperformance when one of these anchor quantities was visi-ble and when neither quantity was visible, Z = 2.201, N = 6,P = 0.028. Despite this pattern, most monkeys still per-formed signiWcantly above chance when no anchor quantitywas visible (Table 2).
We combined all trial data across monkeys to examineperformance as a function of the ratio between the sets. Theratio between the sets was calculated by dividing thesmaller set size by the larger set size, and this value took
Table 1 Percent selection of the larger quantity by each monkey ineach experiment
P values indicate the results of two-tailed sign tests conducted onthe number of trials in which the larger and smaller quantities wereselected
Experiment Gabe GriYn Liam Lily Nala Wren
1 75.000 83.333 81.667 71.667 73.333 78.333
P <0.001 <0.001 <0.001 0.001 <0.001 <0.001
2 85.000 80.000 85.000 81.667 85.000 85.000
P <0.001 <0.001 <0.001 <0.001 <0.001 <0.001
3 76.667 61.667 50.000 51.667 70.000 75.000
P <0.001 0.093 1.000 0.897 0.003 <0.001
123
100 Anim Cogn (2009) 12:97–105
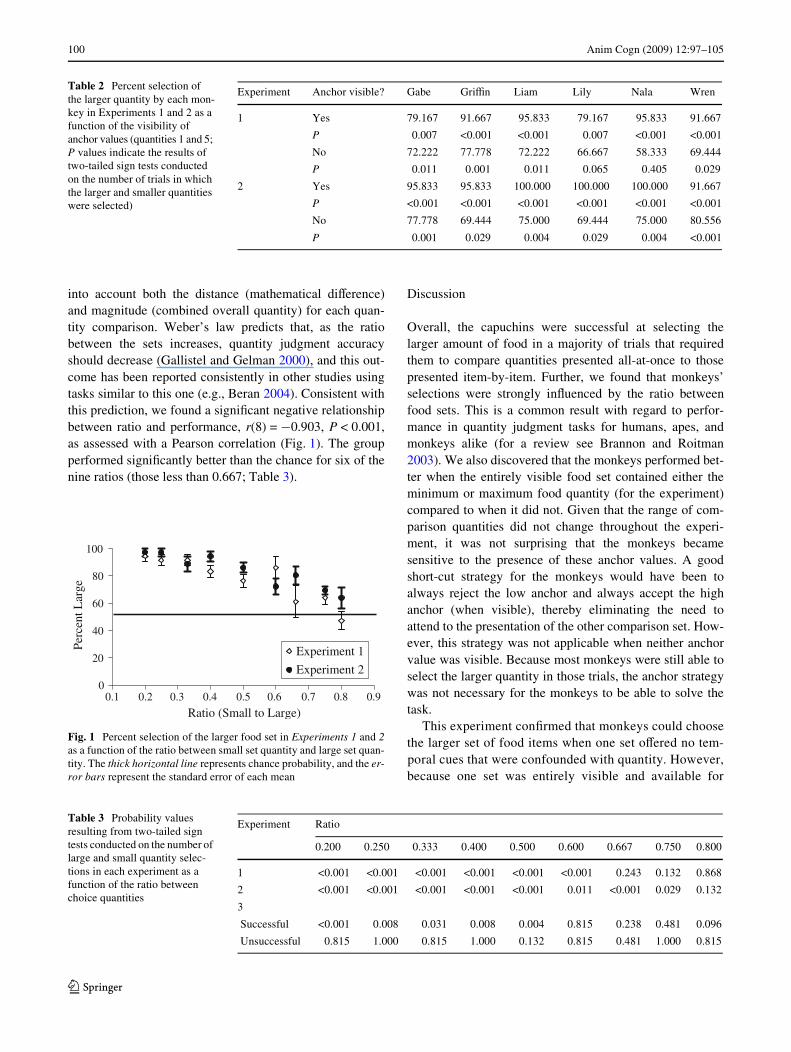
into account both the distance (mathematical diVerence)and magnitude (combined overall quantity) for each quan-tity comparison. Weber’s law predicts that, as the ratiobetween the sets increases, quantity judgment accuracyshould decrease (Gallistel and Gelman 2000), and this out-come has been reported consistently in other studies usingtasks similar to this one (e.g., Beran 2004). Consistent withthis prediction, we found a signiWcant negative relationshipbetween ratio and performance, r(8) = ¡0.903, P < 0.001,as assessed with a Pearson correlation (Fig. 1). The groupperformed signiWcantly better than the chance for six of thenine ratios (those less than 0.667; Table 3).
Discussion
Overall, the capuchins were successful at selecting thelarger amount of food in a majority of trials that requiredthem to compare quantities presented all-at-once to thosepresented item-by-item. Further, we found that monkeys’selections were strongly inXuenced by the ratio betweenfood sets. This is a common result with regard to perfor-mance in quantity judgment tasks for humans, apes, andmonkeys alike (for a review see Brannon and Roitman2003). We also discovered that the monkeys performed bet-ter when the entirely visible food set contained either theminimum or maximum food quantity (for the experiment)compared to when it did not. Given that the range of com-parison quantities did not change throughout the experi-ment, it was not surprising that the monkeys becamesensitive to the presence of these anchor values. A goodshort-cut strategy for the monkeys would have been toalways reject the low anchor and always accept the highanchor (when visible), thereby eliminating the need toattend to the presentation of the other comparison set. How-ever, this strategy was not applicable when neither anchorvalue was visible. Because most monkeys were still able toselect the larger quantity in those trials, the anchor strategywas not necessary for the monkeys to be able to solve thetask.
This experiment conWrmed that monkeys could choosethe larger set of food items when one set oVered no tem-poral cues that were confounded with quantity. However,because one set was entirely visible and available for
Table 2 Percent selection of the larger quantity by each mon-key in Experiments 1 and 2 as a function of the visibility of anchor values (quantities 1 and 5; P values indicate the results of two-tailed sign tests conducted on the number of trials in which the larger and smaller quantities were selected)
Experiment Anchor visible? Gabe GriYn Liam Lily Nala Wren
1 Yes 79.167 91.667 95.833 79.167 95.833 91.667
P 0.007 <0.001 <0.001 0.007 <0.001 <0.001
No 72.222 77.778 72.222 66.667 58.333 69.444
P 0.011 0.001 0.011 0.065 0.405 0.029
2 Yes 95.833 95.833 100.000 100.000 100.000 91.667
P <0.001 <0.001 <0.001 <0.001 <0.001 <0.001
No 77.778 69.444 75.000 69.444 75.000 80.556
P 0.001 0.029 0.004 0.029 0.004 <0.001
Fig. 1 Percent selection of the larger food set in Experiments 1 and 2as a function of the ratio between small set quantity and large set quan-tity. The thick horizontal line represents chance probability, and the er-ror bars represent the standard error of each mean
0
20
40
60
80
100
0.1
Ratio (Small to Large)
Perc
ent L
arge
Experiment 1
Experiment 2
0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9
Table 3 Probability values resulting from two-tailed sign tests conducted on the number of large and small quantity selec-tions in each experiment as a function of the ratio between choice quantities
Experiment Ratio
0.200 0.250 0.333 0.400 0.500 0.600 0.667 0.750 0.800
1 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 0.243 0.132 0.868
2 <0.001 <0.001 <0.001 <0.001 <0.001 0.011 <0.001 0.029 0.132
3
Successful <0.001 0.008 0.031 0.008 0.004 0.815 0.238 0.481 0.096
Unsuccessful 0.815 1.000 0.815 1.000 0.132 0.815 0.481 1.000 0.815
123

Anim Cogn (2009) 12:97–105 101
comparison throughout each trial, it was possible thatmonkeys used one-to-one correspondence of itemsbetween sets as a means for making judgments of relativeamount. In other words, they may have viewed eachsequentially presented item and compared that item on aone-to-one basis to a visible item in the other set, andthen responded based on the outcome of that procedure.This would allow them to succeed without performingany actual summation or enumeration of the sequentialset at all. Therefore, in Experiment 2, we presented thesequential set Wrst and the visible set second so that nosuch correspondence could be used. This change allowedus to assess whether the monkeys could truly form mentalrepresentations of food quantities in order to solve thistask.
Experiment 2
Methods
Participants
We tested the same individuals that participated in Experi-ment 1.
Design and procedure
We used the same procedure that we used in Experiment 1,except that we presented the two sets in opposite order (theitem-by-item set and then the entirely visible set). All otherdetails, such as the number of trials and the set sizes wereidentical to Experiment 1.
Results
All six animals performed signiWcantly above chance asassessed with two-tailed sign tests (Table 1). As a group,capuchins performance in Experiment 2 was none diVer-ent from that in Experiment 1, Z = 1.897, N = 6,P = 0.058, as assessed with a Wilcoxon Matched-PairsSigned-Ranks test. Also, as in Experiment 1, the monkeysperformed signiWcantly better when an anchor was visiblecompared to when no anchor was visible, Z = 2.207,N = 6, P = 0.027. However, all monkeys performed sig-niWcantly better than chance even when no anchor wasvisible (Table 2). Combining all trial data across mon-keys, we found a signiWcant negative relationshipbetween ratio and Experiment 2 performance using aPearson correlation, r(8) = ¡0.942, P < 0.001 (Fig. 1), aswas seen in Experiment 1. The group performed signiW-cantly better than chance for eight out of nine ratios (thoseless than 0.800; Table 3).
Discussion
In Experiments 1 and 2, the monkeys could see one food setin its entirety and could compare this set to the second set,either during or following item-by-item presentation,respectively. The fact that the monkeys performed equallywell in these two experiments indicated that they did nothave to rely on one-to-one correspondence of comparisonitems, but instead could form mental representations ofcomparison sets to solve the task. They also were not com-paring the temporal aspects of set presentation, and theywere capable of enumerating or summing one set presentedacross time with another set presented in its entirety. Giventhe successful performance of the capuchins in comparingstatic sets of food items to sets of food items presented onepiece at a time, the next step was to test their ability to com-pare two sequentially presented sets. Critically, we ensuredthat temporal cues were not available in the new test as wefurther investigated the quantitative skills of these mon-keys.
Experiment 3
Methods
Participants
We tested the same individuals that participated in Experi-ments 1 and 2.
Design and procedure
We used the same general procedure that we used in Exper-iments 1 and 2. However, there were two key diVerences inthe Experiment 3 method. First, we presented both quanti-ties into opaque boxes with lids and thick fabric lining thatprevented auditory feedback (instead of into one shallowbowl and one box). Also, Experimenter 2 dropped each setof food items piece-by-piece into one of the two containers(instead of presenting the Wrst set all-at-once). As in theprevious experiments, Experimenter 2 still presented allitems of one set before presenting any items of the secondset. Also, as in the previous experiments, the trial wasstopped if the monkey turned away from the experimentersduring item-by-item presentation of food sets.
To control for temporal cues, Experimenter 2 alwaystook more time to drop items into the Wrst container com-pared to the second container, regardless of which set con-tained more items. The experimenter did this by holding hisor her hand over the Wrst container for a longer period oftime in comparison to the second container, as well as byvarying the item drop rate between containers as necessary.
123
102 Anim Cogn (2009) 12:97–105
For example, to present a comparison of 1 item versus 5items, in that order, the experimenter would hold his or herhand over the Wrst container for a total of approximately5 s, dropping the single item after approximately 2 s, andthen would drop the set of 5 items one after another,approximately every 500 ms, into the second container.Combined with the fact that left and right positioning ofquantities was randomized across trials, this meant thatthere could be no consistent relationship between set quan-tity and presentation duration (i.e., sometimes the largerquantity took longer to present, and sometimes the smallerquantity took longer to present).
Results
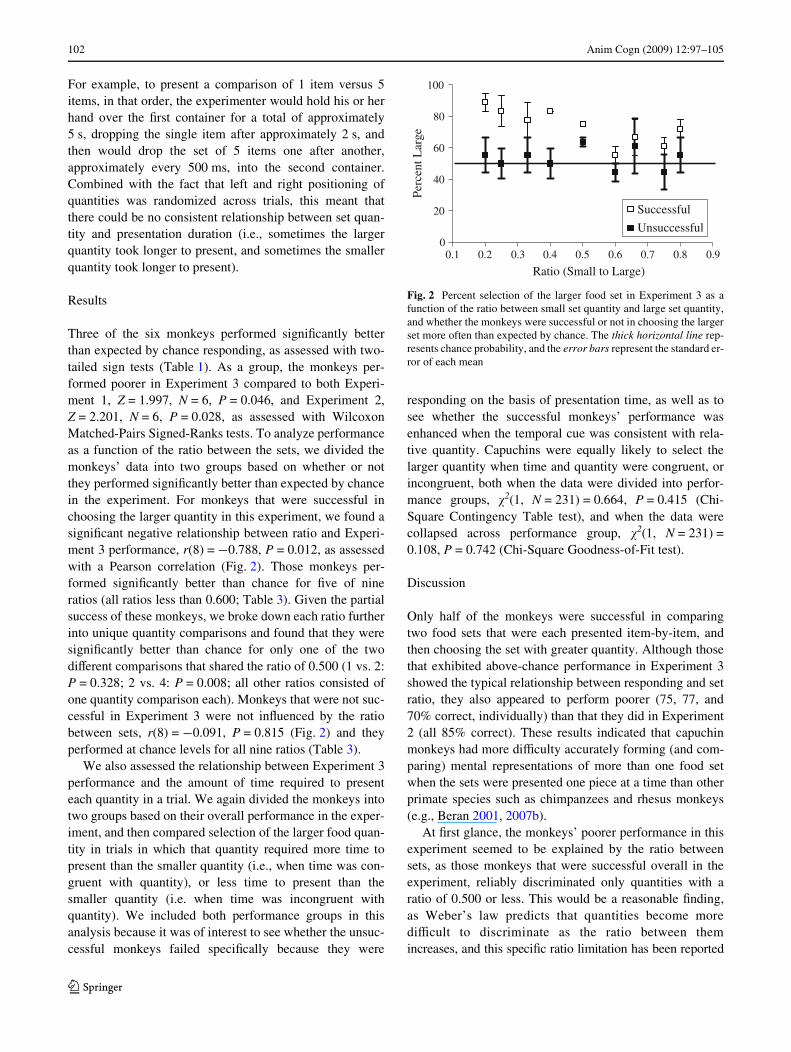
Three of the six monkeys performed signiWcantly betterthan expected by chance responding, as assessed with two-tailed sign tests (Table 1). As a group, the monkeys per-formed poorer in Experiment 3 compared to both Experi-ment 1, Z = 1.997, N = 6, P = 0.046, and Experiment 2,Z = 2.201, N = 6, P = 0.028, as assessed with WilcoxonMatched-Pairs Signed-Ranks tests. To analyze performanceas a function of the ratio between the sets, we divided themonkeys’ data into two groups based on whether or notthey performed signiWcantly better than expected by chancein the experiment. For monkeys that were successful inchoosing the larger quantity in this experiment, we found asigniWcant negative relationship between ratio and Experi-ment 3 performance, r(8) = ¡0.788, P = 0.012, as assessedwith a Pearson correlation (Fig. 2). Those monkeys per-formed signiWcantly better than chance for Wve of nineratios (all ratios less than 0.600; Table 3). Given the partialsuccess of these monkeys, we broke down each ratio furtherinto unique quantity comparisons and found that they weresigniWcantly better than chance for only one of the twodiVerent comparisons that shared the ratio of 0.500 (1 vs. 2:P = 0.328; 2 vs. 4: P = 0.008; all other ratios consisted ofone quantity comparison each). Monkeys that were not suc-cessful in Experiment 3 were not inXuenced by the ratiobetween sets, r(8) = ¡0.091, P = 0.815 (Fig. 2) and theyperformed at chance levels for all nine ratios (Table 3).
We also assessed the relationship between Experiment 3performance and the amount of time required to presenteach quantity in a trial. We again divided the monkeys intotwo groups based on their overall performance in the exper-iment, and then compared selection of the larger food quan-tity in trials in which that quantity required more time topresent than the smaller quantity (i.e., when time was con-gruent with quantity), or less time to present than thesmaller quantity (i.e. when time was incongruent withquantity). We included both performance groups in thisanalysis because it was of interest to see whether the unsuc-cessful monkeys failed speciWcally because they were
responding on the basis of presentation time, as well as tosee whether the successful monkeys’ performance wasenhanced when the temporal cue was consistent with rela-tive quantity. Capuchins were equally likely to select thelarger quantity when time and quantity were congruent, orincongruent, both when the data were divided into perfor-mance groups, �2(1, N = 231) = 0.664, P = 0.415 (Chi-Square Contingency Table test), and when the data werecollapsed across performance group, �2(1, N = 231) =0.108, P = 0.742 (Chi-Square Goodness-of-Fit test).
Discussion
Only half of the monkeys were successful in comparingtwo food sets that were each presented item-by-item, andthen choosing the set with greater quantity. Although thosethat exhibited above-chance performance in Experiment 3showed the typical relationship between responding and setratio, they also appeared to perform poorer (75, 77, and70% correct, individually) than that they did in Experiment2 (all 85% correct). These results indicated that capuchinmonkeys had more diYculty accurately forming (and com-paring) mental representations of more than one food setwhen the sets were presented one piece at a time than otherprimate species such as chimpanzees and rhesus monkeys(e.g., Beran 2001, 2007b).
At Wrst glance, the monkeys’ poorer performance in thisexperiment seemed to be explained by the ratio betweensets, as those monkeys that were successful overall in theexperiment, reliably discriminated only quantities with aratio of 0.500 or less. This would be a reasonable Wnding,as Weber’s law predicts that quantities become morediYcult to discriminate as the ratio between themincreases, and this speciWc ratio limitation has been reported
Fig. 2 Percent selection of the larger food set in Experiment 3 as afunction of the ratio between small set quantity and large set quantity,and whether the monkeys were successful or not in choosing the largerset more often than expected by chance. The thick horizontal line rep-resents chance probability, and the error bars represent the standard er-ror of each mean
0
20
40
60
80
100
0.1
Ratio (Small to Large)
Perc
ent L
arge
Successful
Unsuccessful
0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9
123

Anim Cogn (2009) 12:97–105 103
elsewhere with regard to quantitative decision making innonhuman primates (e.g., Flombaum et al. 2005). However,this explanation is not perfect, because these monkeyssuccessfully discriminated only one of the two quantitypairings characterized by a ratio of 0.500 (2 vs. 4, but not 1vs. 2), whereas Weber’s law states that those two compari-sons should possess an equal level of diYculty with regardto perceptual discrimination.
Another possible explanation for this performance pat-tern was that the successful monkeys were able to reliablychoose the larger quantity only when it was compared to analternative of one item. Of the Wve ratios for which thesemonkeys performed signiWcantly better than expected bychance (0.200 to 0.500), four involved the comparison ofone item to some other quantity (all ratios but 0.400, thecomparison of 2 vs. 5 items).1 However, the fact that thesemonkeys performed signiWcantly above chance when com-paring 2 versus 4 items, but not when comparing 1 versus 2items, contradicts this explanation. This assessment of themonkeys’ performance with regard to individual quantitycomparisons, as opposed to ratio (1 versus 2 and 2 versus 4share the ratio of 0.500), indicates that the monkeys reliablydiscriminated a similar number of comparisons that did anddid not involve the quantity 1 (three and two comparisons,respectively).
All things considered, it seems that the best explanationfor this performance pattern is that monkeys’ respondingwas inXuenced strongly by set ratio, but with regard to theratio 0.500, monkeys’ were inXuenced more by the distancecomponent of Weber’s law than by the magnitude compo-nent. In other words, monkeys treated the quantities 1 and 2as approximate equals due to their closeness in value,whereas monkeys discriminated between the quantities 2and 4 successfully because they were less similar in value.This was true despite the greater overall magnitude associ-ated with the latter set of quantities.
Aside from the diVerence in overall performance betweenthis and previous experiments, we also found that presenta-tion time was neither helpful nor detrimental to monkeys’performance. This Wnding was true for monkeys that weresuccessful overall and those that were unsuccessful overall atchoosing the larger set of food items across trials. Therefore,monkeys were most likely responding on the basis of relativequantity in this experiment as opposed to temporal cues.
General discussion
The primary objective of this study was to test capuchinmonkeys’ capacity to represent food quantities presented
one piece at a time, while controlling for the potentialresponse cue of set presentation duration. In Experiment 1,monkeys succeeded in quantifying one such food quantityand comparing it to another food quantity that presented allitems at once at the beginning of the trial. This method con-trolled for the temporal cue by making it nonexistent forone of the sets (presentation duration may have correlatedwith quantity in the sequentially presented set, but it couldnot for the static set). However, Experiment 1 methodpotentially allowed monkeys to use one-to-one correspon-dence to compare the two sets. Experiment 2 prevented thistype of strategy by withholding the entirely visible set untilthe end of the trial, thereby forcing monkeys to encode andremember the amount of the sequentially presented set inorder to make an accurate judgment. Monkeys continued tochoose the larger food quantities in Experiment 2, and thus,we can conclude that the monkeys were representing oneset of individually presented food items, and using that rep-resentation to make accurate quantity judgments. This Wnd-ing indicates that, with regard to a single set of 1–5 items,capuchins are on par with rhesus macaques’ and chimpan-zees’ representation of nonvisible food quantities (seeBeran 2001, 2004; Beran and Beran 2004; Hauser et al.2000). It remains to be seen whether that performance willcontinue for larger quantities and whether performance willcontinue to obey Weber’s law.
In Experiment 3, however, the monkeys exhibited sig-niWcantly poorer judgments between quantities that wereeach presented one piece at a time. Half of the subjects didnot select the larger quantity more often than expected bychance, and the other half, though successful overall, per-formed poorer than that they did with respect to just onesequentially-presented set. Performance in Experiment 3was most likely poorer than in the earlier experimentsbecause discriminations between quantities of the highestratios, which were expected to be the most diYcult (accord-ing to Weber’s law), were made even more diYcult by theadded requirement of forming and maintaining representa-tions of both quantities (as opposed to only one quantity inthe earlier experiments). This explanation is supported bythe fact that, in Experiment 3, monkeys were only success-ful in selecting the larger quantity when comparisons werecharacterized by relatively low ratios (Fig. 2; Table 3),whereas monkeys successfully discriminated quantity com-parisons involving low and high ratios in Experiment 1 andExperiment 2 (Fig. 1; Table 3). This diVerence in the twotask types (where one involves a static, visible set, whereasthe other oVers no such perceptual constant during the judg-ment response) reXects a diVerence in working memoryload. The resulting change in performance indicates thatcapuchin quantity estimation and comparison is dependenton working memory capacity that appears to be somewhatlimited in comparison to other primate species tested in this
1 This alternative explanation was oVered by a referee during the re-view process, and we are grateful to him or her for the suggestion.
123

104 Anim Cogn (2009) 12:97–105
paradigm. This needs to be investigated more fully, both inother quantitative paradigms, and more broadly in othertasks that require working memory, to assess this possibility.
It seems that capuchins may be less able to represent anddiscriminate multiple sets of individually presented itemsthan other primate species more closely related to humans(e.g., Beran and Beran 2004; Hauser et al. 2000). However,not all apes succeed with this type of task either (Hanus andCall 2007), so there is a reason to presume continuityacross New World and Old World monkeys in these kindsof skills. Our data partially support the results of van Marleet al. (2006) who reported that capuchins made accuratejudgments of this type in a majority of sessions in whichtwo sets of food items were presented piece-by-piece. Someof our monkeys also showed this result, although somefailed despite successfully performing the earlier tasks. Thecritical diVerence between van Marle et al. (2006) methodand our own is that we included a control for the temporalcue, whereas van Marle et al. admittedly allowed quantityand presentation duration to co-vary. Although our high-performing monkeys showed no diVerence in performancebetween temporal conditions, it is possible that, if exposedto a proWtable temporal cue for a longer time (i.e., through-out an entire experiment), they may have come to favor thetemporal cue over the discrete quantity cue. However,because presentation duration was never a proWtable cue inthis or previous experiments for these monkeys, the capu-chins probably learned to ignore it.
Aside from the temporal cue, it still remains to be deter-mined what stimulus feature(s) these and other animals useto make quantity judgments in these kinds of tasks. In thesetasks, item number and overall set amount (i.e., mass, vol-ume, or area) usually co-vary because comparison sets con-sist of nearly identical individual food items. This makes itimpossible to determine whether a numeric or nonnumericquantity judgment is at the root of the animal’s perfor-mance. When faced with this interpretational problem, it isintuitive to suggest that overall food amount is the salientcue, given that sensitivity to food quantity would mostlikely evolve as a result of the pressure to maximize nutri-tional or caloric intake of food sources. This idea has ledsome to suggest that, in these tests and analogous naturalsituations, number is important only as a last-resort strategyto discriminate quantities when more salient nonnumericcues are unavailable (Davis and Memmott 1982). There aresome empirical Wndings to support the application of this“last-resort” hypothesis to the tasks used in the presentstudy. For example, chimpanzees have exhibited the ten-dency to discriminate food sets, presented in either static orsequential fashion, using amount cues such as surface areaand individual item size instead of item number (Beranet al. 2008c). However, this may not be the case whenquantities consist of nonfood items, in which case number
appears to play a more central role in performance (e.g.,Cantlon and Brannon 2007).
It perhaps is parsimonious to favor amount as the domi-nant cue in these kinds of tasks because amount judgmentsare less sophisticated than a truly numerical judgment.However, a recent model of nonverbal quantity representa-tion suggests that all quantity judgments, discrete and con-tinuous, countable and uncountable, can be made using thesame representational system (Gallistel and Gelman 2000).In this model, a construct called the accumulator is incre-mented or updated as each unit of the total quantity (e.g.,individual food item or splash of liquid) is gauged. Whenthe whole quantity has been evaluated, the accumulatorempties its contents into memory and then the next quantitycan be incremented and later compared to the quantity inmemory. Thus, this model suggests that tallying and com-paring two nonnumeric quantities is no less sophisticatedthan tallying and comparing two numeric quantities. Con-sider the work of van Marle et al. (2006) as a potentialapplication of this model to empirical Wndings. Theseresearchers showed that capuchin monkeys discriminatedamounts of solid objects (yogurt raisins) and nonsolid sub-stances (banana puree) in a similar manner with respect tothe ratio between quantities. Thus, one could argue that themonkeys’ mental accumulator was being incremented withapproximate amounts of either raisins or puree, in a similarfashion, to form a representation of each type of quantity.
Given the results of the present investigation, capuchinmonkeys will continue to be valuable to the comparativestudy of numerical cognition and quantity judgments, asthey seem to occupy a middle ground along the continuumof such capacities. While our data and those from vanMarle et al. (2006) jointly suggest that capuchin monkeyscan quantify and compare sequentially presented sets ofstimuli, there are some animal species that seem to showgreater representational capacity with regard to this andother tasks (e.g., Beran 2001, 2004). One question forfuture research that may help place capuchin monkeysalong this continuum of numerical and quantitative compe-tence is whether they can quantify and compare sets of fooditems beyond the range of 1–5, as do some Old World pri-mate species (e.g., Beran 2001; Boysen et al. 1995; Flom-baum et al. 2005). While capuchin monkeys have exhibitedthe ability to order completely visible arrays of items repre-senting quantities beyond this range (Judge et al. 2005), itis unknown whether these monkeys can form representa-tions of larger quantities (e.g., 5–10) and compare them toone another from memory. The results of Experiment 3 ofthe present study suggest that they may be able to do so, aslong as the ratio between these larger comparison quantitiesis relatively small, but nonetheless, more direct evidencewill be required to address this question. More generally,characterizing the underlying processes that are associated
123

Anim Cogn (2009) 12:97–105 105
with diVerent levels of quantiWcation in these animals willbe important for furthering our understanding of the evolu-tionary and physiological underpinnings of numerical cog-nition and quantitative decision making.
Acknowledgments This research was supported by National Insti-tutes of Health Grant HD-38051 to the Language Research Center ofGeorgia State University. We thank John Kelley, Sarah Hunsbergerand Betty Chan for assistance with animal care and management.These experiments complied with the current laws of the United Statesof America.
References
Addessi E, Crescimbene L, Visalberghi E (2007) Do capuchin mon-keys (Cebus apella) use tokens as symbols? Proc R Soc Lond B274:2579–2585
Addessi E, Crescimbene L, Visalberghi E (2008) Food and token quan-tity discrimination in capuchin monkeys (Cebus apella). AnimCogn 11:275–282
Beran MJ (2001) Summation and numerousness judgments of sequen-tially presented sets of items by chimpanzees (Pan troglodytes). JComp Psychol 115:181–191
Beran MJ (2004) Chimpanzees (Pan troglodytes) respond to nonvisi-ble sets after one-by-one addition and removal of food items. JComp Psychol 118:25–36
Beran MJ (2007a) Rhesus monkeys (Macaca mulatta) succeed on acomputerized test designed to assess conservation of discretequantity. Anim Cogn 10:37–45
Beran MJ (2007b) Rhesus monkeys (Macaca mulatta) enumerate largeand small sequentially presented sets of items using analog numer-ical representations. J Exp Psychol Anim Behav Process 33:42–54
Beran MJ (2008) Capuchin monkeys (Cebus apella) succeed in a testof quantity conservation. Anim Cogn 11:109–116
Beran MJ, Beran MM (2004) Chimpanzees remember the results ofone-by-one addition of food items to sets. Psychol Sci 15:94–99
Beran MJ, Harris EH, Evans TA, Klein ED, Chan B, Flemming TM,Washburn DA (2008a) Ordinal judgments of symbolic stimuli bycapuchin monkeys (Cebus apella) and rhesus monkeys (Macacamulatta): the eVects of diVerential and nondiVerential reward. JComp Psychol 122:52–61
Beran MJ, Evans TA, Leighty KA, Harris EH, Rice D (2008b) Sum-mation and quantity judgments of sequentially presented sets bycapuchin monkeys (Cebus apella). Am J Primatol 70:191–194
Beran MJ, Evans TA, Harris EH (2008c) Perception of food amountsby chimpanzees based on the number, size, contour length andvisibility of items. Anim Behav 75:1793–1802
Boysen ST, Bernston GG, Shreyer TA, Hannan MB (1995) Indicatingacts during couting by a chimpanzee (Pan troglodytes). J CompPsychol 109:47–51
Brannon EM, Roitman JD (2003) Nonverbal representations of timeand number in animals and human infants. In: Neck WH (ed)Functional and neural mechanisms of interval timing. CRC Press,Boca Raton, pp 143–182
Brannon EM, Terrace HS (2000) Representation of the numerosities1–9 by rhesus macaques (Macaca mulatta). J Exp Psychol AnimBehav Process 26:31–49
Cantlon JF, Brannon EM (2007) How much does number matter to amonkey (Macaca mulatta)? J Exp Psychol Anim Behav Process33:32–41
Davis H, Memmott J (1982) Counting behavior in animals: a criticalevaluation. Psychol Bull 92:547–571
Flombaum JI, Junge JA, Hauser MD (2005) Rhesus monkeys (Macacamulatta) spontaneously compute addition operations over largernumbers. Cognition 97:315–325
Gallistel CR, Gelman R (2000) Nonverbal numerical cognition: fromreals to integers. Trends Cogn Sci 4:59–65
Hanus D, Call J (2007) Discrete quantity judgments in the great apes(Pan paniscus, Pan troglodytes, Gorilla gorilla, Pongo pygma-eus): the eVect of presenting whole sets versus item-by-item. JExp Psychol Anim Behav Process 121:241–249
Hauser MD, Carey S, Hauser LB (2000) Spontaneous number repre-sentation in semi-free-ranging rhesus monkeys. Proc R Soc LondB 267:829–833
Judge PG, Evans TA, Vyas DK (2005) Ordinal representation of nu-meric quantities by brown capuchin monkeys (Cebus apella). JExp Psychol Anim Behav Process 31:79–94
Muncer SJ (1983) “Conservations” with a chimpanzee. Dev Psychol16:1–11
Suda C, Call J (2005) Piagetian conservation of discrete quantities inbonobos (Pan paniscus), chimpanzees (Pan troglodytes), andorangutans (Pongo pygmaeus). Anim Cogn 8:220–235
Tomonaga M, Matsuzawa T (2000) Sequential responding to Arabicnumerals with wild cards by the chimpanzee (Pan troglodytes).Anim Cogn 3:1–11
van Marle K, Aw J, McCrink K, Santos LR (2006) How capuchin mon-keys (Cebus apella) quantify objects and substances. J Comp Psy-chol 120:416–426
Washburn DA, Rumbaugh DM (1991) Ordinal judgments of numer-ical symbols by macaques (Macaca mulatta). Psychol Sci2:190–193
123





























