Embed Size (px)
Citation preview





168 W.A. Makled, M.R. Langer / Revue de micropaléontologie 53 (2010) 163–173
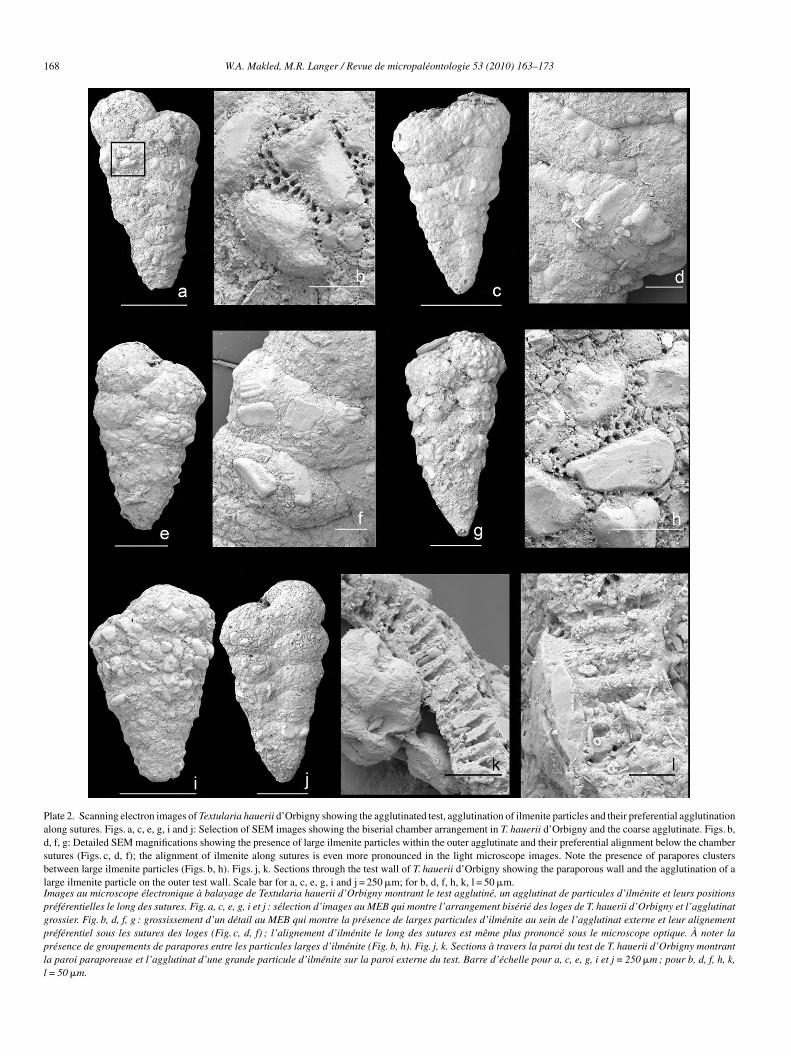
Plate 2. Scanning electron images of Textularia hauerii d’Orbigny showing the agglutinated test, agglutination of ilmenite particles and their preferential agglutinationalong sutures. Figs. a, c, e, g, i and j: Selection of SEM images showing the biserial chamber arrangement in T. hauerii d’Orbigny and the coarse agglutinate. Figs. b,d, f, g: Detailed SEM magnifications showing the presence of large ilmenite particles within the outer agglutinate and their preferential alignment below the chambersutures (Figs. c, d, f); the alignment of ilmenite along sutures is even more pronounced in the light microscope images. Note the presence of parapores clustersbetween large ilmenite particles (Figs. b, h). Figs. j, k. Sections through the test wall of T. hauerii d’Orbigny showing the paraporous wall and the agglutination of alarge ilmenite particle on the outer test wall. Scale bar for a, c, e, g, i and j = 250 �m; for b, d, f, h, k, l = 50 �m.Images au microscope électronique à balayage de Textularia hauerii d’Orbigny montrant le test agglutiné, un agglutinat de particules d’ilménite et leurs positionspréférentielles le long des sutures. Fig. a, c, e, g, i et j : sélection d’images au MEB qui montre l’arrangement bisérié des loges de T. hauerii d’Orbigny et l’agglutinatgrossier. Fig. b, d, f, g : grossissement d’un détail au MEB qui montre la présence de larges particules d’ilménite au sein de l’agglutinat externe et leur alignementpréférentiel sous les sutures des loges (Fig. c, d, f) ; l’alignement d’ilménite le long des sutures est même plus prononcé sous le microscope optique. À noter laprésence de groupements de parapores entre les particules larges d’ilménite (Fig. b, h). Fig. j, k. Sections à travers la paroi du test de T. hauerii d’Orbigny montrantla paroi paraporeuse et l’agglutinat d’une grande particule d’ilménite sur la paroi externe du test. Barre d’échelle pour a, c, e, g, i et j = 250 �m ; pour b, d, f, h, k,l = 50 �m.

W.A. Makled, M.R. Langer / Revue de mic
Fig. 1. Location and sampling sites for Textularia hauerii d’Orbigny, 1846 atBazaruto Island, Mozambique (Indian Ocean).Localisation et sites d’échantillonnages des Textularia hauerii d’Orbigny, 1846sur l’île de Bazaruto, au Mozambique (Océan Indien).
tAAs
Btsdrma
TtonicotTroistt
ststfaPprggabtno
n
Table 1Quantitative distribution of minerals (%) in the size class between 100 and 150 �m asonly represent a minor fraction in this size-range and makes up less than 0.6%.Distribution quantitative des minéraux (%) dans la classe de taille entre 100 et 150 �
724 et 725. À noter que les minéraux d’ilménite représentent une faible fraction dans
Quartz Plagioclase Orthocl
Sample 724 80.31 3.84 0.48Sample 725 77.01 2.91 0.19
ropaléontologie 53 (2010) 163–173 169
est surface of T. hauerii were previously depicted by Heron-llen and Earland (1915) in material collected off the Kerimbarchipelago (Mozambique) but they did not comment on grain
electivity.The source area for the heavy ilmenite (FeTiO3) grains off
azaruto is probably on mainland Mozambique, where some ofhe world’s largest deposits of titanium-bearing heavy mineralands are located (Tyler and Minnitt, 2004). River sedimentsischarged from both the Limpopo and the Save Rivers rep-esent the major pathway for transport from the land to thearine environment of the Bazaruto Archipelago (Armitage et
l., 2006).Besides ilmenite, aragonite is enriched in the tests of
. hauerii (59%) whereas quartz is present only in low concen-rations (7.5% in the tests versus 62.3% in the sediment) andrthoclase grains appear to be completely excluded as aggluti-ated particles (Table 1). The enrichment in aragonite (Table 1)s probably linked to the use of this mineral as a biomineralizedement (Roberts and Murray, 1995). The remaining proportionsf quartz present in the agglutinate of T. hauerii argue againsthe alternate hypothesis that quartz is selectively rejected by. hauerii. The percentage composition of minerals in the sur-ounding sediment is thus not reflected in the agglutinated testsf T. hauerii. In addition, the enhanced concentration of ilmeniten the tests of T. hauerii suggests that there is a preferentialelection for ilmenite. This raises the questions of how selec-ivity is accomplished by a protistan cell in the agglutination ofitanium-containing minerals.
The ilmenite grains incorporated on the test surface range inize between 100 and 150 �m, and are among the larger par-icles in the agglutinate of T. hauerii. Among all grains in theize class between 100 and 150 �m, ilmenite makes up lesshan 0.6% within the sediment. X-ray diffraction analysis oforaminiferal tests, in turn, shows that ilmenite is selectivelygglutinated within the tests of T. hauerii (Table 2; Plate 1 andlate 2), where it makes up more than 26% of the mineral com-onents (Table 2). As the incorporated ilmenite minerals are onlyepresented in minor amounts among the surrounding sedimentrains in their size class (Table 1), the selectivity of ilmeniterains for test construction is not based on grain size but stronglyrgues in favor of specific mineral selection. The absence oflack ilmenite grains in the other agglutinated Foraminifera fromhe same sample sites supports the conclusion, that grain size is
ot the primary driving force that governs the preferential uptakef ilmenite in T. hauerii.To date, however, selectivity of grains according to size can-ot fully be explored: A detailed grain-size analysis of the
counted in sediment samples from site 724 and 725. Note that ilmenite minerals
m telles qu’elles ont été comptées dans les échantillons sédimentaires des sitescette classe de tailles et en constituent moins de 0,6 %.
ase (Microcline) Calcareous Fragments Ilmenite
14.79 0.5819.40 0.48
170 W.A. Makled, M.R. Langer / Revue de micropaléontologie 53 (2010) 163–173
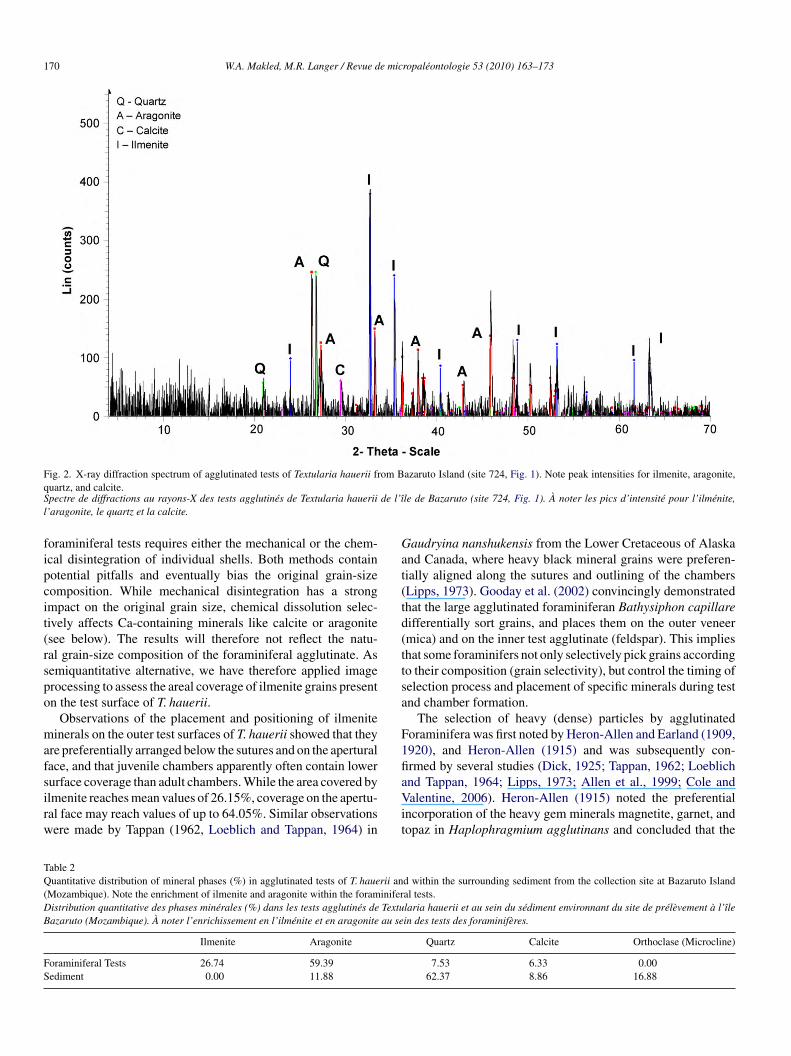
Fig. 2. X-ray diffraction spectrum of agglutinated tests of Textularia hauerii from Bazaruto Island (site 724, Fig. 1). Note peak intensities for ilmenite, aragonite,qS de l’îl
fipcit(rspo
mafsirw
Gat(td(ttsa
F1fi
TQ(DB
FS
uartz, and calcite.pectre de diffractions au rayons-X des tests agglutinés de Textularia hauerii’aragonite, le quartz et la calcite.
oraminiferal tests requires either the mechanical or the chem-cal disintegration of individual shells. Both methods containotential pitfalls and eventually bias the original grain-sizeomposition. While mechanical disintegration has a strongmpact on the original grain size, chemical dissolution selec-ively affects Ca-containing minerals like calcite or aragonitesee below). The results will therefore not reflect the natu-al grain-size composition of the foraminiferal agglutinate. Asemiquantitative alternative, we have therefore applied imagerocessing to assess the areal coverage of ilmenite grains presentn the test surface of T. hauerii.
Observations of the placement and positioning of ilmeniteinerals on the outer test surfaces of T. hauerii showed that they
re preferentially arranged below the sutures and on the aperturalace, and that juvenile chambers apparently often contain lower
urface coverage than adult chambers. While the area covered bylmenite reaches mean values of 26.15%, coverage on the apertu-al face may reach values of up to 64.05%. Similar observationsere made by Tappan (1962, Loeblich and Tappan, 1964) inaVit
able 2uantitative distribution of mineral phases (%) in agglutinated tests of T. hauerii an
Mozambique). Note the enrichment of ilmenite and aragonite within the foraminiferistribution quantitative des phases minérales (%) dans les tests agglutinés de Textuazaruto (Mozambique). À noter l’enrichissement en l’ilménite et en aragonite au se
Ilmenite Aragonite
oraminiferal Tests 26.74 59.39ediment 0.00 11.88
le de Bazaruto (site 724, Fig. 1). À noter les pics d’intensité pour l’ilménite,
audryina nanshukensis from the Lower Cretaceous of Alaskand Canada, where heavy black mineral grains were preferen-ially aligned along the sutures and outlining of the chambersLipps, 1973). Gooday et al. (2002) convincingly demonstratedhat the large agglutinated foraminiferan Bathysiphon capillareifferentially sort grains, and places them on the outer veneermica) and on the inner test agglutinate (feldspar). This implieshat some foraminifers not only selectively pick grains accordingo their composition (grain selectivity), but control the timing ofelection process and placement of specific minerals during testnd chamber formation.
The selection of heavy (dense) particles by agglutinatedoraminifera was first noted by Heron-Allen and Earland (1909,920), and Heron-Allen (1915) and was subsequently con-rmed by several studies (Dick, 1925; Tappan, 1962; Loeblich
nd Tappan, 1964; Lipps, 1973; Allen et al., 1999; Cole andalentine, 2006). Heron-Allen (1915) noted the preferentialncorporation of the heavy gem minerals magnetite, garnet, andopaz in Haplophragmium agglutinans and concluded that the
d within the surrounding sediment from the collection site at Bazaruto Islandal tests.laria hauerii et au sein du sédiment environnant du site de prélèvement à l’îlein des tests des foraminifères.
Quartz Calcite Orthoclase (Microcline)
7.53 6.33 0.0062.37 8.86 16.88
W.A. Makled, M.R. Langer / Revue de micropaléontologie 53 (2010) 163–173 171
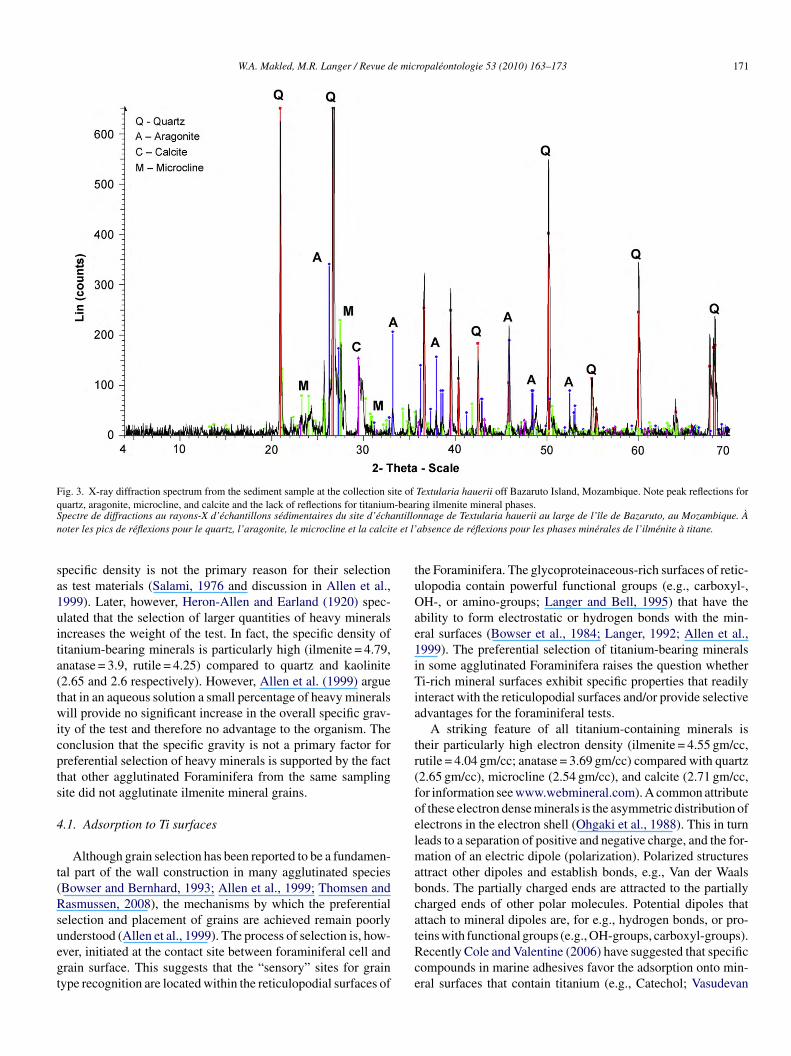
Fig. 3. X-ray diffraction spectrum from the sediment sample at the collection site of Textularia hauerii off Bazaruto Island, Mozambique. Note peak reflections forq -bearS ntillon e et l’
sa1uita(twicpts
4
t(Rsuegt
tuOae1iTia
tr(foelmabca
uartz, aragonite, microcline, and calcite and the lack of reflections for titaniumpectre de diffractions au rayons-X d’échantillons sédimentaires du site d’échaoter les pics de réflexions pour le quartz, l’aragonite, le microcline et la calcit
pecific density is not the primary reason for their selections test materials (Salami, 1976 and discussion in Allen et al.,999). Later, however, Heron-Allen and Earland (1920) spec-lated that the selection of larger quantities of heavy mineralsncreases the weight of the test. In fact, the specific density ofitanium-bearing minerals is particularly high (ilmenite = 4.79,natase = 3.9, rutile = 4.25) compared to quartz and kaolinite2.65 and 2.6 respectively). However, Allen et al. (1999) arguehat in an aqueous solution a small percentage of heavy mineralsill provide no significant increase in the overall specific grav-
ty of the test and therefore no advantage to the organism. Theonclusion that the specific gravity is not a primary factor forreferential selection of heavy minerals is supported by the facthat other agglutinated Foraminifera from the same samplingite did not agglutinate ilmenite mineral grains.
.1. Adsorption to Ti surfaces
Although grain selection has been reported to be a fundamen-al part of the wall construction in many agglutinated speciesBowser and Bernhard, 1993; Allen et al., 1999; Thomsen andasmussen, 2008), the mechanisms by which the preferential
election and placement of grains are achieved remain poorly
nderstood (Allen et al., 1999). The process of selection is, how-ver, initiated at the contact site between foraminiferal cell andrain surface. This suggests that the “sensory” sites for grainype recognition are located within the reticulopodial surfaces oftRce
ing ilmenite mineral phases.nnage de Textularia hauerii au large de l’île de Bazaruto, au Mozambique. Àabsence de réflexions pour les phases minérales de l’ilménite à titane.
he Foraminifera. The glycoproteinaceous-rich surfaces of retic-lopodia contain powerful functional groups (e.g., carboxyl-,H-, or amino-groups; Langer and Bell, 1995) that have the
bility to form electrostatic or hydrogen bonds with the min-ral surfaces (Bowser et al., 1984; Langer, 1992; Allen et al.,999). The preferential selection of titanium-bearing mineralsn some agglutinated Foraminifera raises the question whetheri-rich mineral surfaces exhibit specific properties that readily
nteract with the reticulopodial surfaces and/or provide selectivedvantages for the foraminiferal tests.
A striking feature of all titanium-containing minerals isheir particularly high electron density (ilmenite = 4.55 gm/cc,utile = 4.04 gm/cc; anatase = 3.69 gm/cc) compared with quartz2.65 gm/cc), microcline (2.54 gm/cc), and calcite (2.71 gm/cc,or information see www.webmineral.com). A common attributef these electron dense minerals is the asymmetric distribution oflectrons in the electron shell (Ohgaki et al., 1988). This in turneads to a separation of positive and negative charge, and the for-
ation of an electric dipole (polarization). Polarized structuresttract other dipoles and establish bonds, e.g., Van der Waalsonds. The partially charged ends are attracted to the partiallyharged ends of other polar molecules. Potential dipoles thatttach to mineral dipoles are, for e.g., hydrogen bonds, or pro-
eins with functional groups (e.g., OH-groups, carboxyl-groups).ecently Cole and Valentine (2006) have suggested that specificompounds in marine adhesives favor the adsorption onto min-ral surfaces that contain titanium (e.g., Catechol; Vasudevan
1 e mic
ausLsbPttitIctla
tittoiEdHemOabwtiF
5
Tt
•
•
•
•
•
C
A
boAfaD5tfG
A
l
R
A
A
A
A
A
B
B
B
72 W.A. Makled, M.R. Langer / Revue d
nd Stone, 1996). Other potential partners that may form partic-larly strong complex bonds with titanium-containing mineralurfaces are glycoproteins or carbohydrates (Bowser et al., 1984;anger, 1992; Allen et al., 1999). The formation of particularlytrong bonds to titanium surfaces was shown to be mediatedy the extracellular matrix proteins (Shubayev et al., 2004).ossibly, the strong bonds between the titanium-bearing agglu-
inate and the proteinaceous adhesives of the foraminifer mayhus constitute a selective advantage for the mechanical stabil-ty and hardness of foraminiferal shells. The comparatively largeests of T. hauerii and the abundance of this taxon off Bazarutosland implies that the preferential incorporation of the titanium-ontaining mineral ilmenite may constitute a selective advantagehat increases the mechanical stability of the test, which in turneads to improved protection against potential predators (Langernd Bell, 1995; Langer et al., 1995).
Our study on the preferential agglutination of ilmenite consti-utes the first report of this mineral to be incorporated selectivelyn Foraminifera. It also provides additional evidence thatitanium-containing minerals are selectively incorporated intohe agglutinate of some Foraminifera. The preferential selectionf titanium-containing minerals has previously been reportedn two species of agglutinated Foraminifera: Heron-Allen andarland (1913) were the first to described the directed uptake ofetrital rutile needles (TiO2) in the agglutinate of B. argenteuseron-Allen and Earland (Cole and Valentine, 2006). The pref-
rential and systematic agglutination of the titanium-bearingineral anatase (TiO2) was reported in A. balkwilli Haynes.ur observations of the preferential inclusion of ilmenite thus
dd to the existing evidence that the incorporation of titanium-earing minerals is not a random process but possibly moreidespread than previously thought. Further studies are required
o clarify the role of specific types of bonds and the moleculesnvolved in the process of mineral selection in agglutinatedoraminifera.
. Conclusions
Examination and analysis of the agglutinated particles in. hauerii from Bazaruto Island Archipelago (Mozambique) leado the following principal conclusions:
ilmenite grains are selectively incorporated into the agglu-tinated tests of this biserial foraminifer. Our observationsprovide the first record of ilmenite selection in Foraminifera;the positioning and enhanced concentration of ilmenite in thetests imply a selective mechanism to regulate the timing andplacement of grains during chamber and test formation;the detection of preferential uptake and enhanced concen-tration represent the first record of ilmenite selection inForaminifera;the abundance and size of T. hauerii suggest the possibilitythat the preferential incorporation of the titanium-containing
mineral ilmenite may constitute a selective advantage thatincreases the mechanical stability of the test;further studies involving the interactions between electrostat-ically charged mineral surfaces and pseudopodial compoundsB
ropaléontologie 53 (2010) 163–173
are required to shed further light on the mechanisms thatfacilitate selective agglutination in Foraminifera.
onflict of interest
The authors have not declared any conflict of interest.
cknowledgements
The authors are grateful to Chris Ballhaus, and Bruno Bar-ier for carrying out diffraction analysis. Constructive commentsn an earlier version by Sam Bowser are greatly appreciated.ndy Gooday, Michael Kaminski, and Ann Valentine care-
ully reviewed the manuscript, and provided helpful commentsnd suggestions. This study was supported by a grant from theeutsche Forschungsgemeinschaft awarded to ML (La 884/5-1,-2). Walid Makled is supported by a scholarship from the Egyp-ian Ministry of High Education. We also thank Taniel Danielanor organizing a stimulating and productive meeting in honor oférard Bignot.
ppendix A. Systematic information.
Textularia hauerii d’Orbigny, 1846.1846. Textularia hauerii d’Orbigny, p. 250, p. xv, figs. 13–15.1915. Textularia hauerii d’Orbigny, Heron-Allen and Ear-
and, p. 626, pl. XLVII, figs. 21–23.
eferences
llen, K., Roberts, S., Murray, J.W., 1999. Marginal marine agglutinatedForaminifera: Affinities for mineral phases. Journal of Micropaleontology18, 183–191.
lmogi-Labin, A., Hemleben, C., Meischner, D., Erlenkeuser, H., 1996.Response of Red Sea deep-water agglutinated Foraminifera to deep water-mass changes during the late quaternary. Marine Micropaleontology 28,283–297.
rmitage, S.J., Botha, G.A., Duller, G.A.T., Wintle, A.G., Rebêlo, L.P.,Momade, F.J., 2006. The formation and evolution of the barrierislands of Inhaca and Bazaruto, Mozambique. Geomorphology 82,295–308.
rmynot du Châtelet, E., Recourt, P., Chopin, V., 2008. Mineralogy ofagglutinated benthic Foraminifera; implications for paleo-environmentalreconstructions. Bulletin de la Société géologique de France 179 (6),583–593.
vnimelech, M., 1952. Revision of the tubular Monothalamia. Contributionsfrom the Cushman Foundation for Foraminiferal Research 3, 60-68.
ender, H., 1989. Gehäuseaufbau, Gehäusegenese und Biologie agglutinierterForaminiferen (Sarcodina Textulariina). Jahrbuch Geologische Bunde-sanstalt 132, 259–347.
owser, S.S., Bernhard, J.M., 1993. Structure, bioadhesive distribution andelastic properties of the test of Astrammina rara (protozoa; Foraminifera).Journal of Eukaryotic Microbiology 40, 121–131.
owser, S.S., Bernhard, J.M., Habura, A., Gooday, A.J., 2002. Structure, taxon-omy and ecology of Astrammina triangularis (Earland), an allogromiid-likeagglutinated foraminifer from Explorers Cove, Antarctica. Journal of
Foraminiferal Research 32, 364–374.owser, S.S., Israel, H.A., McGee-Russell, S.M., Rieder, C.L., 1984. Surfacetransport properties of reticulopodia; do intracellular and extracellular motil-ity share a common mechanism? Cell Biology International Reports 8,1051–1062.

e mic
C
C
C
D
D
G
G
G
H
H
H
H
H
H
H
H
H
J
K
L
L
L
L
L
L
M
O
d
R
R
R
S
S
S
S
T
T
T
V
W
on agglutinated Foraminifera. Grzybowski Foundation Special Publication
W.A. Makled, M.R. Langer / Revue d
arpenter, W.B., 1875. The microscope and its revelations, 5th edition, preparedwith the assistance of H.J. Slack: pp. I–XXXII, 1-848. J. and A. Churchill,London.
ole, K.E., Valentine, A.M., 2006. Titanium biomaterial: Titania needles inthe test of the foraminiferan Bathysiphon argenteus. Dalton Transactions 3,430–432.
orliss, B.H., Milliman, J.D., 1981. The use of phillipsite in test construction ofagglutinated deep-sea benthonic Foraminifera. Sedimentology 28, 401–406.
eLaca, T.E., Bernhard, J.M., Reilly, A.A., Bowser, S.S., 2002. Notodendrodeshyalinosphaira (sp. nov.): Structure and autecology of an allogromiid-likeagglutinated foraminifer. Journal of Foraminiferal Research 32, 177–187.
ick, A.B., 1925. On needles of rutile in the test of Bathysiphon argenteus.Transactions of the Edinburgh Geological Society 12, 19–21.
ooday, A.J., Bernhard, J.M., Bowser, S.S., 1995a. The taxonomy and ecologyof Crithionina delacai sp. nov., an abundant large foraminifer from ExplorersCove, Antarctica. Journal of Foraminiferal Research 25, 290–298.
ooday, A.J., Nott, J.A., Davis, S., Mann, S., 1995b. Apatite particles in thetest wall of the large agglutinated foraminifer Bathysiphon major (Protista).Journal of the Marine Biological Association of the United Kingdom 75,469–481.
ooday, A.J., Pond, D.W., Bowser, S.S., 2002. Autecology and trophic strategyof the large agglutinated foraminiferan Bathysiphon capillare de Folin, 1886(Protista) in the bathyal Northern Rockall Trough (NE Atlantic). MarineEcology Progress Series 245, 69–82.
eron-Allen, E., 1915. A short statement upon the theory, and the phenomenaof purpose and intelligence exhibited by the protozoa, as illustrated by selec-tion and behaviour in the Foraminifera. Journal of the Royal MicroscopicalSociety 6, 547–557.
eron-Allen, E., Earland, A., 1909. On a new species of Technitella from theNorth Sea, with some observations upon selective power as exercised bycertain species of arenaceous Foraminifera. Journal of the Quekett Micro-scopical Club, Series 2 10, 403–412.
eron-Allen, E., Earland, A., 1913. Clare Island Survey. Part 64, Foraminifera.Proceedings of the Royal Irish Academy 31, 1–188.
eron-Allen, E., Earland, A., 1915. The Foraminifera of the KerimbaArchipelago (Portuguese East Africa). Part II. In: Transactions of the Zoo-logical Society of London, 20, pp. 543–794.
eron-Allen, E., Earland, A., 1920. An experimental study of the foraminiferalspecies Verneuilina polystropha (Reuss); and some others, being a contribu-tion to a discussion on the origin, evolution and transmission of biologicalcharacters. Proceedings of the Royal Irish Academy B35, 153–177.
ofker, J., 1953. Arenaceous test in Foraminifera-chalk of silica? Micropaleon-tologist 7, 65–66.
olbourn, A.E.L., Kaminski, M.A., 1997. Lower Cretaceous deep-water ben-thic Foraminifera of the Indian Ocean. Grzybowski Foundation SpecialPublication 4, 1–172.
ottinger, L., 1986. Construction, structure and function of foraminiferal shells.In: Leadbeater, B., Riding, R. (Eds.), Biomineralisation in lower plantsand animals. Systematic association special, 30. Clarendon, Oxford, pp.219–239.
ottinger, L., Halicz, E., Reiss, Z., 1993. Recent Foraminiferida from the Gulfof Aqaba, Red Sea. Slovenska Akademija Znanosti in Umetnosti, Ljubljana,Classis IV, Historia Naturalis, Dela Opera 33, pp. 179.
ørgensen, N.O., 1977. Wall structure of some arenaceous Foraminifera fromMaastrichtian White Chalk (Denmark). Journal of Foraminiferal Research7, 313–321.
aminski, M.A., Armitage, D.A., Jones, A.P., Coccioni, R., 2008. Shockeddiamonds in agglutinated Foraminifera from the Cretaceous/Paleogeneboundary, Italy: A preliminary report. In: Kaminski, M.A., Coccioni, R.
(Eds.), Proceedings of the seventh International Workshop on AgglutinatedForaminifera, 13. Grzybowski Foundation Special Publication, pp. 57–61.anger, M.R., 1992. Biosynthesis of glycosaminoglycans in Foraminifera: Achoreography by ribosomes, the endoplasmatic reticulum and the golgicomplex. Marine Micropaleontology 19, 245–255.
W
ropaléontologie 53 (2010) 163–173 173
anger, M.R., Bell, C., 1995. Toxic Foraminifera: Innocent until proven guilty.Marine Micropaleontology 24, 205–214.
anger, M.R., Lipps, J.H., Moreno, G., 1995. Predation on Foraminifera bythe deep-sea scaphopod Fissidentalium megathyris. Deep-Sea Research 42,849–857.
eCalvez, Y., Cesana, D., 1980. Regarding the genus Hemisphaerammina Loe-blich and Tappan (Foraminifera). Cushman Foundation for ForaminiferalResearch Special Publication 19, 215–224.
ipps, J.H., 1973. Test structure in Foraminifera. Annual Review Microbiology27, 471–488.
oeblich, A.R., Tappan, H., 1964. Sarcodina, chiefly “Thecamoebians” andForaminiferida. In: Moore, R.C. (Ed.), Treatise on Invertebrate Paleontol-ogy, Part C, Protista 2. Geological Society of America and University ofKansas Press, pp. C89–C91.
cIlroy, D., Green, O.R., Brazier, B.B., 1994. The world’s oldest foraminiferans.Microscopy and Analysis, November, 13–15.
hgaki, K., Ohgaki, M., Tanaka, K., Marumo, F., Takei, H., 1988. Electron-density distribution in ilmenite-type crystals, IV. Iron (II) titanium (IV)trioxide, FeTiO3. Mineralogica Journal 14, 179–190.
’Orbigny, A., 1846. Foraminifères fossiles du Bassin Tertiaire de Vienne(Autriche). Gide et Compe, Paris.
asband, W.S., 2008. ImageJ, U.S. National Institutes of Health, Bethesda,Maryland, USA, http://rsb.info.nih.gov/ij/, 1997–2008.
eiss, Z., Hottinger, L., 1984. The Gulf of Aqaba. Ecological Micropa-leontology. In: Ecological Studies 50. Springer-Verlag, Heidelberg,354.
oberts, S., Murray, J.W., 1995. Characterization of cement mineralogy inagglutinated Foraminifera (Protista) by Raman spectroscopy. Journal of theGeological Society, London 152, 7–9.
alami, M.B., 1976. Biology of Trochammina cf. T. quadriloba Höglund(1947), an agglutinating foraminifer. Journal of Foraminiferal Research 6,142–153.
hubayev, V.I., Brånemark, R., Steinauer, J., Myers, R.R., 2004. Titaniumimplants induce expression of matrix metalloproteinases in bone duringosseointegration. Journal of Rehabilitation Research and Development 41,757–766.
liter, W.V., 1968. Shell material variation in the agglutinated foraminiferTrochammina pacifica Cushman. Tulane Studies Geology and Paleontology6, 80–84.
mith, M.A., Kaesler, R.L., 1970. Selection of adventitious test mate-rial by Reophax curtus (Foraminiferida). Journal of Paleontology 44,953–957.
appan, H., 1962. Foraminifera from arctic slope of Alaska. United StatesGeological Survey Professional Paper 236-C, 91–209.
homsen, E., Rasmussen, T.L., 2008. Coccolith-agglutinating Foraminiferafrom the early Cretaceous and how they constructed their tests. Journal ofForaminiferal Research 38, 193–214.
yler, R.M., Minnitt, R.C.A., 2004. A review of sub-Saharan heavy mineral sanddeposits: Implications for new projects in Southern Africa. Journal of theSouth African Institute of Mining and Metallurgy 104, 89–100.
asudevan, D., Stone, A.T., 1996. Adsorption of catechols, 2-aminophenols, and1,2-phenylenediamines at the metal (Hydr) oxide–water interface: Effect ofring substituents on the adsorption onto TiO2. Environmental Science andTechnology 30, 1604–1613.
idmark, J.G.V., Henriksson, A.S., 1995. The “orphaned” agglutinatedForaminifera: Gaudryina cribrosphaerellifera n. sp. from the upper Cre-taceous (Maastrichtian) Central Pacific Ocean. In: Kaminski, M.A., Geroch,S., Gasinski, M.A. (Eds.), Proceedings of the fourth International Workshop
3, pp. 292–300.ollenburg, J., 1992. Zur Taxonomie von rezenten benthischen Foraminiferen
aus dem Nansen Becken, Arktischer Ozean. Berichte zur Polarforschung112, 1–137.





























