Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Practice makes transfer of motor skills imperfect
Arnaud Boutin • Arnaud Badets • Robin N. Salesse •
Udo Fries • Stefan Panzer • Yannick Blandin
Received: 12 April 2011 / Accepted: 2 June 2011 / Published online: 14 June 2011
� Springer-Verlag 2011
Abstract We investigated the practice-effects on motor
skill transfer and the associated representational memory
changes that occur during the within-practice and between-
practice phases. In two experiments, participants produced
extension–flexion movements with their dominant right
arm for a limited or prolonged practice session arranged in
either a single- or multi-session format. We tested the
ability of participants to transfer the original pattern
(extrinsic transformation) or the mirrored one (intrinsic
transformation) to the non-dominant left arm, 10 min and
24 h after the practice sessions. Results showed that prac-
tice induces rapid motor skill improvements that are non-
transferable irrespective of the amount of acquisition trials.
Furthermore, the extrinsic component of the skill develops
early and remains the dominant coding system during
practice. Conversely, we found distinct between-practice
memory changes: a limited practice induces an off-line
development of the extrinsic component, whereas a pro-
longed practice session subserves the off-line development
of the intrinsic component (Experiment 2). We provided
further evidence that the long-term representation of the
motor skill also depends on the nature of the practice
session itself: the parsing of practice into multiple sessions
narrows the effector-transfer capacities in comparison to a
single session (Experiment 1). These findings yield theo-
retical and practical implications that are discussed in the
context of recent motor skill learning models.
Introduction
Growing evidence, spanning behavioural and neurophysi-
ological studies, supports the notion that the acquisition of
a new motor skill is mediated by distinct successive phases:
a fast learning, within-practice performance improvements
phase, followed by a slow learning phase consisting of
delayed (off-line), time-dependent performance improve-
ments emerging after practice (e.g., Karni, Meyer, Jezzard,
Adams, Turner, & Ungerleider, 1995; Korman, Raz, Flash,
& Karni, 2003; Robertson, Pascual-Leone, & Press, 2004b;
see also Krakauer & Shadmehr 2006; Stickgold & Walker
2007, for reviews). Current hypotheses state that a specific
and labile memory representation of the motor skill is
formed during practice. This newly memory trace will then
undergo further modifications during a period of consoli-
dation, in the absence of additional practice (e.g., Robertson,
Pascual-Leone, & Miall, 2004a; Walker & Stickgold, 2004;
see Krakauer & Shadmehr, 2006; Robertson, 2009, for a
review). Consolidation refers to the off-line process (see
McGaugh, 2000, for a review) that transforms new and
initially labile memories into more stable representations
that become integrated into the network of pre-existing
long-term motor memories ([24 h; see Krakauer &
Shadmehr, 2006, for a review).
Part of this work was presented at the conference of the North
American Society for the Psychology of Sport and Physical Activity,
Tucson (AZ), 2010.
A. Boutin � A. Badets � R. N. Salesse � Y. Blandin (&)
National Centre of Scientific Research, Centre de Recherches sur
la Cognition et l’Apprentissage, CeRCA, CNRS UMR 6234,
MSHS. Bat A5, University of Poitiers, 5, rue Theodore Lefebvre,
86000 Poitiers, France
e-mail: [email protected]
U. Fries
Department of Human Movement Science,
University of Leipzig, Leipzig, Germany
S. Panzer
IfADo, Leibniz Research Centre for Working Environment
and Human Factors, Dortmund, Germany
123
Psychological Research (2012) 76:611–625
DOI 10.1007/s00426-011-0355-2

Numerous theoretical perspectives proposed that the
representation of motor skills relies on distinct and inde-
pendent coordinate or coding systems (e.g., Criscimagna-
Hemminger, Donchin, Gazzaniga, & Shadmehr, 2003;
Hikosaka, Nakahara, Rand, Sakai, Lu, Nakamura, et al.,
1999; Lange, Godde, & Braun, 2004). Each coding system
contributes to movement production and could produce
specific learning and transfer capacities. While the visually
acquired information about movement and target positions
are initially encoded in an eye-centred, extrinsic world-
based reference frame, the muscular activation patterns are
then encoded in an intrinsic, body-centred reference frame
(e.g., Colby & Goldberg, 1999; Soechting & Flanders,
1989). The intrinsic code is represented in terms of an
internal model of joint representations (Criscimagna-
Hemminger et al., 2003), musculoskeletal forces and
dynamics (Krakauer, Ghilardi, & Ghez, 1999), which takes
into the account of orientation of body segments relative to
each other (Lange et al., 2004; Soechting & Flanders,
1989). The intrinsic coding system is supposed to result in
an effector-dependent representation of the motor skill (i.e.,
without effector transfer capacity) to the extent that bio-
mechanical, neurological, and dynamic properties of the
effectors used on a transfer test are dissimilar to those used
during practice. Conversely, the extrinsic code reflects
Cartesian coordinates of the task space with respect to the
visual display. This code is represented in an effector-
independent representation (i.e., effector transfer capacity)
when the extrinsic coordinates are reinstated during an
effector transfer test even though intrinsic characteristics of
the required movement are altered (Hikosaka et al., 1999).
Many studies have investigated the capacity of indi-
viduals to transfer skills between hands/arms using inter-
manual transfer paradigms (e.g., Cohen, Pascual-Leone,
Press, & Robertson, 2005; Kovacs, Boyle, Gruetzmatcher,
& Shea, 2010; Kovacs, Han, & Shea, 2009; Panzer,
Krueger, Muehlbauer, Kovacs, & Shea, 2009). However,
the vast majority of studies have almost exclusively
focused on the long-term effects of practice on skill
transfer (e.g., Boutin, Fries, Panzer, Shea, & Blandin, 2010;
Kovacs, Muehlbauer, & Shea, 2009), on the practice-rela-
ted memory changes (e.g., Karni, Meyer, Rey-Hipolito,
Jezzard, Adams, Turner, et al., 1998; Korman et al., 2003)
and on the degree of overnight/overday improvements
(e.g., Cohen et al., 2005; Robertson, Press, & Pascual-
Leone, 2005; Witt, Margraf, Bieber, Born, & Deuschl,
2010). But the assumption that the coding systems may be
enhanced in distinct learning phases (within- and between-
practice) has not yet been directly addressed. A protocol is
lacking that allows direct assessment of both within- and
between-practice session improvements, and their relative
contribution to the development of a specific coding sys-
tem. The main purposes of this study were, therefore, to
assess the evolution of the coding systems through practice
(Experiment 1) and to further investigate the rapid and
delayed practice-dependent changes (Experiment 2) that
contribute to the formation of long-term motor skill
representations.
The questions addressed in the current study required
the use of a particular experimental design where partic-
ipants had to perform rehearsed retention and transfer tests
(i.e., testing and re-testing sessions). However, numerous
studies (see Roediger & Butler, 2010; Roediger & Kar-
picke, 2006b, for reviews) in the verbal skill literature
have shown that if the period of time devoted to learning
includes at least one test, performance on a final test is
improved. This finding is known as the testing effect (see
Roediger & Karpicke, 2006a, for a review). Recent
research provided further evidence that testing rehearsal
can be used to further promote transfer on the final test
(e.g., Butler, 2010; McDaniel, Roediger, & McDermott,
2007, for a review). In their review, Roediger and Butler
(2010) argued that testing rehearsal promotes the acqui-
sition and long-term retention of the acquired knowledge
that can be flexibly retrieved and transferred to different
contexts, thus suggesting that the mnemonics benefits of
testing extend beyond the retention of a specific response.
The retrieval of information from memory during testing
may be the central mechanism that produces learning
benefits during a final test (Butler, 2010). One possible
explanation is that combining practice with testing session
provides greater encoding variability than single practice
(McDaniel & Masson, 1985). Thereby, testing rehearsal
can be a potent memory enhancer, not just a neutral event
as usually wrongly considered (e.g., Carpenter, 2009;
Roediger & Butler, 2010; Rohrer, Taylor, & Sholar, 2010,
for similar position). It should be noted that testing-effect
studies were traditionally conducted, with very few
exceptions, using word lists (e.g., McDaniel & Masson,
1985; Wheeler et al. 2003) or educationally relevant sit-
uations (e.g., Roediger & Karpicke, 2006b; Rohrer et al.,
2010) as materials. However, to the best of our knowl-
edge, the extent to which testing rehearsal can interact
with motor skill learning and transfer remains unknown.
Therefore, a condition without testing rehearsal was also
included in all experiments.
We used a continuous dynamic arm movement task that
required participants to make a sequence of extension-
flexion movements with their dominant right arm to
reproduce a specific pattern of displacement over time. To
test whether the contribution of the extrinsic and intrinsic
coding systems to skill learning and transfer depends on the
amount of practice, performance on retention and transfer
tests was systematically evaluated after both limited and
prolonged practice sessions. We used a retention test
requiring the participants to produce the spatial–temporal
612 Psychological Research (2012) 76:611–625
123

pattern with the same arm that was used during the practice
trials and two effector transfer tests with the unpractised
arm (see Kovacs et al., 2009; Panzer et al., 2009 for a similar
procedure). In the ‘‘intrinsic transformation’’ test, the origi-
nal pattern was mirrored so that the sequential movements
remain the same when transferred to the non-dominant left
arm (i.e., the same pattern of muscle activation and limb joint
angles). This test assessed the contribution of the intrinsic
component to transfer and learning. In the ‘‘extrinsic trans-
formation’’ test, the original pattern was preserved but per-
formed with the non-dominant left arm (i.e., the same goal
movement pattern). This test assessed the contribution of the
extrinsic component to transfer and learning because it
implicated the same spatial positions but a different pattern
of muscle activation compared with that used during prac-
tice. It should be noted that ineffective transfer performance
for one or both effector transfer tests suggests that
the extrinsic and/or intrinsic components of the motor skill
were not the dominant coding system(s) for sequence
production.
Experiment 1
The primary purpose of Experiment 1 was to investigate
the evolution of the coding systems with practice. We
evaluated retention and effector transfer performances after
a limited (18 acquisition trials) and a prolonged (135
additional acquisition trials) practice session. This design
involved performing rehearsed retention and transfer tests
(repeated testing condition). Therefore, we included a
condition that required participants to complete the same
amount of practice without retention/transfer tests
between-practice sessions (non-repeated testing condition).
Our predictions for Experiment 1 were based upon
previous theoretical perspectives (e.g., Bapi, Doya, &
Harner, 2000; Hikosaka et al., 1999, 2002; Keele, Ivry,
Mayr, Hazeltine, & Heuer, 2003; Keele, Jennings, Jones,
Caulton, & Cohen, 1995), which suggested that the repre-
sentation of motor skills evolves from an extrinsic (effec-
tor-independent) to an intrinsic (effector-dependent)
dominant coding system with practice. Thus, we predicted
that a limited practice session would yield better perfor-
mance on both the retention and extrinsic transformation
tests than on the intrinsic transformation test. Conversely,
we expected an opposite pattern of results for prolonged
practice with better performance on the intrinsic transfor-
mation test than on the extrinsic transformation test.
Finally, the repeated testing condition should favour
incremental gains in performance from testing to retesting
and, consequently, could modify the relationship between
the amount of practice and the type of coding used to
produce the task (e.g., Roediger & Butler, 2010).
Method
Participants
Thirty self-declared right-handed undergraduate students
(mean age 19.1 years, SD 0.9 years; 12 women) volun-
teered to participate in this study. Each participant was
requested to read and sign an informed consent form prior
to participation in the experiment. None of the participants
had previous experience with the experimental task, and
they were unaware of the specific purposes of the study. A
local ethics committee approved the protocol.
Apparatus
The apparatus consisted of a horizontal lever supported at
one end by a vertical axle that turned in a ball-bearing
support in a manner that was almost frictionless. The
support was fixed on a table facing the participant, allow-
ing the lever to move in the transversal plane over the table
surface. At the other end of the lever, a vertical handle was
fixed. The handle’s position could be adjusted so that when
the participant grasped the handle, his/her elbow was
aligned with the axis of rotation (Fig. 1, top left). A
potentiometer was attached to the handle’s axis of rotation
to record the position, and its output was sampled at
1000 Hz. The potentiometer data were used to provide
lever position information to the participant and stored for
later analysis. A wooden cover was placed over the table to
prevent participants from seeing the lever and their arm. A
video projector was used to display the goal movement
pattern, the on-line position of the lever and the knowledge
of results (KR) on the wall facing the participant. Partici-
pants were seated on a height-adjustable chair, about 2 m
from the wall where a 1.64 9 1.23 m image was projected,
so that their lower arm was at an approximately 80� angle
to the upper arm at the starting position. All aspects of the
experiment were programmed with the Matlab� R2008b
software from the MathWorks� (The MathWorks, Inc.,
Natick, MA), and using the Psychophysics Toolbox
extensions (Brainard, 1997; Pelli, 1997; Kleiner, Brainard,
& Pelli, 2007).
Task, experimental groups and procedures
Participants were randomly assigned to the ‘‘repeated
testing’’ (RT group; N = 15) or ‘‘non-repeated testing’’
(NRT group; N = 15) practice condition. In the RT prac-
tice condition, participants underwent 24-h delayed reten-
tion and transfer tests after both limited and prolonged
practice sessions. In the NRT practice condition, partici-
pants were merely administered 24-h delayed retention and
transfer tests following the prolonged practice session.
Psychological Research (2012) 76:611–625 613
123

Figure 1 (bottom) presents the experimental phases for
each practice condition.
Participants were individually tested in a silent and
dimly lit room. Before beginning the experiment, each
participant received written instructions and additional
verbal information about the task and procedures. They had
to perform a sequence of extension-flexion movements
with their dominant right arm to reproduce the spatial and
temporal aspects of the goal pattern projected in front of
them (Fig. 1, top). The spatial–temporal pattern was cre-
ated by summing two sine waves with similar periods but
different amplitudes. The duration of the goal pattern was
1,500 ms, and the extension–flexion movements required
three reversals. The potentiometer output was sampled for
2,000 ms, but only the first 1,500 ms was retained for later
analysis.
At the beginning of each trial, the goal movement pattern
was displayed on the wall, and participants were asked to
move the lever to the starting position (1� area at the
beginning of the goal movement pattern). One second after
achieving the start position, a tone (50 ms in duration,
3 kHz square wave) indicated that they should perform the
task. The task required moving the lever with the dominant
right arm through a pattern of extension–flexion cycles
(three reversals) to produce the required spatial–temporal
pattern. As soon as the participant started moving, the goal
movement pattern disappeared from the screen, and a cursor
representing the position of the lever was displayed. After a
2-s interval following response completion, KR was pro-
vided by superimposing the goal movement pattern (white)
over the actual pattern (green). In addition, the root-mean-
square error (RMSE) of the actual movement from the goal
movement was displayed on the screen. RMSE is the
deviation of the actual pattern from the goal pattern calcu-
lated from the onset of movement until the end of the first
1,500 ms. RMSE is sensitive to both response bias and
within-participant variability. When presented, KR was
displayed for 5 s. Participants were instructed to reproduce
the goal movement pattern as accurately as possible and
were requested to reduce the RMSE on subsequent trials.
Groups Acquisition 1 Initial testing Acquisition 2 Subsequent testing
Day 1 Day 2 Day 2 Day 2
Repeated testing 2 blocks R - Et - It 15 blocks R - Et - It
Non-repeated testing 2 blocks 15 blocks R - Et - It
Fig. 1 Schematic illustrating the limb and criterion movement
pattern used during acquisition and on the retention and transfer
tests (top). Note that the goal movement pattern was projected onto
the wall in front of the participant. Participants used their dominant
right limb during acquisition and retention. The non-dominant left
limb was used for the extrinsic and intrinsic transformation tests. The
retention test was conducted first, and the order for the two transfer
tests (extrinsic transformation and intrinsic transformation) was
counterbalanced
614 Psychological Research (2012) 76:611–625
123
Participants in the RT and NRT conditions completed
the first acquisition phase (Acquisition 1), which was
composed of two blocks of nine trials each. During this
phase, KR was provided following each trial. Approxi-
mately 24 h after the completion of Acquisition 1, partic-
ipants assigned to the RT practice condition were
administered retention and effector transfer tests, while
their NRT practice condition counterparts were not. The
retention test consisted of one block of nine trials without
KR that required the participants to produce the movement
pattern with the same limb that was used during the prac-
tice trials. With the exception that KR was not provided
during retention, all other procedures remained the same as
those used for acquisition. In addition, two effector transfer
tests without KR were administered in a counterbalanced
order after the retention test (Fig. 1, top). The transfer tests
were performed with the contra-lateral limb (i.e., non-
dominant left arm). In the ‘‘intrinsic transformation’’ test,
the original pattern becomes mirrored so that the sequential
movements remain the same when transferred to the non-
dominant left arm (i.e., the same pattern of muscle acti-
vation and limb joint angles). In the ‘‘extrinsic transfor-
mation’’ test, the original pattern was preserved but
performed with the non-dominant left arm (i.e., the same
goal movement pattern).
With the exception that participants in the NRT practice
condition did not perform retention and transfer tests on
day 2, participants in the RT and NRT conditions contin-
ued practice for an additional acquisition phase (Acquisi-
tion 2). This acquisition phase consisted of fifteen blocks of
nine trials and involved the same experimental procedures
as in Acquisition 1. Note that participants in the RT
practice condition took a 5-min rest interval following
retention and transfer before practicing Acquisition 2.
Twenty-four-hour delayed retention and transfer tests
(extrinsic transformation and intrinsic transformation) were
conducted on day 3 for all participants in the same manner
as the initial testing session experienced in the RT
condition.
Data analysis and measures
Data reduction was performed using Matlab� (The Math-
works�, Inc., Natick, MA, USA). The individual trial time
series were used to compute lever displacement. Angular
displacement time series were filtered with a 2nd order
dual-pass Butterworth filter with a cut-off frequency of
10 Hz. Root-mean-square error was computed to estimate
performance error in achieving the goal movement pattern.
Such a measure is sensitive to both amplitude and timing
errors in the produced movement pattern relative to the
goal movement pattern. To compute RMSE, the difference
between the criterion and the filtered actual movement
pattern was computed at each data point in the time series.
Next, differences for each data point in the time series were
squared, and means of the squared differences were com-
puted on a trial basis. Finally, the square root of the mean
was computed for the final measure of RMSE. Values of
RMSE for individual trials were then averaged to yield a
global estimate of RMSE for each block (nine trials).
Results
Acquisition
Acquisition data were separately analysed. First, Acquisi-
tion 1 data were submitted to a 2 (practice conditions:
repeated testing, non-repeated testing) 9 2 (blocks 1–2)
analysis of variance (ANOVA) with repeated measures on
the last factor. The second analysis was a 2 (practice
conditions: repeated testing, non-repeated testing) 9 15
(blocks 3–17) ANOVA with repeated measures on block
using data from Acquisition 2. All significant effects were
reported at p \ 0.05, unless otherwise stated, and Duncan’s
multiple range test was used for post hoc comparisons.
Partial eta square (gp2) is the effect size reported for all
significant effects (Cohen, 1988). Outliers were removed
from the analysis (±2 SD: about 4%). Mean RMSEs during
the acquisition, retention and transfer phases is displayed in
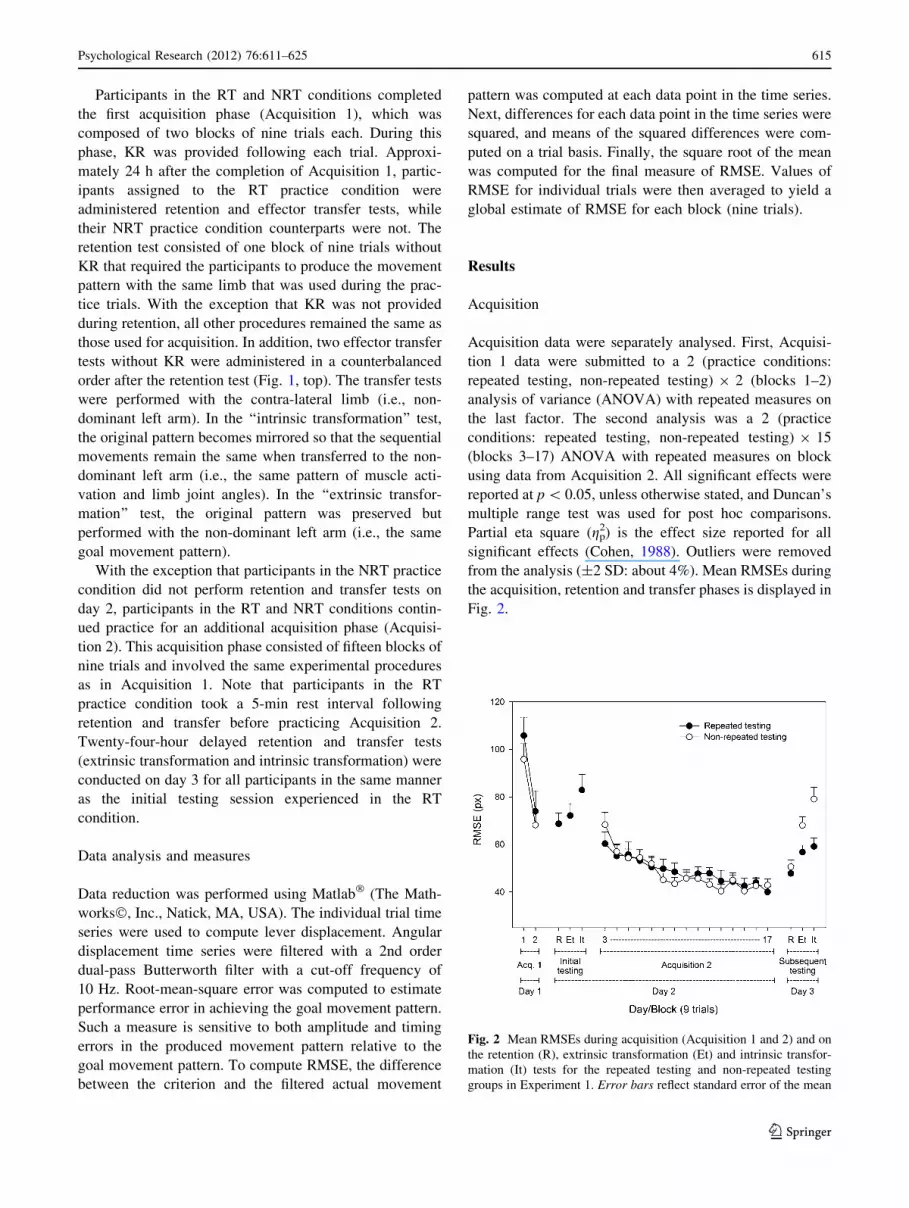
Fig. 2.
Fig. 2 Mean RMSEs during acquisition (Acquisition 1 and 2) and on
the retention (R), extrinsic transformation (Et) and intrinsic transfor-
mation (It) tests for the repeated testing and non-repeated testing
groups in Experiment 1. Error bars reflect standard error of the mean
Psychological Research (2012) 76:611–625 615
123

Acquisition 1
The analysis detected a main effect of block, F(1, 28) =
75.34, gp2 = 0.72, indicating higher RMSE on block 1
(M = 100.84) than on block 2 (M = 71.10). The main
effect of practice condition, F(1, 28) \ 1, and the practice
condition 9 block interaction were not significant, F(1,
28) \ 1.
Acquisition 2
The analysis detected a main effect of block, F(14,
392) = 13.43, gp2 = .32. Duncan’s multiple range test
revealed higher RMSE on block 3 (M = 64.31) compared
with blocks 4–7 (from M = 56.04 to M = 51.18, respec-
tively), which were not different from each other but higher
than blocks 8–17 (from M = 47.44 to M = 41.43,
respectively), which did not differ from each other. The
analysis failed to detect neither a main effect of practice
condition, F(1, 28) \ 1, nor a practice condition 9 block
interaction, F(14, 392) \ 1.
Retention and transfer
Repeated testing group: from testing to retesting
To assess the evolution of the long-term representation of
motor skills with practice, we compared performances of
the RT group on the retention and transfer tests following
limited and prolonged practice. Mean RMSEs on the
retention and transfer tests were analysed in a 2 (amounts
of practice: limited practice, prolonged practice) 9 3
(tests: retention, extrinsic transformation, intrinsic trans-
formation) ANOVA with repeated measures on the two
factors. The analysis indicated main effects of amount of
practice, F(1, 14) = 31.11, gp2 = 0.68, and test, F(2,
28) = 6.17, gp2 = 0.30, but failed to detect a significant
amount of practice 9 test interaction, F(2, 28) = 1.78.
RMSE was higher following the limited (M = 74.59) than
the prolonged practice (M = 54.58). For the main effect of
test, Duncan’s multiple range test indicated that RMSE was
significantly higher on the intrinsic transformation test
(M = 71.03) than on the retention (M = 58.26) and
extrinsic transformation tests (M = 64.47), which did not
differ from each other.
Repeated and non-repeated testing groups: subsequent
testing
Performances of the RT and NRT groups on the retention
and transfer tests following the prolonged practice session
were compared to show evidence for/against a testing
effect. RMSE was analysed in a 2 (practice conditions:
repeated testing, non-repeated testing) 9 3 (tests: reten-
tion, extrinsic transformation, intrinsic transformation)
ANOVA with repeated measures on the last factor.
The analysis revealed main effects of practice condition,
F(1, 28) = 8.09, gp2 = 0.22, test, F(2, 56) = 28.09,
gp2 = 0.50, and a significant practice condition 9 test
interaction, F(2, 56) = 5.03, gp2 = 0.15. RMSE on the
retention tests were not significantly different between the
RT (M = 47.82) and NRT groups (M = 50.59). However,
lower RMSE was found for the RT group than for the NRT
group on the extrinsic and intrinsic transformation tests.
Specifically, for the RT group, post hoc comparisons
indicated that RMSE was significantly lower on the
retention test (M = 47.82) than on the extrinsic (M =
56.78) and intrinsic transformation tests (M = 59.15),
which did not significantly differ from each other. For the
NRT group, RMSE was also significantly lower on the
retention test (M = 50.59) than on the extrinsic
(M = 68.06) and intrinsic transformation tests (M = 79.06).
However, in contrast to the RT practice condition, the anal-
ysis displayed higher RMSE on the intrinsic transformation
test compared with that on the extrinsic transformation test
(p \ 0.05) for participants assigned to the NRT group.
Discussion
Results for the acquisition indicated that the RT and NRT
groups performed similarly on the first and second practice
sessions (Acquisition 1 and Acquisition 2, respectively),
suggesting that participants in both groups improved their
performance during practice. Importantly, this finding
demonstrates that improvements during Acquisition 2 were
not testing-dependent. Indeed, participants in the NRT
practice condition did not express worse performance
during Acquisition 2 compared with their RT practice
condition counterparts. Our findings revealed that testing
rehearsal did not interact with motor skill acquisition.
Similar conclusions can be drawn regarding motor skill
retention. As revealed by performances during retention at
subsequent testing, RMSE on retention tests were identical
for the RT and NRT groups.
However, different patterns of results were observed at
subsequent transfer tests for the RT and NRT groups. This
indicates that the initial testing session, performed in the
RT practice condition, has influenced effector transfer
capacities. This is in line with previous research in the
testing-effect literature (e.g., Butler, 2010; Rohrer et al.,
2010; see Roediger & Butler, 2010; Roediger & Karpicke,
2006a, for reviews), and extend the findings from the
verbal to the motor domain. Specifically, transfer data for
the NRT group indicated that practice produced an effec-
tor-dependent representation (lower RMSE on the retention
616 Psychological Research (2012) 76:611–625
123

test compared with the transfer tests) that remained
extrinsic (lower RMSE on the extrinsic than on the intrinsic
transformation test). Conversely, testing rehearsal resulted
in a multiple coding representation (similar performances
for the RT group on the extrinsic and intrinsic transfor-
mation tests). Interestingly, this finding suggests that test-
ing rehearsal promotes the development of a multiple
coding representation that can be flexibly used from
dominant to non-dominant limb.
Because testing rehearsal interacts with motor skill
transfer, it is therefore not surprising that we did not find
any amount of practice 9 test interaction when analysing
data of the RT group. Indeed, one could assume that
retrieval practice at initial testing might have enhanced
motor skill transfer at subsequent testing, thus precluding
the possibility of additional analysis regarding the evolu-
tion of the motor skill representation from testing to
retesting. Therefore, to assess the motor skill representation
after limited practice, we further conducted a 3 (tests:
retention, extrinsic transformation and intrinsic transfor-
mation) ANOVA with repeated measures on test using the
RT group’s data at initial testing. Analysis revealed a main
effect of test, F(2, 28) = 4.47, gp2 = 0.24, with lower
RMSE on the retention (M = 68.71) and extrinsic trans-
formation tests (M = 72.16) than on the intrinsic trans-
formation test (M = 82.91). These results agree with the
theoretical perspective that an effector-independent repre-
sentation of an extrinsically transformed skill is developed
after few practice trials (Hikosaka et al., 1999, 2002; Keele
et al., 2003, 1995).
To summarise, a key finding of Experiment 1 is that the
long-term representation of the motor skill after multises-
sion practice was qualitatively (i.e., reliance on a particular
coding system) different when participants were confronted
with testing rehearsal or with a single delayed testing
session. Neither the delay between testing sessions nor the
multisession format per se could explain such differences
because the experimental design was exactly the same for
the RT and NRT practice conditions. Two issues could be
considered when interpreting these results: the opportunity
for testing rehearsal and/or the process of motor memory
consolidation that took place during the 24-h interval fol-
lowing the first and second acquisition phase. Therefore,
Experiment 2 sought to further clarify the relative contri-
bution of testing rehearsal and/or the process of memory
consolidation to motor skills coding. An additional purpose
of Experiment 2 was to investigate the rapid and delayed
practice-related memory changes that contribute to the
formation of long-term motor skill representations, using
single practice sessions composed of either 18 acquisition
trials (limited practice) or 153 acquisition trials (18 ? 135;
prolonged practice).
Experiment 2
Experiment 2 was designed to extend the findings of
Experiment 1 from multisession to single session and to
evaluate the off-line practice-related memory changes
following a single practice session. The results of Experi-
ment 1 revealed that testing rehearsal might be an effective
memory enhancer when the testing sessions are adminis-
tered between-practice sessions. Experiment 2 aimed at
determining whether off-line testing-dependent memory
enhancements of the motor skill could be observed from
testing to retesting in the absence of further task
engagement.
Participants from two groups were required to reproduce
the spatial–temporal pattern used in Experiment 1 for either
18 or 153 acquisition trials in a single practice session
before being evaluated on both 10-min and 24-h delayed
retention and transfer tests. As in Experiment 1, two other
groups practised the same goal pattern for either 18 or 153
acquisition trials but with the exception that no 10-min
retention/transfer tests were conducted, thus avoiding a
potential testing effect from 10-min testing to 24-h delayed
retesting. We expected rapid representational memory
changes with practice, as evaluated by 10-min retention
and transfer tests, with the dominance of the extrinsic
coding system after limited practice and conversely of the
intrinsic one after prolonged practice. Additionally, we
predicted that the rapid and practice-dependent memory
changes would extend beyond the practice session, i.e.,
from 10-min to 24-h testing. More specifically, a limited
practice session would favour the off-line development of
the extrinsic component of the motor skill while in con-
trast, a prolonged practice session would emphasize the
off-line development of the intrinsic coding system.
Method
Participants
Sixty self-declared right-handed undergraduate students
(mean age 21.1 years, SD 1.5 years; 21 women) volun-
teered to participate in this study. Each participant was
requested to read and sign an informed consent form prior
to participation in the experiment. None of the participants
had previous experience with the experimental task, and
they were unaware of the specific purposes of the study. A
local ethics committee approved the protocol.
Apparatus
The apparatus was identical to that used in Experiment 1.
Psychological Research (2012) 76:611–625 617
123

Task, experimental groups and procedures
Participants were randomly assigned to one of four groups
(N = 15) defined by the practice condition (repeated testing
or non-repeated testing) and the amount of practice (limited
or prolonged). While assigned to the ‘‘repeated testing’’
(RT) practice condition, participants underwent both
immediate and 24-h delayed retention and transfer tests
following either a limited or a prolonged practice session.
Conversely, while assigned to the ‘‘non-repeated testing’’
(NRT) practice condition, participants were merely
administered 24-h delayed retention and transfer tests fol-
lowing either a limited or a prolonged practice session (see
Table 1). Task and procedures for the acquisition, retention
and transfer tests were conducted in the same manner as
described in Experiment 1, except as noted.
Data analysis and measures
Data reduction was performed in the same manner as in
Experiment 1.
Results
Acquisition
Acquisition data were separately analysed. First, the lim-
ited practice data were submitted to a 2 (practice condi-
tions: repeated testing, non-repeated testing) 9 2 (blocks
1–2) ANOVA with repeated measures on the last factor.
The second analysis was a 2 (practice conditions: repeated
testing, non-repeated testing) 9 17 (blocks 1–17) ANOVA
with repeated measures on block using data from the pro-
longed practice groups. All significant effects were repor-
ted at p \ 0.05, unless otherwise stated, and Duncan’s
multiple range test was used for post hoc comparisons.
Partial eta square (gp2) is the effect size reported for all
significant effects (Cohen, 1988). Outliers were removed
from the analysis (±2 SD: about 4%). Mean RMSEs during
acquisition for the limited and prolonged practice groups
are displayed in Fig. 3.
Limited practice
The analysis revealed a main effect of block, F(1,
28) = 59.89, gp2 = 0.68, indicating that RMSE was higher
on block 1 (M = 102.01) than on block 2 (M = 68.59).
The main effect of practice condition, F(1, 28) \ 1, and the
practice condition 9 block interaction, F(1, 28) \ 1, were
not significant.
Prolonged practice
The analysis detected a main effect of block, F(16,
448) = 36.29, gp2 = 0.56. Duncan’s multiple range test
revealed that RMSE was higher on block 1 (M = 98.54)
than on block 2 (M = 69.59). Further, analysis found
higher RMSE on block 2 than on blocks 3–4 (from
M = 63.18 to M = 57.63, respectively), which was not
different from each other but higher than on blocks 5–13
(from M = 53.90 to M = 47.12, respectively), which did
not differ from each other. From block 14, RMSE remained
stable until the end of acquisition (from M = 45.02 to
M = 43.91, for blocks 14–17 respectively). The analysis
failed to detect neither a main effect of practice condition,
F(1, 28) = 2.37, nor a practice condition 9 block inter-
action, F(16, 448) \ 1.
Retention and transfer
Mean RMSEs on the retention and transfer tests were
analysed in several ways. To assess the coding systems’
time course during practice and memory consolidation, we
separately analysed performances of the RT groups (a) at
10-min testing and performances of the RT and NRT
groups at 24-h testing after (b) limited and (c) prolonged
practice (Fig. 3). Additionally, to investigate the contri-
bution of the consolidation process to the quantitative and
qualitative memory changes, we compared performance
(d) at 10-min testing to performance at 24-h retesting using
data from the RT groups (Fig. 4).
Repeated groups: 10-min testing
Mean RMSEs on the retention and transfer tests were
analysed in a 2 (amounts of practice: limited practice,
prolonged practice) 9 3 (tests: retention, extrinsic trans-
formation, intrinsic transformation) ANOVA with repeated
measures on the last factor. The analysis indicated a main
Table 1 Experimental phases for the repeated testing and non-
repeated testing groups in Experiment 2
Groups Acquisition Immediate
testing
24-h
testing
Day 1 Day 1 Day 2
Limited practice
Repeated testing 2 Blocks R–Et–It R–Et–It
Non-repeated testing 2 Blocks R–Et–It
Prolonged practice
Repeated testing 17 Blocks R–Et–It R–Et–It
Non-repeated testing 17 Blocks R–Et–It
R retention, Et extrinsic transformation, It intrinsic transformation
(order counterbalanced)
618 Psychological Research (2012) 76:611–625
123
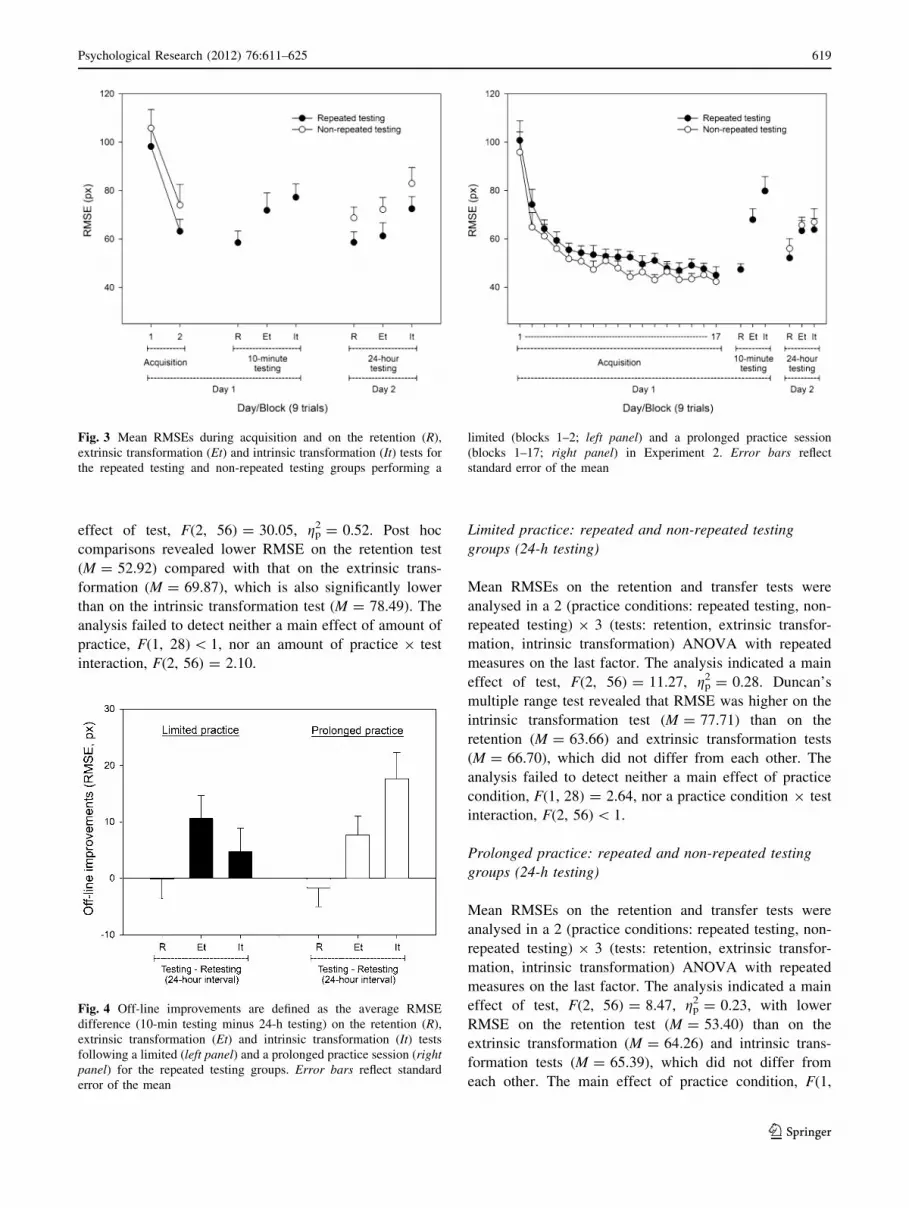
effect of test, F(2, 56) = 30.05, gp2 = 0.52. Post hoc
comparisons revealed lower RMSE on the retention test
(M = 52.92) compared with that on the extrinsic trans-
formation (M = 69.87), which is also significantly lower
than on the intrinsic transformation test (M = 78.49). The
analysis failed to detect neither a main effect of amount of
practice, F(1, 28) \ 1, nor an amount of practice 9 test
interaction, F(2, 56) = 2.10.
Limited practice: repeated and non-repeated testing
groups (24-h testing)
Mean RMSEs on the retention and transfer tests were
analysed in a 2 (practice conditions: repeated testing, non-
repeated testing) 9 3 (tests: retention, extrinsic transfor-
mation, intrinsic transformation) ANOVA with repeated
measures on the last factor. The analysis indicated a main
effect of test, F(2, 56) = 11.27, gp2 = 0.28. Duncan’s
multiple range test revealed that RMSE was higher on the
intrinsic transformation test (M = 77.71) than on the
retention (M = 63.66) and extrinsic transformation tests
(M = 66.70), which did not differ from each other. The
analysis failed to detect neither a main effect of practice
condition, F(1, 28) = 2.64, nor a practice condition 9 test
interaction, F(2, 56) \ 1.
Prolonged practice: repeated and non-repeated testing
groups (24-h testing)
Mean RMSEs on the retention and transfer tests were
analysed in a 2 (practice conditions: repeated testing, non-
repeated testing) 9 3 (tests: retention, extrinsic transfor-
mation, intrinsic transformation) ANOVA with repeated
measures on the last factor. The analysis indicated a main
effect of test, F(2, 56) = 8.47, gp2 = 0.23, with lower
RMSE on the retention test (M = 53.40) than on the
extrinsic transformation (M = 64.26) and intrinsic trans-
formation tests (M = 65.39), which did not differ from
each other. The main effect of practice condition, F(1,
Fig. 3 Mean RMSEs during acquisition and on the retention (R),
extrinsic transformation (Et) and intrinsic transformation (It) tests for
the repeated testing and non-repeated testing groups performing a
limited (blocks 1–2; left panel) and a prolonged practice session
(blocks 1–17; right panel) in Experiment 2. Error bars reflect
standard error of the mean
Fig. 4 Off-line improvements are defined as the average RMSE
difference (10-min testing minus 24-h testing) on the retention (R),
extrinsic transformation (Et) and intrinsic transformation (It) tests
following a limited (left panel) and a prolonged practice session (rightpanel) for the repeated testing groups. Error bars reflect standard
error of the mean
Psychological Research (2012) 76:611–625 619
123

28) \ 1, and the practice condition 9 test interaction, F(2,
56) \ 1, was not significant.
Off-line improvements (from testing to retesting): repeated
testing groups
We compared performance on tests conducted immediately
and tests conducted following a 24-h delay among partic-
ipants who had received either a limited or prolonged
practice session. The difference between these measures
(testing minus retesting) highlighted the off-line develop-
ment of the motor skill; a positive value reflects improved
performance. We measured off-line improvements for the
RT groups using two amounts of practice (limited practice,
prolonged practice) and three retention/transfer tests
(retention, extrinsic transformation, intrinsic transforma-
tion), resulting in a 2 9 3 factorial design with repeated
measures on the last factor. Results are depicted in Fig. 4.
The analysis indicated a main effect of test, F(2, 56) =
6.70, gp2 = 0.19, and a significant amount of practice 9 test
interaction, F(2, 56) = 3.10, gp2 = 0.10. Duncan’s multiple
range test revealed higher off-line improvements after
limited practice on the extrinsic transformation test
(M = 10.58) compared with the retention (M = -0.09)
and intrinsic transformation tests (M = 4.71), which did
not differ from each other. In contrast, following prolonged
practice, post hoc comparisons revealed higher off-line
improvements on the intrinsic transformation test (M =
17.63) than on the retention (M = -1.67) and extrinsic
transformation tests (M = 7.67), which did not differ from
each other.
Discussion
Participants demonstrated a significant decrease in RMSE
across acquisition blocks, suggesting that RT and NRT
groups similarly improved their performances during both
limited and prolonged practice sessions. In addition, anal-
ysis of the retention/transfer data at 24-h testing failed to
detect a significant interaction between the practice con-
ditions and the tests for both limited and prolonged prac-
tice. This suggests that testing rehearsal did not interact
with the off-line development of the coding systems. Our
findings argue for the evidence that testing rehearsal per se
does not induce representational memory changes of the
motor skill if no additional practice trials are performed
between testing sessions. To our knowledge, this is the first
demonstration of the interactive influences between mul-
tisession practice and testing rehearsal on the evolution of
motor skill memories.
However, although the off-line learning phase was not
influenced by testing rehearsal following a single practice
session, we found evidence that the long-term representa-
tion of motor skills depends on the amount of prior prac-
tice. Following a limited practice session, we showed a
significant advantage at 24-h testing of effector transfer for
an extrinsic transformation of the goal movement pattern
(i.e., the same spatial–temporal pattern) compared with an
intrinsic transformation (i.e., the same pattern of muscle
activation and limb joint angles). In contrast, no transfer
capacities were observed for an extrinsic or intrinsic
transformation of the original pattern following a pro-
longed practice session (similar performances on the
extrinsic and intrinsic transformation tests). Interestingly,
these findings revealed that the long-term representation of
a motor skill is based on an extrinsic coding system and at
an effector-independent level after a limited single practice
session, whereas a prolonged single practice session
resulted in a multiple coding and effector-dependent
representation.
Distinct consolidating mechanisms appear therefore to
be dependent on prior practice experience (see also
Trempe & Proteau, 2010). More specifically, we found
robust delayed gains expressed at retesting on the
extrinsic transformation test after a limited single prac-
tice session and on the intrinsic transformation test after
a prolonged single practice session. The present data are
consistent with our initial predictions, which were based
on the rationale that distinct coding systems develop off-
line and at different practice scales. We showed
between-practice memory enhancements involving an
off-line developing effector-independent component that
is represented in extrinsic coordinates early in practice,
and an off-line developing effector-dependent component
that is represented in intrinsic coordinates late in
practice.
General discussion
The main questions addressed by the present study were (1)
whether motor skill learning is mediated by different
coding systems, and (2) how those coding systems evolve
during the rapid and slow learning phases depending on the
organisation of practice and testing sessions. This study
sought to determine the underlying processes that mediate
the formation of motor skill memories. As expected, in two
experiments we showed that practice triggers within-prac-
tice and between-practice session reorganisations of the
motor skill representation, yielding distinct transfer
capacities. Our findings revealed that practice induces rapid
and non-transferable (i.e., effector-dependent representa-
tion) motor skill improvements irrespective of the amount
of acquisition trials. We found that the extrinsic component
of the skill develops early and remains the dominant coding
620 Psychological Research (2012) 76:611–625
123

system during practice. Conversely, results revealed latent
and practice-dependent reorganisations of the motor skill
during the off-line period: a limited practice induced an
off-line development of the extrinsic component, whereas a
prolonged practice session improved an off-line develop-
ment of the intrinsic component. Additionally, we provided
evidence that the long-term representation of the newly
acquired skill is also dependent on the nature of the prac-
tice session itself. Specifically, the parsing of practice into
multiple sessions provides participants with a stronger
intrinsic skill representation, which is less susceptible to
transfer in comparison to the outcome of a single practice
session. Importantly, however, we found interactive influ-
ences between multisession practice and testing rehearsal.
Results suggest that the alternation between-practice and
testing sessions provides greater transfer capacities than
does the mere repetition of practice sessions themselves.
Each of these issues will be discussed in the following
sections.
Practice and its effects on motor skill transfer
To investigate the rapid (10-min testing) and latent (24-h
testing) reorganisations of the motor skill representation
and the practice-effects on motor skill transfer, we eval-
uated the ability of participants to transfer the original
spatial–temporal pattern (extrinsic transformation) or the
mirrored one (intrinsic transformation) to the unpractised
non-dominant left arm following limited and prolonged
practice. It is important to note that the present findings
are considered for dominant arm practice but may not
generalize to practice with the non-dominant arm. Indeed,
we used unidirectional transfer tests from the dominant
practised arm to the non-dominant unpractised arm. Con-
sequently, the present finding can only be interpreted for a
motor improvement of the dominant arm. In the same
vein, recent studies (e.g., Kovacs et al., 2009; Sainburg,
2005; Stockel, Weigelt, & Krug, 2011; Wang & Sainburg,
2006) provided evidence for inter-manual transfer asym-
metries depending on the limb used during practice, i.e.,
from the dominant limb to the non-dominant limb or vice
versa.
Thus, on the one hand, our results indicated an early
setting-up of an effector-dependent representation, as
revealed by the 10-min retention/transfer tests (Experi-
ment 2). Specifically, the better performances observed on
the retention test compared with the extrinsic and intrinsic
transformation tests provide evidence for within-practice
improvements during the acquisition of skilled perfor-
mance, which is a fast and non-transferable (unpractised
arm) process. This rapid motor skill’s reorganisation arises
irrespective of the amount of practice. On the other hand,
unlike the rapid learning phase, we showed latent and
practice-dependent reorganisations of the motor skill.
After limited practice, the slow learning phase enables the
latent reorganisation of the skill memory towards an
effector-independent representation of the extrinsically
transformed skill. Conversely, following prolonged prac-
tice, the slow learning phase maintained the transfer
capacities to an effector-dependent level based on a mul-
tiple coding representation of the motor task (similar
performances on the extrinsic and intrinsic transformation
tests).
Lange et al. (2004) reported findings about the neural
processes underlying the acquisition and effector transfer
of motor skills. For instance, when participants performed
the original motor task with their unpractised left arm (here
referred to as the extrinsic transformation test), they found
increased neural activity in restricted cortical regions and
decreased activation of cortical networks. They concluded
that processes involved in the modification of the intrinsic
motor skill component included fronto-central areas as well
as the supplementary motor area. Conversely, when par-
ticipants performed the mirrored motor task with their left
arm (here referred to as the intrinsic transformation test),
they indicated that the processes underlying the modifica-
tion of the extrinsic motor skill component are controlled
by intensified parieto-frontal information flow. The pro-
duction of the mirrored motor task was associated with
suppressed cortical activity, which presumably reflects the
inhibited recall of the learned extrinsic motor skill
component.
This interpretation gains support from our results but
only during the rapid learning phase. Indeed, we showed
that the extrinsic component of the motor skill develops
early and remains the dominant coding system during
practice. Because the acquired extrinsic component is not
the appropriate coding system to perform the intrinsic
transformation test, the worse performances observed on
the intrinsic transformation test might reflect the inhibition
of the learned extrinsic component. In contrast, we
observed distinct and practice-dependent representations of
the motor skill in long-term memory, suggesting that
practice initiates latent and distinct processes during con-
solidation. The process of memory consolidation during the
off-line period is therefore differently affected by the
amount of prior practice experience.
Distinct consolidating mechanisms
The present findings confirmed the relevance of the slow
learning phase for the formation of long-term motor skill
representations. The effectiveness of the consolidation
process for motor skill learning has been extensively
Psychological Research (2012) 76:611–625 621
123

documented in the literature (e.g., Diekelmann & Born,
2010; McGaugh, 2000, for reviews). It is suggested that
consolidation refers to a process that transforms new and
labile memories encoded during practice into more stable
representations that become integrated into the network
of pre-existing long-term memories. In this view, the
present study shows for the first time that practice trig-
gers distinct consolidating mechanisms during the off-line
period. Indeed, we observed an off-line developing
effector-independent component that is represented in
extrinsic coordinates following few practice trials, and an
off-line developing effector-dependent component that is
represented in intrinsic coordinates with extensive
practice.
Recent research using imaging techniques suggested
that motor memory consolidation involves a reorganisation
of the motor skill representation residing predominantly in
M1 (e.g., Li et al. 2001; Sanes & Donoghue, 2000). The
off-line representational memory changes, presumably
initiated during practice, could be mediated by functionally
distinct subpopulations of neurons within M1 (Steele &
Penhune, 2010) or by the recruitment of additional M1
units into a local network that specifically represents the
trained motor skill with prolonged practice (Karni et al.,
1995). We therefore propose that the motor memory con-
solidation is not a single process. There might be multiple
routes to support off-line learning and engagement of these
distinct mechanisms (coding systems) is determined by
prior practice experience. It is plausible that both of these
mechanisms operate together to mediate off-line memory
processing, with one tuned to the more rapidly learned
extrinsic component of the task (following limited practice)
and the other tuned to the more slowly learned intrinsic
component of the task (following prolonged practice). This
off-line memory processing could provide the key basis for
the formation of motor skill representations in long-term
memory.
Therefore, the current study provides a critical insight
into the off-line memory processing framework. Observing
motor skill memory improvements after a single practice
session challenges the prevailing notion that the released
time is critical for motor memory consolidation. However,
it appears that single and multiple practice sessions make
distinct contributions to long-term motor skill memory
during the off-line period.
Single versus multisession practice
In the present study, we demonstrated that the long-term
representation of motor skills after multisession practice
(Experiment 1) was more resistant to transfer compared
with a single practice session (Experiment 2) when no
testing rehearsal was allowed between-practice sessions
(i.e., NRT groups).1 The combined results of Experiments
1 and 2 revealed that a single practice session promotes the
development of a multiple coding representation of the
motor skill, while the parsing of practice into multiple
sessions restricts the development of the intrinsic coding
system. This demonstrates that the long-lasting multises-
sion practice improvements cannot be conceptualised as
the sum of incremental gains expressed following single
practice sessions. That is, we postulate that distinct long-
lasting practice gains would be observed depending on
whether the amount of acquisition trials is performed
during a unique single practice session or over multiple
practice sessions.
Furthermore, we showed in Experiment 1 that the long-
term representation of the motor skill underwent qualitative
changes when comparing participants who rehearsed
retention and transfer tests (RT condition) with those who
only performed single delayed retention and transfer tests
(NRT condition). In accordance with previous research in
the testing-effect literature (e.g., Butler, 2010; Rohrer et al.,
2010), our results revealed that testing rehearsal promotes
the development of a multiple coding representation of the
motor task. This suggests that each of the extrinsic and
intrinsic codes potentially contribute to retention and
transfer performance (see also Kovacs et al., 2009). Thus,
performing several practice sessions could sustain the
development of a multiple coding representation based on
extrinsic and intrinsic components, but only when partici-
pants rehearsed retention and transfer tests between-prac-
tice sessions. In this condition, the retention and transfer
performances were determined by multiple coding systems
that acted together to enhance motor skill performance
rather than the pure dominance of a single most salient
coding system.
Our results in Experiment 1 suggest that the reactivation
and updating of existing consolidated motor memories (i.e.,
during the initial retention and transfer tests, respectively)
may be critical for further memory modifications.
1 1. We compared retention and transfer performances of the NRT
groups following a prolonged practice session arranged in a single
and multisession format (Experiments 2 and 1, respectively). We
conducted a 2 (practice formats: multisession, single session) 9 3
(tests: retention, extrinsic transformation, intrinsic transformation)
ANOVA with repeated measures on test using data from the NRT
groups. The analysis revealed a main effect of test, F(2, 56) = 18.28,
gp2 = 0.39, and a practice format 9 test interaction, F(2, 56) = 3.46,
gp2 = 0.11. Duncan’s multiple range test indicated that RMSE was
significantly higher on the intrinsic transformation test following
multisession practice (M = 79.06) than following a single practice
session (M = 66.96). No performance difference was observed
between groups on the retention (M = 56.20 and 50.59 following a
single and multisession practice, respectively) and extrinsic transfor-
mation tests (M = 65.69 and 68.06 following a single and multises-
sion practice, respectively). The main effect of practice format was
not significant, F(1, 28) \ 1.
622 Psychological Research (2012) 76:611–625
123

Likewise, recent research suggests that following their
initial acquisition and consolidation, memories can be
further modified (reconsolidated) after being reactivated
during retrieval or after additional practice. Specifically,
the reactivation of an existing motor memory yields addi-
tional strengthening and off-line gains (Walker et al. 2003).
This process, known as ‘‘reconsolidation’’ (e.g., Dudai &
Eisenberg, 2004; Przybyslawski & Sara, 1997; Stickgold &
Walker, 2007), has been suggested to enable the updating
of the internal neural representation of a previously
acquired memory (e.g., Dudai, 2006; Sara, 2000). The
phenomenon of memory reconsolidation led Censor et al.
(2010) to propose a model for human motor memory
modification. This model differentiates between what the
authors referred to as the ‘‘executing storage domain’’
(primary motor cortex) and the ‘‘core storage domain’’. The
brain areas supposed to be involved as a part of the core
storage domain which may include the cerebellum, stria-
tum and/or other motor-related cortical areas (e.g., Doyon,
Song, Karni, Lalonde, Adams, & Ungerleider, 2002;
Ungerleider, Doyon, & Karni 2002; Shadmehr & Holcomb,
1997) and the hippocampus (e.g., Albouy, Sterpenich,
Balteau, Vandewalle, Desseilles, Dang-Vu, et al., 2008).
According to this model, when the memory is reactivated,
recurrent output from the core storage domain to the exe-
cuting storage domain (which interacts with the environ-
ment) enables further memory modifications. In light of
this model, our findings suggest that testing rehearsal, when
combined to more than a single practice session, enables
the modification of existing consolidated motor memories.
This implies that testing rehearsal influences the repre-
sentation of motor skills through the process of memory
reconsolidation.
Finally, the present findings demonstrated that the
combination of practice and testing sessions provides
greater effector transfer capacities than does practice
alone–an important finding in light of the potential
behavioural and motor-related applications of the testing
effect. Indeed, our findings are important not only for
theoretical but also for practical reasons related to training
protocols designed to improve learning, as in the case
where one limb is injured, for instance. From a practical
standpoint, this research has the potential to impact the
design of training and learning therapy programs, and thus
to (re)shape the development of new learning and
relearning programs. More specifically, when elaborating
training protocols on multiple practice sessions, alternating
practice and testing sessions seems to be the best condition
for long-lasting retention of skill memories and for
enhancing transfer capacities. Obviously, the benefits of
testing rehearsal on motor skill transfer have yet to be fully
exploited.
Summary
Our findings revealed that practice does trigger rapid
(within-practice) and latent (between-practice) reorganisa-
tions of the new motor skill representation, yielding distinct
transfer capacities. While the extrinsic component of the
motor skill develops early and remains the dominant cod-
ing system during practice, we showed that practice leads
to the off-line development of distinct coding systems.
Only few practice trials induces an off-line development of
the extrinsic component of the motor skill, whereas
extensive practice subserves the off-line development of
the intrinsic component. Another important finding is that
practice makes transfer of motor skills imperfect, irre-
spective of the amount of acquisition trials and the practice
session format (single session vs. multisession practice).
Our results indicated, however, that the alternation between
practice and testing sessions provides greater transfer
capacities than does the mere repetition of practice
sessions.
Acknowledgments This work was supported by a grant from the
German Research Foundation (PA 774/8-1) and the Agence Nationale
de la Recherche (ANR-08-FASHS-14).
References
Albouy, G., Sterpenich, V., Balteau, E., Vandewalle, G., Desseilles,
M., Dang-Vu, T., et al. (2008). Both the hippocampus and
striatum are involved in consolidation of motor sequence
memory. Neuron, 58, 261–272.
Bapi, R. S., Doya, K., & Harner, A. M. (2000). Evidence for effector
independent and dependent representations and their differential
time course of acquisition during motor sequence learning.
Experimental Brain Research, 132, 149–162.
Boutin, A., Fries, U., Panzer, S., Shea, C. H., & Blandin, Y. (2010).
Role of action observation and action in sequence learning and
coding. Acta Psychologica, 135, 240–251.
Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision,10, 433–436.
Butler, A. C. (2010). Repeated testing produces superior transfer of
learning relative to repeated studying. Journal of ExperimentalPsychology. Learning, Memory, and Cognition, 36, 1118–1133.
Carpenter, S. K. (2009). Cue strength as a moderator of the testing
effect: The benefits of elaborative retrieval. Journal of Exper-imental Psychology. Learning, Memory, and Cognition, 35,
1563–1569.
Censor, N., Dimyan, M. A., & Cohen, L. G. (2010). Modification of
existing human motor memories is enabled by primary cortical
processing during memory reactivation. Current Biology, 20,
1545–1649.
Cohen, J. (1988). Statistical power analysis for the behavioralsciences (2nd ed.). Hilsdale, Nj: Erlbaum.
Cohen, D. A., Pascual-Leone, A., Press, D. Z., & Robertson, E. M.
(2005). Off-line learning of motor skill memory: a double
dissociation of goal and movement. Proceedings of the NationalAcademy of Sciences of the United States of America, 102,
18237–18241.
Psychological Research (2012) 76:611–625 623
123

Colby, C. L., & Goldberg, M. E. (1999). Space and attention in
parietal cortex. Annual Review of Neuroscience, 22, 319–349.
Criscimagna-Hemminger, S. E., Donchin, O., Gazzaniga, M. S., &
Shadmehr, R. (2003). Learned dynamics of reaching movements
generalize from dominant to nondominant arm. Journal ofNeurophysiology, 89, 168–176.
Diekelmann, S., & Born, J. (2010). The memory function of sleep.
Nature Reviews Neuroscience, 11, 114–126.
Doyon, J., Song, A. W., Karni, A., Lalonde, F., Adams, M. M., &
Ungerleider, L. G. (2002). Experience-dependent changes in
cerebellar contributions to motor sequence learning. Proceedingsof the National Academy of Sciences of the United States ofAmerica, 99, 1017–1022.
Dudai, Y. (2006). Reconsolidation: the advantage of being refocused.
Current Opinion in Neurobiology, 16, 174–178.
Dudai, Y., & Eisenberg, M. (2004). Rites of passage of the engram:
reconsolidation and the lingering consolidation hypothesis.
Neuron, 44, 93–100.
Hikosaka, O., Nakahara, H., Rand, M. K., Sakai, K., Lu, X.,
Nakamura, K., et al. (1999). Parallel neural networks for
learning sequential procedures. Trends in Neuroscience, 22,
464–471.
Hikosaka, O., Rand, M. K., Nakamura, K., Miyachi, S., Kitaguchi, K.,
Sakai, K., et al. (2002). Long-term retention of motor skill in
macaque monkeys and humans. Experimental Brain Research,147, 494–504.
Karni, A., Meyer, G., Jezzard, P., Adams, M. M., Turner, R., &
Ungerleider, L. G. (1995). Functional MRI evidence for adult
motor cortex plasticity during motor skill learning. Nature, 377,
155–158.
Karni, A., Meyer, G., Rey-Hipolito, C., Jezzard, P., Adams, M. M.,
Turner, R., et al. (1998). The acquisition of skilled motor
performance: fast and slow experience-driven changes in
primary motor cortex. Proceedings of the National Academy ofSciences of the United States of America, 95, 861–868.
Keele, S. W., Ivry, R., Mayr, U., Hazeltine, E., & Heuer, H. (2003).
The cognitive and neural architecture of sequence representation.
Psychological Review, 110, 316–339.
Keele, S. W., Jennings, P., Jones, S., Caulton, D., & Cohen, A.
(1995). On the modularity of sequence representation. Journal ofMotor Behavior, 27, 17–30.
Kleiner, M., Brainard, D., & Pelli, D. G. (2007). What’s new in
Psychtoolbox-3? Perception, 36 (ECVP Abstract Supplement).
Korman, M., Raz, N., Flash, T., & Karni, A. (2003). Multiple shifts in
the representation of a motor sequence during the acquisition of
skilled performance. Proceedings of the National Academy ofSciences of the United States of America, 100, 12492–12497.
Kovacs, A. J., Boyle, J., Gruetzmatcher, N., & Shea, C. H. (2010).
Coding of on-line and pre-planned movement sequences. ActaPsychologica, 133, 119–126.
Kovacs, A. J., Han, D.-W., & Shea, C. H. (2009a). The representation
of movement sequences is related to task characteristics. ActaPsychologica, 132, 54–61.
Kovacs, A. J., Muehlbauer, T., & Shea, C. H. (2009b). The coding of
movement sequences. Journal of Experimental Psychology:Human Perception and Performance, 35, 390–407.
Krakauer, J. W., Ghilardi, M. F., & Ghez, C. (1999). Independent
learning of internal models for kinematic and dynamic control of
reaching. Nature Neuroscience, 2, 1026–1031.
Krakauer, J. W., & Shadmehr, R. (2006). Consolidation of motor
memory. Trends in Neurosciences, 29, 58–64.
Lange, R. K., Godde, B., & Braun, C. (2004). EEG correlates of
coordinate processing during intermanual transfer. ExperimentalBrain Research, 159, 161–171.
Li, C. S. R., Padoa-Schioppa, C., & Bizzi, E. (2001). Neuronal
correlates of motor performance and motor learning in the
primary motor cortex of monkeys adapting to an external force
field. Neuron, 30, 593–607.
McDaniel, M. A., & Masson, M. E. J. (1985). Alterning memory
representations through retrieval. Journal of Experimental Psy-chology. Learning, Memory, and Cognition, 11, 371–385.
McDaniel, M. A., Roediger, H. L., I. I. I., & McDermott, K. B.
(2007). Generalizing test-enhanced learning from the laboratory
to the classroom. Psychonomic Bulletin & Review, 14, 200–206.
McGaugh, J. L. (2000). Memory—a century of consolidation.
Science, 287, 248–251.
Panzer, S., Krueger, M., Muehlbauer, T., Kovacs, A. J., & Shea, C. H.
(2009). Inter-manual transfer and practice: coding of simple
motor sequences. Acta Psychologica, 131, 99–109.
Pelli, D. G. (1997). The VideoToolbox software for visual psycho-
physics: transforming numbers into movies. Spatial Vision, 10,
437–442.
Przybyslawski, J., & Sara, S. J. (1997). Reconsolidation of memory
after its reactivation. Behavioural Brain Research, 84, 241–246.
Robertson, E. M. (2009). From creation to consolidation: A novel
framework for memory processing. PLoS Biology, 7, 11–19.
Robertson, E. M., Pascual-Leone, A., & Miall, R. C. (2004a). Current
concepts in procedural consolidation. Nature Reviews Neurosci-ence, 5, 576–582.
Robertson, E. M., Pascual-Leone, A., & Press, D. Z. (2004b).
Awareness modifies the skill-learning benefits of sleep. CurrentBiology, 14, 208–212.
Robertson, E. M., Press, D. Z., & Pascual-Leone, A. (2005). Off-line
learning and the primary motor cortex. The Journal of Neuro-science, 25, 6372–6378.
Roediger, H. L., I. I. I., & Butler, A. C. (2010). The critical role of
retrieval practice in long-term retention. Trends in CognitiveSciences, 15, 20–27.
Roediger, H. L., I. I. I., & Karpicke, J. D. (2006a). Test-enhanced
learning: taking memory tests improves long-term retention.
Psychological Science, 17, 249–255.
Roediger, H. L., I. I. I., & Karpicke, J. D. (2006b). The power of
testing memory: basic research and implications for educational
practice. Perspectives on Psychological Science, 3, 181–210.
Rohrer, D., Taylor, K., & Sholar, B. (2010). Tests enhance the
transfer of learning. Journal of Experimental Psychology.Learning, Memory, and Cognition, 36, 233–239.
Sainburg, R. L. (2005). Handedness: differential specializations for
control of trajectory and position. Exercise and Sport SciencesReviews, 33, 206–213.
Sanes, J. N., & Donoghue, J. P. (2000). Plasticity and primary motor
cortex. Annual Review of Neuroscience, 23, 393–415.
Sara, S. J. (2000). Retrieval and reconsolidation: toward a neurobi-
ology of remembering. Learning and Memory, 7, 73–84.
Shadmehr, R., & Holcomb, H. H. (1997). Neural correlates of motor
memory consolidation. Science, 277, 821–825.
Soechting, J. F., & Flanders, M. (1989). Errors in pointing are due to
approximations in sensorimotor transformations. Journal ofNeurophysiology, 62, 595–608.
Steele, C. J., & Penhune, V. B. (2010). Specific increases within
global decreases: a functional magnetic resonance imaging
investigation of five days of motor sequence learning. TheJournal of Neuroscience, 30, 8332–8341.
Stickgold, R., & Walker, M. P. (2007). Sleep-dependent memory
consolidation and reconsolidation. Sleep Medicine, 8, 331–343.
Stockel, T., Weigelt, M., & Krug (2011). Acquisition of a complex
basketball dribbling task in school children as a function of
bilateral practice order. Research Quarterly for Exercise andSport, 82, 188–197.
Trempe, M., & Proteau, L. (2010). Distinct consolidation outcomes in
a visuomotor adaptation task: off-line leaning and persistent
after-effect. Brain and Cognition, 73, 135–145.
624 Psychological Research (2012) 76:611–625
123

Ungerleider, L. G., Doyon, J., & Karni, A. (2002). Imaging brain
plasticity during motor skill learning. Neurobiology of Learningand Memory, 78, 553–564.
Walker, M. P., Brakefield, T., Hobson, J. A., & Stickgold, R. (2003).
Dissociable stages of human memory consolidation and recon-
solidation. Nature, 425, 616–620.
Walker, M. P., & Stickgold, R. (2004). Sleep-dependent learning and
memory consolidation. Neuron, 44, 121–133.
Wang, J., & Sainburg, R. L. (2006). Interlimb transfer of visuomotor
rotations depends on handedness. Experimental Brain Research,175, 223–230.
Wheeler, M. A., Ewers, M., & Buonanno, J. F. (2003). Different rates
of forgetting following study versus test trials. Memory, 11,
571–580.
Witt, K., Margraf, N., Bieber, C., Born, J., & Deuschl, G. (2010).
Sleep consolidates the effector-independent representation of a
motor skill. Neuroscience, 171, 227–234.
Psychological Research (2012) 76:611–625 625
123





























