Embed Size (px)
Citation preview

Plugging into the network: belowground connections between germlings andextraradical mycelium of arbuscular mycorrhizal fungi
Cristiana SbranaCNR, Institute of Biology and Agrobiotechnology UOSPisa, Via del Borghetto 80, 56124 Pisa, Italy
Paola FortunaManuela Giovannetti1
University of Pisa, Department of Crop Plant Biology,Via del Borghetto 80, 56124 Pisa, Italy
Abstract: Arbuscular mycorrhizal fungi (AMF) areobligate biotrophs; nevertheless their spores cangerminate in the absence of host plants. Suchinconsistent behavior is balanced by diverse survivalstrategies. The ability of AM fungal hyphae to fusemight represent a fundamental survival strategybecause germlings could plug into compatible my-corrhizal networks, thus gaining access to plant-derived carbon before asymbiotic growth arrest. Anin vivo experimental system was used to growextraradical mycelium produced by Glomus mosseaecolonizing three different plant species and germlingsof the same isolate. After symbiotic and asymbioticmycelia came into contact we showed that germlinghyphae fused with symbiotic network hyphae andestablished protoplasm connections with nuclei oc-curring in fusion bridges. The frequency of anasto-moses between germling and symbiotic hyphae was4.9–23.9%. Prefusion and postfusion incompatibleresponses, with protoplasm withdrawal in interactinghyphae, were evident in some hyphal contacts. Giventhe multigenomic nature of AMF, the mingling ofgermling nuclei with those of the mycorrhizalnetwork through perfect fusions might represent ameans for the maintenance of genetic diversity in theabsence of sexual recombination.
Key words: anastomosis, arbuscular mycorrhizalfungi (AMF), Glomus mosseae, nuclear mingling,survival strategy, vegetative compatibility and incom-patibility
INTRODUCTION
Arbuscular mycorrhizal (AM) fungi establish mutual-istic symbioses with most land plant species, improv-ing plant nutrition and fitness and affecting ecosys-tem functioning and biodiversity (van der Hejiden et
al. 1998, Smith and Read 2008). AM fungi (AMF) areobligate biotrophs unable to complete their life cycleand reproduce apart from host plants, although theirspores can germinate in different environmentalconditions in the absence of hosts. Such inconsistentbehavior, which could represent a strong selectivedisadvantage, is puzzling because AMF are consideredevolutionary successful living fossils, having survived460 000 000 y (Morton 1990, Simon et al. 1993, Remyet al. 1994, Phipps and Taylor 1996, Redecker et al.2000), supporting the view of their fundamental rolein land colonization by plants (Pirozynski andMalloch 1975, Pyrozynski 1981).
The life cycle of AMF involves diverse survivalstrategies allowing them to overcome their obligatebiotrophic status and the lack of a mechanismcontrolling spore germination by host plants (Gio-vannetti 2001). The first survival strategy is represent-ed by the wide host range (, 80% of land plantspecies), which increases the chance that myceliumfrom a single germinated spore will come into contactand colonize a host root. Such a strategy, relyingentirely on chance, appears to be a weak explanationfor 460 000 000 y (see above) to possibly 980 000 000 y(Blair 2009) of existence.
A second evolutionary mechanism developed byAMF allows for the survival of spores that germinatedin the absence of host-derived signals. The coenocyticmycelial networks produced by germlings rapidlyundergo growth arrest accompanied by protoplasmwithdrawal from hyphal tips and resource reallocationtoward mother spores (Mosse 1959, Hepper 1983,Becard and Piche 1989, Logi et al. 1998). Thisphenomenon likely is important to maintain sporereserves for retention of viability and a capacity togerminate again (Beilby and Kidby 1980, Koske 1981,Tommerup 1984, Logi et al. 1998). Energy-savingmechanisms let AMF discriminate host from non-hostroots; AMF hyphae undergo a distinctive pattern ofhyphal morphogenesis only after perceiving host-derived signals (Giovannetti et al. 1993b, 1994; Bueeet al. 2000; Akiyama et al. 2005). AMF also regulateappressoria differentiation, which occurs only on thesurface of host roots (Glenn et al. 1988, Avio et al.1990, Giovannetti et al. 1993a, Gollotte et al. 1993).
Many findings reported the ability of AM fungalhyphae originating from germlings (Giovannetti et al.1999, Giovannetti and Sbrana 2001, Croll et al. 2009)and from extraradical networks (Giovannetti et al.
Submitted 23 Apr 2010; accepted for publication 27 Sep 2010.1 Corresponding author. E-mail: [email protected]
Mycologia, 103(2), 2011, pp. 307–316. DOI: 10.3852/10-125# 2011 by The Mycological Society of America, Lawrence, KS 66044-8897
307

2004) to anastomose when belonging to the sameisolate and to genetically non-identical conspecificisolates. The ability of AM fungal mycelium toanastomose might represent a survival strategy forgermlings growing in the absence of the host. Ifgermling hyphae are better able to contact the largeextraradical networks, as opposed to reaching hostroots, hyphal anastomosis would allow incorporationinto compatible mycorrhizal mycelia. This potentiallylets germlings access plant-derived carbon beforegrowth arrest and increases probability of survival. Asimilar event has been described to occur inectomycorrhizal fungi, between monokaryotic germl-ings and dikaryotic hyphae (Gardes et al. 1990,Kytoviita 2000), but has not been observed in AMF.AM fungal germlings recognize hyphae belonging togeographically different conspecific isolates as non-self leading to incompatible reactions that preventhyphal fusion (Giovannetti et al. 2003). Anastomosisis observed only when germlings of genetically similarAM fungal isolates interact (Croll et al. 2009).
To date no study has demonstrated interactionsbetween asymbiotic and symbiotic hyphae in AMF.AM fungal mycelium shows differential colonizationability and gene expression depending on whether itoriginates from asymbiotic versus symbiotic phase(Harrison and van Buuren 1995, David-Schwartz et al.2001, Breuninger and Requena 2004). However theability of hyphae produced by germlings to interactand fuse with symbiotic mycelium is unknown.
In this work an in vivo two-dimensional experimen-tal system allowing the analysis of intact extraradicalmycorrhizal networks produced by the AM fungusGlomus mosseae living in symbiosis with three hosts(Petroselinum hortense, Solanum melongena and Pru-nus cerasifera) was used to investigate the ability ofgermlings to plug into the symbiotic mycelial net-works by means of hyphal fusions. To this aim weassessed (i) the occurrence and frequency of anasto-moses between germlings and extraradical hyphae,(ii) the occurrence and frequency of hyphal incom-patibility responses, (iii) viability and nuclear pres-ence in anastomosing hyphae.
MATERIALS AND METHODS
Fungal material.—The AM fungal species used was Glomusmosseae (Nicol. & Gerd.) Gerdemann & Trappe(Rothamsted isolate, local code IMA1), obtained from potcultures maintained in the collection of the MicrobiologyLaboratory at the Department of Crop Plant Biology,University of Pisa, Italy. Voucher specimens of this isolateare deposited in the Herbarium of the Department ofBiology, University of Pisa, Herbarium Horti Botanici Pisani(PI), as PI-HMZ4. Sporocarps of G. mosseae were extractedfrom pot culture soil by wet sieving and decanting, through
a set of nested grids. Sporocarps retained on 100 mm sievewere flushed into Petri dishes and manually collected withforceps under a dissecting microscope (Leica, Milano,Italy).
Sporocarps to be used as inoculum for host plants werewashed by vortex in sterile distilled water (SDW) for 20 s,rinsed three times in SDW and germinated in the dark at24 C between two 47 mm cellulose nitrate MilliporeTM
membranes (0.45 mm diam pore) placed on moist sterilequartz grit in 14 cm diam Petri dishes. Fifteen sporocarpswere used for each membrane sandwich.
Sporocarps to be used for asymbiotic/symbiotic myceliumanastomosis tests were washed and rinsed in SDW asdescribed above and placed on a single 90 mm MilliporeTM
membrane on moist sterile quartz grit in 14 cm diam Petridishes sealed with Parafilm and incubated in the dark at24 C until germinating hyphae were observed.
Plant material.—Petroselinum hortense Hoffm. and Solanummelongena L. plants were obtained from sterile seedsgerminated in moist sterile grit, whereas MrS 2/5, aselection of Prunus cerasifera Ehrh., plants were obtainedfrom in vitro propagation. The root systems of both 20 d oldplantlets from seeds and micropropagated rooted plantswere washed carefully and individually sandwiched between47 mm MilliporeTM membranes containing germinatedsporocarps. Plants were placed into 10 cm diam pots, andthe sandwiched root systems were buried in sterile quartzgrit. Pots were closed in sun transparent bags (SigmaAldrich s.r.l., Milano, Italy) and maintained in a growthchamber with 24 C day and 21 C night temperature and 16/8 h light/dark cycle. Fourteen plants were prepared foreach species.
Experimental system.—After 42 d growth the roots weregently removed from the sandwich by immersion in waterand checked for the presence of extraradical hyphae, sporesand sporocarps, which were carefully plucked with forcepsunder a dissecting microscope. The occurrence of mycor-rhizal symbiosis was confirmed by autofluorescence ofintraradical fungal structures in fresh whole roots mountedin water and observed under a Reichert-Jung (Vienna,Austria) polyvar microscope equipped with epifluorescenceoptics (HBO 200 mercury lamp; Osram, Munich, Germany)with the B1 filter combination (BP 450–495, LP 520, DS510) (Vierheilig et al. 1999). The roots of each plant werearranged between two 90 mm MilliporeTM membranes onmoist sterile quartz grit in a 14 cm diam Petri dish, whichwas sealed with Parafilm. Ten replicate membranes wereprepared for each plant species. Petri dishes were main-tained in a growth chamber at 24 C day and 21 C night with16/8 h light/dark cycle, and at intervals the sandwicheswere carefully opened to check the development ofmycorrhizal extraradical mycelium. As soon as hyphaeappeared to spread from roots on the membranesgerminated sporocarps were placed nearby (about 5 mmapart, seven sporocarps per plant). Petri dishes were sealed,plants were incubated as previously described and harvestedwhen symbiotic and asymbiotic mycelia came into contact,after about 7–10 d, depending on the host plant species.
308 MYCOLOGIA

Occurrence and frequency of contacts between germlings andmycorrhizal networks.—Membranes bearing root systemsand mycelia were stained for histochemical localization ofsuccinate dehydrogenase (SDH) activity (Smith and Giani-nazzi-Pearson 1990). Deposition of formazan salts in hyphaeallowed the assessment of hyphal viability and protoplasmiccontinuity established between fusing hyphae. After SDHtest the membranes were stained with trypan blue in lacticacid (0.05%) to view mycelium under a stereomicroscope.Areas of contact between symbiotic and asymbiotic hyphaewere cut away from the larger membrane, mounted onmicroscope slides and examined under the polyvar micro-scope. Hyphal contacts were counted at magnifications of125–5003 and verified at 12503.
Classifying hyphal contacts. Hyphal contacts were classifiedin four categories. Perfect fusions were recorded whenhyphae anastomosed and protoplasm continuity could beverified by SDH staining. Prefusion incompatibility was scoredwhen incompatibility signs were observed before anastomosis.Hyphae underwent morphological changes indicative of thefirst step of anastomosis formation (swelling of hyphal tip,hyphal homing), but the protoplasm of one hypha retractedand septa were observed. Postfusion incompatibility wasscored when incompatibility signs were observed only afteranastomosis. Protoplasm withdrawal occurred in one or bothhyphae, and septa isolating fused hyphal compartments wereformed. Hyphal contacts were scored as non-interacting whenhyphae in close contact did not undergo any morphologicalchange and no fusions were observed.
Detection of nuclei in hyphal bridges between germlings andmycorrhizal networks.—To view the occurrence and locationof nuclei in anastomosed hyphae some membranes werestained with 5 mg/mL diamidinophenylindole (DAPI) in a1 : 1 (v/v) water : glycerol solution and observed underepifluorescence with the polyvar microscope using the filtercombination U1 (BP 330–380, LP 418, DS 420). Themicroscope was equipped with a 3 CCD color video cameraconnected to a personal computer with image acquiringsoftware (Pinnacle Studio 8). To view hyphal contacts somemembranes were stained with DAPI and mounted in a0.01% (w/v) solution of calcofluor white (Sigma Aldrichs.r.l.) (Sbrana et al. 1995) and observed under epifluores-cence with the filter combination U1.
Data analysis. The frequency of contacts in each categoryof hyphal contact (as reported in Classifying hyphal contacts)was calculated as a percentage of the total hyphal contactsobserved. Calculation of 99% confidence intervals wasperformed. Percentage data were subjected to arcsintransformation before carrying out two-way ANOVA analysisto assess the significance of the two factors involved (mycelialstage and host plant species) on each category of hyphalcontact and one way ANOVA to detect significant differencesbetween data obtained by different plant species or differentmycelial stages within each category of hyphal contact.
RESULTS
Occurrence of anastomoses between germlings andmycorrhizal networks.—Our experimental system al-
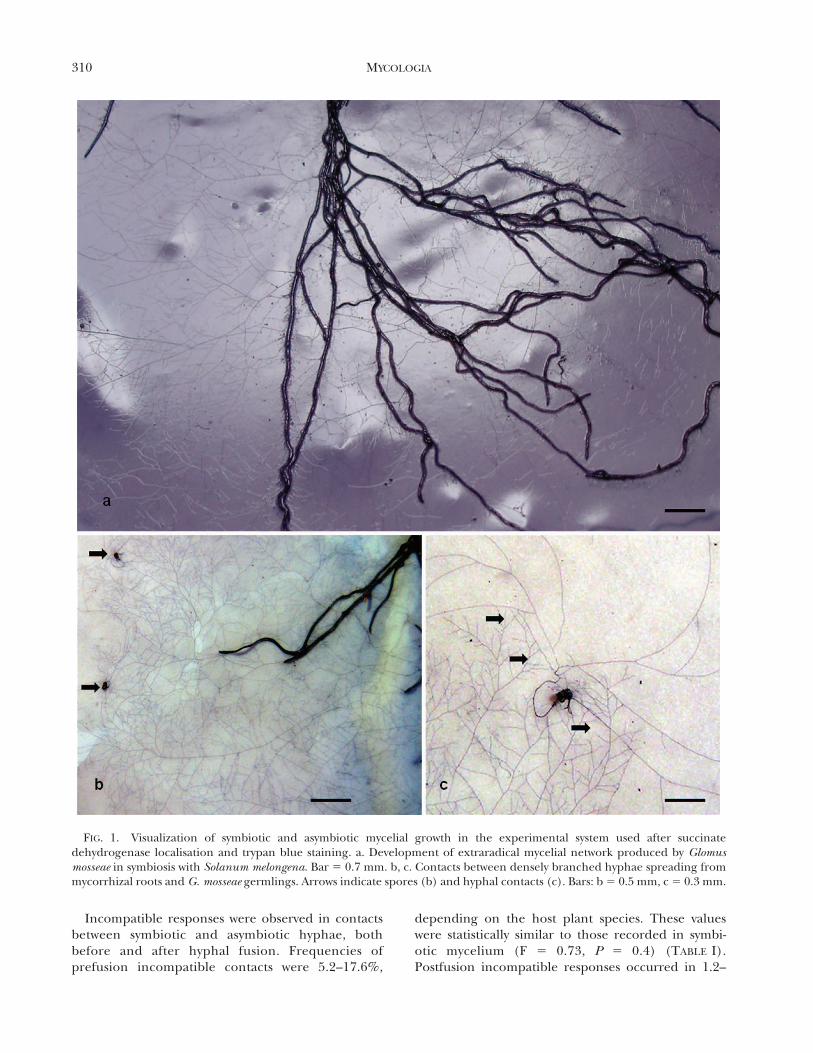
lowed the detection of anastomoses between intactextraradical hyphal networks originating from mycor-rhizal plant species and asymbiotic mycelium origi-nating from germinated spores. Viability of themycorrhizal network formed by host plants was nearly100% as determined by localization of SDH activity.Contacting hyphae were detected after SDH, trypanblue (FIG. 1a–c) and DAPI staining and completefusion of hyphal walls, protoplasm continuity andoccurrence of nuclei in the middle of hyphal bridgescharacterized perfect anastomoses (FIG. 2a, b). Toassess anastomoses, contacting hyphae observedunder the dissecting microscope were traced back tothe origin to check whether the contact was betweenhyphae originating from different sources (germlingand extraradical network) or from the same. Al-though many contacts resulted in no hyphal interac-tion (FIG. 2c), some hyphal contacts produced in-compatible interactions either before or after hyphalfusion. In prefusion incompatibility protoplasm with-drawal occurred in the approaching hypha aftercontact, whereas in postfusion incompatibility proto-plasm withdrawal and septa formation occurred inhyphae that underwent fusion (FIG. 2d, e). Whenincompatible fusions occurred the development of athin-walled intrahyphal hypha often was observed inthe recipient hypha (FIG. 2f).
Frequency of contacts between germlings andmycorrhizal networks.—Percentages of perfect anasto-moses occurring between germlings and extraradicalfungal networks spreading from S. melongena and P.hortense were respectively 23.9% (56 fusions, 234contacts) and 15.4% (14 fusions, 91 contacts). Lowerfrequencies were obtained when using P. cerasifera MrS2/5 as host plant with an average value of 4.9% (16fusions, 325 contacts) (TABLE I). One-way ANOVAshowed significant differences between perfect fusionfrequencies found for P. cerasifera and S. melongena (F5 5.31, P , 0.01). Two-way ANOVA showed thatfrequencies of perfect anastomosis occurring incontacts between germlings and extraradical fungalnetworks and those of anastomoses occurring withinextraradical networks were significantly different (F 5
46.87, P , 0.0001). Most hyphal contacts betweengermlings and extraradical fungal networks did notundergo anastomosis and were recorded as non-interacting. Frequencies were respectively 88.6%,57.1% and 46.6% for P. cerasifera MrS 2/5, P. hortenseand S. melongena (TABLE I) with significant differencesbetween P. cerasifera MrS 2/5 and the other hosts (F 5
8.54, P , 0.001). Two-way ANOVA results showedsignificant differences between such non-interactingcontact frequencies and those recorded within thesymbiotic network (F 5 8.78, P 5 0.005).
SBRANA ET AL.: MYCORRHIZAL NETWORKS 309
Incompatible responses were observed in contactsbetween symbiotic and asymbiotic hyphae, bothbefore and after hyphal fusion. Frequencies ofprefusion incompatible contacts were 5.2–17.6%,
depending on the host plant species. These valueswere statistically similar to those recorded in symbi-otic mycelium (F 5 0.73, P 5 0.4) (TABLE I).Postfusion incompatible responses occurred in 1.2–
FIG. 1. Visualization of symbiotic and asymbiotic mycelial growth in the experimental system used after succinatedehydrogenase localisation and trypan blue staining. a. Development of extraradical mycelial network produced by Glomusmosseae in symbiosis with Solanum melongena. Bar 5 0.7 mm. b, c. Contacts between densely branched hyphae spreading frommycorrhizal roots and G. mosseae germlings. Arrows indicate spores (b) and hyphal contacts (c). Bars: b 5 0.5 mm, c 5 0.3 mm.
310 MYCOLOGIA
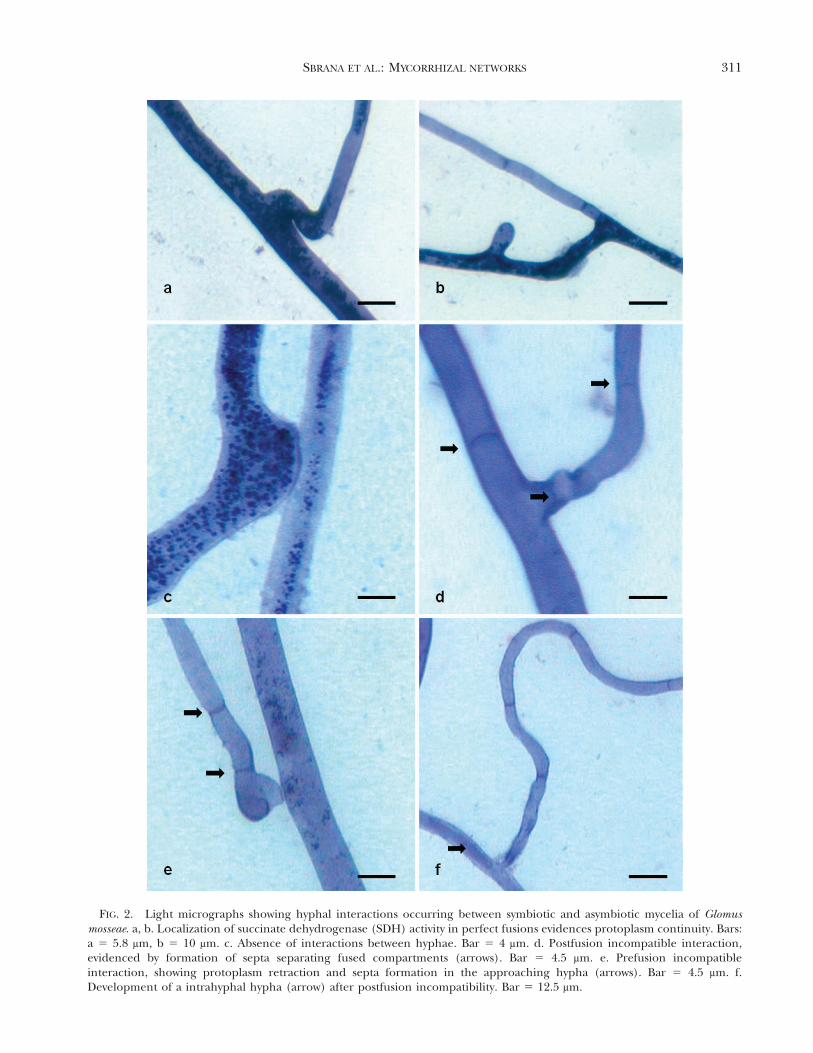
FIG. 2. Light micrographs showing hyphal interactions occurring between symbiotic and asymbiotic mycelia of Glomusmosseae. a, b. Localization of succinate dehydrogenase (SDH) activity in perfect fusions evidences protoplasm continuity. Bars:a 5 5.8 mm, b 5 10 mm. c. Absence of interactions between hyphae. Bar 5 4 mm. d. Postfusion incompatible interaction,evidenced by formation of septa separating fused compartments (arrows). Bar 5 4.5 mm. e. Prefusion incompatibleinteraction, showing protoplasm retraction and septa formation in the approaching hypha (arrows). Bar 5 4.5 mm. f.Development of a intrahyphal hypha (arrow) after postfusion incompatibility. Bar 5 12.5 mm.
SBRANA ET AL.: MYCORRHIZAL NETWORKS 311

14.9% total contacts between symbiotic and asymbio-tic mycelia. However this category of hyphal responsewas never observed in contacts between symbiotichyphae (TABLE I).
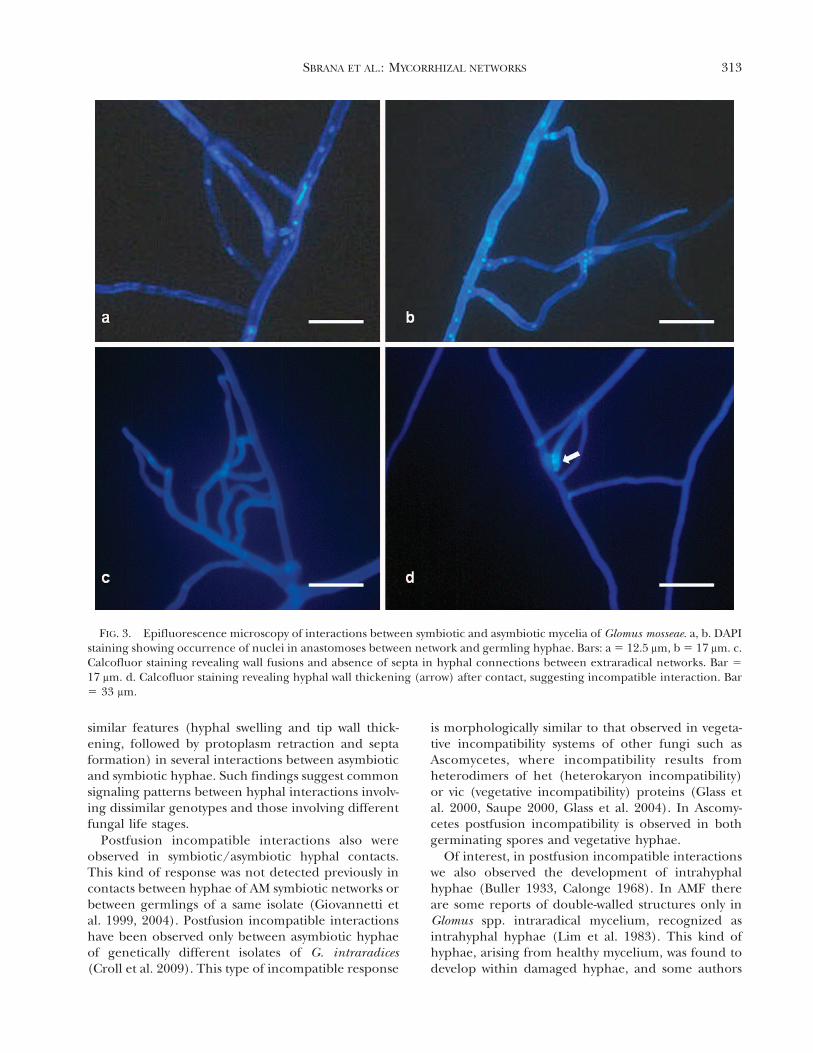
Nuclear occurrence in hyphal bridges between germlingsand mycorrhizal networks.—In perfect anastomosesDAPI staining and epifluorescence microscopyshowed that the process of hyphal anastomosisbetween germlings and extraradical hyphae involvedthe migration of nuclei, via the fusion bridge (FIG. 3a,b). In this type of hyphal contact calcofluor stainingfailed to detect signs of hyphal incompatibilityreactions, such as septa development or hyphal lysis,either before or after hyphal fusions (FIG. 3c). On theother hand, development of retraction septa and/ortip wall thickening were observed in postcontact andpostfusion incompatible responses (FIG. 3d).
DISCUSSION
Our results showed for the first time the occurrenceof hyphal fusions between AM networks spreadingfrom plant roots and germlings of the same AMfungal isolate. Such linkages were developed andviewed with a system in which plants and AMF aregrown in vivo. Our observations show that AMFgerminated spores are able to plug into a pre-existingmycorrhizal network. These findings provide anadditional explanation to the interesting ability ofAMF spores to germinate in the absence of hosts,despite being obligate biotrophs.
Our data suggest that anastomoses between germl-ings and AM network hyphae might represent adistinctive survival strategy because germlings canplug into the large AM networks exploring the soil,thus gaining immediate access to plant-derivedcarbon and overcoming growth arrest, enhancingtheir survival chances.
In this work we observed perfect anastomosesbetween germlings and extraradical hyphae of theAM fungus G. mosseae. SDH activity confirmed theestablishment of protoplasm connections, which arefundamental for the maintenance of physiologicalcontinuity (Schubert et al. 1987, Hamel et al. 1990,Giovannetti and Sbrana 2001). Perfect fusions alsowere confirmed by detection of nuclei in the middleof fusion bridges through DAPI staining and epi-fluorescence microscopy.
The number of hyphal contacts leading to anasto-mosis formation recorded between germling andnetwork hyphae was much lower than reportedbetween germlings of the same spores and ofdifferent spores of the same isolate (Giovannetti etal. 1999), between extraradical hyphae of the sameAM network (TABLE I; Giovannetti et al. 2001) orbetween contiguous AM networks interconnectingplants of different species, genera and families(Giovannetti et al. 2004). The low anastomosisfrequencies found between germling and networkhyphae compare well with those recently reportedbetween genetically different isolates of Glomusintraradices, where perfect fusions occurred in 1–10.3% of contacts (Croll et al. 2009).
Of note, a plant effect on anastomosis formationwas observed with larger numbers of non-interactingcontacts when germlings interacted with Prunuscerasifera MrS2/5 extraradical networks compared toS. melongena and P. hortense ones. A significant planteffect on extraradical mycelium density and anasto-mosis frequency was observed previously, suggestingthat nutritional or physiological characteristics ofplant partners might affect anastomosis within amycorrhizal network (Giovannetti et al. 2004).
Prefusion incompatibility was detected betweengermlings of geographically different isolates ofGlomus mosseae (Giovannetti et al. 2003). In this workthis phenomenon occurred with morphologically
TABLE I. Percentage frequencies of hyphal interactions between spore germlings (asymbiotic) and extraradical mycelium(symbiotic) of Glomus mosseae in symbiosis with different host plants and between hyphae of the same symbiotic mycelium
Between asymbiotic-symbiotic hyphae
Perfectfusions
Prefusionincompatibility
Postfusionincompatibility
No hyphalinteraction
Petroselinum hortense 15.4 (8.7–24.5)a 17.6 (10.4–26.9) 9.9 (4.6–17.9) 57.1 (46.3–67.5)Prunus cerasifera 4.9 (2.8–7.9) 5.2 (3.1–8.2) 1.2 (0.3–3.1) 88.6 (84.6–91.9)Solanum melongena 23.9 (18.6–29.9) 14.5 (10.3–19.7) 14.9 (10.6–20.2) 46.6 (40.1–53.2)
Between symbiotic hyphae
Petroselinum hortense 51.9 (42.9–60.9) 2.4 (0.5–6.7) 0b (0–2.9) 37.8 (29.3–46.8)Prunus cerasifera 50.5 (43.4–57.6) 8.9 (5.4–13.7) 0 (0–1.8) 40.6 (33.8–47.7)Solanum melongena 47.0 (35.9–58.3) 8.4 (3.5–16.6) 0 (0–4.3) 44.6 (33.7–55.9)
a Confidence limits for means (P 5 0.01).b Zero values obtained for postfusion incompatibility within symbiotic mycelium were removed from the dataset for ANOVA
because they have no variance.
312 MYCOLOGIA
similar features (hyphal swelling and tip wall thick-ening, followed by protoplasm retraction and septaformation) in several interactions between asymbioticand symbiotic hyphae. Such findings suggest commonsignaling patterns between hyphal interactions involv-ing dissimilar genotypes and those involving differentfungal life stages.
Postfusion incompatible interactions also wereobserved in symbiotic/asymbiotic hyphal contacts.This kind of response was not detected previously incontacts between hyphae of AM symbiotic networks orbetween germlings of a same isolate (Giovannetti etal. 1999, 2004). Postfusion incompatible interactionshave been observed only between asymbiotic hyphaeof genetically different isolates of G. intraradices(Croll et al. 2009). This type of incompatible response
is morphologically similar to that observed in vegeta-tive incompatibility systems of other fungi such asAscomycetes, where incompatibility results fromheterodimers of het (heterokaryon incompatibility)or vic (vegetative incompatibility) proteins (Glass etal. 2000, Saupe 2000, Glass et al. 2004). In Ascomy-cetes postfusion incompatibility is observed in bothgerminating spores and vegetative hyphae.
Of interest, in postfusion incompatible interactionswe also observed the development of intrahyphalhyphae (Buller 1933, Calonge 1968). In AMF thereare some reports of double-walled structures only inGlomus spp. intraradical mycelium, recognized asintrahyphal hyphae (Lim et al. 1983). This kind ofhyphae, arising from healthy mycelium, was found todevelop within damaged hyphae, and some authors
FIG. 3. Epifluorescence microscopy of interactions between symbiotic and asymbiotic mycelia of Glomus mosseae. a, b. DAPIstaining showing occurrence of nuclei in anastomoses between network and germling hyphae. Bars: a 5 12.5 mm, b 5 17 mm. c.Calcofluor staining revealing wall fusions and absence of septa in hyphal connections between extraradical networks. Bar 5
17 mm. d. Calcofluor staining revealing hyphal wall thickening (arrow) after contact, suggesting incompatible interaction. Bar5 33 mm.
SBRANA ET AL.: MYCORRHIZAL NETWORKS 313

suggested that nearby cross-wall septa might be thefocal point for differentiation (States 1975).
Our findings on hyphal incompatibility and on AMfungal differential behavior in anastomosis patternscould be explained by physiological differencesbetween germling hyphae and extraradical myceliarepresenting highly different developmental phasesof AMF. Large differences in gene expression,physiological characteristics and host recognitionability were observed between asymbiotic and symbi-otic AMF hyphae (Requena et al. 2000; Bago et al.2002; Requena et al. 2002, 2003; Breuninger andRequena 2004), suggesting that such diversity alsocould exist for hypha-hypha recognition patterns.Because the genetic determinants of hyphal fusionare still to be unraveled in AMF our findings suggestthat physiological differences between germlings andsymbiotic hyphae play an important role in determin-ing the outcome of fusion.
Co-existing sequence variants detected in singlespores, both for multicopy and single copy genes(Sanders et al. 1995, Lloyd-Macgilp et al. 1996,Helgason et al. 2003, Koch et al. 2004), stronglysuggested the multigenomic state of AMF (Bever andWang 2005, Hijri and Sanders 2005). In this work wedetected nuclei in fusion bridges between asymbioticand symbiotic hyphae, confirming previous data onnuclear migration during anastomosis, which couldrepresent a means for the maintenance of geneticdiversity in the absence of sexual recombination(Giovannetti et al. 2004, Bever and Wang 2005, Kuhnet al. 2001). Although we did not test whether suchfusions can occur between symbiotic and asymbiotichypha of genetically different AMF isolates, Croll et al.(2009) provided evidence suggesting that nuclei canbe exchanged between genetically different isolates ofAMF. Such data showed that the establishment ofviable protoplasmic connections allowed the migra-tion of nuclei through hyphal bridges, even in thepresence of low anastomosis frequencies.
In conclusion the ability of AMF germlings growingin the absence of the host to plug into a compatibleextraradical mycelium by means of viable, perfectanastomoses can let them access nutrient resourcesflowing in the mycorrhizal network and enhancesurvival chances.
ACKNOWLEDGMENTS
We thank two anonymous reviewers and the editor forhelpful and valuable suggestions. We are grateful for the helpof Dr Patrizia Strani who took part in preliminary experi-ments. PF was financially supported by doctoral funds of theUniversity of Pisa. This work was financially supported by aUniversity of Pisa grant (Fondi di Ateneo) and by CNR.
LITERATURE CITED
Akiyama K, Matsuzaki KI, Hayashi H. 2005. Plant sesquiter-penes induce hyphal branching in arbuscular mycor-rhizal fungi. Nature 435:824–827, doi:10.1038/nature03608
Avio L, Sbrana C, Giovannetti M. 1990. The response ofdifferent species of Lupinus to VAM endophytes.Symbiosis 9:321–323.
Bago B, Zipfel W, Williams RM, Jun J, Arreola R, LammersPJ, Pfeffer PE, Shachar Hill Y. 2002. Translocation andutilization of fungal storage lipid in the arbuscularmycorrhizal symbiosis. Plant Physiol 128:108–124,doi:10.1104/pp.010466
Becard G, Piche Y. 1989. Fungal growth stimulation by CO2
and root exudates in vesicular-arbuscular mycorrhizalsymbiosis. Appl Environ Microb 55:2320–2325.
Beilby JP, Kidby DK. 1980. Biochemistry of ungerminatedand germinated spores of the vesicular-arbuscularmycorrhizal fungus, Glomus caledonium: changes inneutral and polar lipids. J Lipid Res 21:739–750.
Bever JD, Wang M. 2005. Arbuscular mycorrhizal fungi—hyphal fusion and multigenomic structure. Nature 433:E3–E4, doi:10.1038/nature03294
Blair JE. 2009. Fungi. In: Hedges SB, Kumar S, eds. Thetimetree of life. Oxford, UK: Oxford Univ. Press. p 215–219.
Breuninger M, Requena N. 2004. Recognition events in AMsymbiosis: analysis of fungal gene expression at theearly appressorium stage. Fungal Genet Biol 41:794–804, doi:10.1016/j.fgb.2004.04.002
Buee M, Rossignol M, Jauneau A, Ranjeva R, Becard G.2000. The pre-symbiotic growth of arbuscular mycor-rhizal fungi is induced by a branching factor partiallypurified from plant root exudates. MPMI 13:693–698,doi:10.1094/MPMI.2000.13.6.693
Buller AHR. 1933. Researches on fungi. London: Long-mans, Green & Co. p 33–49.
Calonge FD. 1968. Origin and development of intrahyphalhyphae in Sclerotinia fructigena. Mycologia 60:932–942,doi:10.2307/3757395
Croll D, Giovannetti M, Koch AM, Sbrana C, Ehinger M,Lammers PJ, Sanders IR. 2009. Nonself vegetativefusion and genetic exchange in the arbuscular mycor-rhizal fungus Glomus intraradices. New Phytol 181:924–937, doi:10.1111/j.1469-8137.2008.02726.x
David Schwartz R, Badani H, Smadar W, Levy AA, Galili G,Kapulnik Y. 2001. Identification of a novel geneticallycontrolled step in mycorrhizal colonization: plantresistance to infection by fungal spores but notextraradical hyphae. Plant J 27:561–569, doi:10.1046/j.1365-313X.2001.01113.x
Gardes M, Wong KKY, Fortin JA. 1990. Interactions betweenmonokaryotic and dikaryotic isolates of Laccaria bicoloron roots of Pinus banksiana. Symbiosis 8:233–250.
Giovannetti M. 2001. Survival strategies in arbuscularmycorrhizal symbionts. In: Seckbach J, ed. Symbiosismechanisms and model systems. Dordrecht, the Neth-erlands: Academic Publisher. p 185–196.
———, Avio L, Sbrana C, Citernesi AS. 1993a. Factors
314 MYCOLOGIA

affecting appressorium development in the vesicular-arbuscular mycorrhizal fungus Glomus mosseae (Nicol.& Gerd.) Gerd. & Trappe. New Phytol 123:114–122.
———, Azzolini D, Citernesi AS. 1999. Anastomosisformation and nuclear and protoplasmic exchange inarbuscular mycorrhizal fungi. Appl Environ Microb 65:5571–5575.
———, Fortuna P, Citernesi AS, Morini S, Nuti MP. 2001.The occurrence of anastomosis formation and nuclearexchange in intact arbuscular mycorrhizal networks.New Phytol 151:717–724, doi:10.1046/j.0028-646x.2001.00216.x
———, Sbrana C. 2001. Self and non-self responses inhyphal tips of arbuscular mycorrhizal fungi. In:Geitmann A, Cresti M, eds. Cell biology of plant andfungal tip growth, NATO Science Series I: life andbehavioural sciences. Vol. 328. Amsterdam, the Nether-lands: IOS Press. p 221–231.
———, ———, Avio L, Citernesi AS, Logi C. 1993b.Differential hyphal morphogenesis in arbuscular my-corrhizal fungi during pre-infection stages. New Phytol125:587–594, doi:10.1111/j.1469-8137.1993.tb03907.x
———, ———, ———, Strani P. 2004. Patterns ofbelowground plant interconnections established bymeans of arbuscular mycorrhizal networks. New Phytol164:175–181, doi:10.1111/j.1469-8137.2004.01145.x
———, ———, Logi C. 1994. Early processes involved inhost recognition by arbuscular mycorrhizal fungi. NewPhytol 127:703–709, doi:10.1111/j.1469-8137.1994.tb02973.x
———, ———, Strani P, Agnolucci M, Rinaudo V, Avio L.2003. Genetic diversity of isolates of Glomus mosseaefrom different geographic areas detected by vegetativecompatibility testing and biochemical and molecularanalysis. Appl Environ Microb 69:616–624, doi:10.1128/AEM.69.1.616-624.2003
Glass NL, Jacobson DJ, Shiu PKT. 2000. The genetics ofhyphal fusion and vegetative incompatibility in fila-mentous ascomycete fungi. Annu Rev Genet 34:165–186, doi:10.1146/annurev.genet.34.1.165
———, Rasmussen C, Roca MG, Read ND. 2004. Hyphalhoming, fusion and mycelial interconnectedness.Trends Microbiol 12:135–141, doi:10.1016/j.tim.2004.01.007
Glenn MG, Chew FS, Williams PH. 1988. Influence ofglucosinolate content of Brassica (Cruciferae) roots ongrowth of vesicular-arbuscular mycorrhizal fungi. NewPhytol 110:217–225, doi:10.1111/j.1469-8137.1988.tb00255.x
Gollotte A, Gianinazzi-Pearson V, Giovannetti M, Sbrana C,Avio L, Gianinazzi S. 1993. Cellular localization andcytochemical probing of resistance reactions to arbus-cular mycorrhizal fungi in a ‘‘locus a’’ myc2 mutant ofPisum sativum L. Planta 191:112–122, doi:10.1007/BF00240902
Hamel C, Flyes H, Smith DL. 1990. Measurement ofdevelopment of endomycorrhizal mycelium using threedifferent vital stains. New Phytol 115:297–302,doi:10.1111/j.1469-8137.1990.tb00455.x
Harrison MJ, van Buuren ML. 1995. A phosphate transport-
er from the mycorrhizal fungus Glomus versiforme.Nature 378:626–629, doi:10.1038/378626a0
Helgason T, Watson IJ, Young JPW. 2003. Phylogeny of theGlomerales and Diversisporales (Fungi: Glomeromy-cota) from actin and elongation factor 1-alphasequences. FEMS Microbiol Lett 229:127–132,doi:10.1016/S0378-1097(03)00802-4
Hepper CM. 1983. Limited independent growth of avesicular-arbuscular mycorrhizal fungus in vitro. NewPhytol 93:537–542, doi:10.1111/j.1469-8137.1983.tb02704.x
Hijri M, Sanders IR. 2005. Low gene copy number showsthat arbuscular mycorrhizal fungi inherit geneticallydifferent nuclei. Nature 433:160–163, doi:10.1038/nature03069
Koch AM, Kuhn G, Fontanillas P, Fumagalli L, Goudet I,Sanders IR. 2004. High genetic variability and low localdiversity in a population of arbuscular mycorrhizalfungi. Proc Natl Acad Sci USA 101:2369–2374,doi:10.1073/pnas.0306441101
Koske RE. 1981. Gigaspora gigantea: observations on sporegermination of a VA- mycorrhizal fungus. Mycologia 73:288–300, doi:10.2307/3759650
Kuhn G, Hijri M, Sanders IR. 2001. Evidence for theevolution of multiple genomes in arbuscular mycorrhi-zal fungi. Nature 414:745–748, doi:10.1038/414745a
Kytoviita MM. 2000. Do symbiotic fungi refresh themselvesby incorporating their own or closely related sporesinto existing mycelium? Oikos 90: 606– 608,doi:10.1034/j.1600-0706.2000.900318.x
Lim LL, Fineran BA, Cole ALJ. 1983. Ultrastructure of intra-hyphal hyphae of Glomus fasciculatum (thaxter)Gerdemann & Trappe in roots of white clover(Trifolium repens L.). New Phytol 95:231–239,doi:10.1111/j.1469-8137.1983.tb03489.x
Lloyd-Macgilp SA, Chambers SM, Dodd JC, Fitter AH,Walker C, Young JPW. 1996. Diversity of the ribosomalinternal transcribed spacers within and among isolatesof Glomus mosseae and related mycorrhizal fungi. NewPhytol 133:103–111, doi:10.1111/j.1469-8137.1996.tb04346.x
Logi C, Sbrana C, Giovannetti M. 1998. Cellular eventsinvolved in survival of individual arbuscular mycorrhi-zal symbionts growing in the absence of the host. ApplEnviron Microb 64:3473–3479.
Morton JB. 1990. Evolutionary relationships among arbus-cular mycorrhizal fungi in the Endogonaceae. Mycolo-gia 82:192–207, doi:10.2307/3759848
Mosse B. 1959. The regular germination of resting sporesand some observations on the growth requirements ofan Endogone sp. causing vesicular-arbuscular mycor-rhiza. Trans Br Mycol Soc 42:273–286, doi:10.1016/S0007-1536(56)80033-8
Phipps CJ, Taylor TN. 1996. Mixed arbuscular mycorrhizaefrom the Triassic of Antarctica. Mycologia 88:707–714,doi:10.2307/3760964
Pirozynski KA. 1981. Interactions between fungi and plantsthrough the ages. Can J Bot 59:1824–1827.
———, Malloch DW. 1975. The origin of land plants: a
SBRANA ET AL.: MYCORRHIZAL NETWORKS 315

matter of mycotrophism. Bio Syst 6:153, doi:10.1016/0303-2647(75)90023-4
Redecker D, Kodner R, Graham LE. 2000. Glomalean fungifrom the Ordovician. Science 289:1920–1921,doi:10.1126/science.289.5486.1920
Remy W, Taylor TN, Hass H, Kerp H. 1994. Four hundredmillion-year-old vesicular arbuscular mycorrhizae. PNatl Acad Sci USA 91:11841–11843, doi:10.1073/pnas.91.25.11841
Requena N, Breuninger M, Franken P, Ocon A. 2003.Symbiotic status, phosphate and sucrose regulate theexpression of two plasma membrane H+ 2ATPase genesfrom the mycorrhizal fungus Glomus mosseae. PlantPhysiol 132:1540–1549, doi:10.1104/pp.102.019042
———, Mann P, Franken P. 2000. A homolog of the cellcycle check point TOR2 from Saccharomyces cerevisiaeexists in the arbuscular mycorrrhizal fungus Glomusmosseae. Protoplasma 212:89–98, doi:10.1007/BF01279350
———, ———, Hampp R, Franken P. 2002. Early develop-mentally regulated genes in the arbuscular mycorrhizalfungus Glomus mosseae: identification of GmGIN1, anovel gene with homology to the C-terminus ofmetazoan hedgehog proteins. Plant Soil 244:129–139,doi:10.1023/A:1020249932310
Sanders IR, Alt M, Groppe K, Boller T, Wiemken A. 1995.Identification of ribosomal DNA polymorphismsamong and within spores of the Glomales: applicationto studies on the genetic diversity of arbuscularmycorrhizal fungal communities. New Phytol 130:419–427, doi:10.1111/j.1469-8137.1995.tb01836.x
Saupe SJ. 2000. Molecular genetics of heterokaryonincompatibility in filamentous ascomycetes. MicrobiolMol Biol Rev 64:489–502, doi:10.1128/MMBR.64.3.489-502.2000
Sbrana C, Avio L, Giovannetti M. 1995. The occurrence ofcalcofluor and lectin binding polysaccharides in the
outer wall of AM fungal spores. Mycol Res 99:1249–1252, doi:10.1016/S0953-7562(09)80287-6
Schubert A, Marzachı C, Mazzitelli M, Cravero MC,Bonfante-Fasolo P. 1987. Development of total andviable extraradical mycelium in the vesicular-arbuscularmycorrhizal fungus Glomus clarum Nicol. & Schenck.New Phytol 107:183–190, doi:10.1111/j.1469-8137.1987.tb04892.x
Simon L, Bousquet J, Levesque RC, Lalonde M. 1993.Origin and diversification of endomycorrhizal fungiand coincidence with vascular land plants. Nature 363:67–69, doi:10.1038/363067a0
Smith SE, Gianinazzi-Pearson V. 1990. Phosphate uptakeand arbuscular activity in mycorrhizal Allium cepa L.:effects of photon irradiance and phosphate nutrition.Aust J Plant Physiol 17:177–188, doi:10.1071/PP9900177
———, Read DJ. 2008. Mycorrhizal simbiosis. London:Academic Press. 787 p.
States JS. 1975. Intrahyphal hyphae in the BasidiomyceteGloeophyllum (Lenzites) saepiarium. Mycologia 67:417–420, doi:10.2307/3758433
Tommerup IC. 1984. Persistence of infectivity by germinat-ed spores of vesicular-arbuscular mycorrhizal fungi insoil. Trans Br Mycol Soc 82:275–282, doi:10.1016/S0007-1536(84)80069-8
van der Heijden MGA, Klironomos JN, Ursic M, MoutoglisP, Streitwolf-Engel R, Boller T, Wiemken A, Sanders IR.1998. Mycorrhizal fungal diversity determines plantbiodiversity, ecosystem variability and productivity.Nature 396:69–72, doi:10.1038/23932
Vierheilig H, Bockenhoff A, Knoblauch M, Juge C, van BelA, Piche Y, Grundler FMW, Wyss U. 1999. In vivoobservations of the arbuscular mycorrhizal fungusGlomus mosseae in roots by confocal laser scanningmicroscopy. Mycol Res 103:311–314, doi:10.1017/S0953756298007576
316 MYCOLOGIA


























