Embed Size (px)
Citation preview

452
Mycologia, 96(3), 2004, pp. 452–462.q 2004 by The Mycological Society of America, Lawrence, KS 66044-8897
Differential morphogenesis of the extraradical mycelium of an arbuscularmycorrhizal fungus grown monoxenically on spatially heterogeneous
culture media
Bert Bago1
Centro de Investigaciones sobre Desertificacion (CSIC/UV/GV), Camı de la Marjal s/n, 46470 Albal(Valencia), Spain
Custodia CanoConcepcion Azcon-Aguilar
Departamento de Microbiologıa del Suelo y SistemasSimbioticos, Estacion Experimental del Zaidın (CSIC),calle Profesor Albareda 1, 18008 Granada, Spain
Julie SamsonAndrew P. CoughlanYves Piche
Centre de Recherche en Biologie Forestiere (CRBF),Pavillon Charles-Eugene-Marchand, Universite Laval,Quebec, G1K 7P4 Canada
Abstract: A new in vitro experimental system was de-veloped to study the morphogenesis of discrete re-gions of a single extraradical mycelium of the arbus-cular mycorrhizal (AM) fungus Glomus intraradices,growing simultaneously in six different agar-basedmedia. The media were (i) unamended water agar(WA), (ii) WA1PO4
32 (PO432), (iii) WA1NO3
2
(NO32), (iv) WA1NH4
1 (NH41), (v) WA1NH4
1
1MES (NH411MES) and (vi) minimal medium (M,
complete nutrients). Each medium was amendedwith the pH indicator bromocresol purple. The ex-traradical mycelium of the fungus showed between-treatment differences in morphogenesis, architec-ture, formation of branched absorbing structures(BAS) and sporulation. Extraradical hyphae that de-veloped in WA or PO4
32 compartments exhibited aneconomic development pattern, in which runner hy-phae radially extended the external colony. Extra-radical hyphal growth in the NO3
2 compartments wascharacterized by increased formation of runner hy-phae, BAS and spores and an alkalinization of themedium. In the two NH4
1-amended media (NH41,
NH411MES), sporulation was suppressed and consid-
erable morphological changes were noted. These re-sults show the plasticity of G. intraradices that lets itefficiently exploit an heterogeneous substrate.
Accepted for publication November 10, 2003.1 Corresponding author. E-mail: [email protected] address: Departamento de Microbiologıa del Suelo, Siste-mas Simbioticos, Estacion Experimental del Zaidın (CSIC), calleProfesor Albareda 1, 18008, Granada, Spain.
Key words: branched absorbing structures (BAS),Glomus intraradices, monoxenic culture, nitrogensource, pH, sporulation
INTRODUCTION
More than 80% of land plants are estimated to formarbuscular mycorrhizal (AM) associations (Smith andRead 1997) with soil-borne fungi in the phylumGlomeromycota (Schussler et al 2001). The wide en-vironmental tolerance of many taxa suggests that theyhave the capacity to adapt to different environmentalconditions.
Arbuscular mycorrhizal fungi are obligate bio-trophs, being unable to complete their life cycle inthe absence of a host plant (Azcon-Aguilar et al1998). After contact with a suitable host, inter- and/or intracellular colonization of the root cortex occurs(Bonfante-Fasolo 1984, 1987, Smith and Smith 1997).Establishment of the symbiosis allows the completionof the AM fungal life cycle. This involves formationof an extraradical hyphal phase that colonizes the soilin the vicinity of the host root. These hyphae formcharacteristic structures including branched absorb-ing structures (BAS, formerly named arbuscule-likestructures, ALS; Bago et al 1998b, c), spore-associatedBAS (BAS-s; Bago et al 1998b, c) and spores. Theextraradical mycelial network increases the nutrientuptake surface of the host plant and allows a moreefficient extraction of phosphorus, nitrogen and cer-tain micronutrients (Smith and Read 1997).
Several studies have highlighted the metabolic ca-pacities of the AM extraradical phase (reviewed byJakobsen 1995; see also Bago et al 1996, 2002, 2003,Olsson et al 2002). However, little is known about theeffect of the heterogeneous soil environment on theextraradical mycelium (ERM) and its subsequent ef-fects on nutrient uptake by AM fungi. Friese and Al-len (1991) were the first to study extraradical hyphalarchitecture in rhizospheric soil. However recentstudies by Bago et al (1998a, b, c) using AM mono-xenic cultures (in vitro dual AM fungus and root-organ cultures) indicated that the AM extraradicalmycelium is more complex than was reported byFriese and Allen (1991). Cui and Caldwell (1996a, b)showed that extraradical AM hyphae are equally ef-ficient in transporting soil phosphate and nitrate, ir-respective of the patchiness of the substrate. There-

453BAGO ET AL: DIFFERENTIAL MORPHOGENESIS OF AMF
fore, we hypothesize that hyphal development andcolony architecture of AM fungi vary on a microscaleto maximize growth and nutrient uptake within het-erogeneous substrates. Such responses have beenshown to occur with other soil-borne fungi such asectomycorrhizal (Bending and Read 1995) and sap-rophytic fungi (Bailey et al 2000, Fomina et al 2003,Ritz 1995). In particular, the work by Bailey et al(2000) indicates that fungal colony development ina given substrate depends mainly on nutrient distri-bution and availability but that it also is influencedby the intrinsic morphogenesis of the fungal colony,which includes the radial density of hyphal growth,aggregation into strands and the degree of branchingand anastomosing. Our hypothesis also is supportedby a recent study that shows the differential growingstrategy of various AM fungi when colonizing and ex-ploiting a given substrate (Smith et al 2000). The aimof the present work was to study morphological anddevelopmental changes to the architecture of the ex-traradical mycelium of Glomus intraradices Smith andSchenk subjected to different nutritional conditionsin a patchy environment.
MATERIALS AND METHODS
Experimental system and biological material.—Under sterileconditions the top of a 60 mm Petri plate was placed in themiddle of a 150 mm Petri plate and six tops from 35 mmPetri plates were arranged around it (FIG. 1a). The central(60 mm) plate was filled with 20 mL of sterilized (121 C,20 min) minimal medium (Chabot et al 1992). The nutri-ent content of this medium (in mg/L of distilled water andmolecular amounts) was: MgSO4·7H2O, 731 (29.6 mM);KNO3, 80 (7.9 mM); Ca(NO3)2·4H2O, 288 (12.2 mM) (totalN content 5 45.23 mg/L); KCl, 65 (8.71 mM); KH2PO4, 4.8(1.37 mM); Na-FeEDTA, 8 (4.36mM); KI, 0.75 (4.51 mM);MnCl2·4H2O, 6 (30.3 mM); ZnSO4·7H2O, 2.65 (9.2 mM);H3BO3, 1.5 (24.3 mM); CuSO4·5H2O, 0.13 (0.52 mM);Na2MoO4·2H2O, 0.0024 (9.9 mM); glycine, 3 (3.99 mM); thi-amine, 0.1 (29.7 mM); pyridoxine, 0.1 (48.6 mM); nicotinicacid, 0.5 (0.4 mM); myoinositol, 50 (27.7 mM); and sucrose,10 g/L (29.2 mM). The central plate, referred to as theculture compartment (FIG. 1a, CC), was used to establishthe mycorrhizal symbiosis. This consisted of a monoxenicculture (Becard and Fortin 1988) of the AM fungus Glomusintraradices Schenck & Smith (DAOM 197198, Biosystemat-ic Research Centre, Ottawa, Canada), grown with a carrot(Daucus carota L.) root-organ culture (clone DC-2). Mono-xenic cultures were started by placing a 5 cm-long root inthe culture compartment. A cube of medium from a pre-vious monoxenic culture, containing approximately 100spores, fragments of mycorrhizal roots and external hyphae,was placed near the root apex.
The six 35 mm plates, referred to as the hyphal com-partments (FIG. 1a, HC), were filled with 7 mL of one ofsix different sterilized (121 C, 20 min) culture media. These
media consisted of 0.8% Bacto-Difco agar in distilled watereither (i) unamended (water agar [WA]) or amended with(ii) 4.8 mg/L KH2PO4, (iii) 326 mg/L KNO3 (45.23 mg/Ltotal nitrogen), (iv) 214 mg/L (NH4)2SO4 (45.23 mg/L to-tal nitrogen), (v) 214 mg/L (NH4)2SO4 in a MES solution(10 mM, pH 5 6) and (vi) nutrients to provide the samecomposition of the minimal medium. All six hyphal com-partment media were amended (1%) with a solution (1%in distilled, sterile water; Bago et al 1996) of the pH indi-cator bromocresol purple to follow pH changes during thestudy (Bago et al 1996). After autoclaving, pH values of thedifferent culture media were: minimal medium, 5.7; PO4
32,6.0; NO3
2, 6.0; NH41, 6.2; NH4
11MES, 5.8; and WA, 6.2.Samples (n 5 5) of the six media in which neither rootsnor the fungus were cultured (referred to as color controls)were maintained under identical conditions as the cultures.
The space between the six hyphal compartments and theculture compartment was filled with bromocresol purple-free WA (pH 6.0) (FIG. 1a, Interm), so that hyphae thatgrew out of the culture compartment could develop in WAmedium and then into the different test media. Ten repli-cates were set up and the plates incubated in the dark at25 C for 84 d. Roots periodically were cut to prevent themdeveloping in the surrounding WA media or HC.
Observation and measurements of fungal growth.—At the endof the experiment, mean total runner hyphal length wascalculated using a 2 3 2 mm grid (Marsh 1971) and thetotal numbers of BAS and spores were determined. All mea-surements were done under a Nikon AFX stereomicro-scope. Microphotographs (Kodak 100 ISO film) of hyphalmorphology and extraradical structures in the different cul-ture media were taken with a Leica DMRB microscope fit-ted with a Leica MPS-60. Results were analyzed statisticallyusing the Fisher’s Protected LSD test (P # 0.05).
pH measurements.—Nondestructive pH measurements ofthe HCs were made according to Bago et al (1996). Absor-bance of the culture media contained in each HC was mea-sured (Shimadzu UV-visible spectrophotometer, UV 1603),without removing them from the Petri plates, at two wave-lengths, 589 and 429 nm. The ratio between the two mea-sured absorbances (i.e., A589/A429) was obtained, and thepH of the medium, y, was calculated using
A589y 5 2.24·log 1 5.661 2A429
RESULTS
Culture development and pH changes.—All six brom-ocresol purple amended culture media initially hada similar pH (approximately 6, see Methods) andwere reddish-orange. However, after 7 d of incuba-tion the color changed to yellow in the NH4
1 HC,both in the color controls (plates that contained noroot or fungus) and in plates that contained rootsand AM fungus, indicating a strong acidification ofthe medium in these HC. Because this drop in pHtook place before fungal development in the HC, the

454 MYCOLOGIA
TABLE I. Estimate of extraradical mycelium development in four representative Petri plates after 42 d of culture
Rootcompartment
Interm.agar
Hyphal compartments
M PO432 NO32 NH41 NH41 1 MES WA
Plate 1DevelopmentpH
**ii **** Absiii
5.5Abs5.5
Abs/Esp6.9
Sparse4.5
Abs. Interconn.6.0
Abs5.5
Plate 2DevelopmentpH
** **** Abs6.4
Abs5.5
Abs/Esp6.5
Sparse4.5
Abs. Interconn.6.1
Abs5.5
Plate 3DevelopmentpH
* *** Diffuse6.0
Abs6.0
Abs/Esp6.5
Sparse4.6
Abs. Interconn.6.0
Abs5.5
Plate 4DevelopmentpH
**/*** **** Diffuse6.0
Abs5.5
Abs6.7
Sparse4.6
Abs. Interconn.5.9
Abs5.5
i Hyphal compartments containing the different media tested.ii Estimates were quantified based on a subjective scale ranging from one asterisk (approximately 200 spores per cm2 of
medium), and five asterisks (approximately 1000 spore/cm2 of medium).iii Abs, absorptive phase; Abs/Esp, transition between absorptive to sporulative phases; Sparse, few runner hyphae with
atrophied BAS; Abs. Interconn., profusely branched mycelium with interconnected hyphae; Diffuse, tangled, undulatedhyphae with less compact BAS.
TABLE II. Runner hyphal length, structures/cm of runner hyphae and pH of the media in the different hyphal compart-ments of the experimental system after 84 d of culture
Runner hyphalength (cm/cm2) BAS/cm2 Spore/cm2
pH
Color controls 1 AMFi
MPO4
32
NO32
NH41
NH41 1 MESWA
68.3 aii
56.4 a125.3 b33.7 a
143.9 b64.0 a
254.9 ab254.1 ab636.0 c72.0 a—iv
288.0 b
27.8 a13.6 a
368.0 b0.0 a0.0 a
61.3 a
5.9 Aiii
5.7 A5.9 A5.3 A5.9 A5.7 A
6.4 B d5.6 A b6.9 B e4.9 B a6.0 A c5.4 A b
i Hyphal compartments in which the extraradical mycelium (ERM) of fungus had developed (n 5 10).ii Different letters correspond to statistical differences between means (Fisher’s Protected LSD test, (P # 0.05).iii Different capital letters correspond to significant pH differences between media sustaining ERM growth compared to
their respective color controls.iv Extraradical mycelium was too profusely branched for BAS to be properly quantified.
extraradical mycelium developing in NH41 treat-
ments initially faced acidic conditions. By contrast, incolor control plates of NH4
11MES, color remainedunchanged.
After 28 d, extraradical hyphae extensively had col-onized the 10 culture compartments. Approximately14 d later, extraradical hyphae crossed over the plas-tic boundary of the culture compartments and de-veloped in the external, bromocresol-free WA media(TABLE I). Extraradical hyphae also began to developin the different HC. We observed an alkalinization(color change to purplish-violet) in the NO3
2 HC col-onized by extraradical hyphae. A similar but weaker
color change occurred in the minimal medium(which also contained NO3
2 as the sole nitrogensource). No detectable color (pH) changes occurredin compartments containing PO4
32 or WA after col-onization by extraradical hyphae or in those contain-ing the buffered NH4
1 medium (NH411MES). Un-
buffered ammonium-amended compartments(NH4
1) remained yellow (acidic) throughout the ex-periment.
After 84 d, spectrophotometric pH measurementsof the HC confirmed the above observations (TABLE
II). Results showed that hyphae developing in com-partments containing NO3
2 raised the pH of the me-

455BAGO ET AL: DIFFERENTIAL MORPHOGENESIS OF AMF
dium by 0.5 (minimal medium) and 1.0 (NO32) com-
pared to their respective color control plates. Theyellow observed in NH4
1 HC, where some extra-radical hyphae were developing, corresponded to apH decrease of 0.4 when compared to the color con-trol plates but of 1.3 pH units when compared to theinitial pH of the medium (before the incubation pe-riod, see Methods). The pH of the rest of the HCdid not differ by more than 0.1 from the pH of theirrespective color control plates, except for WA com-partments, which showed a decrease of 0.3.
Fungal development and hyphal morphogenesis.—Afterthe 84 d culture period, extraradical hyphae had de-veloped extensively in the bromocresol-free WA sur-rounding the HC and this WA medium contained ahigh number of BAS and spores (FIG. 1b). This con-trasted with the reduced fungal development in theHC, particularly in the NH4
1 treatments (see below).When developing in WA (FIG. 1c, d), PO4
32 (FIG.1e, f) or NO3
2 (FIG. 2e, f), the extraradical myceliumwas highly organized. The mycelium consisted ofthick (10–15 mm diam), leading runner (Friese andAllen 1991, Bago et al 1998b) hyphae that branchedat approximately 458 (FIG. 1c, e) to produce thinner(1–7 mm diam), higher order, runner hyphae, re-sulting in a radial extension of the mycelium (FIG.1e). Under this developmental pattern, runner hy-phae showed apical dominance and constituted theframework of the colony. Branched absorbing struc-tures (BAS) and spore-associated BAS (BAS-s; Bagoet al 1998b, c) developed at relatively regular inter-vals on second and higher order runner hyphae(FIGS. 1d, f, 2f). Spores also were produced common-ly by BAS-s (Bago et al 1998b) along runner hyphae(FIG. 2c–f).
In contrast to this well-organized developmentalpattern, on initial contact with the minimal medium,extraradical hyphae showed a more diffuse growthpattern (FIG. 2a–c). Apical dominance was lost, andthe mycelium frequently appeared tangled or inter-woven (FIG. 2a, b, arrows; FIG. 2c). Leading runnerhyphae were barely distinguishable (FIG. 2a, b) andBAS were less compact, merging within the profuselybranched fungal colony. However, after 1–2 wk apicaldominance recovered (FIG. 2d). Highly branched,leading runner hyphae re-appeared, and BAS formedat regular intervals (FIG. 2d, arrows).
Mycelial architecture was altered markedly whenthe fungus developed in the acidic NH4
1 medium(FIG. 3a–d). Leading runner hyphae still branched atapproximately 458 (FIG. 3a), but branches were thick-er (approximately 10–12 mm diam versus 5–7 mm inother treatments, see above) and colony architectureappeared disrupted. Cytoplasmic protrusions oc-
curred at different sites, especially at apices, whichfrequently appeared lysed (FIG. 3b, c, arrows). Run-ner hyphae occasionally formed short ramificationsat irregular intervals (possibly atrophied BAS; FIG.3b, white arrows). Coils of hyphae, which consistedof closely aligned, cytoplasm-filled hyphae, were ob-served in some of the replicates (FIG. 3c, d). In theMES-amended compartments, morphological chang-es also were evident but were different than thoseobserved in NH4
1 treatment (FIG. 3e, h); upon con-tact with the NH4
11MES medium, the leading run-ner hyphae lost apical dominance (FIG. 3e, arrow)and branched so profusely that it was impossible tofollow its growing pattern (FIG. 3h). Extraradical hy-phae became much thinner and interwoven (FIG.3h) and growth was restricted to the medium closestto the boundary of the HC, while the center of thecompartment remained almost uncolonized.
BAS that formed in compartments containing min-imal medium (FIG. 2d) were 1.5–2 times larger thanin the other treatments and less compact than thosethat developed in WA (FIG. 1d), PO4
32 (FIG. 1f) andNO3
2 (FIG. 2f) compartments. In NH411MES treat-
ments, BAS were indistinguishable due to the profusebranching of the fungus (FIG. 3h; see TABLE II, foot-note iv), and as outlined above in NH4
1 compart-ments BAS were rare (TABLE II) and atrophied (FIG.3b). The formation of BAS was stimulated most inNO3
2, whereas WA, minimum medium and PO432
treatments showed an intermediate degree of BASformation (TABLE II).
Sporulation was increased significantly in the NO32
treatment (TABLE II, FIG. 2e). This effect also wasobserved in preliminary tests using similar experi-mental conditions but with monoxenic cultures of G.intraradices on untransformed tomato (Lycopersiconesculentum Mill.) root organs. Sporulation was lowerin WA compartments, further reduced in minimalmedium and PO4
32 and suppressed in NH41 com-
partments (either buffered or unbuffered, TABLE II).Although variable in all the media tested, spore sizewas particularly heterogeneous in minimal medium(FIG. 2c; 84% of them ranged from 15 to 40 mm,14%, from 40 to 65 mm). However, spores lookedviable and their lipid contents appeared similar ir-respective of size. In all replicates, G. intraradicespreferentially sporulated (1053 6 330 spore/cm2) inthe bromocresol-free WA medium surrounding theHC (FIGS. 1b, c, 3e). The morphological features de-scribed throughout were consistent in the 10 repli-cate plates.
DISCUSSION
The evolution of a mycelial habit in fungi let themdevelop diverse feeding strategies and exploit a wide
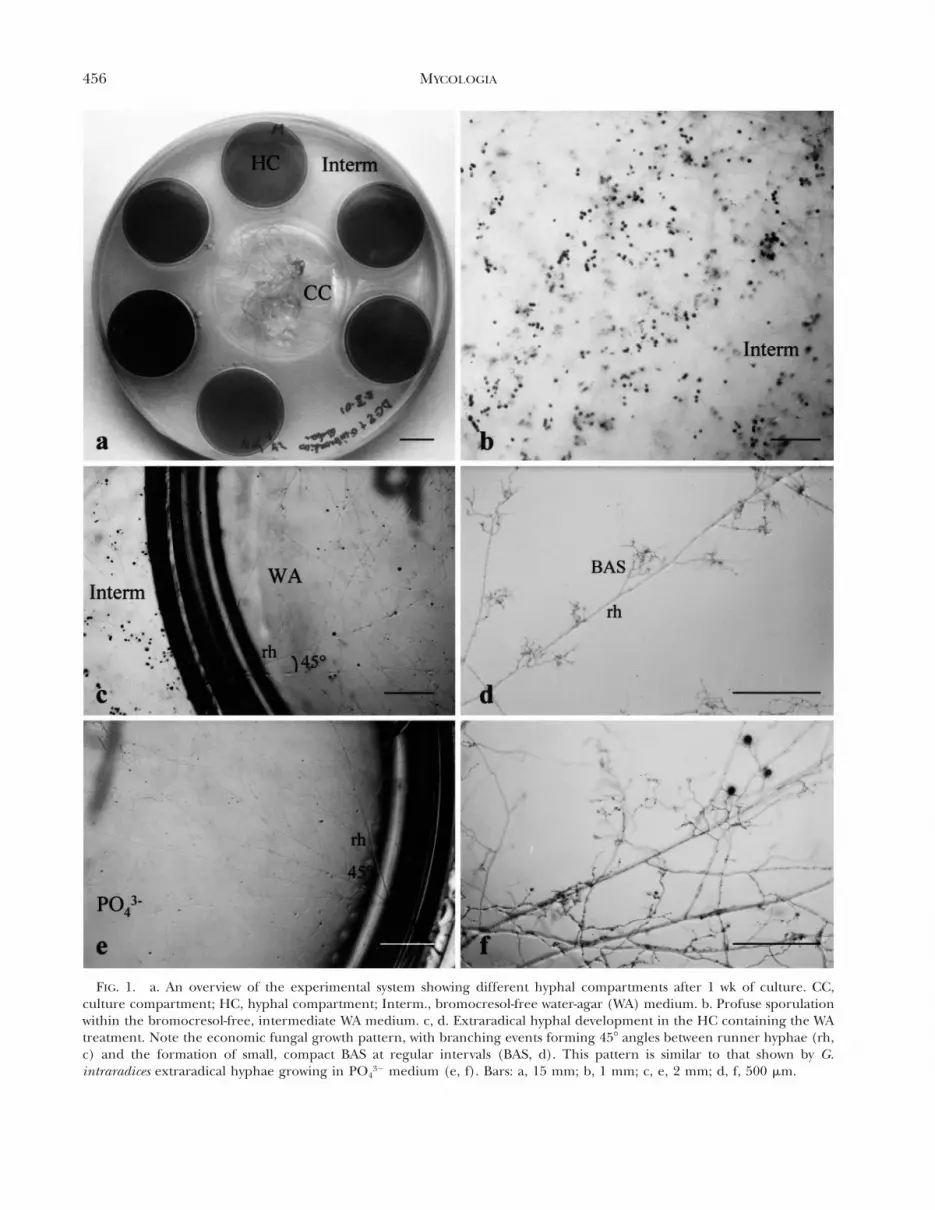
456 MYCOLOGIA
FIG. 1. a. An overview of the experimental system showing different hyphal compartments after 1 wk of culture. CC,culture compartment; HC, hyphal compartment; Interm., bromocresol-free water-agar (WA) medium. b. Profuse sporulationwithin the bromocresol-free, intermediate WA medium. c, d. Extraradical hyphal development in the HC containing the WAtreatment. Note the economic fungal growth pattern, with branching events forming 458 angles between runner hyphae (rh,c) and the formation of small, compact BAS at regular intervals (BAS, d). This pattern is similar to that shown by G.intraradices extraradical hyphae growing in PO4
32 medium (e, f). Bars: a, 15 mm; b, 1 mm; c, e, 2 mm; d, f, 500 mm.

457BAGO ET AL: DIFFERENTIAL MORPHOGENESIS OF AMF
FIG. 2. Morphological features of G. intraradices external mycelium developing in minimal medium (a–d) and NO32 (e,
f) media. a. Extraradical hyphae growing in minimal medium show a diffuse growth pattern (arrow). On first contact withthe minimal medium (b) extraradical hyphae lose apical dominance (arrow) and hyphae frequently appear undulate andtangled (c). However, with time recovery of apical dominance is noted (d) and large BAS are formed (arrows). In contrastto this, when developing in NO3
2 medium runner hyphae show a clear apical dominance and profuse sporulation (e). Undersuch nutrient conditions small, compact BAS, BAS-s and spores are formed (f) by the runner hyphae at regular intervals.Bars: a, e, 3 mm; b, d, 1 mm; c, 500 mm; f, 250 mm.

458 MYCOLOGIA
FIG. 3. Morphological features of G. intraradices external mycelium developing in NH41 (a–d) and NH4
11MES (e, h)media. When developing in NH4
1 hyphal compartments extraradical hyphae showed a certain apical dominance on firstcontact with the culture medium and branching at 458 angles was observed (a). However, the fungus soon lost apicaldominance and formed coarse hyphae (b) with frequent cytoplasmic protrusions, especially at the apices (black arrows).Rarely, atrophied BAS were observed (white arrows) (c). Coils of hyphae, which frequently bound together, were formedoccasionally (d). On initial contact with the NH4
11MES medium G. intraradices external hyphae also lost apical dominance(e, arrow), however, they branched profusely. Hyphae became thin and tangled (h) and BAS were impossible to distinguishand quantify. Bars: a, f, g, 3 mm; b, c, 100 mm; d, 50 mm; e, 5 mm; h, 500 mm.
range of substrates (Kendrick 1992). Indeed, fungiexhibit marked phenotypic plasticity, quickly adapt-ing to changing conditions, and their hyphae are par-ticularly well suited to heterogeneous environments.
In particular, arbuscular mycorrhizal (AM) fungi ex-hibit two distinct mycelial phases: an intraradicalphase (intraradical mycelium, IRM), developing un-der relatively homogeneous conditions in the root

459BAGO ET AL: DIFFERENTIAL MORPHOGENESIS OF AMF
cortex (Smith and Read 1997), and an extraradicalphase (extraradical mycelium, ERM), developing un-der temporally and spatially heterogeneous condi-tions in the soil. Intraradical hyphae alter their mor-phology, physiology and probably their genetic ex-pression to meet the constraints imposed by host rootcells (Smith and Smith 1997). This prevents rejectionof the association and allows establishment of a fullyfunctional symbiosis. The extraradical mycelium ofAM fungi also might be expected to adapt its growthpattern and metabolic capabilities to explore and ex-ploit different soil microenvironments (Bago 2000).
The extent of spread of the extraradical AM hy-phae depends on the fungal species and environ-mental conditions (van Bruggen et al 2000, Smith etal 2000) ranging from soil phosphorus concentration(Abbott et al 1984, Abbott and Robson 1985) to at-mospheric CO2 (Klironomos et al 1998). Cui andCaldwell (1996a, b) demonstrated that the AM fungalERM facilitates the acquisition of phosphate by plantsgrowing in enriched soil patches. The results pre-sented here demonstrate that different zones of thesame AM ERM can develop differently to exploreand exploit the surrounding substrate. These mor-phogenetic changes are localized (and sometimestransient) within the fungal colony and are influ-enced directly by the presence of a nutrient (or com-bination of nutrients). It is interesting to note thatthe growth of some hyphae into one hyphal com-partment (HC) did not influence hyphal morpho-genesis of the rest of the colony. The plasticity of theERM might be an important strategy for adaptationand survival in a diverse range of ecosystems andwithin heterogeneous substrates. Although the ex-perimental system used (monoxenic AM cultures) issomewhat artificial, our results, as well as those ofother previously published studies that also usedmonoxenic cultures, seem to reflect accurately themorphogenetic processes known to occur in AM fun-gi when growing in soil.
When growing with little (NO32, PO4
32) or no(WA) nutrients, extraradical AM hyphae exhibited awell organized and economic development pattern,probably designed to maximize exploration and ex-ploitation of the medium and allow the productionof spores outside the zone of influence of the colo-nized root. This pattern, similar to that described byBago et al (1998a, b), is based on the formation ofrunner hyphae, which radially extend the fungal col-ony, and from which small, compact branched ab-sorbing structures (BAS) develop at regular intervals.These latter structures have been proposed as pref-erential nutrient uptake structures (Bago et al1998c). Thus, while runner hyphae rapidly extendthe colony to find either zones with more nutrition
within the substrate or a new host root to colonize,BAS immediately form to absorb nutrients and sporesdevelop in older parts of the fungal colony that prob-ably have switched from assimilative to reproductivemetabolism. The spore contents are mainly carbo-naceous compounds (stored as lipids and glycogen;Beilby and Kidby 1980, Jabaji-Hare 1988, Bonfante etal 1994), which are acquired by the intraradical hy-phae from the host plant (Shachar-Hill et al 1995)and transported toward the extraradical mycelium(Pfeffer et al 1999, Bago et al 2002, 2003). However,spore formation also requires other nutrients, one ofthe most important being nitrogen, a principal com-ponent of chitin, which is abundant in the spore wall.The idea that nitrogen is taken up by the fungus(probably via BAS) from the culture medium duringassimilative growth is supported by the enhancedBAS production observed on extraradical hyphaegrowing in NO3
2-amended media. A further indica-tion of NO3
2 uptake by the fungus in the HC comesfrom the observed alkalinization of the NO3
2-con-taining media (NO3
2, minimal medium), which alsowas observed by Bago et al (1996). The absence ofpH changes in PO4
32 medium suggests that othermechanisms apart from symport/antiport were activeon the fungal hyphae in this treatment (e.g., organicacid excretion), which could balance the otherwiseexpected alkalinization of the culture medium.
Fungal sporulation can be enhanced by increasingthe C:N ratio of the culture media (Delatorre andCardenascota 1996, Engelkes et al 1997, Pascual et al1997, Yu et al 1998). After 84 d, the presence of anadequate supply of carbohydrates (exported from theintraradical mycelium) combined with a possible de-pletion of media nitrogen (through fungal NO3
2 up-take, Bago et al 1996) could explain the observedincrease in spore production in the NO3
2 HC. Al-though NO3
2 is the sole nitrogen source in minimalmedium HC, this culture medium also contains su-crose; such an exogenous carbon source, which theextraradical fungal structures are unable to acquireand metabolize (Pfeffer et al 1999, Bago et al 2002),seems to hinder spore production by G. intraradicesERM, as has been shown by St.-Arnaud et al (1996).
When a balanced mix of mineral nutrients is avail-able in the medium (e.g., in minimal medium HC)uptake becomes the principal function of the myce-lium, rather than exploration or reproduction. Inten-sive substrate exploitation probably was achieved byincreased hyphal branching and a temporary loss ofapical dominance by the runner hyphae in the min-imal medium treatment. The fact that hyphae werein an assimilative rather than in a reproductive phasecould explain the low spore number in this treat-ment. When growing in batch culture, fungal growth

460 MYCOLOGIA
is unrestricted until nutrient depletion or alterationof other culture conditions inhibit it (Trinci et al1994). It is interesting to note that after a certaintime the morphogenesis of some hyphae in the min-imal medium compartments changed to resemblethose growing in low-nutrient media. This probablyis explained by nutrient depletion of the minimalmedium.
Although initially adjusted to 6.2, the pH of NH41
HC fell sharply before fungal development withinthem. This decline probably was caused by the for-mation of carbonic acid (CO2 1 H2O → HCO3
2 1H1) in the NH4
1-amended media after a build up ofCO2 produced by the roots and hyphae in the culturecompartment. Although production of this weak acidwas not enough to reduce the pH in the other cul-ture media, the fact that an (NH4)2SO4 solution inwater trends to be acidic (as observed in the NH4
1
color controls, which spontaneously drop almost 1pH unit; TABLE II) seemed to be enough to shift thealready weak chemical equilibrium of the medium.
Development of external hyphae of G. intraradicesin NH4
1 HC reflects the adaptive changes of this fun-gus when growing under adverse conditions. Chang-es in extraradical fungal morphology (e.g., infre-quent and deformed BAS, formation of coiled hy-phae and protruded apices and a suppression ofsporulation) were observed when NH4
1 was the onlynitrogen form in the culture medium under acidicconditions. AM colonization has been reported inplants growing in mine spoils at pH 2.7 (Daft et al1975, see also Heijne et al 1996). Using monoxeniccultures of G. intraradices Coughlan (1998) foundthat external hyphae stopped growing immediatelyon reaching HC containing WA medium adjusted topH 4. Clark and Zeto (1996) reported a sharp re-duction in intraradical arbuscules or vesicles in Glo-mus-colonized maize roots when plants were grownin acidic soil, results supported by Tiwari et al (2002)using AM monoxenic cultures. Finally, van Aarle etal (2002) have shown that low pH negatively influ-enced both extraradical hyphal spread and (possibly)spore formation in two AM fungi grown in soil, eventhough the roots of the host plants were not exposedto the same pH. These reports agree with the resultsobtained in our work. However, it remains unclearwhether the morphological changes presented herewere caused by low pH alone or whether the nitrogensource also played a role.
In an attempt to clarify this point, we preparedNH4
1 HC containing 10 mM of the pH buffer MES.However, the results obtained for this treatment (i.e.,loss of apical dominance and profuse runner hyphalgrowth) suggests that the changes observed werecaused by the MES buffer rather than pH stabiliza-
tion. Vilarino et al (1997) observed a slight increasein root colonization and a strong effect on ERM de-velopment when AM soil cultures were amended withMES. The authors attributed these results first to apossible effect of the buffer on soil micro-organisms.However, the monoxenic conditions used in the pre-sent study indicate that the enhanced growth ob-served in the MES-buffered medium must be due, atleast in part, to other mechanisms. Vilarino et al(1997) also suggested that sulfur in the MES buffer(32 g per mol) might be responsible for the in-creased growth. This hypothesis, supported by pre-vious studies performed in AM fungi grown axeni-cally in the presence of sulfuric compounds (Hepper1984, Bago 1990) seems more likely. In our study 2.6mg sulfur per HC were added as MES. Therefore, itis possible that sulfur has an important effect on ERMdevelopment, but more research is needed to provethis.
Branched absorbing structures are formed by ex-traradical hyphae of AM fungi only after a successfulsymbiosis has been established with a host root (Bagoet al 1998c). Our observations support the idea thatBAS are intrinsic features of extraradical hyphae be-cause they were formed under all experimental con-ditions tested. External conditions, however, stronglyinfluenced their morphology. In contrast, spore for-mation seems to depend on nutrient (mainly nitro-gen) availability and/or environmental conditions,with low pH and/or the presence of ammonium sup-pressing sporulation.
Finally, with regard to sporulation, why did sporeproduction increase threefold in the bromocresol-free WA surrounding the HC, compared to the max-imum sporulation rate observed within a HC(NO3
2)? Although a negative effect of bromcresolpurple on spore formation cannot be ruled out, it isprobable that the extraradical hyphae of AM fungidifferentiate into an exploitative (absorptive) myce-lium in zones where nutrients are available. The nu-trients absorbed then would be moved to older zonesof the fungal colony where sporulation can be initi-ated. Such sporulation likely is a consequence of var-ious nutritional and physiological conditions, such asthe above-mentioned increased C:N ratio and/or col-ony aging.
In conclusion, our results highlight the ability ofthe ERM of Glomus intraradices to adapt its hyphalmorphology and architecture in discrete micrositesto efficiently exploit a given substrate. These resultsshould be further tested by using different AM fungalisolates. New studies to clarify the discrete influenceof nutrients, pH and environmental conditions onthe differential morphogenesis of the AM extraradi-cal mycelium also are needed.

461BAGO ET AL: DIFFERENTIAL MORPHOGENESIS OF AMF
ACKNOWLEDGMENTS
This work was supported by two postdoctoral grants to B.B.(Direccion General de Investigacion Cientıfica y Tecnica,Spain, and Ministere de l’Education, Quebec, Canada). Thefinancial assistance provided by the NSERC to Y.P. also isgratefully acknowledged.
LITERATURE CITED
Abbott LK, Robson AD. 1985. Formation of external hyphaein soil by four species of vesicular-arbuscular mycorrhi-zal fungi. New Phytol 99:245–255.
, de Boer G. 1984. The effect of phosphorus on theformation of hyphae in soil by the vesicular-arbuscularmycorrhizal fungus, Glomus fasciculatum. New Phytol97:437–446.
Azcon-Aguilar C, Bago B, Barea JM. 1998. Saprophyticgrowth of AMF. In: Varma A, Hock B, eds. Mycorrhiza:structure, function, molecular biology and biotechnol-ogy. 2nd ed. Berlin: Springer-Verlag. 391–407.
Bago B. 1990. Efecto de distintos compuestos azufrados enel desarrollo independiente del hongo formador demicorrizas vesıculo-arbusculares Glomus mosseae [Mas-ter’s Thesis]. Granada, Spain: Universidad de Granada.108 pp.
. 2000. Putative sites for nutrient uptake in arbuscularmycorrhizal fungi. Plant Soil 226:263–274.
, Azcon-Aguilar C, Piche Y. 1998a. Extraradical myce-lium of arbuscular mycorrhizae: the concealed exten-sion of roots. In: Flores HE, Lynch JP, Eissenstat D, eds.Radical biology: advances and perspectives on the func-tion of plant roots. Current Topics in Plant Physiology,ASPP Series, 18:502–505.
, , . 1998b. Architecture and develop-mental dynamics of the external mycelium of the ar-buscular mycorrhizal fungus Glomus intraradices grownunder monoxenic conditions. Mycologia 90:52–62.
, , Goulet A, Piche Y. 1998c. Branched absorb-ing structures (BAS): a feature of the extraradical my-celium of symbiotic arbuscular mycorrhizal fungi. NewPhytol 139:375–388.
, Shachar-Hill Y, Pfeffer PE. 2000. Carbon metabolismand transport in arbuscular mycorrhizas. Plant Physiol124:949–957.
, Pfeffer PE, Abubaker J, Jun J, Allen JW, BrouilletteJ, Douds DD, Lammers PJ, Shachar-Hill Y. 2003. Carbonexport from arbuscular mycorrhizal roots involves thetranslocation of glycogen as well as lipid. Plant Physiol131:1496–1507.
, , Douds DD Jr, Brouillette J, Becard G, Shach-ar-Hill Y. 1999. Carbon metabolism in spores of the ar-buscular mycorrhizal fungus Glomus intraradices as re-vealed by nuclear magnetic resonance spectroscopy.Plant Physiol 121:263–271.
, Vierheilig H, Piche Y, Azcon-Aguilar C. 1996. Nitratedepletion and pH changes induced by the extraradicalmycelium of the arbuscular mycorrhizal fungus Glomusintraradices grown in monoxenic culture. New Phytol133:273–280.
, Zipfel W, Williams RC, Jun J, Arreola R, Pfeffer PE,Lammers PJ, Shachar-Hill Y. 2002. Translocation and uti-lization of fungal storage lipid in the arbuscular mycor-rhizal symbiosis. Plant Physiol 128:108–124.
Bailey DJ, Otten W, Gilligan CA. 2000. Saprotrophic invasionby the soil-borne fungal plant pathogen Rhizoctonia so-lani and percolation thresholds. New Phytol 146:535–544.
Becard G, Fortin A. 1988. Early events of vesicular-arbuscularmycorrhiza formation on Ri T-DNA transformed roots.New Phytol 108:211–218.
Beilby JP, Kidby DK. 1980. Biochemistry of ungerminatedand germinated spores of the vesicular-arbuscular my-corrhizal fungus Glomus caledonium: changes in neutraland polar lipids. J Lipid Res 21:739–750.
Bending GD, Read DJ. 1995. The structure and function ofthe vegetative mycelium of ectomycorrhizal plants. V.Foraging behaviour and translocation of nutrients fromexploited litter. New Phytol 130:401–409.
Bonfante P, Balestrini R, Mendgen K. 1994. Storage and se-cretion processes in the spore of Gigaspora margaritaBecker & Hall as revealed by high-pressure freezing andfreeze substitution. New Phytol 128:93–101.
Bonfante-Fasolo P. 1984. Anatomy and morphology of VA my-corrhizae In: Powell CL, Bagyaraj DJ, eds. VA mycorrhi-za. Boca Raton, Florida: CRC Press. p 5–33.
. 1987. Vesicular-arbuscular mycorrhizae: fungus-plantinteractions at the cellular level. Symbiosis 3:249–268.
Chabot S, Becard G, Piche Y. 1992. Life cycle of Glomus in-traradix in root organ culture. Mycologia 84:315–321.
Clark RB, Zeto SK. 1996. Growth and root colonization ofmycorrhizal maize grown on acid and alkaline soil. SoilBiol Biochem 28:1505–1511.
Coughlan AP. 1998. pH and nutrient availability influenceextraradical hyphal morphogenesis and substrate ame-lioration of the arbuscular mycorrhizal fungus Glomusintraradices in monoxenic culture [Master’s thesis]. Que-bec, Canada: Universite Laval. 325 pp.
Cui M, Caldwell MM. 1996a. Facilitation of plant phosphateacquisition by arbuscular mycorrhizas from enriched soilpatches. I. Roots and hyphae exploiting the same soilvolume. New Phytol 133:453–460.
, . 1996b. Facilitation of plant phosphate ac-quisition by arbuscular mycorrhizas from enriched soilpatches. II. Hyphae exploiting root-free soil. New Phytol133:461–467.
Daft MJ, Hacskaylo E, Nicolson TM. 1975. Arbuscular my-corrhizas in plants colonizing coal spoils in Scotland andPennsylvania. In: Sanders FE, Mosse B, Tinker PB, eds.Endomycorrhizas. London: Academic Press. p 581–592.
Delatorre M, Cardenascota HM. 1996. Production of Paeci-lomyces fumosoroseus conidia in submerged culture. En-tomophaga 41:443–453.
Engelkes CA, Nuclo RL, Fravel DR. 1997. Effect of carbon,nitrogen and C:N ratio on growth, sporulation and bio-control efficacy of Talaromyces flavus. Phytopathology87:500–505.
Fomina M, Ritz K, Gadd GM. 2003. Nutritional influence onthe ability of fungal mycelia to penetrate toxic metal-containing domanis. Mycol Res 107:861–871.

462 MYCOLOGIA
Friese CF, Allen MF. 1991. The spread of VA mycorrhizalfungal hyphae in the soil: inoculum types and externalhyphal architecture. Mycologia 83:409–418.
Heijne B, Vandam D, Heil GW, Bobbink R. 1996. Acidifica-tion effects on vesicular-arbuscular mycorrhizal (VAM)infection, growth and nutrient uptake of establishedheathland herb species. Plant Soil 179:197–206.
Hepper CM. 1984. Inorganic sulphur nutrition of the vesic-ular-arbuscular mycorrhizal fungus Glomus caledonium.Soil Biol Biochem 16:669–671.
Jabaji-Hare S. 1988. Lipid and fatty acid profiles of some ve-sicular-arbuscular mycorrhizal fungi: contribution to tax-onomy. Mycologia 80:622–629.
Jakobsen I. 1995. Transport of phosphorus and carbon in VAmycorrhizas. In: Varma A, Hock B, eds. Mycorrhiza:structure, function, molecular biology and biotechnol-ogy. Berlin: Springer-Verlag. p 297–323.
Kendrick B. 1992. The fifth kingdom. Newburyport; Massa-chusetts: Focus Texts. 406 pp.
Klironomos JN, Ursic M, Rillig M, Allen MF. 1998. Interspe-cific differences in the response of arbuscular mycor-rhizal fungi to Artemisia tridentata grown under elevatedatmospheric CO2. New Phytol 138:599–605.
Marsh BAB. 1971. Measurement of length in random ar-rangement of lines. J Appl Ecol 8:265.
Neiderhofer MA, Schenck NC. 1987. Morphological diversitywithin a single vesicular-arbuscular mycorrhizal fungus(VAMF) in a mixed hardwood forest in north Florida.318. In: Sylvia DM, Hung LL, Graham JH, eds. Mycor-rhizae in the next decade, practical application and re-search priorities. Proceedings of the 7th. NACOM, Uni-versity of Florida, Gainesville.
Olsson PA, van Aarle IM, Allaway WG, Ashford AE, RouhierH. 2002. Phosphorus effects on metabolic processes inmonoxenic arbuscular mycorrhiza cultures. Plant Phy-siol 130:1162–1171.
Pascual S, Melgarejo P, Magan N. 1997. Induction of sub-merged conidiation of the biocontrol agent Penicilliumoxalicum. App Microbiol Biotechnol 48:389–392.
Pfeffer PE, Douds DD Jr, Becard G, Shachar-Hill Y. 1999.Carbon uptake and the metabolism and transport of lip-ids in and arbuscular mycorrhiza. Plant Phys 120:587–598.
Ritz K. 1995. Growth responses of some fungi to spatiallyheterogeneous nutrients. FEMS Microbiol Ecol 16:269–280.
Schussler A, Gehrig H, Schwarzott D, Walker C. 2001. Anal-
ysis of partial Glomales SSU rRNA gene sequences: im-plications for primer design and phylogeny. Mycol Res105:5–15.
Shachar-Hill Y, Pfeffer PE, Douds D, Osman SF, Doner LW,Ratcliffe RG. 1995. Partitioning of intermediate carbonmetabolism in VAM colonized leek. Plant Physiol 108:7–15.
Smith FA, Jakobsen I, Smith SE. 2000. Spatial differences inacquisition of soil phosphate between two arbuscularmycorrhizal fungi in symbiosis with Medicago truncatula.New Phytol 147:357–366.
, Smith SE. 1997. Structural diversity in (vesicular)-arbuscular mycorrhizal symbioses. New Phytol 137:373–388.
Smith SE, Read DJ. 1997. Mycorrhizal symbiosis. San Diego,California: Academic Press. 605 pp.
St-Arnaud M, Hamel C, Vimard B, Caron M, Fortin JA. 1996.Enhanced hyphal growth and spore production of thearbuscular mycorrhizal fungus G. intraradices in an invitro system in the absence of host roots. Mycol Res 100:328–332.
Tiwari P, Prakash A, Adholeya A. 2002. Effect of pH on AMfungus Glomus intraradices and Ri T-DNA transformedcarrot roots as host under in vitro. Abstracts of the 7thInternational Mycological Congress, Oslo, Norway.http://www.uio.no/conferences/imc7/Abstract no. 1191
Trinci APJ, Wiebe MG, Robson GD. 1994. The mycelium asan integrate entity. In: Wessels JGH, Meinhardt F, eds.The Mycota I: growth, differenciation and sexuality.Berlin: Springer-Verlag. p 175–193.
van Aarle IM, Olsson PA, Soderstrom B. 2002. Arbuscularmycorrhizal fungi respond to the substrate pH of theirextraradical mycelium by altered growth and root col-onization. New Phytol 155:173–182.
van Bruggen AHC, Termorshuizen AJ, Semenov AM. 2000.Hyphal growth and colony expansion. New Phytol 146:355–356.
Vilarino A, Frey B, Schuepp H. 1997. MES [2-(N-morpho-line)-ethane sulphonic acid] buffer promotes thegrowth of external hyphae of the arbuscular mycorrhi-zal fungus Glomus intraradices in alkaline sand. BiolFertil Soils 25:79–81.
Yu X, Hallett SG, Sheppard J, Watson AK. 1998. Effects ofcarbon concentration and carbon-to-nitrogen ratio ongrowth, conidiation, spore germination and efficacy ofthe potential bioherbicide Colletotrichum coccodes. J In-dustrial Microbiol Biotechnol 20:333–338.





























