Embed Size (px)
Citation preview

Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
Contemporary HypnosisContemp. Hypnosis (2009)Published online in Wiley InterScience(www.interscience.wiley.com) DOI: 10.1002/ch.386
PLASTICITY CHANGES IN THE BRAIN IN HYPNOSIS AND MEDITATION
Ulrike Halsband,1 Susanne Mueller,1 Thilo Hinterberger,2 Simon Strickner1
1 University of Freiburg, Department of Psychology, Neuropsychology, Freiburg, Germany, 2 University Medical Center Freiburg, Department of Environmental Health Sciences, Freiburg, Germany
Abstract
Neuroscientifi c evidence interprets both hypnotic trance induction and different medita-tion traditions as modifi ed states of consciousness that emphasize attention, concentra-tion and the letting go of thoughts, but they differ in terms of sensory input, processing, memory, and the sense of time. Furthermore, hypnosis is based on the suggestibility of a person and meditation on mindfulness; therefore it is not surprising to fi nd differential brain plasticity changes. We analysed shared and non-shared neural substrates using electroencephalography (EEG), positron emission tomography (PET), and functional magnetic resonance imaging (fMRI). Most pronounced EEG changes were in deep as compared to light hypnosis (step-by-step induction) and in arm levitation where sug-gested movement was perceived as external. In a within-subject-design changes in brain activity during hypnosis and Tibetan Buddhist meditation were compared. High ampli-tudes in alpha frequency bands were most pronounced with meditation at frontal posi-tions and with hypnosis in central and temporal locations. Signifi cantly greater activity in theta 2 band was observed only with hypnosis in both hemispheres. PET cerebral activation patterns of imagery-mediated learning were analysed in hypnosis in a within-subject-design. Compared with baseline the learning of high-imagery words was associ-ated with (i) more pronounced bilateral activation in the occipital cortex and prefrontal areas and (ii) improved memory performance. Visual illusion in hypnosis was studied with fMRI, analysed with Granger Causality Mapping, showing changes in the effective connectivity relations of fusiform gyrus, anterior cingulate cortex and intraparietal sulcus. Little is known about the neurobiological basis of the process of enhancing cogni-tive and emotional traits in meditation. In a longitudinal fMRI study attention abilities through intensive Soto-Zen meditation were investigated before (baseline), after training (6 months) and at follow-up (9 months). After six months differences were observed in the left inferior and left superior frontal gyrus; after 9 months activations in the left precuneus. Taken together, the fi ndings advance understanding of the neural mechanisms that underlie hypnosis and meditation. Further studies with a greater sample size are needed to explore the differences and commonalities of hypnosis and different meditation techniques. Copyright © 2009 British Society of Experimental & Clinical Hypnosis. Published by John Wiley & Sons, Ltd.
Key words: brain plasticity, EEG, fMRI, hypnosis, meditation, PET

Halsband et al.
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
Introduction: What is essential in hypnosis and meditation? A neurobiological perspective
A major problem in defi ning hypnosis and meditation is the ambiguity of the terms. ‘Hypnosis’ was derived from the Greek word ‘hypnos’ meaning sleep1 and is character-ized by focused attention, a heightened compliance with suggestion, an awareness of internal images and a reduced ability to think critically. Hypnosis extends to different aspects of the subject’s personal awareness and may turn their experience into a different form of reality (Revenstorf, 1996; Peter, 2009).
The term ‘meditation’ comes from the Latin ‘meditatio’ which originally indicated every type of physical or intellectual exercise. Meditation generically refers to an extremely wide range of practices including Buddhist, Christian, Islamic, Hindu and Jewish traditions. Thus, defi ning meditation involves the need for a precise understand-ing of meditation as a scientifi c explanandum taking into account the importance of various traditions (Lutz et al., 2007). Furthermore, certain aspects that have been described as ‘self-hypnosis’ by one scientist might well be described as ‘meditation’ by another scientist and vice versa (Tart, 2001).
It is important to take the problem of terminology into consideration when the impact of meditation and hypnosis on brain function, attentional and emotional states is inves-tigated. The word ‘meditation’ refers to a broad variety of practices, and failure to make distinctions would be akin to the use of the term ‘sport’ to refer to all sport disciplines as if they were essentially the same (Lutz et al., 2008). Hence, descriptions of various meditation practices imply that there are differences in brain function associated with different types of meditation (Lutz et al., 2004). Lutz et al. (2008) suggested a useful framework in which meditation techniques are grouped into two main categories:
1) focused attention; directing and sustaining attention on a selected object, such as breathing, scriptural passage, mantra, religious pictures, etc. A typical example is Shamatha2, a single pointed, focusing, pacifying and calming meditation technique. The key concept is the concentration of mind. If the mind is wandering meditators show a disengagement of attention from distraction and a shift of attention back to the selected object.
2) open monitoring: no explicit focus on objects. This meditation technique is character-ized by meta-attention. Vipassana is an example of a form of meditation that includes any meditation technique that cultivates insight including contemplation, introspec-tion, analytic meditation and observations about experience.
Common aspects and differences between hypnosis and meditation techniques
The search for the neural correlates that characterize hypnosis and different forms of meditation is a topic of great interest in neuroscience. From a neurobiological point of view, both hypnotic trance induction (e.g. Rainville et al., 2002; Halsband, 2006) and meditation (e.g. Holroyd, 2003; Lutz et al., 2007) can be interpreted as a modifi ed state of consciousness which refl ects a dynamic change of brain activity. The experiencer feels a qualitative alteration in the overall pattern of mental functioning such that their con-sciousness is radically different from the way it functions ordinarily (Tart, 1972); this is linked to plasticity chances in the brain. Consciousness is a multifaceted concept. Accord-ing to Laureys (2005) it can be viewed as having two major components: i) awareness

Plasticity changes in the brain
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
of the environment and of the self (i.e. the content of consciousness); and ii) the level of vigilance or arousal. Hypnosis and different kinds of meditation traditions emphasize attentional focus, concentration and letting go of thoughts. So, at a fi rst glance hypnosis and meditation seem to have lots in common.
However, there are also crucial behavioural differences between hypnosis and medita-tion. A main difference is that hypnosis is based on the suggestibility of a person and meditation is focused on mindfulness (Holroyd, 2003). Table 1 directly compares hyp-nosis and two kinds of meditation techniques: focused attention, Shamatha; and open monitoring, Vipassana. The table was modifi ed and extended after Tart (2000, 2001) and it shows differences and commonalities in terms of interactions, sensory input and pro-cessing, concentration, memory, sense of time and stress coping.
In hypnosis a special rapport with the hypnotist is of crucial importance, whereas in the meditation context independence from needing social relationships is expected. The special rapport between the hypnotist and the hypnotized person as well as the absorption of the voice of the hypnotist takes a key role, whereas such interpersonal interactions are not of relevance for meditation techniques. Meditation, however, interacts with indepen-dence, empathy for all living beings and bliss.
In hypnosis sensory processing is limited and determined by suggestions. Sensory processing is focused and deliberately controlled in Shamatha meditation. In Vipassana the input/processing is mainly aware-controlled.
Hypnotic trance is accompanied by a heightened suggestibility, in which suppressed memories may be experienced. In hypnotic trance age-regression or age-progression can be used therapeutically to allow the subjects to experience or re-experience all forms of inner sensory, perceptual or emotional events. Concerning memory processing, results
Table 1. Comparison of hypnosis with Shamatha and Vipassana meditation
Hypnosis
Meditation
Shamathafocused attention
Vipassanaopen monitoring
Interactions special rapport independence, empathy for all living beings, bliss
Input/Processing limited and determined by the suggestion
focused, deliberately controlled
meta-attention, aware-controlled
Concentration increased concentration Memory explicit learning:
increased memory for high-imagery material, reduced memory for abstract material
improved implicit learning
inactive, focus on the present
Sense of Time time distortion, progression/regression
focus on the here and now
Stress Coping reduction of stress

Halsband et al.
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
showed an improved recall of high-imagery word pairs under hypnosis, but a decrease in the ability to learn abstract word pairs (Halsband, 2004, 2006). Furthermore, the use of implicit knowledge and implicit information processing play a key function in hyp-nosis. In contrast, memory does not play a major role in meditation: the meditator is asked to remain in the ‘here and now’; the main focus of meditation is the present.
Both, hypnosis and different forms of meditation practices show benefi cial effects for coping with stress. It is possible to relax one’s mind, soul and body. The techniques can also be used as prevention methods to avoid harmful effects of stress induction. People suffering from stress are often exhausted, lack concentration and have a weakened immune system. Using meditation and hypnosis people have a powerful tool to calm down and to disentangle themselves from their stressful environment.
From a neurobiological point of view a critical question to be addressed is as follows: what are the shared and non-shared neural substrates in hypnosis and in different forms of meditation?
Neurophysiology: EEG studies
HypnosisAttentional control, concentration, imagination, mental relaxation, altered perception of the environment, disengagement of the discursive, and critical analytical reasoning are all characteristic elements of hypnosis (Halsband, 2008). Numerous EEG studies showed changes in neuroelectrical activity. There is evidence for a higher proportion of occipital alpha waves in high suggestibles as compared to lows (e.g. London et al., 1968; Bakan and Svorad, 1969; Engstrom et al., 1970; Ulett et al., 1972; Morgan et al., 1974; Edmon-ston and Grotevant, 1975), but this fi nding was not replicated by some other studies (Barabasz, 1983; Perlini and Spanos, 1991).
Signifi cantly greater activity in high alpha (11.5–13.45 Hz), beta (16.5–25 Hz) and high theta (5.5–7.5 Hz) band was reported in highs in the right parietal cortex (Crawford et al., 1996). Several authors reported an increase of the power in the theta frequency band (Tebecis et al., 1975; Crawford, 1990; Sabourin et al., 1990; Graffi n et al., 1995; De Pascalis et al., 1998). In contrast to meditation, increased beta activity in the right occipi-tal cortex was measured (Ulett et al., 1972) and a signifi cant power increase in the right parietal region (Crawford et al., 1996). In addition, there is evidence for an increase in the gamma band power around 40 Hz – more pronounced over the right than the left hemisphere (De Pascalis et al., 1998) and in the parieto-midline-to-right temporal areas (Schnyer and Allen, 1995).
High-hypnotizables – as compared to lows – also produced a higher theta 1 amplitude (4–6 Hz) in bilateral frontal and right posterior areas (De Pascalis et al., 1998). In the bilateral frontal cortex the same subjects showed a smaller alpha 1 (8.25–10 Hz) ampli-tude. High and low hypnotizables were also distinguished by means of changes in mis-match negativity across hypnosis and pre- and post-hypnosis conditions (Jamieson et al., 2005). Isotani et al. (2001) made an important point: already before hypnosis was induced, high and low susceptible subjects were in different brain electric states. In high-hypnotizables posterior brain activations were most pronounced whereas lows pre-sented with anterior weighted brain activation patterns.
Fingelkurts et al. (2007) published a single case study of a highly susceptible subject (virtuoso). During hypnosis there were alterations in all studied frequency bands (delta, theta, alpha, beta, and gamma) which were stable after one year. Results gave evidence

Plasticity changes in the brain
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
for local and remote cortex functional connectivity changes. Interestingly, the authors reported a disruption in the functional synchrony among neural assemblies within the left frontal cortex which is consistent with Gruzelier s fi ndings (2000) of hypofrontality and left-hemisphere inhibition.
Taken together the results are very heterogeneous. This may be partly caused by the great variability in intracerebral source location, EEG dimensionality, the technology and methods of analysis used as well as by the different ways (e.g. direct/indirect sug-gestions, confusion techniques) and various stages of trance induction (e.g. auto-focus-ing, arm levitation, deep trance experience). Most studies failed to disentangle the different state patterns of the brain during specifi c periods of the trance induction (see next chapter).
Arm levitation and deep trance inductionWe present a single case EEG-study on a highly susceptible person (male/aged 25)3 who was guided through a hypnotic trance induction. The experimental setting comprised the measurement of 64 channel EEG plus peripheral physiological measures. The par-ticipant was seated on a chair approximately one meter in front of the hypnotist. A 15 min baseline session was recorded consisting of 5 min sitting relaxed with eyes open, 5 min with eyes closed, and 5 min of reading a text from a book. Subsequently a hypnotic induction (26 min) was recorded, subdivided into the following phases: i) auto-focussing and introduction, ii) fostering mental and physical relaxation, iii) arm levitation, iv) step-by-step instructions to allow the subject to enter a deeper state of trance (counting from 1 to 10), v) metaphoric story, trance anchoring, and vi) termination, when the hypnotic trance was brought to an end.
For statistical analysis, the EEG data were fi rst corrected for eye movements. Phases with large amplitude movement artefacts were removed from the analysis. A Fast Fourier Transform (FFT) was applied using a window size of 2 seconds. The resulting Fourier amplitudes were converted into spectral power by squaring. The power spectral density (PSD) in the bands of interest was defi ned as the mean of the Fourier coeffi cients of 4 to 7.5 Hz (theta band), 8 to 12 Hz (alpha band), and 25.5 to 70 Hz (gamma band). The region from 47 to 53 Hz was ignored due to the possible 50 Hz humming artefact. In the following, the results of the two phases with the most profound changes in brain activa-tions are reported: stepwise trance induction and arm levitation phase. These two phases led to distinct changes in theta, alpha and gamma frequency bands. Full details will appear later (Hinterberger and Halsband, to be submitted).
Hypnotic suggestions may dissociate physical movements from normal conscious volition (Kirsch and Lynn, 1997; Weitzenhoffer, 2000; Heap and Aravind, 2002). A typical example is arm levitation, which is an important sign for verifying a trance induction. It is a frequently demonstrated hypnotic phenomenon in which self-induced movements are attributed to an external source (Oakley, 1999; Heap and Aravind, 2002; Raz and Shapiro, 2002; Blakemore et al., 2003;). The experience of such an anomalous control is different from the normal conscious experience of a similar movement pro-duced intentionally (Haggard et al., 2004). We gave the hypnotized subject the suggestion that his arm is being raised upwards by a helium balloon. This suggestion resulted in the appropriate movement. The left arm of our subject started to rise on itself after the suggestion and remained upraised.
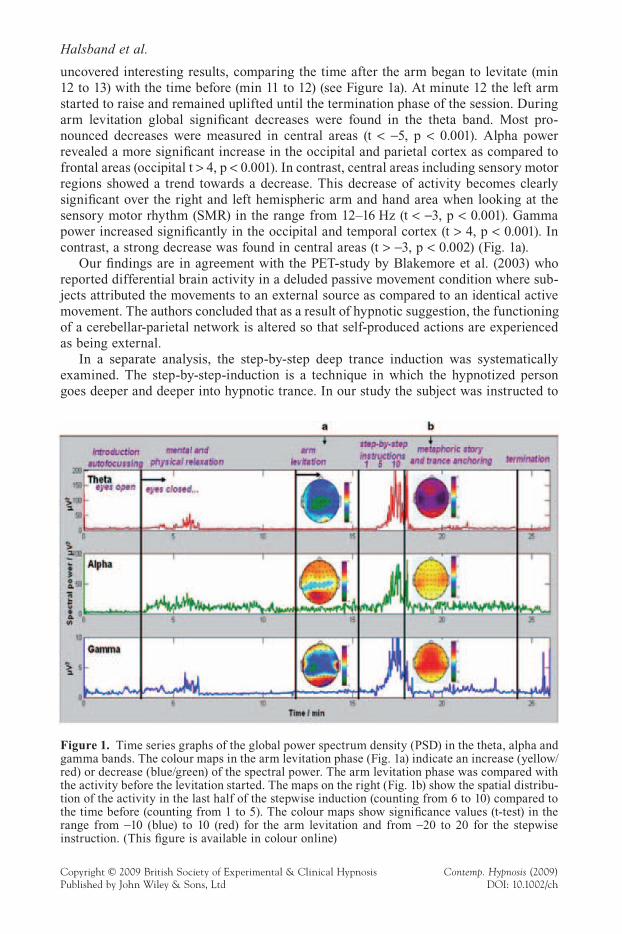
During the arm levitation phase changes in the global PSD could not be clearly observed on the time series graph. However, a brain mapping statistical analysis
Halsband et al.
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
uncovered interesting results, comparing the time after the arm began to levitate (min 12 to 13) with the time before (min 11 to 12) (see Figure 1a). At minute 12 the left arm started to raise and remained uplifted until the termination phase of the session. During arm levitation global signifi cant decreases were found in the theta band. Most pro-nounced decreases were measured in central areas (t < !5, p < 0.001). Alpha power revealed a more signifi cant increase in the occipital and parietal cortex as compared to frontal areas (occipital t > 4, p < 0.001). In contrast, central areas including sensory motor regions showed a trend towards a decrease. This decrease of activity becomes clearly signifi cant over the right and left hemispheric arm and hand area when looking at the sensory motor rhythm (SMR) in the range from 12–16 Hz (t < !3, p < 0.001). Gamma power increased signifi cantly in the occipital and temporal cortex (t > 4, p < 0.001). In contrast, a strong decrease was found in central areas (t > !3, p < 0.002) (Fig. 1a).
Our fi ndings are in agreement with the PET-study by Blakemore et al. (2003) who reported differential brain activity in a deluded passive movement condition where sub-jects attributed the movements to an external source as compared to an identical active movement. The authors concluded that as a result of hypnotic suggestion, the functioning of a cerebellar-parietal network is altered so that self-produced actions are experienced as being external.
In a separate analysis, the step-by-step deep trance induction was systematically examined. The step-by-step-induction is a technique in which the hypnotized person goes deeper and deeper into hypnotic trance. In our study the subject was instructed to
Figure 1. Time series graphs of the global power spectrum density (PSD) in the theta, alpha and gamma bands. The colour maps in the arm levitation phase (Fig. 1a) indicate an increase (yellow/red) or decrease (blue/green) of the spectral power. The arm levitation phase was compared with the activity before the levitation started. The maps on the right (Fig. 1b) show the spatial distribu-tion of the activity in the last half of the stepwise induction (counting from 6 to 10) compared to the time before (counting from 1 to 5). The colour maps show signifi cance values (t-test) in the range from !10 (blue) to 10 (red) for the arm levitation and from !20 to 20 for the stepwise instruction. (This fi gure is available in colour online)

Plasticity changes in the brain
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
go on an imaginary journey in hypnosis. The further the participant had to walk in imagination downstairs the deeper was the level of experienced hypnotic trance.
In the deep hypnotic condition (stepwise induction 6–10) differential brain activity changes occurred that were not present in light hypnotic conditions. During the last half of the stepwise trance induction phase a highly signifi cant increase in all frequency bands was observed. The participant was sitting completely motionless. For a more detailed examination a statistical comparison (t-test) was calculated for each electrode separately between the time interval counting from six to ten (44 seconds) and counting from one to fi ve (50 seconds). The colour maps in Figure 1b show highly signifi cant increases in all frequency bands. In the theta band (t > 10.0, p < 0.001) a strong global increase was measured with a special emphasis on sensory motor areas bilaterally. In the alpha band the increases were less pronounced (t > 3, p < 0.002 over sensory motor areas) but sig-nifi cant – again with a higher activation rate bilaterally in sensory motor areas. In addi-tion, within the gamma band strongest activations were recorded in parietal, central, and frontal brain regions (t > 4, p < 0.001 over central areas) (Fig. 1b).
Our fi ndings are in agreement with the results by Katayama et al. (2007) who reported differences in brain activity in deep and light hypnosis. Furthermore, our most pro-nounced increases in activity within the sensory-motor areas in alpha-, gamma- and theta frequency bands should be interpreted in the context of motor imagery. Interestingly, other fi ndings confi rm a relationship between motor imagery and hypnotic responding (Konradt et al., 2005).
MeditationDuring meditation sensory input is diminished (Carrington, 1998; Dietrich, 2003). In meditation sustained concentration and heightened awareness can be achieved by focus-ing attention on mantra, breathing rhythm, or a number of other internal or external events (Herzog et al., 1990; Lou et al., 1999; Lazar et al., 2000; Newberg et al., 2001; Dietrich, 2003). This is accompanied by changes in neuroelectrical activity and indicated by increases in alpha, gamma and theta waves (e.g. Anand et al., 1961; Banquet, 1972; Corby et al., 1978; Benson et al., 1990). Under meditation – as compared to relaxation – an increase in alpha amplitudes was observed (Wallace, 1970; Wallace and Benson, 1972; Banquet, 1973; Glueck and Stroebel, 1975) which appeared to be most pronounced in the frontal cortex (Kesterson, 1989; Sudsuang et al., 1991; Jevning et al., 1992). An increase in theta power was reported by several authors in different types of meditation (e.g. Pan et al., 1994 for Qi Gong; Kubota et al., 2001 for Zen breath counting; Aftanas and Golocheikine, 2002 for Sahaja yoga; Kjaer et al., 2002 for yoga nidra). Lutz et al. (2004) found in eight long-term practitioners of the Kagyüpa- and Nyingmapa- schools (compared to 10 non-practising students) a high amplitude activity and a marked phase synchronization in the gamma-band (between 25 and 42 Hz), especially in lateral fronto-parietal locations (see also Banquet, 1973).
Coromaldi et al. (2004) investigated the EEG-activity in a Zen-master – a highly experienced subject – during deep meditation. The results of the deepest meditation stage showed an increase in the alpha band (8–13 Hz) and theta-power (4–7 Hz) at all locations and most prominent in the left parietal cortex. In contrast, there was a reduction of beta-activity (15–28 Hz) over the right hemisphere. Aftanas and Golosheikine (2005) made an important point: in long-term meditators (Sahaja yoga tradition) changes in EEG activity were dynamical and dependent on the arousal level. Increasing the arousal level (viewing aversive video clips) desynchronized activities in theta and alpha frequency bands.
Halsband et al.
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
Taken together the results are controversial. There are different kinds of meditation techniques and therefore the fi ndings are heterogeneous. The heterogeneity is addition-ally caused by the great variability in the degree of experience in meditation (Halsband, 2008). It is also diffi cult to compare recent and older studies because technology and analytical procedures have changed.
Intersection: hypnosis and meditationThere is an increasing interest in plasticity changes in the brain in hypnosis compared to different types of meditation (Holroyd, 2003; Otani, 2003; Grant and Rainville, 2005). In the present study a within-subject design was used to allow for a direct comparison of brain changes in hypnosis and a Tibetan form of meditation (Vajrayana-Buddhism).
EEG activity was recorded under hypnosis and meditation in a high-suggestible (according to the German version of the Harvard Scale of Hypnotic Suggestibility) male subject (43 years) who practised meditation for 18 years and has started every day with meditation exercises. 32 channel EEG was recorded with an electrode cap at standard positions of the extended international 10/20 system. Data were analyzed with the Fast-Fourier-Transformation for the following frequency bands: theta 1 (3.71–5.57 Hz), theta 2 (5.57–7.42 Hz), alpha 1 (7.42–9.28 Hz), alpha 2 (9.28–11.13 Hz), beta (13–30 Hz) and gamma (30–49 Hz). The frequency windows were defi ned individually as suggested by Klimesch (1999) and Aftanas and Golocheikine (2001) using alpha-peak-frequency of the subject as a reference point.
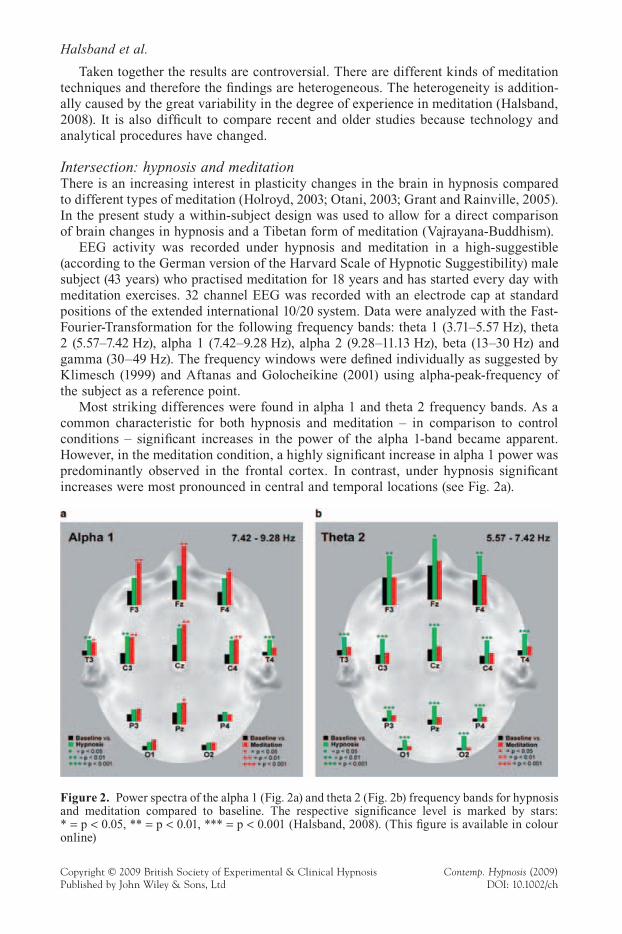
Most striking differences were found in alpha 1 and theta 2 frequency bands. As a common characteristic for both hypnosis and meditation – in comparison to control conditions – signifi cant increases in the power of the alpha 1-band became apparent. However, in the meditation condition, a highly signifi cant increase in alpha 1 power was predominantly observed in the frontal cortex. In contrast, under hypnosis signifi cant increases were most pronounced in central and temporal locations (see Fig. 2a).
Figure 2. Power spectra of the alpha 1 (Fig. 2a) and theta 2 (Fig. 2b) frequency bands for hypnosis and meditation compared to baseline. The respective signifi cance level is marked by stars: * = p < 0.05, ** = p < 0.01, *** = p < 0.001 (Halsband, 2008). (This fi gure is available in colour online)

Plasticity changes in the brain
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
Interestingly, signifi cantly greater activity in theta 2 band was observed only under hypnosis, but not under Tibetan meditation. Theta band activity changes in hypnosis were observable in both hemispheres (see Fig. 2b).
Taken together brain plasticity changes in hypnosis can be clearly differentiated from neurophysiological changes in meditation. Further research is needed to systematically compare shared and non-shared neuronal substrates in different kinds of meditation tra-ditions and different levels of hypnotic trance.
Brain imaging tools in neuroscience
Functional magnetic resonance imaging (fMRI) and positron emission tomography (PET)A major breakthrough in the study of the cortical plasticity during hypnosis and medita-tion was the use of modern brain imaging techniques. In this fascinating world of neu-roscience, efforts for an improved understanding of the brain mechanisms involved were made by the use of positron emission tomography (PET) and functional magnetic reso-nance imaging (fMRI). The PET tomograph is an external detection system that maps the distribution of an injected radiotracer in three-dimensional space. The masses of the positron and electron are converted into two photons emitted in directly opposite direc-tions (180°) with the same energy. By contrast, fMRI is a non-invasive method based on the increase in blood fl ow to the local vasculature that accompanies neural activity in the brain. Changes in the oxygenation level of the blood, the so-called BOLD (Blood Oxygenation Level Dependent) effect, occurs as a consequence of neuronal activity. The magnitude of change in signal intensity can be used as an indirect measure of excitary input to neurons (Logothetis et al., 2001). A detailed description of brain imaging tools was given by Otte and Halsband (2006). Earlier, we reported a preponderance of com-monalities in the activation patterns yielded with fMRI and PET (e.g. Halsband et al., 1998, 2002; Krause et al., 1999a, b; Mottaghy et al., 1999, 2000; Schmidt et al., 2002); these fi ndings were confi rmed by other studies (e.g. Schall et al., 2003).
The decision to choose one or another neuroimaging modality depends on the task of interest. Major advantages of fMRI are 1) that this technique does not need radioactive tracers which is of value especially in studies with repetitive measurements and follow-up investigations; and 2) a better spatial and time resolution. However, special skills are needed by the experimenter to ensure that subjects would relax with their heads restrained in the noisy MRI scanner environment. Although it is possible to reduce the noise level to about 70 decibels with the help of a special fMRI-compatible headset, this background noise remains an inadequate interference. In fMRI-studies on hypnotic trance induction we found it useful to keep the noise level stable in order not to interrupt the state of deep relaxation (Halsband, 2009).
There is hope for an introduction of a ‘silent fMRI’. This promising new approach was introduced by Schmitter et al. (2008). The authors reported about a new low-noise echo-planar imaging (EPI) that is optimized for auditory fMRI measurements. The sequence produces a narrow-band acoustic frequency spectrum by using a sinusoidal readout echo train and a constant phase encoding gradient. This could be a most useful tool for future studies on hypnosis and meditation.
HypnosisSeveral authors have shown that plastic changes in neuronal activity occur after hypno-tic trance induction (e.g. Grond et al., 1995; Crawford et al., 1998; Szechtman et al.,

Halsband et al.
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
1998; Maquet et al., 1999; Rainville et al., 1999, 2002; Faymonville et al., 2000; Kosslyn, et al., 2000; Halsband, 2004; 2006; Spiegel and Kosslyn, 2004; Egner et al., 2005).
In a PET-study by Maquet et al. (1999) subjects were allowed to listen to pleasant autobiographical memories. During hypnosis signifi cant activations were observed in a complex neural network including occipital, parietal, precentral, prefrontal and cingulate cortices. Using PET, Rainville et al. (2002) reported that hypnotic relaxation involved an increase in occipital regional Cerebral Blood Flow (rCBF), a decrease in cortical arousal and a reduction in cross-modality suppression (disinhibition). In contrast, increases in mental absorption during hypnosis were associated with rCBF increases in a distributed network of cortical and subcortical structures previously described as the brain’s atten-tional system.
Using event-related fMRI and EEG coherence measures, Egner et al. (2005) were able to show that individual differences in hypnotic susceptibility are linked with the effi ciency of the frontal attention system, and that the hypnotized condition is character-ized by a functional dissociation of confl ict monitoring and cognitive control processes. Muzur et al. (2006) concluded that hypnosis and suggestion are methods of external manipulation with frontal-lobe functions. The effectiveness of hypnosis in producing analgesia has been well documented (e.g. Faymonville et al., 1997, 2000, 2006; Rainville et al., 1999). Faymonville and co-workers came to the conclusion that the midcingulate cortex (Brodman area 24) mediates the hypnosis-induced analgesia. They reported that the reduced nociception under hypnosis is mediated by an increased functional connec-tivity between the midcingulate cortex and insular, pregenual, frontal and pre-SMA regions as well as brainstem, thalamus and basal ganglia.
Hypnotic visual illusionIn hypnosis, perceptual changes may occur. This is accompanied by changes in brain activation. Using PET, Kosslyn et al. (2000) reported that hypnotic illusion of colour induced blood fl ow changes consistent with actually observing colour. It was found that colour areas were activated under hypnosis when subjects were asked to perceive colour, whether they were actually shown the colour or a grey-scale stimulus. It was concluded that among highly hypnotizable subjects, observed changes in subjective experience in hypnosis were refl ected by changes in brain function.
But, critically speaking, in the study by Kosslyn et al. (2000) colour change sugges-tions were only given in hypnosis, whereas imagination instructions were given outside hypnosis. Hence it remains unclear whether differences between the two conditions refl ect the effects of suggestion or hypnotic trance or a combination of the two. In other words, it is not possible to conclude from the data that hypnotic trance was the factor that created the differences in brain activity. In cooperation with the University of Maas-tricht we used fMRI and an experimental design in which the same suggestions were given in and outside hypnosis (Otto, 2007). RCBF changes were measured using fMRI with short time repetition to obtain high temporal resolution data. Granger Causality Mapping (Roebroeck et al., 2005) was used to identify voxel time courses for inferring directions of neural interactions and information fl ow.
Three subjects were asked to see a grey-scale in colour or a colour pattern in a grey scale (visual illusion) or to see a colour pattern in colour and a grey scale as a grey scale (natural suggestion). The same instructions were given in hypnosis and without hypnosis.
Plasticity changes in the brain
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
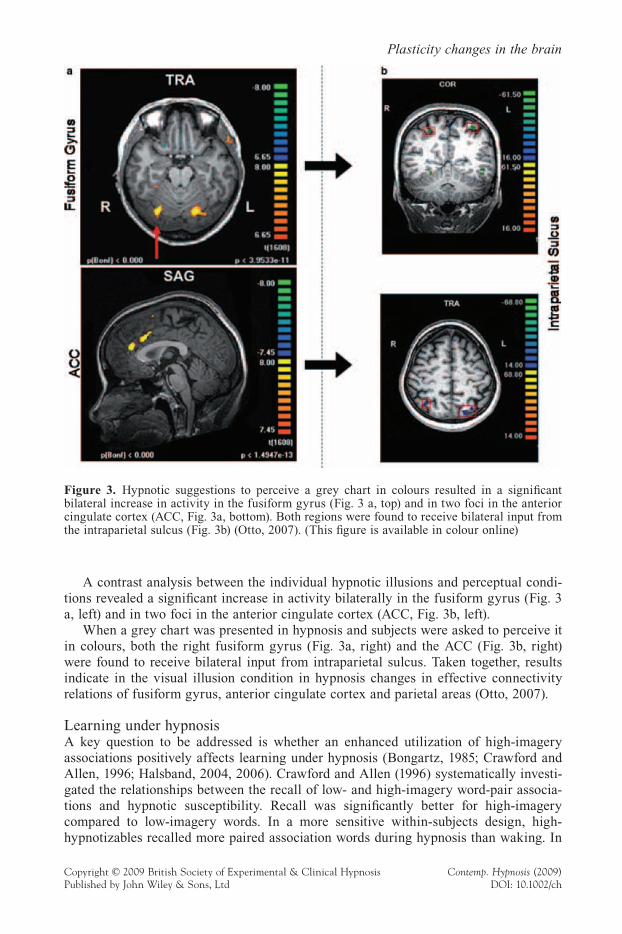
A contrast analysis between the individual hypnotic illusions and perceptual condi-tions revealed a signifi cant increase in activity bilaterally in the fusiform gyrus (Fig. 3 a, left) and in two foci in the anterior cingulate cortex (ACC, Fig. 3b, left).
When a grey chart was presented in hypnosis and subjects were asked to perceive it in colours, both the right fusiform gyrus (Fig. 3a, right) and the ACC (Fig. 3b, right) were found to receive bilateral input from intraparietal sulcus. Taken together, results indicate in the visual illusion condition in hypnosis changes in effective connectivity relations of fusiform gyrus, anterior cingulate cortex and parietal areas (Otto, 2007).
Learning under hypnosisA key question to be addressed is whether an enhanced utilization of high-imagery associations positively affects learning under hypnosis (Bongartz, 1985; Crawford and Allen, 1996; Halsband, 2004, 2006). Crawford and Allen (1996) systematically investi-gated the relationships between the recall of low- and high-imagery word-pair associa-tions and hypnotic susceptibility. Recall was signifi cantly better for high-imagery compared to low-imagery words. In a more sensitive within-subjects design, high- hypnotizables recalled more paired association words during hypnosis than waking. In
Figure 3. Hypnotic suggestions to perceive a grey chart in colours resulted in a signifi cant bilateral increase in activity in the fusiform gyrus (Fig. 3 a, top) and in two foci in the anterior cingulate cortex (ACC, Fig. 3a, bottom). Both regions were found to receive bilateral input from the intraparietal sulcus (Fig. 3b) (Otto, 2007). (This fi gure is available in colour online)

Halsband et al.
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
contrast, lows did not differ. However, in a between-subjects design, hypnotic level was not a moderator of performance during hypnosis. Taken together, the fi ndings by Craw-ford and Allen (1996) are controversial and do not allow a general conclusion that the use of imagery strategies in high-hypnotizables increases their learning ability. Further-more, the neural correlates of learning under hypnosis remained unclear.
Using O–15 water PET in a within-subject design the neural mechanisms of encoding and retrieval of high-imagery words in high-hypnotizables were systematically investi-gated under hypnosis and in the waking state (Halsband, 2006). Subjects were assessed on the German version of the Harvard Group Scale of Hypnotic Susceptibility (HGSHS Form A). Seven highly hypnotizable subjects with a susceptibility score >7 were assigned to the PET study. Their mean age was 25.4 years (sd 3.1).
A verbal episodic memory task was used. During encoding subjects were visually presented 12 word pairs. Words implemented in the study were two-syllable German high-imagery nouns that were of high frequency (Meier, 1964). Afterwards, the subjects were asked to retrieve the corresponding word-pair associate after having been randomly presented the fi rst of the two words of each word pair (retrieval). Two reference condi-tions were used, either containing 12 single nonsense words (two-syllable pseudo words that obey German spelling rules) or 12 nonsense word pairs.
The word pairs were semantically unrelated and therefore diffi cult to associate (e.g. monkey-candle). Word pairs were of high-imagery according to a German linguistic database of 800 nouns on a scale between 6 and 7 (Baschek et al., 1977).
Results indicate during the encoding phase in hypnosis a most pronounced occipital activation and an increased prefrontal activity When word pairs were retrieved previ-ously learned under hypnosis, a stronger activation in the prefrontal cortex and cerebel-lum, as well as an additional bilateral activation in the occipital lobe were reported (Halsband, 2006).
Highly hypnotizables benefi t from hypnosis when they have to acquire word pairs with high-imagery content. Halsband and Herfort (2007) found that highly hypnotizable subjects show a better learning performance of high-imagery word-pair associations than do low-hypnotizable subjects. However, the ability to retrieve abstract word-pair associa-tions (e.g. wisdom-moral) strongly decreased when encoding took place in trance.
MeditationAttentional regulation is a common cognitive function associated with divergent medita-tion methods (Cahn and Polich, 2006). Several authors have demonstrated that meditation practice alters brain activity in areas important for sensory, cognitive and emotional processing (e.g. Newberg and Iversen, 2003; Lazar et al., 2005; Doraiswami and Xiong, 2007; Jha et al., 2007; Srinivasan and Baijal, 2007) concluded that an increased activity in the bilateral prefrontal cortex and the cingulate gyrus already appears at an early stage of meditation. Using fMRI, Baron et al. (2007) tested subjects with at least four years of regular meditative practices from different meditative traditions (Tibetan Buddhists, Zen Buddhists, Yoga practitioners) and different experiences in meditation practices and durations. Results indicate that brain activities in the dorsal lateral prefrontal cortex and anterior cingulate cortex varied over the time of a meditation session and differed between long- and short-term practitioners. In the more practiced subjects regional brain activations correlated with better sustained attention and attentional error monitoring.
With regard to focused attention meditation (example: Shamatha) neural activations associated with selective attention including the temporal-parietal junction, intraparietal sulcus, ventro-lateral prefrontal cortex and frontal eye-fi elds are of crucial importance

Plasticity changes in the brain
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
(Corbetta and Shulman, 2002). Furthermore, brain areas engaged in sustaining attention, such as right frontal and parietal cortex and the thalamic structures, are involved (Kastner et al., 1999). Long term meditators showed less amygdala activation during meditation compared to novices (Slagter et al., 2007). Interestingly neural networks associated with confl ict monitoring, i.e. the dorsal anterior cingulate cortex and dorsolateral prefrontal cortex seem to play an important role in both focused meditation and hypnosis (Coull, 1998; Raz et al., 2006).
In open monitoring, meta-attention meditation (for example, Vipassana), the aware-ness of the subjective features of a given moment and its emotional tone are of crucial importance. One may therefore argue that brain regions involved in focusing or sustain-ing attention onto a specifi c object are of less importance. But instead processes that rely on meta-representation in the brain are critically involved including the anterior insula, somatosensory cortex and anterior cingulate cortex (Damasio, 2000; Craig, 2000). Recently, 15 Vipassana meditators were compared with 15 non-meditators. Results indi-cate stronger activation in the rostral anterior cingulate cortex and bilaterally in the dorsal medial prefrontal cortex (Hoelzel et al., 2007).
So far only a few studies are available that specifi cally compared aspects of anatomi-cal correlates between meditators and non-meditators (Lazar et al., 2005; Hoelzel et al., 2008; Luders et al., 2009; Vestergaard-Poulsen et al., 2009). Lazar et al. (2005) reported that brain regions associated with attention, interoception and sensory processing were thicker in long-term meditators (Vipassana) as compared to a non-meditating control group including the prefrontal cortex and anterior insula. Differences in prefrontal cortex thickness were most pronounced in the most experienced and older subjects.
Hoelzel et al. (2008) found differences in grey matter concentration associated with long-term meditation (Vipassana). These fi ndings are consistent with the results by Vestergaard-Poulsen et al. (2009) who reported increased grey matter density in medita-tors in the left superior and inferior frontal gyri, the left fusiform gyrus, the lower brain stem and the bilateral anterior lobes of the cerebellum. Most recently Luders et al. (2009) found larger hippocampal and frontal volumes of grey matter in long-term meditators (range of meditation: 5–46 years) using different meditation techniques (Zazen, Samatha, Vipassana, etc.). Taken together the results suggest that long-term meditation can induce changes in brain structure. However, it remains an open question whether alterations in brain functions like enhanced attentional capacities are caused by long-term meditation training itself or by individual personality differences. Therefore longitudinal studies are needed to follow individuals over time in response to mental training.
Zen meditation and attention: a longitudinal studyTaken together, there have been two major approaches in meditation research: 1) studies on advanced and highly skilled meditators, and 2) studies on novices who usually started meditation training in a clinical setting as adjuvant therapy. However, as yet little is known about the neurobiological basis of the process of enhancing cognitive and emo-tional traits. Therefore we tried to bridge the gap between the two before mentioned experimental designs by conducting a longitudinal study on meditation training with different types of assessment strategies including fMRI.
We conducted an fMRI study to examine longitudinal meditation effects in a within-subject design. Seven persons without previous meditative experience but the motivation to take part in an intensive Zen meditation training (Soto) during 6 months were sys-tematically evaluated. Assessments (psychological, neurophysiological) were conducted before (baseline), after the training (6 months) and in a follow-up (9 months). Our main
Halsband et al.
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
focus was on the process of enhancing attention abilities through meditation. By studying the neural mechanisms underlying attention we also analyzed their relation to changes of emotional and socio-behavioural traits. Full details will appear in Mueller and Hals-band (to be submitted).
A useful experimental paradigm in studying the neural correlates of (focused) atten-tion and conscious perception – as described in the study of Carter et al. (2005) – is binocular rivalry. Binocular rivalry involves perceptual alternations between competing monocular images. Because transitions between each monocular view occur without any change of the physical stimulus, perceptual processes can be clearly distinguished from those due to stimulus characteristics. While this paradigm has traditionally been of inter-est to visual scientists, it has recently gained wider prominence as a tool for studying visual consciousness and focused attention. It could be shown in previous studies that observers were able to voluntarily control the alternation rate of the competing images. We used face and house images as stimuli which were adopted with friendly permission from previous fMRI work (Tong et al., 1998; Meng and Tong, 2004).4 Based on the biased competition model of selective attention we expected our participants to enhance the attended percept while suppressing the unattended one. Moreover we expected an improvement in the ability to voluntarily control the duration of the attended percept during the course of the six months intensive meditation training.
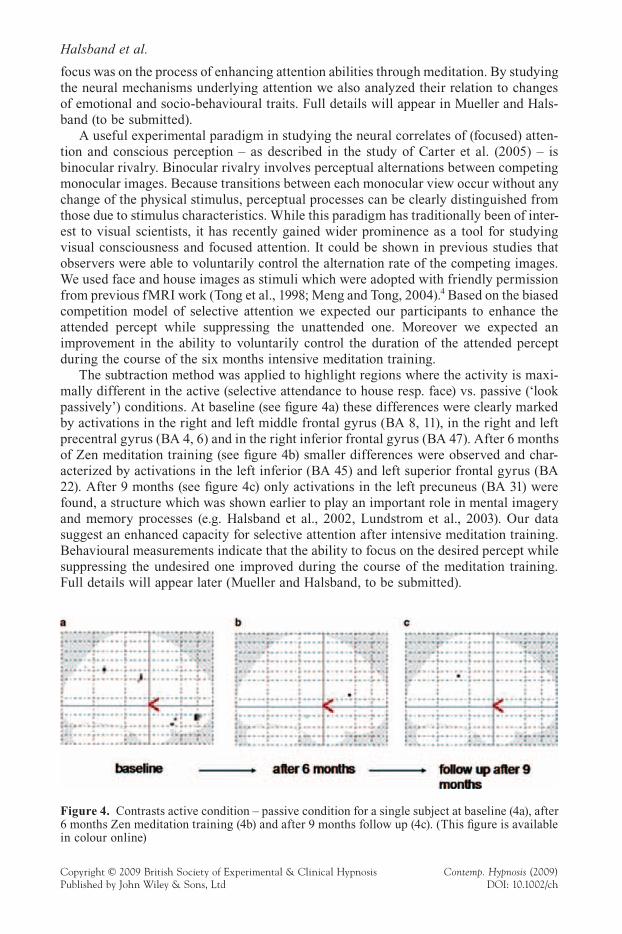
The subtraction method was applied to highlight regions where the activity is maxi-mally different in the active (selective attendance to house resp. face) vs. passive (‘look passively’) conditions. At baseline (see fi gure 4a) these differences were clearly marked by activations in the right and left middle frontal gyrus (BA 8, 11), in the right and left precentral gyrus (BA 4, 6) and in the right inferior frontal gyrus (BA 47). After 6 months of Zen meditation training (see fi gure 4b) smaller differences were observed and char-acterized by activations in the left inferior (BA 45) and left superior frontal gyrus (BA 22). After 9 months (see fi gure 4c) only activations in the left precuneus (BA 31) were found, a structure which was shown earlier to play an important role in mental imagery and memory processes (e.g. Halsband et al., 2002, Lundstrom et al., 2003). Our data suggest an enhanced capacity for selective attention after intensive meditation training. Behavioural measurements indicate that the ability to focus on the desired percept while suppressing the undesired one improved during the course of the meditation training. Full details will appear later (Mueller and Halsband, to be submitted).
Figure 4. Contrasts active condition – passive condition for a single subject at baseline (4a), after 6 months Zen meditation training (4b) and after 9 months follow up (4c). (This fi gure is available in colour online)

Plasticity changes in the brain
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
Conclusions and future perspectives
This study examined the brain mechanism of hypnosis and meditation. We analysed shared and non-shared neural substrates and studied brain plasticity changes using func-tional magnetic resonance imaging (fMRI) and positron emission tomography (PET). We also used electroencephalography (EEG). The decision to choose one or the other brain imaging technique depended on the task of interest.
HypnosisHypnosis is a state of focused attention, concentration and inner absorption (Laureys et al., 2004). It is generally established by an induction procedure. Under hypnosis the imagery content is polymodal and perceptual changes may occur. As yet, the neural mechanisms underlying hypnosis remain poorly understood.
Using fMRI visual illusion effects in hypnosis were systematically investigated and analysed with Granger Causality Mapping. Perceptual illusions were accompanied by changes in brain activations in fusiform gyrus, anterior cingulate cortex and parietal areas. In a separate study, EEG was used to analyse the mechanisms of a hypnotic trance induction.
MeditationMeditation is a very heterogeneous term and it refers to a broad variety of different practices. Therefore it is not surprising that there are differences in brain function associ-ated with different types of meditation techniques. We adopted the framework by Lutz et al. (2008) and compared brain activation patterns in meditation techniques with focused attention (example: Shamatha) and open monitoring, meta-attention (example: Vipassana).
We conducted a longitudinal study on Zen meditation training with different types of assessment strategies including fMRI. Our main focus was on the process of enhanced sustained attention abilities through meditation as well as developing a mindful attitude. The experimental paradigm used in combination with fMRI was ‘binocular rivalry’ which has recently gained wider prominence as a tool for studying (visual) consciousness and focused attention. Our fi ndings suggest an enhanced capacity for selective attention after a 6-month meditation training.
Comparison of hypnosis and meditation techniquesCommon aspects and differences between hypnosis and meditation techniques were critically discussed. As a common denominator, both hypnosis and different kinds of meditation traditions emphasize attention, concentration and letting go of thoughts. One of the main purposes of both hypnosis and meditation techniques are to understand and gain control of our emotions. Both methods show benefi cial effects for stress reduction and help us to improve our immune system. But this common goal is achieved by dif-ferent means. Whereas in hypnosis a special rapport with the hypnotist and suggestions play a central role, it is independence from needing social relationships that plays a key role in meditation exercises. In addition, it was argued that hypnosis and meditation show differences in terms of sensory input, processing, memory, and sense of time. Therefore it is not surprising to fi nd brain plasticity changes in hypnosis that can be clearly differ-entiated from neurophysiological changes in meditation.
Using EEG changes in brain activity in hypnosis and Vipassana meditation these were directly compared in a within-subject design. Striking differences were found in

Halsband et al.
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
alpha and theta frequency bands. Further research is needed to disentangle plasticity changes in hypnosis and different types of meditation and to systematically compare shared and non-shard neuronal substrates in different stages of hypnotic trance induction.
One main purpose of our current studies was to try to understand different states of consciousness in terms of the subjective meanings individual subjects bring to them. Some of the fi ndings are based on single-case studies. Single case studies are a challeng-ing approach, e.g. to analyze in detail the progression of a hypnotic trance or to directly compare within the same subject the different brain activations in hypnosis and medita-tion. The principles of evidence based practise in single case studies are well established (Crombie, 1996) and accepted as a legitimate and useful method research (Klein and Myers, 1999). However, we are well aware of the fact that these fi ndings are limited and diffi cult to generalize. There are individual differences in the experience of a hypnotic trance and meditation skills which are not taken into account. Thus, further studies with a greater sample size are needed to explore the differences and commonalities of hyp-nosis and different meditation techniques.
In an integrative working programme, PET and/or fMRI could be used to measure regional activation effects in combination with neurophysiological recordings of the brain (EEG, magnetoencephalography, MEG). This ensures simultaneously a high spatial (fMRI, PET) and a high temporal resolution (EEG, MEG) in the range of milliseconds.
Another interesting future perspective is the integration of the analysis of neuro-chemical changes, e.g. stress hormones (cortisol, "-endorphin) and neurotransmitters with functional brain imaging techniques. Recently a new system was introduced (Siemens MR-PET) which is capable of performing simultaneously measurements of anatomy, functionality and biochemistry. MR-PET holds great promises for differentiat-ing the functional and biochemical basis of hypnosis and meditation.
Notes1 Nowadays it has been well established that distinct differences exist between hypnosis, sleep and
normal waking consciousness.2 The semantic meaning of shama is ‘pacifi cation’, ‘the slowing or cooling down’, or ‘rest’ (Ray,
2004).3 The recordings were carried out using a 72 channel QuickAmp EEG amplifi er system (Brainprod-
ucts GmbH, Munich, Germany) and a 64 channel electrode cap with actively shielded Ag/AgCl electrodes (ANT, Netherlands). EEG was recorded from DC to 70 Hz at a sampling rate of 250 S/s and a resolution of 0.07 microvolt. Additionally, ECG, vertical and horizontal eye movements, respiration and skin conductance from the non-dominant hand were recorded.
4 Perception of face and house images is associated with the activation of two specifi c regions. One of them, the fusiform face area (FFA) responds at least twice as strongly to faces as to other non-face stimuli. In contrast, perception of houses and places is strongly associated with a bilateral activation in the parahippocampal area (PPA).
Acknowledgements
The studies on hypnosis were supported by the Milton H. Erickson Gesellschaft für Klinische Hypnose e.V. (MEG, Germany), the Milton Erickson Stiftung (Germany), the Deutsche Gesellschaft für Zahnärztliche Hypnose e.V. (DGZH) and the Deutsche Gesell-schaft für Hypnose und Hypnotherapie e.V. (DGH) to Ulrike Halsband and Simon Strickner. The research on meditation was supported by Bial, Portugal (Grant No. 147 /

Plasticity changes in the brain
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
06) to Ulrike Halsband and Susanne Mueller and by the Samueli Institute for Information Biology (SIIB, CA, USA) and the Beckley Foundation (Oxford, UK) to Thilo Hinterberger.
ReferencesAftanas LI, Golocheikine SA (2001) Human anterior and frontal midline theta and lower alpha
refl ect emotionally positive state and internalized attention: high-resolution EEG investigation of meditation. Neuroscience Letters 310: 57–60.
Aftanas LI, Golocheikine SA (2002) Non-linear dynamic complexity of the human EEG during meditation. Neuroscience Letters 330: 143–6.
Aftanas LI, Golocheikine SA (2005) Impact of regular meditation practice on EEG activity at rest and during evoked negative emotions. International Journal of Neuroscience 115: 433–43.
Anand B, Chhina G, Singh B (1961) Some aspects of electroencephalographic studies in yogis. Electroencephalography and Clinical Neurophysiology 13: 452–6.
Bakan P, Svorad D, (1969) Resting EEG alpha and asymmetry of refl ective lateral eye movements. Nature 223: 975–6.
Banquet JP (1972) EEG and meditation. Electroencephalography and Clinical Neurophysiology 33: 454.
Banquet JP (1973) Spectral analysis of the EEG in meditation. Electroencephalography and Clini-cal Neurophysiology 35: 143–51.
Barabasz AF (1983) EEG alpha-hypnotizability correlations are not simple covariates of subject self-selection. Biological Psychology 17: 169–72.
Baron Short E, Kose S, Mu Q, Borckardt J, Newberg A, George MS, Kozel FA (2007) Regional brain activation during meditation shows time and practice effects: an exploratory fMRI study. Evidence Based Complementary and Alternative Medicine. (Epub ahead of print)
Baschek IL, Bredenkamp J, Oehrle B, Wippich W (1977) Bestimmung der Bildhaftigkeit (I), Konkretheit (C) und der Bedeutungshaltigkeit (M) von 800 Substantiven. Zeitschrift für Experimentelle und Angewandte Psychologie 24: 353–96.
Benson H, Malhotra MS, Goldman RF, Jacobs GD, Hopkins PJ (1990) Three case reports of the metabolic and electroencephalographic changes during advanced Buddhist meditation tech-niques. Behavioral Medicine 16: 90–5.
Blakemore SJ, Oakley DA, Frith CD (2003) Delusions of alien control in the normal brain. Neu-ropsychologia 41: 1058–67
Bongartz W (1985) Encoding of high- and low-imagery nouns during hypnotic age regression. Experimentelle Klinische Hypnose I: 143–51.
Cahn R, Polich J (2006) Meditation states and traits: EEG, ERP, and neuroimaging studies. Psy-chological Bulletin 132: 180–211.
Carrington P (1998) The Book of Meditation: The Complete Guide to Modern Meditation (rev ed) New York: Element Books.
Carter OL, Presti DE, Callistemon C, Ungerer Y, Liu GB, Pettigrew JD (2005) Meditation alters perceptual rivalry in tibetan buddhist monks. Current Biology 15: 412–13.
Corbetta M, Shulman GL (2002) Control of goal-directed and stimulus-driven attention in the brain. Nature Reviews Neuroscience 3: 201–15.
Corby JC, Roth WT, Zarcone VP jr, Kopell BS (1978) Psychophysiological correlates of the practice of tantric yoga meditation. Archives of General Psychiatry 35: 571–7.
Coromaldi E, Basar-Eroglu C, Stadler MA (2004) EEG-Rhythmen während tiefer Meditation: Eine Einzelfallstudie mit einem Zen-Meister. Hypnose und Kognition 21: 61–77.
Coull JT (1998) Neural correlates of attention and arousal: insights from electrophysiology, func-tional neuroimaging and psychopharmacology. Progress in Neurobiology 55: 343–61.
Craig AD (2000) How do you feel? Interoception: the sense of the physiological condition of the body. Nature Reviews Neuroscience 3: 655–66.

Halsband et al.
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
Crawford HJ (1990) Cognitive and psychophysiological correlates of hypnotic responsiveness and hypnosis. In: ML Fass, DP Brown (eds) Creative Mastery in Hypnosis and Hypnoanalysis. New York: Plenum, 47–54.
Crawford HJ, Allen SN (1996) Paired- associated learning and recall of high and low imagery words: moderating effects of hypnosis, hypnotic susceptibility level, and visualization abili-ties. American Journal of Psychology 109: 353–72.
Crawford HJ, Corby JC, Kopell BS (1996) Auditory event-related potentials while ignoring tone stimuli: attentional differences refl ected in stimulus intensity and latency responses in low and highly hypnotizable persons. International Journal of Neuroscience 85: 57–69.
Crawford HJ, Knebel T, Kaplan L, Vendemia JM, Xie M, Jamison S, Pribram KH (1998) Hypnotic analgesia: 1. Somatosensory event-related potential changes to noxious stimuli and 2. Transfer learning to reduce chronic low back pain. International Journal of Clinical and Experimental Hypnosis 46: 92–132.
Crombie IK (1996) The Pocket Guide to Critical Appraisal. London: BMJ Publishing Group.Damasio AR (2000) The Feeling of What Happens: Body and Emotion in the Making of Con-
sciousness. San Diego: Harcourt Brace & Co.De Pascalis V, Ray WJ, Tranquillo I, D Amico D (1998) EEG activity and heart rate during recall
of emotional events in hypnosis: relationships with hypnotizability and suggestibility. Inter-national Journal of Psychophysiology 29: 255–75.
Dietrich A (2003) Functional neuroanatomy of altered states of consciousness: thetransient hypo-frontality hypothesis. Consciousness and Cognition 12: 231–56.
Doraiswami PM, Xiong CL (2007) Does meditation enhance cognition and brain longevity? Annals of the New York Academy of Science [Electronic Publication ahead of print].
Edmonston WE, Grotevant WR (1975) Hypnosis and alpha density. American Journal of Clinical Hypnosis 17: 221–32.
Egner T, Jamieson G, Gruzelier J (2005) Hypnosis decouples cognitive control from confl ict monitoring processes of the frontal lobe. NeuroImage 27: 969–78.
Engstrom DR, London P, Hart JT (1970) Hypnotic susceptibility increased by EEG alpha training. Nature 227: 1261–2.
Faymonville ME, Boly M, Laureys S (2006) Functional neuroanatomy of the hypnotic state.Journal of Physiology (Paris) 99: 463–9.
Faymonville ME, Laureys S, Degueldre C, DelFiore G, Luxen A, Franck G, Lamy M, Maquet P (2000) Neural mechanisms of antinociceptive effects of hypnosis. Anesthesiology 92: 1257–67.
Faymonville ME, Mambourg PH, Joris J, Vrijens B, Fissette J, Albert A, Lamy M (1997)Psycho-logical approaches during conscious sedation. Hypnosis versus stress reducingstrategies: a prospective randomized study. Pain 73: 361–7.
Fingelkurts AA, Fingelkurts AA, Kallio S, Revonsuo A (2007) Cortex functional connectivity as a neurophysiological correlate of hypnosis: an EEG case study. Neuropsychologia 45: 1452–62.
Glueck BC, Stroebel CF (1975) Biofeedback and meditation in the treatment of psychiatric ill-nesses. Comprehensive Psychiatry 16: 303–21.
Graffi n NF, Ray WJ, Lundy R (1995) EEG concomitants of hypnosis and hypnotic susceptibility. Journal of Abnormal Psychology 104: 123–31.
Grant JA, Rainville P (2005) Hypnosis and meditation: similar experiential changes and shared brain mechanisms. Medical Hypotheses 65: 625–6.
Grond M, Pawlik G, Walter H, Lesch OM, Heiss W (1995) Hypnotic catalepsy-induced changes of regional cerebral glucose metabolism. Psychiatry Research: Neuroimaging 61: 173–9.
Gruzelier JH (2000) Redefi ning hypnosis: theory, methods and integration. Contemporary Hyp-nosis 17: 51–70.
Haggard P, Cartledge P, Meilyr D, Oakley DA (2004) Anomalous control: When ‘free-will’ is not conscious. Consciousness and Cognition 13: 646–54.

Plasticity changes in the brain
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
Halsband U (2004) Mechanismen des Lernens in Trance: funktionelle Bildgebung und Neuro-psychologie. Hypnose und Kognition 21: 11–38.
Halsband U (2006) Learning in trance: functional brain imaging studies and neuropsychology. Journal of Physiology (Paris) 99 4–6: 470–82.
Halsband U (2008) Hypnose und Meditation: Was passiert in unserem Gehirn? Suggestionen 1: 6–24.
Halsband U (2009) Neurobiologie der Hypnose. In: D Revensdorf , B Peter (eds) Hypnose in Psychotherapie, Psychosomatik und Medizin. Heidelberg: Springer.
Halsband U, Herfort A (2007) Neurobiologische Grundlagen der medizinischen Hypnose. S. Schulz-Stübner (eds) Medizinische Hypnose, Schattauer, Chapter 2: 7–38.
Halsband U, Krause BJ, Schmidt D, Herzog H, Tellmann L, Müller-Gärtner HW (1998) Encoding and retrieval in declarative learning: a positron emission tomography study. Behavioural Brain Research 97: 69–78.
Halsband U, Krause BJ, Sipilä H, Teräs M, Laihinen A (2002) PET studies on the memory pro-cessing of word pairs in bilingual Finnish-English subjects. Behavioural Brain Research 132: 47–57.
Heap M, Aravind KK (2002) Hartland’s Medical and Dental Hypnosis, 4th edn. Edinburgh: Churchill Livingston.
Herzog H, Lele VR, Kuwert T, Langen K-J, Kops ER, Feinendegen LE (1990) Changed pattern of regional glucose metabolism during yoga meditative relaxation. Neuropsychobiology 23: 182–7.
Hinterberger T, Halsband U (to be submitted) Hypnotic trance induction: an EEG study.Hoelzel BK, Ott U, Hempel H, Hackl A, Wolf K, Stark R, Vaitl D (2007) Differential engagement
of anterior cingulate and adjacent medial frontal cortex in adept meditators and non-medita-tors. Neuroscience Letters 421: 16–21.
Hoelzel BK, Ott U, Gard T, Hempel H, Weygandt M, Morgen K, Vaitl D (2008) Investigation of mindfulness meditation practitioners with voxel-based morphometry. Social Cognitive andAffective Neuroscience 3: 55–61.
Holroyd J (2003) The science of meditation and the state of hypnosis. American Journal of Clini-cal Hypnosis 46(2): 109–28.
Isotani T, Lehmann D, Pascual-Marqui RD, Kochi K, Wackermann J, Saito N, Yagyu T, Kinoshita T, Sasada K (2001) EEG source localization and global dimensional complexity in high- and low-hypnotizable subjects: a pilot study. Neuropsychobiology 44: 192–8.
Jamieson GA, Dwivedi P, Gruzelier JH (2005) Changes in mismatch negativity across pre- hypnosis, hypnosis and post-hypnosis conditions distinguish high from low hypnotic sus-ceptibility groups. Brain Research Bulletin 67: 298–303.
Jevning R, Wallace RK, Beidebach M (1992) The physiology of meditation: a review. A wakeful hypometabolic integrated response. Neuroscience and Biobehavioral Reviews 16: 415–24.
Jha AP, Krompinger J, Baime MJ (2007) Mindfulness training modifi es subsystems of attention. Cognitive Affective and Behavioral Neuroscience 7: 109–19.
Kastner S, Pinsk MA, De Weerd P, Desimone R, Ungerleider LG (1999) Increased activity in human visual cortex during directed attention in the absence of visual stimulation. Neuron 22: 751–61.
Katayama H, Gianotti LR, Isotani T, Faber PL, Sasada K, Kinoshita T, Lehmann D (2007) Classes of multichannel EEG microstates in light and deep hypnotic conditions. Brain Topography 20(1): 7–14.
Kesterson J (1989) Metabolic rate, respiratory exchange ratio and apneas during meditation. American Journal of Physiology R256: 632–8.
Kirsch I, Lynn SJ (1997) Hypnotic involuntariness and the automaticity of everyday life. Ameri-can Journal of Clinical Hypnosis 40: 329–48.
Kjaer TW, Bertelsen C, Piccini P, Brooks D, Alving J, Lou HC (2002) Increased dopamine tone during meditation-induced change of consciousness. Brain Research and Cognition 13: 255–9.

Halsband et al.
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
Klein H and Myers M (1999) A set of principles for conducting and evaluating interpretive fi eld studies in information systems. MIS Quarterly 23(1): 67–93.
Klimesch W (1999) EEG alpha and theta oscillations refl ect cognitive and memory performance: a review and analysis. Brain Research. Brain Research Reviews 29: 169–95.
Konradt B, Deeb S, Scholz OB (2005) Motor imagery in hypnosis: accuracy and duration of motor imagery in waking and hypnotic state. International Journal of Clinical and Experi-mental Hypnosis 53(2): 148–69.
Kosslyn SM, Thompson WL, Costantini–Ferrando MF, Alpert NM, Spiegel D (2000) Hypnotic visual illusion alters color processing in the brain. American Journal of Psychiatry 157(8): 1279–84.
Krause BJ, Horwitz B, Taylor JG, Schmidt D, Mottaghy FM, Herzog H, Halsband U, Müller-Gärtner H (1999a) Network analysis in episodic encoding and retrieval of word-pair associates: a PET study. European Journal of Neuroscience 11: 3293–301.
Krause BJ, Schmidt D, Mottaghy FM, Taylor J, Halsband U, Herzog H, Tellmann L, Müller-Gärtner HW (1999b) Episodic retrieval activates the precuneus irrespective of the imagery content of word pair associates. A PET study. Brain 122: 255–63.
Kubota Y, Sato W, Toichi M, Murai T, Okada T, Hayashi A, Sengoku A (2001) Frontal midline theta rhythm is correlated with cardiac autonomic activities during the perfor-mance of an attention demanding meditation procedure. Cognitive Brain Research 11: 281–7.
Laureys S (2005) The neural correlate of (un)awareness: lessons from the vegetative state. Trends in Cognitive Sciences 9: 556–9.
Laureys S, Maquet P, Faymonville ME (2004) Brain function druing hypnosis. In: A Otte, K Audenaert, K Peremans, K van Heeringen, RA Dierckx (eds) Nuclear Medicine in Psychiatry. Berlin: Springer-Verlag, 507–19.
Lazar SW, Bush G, Gollub RL, Fricchione GL, Khalsa G, Benson H (2000) Functional brain mapping of the relaxation response and meditation. Neuroreport 11: 1581–5.
Lazar SW, Kerr CE, Wasserman RH, Gray JR, Greve DN, Treadway MT, McGarvey M, Quinn BT, Dusek JA, Benson H, Rauch SL, Moore CI, Fischl B (2005) Meditation experience is associated with increased cortical thickness. Neuroreport 16: 1893–7.
Logothetis NK, Pauls J, Augath M, Trinath T, Oeltermann A (2001) Neurophysiological investiga-tion of the basis of the fMRI signal. Nature 412: 150–7.
London P, Hart JT, Leibovitz MP (1968) EEG alpha rhythms and susceptibility to hypnosis. Nature 219: 71–2.
Lou HC, Kjaer TW, Friberg L, Wildschiodtz G, Holm S, Nowak M (1999) A 150-H20 PET study of meditation and the resting state of normal consciousness. Human Brain Mapping 7: 98–105.
Luders E, Toga AW, Lepore N, Glaser C (2009) The underlying and atomical correlates of long-term meditation: larger hippocampal and frontal volumes of gray matter. Neuroimage 45: 672–8.
Lundstrom BN, Petersson KM, Andersson J, Johansson M, Fransson P, Ingvar M (2003) Isolating the retrieval of imagined pictures during episodic memory: activation of the left precuneus and left prefrontal cortex. NeuroImage 20: 1934–43.
Lutz A, Dunne JD, Davidson RJ (2007) Meditation and the neuroscience of consciousness: an introduction. In: P Zelazo, M Moscovitch, E Thompson (eds) Cambridge Handbook of Con-sciousness. Cambridge: Cambridge University Press, 499–554.
Lutz A, Greischar LL, Rawlings NB, Ricard M, Davidson RJ (2004) Long-term meditators self-induce high-amplitude gamma synchrony during mental practice. Proceedings of the National Academy of Sciences USA 101: 16369–73.
Lutz A, Slagter HA, Dunne JD, Davidson RJ (2008) Attention regulation and monitoring in meditation. Trends in Cognitive Science 12: 163–9.
Maquet P, Faymonville ME, Degueldre C, Delfi ore G, Franck G, Luxen A, Lamy M (1999) Func-tional neuroanatomy of hypnotic state. Biological Psychiatry 45: 327–33.
Meier H (1964) Deutsche Sprachstatistik. Hildesheim: Georg Ohms-Verlag.

Plasticity changes in the brain
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
Meng M, Tong F (2004) Can attention selectively bias bistable perception? Differences between binocular rivalry and ambiguous fi gures. Journal of Vision 4: 539–51.
Morgan AH, MacDonald H, Hilgard ER (1974) EEG alpha: lateral asymmetry related to task and hypnotizability. Psychophysiology 11: 275–82.
Mottaghy FM, Krause BJ, Schmidt D, Hautzel H, Herzog H, Shah NJ, Halsband U, Müller-Gärtner HW (2000) Comparison of PET and fMRI activation patterns during declarative memory processes. Nuklearmedizin 39: 196–203.
Mottaghy FM, Shah NJ, Krause BJ, Schmidt D, Halsband U, Jäncke L, Müller-Gärtner HW (1999) Neuronal correlates of encoding and retrieval in episodic memory during a paired word association learning task: a functional magnetic resonance imaging study. Experimental Brain Research 128: 332–42.
Mueller S, Halsband U (to be submitted) Cognitive and affective trait effects of intensive zen-meditaion training on brain and behaviour. An event-related longitudinal fMRI study.
Muzur A (2006) Hypnosis and modern frontal-lobe concepts – a sketch for a review and an invi-tation to one particularly promising fi eld. Collegium Antropologicum 30: 205–211.
Newberg AB, Alavi A, Baime M, Pourdehnad M, Santanna J, d Aquili E (2001) The measurement of regional cerebral blood fl ow during the complex cognitive task of meditation. A preliminary SPECT study. Psychiatry Research 106: 113–22.
Newberg AB, Iversen J (2003) The neural basis of the complex mental task of meditation: neurotransmitter and neurochemical considerations. Medical Hypotheses 61: 282–91.
Oakley DA (1999) Hypnosis and conversion hysteria: a unifying model. Cognitive Neuropsychia-try 4: 243–65.
Otani A (2003) Eastern meditative techniques and hypnosis: a new synthesis. American Journal of Clinical Hypnosis 46: 97–108.
Otte A, Halsband U (2006) Brain imaging tools in neurosciences. Journal of Physiology (Paris) 99: 281–92.
Otto T (2007) Effective connectivity changes in hypnotic visual illusion. Master Thesis, Univer-sity of Maastricht. A study carried out by Otto T, Halsband U, Goebel R.
Pan W, Zhang L, Xia Y (1994) The difference in EEG theta waves between concentrative and non-concentrative qigong states – a power spectrum and topographic mapping study. Journal of Traditional Chinese Medicine 14: 212–18.
Perlini AH, Spanos NP (1991) EEG alpha methodologies and hypnotizability: a critical review. Psychophysiology 28: 511–30.
Peter B (2009) Hypnose und die Konstruktion von Wirklichkeit. In: D Revensdorf, B Peter (eds) Hypnose in Psychotherapie, Psychosomatik und Medizin. Ein Manual für die Praxis. Heidel-berg: Springer, 32–40.
Rainville P, Hofbauer RK, Bushnell MC, Duncan GH, Price DD (2002) Hypnosis modulates activity in brain structures involved in the regulation of consciousness. Journal of Cognitive Neuroscience 14: 887–901.
Rainville P, Hofbauer RK, Paus T, Duncan GH, Bushnell MC, Price DD (1999) Cerebral mecha-nisms of hypnotic induction and suggestion. Journal of Cognitive Neuroscience 11: 110–25.
Ray, Reginald A. (ed.) (2004) In the Presence of Masters: Wisdom from 30 Contemporary Tibetan Buddhist Teachers. Boston, Massachusetts: Shambhala.
Raz A, Fan J, Posner MI (2006) Neuroimaging and genetic associations of attentional and hyp-notic processes. Journal of Physiology (Paris). 99(4–6): 483–91.
Raz A, Shapiro T (2002) Hypnosis and neuroscience: cross talk between clinical and cognitive research. Archives of General Psychiatry 59: 85–90.
Revenstorf D (1996) Klinische Hypnose – Gegenwärtiger Stand der Theorie und Empirie. Psy-chotherapie, Psychosomatik und Medizinische Psychologie 49: 5–13.
Roebroeck A, Formisano E, Goebel R (2005) Mapping directed infl uence over the brain using Granger causality and fMRI. NeuroImage 25: 230–42.
Sabourin ME, Cutcomb SD, Crawford HJ, Pribram K (1990) EEG correlates of hypnotic suscep-tibility and hypnotic trance: spectral analysis and coherence. International Journal of Psycho-physiology 10: 125–42.

Halsband et al.
Copyright © 2009 British Society of Experimental & Clinical Hypnosis Contemp. Hypnosis (2009)Published by John Wiley & Sons, Ltd DOI: 10.1002/ch
Schall U, Johnston P, Lagopoulos J, Juptner M, Jentzen W, Thienel R, Dittmann-Balcar A, Bender S, Ward PB (2003) Functional brain maps of Tower of London performance: positron emission tomography and functional magnetic resonance imaging study. NeuroImage 20: 1154–61.
Schmidt D, Krause BJ, Mottaghy FM, Halsband U, Herzog H, Tellmann L, Müller-Gärtner HW (2002) Brain systems engaged in encoding and retrieval of word-pair associates independent of their imagery content or presentation modalities. Neuropsychologia 40: 457–70.
Schmitter S, Diesch E, Amann M, Kroll A, Moayer M, Schad LR (2008) Silent echo-planar imaging for auditory fMRI. Magnetic Resonance Materials in Physics, Biology and Medicine 21: 317–25.
Schnyer DM, Allen JJ (1995) Attention-related electroencephalographic and event-related poten-tial predictors of responsiveness to suggested posthypnotic amnesia. International Journal of Clinical and Experimental Hypnosis 43: 295–315.
Slagter HA, Lutz A, Greischar LL, Francis AD, Nieuwenhuis S, Davis JM and Davidson RJ (2007) Mental training affects distribution of limited brain resources. PLoS Biology 5, e138.
Spiegel D, Kosslyn SM (2004) Glauben ist Sehen: Die Neurophysiologie der Hypnose. Hypnose und Kognition 21(1–2): 119–37.
Srinivasan N, Baijal S, (2007) Concentrative meditation enhances preattentive processing: a mismatch negativity study. Neuroreport 18: 1709–12.
Sudsuang R, Chentanez V, Veluvan K (1991) Effects of buddhist meditation on serum cortisol and total protein levels, blood pressure, pulse rate, lung volume and reaction time. Physiology and Behavior 50: 543–8.
Szechtman H, Woody E, Bowers KS, Nahmias C (1998) Where the imaginal appears real: a posi-tron emission tomography study. Proceedings of the National Academy of Sciences USA 95: 1956–60.
Tart CT (1972) States of consciousness and state-specifi c sciences. Science 176: 1203–10.Tart CT (2000) Investigating altered states of consciousness on their own terms: a proposal for
the creation of state-specifi c sciences. International Journal of Parapsychology 11(1): 7–41.Tart CT (2001) Meditation: some kind of (self)hypnosis? A deeper look. Speech at the 109th
Annual Convention of the American Psychological Association at San Francisco, August.Tebecis AK, Provins KA, Farnbach RW, Pentony P (1975) Hypnosis and the EEG: a quantitative
investigation. Journal of Nervous and Mental Disease 161: 1–17.Tong F, Nakayama K, Vaughan JT, Kanwisher N (1998) Binocular rivalry and visual awareness
in human extrastriate cortex. Neuron 21: 753–9.Ulett GA, Akpinar S, Itil TM (1972) Hypnosis: physiological, pharmacological reality. American
Journal of Psychiatry 128: 799–805.Vestergaard-Poulsen P, van Beek M, Skewes J, Bjarkam CR, Stubberup M, Bertelsen J, Roep-
storff A (2009) Long-term meditation is associated with increased gray matter density in the brain stem. Neuroreport 20: 170–4.
Wallace RK (1970) Physiological effects of transcendental meditation. Science 167: 1751–4.Wallace RK, Benson H (1972) The physiology of meditation. Scientifi c American 226: 85–90.Weitzenhoffer AM (2000) The Practice of Hypnotism, 2nd edn. New York: Wiley.
Address for correspondence:Prof. Dr Ulrike Halsband (D. Phil., Oxon)NeuropsychologyDepartment of PsychologyUniversity of FreiburgEngelbergerstr. 41D-79106 FreiburgGermanyTel: +49-761-203-2475 /-2473 (offi ce)Email: [email protected]





























