Embed Size (px)
Citation preview

Neuroscience Letters 371 (2004) 18–23
Perceptual learning modulates electrophysiological and psychophysicalresponse to visual texture segmentation in humans
Clara Cascoa,∗, Gianluca Campanaa, Alba Griecoa, Giorgio Fuggettab
a Dipartimento di Psicologia Generale, Universit`a di Padova, Via Venezia, 8, 35131 Padova, Italyb Dipartimento di Scienze Neurologiche e della Visione, Universit`a di Verona, Italy
Received 14 April 2004; received in revised form 2 July 2004; accepted 2 August 2004
Abstract
We investigated the mechanisms that allow, via perceptual learning, selective modulation of a visual line-texture figure saliency in accordancewith task relevance. Learning-dependent saliency increase was inferred by increased accuracy in orientation discrimination with task repetition.As a result of learning, accuracy increase was more pronounced when local and global orientation of the texture figure conflicted, and reachedceiling in both conflict and conflict-free conditions. This psychophysical effect was associated with a decrease in amplitude of negative VEPc componenti t channels,a©
K
LsspiHevltloshdcpv
tory
g thesherlas-ed
tednce
d bye incor-ith aeasemea-itherthe
romy as-ctly
0d
omponents in the configurations where global and local orientation conflicted, and to a weak increase of VEP’s earliest negativen the conflict-free condition. The VEP result is a direct demonstration that learning, in addition to increasing response of relevanlso reduces the weight of channels whose receptive field size and orientation tuning conflict with the task.2004 Elsevier Ireland Ltd. All rights reserved.
eywords:Perceptual learning; Textures; VEPs; Psychophysics
ittle is known about the mechanisms that allow the visualystem to increase the salience of behaviourally importanttimuli and to decrease salience of irrelevant stimuli. Somehysiological evidence in both audition[13] and vision[6]
ndicates that receptive field tuning can be modulated via aebbian learning mechanism. In line with these studies, wexplored the mechanisms that allow implicit processing ofisual information in humans to be modulated by perceptualearning according to task relevance. The processes inves-igated are those involved in figure-ground segmentation ofine-texture stimuli, on the basis of local orientation contrastr global orientation. This is a well-suited stimulus for as-essing how saliency is modulated by learning, because itas been shown that saliency of local and global orientationepends on orientation contrast[18] and is increased by per-eptual learning[12]. Here we asked how global and localrocesses are modulated by learning depending on task rele-ance. In accordance with Hebb’s rule[10], learning produces
∗ Corresponding author. Tel.: +39 0498276611; fax: +39 0498276600.E-mail address:[email protected] (C. Casco).
long-lasting regulation of visual responses, either excitaor inhibitory, and furthermore, it has been proposed[1] thatlearning can have the effect of increasing or decreasinstrength of inhibitory connections amongst channels. Doand Lu[5] suggest that perceptual learning might reflect pticity in the relative activity of different visual channels tunto orientation and spatial frequency. Both facilitatory[5,8]and inhibitory[16] effects of learning are well-documenpsychophysically, but no direct electrophysiological evideof inhibitory effect of learning has yet been provided.
Here we asked not only whether saliency is increaselearning from trial repetition, but also whether an increassaliency is associated with an increase in amplitude oftical response (VEPs) to relevant texture features and wdecrease for irrelevant ones. The effect of saliency incrdue to trial repetition was psychophysically assessed bysuring accuracy in an orientation discrimination task, eglobal or local, as a function of block number. Indeed,saliency of a single or group of elements emerging fan array of texture elements can be psychophysicallsessed either directly from subjective reports, or indire
304-3940/$ – see front matter © 2004 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.neulet.2004.08.005

C. Casco et al. / Neuroscience Letters 371 (2004) 18–23 19
Fig. 1. Stimuli. Texture consisted of white vertical line elements 19′ longarranged on a diamond raster, with raster step of 30.5′ and jittered aroundtheir raster centre by 0–2.7′. The segmented texture bar comprised 6× 24line elements tilted 45◦ clockwise or counterclockwise at random. In theparallel condition (a) the bar had the same orientation as its line elements,while in the orthogonal configuration (b) it had opposite orientation.
by measuring speed and accuracy of detection or discrimina-tion [18].
We based the assessment of neural correlates of the ef-fect of perceptual learning on the evidence that variationsin D-VEP amplitudes (obtained as the difference betweenuniform- and segmentation-texture VEPs) are specifically as-sociated with texture segmentation[12,14].
Observers viewed line-texture bars, oriented in either thesame (parallel,Fig. 1a) or opposite (orthogonal,Fig. 1b) di-rection to their elements segregating out from a uniform tex-ture background. Separate groups performed a 2AFC taskin which they had to discriminate either the global (bar) orlocal (elements) orientation. When parallel, both global andlocal orientations are relevant to task, whereas when orthog-onal either local or global conflict, depending on the task.Perceptual learning was inferred from block-dependent vari-ation of both D-VEP early components and percentage cor-rect responses for the various blocks of trials in independentgroups. To analyse the effect of perceptual learning, we first
considered the possibility that observers were not blind toorientation conflicting with that explicitly judged (either lo-cal or global). This was indeed the case, since accuracy waslower when texture elements were orthogonal (conflicting) toglobal bar, an effect that disappeared with stimulus repetition.We then asked whether the improvement in accuracy withstimulus repetition was associated with a modulation of therelated D-VEP amplitude and whether this modulation wasin the same or in a different direction in the two conditions.Our results showed that increased accuracy with perceptuallearning was associated with opposite electrophysiologicaleffects depending on the configuration: an amplitude increasein conflict-free configurations, suggesting facilitation, and adecrease in the conflict configuration, suggesting inhibitionof mechanisms with receptive field size and orientation tun-ing conflicting with the task.
Four groups of 10 observers, aged 20–30 years, selected onthe basis of absence of astigmatism, participated voluntarilyin the experiments after approval by the human subject reviewboard of the University of Padova and in accordance with theHelsinki Declaration.
Different groups of observers had to be used in differenttasks of both psychophysical and VEP experiments to re-duce possible artefacts due to generalisation of learning[2].Two-way ANOVA, with block and stimulus configurationsa al ande
me ark-ew -p round( thes r bothve
an-d nalt
ockso ames whichw fromt 6 ms(
twok mis-s onses.
tri-a ringt of thec achp d asa theu ures
s main factors were used to analyse the psychophysiclectrophysiological data.
Stimuli were generated by a PC, displayed on a 15′′ colouronitor (70 Hz vertical refresh) viewed through a 16◦ diam-ter circular aperture from a distance of 57 cm in a dned room. The resolution of the monitor was 640× 480ith square pixel 2.7× 2.7 arc min. Textures were comosed of white line elements, presented on a dark backg0.6 cd/m2). The look-up table was set in such a way thatpace average luminance of the texture was matched foertical (11.45 cd/m2), and 45◦ and 135◦ (11.51 cd/m2) ori-ntation of the texture line elements.
In each block of trials, four stimulus conditions were romly interleaved: bar-oriented right (parallel or orthogo
o its elements) or left (parallel or orthogonal).The psychophysical experiments consisted of five bl
f 64 trials each, performed consecutively during the session. Each trial consisted of a three-frame sequence,as perceived as a texture bar appearing or disappearing
he uniform texture background. Frame duration was 2with 2977 ms interval).
Observers performed a 2AFC task every trial by usingeys. An acoustic feedback was given for errors and oions. Performance was defined as percent correct resp
The VEP experiment consisted of two blocks of 200ls. The two blocks were performed consecutively du
he same session. Onset–offset stimulation consistedyclical alternation of segmented and uniform texture, eresented for 840 ms with no interval, which was perceivetexture bar cyclically appearing and disappearing fromniform texture background. Note that these long expos
20 C. Casco et al. / Neuroscience Letters 371 (2004) 18–23
Fig. 2. Trial events. Events in a single trial consisted in the alternation of thesegmented (parallel or orthogonal bar, tilted clockwise or counterclockwise)and uniform texture. During the blank interval between trials, presented ei-ther every trial (in the psychophysical experiment) or every three trials onaverage (in the VEP experiments), the subject was asked to discriminate ei-ther the local orientation gradient or the global bar while maintaining fixationon a central dot.
allowed to obtain VEPs with sufficiently large amplitudes,so that all components (included those related to texture seg-mentation) could be easily seen on VEP traces, if present[3].However, such long exposure times were inadequate for psy-chophysical testing since they produced ceiling performancein both configurations and tasks.
Individual D-VEPs were determined by algebraic sub-traction of the background-stimulus VEP from eachsegmentation-stimulus VEP; the four D-VEP peaks (N1, P1,N2, N3) were identified as the largest peak within the ap-propriate temporal window (60–90, 91–120, 121–160, and161–240 ms) (Fig. 2). After presentation of three texture bar-uniform texture pairs on average, the onset–offset stimulationwas suspended and the monitor remained dark until responsewas given. The observer’s response re-started the onset–offsestimulation, after 2000 ms of a uniform texture display to pre-pare fixation.
The electroencephalogram (EEG) was recorded fromAg/AgCl-coated cup electrodes placed at Oz and left (ref-erence) and right (ground) earlobes, in accordance with theinternational 10/20 system. Electrode impedance was heldbelow 5 k�. The EEG was amplified (BM 623) and digitallyconverted (CED 1401) under control of a second PC. Stimu-lation and recording onset were synchronised using the ver-tical retrace signal of the monitor that displaced the stimulus.T d at1 ands linew
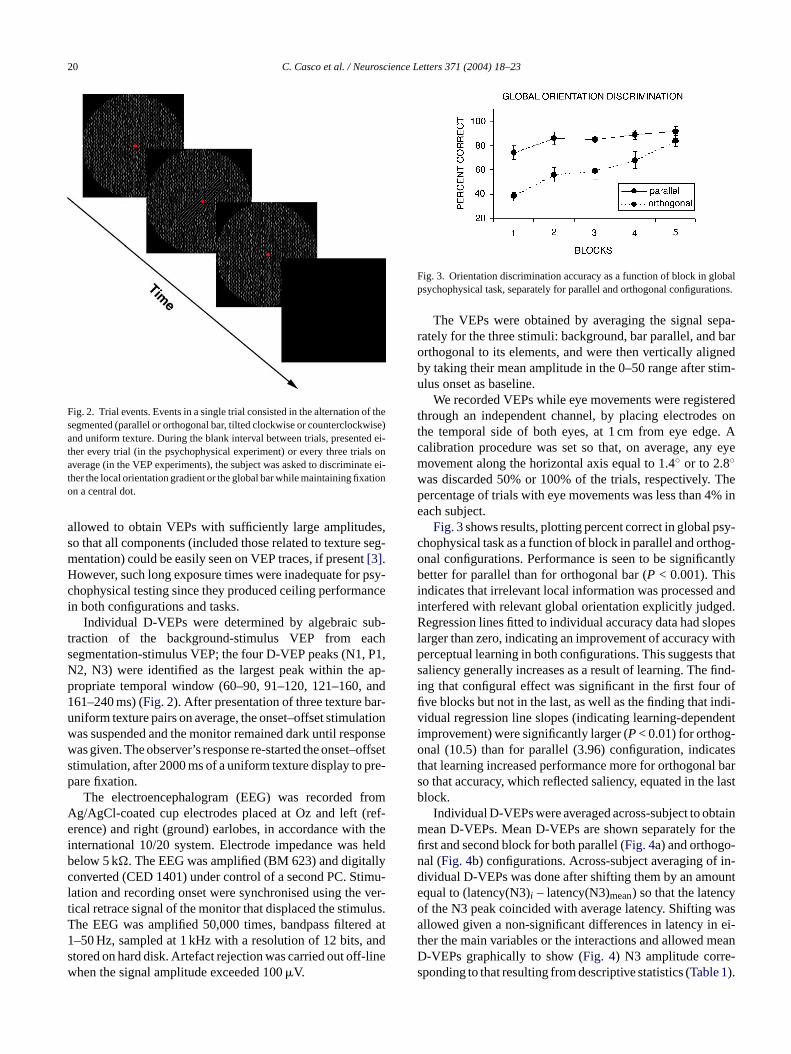
Fig. 3. Orientation discrimination accuracy as a function of block in globalpsychophysical task, separately for parallel and orthogonal configurations.
The VEPs were obtained by averaging the signal sepa-rately for the three stimuli: background, bar parallel, and barorthogonal to its elements, and were then vertically alignedby taking their mean amplitude in the 0–50 range after stim-ulus onset as baseline.
We recorded VEPs while eye movements were registeredthrough an independent channel, by placing electrodes onthe temporal side of both eyes, at 1 cm from eye edge. Acalibration procedure was set so that, on average, any eyemovement along the horizontal axis equal to 1.4◦ or to 2.8◦was discarded 50% or 100% of the trials, respectively. Thepercentage of trials with eye movements was less than 4% ineach subject.
Fig. 3shows results, plotting percent correct in global psy-chophysical task as a function of block in parallel and orthog-onal configurations. Performance is seen to be significantlybetter for parallel than for orthogonal bar (P < 0.001). Thisindicates that irrelevant local information was processed andinterfered with relevant global orientation explicitly judged.Regression lines fitted to individual accuracy data had slopeslarger than zero, indicating an improvement of accuracy withperceptual learning in both configurations. This suggests thatsaliency generally increases as a result of learning. The find-ing that configural effect was significant in the first four offive blocks but not in the last, as well as the finding that indi-v denti -o test l bars e lastb
btainm r thefi -n in-d unte yo wasa ei-t eanD -s
he EEG was amplified 50,000 times, bandpass filtere–50 Hz, sampled at 1 kHz with a resolution of 12 bits,tored on hard disk. Artefact rejection was carried out off-hen the signal amplitude exceeded 100�V.
t
idual regression line slopes (indicating learning-depenmprovement) were significantly larger (P< 0.01) for orthognal (10.5) than for parallel (3.96) configuration, indica
hat learning increased performance more for orthogonao that accuracy, which reflected saliency, equated in thlock.
Individual D-VEPs were averaged across-subject to oean D-VEPs. Mean D-VEPs are shown separately fo
rst and second block for both parallel (Fig. 4a) and orthogoal (Fig. 4b) configurations. Across-subject averaging ofividual D-VEPs was done after shifting them by an amoqual to (latency(N3)i – latency(N3)mean) so that the latencf the N3 peak coincided with average latency. Shiftingllowed given a non-significant differences in latency in
her the main variables or the interactions and allowed m-VEPs graphically to show (Fig. 4) N3 amplitude correponding to that resulting from descriptive statistics (Table 1).
C. Casco et al. / Neuroscience Letters 371 (2004) 18–23 21
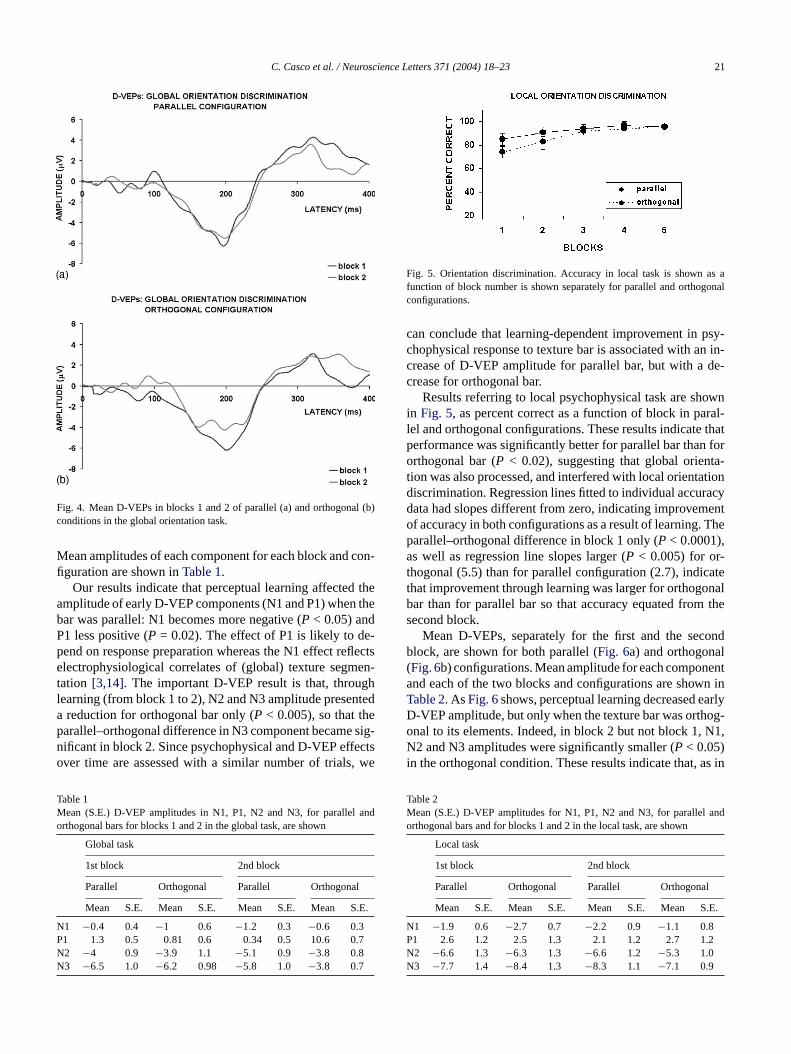
Fig. 4. Mean D-VEPs in blocks 1 and 2 of parallel (a) and orthogonal (b)conditions in the global orientation task.
Mean amplitudes of each component for each block and con-figuration are shown inTable 1.
Our results indicate that perceptual learning affected theamplitude of early D-VEP components (N1 and P1) when thebar was parallel: N1 becomes more negative (P < 0.05) andP1 less positive (P = 0.02). The effect of P1 is likely to de-pend on response preparation whereas the N1 effect reflectselectrophysiological correlates of (global) texture segmen-tation [3,14]. The important D-VEP result is that, throughlearning (from block 1 to 2), N2 and N3 amplitude presenteda reduction for orthogonal bar only (P < 0.005), so that theparallel–orthogonal difference in N3 component became sig-nificant in block 2. Since psychophysical and D-VEP effectsover time are assessed with a similar number of trials, we
Table 1Mean (S.E.) D-VEP amplitudes in N1, P1, N2 and N3, for parallel andorthogonal bars for blocks 1 and 2 in the global task, are shown
Global task
1st block 2nd block
Parallel Orthogonal Parallel Orthogonal
Mean S.E. Mean S.E. Mean S.E. Mean S.E.
N1 −0.4 0.4 −1 0.6 −1.2 0.3 −0.6 0.3P1 1.3 0.5 0.81 0.6 0.34 0.5 10.6 0.7NN
Fig. 5. Orientation discrimination. Accuracy in local task is shown as afunction of block number is shown separately for parallel and orthogonalconfigurations.
can conclude that learning-dependent improvement in psy-chophysical response to texture bar is associated with an in-crease of D-VEP amplitude for parallel bar, but with a de-crease for orthogonal bar.
Results referring to local psychophysical task are shownin Fig. 5, as percent correct as a function of block in paral-lel and orthogonal configurations. These results indicate thatperformance was significantly better for parallel bar than fororthogonal bar (P < 0.02), suggesting that global orienta-tion was also processed, and interfered with local orientationdiscrimination. Regression lines fitted to individual accuracydata had slopes different from zero, indicating improvementof accuracy in both configurations as a result of learning. Theparallel–orthogonal difference in block 1 only (P < 0.0001),as well as regression line slopes larger (P < 0.005) for or-thogonal (5.5) than for parallel configuration (2.7), indicatethat improvement through learning was larger for orthogonalbar than for parallel bar so that accuracy equated from thesecond block.
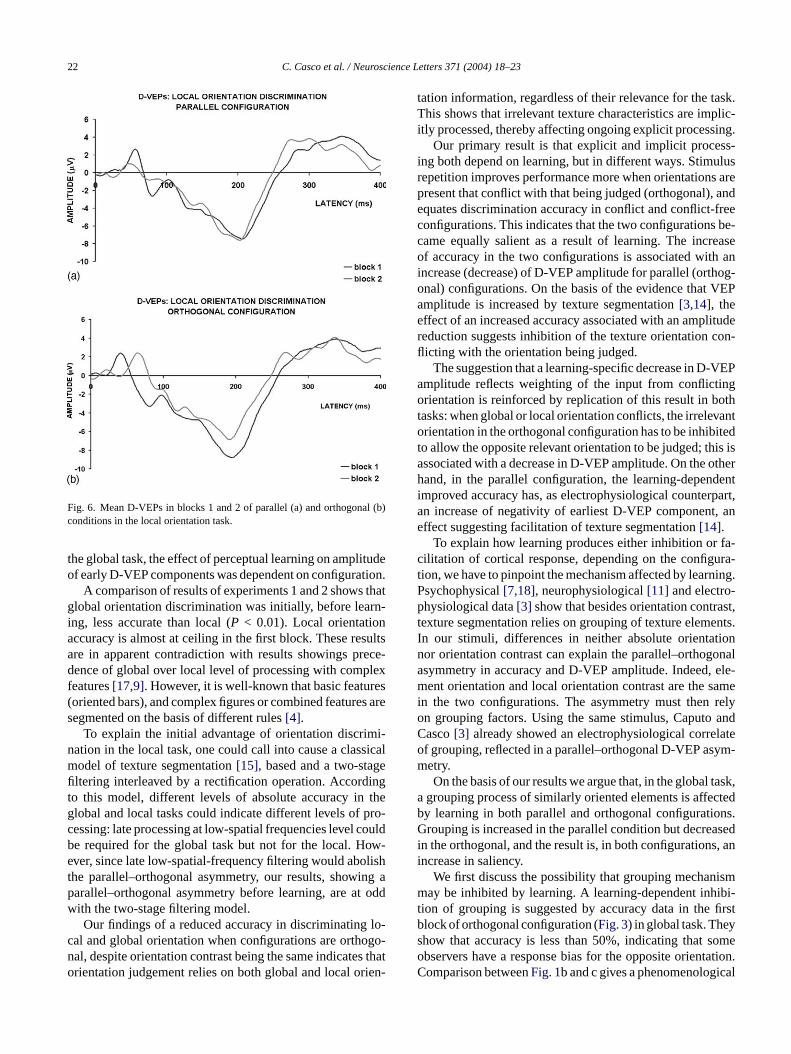
Mean D-VEPs, separately for the first and the secondblock, are shown for both parallel (Fig. 6a) and orthogonal(Fig. 6b) configurations. Mean amplitude for each componentand each of the two blocks and configurations are shown inTable 2. AsFig. 6shows, perceptual learning decreased earlyD-VEP amplitude, but only when the texture bar was orthog-o N1,Ni as in
TM ando
E.
NPNN
2 −4 0.9 −3.9 1.1 −5.1 0.9 −3.8 0.83 −6.5 1.0 −6.2 0.98 −5.8 1.0 −3.8 0.7
nal to its elements. Indeed, in block 2 but not block 1,2 and N3 amplitudes were significantly smaller (P < 0.05)
n the orthogonal condition. These results indicate that,
able 2ean (S.E.) D-VEP amplitudes for N1, P1, N2 and N3, for parallelrthogonal bars and for blocks 1 and 2 in the local task, are shown
Local task
1st block 2nd block
Parallel Orthogonal Parallel Orthogonal
Mean S.E. Mean S.E. Mean S.E. Mean S.
1 −1.9 0.6 −2.7 0.7 −2.2 0.9 −1.1 0.81 2.6 1.2 2.5 1.3 2.1 1.2 2.7 1.22 −6.6 1.3 −6.3 1.3 −6.6 1.2 −5.3 1.03 −7.7 1.4 −8.4 1.3 −8.3 1.1 −7.1 0.9
22 C. Casco et al. / Neuroscience Letters 371 (2004) 18–23
Fig. 6. Mean D-VEPs in blocks 1 and 2 of parallel (a) and orthogonal (b)conditions in the local orientation task.
the global task, the effect of perceptual learning on amplitudeof early D-VEP components was dependent on configuration.
A comparison of results of experiments 1 and 2 shows thatglobal orientation discrimination was initially, before learn-ing, less accurate than local (P < 0.01). Local orientationaccuracy is almost at ceiling in the first block. These resultsare in apparent contradiction with results showings prece-dence of global over local level of processing with complexfeatures[17,9]. However, it is well-known that basic features(oriented bars), and complex figures or combined features aresegmented on the basis of different rules[4].
To explain the initial advantage of orientation discrimi-nation in the local task, one could call into cause a classicalmodel of texture segmentation[15], based and a two-stagefiltering interleaved by a rectification operation. Accordingto this model, different levels of absolute accuracy in theglobal and local tasks could indicate different levels of pro-cessing: late processing at low-spatial frequencies level couldbe required for the global task but not for the local. How-ever, since late low-spatial-frequency filtering would abolishthe parallel–orthogonal asymmetry, our results, showing aparallel–orthogonal asymmetry before learning, are at oddwith the two-stage filtering model.
Our findings of a reduced accuracy in discriminating lo-cal and global orientation when configurations are orthogo-n s thato ien-
tation information, regardless of their relevance for the task.This shows that irrelevant texture characteristics are implic-itly processed, thereby affecting ongoing explicit processing.
Our primary result is that explicit and implicit process-ing both depend on learning, but in different ways. Stimulusrepetition improves performance more when orientations arepresent that conflict with that being judged (orthogonal), andequates discrimination accuracy in conflict and conflict-freeconfigurations. This indicates that the two configurations be-came equally salient as a result of learning. The increaseof accuracy in the two configurations is associated with anincrease (decrease) of D-VEP amplitude for parallel (orthog-onal) configurations. On the basis of the evidence that VEPamplitude is increased by texture segmentation[3,14], theeffect of an increased accuracy associated with an amplitudereduction suggests inhibition of the texture orientation con-flicting with the orientation being judged.
The suggestion that a learning-specific decrease in D-VEPamplitude reflects weighting of the input from conflictingorientation is reinforced by replication of this result in bothtasks: when global or local orientation conflicts, the irrelevantorientation in the orthogonal configuration has to be inhibitedto allow the opposite relevant orientation to be judged; this isassociated with a decrease in D-VEP amplitude. On the otherhand, in the parallel configuration, the learning-dependenti rpart,a t, ane
fa-c ura-t ing.P -p ast,t ents.I tionn onala ele-m amei relyo andC lateo ym-m
task,a ctedb ns.G asedi s, ani
ismm ibi-t firstb ys omeo ation.C cal
al, despite orientation contrast being the same indicaterientation judgement relies on both global and local or
mproved accuracy has, as electrophysiological counten increase of negativity of earliest D-VEP componenffect suggesting facilitation of texture segmentation[14].
To explain how learning produces either inhibition orilitation of cortical response, depending on the configion, we have to pinpoint the mechanism affected by learnsychophysical[7,18], neurophysiological[11] and electrohysiological data[3] show that besides orientation contr
exture segmentation relies on grouping of texture elemn our stimuli, differences in neither absolute orientaor orientation contrast can explain the parallel–orthogsymmetry in accuracy and D-VEP amplitude. Indeed,ent orientation and local orientation contrast are the s
n the two configurations. The asymmetry must thenn grouping factors. Using the same stimulus, Caputoasco[3] already showed an electrophysiological corref grouping, reflected in a parallel–orthogonal D-VEP asetry.On the basis of our results we argue that, in the global
grouping process of similarly oriented elements is affey learning in both parallel and orthogonal configuratiorouping is increased in the parallel condition but decre
n the orthogonal, and the result is, in both configurationncrease in saliency.
We first discuss the possibility that grouping mechanay be inhibited by learning. A learning-dependent inh
ion of grouping is suggested by accuracy data in thelock of orthogonal configuration (Fig. 3) in global task. Thehow that accuracy is less than 50%, indicating that sbservers have a response bias for the opposite orientomparison betweenFig. 1b and c gives a phenomenologi

C. Casco et al. / Neuroscience Letters 371 (2004) 18–23 23
demonstration that global orientation gradient resulting fromgrouping may or may not conflict with the orientation thatobservers are asked to discriminate. The other possibility isthat a facilitatory effect of learning on grouping operationscould explain the results obtained with the parallel configura-tion, where local and global orientation do not conflict. Herethe improvement in accuracy was associated with a weak butsignificant increase in negativity for the earliest D-VEP com-ponent, but only in the global orientation discrimination task,suggesting grouping facilitation in global texture segmenta-tion. If such a facilitation accounted for learning effect onD-VEP in parallel condition, this would explain why only inthe global task N1 is enhanced by learning. Electrophysio-logical correlates of texture segregation has been provided[14] and Caputo and Casco[3] showed a specific compo-nent associated with grouping. As previous studies suggest,an learning-dependent facilitatory effect, specific for parallelcondition, could result from increased response of a binding[2] or grouping mechanism, due to modulation of the strengthin intracortical connections amongst channels at very earlylevels of cortical processing.
In the local orientation task instead, orientation judgmentrelies on orientation gradient not on orientation contrast. Wesuggest that cues resulting from global texture figure, in turnresulting from orientation contrast, have to be inhibited int ientj tedb ar-l nly.
nta-t e re-s putf arn-i ofp rel-e horte sin-g -c asksa sy-c racyd re-d calt
thel rialsw ning[ . Wea thep n linew thet in-t
To conclude, although a learning-dependent increase inrepresentation of one channel would be expected to producea corresponding decrease in representation of the others, veryfew studies have tested the hypothesis of inhibition[13]. Ourpresent results provide the first combined psychophysical andelectrophysiological evidence.
Acknowledgements
This study was supported by grants from PRIN (2001,2003); ATENEO, 2002; MIUR 60% to Clara Casco.
References
[1] Y. Adini, D. Sagi, M. Tsodyks, Excitatory-inhibitory network in thevisual cortex: psychophysical evidence, Proc. Natl. Acad. Sci. U.S.A.94 (1997) 10426–10431.
[2] G. Campana, C. Casco, Learning in combined-feature search: speci-ficity to orientation, Percept. Psychophys. 65 (2003) 1197–1207.
[3] G. Caputo, C. Casco, A visual evoked potential correlate of globalfigure-ground segmentation, Vision Res. 39 (1999) 1597–1610.
[4] C. Casco, G. Campana, Spatial interactions in simple and combinedfeatures visual search, Spat. Vis. 12 (1999) 467–483.
[5] B.A. Dosher, Z.L. Lu, Mechanisms of perceptual learning, VisionRes. 39 (1999) 3197–3221.
y ofdult
mans. 33
urr.
g ofrosci.
[ .[ ent
s in995)
[ rim-atl.
[ led
[ n isence
[ arly
[ ing,
[ res in
[ par-993)
he local tasks because they conflict with orientation gradudgment. This inhibition, specific for local task, is reflecy VEP results, showing that the inhibitory effect in the e
iest VEP component (N1) is present for the local task oOur interpretation of learning effects in texture segme
ion agrees with the view that learning may increase thponse within an orientation channel by weighting the inrom different channels, according to task relevance. Leng could lead to a reduction in orientation bandwidtherceptual filters by pruning or reducing inputs from irvant channels: the orientation bandwidth, which for sxperience spans several channels, may narrow to ale relevant channel after learning[5]. The inhibition ocurs at different spatial frequency scales in the two tnd this is supported by the finding that a similar phophysical effect due to learning (reduction of accuifference in the two configurations) is associated with auction of VEP amplitude of earlier components in the lo
ask.It is important to make a final remark concerning
earning paradigm we adopted. Since relatively few tere used, our effects are likely to be due to fast-lear
12], and cannot be extended to slow, enduring learningre now investigating long-lasting effects of learning onarallel–orthogonal asymmetry. The present results are iith fast perceptual learning effects of a modification of
uning properties of individual channels or modulation oferaction between channels[8].
[6] D. Edying, G. Schweigart, U.T. Eysel, Spatio-temporal plasticitcortical receptive fields to repetitive visual stimulation in the acat, Neuroscience (2002) 112.
[7] D.J. Field, A. Hayes, R.F. Hess, Contour integration by the huvisual system: evidence for a local association field, Vision Re(1993) 173–193.
[8] C.D. Gilbert, Learning: neural dynamics of perceptual learning, CBiol. 4 (1994) 627–629.
[9] S. Han, S. Fan, L. Chen, Y. Zhuo, On the different processinwholes and parts: a psychophysiological analysis, J. Cogn. Neu9 (1997) 687–698.
10] D. Hebb, The Organization of Behavior, Wiley, New York, 194911] M.K. Kapadia, M. Ito, C.D. Gilbert, G. Westheimer, Improvem
in visual sensitivity by changes in local context: parallel studiehuman observers and in V1 of alert monkeys, Neuron 15 (1843–856.
12] A. Karni, D. Sagi, Where practice makes perfect in texture discination: evidence for primary visual cortex plasticity, Proc. NAcad. Sci. U.S.A. 88 (1991) 4966–4970.
13] M.P. Kilgard, M.M. Merzenich, Cortical map reorganization enabby nucleus basalis activity, Science 279 (1998) 1714–1718.
14] V.A. Lamme, B.W. Van Dijk, H. Spekreijse, Texture segregatioprocessed by primary visual cortex in man and monkey. Evidfrom VEP experiments, Vision Res. 32 (1992) 797–807.
15] J. Malik, P. Perona, Preattentive texture discrimination with evision mechanisms, J. Opt. Soc. Am. A (1990) 7.
16] C.P. May, M.J. Kane, L. Hasher, Determinants of negative primPsychol. Bull. 118 (1995) 35–54.
17] D. Navon, Forest before trees: The precedence of global featuvisual perception, Cogn. Psychol. 9 (1977) 353–383.
18] H.C. Nothdurft, The role of features in preattentive vision: comison of orientation, motion and color cues, Vision Res. 33 (11937–1958.















![[Evaluation of the severity of clinical illness: a psychophysical approach]](https://img.dokumen.tips/doc/110x75/635f1b6fac6942764f03df81/evaluation-of-the-severity-of-clinical-illness-a-psychophysical-approach.jpg)













