Embed Size (px)
Citation preview

SYSTEMATICS OF THE CAMBRIAN TRILOBITE
FAMILY NEPEIDAE, WITH REVISION OF
AUSTRALIAN SPECIES
by JOHN R. PATERSONCentre for Ecostratigraphy and Palaeobiology, Department of Earth and Planetary Sciences, Macquarie University, North Ryde, NSW 2109, Australia;
e-mail: [email protected]
Typescript received 6 June 2003; accepted in revised form 16 January 2004
Abstract: Taxonomy of the Cambrian trilobite family
Nepeidae is revised. Morphometric analyses of the genera
Nepea and Penarosa demonstrate that use of proportions
of linear dimensions of the cranidium to differentiate
species is invalid, and that infrageneric variation is con-
tinuous. The taxonomy and biostratigraphy of all Austra-
lian species of Nepeidae is revised. Species considered valid
herein include: Nepea narinosa (type species), N. tonsillata,
N. nans, Penarosa retifera (type species), P. elaticeps,
P. rhinodelphis, P. netenta, Loxonepea loxophrys (type spe-
cies) and Ferenepea hispida (type species). Folliceps is con-
sidered to be a junior subjective synonym of Nepea,
Trinepea is regarded as a junior subjective synonym of
Penarosa, and Ascionepea is considered to be a junior sub-
jective synonym of Ferenepea.
Key words: Nepeidae, Australia, Cambrian, morphometrics,
biostratigraphy.
The Cambrian trilobite family Nepeidae Whitehouse,
1939 represents a distinct group within the Ptychopario-
idea. Once thought to be endemic to Australia (Opik
1963a), nepeids are now known also from Antarctica,
China, New Zealand and Kazakhstan (Jago in Brock et al.
2000). The family is well represented with seven genera
and 23 formally described species (excluding those left
under open nomenclature); however, the literature is lim-
ited to fewer than a dozen papers, including Whitehouse
(1939), Opik (1963a, 1967, 1970), Palmer and Gatehouse
(1972), Jell (1977), Jell and Robison (1978) and Xiang
and Zhang (1985). Whitehouse (1939) erected the family
Nepeidae and genus Nepea, with type species Nepea nari-
nosa. Opik (1967, 1970) named all but five of the known
species of Nepeidae. Initially, Opik (1963a) revised the
concept of the Nepeidae, in addition to the cephalic mor-
phology of N. narinosa. He later described two new gen-
era from the Mindyallan fauna of north-west Queensland
(Opik 1967), and subsequently published the most com-
prehensive study on the group (Opik 1970), a monograph
documenting four genera and 25 species (ten of which
are left under open nomenclature) from the Middle Cam-
brian of north-west Queensland and the Northern Terri-
tory. Palmer and Gatehouse (1972) described Trinepea, a
new genus from Antarctica, the first record of nepeids
outside Australia. Jell (1977) described a new species of
Penarosa from north-west Queensland, and placed Trine-
pea in synonymy with Penarosa. Jell and Robison (1978),
in re-documenting a previously known Whitehouse
(1936, 1939) locality near Thorntonia Station, north-west
Queensland, recorded Penarosa retifera from a Middle
Cambrian trilobite faunule of the Currant Bush Lime-
stone. Xiang and Zhang (1985) described two new species
and one unnamed species of Nepea from China. Fortey
and Owens (1997) and Fortey and Hughes (1998) have
discussed the functional morphology of the preglabellar
boss in nepeids, and Jago (in Brock et al. 2000) provided
a synthesis on the stratigraphical and geographical distri-
bution of the group.
This study represents a taxonomic revision of the Aus-
tralian Nepeidae, with a re-examination of material col-
lected and described by F. W. Whitehouse, A. A. Opik
and P. A. Jell, in addition to new material I have collected
from the Middle Cambrian Gowers Formation, Georgina
Basin, north-west Queensland. The study includes a
morphometric analysis of the entire suite of specimens
(including new collections) upon which Whitehouse
(1939), Opik (1970) and Jell (1977) based their species
designations. Morphometric analysis was conducted in an
attempt to resolve fully the taxonomic status of all Aus-
tralian species of Nepea and Penarosa. The biostratigraphy
and geographical distribution of the group is also revised.
[Palaeontology, Vol. 48, Part 3, 2005, pp. 479–517]
ª The Palaeontological Association 479

BIOSTRATIGRAPHY
Australian occurrences
The nepeid trilobites documented in this paper are all
from Middle–Upper Cambrian strata exposed in the
north-west Queensland and eastern Northern Territory
regions of the Georgina Basin, and one occurrence in the
eastern Amadeus Basin in the Northern Territory. The
stratigraphy and correlation of these regions is illustrated
in Text-figure 1. The current Middle–Upper Cambrian
(Floran–Mindyallan) biostratigraphic scheme is largely
that developed by Opik (1961, 1963b, 1967, 1979) from
sampling of surface outcrop mostly from the Queensland
portion of the Georgina Basin. As outcrop in this basin is
generally poor and because measurable sections are lack-
ing, most of the information comes from spot localities
(J. Laurie, pers. comm. 2003). Because many of the
nepeid species erected by Opik (1967, 1970) are synony-
mized herein (see ‘Systematic palaeontology’), the bio-
stratigraphical ranges for some valid species have been
extended as a result. The ranges of all Australian species
considered valid herein are illustrated in Text-figure 2,
plotted against the northern Australian Cambrian bio-
stratigraphic scheme (sensu Geyer and Shergold 2000).
Locality and associated age information is presented in
the Appendix (section 1). Nepeids have also been repor-
ted from Cambrian successions in New South Wales
(Opik 1970), Victoria (Thomas and Singleton 1956;
Edwards et al. 1998), South Australia (Jago et al. 2002)
and Tasmania (Laurie et al. 1995), but none of the mater-
ial has been illustrated or discussed in detail, and apart
from Tasmanian, South Australian and Victorian occur-
rences (see Laurie et al. 1995, Jago et al. 2002 and
Edwards et al. 1998, respectively), field locality data are
insufficient for re-collection and the whereabouts of col-
lected specimens is unknown.
The oldest nepeid occurrence in Australia is represen-
ted by Nepea? sp. nov. NC, described by Opik (1970).
Locality information for this species is limited, with Opik
(1970) only recording that specimens came from a black
limestone bed in the Ross River Gorge, east of Alice
Springs, Northern Territory. This black limestone bed
represents part of the Giles Creek Dolomite (J. Laurie,
TEXT -F IG . 1 . Correlation of Cambrian successions in the north-west Queensland and eastern Northern Territory regions of the
Georgina Basin and eastern Amadeus Basin.
480 P A L A E O N T O L O G Y , V O L U M E 4 8

pers. comm. 2003). The co-occurrence of Xystridura indi-
cates an early Middle Cambrian (Ordian ⁄Early Templeto-
nian) age. It is doubtful that this species belongs to
Nepea. The apparent clavate shape of the preglabellar boss
and strongly tapered glabella (Opik 1970, pl. 7, fig. 4) is
characteristic of Loxonepea, but the fragmentary nature of
the specimens makes precise taxonomic assessment diffi-
cult.
Penarosa ranges from the Late Templetonian ⁄Floran
(Acidusus atavus Zone) to the early Undillan (Ptychagnos-
tus punctuosus Zone). Penarosa elaticeps and P. rhinodel-
phis occur with agnostoids typical of the Acidusus atavus
Zone (Opik 1979) from a single locality (M179) in the
lower part of the Currant Bush Limestone on Lancewood
Creek, Mount Drummond area, Northern Territory.
The type species of Penarosa, P. retifera, is widespread
and ranges from the Acidusus atavus Zone to the Ptychag-
nostus punctuosus Zone. The lowest stratigraphical occur-
rence of P. retifera is from the Arthur Creek Formation
(locality H4), near Huckitta Station, close to the Arthur
River, Northern Territory. Opik (1970) regarded speci-
mens from this locality as belonging to the Acidusus ata-
vus Zone. Specimens of P. retifera commonly occur in the
upper parts of the Currant Bush Limestone that crops
out in the Mount Drummond area of the Northern Terri-
tory (localities M180 and M186), and the Camooweal and
Lawn Hill areas of north-west Queensland (localities
M28, M123, M124, M130, M393, UQL256, UQL463).
Associated agnostoids from these localities indicate an age
spanning the Euagnostus opimus and Ptychagnostus punc-
tuosus zones (Jell and Robison 1978; Opik 1979; Laurie
1988). Specimens of P. retifera also occur in the coeval
and laterally interfingering Age Creek Formation in the
Morstone–Thorntonia area of north-west Queensland
(localities M157 and M160), and are abundant in the
Gowers Formation at Thorntonia Station (locality GF1),
which underlies the Currant Bush Limestone (Southgate
1986; Shergold and Southgate 1986). Agnostoids from the
Age Creek and Gowers formations are indicative of the
Euagnostus opimus Zone (Opik 1979).
Penarosa netenta is restricted to the Euagnostus opimus
Zone (Opik 1970, 1979; Jell 1977). This species occurs
only at Thorntonia Station, north-west Queensland, in
the vicinity of Chummy Bore (localities M412, QML136,
QML152). Opik (1970, 1979) regarded the shales, dolo-
mitic limestones and chert at Chummy Bore as represent-
ing the Inca Formation, while de Keyser and Cook (1972;
see also Jell 1977) coined the name Chummy Bore
TEXT -F IG . 2 . Biostratigraphic ranges of valid Australian species of Nepeidae plotted against Australian Cambrian stages and trilobite
biozones.
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 481

Formation for these rocks. Southgate (1986) conducted
detailed lithostratigraphic mapping and sedimentological
studies in this area and considered these rocks to repre-
sent part of the Gowers Formation, and regarded the
Chummy Bore Formation as being of doubtful utility.
Loxonepea loxophrys occurs in the Acidusus atavus Zone
portion of the Age Creek Formation (locality M122), and
in the Euagnostus opimus Zone portion of the Currant
Bush Limestone (locality M124), Camooweal area, north-
west Queensland (Opik 1970, 1979).
Nepea is a particularly long-ranging genus, spanning
the Late Templetonian ⁄Floran (Euagnostus opimus Zone)
to the Boomerangian (Lejopyge laevigata Zone). Nepea
nans (formerly Folliceps nans) represents the oldest spe-
cies, occurring in the Euagnostus opimus Zone (Opik
1979; Laurie 1988) of the Currant Bush Limestone in the
Camooweal area, north-west Queensland (localities M123,
M124).
Nepea tonsillata ranges within the Ptychagnostus punc-
tuosus and Goniagnostus nathorsti zones, and is known
only from a single locality (M54) in the V-Creek Lime-
stone, which crops out in Douglas Creek, south of Undil-
la homestead, north-west Queensland (Opik 1970). Opik
(1970, 1979) considered N. tonsillata and associated
agnostoids indicative of the Goniagnostus nathorsti Zone.
However, Opik (1979) listed only Doryagnostus magister
(¼ D. incertus) and Hypagnostus clipeus at locality M54.
D. incertus has an observed range from the punctuosus to
nathorsti zones in Gondwanan and Baltic regions (Wester-
gard 1946; Robison 1978; Peng and Robison 2000), and
thus N. tonsillata should be considered to range some-
where within these zones.
Nepea narinosa ranges from the Doryagnostus deltoides
Zone to the Lejopyge laevigata Zone. The lowest strati-
graphical occurrence of N. narinosa comes from the
V-Creek Limestone (locality M41) in the Camooweal
region of north-west Queensland, associated with agnos-
toids indicative of the Doryagnostus deltoides Zone
(Opik 1979; Laurie 1988). Specimens are rare in the
conformably overlying Mail Change Limestone. Speci-
mens of N. narinosa are abundant throughout the Split
Rock Sandstone, which has a diachronous contact
against both the Mail Change and the V-Creek lime-
stones in the Camooweal and Mount Isa regions of
north-west Queensland (localities M133, M141, M226,
M276, M344, M417, M421). Whitehouse (1939) origin-
ally described N. narinosa from the type locality of the
Split Rock Sandstone at Split Rock Waterhole on
Waroona Creek, north-west Queensland; this locality is
equivalent to Opik’s (1970) locality M417. Agnostoids
from the Split Rock Sandstone are indicative of the
Goniagnostus nathorsti Zone (Opik 1979). Specimens of
N. narinosa are also abundant in the Steamboat Sand-
stone (localities D54, D95, D108), which crops out in
the vicinity of Quita Creek in the Urandangi region of
north-west Queensland. Opik (1967, 1970, 1979) regar-
ded the agnostoids and polymerid trilobites from the
Steamboat Sandstone as belonging to the Lejopyge lae-
vigata Zone.
All Mindyallan species described by Opik (1967) are
herein considered to be synonymous, with Ferenepea
hispida representing the valid type species. For this rea-
son, the use of Ascionepea janitrix as a zonal taxon is
disregarded. Opik (1967) established what he called the
‘Zone of Passage’ (or the ‘Damesella torosa-Ascionepea
janitrix Zone’), which represented a transitional zone
from the Middle to Late Cambrian. Opik (1967, p. 8)
defined this zone as ‘a fauna [including A. janitrix] that
occurs in the span beginning with the last known Lejo-
pyge laevigata and ending with the first appearance of
Erediaspis eretes and its associates’. However, Daily and
Jago (1975) demonstrated that Lejopyge cos, which Opik
(1967) had recorded from this zone, is a synonym of
Lejopyge laevigata, and thus extended the range of that
species in Australia to the Acmarhachis quasivespa Zone,
therefore placing the Middle ⁄Late Cambrian boundary
within the latter zone. Furthermore, Daily and Jago
(1975) noted that Damesella torosa, which according to
Opik (1967) was confined to the ‘Zone of Passage’,
extended into the Erediaspis eretes Zone. As all Mindy-
allan nepeid species are synonymous with Ferenepea his-
pida, this species ranges from the Lejopyge laevigata
Zone to the Glyptagnostus stolidotus Zone (Opik 1967).
Therefore, F. hispida first appears at the top of the
Steamboat Sandstone (locality G103), in the Mungere-
bar–Mindyalla area of north-west Queensland. As noted
previously, the agnostoids from the Steamboat Sand-
stone are indicative of the Lejopyge laevigata Zone
(Opik 1967). Specimens of F. hispida occur throughout
the overlying Mungerebar Limestone, which crops out
at numerous localities (G8–10, G114, G119, G127,
G150, G417, G429) in the Mungerebar–Mindyalla
region of north-west Queensland. Agnostoids and poly-
merid trilobites suggest an early Mindyallan (Erediaspis
eretes Zone) age for the lower part of the Mungerebar
Limestone, extending into the Acmarhachis quasivespa
Zone in the upper part of the unit (Opik 1967).
Specimens of F. hispida are common in the light-
coloured shales of the O’Hara Shale in the Selwyn
Range, Duchess area, north-west Queensland (localities
D6, D28–29). The very diverse agnostoid and
polymerid faunas at locality D29 are indicative of the
Glyptagnostus stolidotus Zone (Opik 1967). Opik (1967)
also recorded F. hispida from the Georgina
Limestone (locality W1) in the Glenorminston–Sylvester
Creek area, north-west Queensland, associated
with a fauna indicative of the Glyptagnostus stolidotus
Zone.
482 P A L A E O N T O L O G Y , V O L U M E 4 8

Other Gondwanan occurrences
The oldest known occurrence of the Nepeidae outside
Australia is that of Trinepea (¼ Penarosa) trinodus from
the lower part of the Nelson Limestone in the Neptune
Range, Transantarctic Mountains, Antarctica (Palmer and
Gatehouse 1972). This species is a constituent of the
Amphoton oatesi faunule, which Cooper and Shergold
(1991) considered to be Floran–Undillan in age.
Nepea brevica and N. xinjiangensis have been described
from the Kensay Formation, northern Tianshan, Xinjiang,
North China (Xiang and Zhang 1985). These species
occur in the ‘upper trilobite assemblage’ within the unit
and are considered to be Undillan in age, spanning the
Ptychagnostus punctuosus–Goniagnostus nathorsti zones.
Undescribed nepeids have also been reported to occur
in limestone lenses within the Tasman Formation, north-
west Nelson, New Zealand, and are associated with coryn-
exochids, dolichometopids, solenopleurids, kopturids and
Pianaspis, which suggest a late Undillan age (Cooper in
Cooper and Bradshaw 1985; Shergold et al. 1985).
MORPHOMETRIC ANALYSIS
The great pioneering works of Opik (1963a, 1967, 1970)
were pivotal to our knowledge of the Nepeidae; however,
he erected an unusually large number of species, in par-
ticular of Nepea and Penarosa (Opik 1970). Fortey and
Hughes (1998, p. 647) have commented that ‘many of the
species erected by Opik are known from small numbers
of specimens, and some from unique ones’. Hughes and
Labandeira (1995) stated that insufficient sampling may
give rise to the appearance of discrete morphological
character states, which can be misinterpreted to have
taxonomic significance. Some of Opik’s (1970) localities
show the co-occurrence of several species of one genus at
a single locality, for example the type locality (M179) for
Penarosa elaticeps, P. meniscops, P. rhinodelphis and
P. zeabunda. In addition, species from M179 are based on
no more than five specimens. Labandeira and Hughes
(1994) noted that taxonomic errors result when intraspe-
cific variations are mistaken for interspecific differences,
leading to oversplit taxa containing species based on
minor and ⁄or inconsistent differences. In an attempt to
resolve fully the taxonomic status of all Australian species
of Nepea and Penarosa a detailed morphometric analysis
was conducted.
Bivariate and multivariate analyses were undertaken to
determine the validity of using linear characters to differ-
entiate species of Nepeidae. Methods used in this analysis
are similar to those presented by Hughes and Jell (1992)
and Hughes (1994). Material analysed includes all speci-
mens documented by Whitehouse (1939), Opik (1970)
and Jell (1977), plus additional material collected but not
documented or figured by Opik, and new collections of
mine (Appendix, section 2). Analysis of this pooled sam-
ple is designed to reveal both inter- and intraspecific vari-
ation and locality-related variation. A total of 117
cranidia were assessed based on nine linear measurements
(see Text-fig. 3). Symbols and terminology for sclerite
dimensions follow Shaw (1957); however, J2 here repre-
sents the maximum cranidial border width (tr.). The pre-
glabellar boss length and width (PBL and PBW,
respectively) have only been analysed bivariately for
Nepea. These nine linear measurements were selected
because they represent the morphological characters that
Opik (1970) considered to be taxonomically significant in
TEXT -F IG . 3 . Linear dimensions measured on cranidia of Penarosa and Nepea. Abbreviations follow Shaw (1957): B1, occipital–
glabellar length (sag.); F1, frontal area length (sag.); G1, cranidial border length (sag.); J2, maximum cranidial border width (tr.); J5,
post-palpebral cranidial width (tr.); K, occipital lobe width (tr.); K5, anterior glabellar width (tr.); PBL, maximum length of
preglabellar boss (sag.); PBW, maximum width of preglabellar boss (tr.).
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 483

determining different species. Linear distances were meas-
ured to the nearest tenth of a millimetre using Vernier
calipers. In cases where measurements were taken from
incomplete specimens with one side of an axially symmet-
rical structure missing, measurements obtained from the
complete half were doubled following the methods of
Labandeira and Hughes (1994. p. 493).
Bivariate analysis
A set of bivariate scatter plots have been established for
Penarosa and Nepea (see Text-figs 4–6); Folliceps is inclu-
ded with Nepea due to suspected synonymy. The mor-
phological variation in each of the nine linear characters
is assessed with reference to a standard measure for size,
in this case the occipital-glabellar length (B1) following
the methods of Hughes (1994, pp. 23–24). Data from lin-
ear characters were analysed using the Pearson product-
moment correlation coefficient (r) to determine the
extent to which two variables covary during growth. The
coefficient varies between zero and one, with zero indica-
ting no correlation and one indicating perfect linear cor-
relation. A more detailed definition and explanation of
this coefficient is provided by Imbrie (1956, p. 233) and
Shaw (1956, p. 1215). The reduced major axis (RMA)
was also calculated to determine growth relationships of a
pair of variables; both axes on the bivariate plots have
been log-transformed to determine RMA values. RMA
values indicate whether linear characters display allomet-
ric or isometric growth. Values significantly different
from 1Æ0 (at 95% confidence levels) indicate allometry,
and values equal to or not significantly different from 1Æ0indicate isometry. For further details of RMA, see Imbrie
(1956), Hayami and Matsukuma (1970) and Hughes
(1994). The Pearson product-moment correlation coeffi-
cient and RMA were calculated using PAST, version 1Æ01,
by Øyvind Hammer and David Harper (for details see
Hammer et al. 2001); r and RMA values for Penarosa and
Nepea are presented in Tables 1 and 2, respectively.
Results of bivariate analysis
The bivariate relationships of Penarosa and Nepea illus-
trate that the majority of the linear characters assessed,
with the exception of the cranidial border length (G1), do
not allow differentiation of more than one species for
each genus. The main reasons for this are: (1) all speci-
mens fall within a single trendline; (2) specimens of each
species in the pooled type-suite show some variation and
lie within the range of variation of other species; and (3)
the majority of type-suite specimens lie within the range
of variation shown by the locality-based collections. This
interpretation is further supported by the consistently
high correlation coefficient values for each character (i.e.
r > 0Æ85). This analysis demonstrates that these variations
are not species specific and in most cases represent allo-
metric growth patterns in these characters.
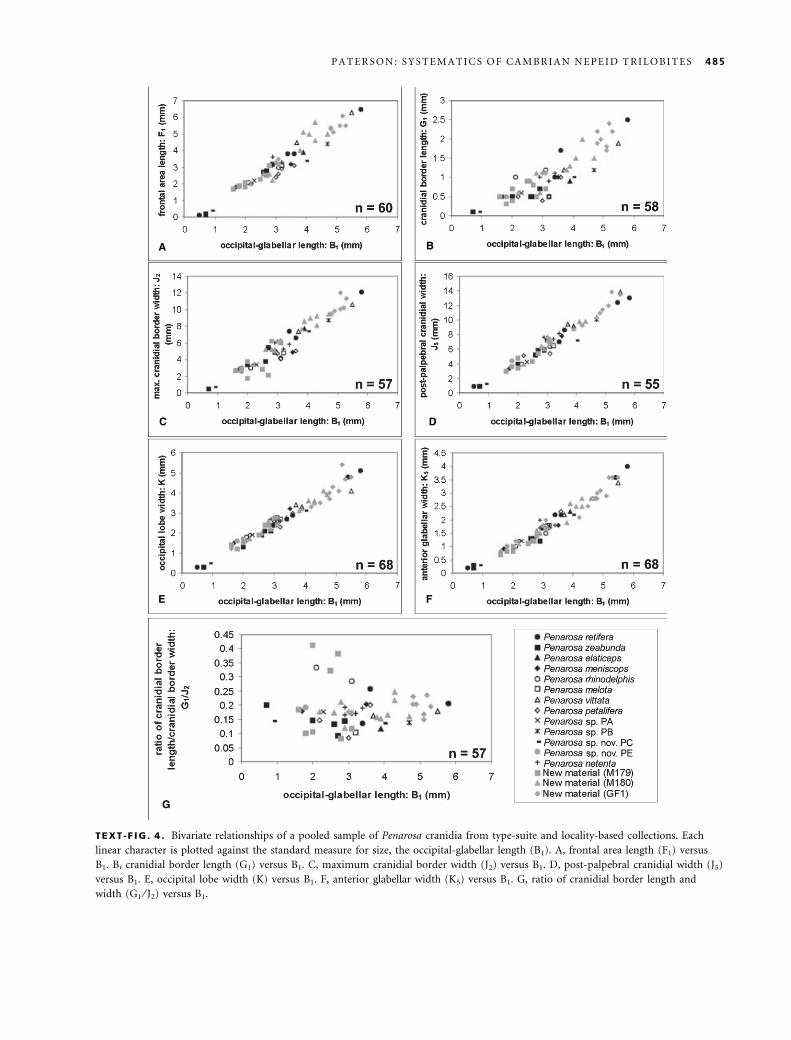
Penarosa. Opik (1970) was consistent in using linear
dimensions of the cranidium to differentiate between spe-
cies of Penarosa. In particular, he emphasized the relative
width of the interocular cheeks, length and width of the
frontal area, and shape of the glabella (i.e. anterior and
posterior width) to differentiate species. However, as seen
in Text-figure 4, all specimens from Opik’s (1970) type-
suite, specimens of P. netenta and those from new collec-
tions show tight clustering about a single trendline and
display continuous and gradational variation throughout
the sample, suggesting there is no justification for the
recognition of more than one morphotype. The relatively
low amount of variance within frontal area length (F1),
cranidial border width (J2), post-palpebral cranidial width
(J5), anterior glabellar width (K5), and occipital lobe
width (K) is also supported by the very high correlation
coefficients (r > 0Æ95) of each character (Table 1). The
RMA values indicate allometric growth in all characters
(Table 1).
The bivariate relationship between cranidial border
length (G1) and occipital-glabellar length (B1) shows that
there is a significant amount of scatter between these vari-
ables, suggesting marked size-independent variation (i.e.
variation is continuous throughout the sample and is not
correlated with overall size). The relatively low correlation
coefficient for the total sample reflects the high degree of
variability in the cranidial border length (r ¼ 0Æ888).
Growth of the cranidial border length is strongly posi-
tively allometric, indicating the relative size of the crani-
dial border length drastically increases throughout
holaspid ontogeny. The variation in cranidial border
length within and between species and locality-based col-
lections would suggest that growth was somewhat more
flexible than in other linear characters. However, speci-
mens of P. rhinodelphis appear to plot along the top of
the trendline (Text-fig. 4B). In plotting the ratio of crani-
dial border length ⁄ cranidial border width (G1 ⁄ J2) against
occipital-glabellar length (B1), type specimens of P. rhino-
delphis and three other specimens from locality M179
(the type locality of P. rhinodelphis) become isolated from
the major trendline, thus suggesting a separate species
(Text-fig. 4G). In pooling specimens of Opik’s (1970)
type-suite of species from M179 and new material from
this locality, plotting the ratio of G1 ⁄ J2 against B1 (Text-
fig. 5) yielded similar results to that of the total sample
(Text-fig. 4G). This relationship illustrates the discrimin-
ation of two distinct species consisting of P. zeabunda-
elaticeps and P. rhinodelphis at locality M179; P. meniscops
484 P A L A E O N T O L O G Y , V O L U M E 4 8
A B
C
E F
G
D
TEXT -F IG . 4 . Bivariate relationships of a pooled sample of Penarosa cranidia from type-suite and locality-based collections. Each
linear character is plotted against the standard measure for size, the occipital-glabellar length (B1). A, frontal area length (F1) versus
B1. B, cranidial border length (G1) versus B1. C, maximum cranidial border width (J2) versus B1. D, post-palpebral cranidial width (J5)
versus B1. E, occipital lobe width (K) versus B1. F, anterior glabellar width (K5) versus B1. G, ratio of cranidial border length and
width (G1 ⁄ J2) versus B1.
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 485
could be included in either group. However, trendlines in
Text-figures 4G and 5 suggest that in specimens of P. rhi-
nodelphis with an occipital-glabellar length longer than
about 3Æ5 mm, the ratio would stabilize with other species
of Penarosa and remain constant thereafter. One possible
explanation is that the trends in the two morphs from
M179 represent intraspecific heterochrony. Hughes (1994)
observed intraspecific heterochrony in Dikelocephalus,
where one particular population (‘Arcadia’) possessed
extraordinarily large eyes and long pygidial spines in
young holaspides, suggesting retention of juvenile features
in adulthood. However, by the late holaspid stage, Arca-
dia specimens are indistinguishable from specimens from
other populations, and hence this developmental differ-
ence only occurs in younger individuals. Hughes (1994)
could not explain this developmental shift in Dikelocepha-
lus because its preholaspid ontogeny is unknown. How-
ever, with regards to Penarosa, of the few meraspid
specimens from M179 illustrated by Opik (1970, pl. 17,
figs 5–6), the cranidial border does not display the same
ratio of length to width as in younger holaspides of P. rhi-
nodelphis, suggesting that the P. rhinodelphis morphotype
does not represent the prolonged expression of juvenile
features (i.e. an intraspecific heterochronic shift), and
thus exemplifies a distinct species.
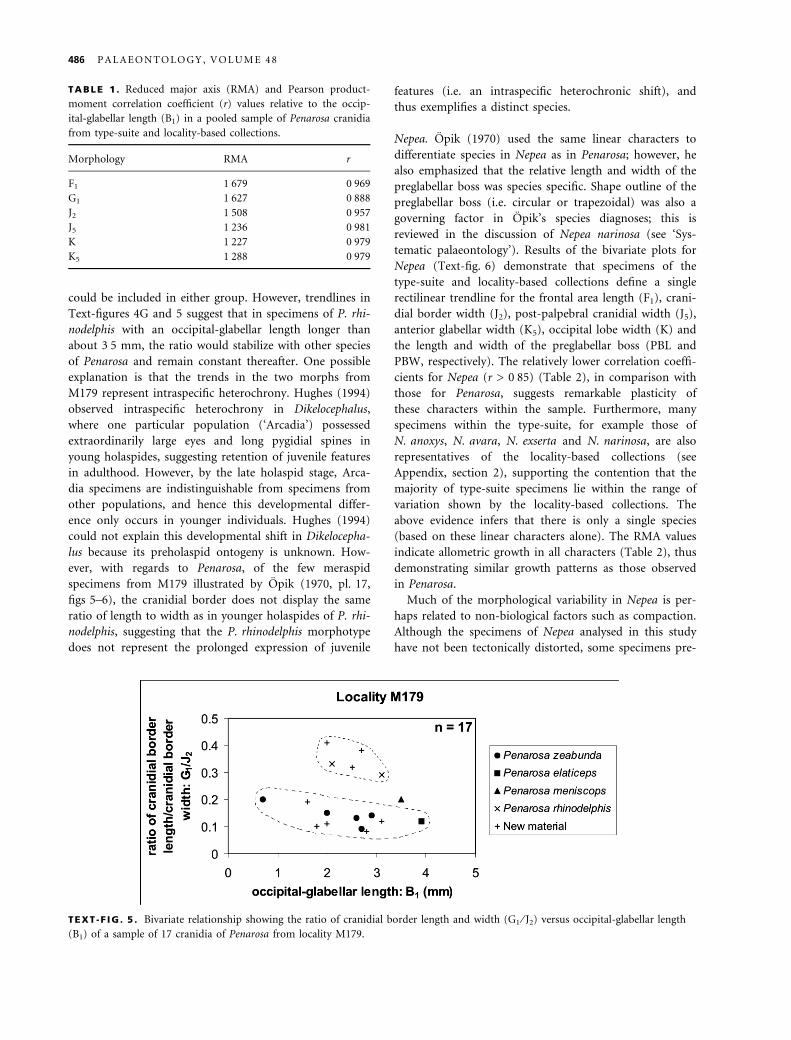
Nepea. Opik (1970) used the same linear characters to
differentiate species in Nepea as in Penarosa; however, he
also emphasized that the relative length and width of the
preglabellar boss was species specific. Shape outline of the
preglabellar boss (i.e. circular or trapezoidal) was also a
governing factor in Opik’s species diagnoses; this is
reviewed in the discussion of Nepea narinosa (see ‘Sys-
tematic palaeontology’). Results of the bivariate plots for
Nepea (Text-fig. 6) demonstrate that specimens of the
type-suite and locality-based collections define a single
rectilinear trendline for the frontal area length (F1), crani-
dial border width (J2), post-palpebral cranidial width (J5),
anterior glabellar width (K5), occipital lobe width (K) and
the length and width of the preglabellar boss (PBL and
PBW, respectively). The relatively lower correlation coeffi-
cients for Nepea (r > 0Æ85) (Table 2), in comparison with
those for Penarosa, suggests remarkable plasticity of
these characters within the sample. Furthermore, many
specimens within the type-suite, for example those of
N. anoxys, N. avara, N. exserta and N. narinosa, are also
representatives of the locality-based collections (see
Appendix, section 2), supporting the contention that the
majority of type-suite specimens lie within the range of
variation shown by the locality-based collections. The
above evidence infers that there is only a single species
(based on these linear characters alone). The RMA values
indicate allometric growth in all characters (Table 2), thus
demonstrating similar growth patterns as those observed
in Penarosa.
Much of the morphological variability in Nepea is per-
haps related to non-biological factors such as compaction.
Although the specimens of Nepea analysed in this study
have not been tectonically distorted, some specimens pre-
TEXT -F IG . 5 . Bivariate relationship showing the ratio of cranidial border length and width (G1 ⁄ J2) versus occipital-glabellar length
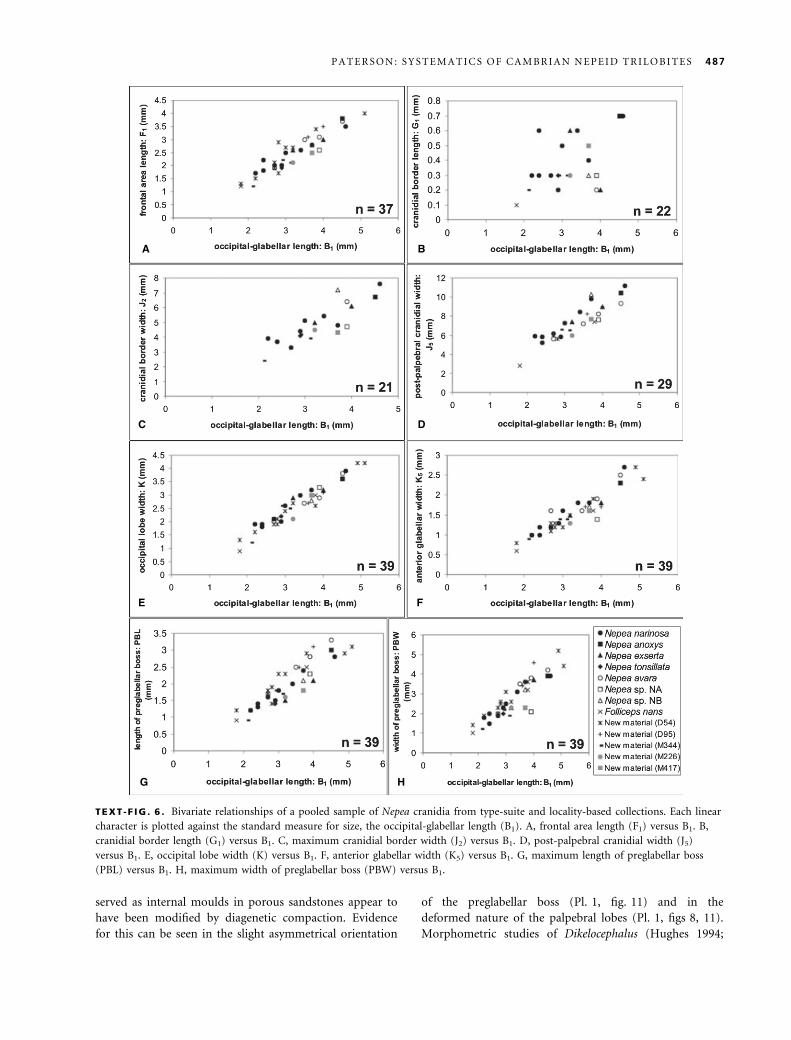
(B1) of a sample of 17 cranidia of Penarosa from locality M179.
TABLE 1 . Reduced major axis (RMA) and Pearson product-
moment correlation coefficient (r) values relative to the occip-
ital-glabellar length (B1) in a pooled sample of Penarosa cranidia
from type-suite and locality-based collections.
Morphology RMA r
F1 1Æ679 0Æ969
G1 1Æ627 0Æ888
J2 1Æ508 0Æ957
J5 1Æ236 0Æ981
K 1Æ227 0Æ979
K5 1Æ288 0Æ979
486 P A L A E O N T O L O G Y , V O L U M E 4 8
served as internal moulds in porous sandstones appear to
have been modified by diagenetic compaction. Evidence
for this can be seen in the slight asymmetrical orientation
of the preglabellar boss (Pl. 1, fig. 11) and in the
deformed nature of the palpebral lobes (Pl. 1, figs 8, 11).
Morphometric studies of Dikelocephalus (Hughes 1994;
A B
DC
E F
HG
TEXT -F IG . 6 . Bivariate relationships of a pooled sample of Nepea cranidia from type-suite and locality-based collections. Each linear
character is plotted against the standard measure for size, the occipital-glabellar length (B1). A, frontal area length (F1) versus B1. B,
cranidial border length (G1) versus B1. C, maximum cranidial border width (J2) versus B1. D, post-palpebral cranidial width (J5)
versus B1. E, occipital lobe width (K) versus B1. F, anterior glabellar width (K5) versus B1. G, maximum length of preglabellar boss
(PBL) versus B1. H, maximum width of preglabellar boss (PBW) versus B1.
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 487

Labandeira and Hughes 1994) have demonstrated that
specimens from both sandstone and siltstone lithologies
that have undergone compaction do not exhibit any sig-
nificant separation in morphological trends. However, the
overall convexity of Dikelocephalus is considerably lower
than that found in Nepea, and it is to be expected that
compaction would have little effect on the measurements
of a trilobite with low convexity. Hughes (1994) noted
that orthogonal dimensions on arched (i.e. convex) surfa-
ces are most likely to be distorted by compression and
deformation. The best examples of such dimensions in
Nepea are the length and width of the strongly con-
vex preglabellar boss (PBL and PBW, respectively). The
correlation coefficients for PBL and PBW are the lowest
of all other pairwise correlation coefficients, with the
exception of the length and width of the cranidial border
(discussed below) (Table 2). This demonstrates that com-
paction probably accounts for at least some variation
within Nepea. The significant degree of variation in the
cranidial border length (G1) (Text-fig. 6B) is probably the
result of compaction of the preglabellar boss directly
influencing the relative length of the cranidial border. In
specimens of Nepea where compaction has resulted in the
anterior expansion of the preglabellar boss, the relative
length of the cranidial border will become shorter (sag.)
as the anterior margin of the boss encroaches onto the
border. This explains the significantly scattered distribu-
tion of specimens in the bivariate plot for the cranidial
border length (G1) and occipital-glabellar length (B1)
(Text-fig. 6B), and the extremely low correlation coeffi-
cient of G1 (r ¼ 0Æ464). Further evidence to support com-
pression in specimens of Nepea is discussed in the
revision of non-linear characters below.
Results of the bivariate analysis demonstrate that only
two species of Penarosa and one species of Nepea exist
when based solely on the linear characters assessed. They
also demonstrate how infrageneric and intraspecific vari-
ation and growth patterns in the linear dimensions of
holaspides (in addition to the effects of diagenetic proces-
ses such as compaction) was mistaken by Opik for inter-
specific differences when dealing with small sample sizes.
Multivariate analysis
Hughes (1994) noted that it is possible that subtle but
discrete patterns of covariance among sclerite dimensions,
which would reveal the presence of more than one spe-
cies, might not be detected by bivariate analysis. For this
reason, principal components analysis (PCA) was conduc-
ted for Penarosa and Nepea to determine possible further
morphological trends in the linear dimensions tested in
the bivariate analysis. These analyses only used specimens
that possess a complete data set, i.e. specimens from
which a measurement can be obtained for each character.
For a detailed summary of PCA, see Hughes (1994,
pp. 41–42).
Results of multivariate analysis
As noted by Hughes (1994), the overall importance of
each principal component (i.e. how much of the total
variation is accounted for by each principal component)
is represented by the eigen value. The contribution of
each input variable to each principal component is
referred to as the eigenweight (¼ eigenvector). The first
principal component (PC1) represents a discriminator of
size and accounts for the majority of variation when deal-
ing with the linear dimensions of a sample (Reyment
et al. 1984; Hughes 1994). Therefore it is important to
investigate subsequent principal components in an
attempt to detect the presence of discrete clusters that
may represent interspecific covariance within a sample.
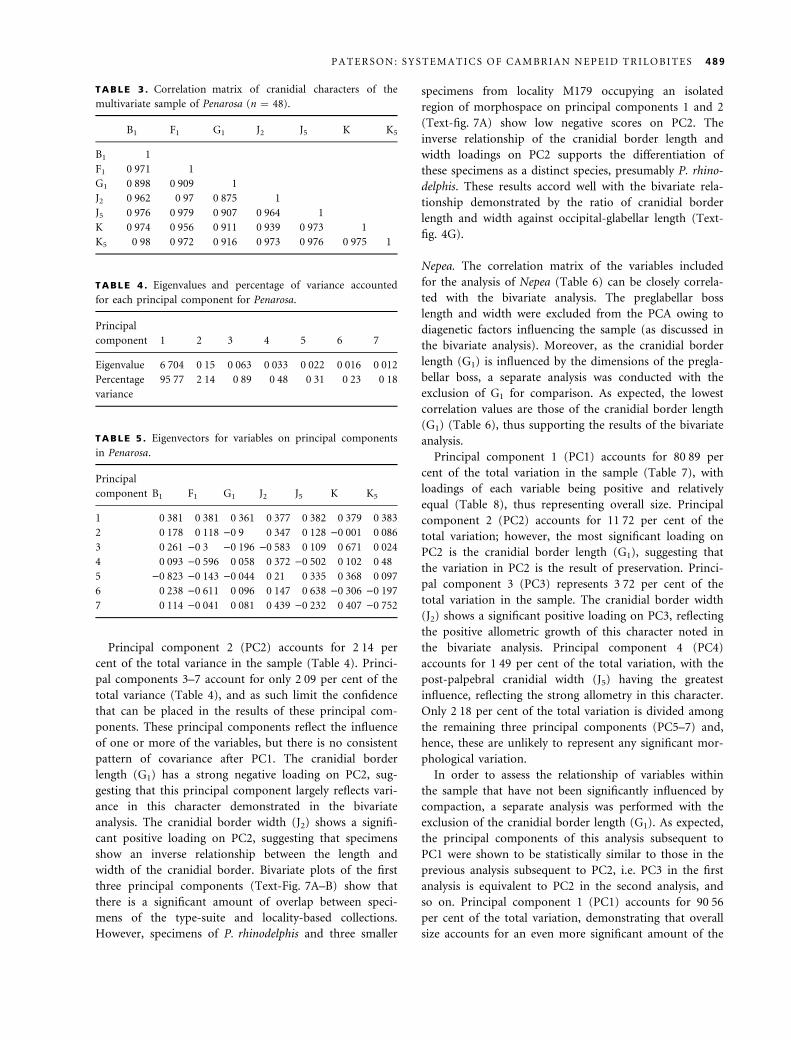
Penarosa. The correlation matrix of the variables used for
the multivariate sample for Penarosa (Table 3) shows a
strong relationship between all pairs and a concordance
with the results of the bivariate analysis. The consistently
lower correlation values of the cranidial border length
(G1) with other variables reflect the growth patterns
observed in the bivariate analysis.
Results for Penarosa show that the first principal com-
ponent (PC1) accounts for 95Æ77 per cent of the total
variation in the sample (Table 4). The loadings of each
variate on PC1 are all positive and equal (Table 5), sug-
gesting that PC1 represents overall size. Therefore, as PC1
accounts for a considerable percentage of the total vari-
ation and largely reflects overall size, size differences
account for the majority of variation in the sample. This
can be attributed to the fact that the sample incorporates
a wide range of holaspid instars, as noted by Hughes and
Jell (1992) and Hughes (1994).
TABLE 2 . Reduced major axis (RMA) and Pearson product-
moment correlation coefficient (r) values relative to the occip-
ital-glabellar length (B1) in a pooled sample of Nepea cranidia
from type-suite and locality-based collections.
Morphology RMA r
F1 1Æ245 0Æ928
G1 1Æ981 0Æ464
J2 1Æ247 0Æ855
J5 1Æ235 0Æ904
K 1Æ293 0Æ964
K5 1Æ249 0Æ929
PBL 1Æ334 0Æ873
PBW 1Æ459 0Æ899
488 P A L A E O N T O L O G Y , V O L U M E 4 8
Principal component 2 (PC2) accounts for 2Æ14 per
cent of the total variance in the sample (Table 4). Princi-
pal components 3–7 account for only 2Æ09 per cent of the
total variance (Table 4), and as such limit the confidence
that can be placed in the results of these principal com-
ponents. These principal components reflect the influence
of one or more of the variables, but there is no consistent
pattern of covariance after PC1. The cranidial border
length (G1) has a strong negative loading on PC2, sug-
gesting that this principal component largely reflects vari-
ance in this character demonstrated in the bivariate
analysis. The cranidial border width (J2) shows a signifi-
cant positive loading on PC2, suggesting that specimens
show an inverse relationship between the length and
width of the cranidial border. Bivariate plots of the first
three principal components (Text-Fig. 7A–B) show that
there is a significant amount of overlap between speci-
mens of the type-suite and locality-based collections.
However, specimens of P. rhinodelphis and three smaller
specimens from locality M179 occupying an isolated
region of morphospace on principal components 1 and 2
(Text-fig. 7A) show low negative scores on PC2. The
inverse relationship of the cranidial border length and
width loadings on PC2 supports the differentiation of
these specimens as a distinct species, presumably P. rhino-
delphis. These results accord well with the bivariate rela-
tionship demonstrated by the ratio of cranidial border
length and width against occipital-glabellar length (Text-
fig. 4G).
Nepea. The correlation matrix of the variables included
for the analysis of Nepea (Table 6) can be closely correla-
ted with the bivariate analysis. The preglabellar boss
length and width were excluded from the PCA owing to
diagenetic factors influencing the sample (as discussed in
the bivariate analysis). Moreover, as the cranidial border
length (G1) is influenced by the dimensions of the pregla-
bellar boss, a separate analysis was conducted with the
exclusion of G1 for comparison. As expected, the lowest
correlation values are those of the cranidial border length
(G1) (Table 6), thus supporting the results of the bivariate
analysis.
Principal component 1 (PC1) accounts for 80Æ89 per
cent of the total variation in the sample (Table 7), with
loadings of each variable being positive and relatively
equal (Table 8), thus representing overall size. Principal
component 2 (PC2) accounts for 11Æ72 per cent of the
total variation; however, the most significant loading on
PC2 is the cranidial border length (G1), suggesting that
the variation in PC2 is the result of preservation. Princi-
pal component 3 (PC3) represents 3Æ72 per cent of the
total variation in the sample. The cranidial border width
(J2) shows a significant positive loading on PC3, reflecting
the positive allometric growth of this character noted in
the bivariate analysis. Principal component 4 (PC4)
accounts for 1Æ49 per cent of the total variation, with the
post-palpebral cranidial width (J5) having the greatest
influence, reflecting the strong allometry in this character.
Only 2Æ18 per cent of the total variation is divided among
the remaining three principal components (PC5–7) and,
hence, these are unlikely to represent any significant mor-
phological variation.
In order to assess the relationship of variables within
the sample that have not been significantly influenced by
compaction, a separate analysis was performed with the
exclusion of the cranidial border length (G1). As expected,
the principal components of this analysis subsequent to
PC1 were shown to be statistically similar to those in the
previous analysis subsequent to PC2, i.e. PC3 in the first
analysis is equivalent to PC2 in the second analysis, and
so on. Principal component 1 (PC1) accounts for 90Æ56
per cent of the total variation, demonstrating that overall
size accounts for an even more significant amount of the
TABLE 3 . Correlation matrix of cranidial characters of the
multivariate sample of Penarosa (n ¼ 48).
B1 F1 G1 J2 J5 K K5
B1 1
F1 0Æ971 1
G1 0Æ898 0Æ909 1
J2 0Æ962 0Æ97 0Æ875 1
J5 0Æ976 0Æ979 0Æ907 0Æ964 1
K 0Æ974 0Æ956 0Æ911 0Æ939 0Æ973 1
K5 0Æ98 0Æ972 0Æ916 0Æ973 0Æ976 0Æ975 1
TABLE 4 . Eigenvalues and percentage of variance accounted
for each principal component for Penarosa.
Principal
component 1 2 3 4 5 6 7
Eigenvalue 6Æ704 0Æ15 0Æ063 0Æ033 0Æ022 0Æ016 0Æ012
Percentage
variance
95Æ77 2Æ14 0Æ89 0Æ48 0Æ31 0Æ23 0Æ18
TABLE 5 . Eigenvectors for variables on principal components
in Penarosa.
Principal
component B1 F1 G1 J2 J5 K K5
1 0Æ381 0Æ381 0Æ361 0Æ377 0Æ382 0Æ379 0Æ383
2 0Æ178 0Æ118 )0Æ9 0Æ347 0Æ128 )0Æ001 0Æ086
3 0Æ261 )0Æ3 )0Æ196 )0Æ583 0Æ109 0Æ671 0Æ024
4 0Æ093 )0Æ596 0Æ058 0Æ372 )0Æ502 0Æ102 0Æ48
5 )0Æ823 )0Æ143 )0Æ044 0Æ21 0Æ335 0Æ368 0Æ097
6 0Æ238 )0Æ611 0Æ096 0Æ147 0Æ638 )0Æ306 )0Æ197
7 0Æ114 )0Æ041 0Æ081 0Æ439 )0Æ232 0Æ407 )0Æ752
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 489
total variation in the sample when diagenetically influ-
enced variables are removed. Principal component 2
(PC2) accounts for 4Æ38 per cent of the total variation.
Loadings of variables on PC2 are very similar to PC3 in
the first analysis, suggesting that the cranidial border
width (J2), occipital-glabellar length (B1) and occipital
lobe width (K) significantly influence this component.
Principal component 3 (PC3) accounts for 1Æ74 per cent
of the total variation, with loadings on variables that cor-
respond to those on PC4 in the first analysis, thus reflect-
ing influences by the post-palpebral cranidial width (J5),
occipital-glabellar length (B1), frontal area length (F1),
and, to a lesser extent, the occipital lobe width (K) and
anterior glabellar width (K5). Disregarding overall size
variation and diagenetic factors influencing principal
components, what seems to be driving the majority of
variation in other principal components are those varia-
bles that display strong allometric growth.
Bivariate plots of the first four principal components
from the first analysis (Text-fig. 8A–C) show a consider-
able amount of overlap among specimens. Unfortunately,
only 20 specimens from the type-suite and locality-based
collections were sufficiently well preserved to be included
in the analysis. Furthermore, over half of these specimens
represent N. narinosa or have come from associated local-
ities (e.g. M417 and M226; see Appendix, section 2). All
other species from the type-suite are represented by only
one or two specimens, therefore making it difficult to dif-
ferentiate discrete clusters that may represent distinct spe-
cies. Hence, there are insufficient grounds for recognizing
more than a single species of Nepea in the sample.
SYSTEMATIC PALAEONTOLOGY
Specimens used in this study come from the Commonwealth
Palaeontological Collection (prefix CPC) housed at Geoscience
Australia, Canberra, and the palaeontological collections of the
Queensland Museum, Brisbane (prefixes QMF and UQF). Mor-
phological terminology follows that of Whittington et al. (1997)
and Shaw (1957). Symbols in synonymy lists follow Matthews
A B
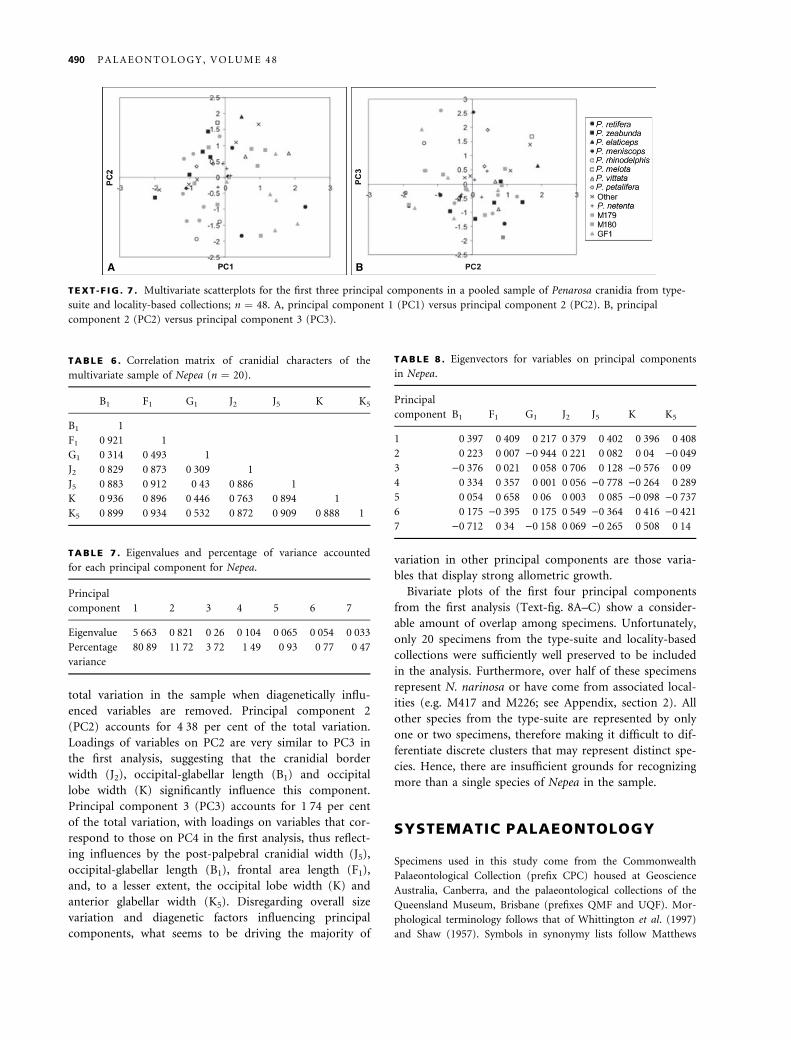
TEXT -F IG . 7 . Multivariate scatterplots for the first three principal components in a pooled sample of Penarosa cranidia from type-
suite and locality-based collections; n ¼ 48. A, principal component 1 (PC1) versus principal component 2 (PC2). B, principal
component 2 (PC2) versus principal component 3 (PC3).
TABLE 6 . Correlation matrix of cranidial characters of the
multivariate sample of Nepea (n ¼ 20).
B1 F1 G1 J2 J5 K K5
B1 1
F1 0Æ921 1
G1 0Æ314 0Æ493 1
J2 0Æ829 0Æ873 0Æ309 1
J5 0Æ883 0Æ912 0Æ43 0Æ886 1
K 0Æ936 0Æ896 0Æ446 0Æ763 0Æ894 1
K5 0Æ899 0Æ934 0Æ532 0Æ872 0Æ909 0Æ888 1
TABLE 7 . Eigenvalues and percentage of variance accounted
for each principal component for Nepea.
Principal
component 1 2 3 4 5 6 7
Eigenvalue 5Æ663 0Æ821 0Æ26 0Æ104 0Æ065 0Æ054 0Æ033
Percentage
variance
80Æ89 11Æ72 3Æ72 1Æ49 0Æ93 0Æ77 0Æ47
TABLE 8 . Eigenvectors for variables on principal components
in Nepea.
Principal
component B1 F1 G1 J2 J5 K K5
1 0Æ397 0Æ409 0Æ217 0Æ379 0Æ402 0Æ396 0Æ408
2 0Æ223 0Æ007 )0Æ944 0Æ221 0Æ082 0Æ04 )0Æ049
3 )0Æ376 0Æ021 0Æ058 0Æ706 0Æ128 )0Æ576 0Æ09
4 0Æ334 0Æ357 0Æ001 0Æ056 )0Æ778 )0Æ264 0Æ289
5 0Æ054 0Æ658 0Æ06 0Æ003 0Æ085 )0Æ098 )0Æ737
6 0Æ175 )0Æ395 0Æ175 0Æ549 )0Æ364 0Æ416 )0Æ421
7 )0Æ712 0Æ34 )0Æ158 0Æ069 )0Æ265 0Æ508 0Æ14
490 P A L A E O N T O L O G Y , V O L U M E 4 8

(1973). Photographed specimens have been coated with black,
water-based ink and then coated with ammonium chloride.
Stereo pairs were photographed using a specially constructed
platform, with a tilt of 4Æ5 degrees either side of the horizontal.
Class TRILOBITA Walch, 1771
Order PTYCHOPARIIDA Swinnerton, 1915
Suborder PTYCHOPARIINA Richter, 1932
Superfamily PTYCHOPARIOIDEA Matthew, 1887
Family NEPEIDAE Whitehouse, 1939
Revised diagnosis. Cranidium strongly vaulted, convex (sag.
and tr.); glabella gently to moderately tapered with two to
three pairs of deep, lateral glabellar furrows, glabellar length
(sag.) between 50 and 70 per cent cranidial length; medial
preglabellar boss occupying entire length (sag.) of preglabel-
lar field; wide fixigenae, post-palpebral cranidial width (tr.)
between 105 and 125 per cent cranidial length (sag.); eye rid-
ges well developed and bifurcate; posterolateral projections
of fixigenae well developed and strongly downsloping. Lib-
rigenae with long genal spines. Thorax with at least 20 seg-
ments; wide (tr.) pleurae. Pygidium small (micropygous).
Remarks. The history of the higher-level classification of
the Nepeidae is brief and vague. Whitehouse (1939)
erected the family, placing it in the Ptychopariina, but
did not discuss the classification any further, except to
mention that the group is closely related to the Ptycho-
pariidae. However, he did suggest that several genera,
including Acrocephalops, Alokistocare, Eldoradia and Bola-
spis, be placed in the Nepeidae, based primarily on the
possession of a preglabellar boss. Lochman-Balk (in Harr-
ington et al. 1959) elevated the Nepeidae to superfamilial
level (i.e. the Nepeacea), but with no justification. Loch-
man-Balk did not, however, recognize the genera that
Whitehouse (1939) considered members of the group,
therefore treating Nepea as being the sole representative
of the family, and indeed the superfamily. Opik (1963a)
reserved the use of Nepeacea, and gave a brief history of
the classification of the Nepeidae. Subsequently, Opik
(1967) regarded the Nepeacea as consisting of two famil-
ies, the Nepeidae and Menomoniidae, and suggested that
the genus Bolaspidella be transferred from the Menomo-
niidae to the Nepeidae. Opik (1970) later retained his
earlier views of the Nepeacea, and further suggested the
possible inclusion of the genus Periommella in the Nepei-
dae. I share the views of Robison (1988) and Pratt
(1992) in that the Nepeidae bears little in common with
the menomoniids, based on overall morphology, biogeog-
raphy and biostratigraphy, and that it should only
A B
C
TEXT -F IG . 8 . Multivariate scatterplots for the first four principal components in a pooled sample of Nepea cranidia from type-suite
and locality-based collections; n ¼ 20. A, principal component 1 (PC1) versus principal component 2 (PC2). B, principal component
2 (PC2) versus principal component 3 (PC3). C, principal component 3 (PC3) versus principal component 4 (PC4).
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 491

contain the genera Nepea, Penarosa, Loxonepea and Fer-
enepea.
The higher-level classification of the Nepeidae presented
here follows that of Fortey (1997) in placing the group
within the superfamily Ptychoparioidea. As discussed by
Fortey, the definition of the Ptychopariina is problematic,
arising from issues such as the conservative and intergra-
dational bauplan of taxa, and the absence of distinctive
morphological characters, stratigraphical and geographical
criteria that have resulted in excessive splitting, and a pro-
liferation of taxa with imprecise morphological definitions.
For these reasons, phylogenetic relationships within the
Ptychopariina are poorly understood. Therefore, it is best
to maintain a conservative approach when dealing with
groups (especially superfamilies and families) within the
Ptychopariina, at least until a comprehensive phylogenetic
treatment can be conducted.
Genus NEPEA Whitehouse, 1939
1939 Nepea Whitehouse, p. 211.
1963 Nepea Whitehouse; Opik, p. 313.
1959 Nepea Whitehouse; Lochman-Balk in Harrington et al.,
p. O251.
1970 Nepea Whitehouse; Opik, p. 9.
1970 Folliceps; Opik, p. 20.
Type species. Nepea narinosa Whitehouse, 1939, p. 212.
Other species. Nepea tonsillata Opik, 1970; Nepea nans (Opik,
1970); Nepea xinjiangensis Xiang and Zhang, 1985; Nepea brevica
Xiang and Zhang, 1985.
Revised diagnosis. Cranidium semicircular, length (sag.)
75 per cent post-palpebral cranidial width (tr.); glabella
gently tapered, width (tr.) at anterior margin 60 per cent
occipital ring width, glabellar length (sag.) 55 per cent
sagittal cranidial length, S1–S2 directed posteriorly, S3
directed laterally; lateral occipital lobes separated from
posterior cranidial border by shallow furrow; preglabellar
boss trapeziform; eye ridge slightly arched anteriorly,
directed slightly anteriorly abaxially.
Remarks. Comparison of Chinese species of Nepea erected
by Xiang and Zhang (1985), N. xinjiangensis and N. brev-
ica with Australian species is difficult because the Chinese
species represent small individuals, possibly early holaspid
stages, with the largest specimen having a maximum cra-
nidial length of 3 mm. Furthermore, N. xinjiangensis and
N. brevica are probably conspecific, based on their identi-
cal cranidial morphology and co-occurrence in the Ken-
say Formation. Further investigation of these species is
required, but that is not attempted here.
Nepea narinosa Whitehouse, 1939
Plate 1, figures 1–12; Plate 2, figures 1–8
v*1939 Nepea narinosa Whitehouse, p. 212, pl. 22, figs 15a–b,
16.
1959 Nepea narinosa Whitehouse; Lochman-Balk in Harr-
ington et al., p. O251, fig. 188.
v1970 Nepea narinosa Whitehouse; Opik, p. 9, pl. 1,
figs 1a)4; pl. 2, figs 1–6; pl. 3, fig. 1.
v1970 Nepea anoxys Opik, p. 14, pl. 3, fig. 4a–b.
v1970 Nepea exserta Opik, p. 15, pl. 4, figs 1–3.
v?1970 Nepea sp. NA Opik, p. 16, pl. 3, fig. 5.
v1970 Nepea sp. NB Opik, p. 17, pl. 3, fig. 3a–b.
v1970 Nepea sp. cf. exserta Opik, p. 17, pl. 3, fig. 2.
v1970 Nepea avara Opik, p. 18, pl. 6, figs 1a)5.
Holotype. UQF3335, internal mould of cranidium from the Split
Rock Sandstone, locality M417 (Pl. 1, fig. 1).
Other material. Thirty-nine cranidia: paratype, UQF3336;
CPC4740–4741, 7272–7274, 7276, 7278, 7280–7284, 7286–7289,
37065–37078, 37105–37111, 37115; three librigenae: CPC4742,
7275, 7277; one partial exoskeleton: CPC7279.
Revised diagnosis. Nepea with occipital ring not elevated
above glabella; dorsal elevation of palpebral lobe increas-
ing posteriorly; long intergenal spine positioned on the
posterolateral corner of the posterolateral projection,
length approximately 130 per cent of the sagittal cranidial
length; posterior border furrow deep, wide (exsag.), rap-
idly expanding at distal end of furrow.
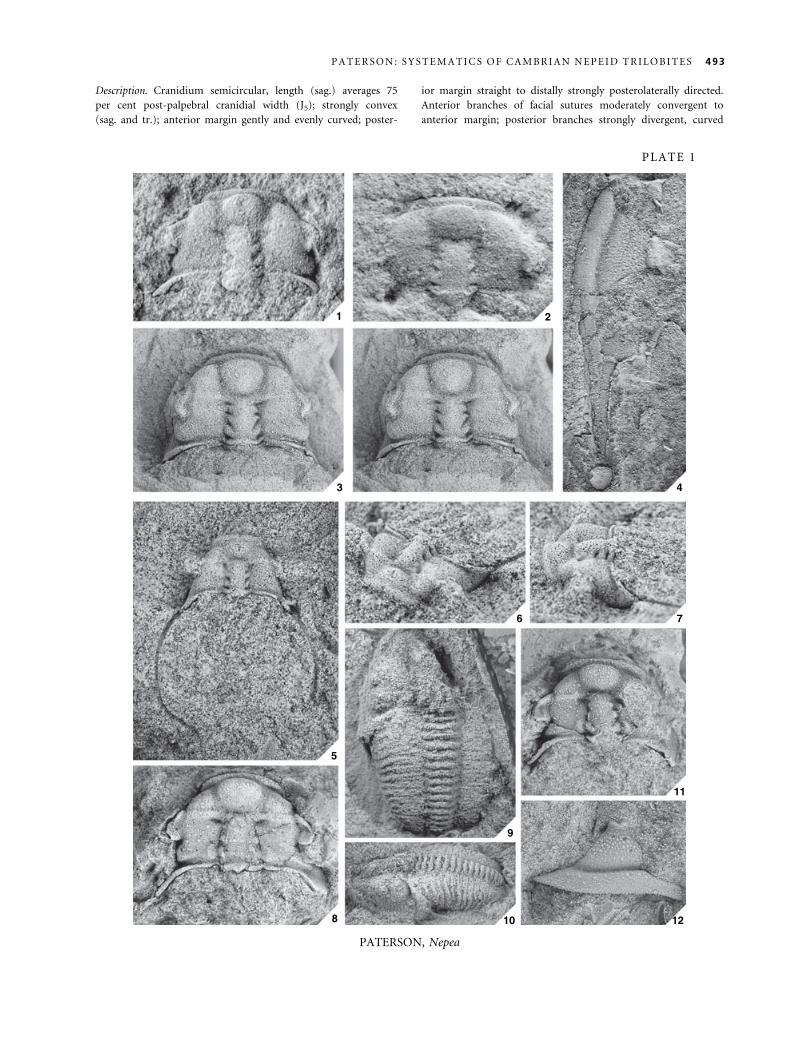
EXPLANATION OF PLATE 1
Figs 1–12. Nepea narinosa Whitehouse, 1939. 1, UQF3335, holotype, internal mould of cranidium; M417; · 4Æ5. 2, UQF3336, paratype,
internal mould of cranidium; M417; · 3. 3, CPC4741, internal mould of cranidium, stereo pair, holotype of Nepea anoxys Opik,
1970; M417; · 3. 4, CPC7275, latex cast of external mould of librigena; M226; · 4Æ5. 5–7, CPC7284, internal mould of cranidium,
holotype of Nepea exserta Opik, 1970; M344. 5, dorsal view; · 3. 6, anterolateral view; · 3Æ5. 7, oblique lateral view; · 3Æ5. 8,
CPC7278, internal mould of cranidium; M226; · 6Æ5. 9–10, CPC7279, latex cast of external mould of partial exoskeleton; M276. 9,
dorsal view; · 4Æ5. 10, oblique lateral view; · 3. 11, CPC7280, internal mould of cranidium; Mail Change Limestone, O’Shannassy
River, Morstone, north-west Queensland; · 4Æ5. 12, CPC7277, latex cast of external mould of librigena; M226; · 5.
492 P A L A E O N T O L O G Y , V O L U M E 4 8
Description. Cranidium semicircular, length (sag.) averages 75
per cent post-palpebral cranidial width (J5); strongly convex
(sag. and tr.); anterior margin gently and evenly curved; poster-
ior margin straight to distally strongly posterolaterally directed.
Anterior branches of facial sutures moderately convergent to
anterior margin; posterior branches strongly divergent, curved
1 2
3 4
5
6 7
8
9
10
11
12
PLATE 1
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 493
PATERSON, Nepea

to distal end of posterolateral projection. Glabella gently
tapered, width (tr.) at anterior margin (K5) averages 60 per
cent occipital lobe width (K); moderately convex (sag.),
strongly convex (tr.); frontal lobe slightly rounded; glabellar
length (B1) (sag.) averages 55 per cent sagittal cranidial length.
Axial furrow shallow, deepening anteriorly; preglabellar furrow
deep across entire width, and wide (sag. and exsag.). Lateral
glabellar furrows deep, S1 strongly directed posteriorly at
approximately 30 degrees to axial furrow, S2 subparallel to,
and shorter (tr.) than S1, S3 directed laterally, and shorter (tr.)
than S2. Occipital ring not elevated above glabella, strongly
convex (tr.), length (sag.) averages 10 per cent glabellar length
(B1); short occipital spine with broad (tr.) base; posterior mar-
gin straight medially to slightly anteriorly directed laterally; lat-
eral parts of occipital lobe directed slightly anteriorly, separated
from posterior cranidial border by shallow furrow. S0 curved
slightly anteriorly, wide (sag.), shallow medially to deep later-
ally. Frontal area length (sag.) averages 45 per cent cranidial
length. Preglabellar field convex, downsloping anteriorly; medial
trapeziform boss strongly convex (sag. and tr.), dorsal surface
flattened to slightly rounded, occupying entire length (sag.) of
preglabellar field, length averages 35 per cent cranidial length,
widest (tr.) anteriorly and becoming narrower posteriorly,
maximum width (tr.) averages 50 per cent cranidial length.
Anterior border convex (sag. and exsag.), flattened dorsally,
slightly narrowing laterally, length (sag.) approximately 20 per
cent frontal area length. Anterior border furrow shallow. Fixi-
gena convex (tr.), preocular area strongly downsloping. Palpe-
bral lobe crescentic, length (exsag.) approximately 50 per cent
glabellar length (B1); anterior end located slightly anterior of
anterior margin of glabella, posterior end located opposite L2,
dorsal elevation of palpebral lobe increasing posteriorly; palpe-
bral furrow shallow. Eye ridge of moderate relief, slightly
arched anteriorly, bifurcates near axial furrow; eye ridge direc-
ted slightly anteriorly abaxially; proximal end connected to
frontal lobe, distal end separated from palpebral lobe by palpe-
bral furrow. Palpebral and postocular areas of fixigena down-
sloping adaxially. Baccula elliptical, moderately developed.
Posterolateral projection of fixigena strongly downsloping; long
intergenal spine positioned on the posterolateral corner of the
posterolateral projection, length approximately 130 per cent of
the sagittal cranidial length. Posterior border convex (exsag.),
moderately expanding distally; border furrow deep, wide (ex-
sag.), slightly directed posterolaterally, rapidly expanding at di-
stal end of furrow.
Librigena long and narrow, width averages 25 per cent length
with spine; lateral margin moderately curved. Genal field tra-
pezoidal, width averages 70 per cent librigenal width. Border
moderately convex (tr.), width averages 30 per cent librigenal
width; lateral border furrow deep and wide (tr.). Genal spine
with narrow base, curved adaxially, occupying approximately 40
per cent total length of librigena.
Rostral plate and hypostome unknown.
Thorax with at least 20 thoracic segments preserved on avail-
able specimen (CPC7279), moderately decreasing in width poste-
riorly. Axial furrows well defined, of moderate depth. Axial ring
width (tr.) approximately 65 per cent width of pleura. Thoracic
pleura projecting horizontally to fulcrum, then downward and
posterolaterally to distal end; pleural furrow short (exsag.); pleu-
ral end rounded.
Pygidium unknown.
Exoskeletal surface densely covered with granules, including
the dorsal surface of the palpebral lobes; frontal area of the cra-
nidium and genal field of librigena covered in genal caeca.
Remarks. Characters that Opik (1970) frequently used to
differentiate Nepea narinosa from other species of Nepea
include: (1) the size and shape of the preglabellar boss; (2)
size and structure of the palpebral lobes; and (3) the
development of the eye ridges. With regards to the size
and shape of the preglabellar boss in Nepea, bivariate ana-
lysis of the linear dimensions of the boss (PBL and PBW,
see Text-fig. 6G–H) has shown that the relative length and
width of the preglabellar boss cannot be used to discrim-
inate species; this apparent plasticity in the size of the boss
is the result of compaction. Opik (1970) differentiated
species of Nepea as either having a circular boss (e.g.
N. exserta, N. narinosa and N. tonsillata) or a trapezoidal
boss (e.g. N. anoxys and N. avara). However, in examin-
ing supplementary material from localities containing spe-
cies with a ‘circular boss’ sensu Opik (e.g. M417, M226,
M344), the majority of specimens exhibit a subtrapezoidal
boss. In fact, some specimens of N. narinosa illustrated by
Opik (1970, pl. 1, fig. 3; pl. 2, fig. 2; pl. 3, fig. 1) show a
subtrapezoidal boss. Therefore, the shape of the preglabel-
lar boss in Nepea shows significant intra- and interpopula-
tional variation, and hence is an unreliable diagnostic
character for species within the genus.
The palpebral lobes of species from Opik’s (1970) type-
suite do vary considerably; however, this variation can be
explained by factors of preservation and ontogeny. Opik
(1963a, 1970) described the unusual structure of the palp-
ebral lobe in Nepea narinosa, which he termed the ‘pixie
ear’. The palpebral lobe was given this name because the
anterior portion of the lobe is narrow and the posterior
portion of the lobe in many specimens slopes up abaxially
(Pl. 1, figs 8, 11). However, there are specimens from sev-
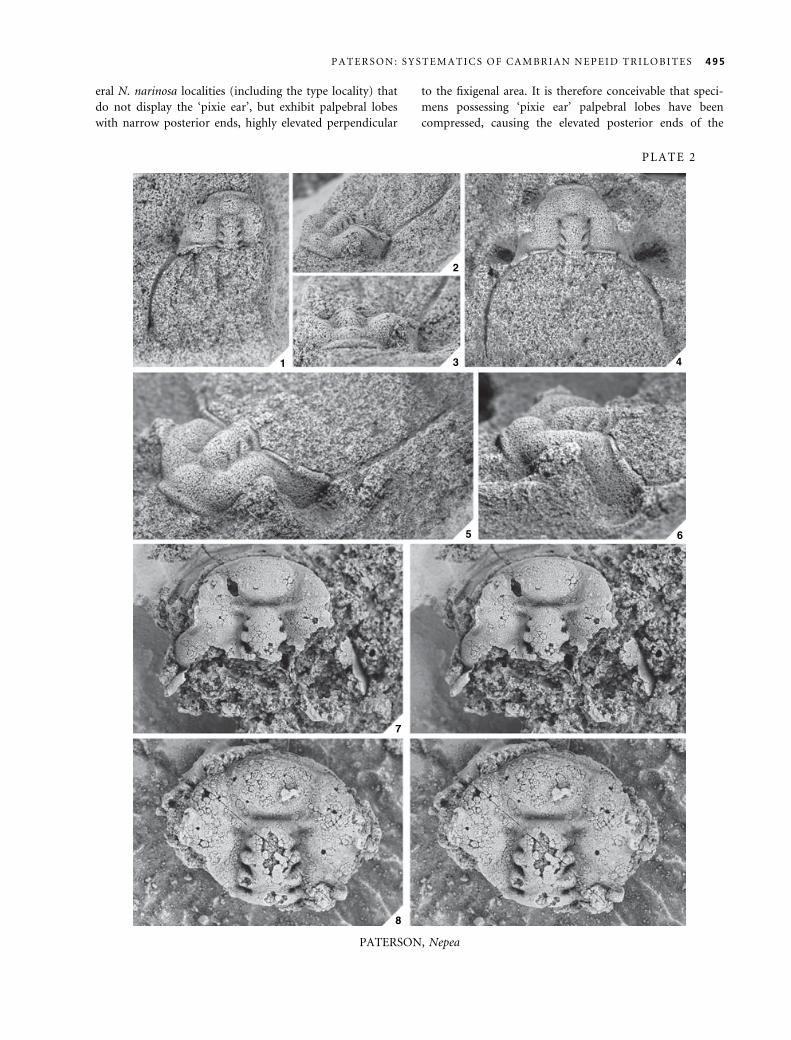
EXPLANATION OF PLATE 2
Figs 1–8. Nepea narinosa Whitehouse, 1939. 1–3, CPC4740, internal mould of cranidium; M417. 1, dorsal view; · 4. 2, anterolateral
view; · 3Æ5. 3, anterior view; · 4Æ5. 4–6, CPC37107, internal mould of cranidium; M344. 4, dorsal view; · 3Æ5. 5, anterolateral
view; · 5. 6, lateral view; · 5Æ5. 7, CPC7286, silicified cranidium, stereo pair, holotype of Nepea avara Opik, 1970; D108; · 5.
8, CPC7287, silicified cranidium, stereo pair; D108; · 6Æ5.
494 P A L A E O N T O L O G Y , V O L U M E 4 8
eral N. narinosa localities (including the type locality) that
do not display the ‘pixie ear’, but exhibit palpebral lobes
with narrow posterior ends, highly elevated perpendicular
to the fixigenal area. It is therefore conceivable that speci-
mens possessing ‘pixie ear’ palpebral lobes have been
compressed, causing the elevated posterior ends of the
1
2
3 4
5 6
7
8
PATERSON, Nepea
PLATE 2
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 495

lobes to become bent over. The relative size of the palpe-
bral lobe among species of Nepea is simply explained by
holaspid ontogeny. Because of the deformed nature or
absence of palpebral lobes in many specimens, it is not
possible to assess the growth pattern of the lobes bivari-
ately. However, in comparing the relative length of the
palpebral lobes with the overall length of the cranidium,
smaller specimens of Nepea possess large lobes, and larger
specimens display relatively smaller lobes, suggesting an
allometric growth pattern. It is unlikely that the palpebral
lobes are of taxonomic significance in Nepea.
Opik (1970) also utilized the structure of the eye ridges
to differentiate species of Nepea. Most of Opik’s species
(i.e. N. avara, N. exserta, N. narinosa and N. tonsillata)
possess eye ridges that are almost perpendicular to the
axial plane. However, Opik (1970, p. 18) differentiated
N. avara from other species by its ‘abaxially fading ocular
ridges’. The ‘fading’ of the abaxial portion of the eye
ridge in N. avara is simply a result of preservation. The
silicified type specimens from locality D108 illustrated by
Opik (1970, pl. 6, figs 1a–c, 2) display extensive beekite
rings on the surface, which commonly obscure details of
ornament in silicified fossils, suggesting that the eye rid-
ges may have been lost or diminished during silicification
(Pl. 2, figs 7–8). Furthermore, examination of specimens
illustrated by Opik (1970, pl. 6, figs 3–5) and supple-
mentary material from localities D54 and D95 show eye
ridges that extend from the anterolateral corners of
the glabella to the palpebral lobes, demonstrating that
N. avara does not possess eye ridges that ‘fade’ abaxially.
Opik distinguished N. anoxys from other species of Nepea
by its oblique eye ridges with divergent branches (Pl. 1,
fig. 3). Unfortunately, the holotype of N. anoxys
(CPC4741) is the only specimen of this species, thus mak-
ing observations of intraspecific variation impossible.
However, the type locality of N. anoxys is also the type
locality for N. narinosa (i.e. M417), and no other speci-
mens from locality M417 display oblique eye ridges with
divergent branches. The holotype of N. anoxys also has a
flattened appearance, implying that the eye ridges have
been skewed during compaction. Therefore, N. anoxys is
undoubtedly a synonym of N. narinosa.
Placement of Nepea sp. NA in synonymy with N. nari-
nosa is speculative. The bivariate analysis reveals that
Nepea sp. NA exhibits a slightly more tapered glabella
and narrower (tr.) preglabellar boss than in other species
of Nepea. Unfortunately, the posterior cranidial border of
Nepea sp. NA (Opik 1970, pl. 3, fig. 5) is poorly pre-
served and possibly flattened, and the distal portions of
the posterolateral projections of the fixigena are covered
by chert, thus making comparisons with other species
impossible.
Opik (1970) noted that the long glabella and short
frontal area distinguished Nepea sp. NB from N. narinosa,
but bivariate analysis does not support this assertion (see
Text-fig. 6A). As a consequence, Nepea sp. NB is placed
in synonymy with N. narinosa.
Stratigraphical occurrence. Middle Cambrian (Undillan–Boomer-
angian). Doryagnostus deltoides Zone, V-Creek Limestone, local-
ity M41. Goniagnostus nathorsti Zone, Split Rock Sandstone,
localities M133, M141, M226, M276, M344, M417, M421. Lejo-
pyge laevigata Zone, Steamboat Sandstone, localities D54, D95,
D108.
Nepea tonsillata Opik, 1970
Text-figure 9A–D
v*1970 Nepea tonsillata Opik, p. 15, pl. 5, figs 1–3.
Holotype. CPC7285, cranidium from the V-Creek Limestone,
locality M54 (Text-fig. 9A–D).
Revised diagnosis. Nepea with occipital ring elevated
above remainder of glabella; dorsal elevation of palpebral
lobe uniform from anterior to posterior; short intergenal
spine positioned on the posterior cranidial border posteri-
orly adjacent to the distal end of the posterior border fur-
row, length equal to sagittal length of glabella; posterior
border furrow deep, wide (exsag.), weakly expanding at
distal end of furrow.
Remarks. Although the holotype of N. tonsillata
(CPC7285) represents the only specimen of this species,
the position of the intergenal spine in this specimen dif-
fers significantly from specimens of N. narinosa as to war-
rant specific differentiation. The intergenal spine in
N. tonsillata is positioned on the posterior cranidial bor-
der posteriorly adjacent to the distal end of the posterior
border furrow, whereas the intergenal spine in N. narin-
osa is positioned on the posterolateral corner of the post-
erolateral projection. Moreover, the length of the
intergenal spine in N. tonsillata (see Text-fig. 9B–C) is
considerably shorter than that of N. narinosa (see Pl. 1,
fig. 5; Pl. 2, figs 1–2, 4–5). Examination of the holotype
of N. tonsillata shows that the intergenal spine tapers
enough to indicate that its total length would be equal to
that of the glabellar length (sag.), whereas in N. narinosa
the length of the intergenal spine is 130 per cent of the
sagittal cranidial length.
Stratigraphical occurrence. Middle Cambrian (Undillan). Ptychag-
nostus punctuosus–Goniagnostus nathorsti zones, V-Creek Lime-
stone, locality M54.
496 P A L A E O N T O L O G Y , V O L U M E 4 8
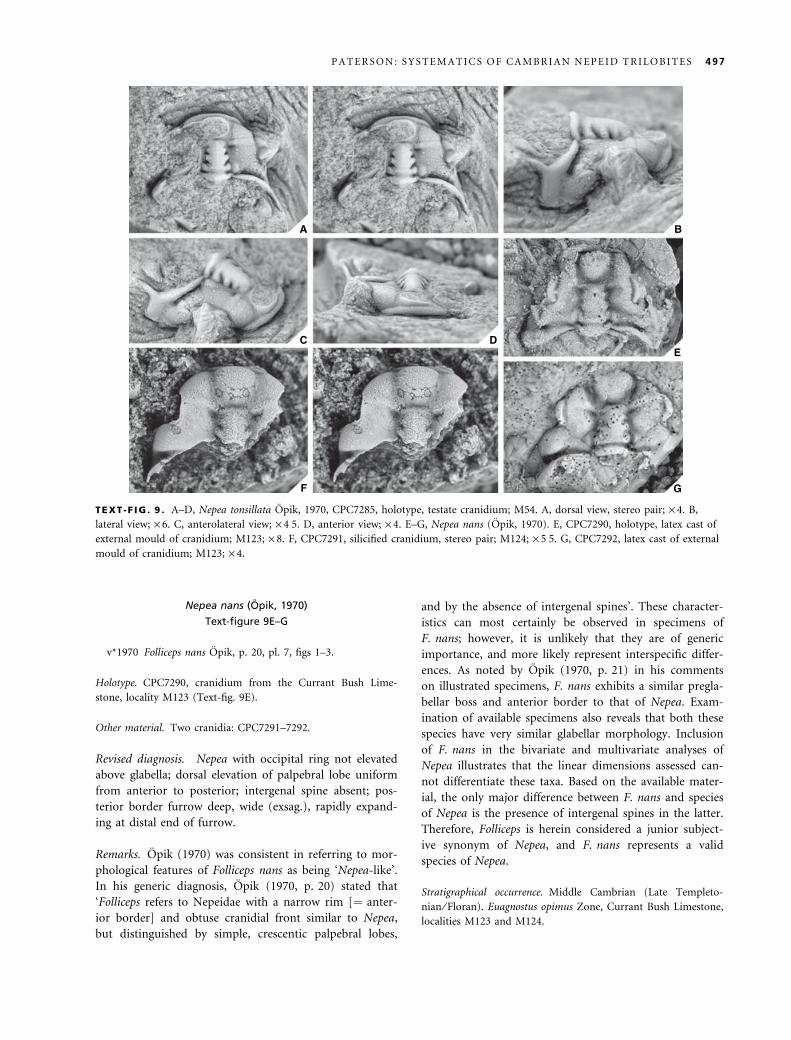
Nepea nans (Opik, 1970)
Text-figure 9E–G
v*1970 Folliceps nans Opik, p. 20, pl. 7, figs 1–3.
Holotype. CPC7290, cranidium from the Currant Bush Lime-
stone, locality M123 (Text-fig. 9E).
Other material. Two cranidia: CPC7291–7292.
Revised diagnosis. Nepea with occipital ring not elevated
above glabella; dorsal elevation of palpebral lobe uniform
from anterior to posterior; intergenal spine absent; pos-
terior border furrow deep, wide (exsag.), rapidly expand-
ing at distal end of furrow.
Remarks. Opik (1970) was consistent in referring to mor-
phological features of Folliceps nans as being ‘Nepea-like’.
In his generic diagnosis, Opik (1970, p. 20) stated that
‘Folliceps refers to Nepeidae with a narrow rim [¼ anter-
ior border] and obtuse cranidial front similar to Nepea,
but distinguished by simple, crescentic palpebral lobes,
and by the absence of intergenal spines’. These character-
istics can most certainly be observed in specimens of
F. nans; however, it is unlikely that they are of generic
importance, and more likely represent interspecific differ-
ences. As noted by Opik (1970, p. 21) in his comments
on illustrated specimens, F. nans exhibits a similar pregla-
bellar boss and anterior border to that of Nepea. Exam-
ination of available specimens also reveals that both these
species have very similar glabellar morphology. Inclusion
of F. nans in the bivariate and multivariate analyses of
Nepea illustrates that the linear dimensions assessed can-
not differentiate these taxa. Based on the available mater-
ial, the only major difference between F. nans and species
of Nepea is the presence of intergenal spines in the latter.
Therefore, Folliceps is herein considered a junior subject-
ive synonym of Nepea, and F. nans represents a valid
species of Nepea.
Stratigraphical occurrence. Middle Cambrian (Late Templeto-
nian ⁄ Floran). Euagnostus opimus Zone, Currant Bush Limestone,
localities M123 and M124.
A B
C DE
F G
TEXT -F IG . 9 . A–D, Nepea tonsillata Opik, 1970, CPC7285, holotype, testate cranidium; M54. A, dorsal view, stereo pair; · 4. B,
lateral view; · 6. C, anterolateral view; · 4Æ5. D, anterior view; · 4. E–G, Nepea nans (Opik, 1970). E, CPC7290, holotype, latex cast of
external mould of cranidium; M123; · 8. F, CPC7291, silicified cranidium, stereo pair; M124; · 5Æ5. G, CPC7292, latex cast of external
mould of cranidium; M123; · 4.
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 497

Genus PENAROSA Opik, 1970
1970 Penarosa Opik, p. 24.
1972 Trinepea Palmer and Gatehouse, p. D25.
1977 Penarosa Opik; Jell, p. 119.
1978 Penarosa Opik; Jell and Robison, p. 17.
Type species. Penarosa retifera Opik, 1970, p. 25.
Other species. Penarosa elaticeps Opik, 1970; Penarosa rhinodel-
phis Opik, 1970; Penarosa trinodus (Palmer and Gatehouse,
1972); Penarosa netenta Jell, 1977.
Revised diagnosis. Cranidium trapeziform or subtriangu-
lar, length (sag.) 95 per cent post-palpebral cranidial width
(tr.); glabella gently tapered, width (tr.) at anterior margin
75 per cent occipital ring width, glabellar length (sag.) 50
per cent cranidial length, S1 and S2 directed posteriorly,
S3 directed anteriorly; lateral occipital lobes separated
from posterior cranidial border by shallow or deep furrow;
preglabellar boss subtriangular; eye ridge slightly arched
anteriorly, almost perpendicular to axial line.
Remarks. Jell (1977, p. 119) discussed the taxonomic sta-
tus of Trinepea, concluding that it is a junior synonym of
Penarosa. The morphogenesis of Penarosa has been des-
cribed in detail by Opik (1970, pp. 43–44) and will not
be elaborated upon here.
Penarosa retifera Opik, 1970
Plate 3, figures 1–12; Text-figure 10A–E
v*1970 Penarosa retifera Opik, p. 25, pl. 8, figs 1a)2b; pl. 9,
figs 1–2, 4.
v1970 Penarosa sp. PA aff. retifera Opik, p. 29, pl. 9, fig. 5a–
b.
v1970 Penarosa vittata Opik, p. 29, pl. 10, fig. 2; pl. 15,
figs 1a)2.
v1970 Penarosa sp. PB aff. vittata Opik, p. 30, pl. 15, fig. 3.
v1970 Penarosa petalifera Opik, p. 37, pl. 11, fig. 4; pl. 16,
figs 1a–c, 3a)5; p. 38, text-fig. 13.
v1970 Penarosa melota Opik, p. 40, pl. 14, fig. 1a–b.
v1970 Penarosa sp. nov. PD aff. vittata Opik, p. 42, pl. 7,
fig. 6.
v1970 Penarosa sp. nov. PE Opik, p. 42, pl. 7, fig. 7.
1978 Penarosa retifera Opik; Jell and Robison, p. 17, pl. 4,
figs 17–19.
Holotype. CPC7297, cranidium from the Currant Bush Lime-
stone, locality M180 (Pl. 3, figs 1–6).
Other material. Forty-eight cranidia: CPC7295–7296, 7298–7299,
7302–7303, 7312, 7316, 7319–7322, 7324–7326, 7333–7335,
37079, 37091–37093, 37095–37104, 37116–37118, 37120–37126,
37128–37129, 37132–37136; seven librigenae: CPC7304–7305,
37094, 37119, 37127, 37130–37131; two partial exoskeletons:
CPC7300, 7318.
Revised diagnosis. Penarosa with trapeziform cranidium;
short posteromedial occipital spine with broad (tr.) base;
lateral occipital lobe separated from posterior cranidial
border by shallow furrow; baccula elliptical, moderately
developed, length (exsag.) equal to length of L1; anterior
border moderately to strongly concave, strongly upturned,
narrows laterally; anterior border furrow very shallow and
poorly defined; posterior border furrow narrow (exsag.),
less than width (sag.) of occipital ring. Cranidial surface,
except for the palpebral lobes and posterior border fur-
row, covered in granules of varying sizes; glabella and
occipital ring densely covered in small granules; proximal
portion of palpebral and postocular areas of fixigena cov-
ered in moderately spaced, medium-sized granules,
becoming smaller and more closely spaced abaxially; post-
erolateral projection of fixigena covered in small granules
and genal caeca; frontal area evenly covered in small and
medium-sized granules and genal caeca.
Description. Cranidium trapeziform, length (sag.) averages 95
per cent post-palpebral cranidial width (J5); strongly convex
(sag. and tr.); anterior margin moderately and evenly curved;
posterior margin straight, distally strongly posterolaterally
directed. Anterior branches of facial sutures subparallel to
about level with midlength of frontal area (exsag.), then con-
vergent to anterior margin; posterior branches strongly diver-
gent, curved to distal end of posterolateral projection then
moderately convergent to posterior margin. Glabella gently
tapered, width (tr.) at anterior margin (K5) averages 75 per
cent occipital lobe width (K); moderately convex (sag.),
strongly convex (tr.); frontal lobe bluntly rounded; glabellar
length (B1) (sag.) averages 50 per cent cranidial length. Axial
furrow deep, shallower posteriorly, and wide (tr.); preglabellar
EXPLANATION OF PLATE 3
Figs 1–12. Penarosa retifera Opik, 1970. 1–6, CPC7297, holotype, testate cranidium; M180. 1, dorsal view, stereo pair; · 2. 2, lateral
view; · 2Æ5. 3, oblique lateral view; · 2. 4, anterior view; · 2Æ5. 5, anterolateral view; · 2. 6, enlargement of glabella; · 4. 7,
CPC7304, testate librigena; M180; · 3. 8–10, CPC7300, testate partial exoskeleton; M393. 8, dorsal view; · 2. 9, enlargement
of articulated axial rings; · 4Æ5. 10, enlargement of articulated pleurae; · 4. 11, CPC7298, testate cranidium, stereo pair;
M186; · 3. 12, CPC7316, testate cranidium, stereo pair, holotype of Penarosa melota Opik, 1970; M124; · 3Æ5.
498 P A L A E O N T O L O G Y , V O L U M E 4 8
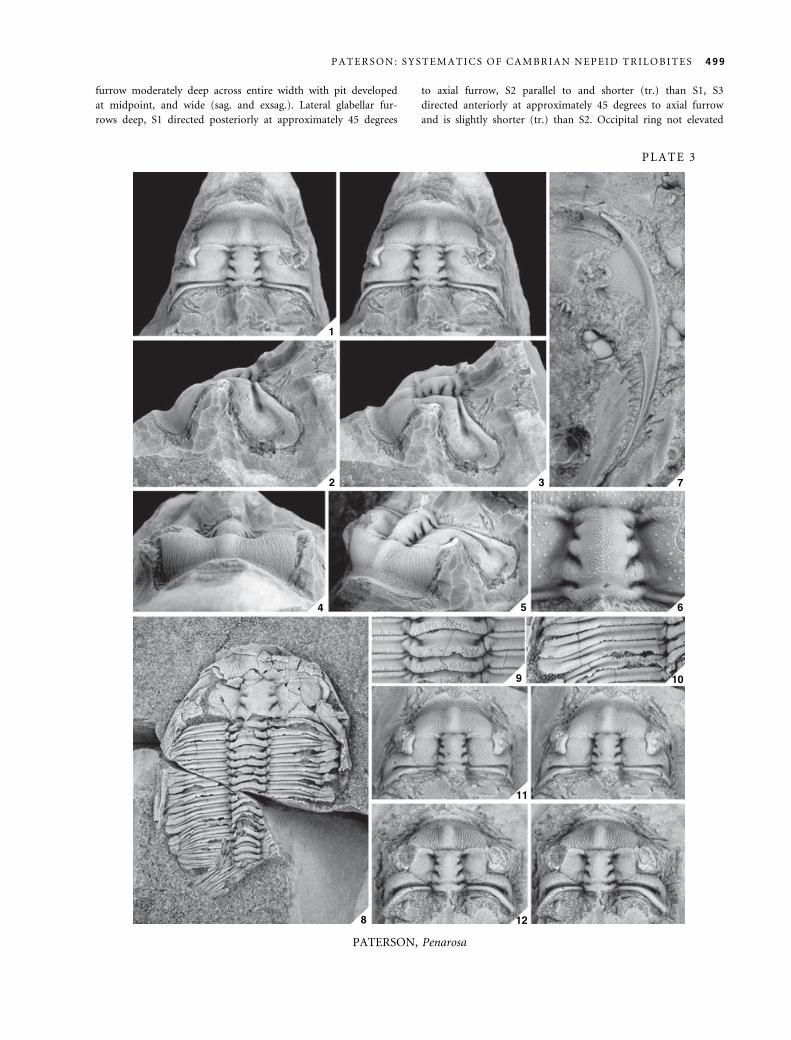
furrow moderately deep across entire width with pit developed
at midpoint, and wide (sag. and exsag.). Lateral glabellar fur-
rows deep, S1 directed posteriorly at approximately 45 degrees
to axial furrow, S2 parallel to and shorter (tr.) than S1, S3
directed anteriorly at approximately 45 degrees to axial furrow
and is slightly shorter (tr.) than S2. Occipital ring not elevated
1
2 3
4 5 6
7
8
9 10
11
12
PLATE 3
PATERSON, Penarosa
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 499

above remainder of glabella, strongly convex (tr.), length (sag.)
averages 15 per cent glabellar length (B1); short posteromedial
occipital spine with broad (tr.) base; posterior margin straight
medially and slightly anteriorly directed laterally; lateral parts
of occipital lobe directed slightly anteriorly, separated from
posterior cranidial border by shallow furrow. S0 curved slightly
anteriorly medially, wide (sag.), shallow medially to deep later-
ally. Frontal area length (sag.) averages 50 per cent cranidial
length (sag.). Preglabellar field convex, strongly anteriorly
downsloping, length (sag.) averages 60 per cent frontal area
length; medial subtriangular boss of moderate relief occupying
entire length (sag.) of preglabellar field, slightly extending onto
anterior border, posterior width slightly narrower (tr.) than
anterior glabellar width (K5), slightly tapering anteriorly. Anter-
ior border moderately to strongly concave, strongly upturned,
moderately narrowing laterally, length (sag.) averages 35 per
cent frontal area length. Anterior border furrow very shallow
and poorly defined. Fixigena moderately convex (tr.), preocular
area strongly downsloping. Palpebral lobe reniform, length (ex-
sag.) 50 per cent glabellar length (B1), width approximately 40
per cent lobe length; anterior end located opposite frontal lobe,
posterior end located opposite S1 or L1; palpebral furrow
moderately deep. Eye ridge of moderate relief, slightly arched
anteriorly, bifurcates near axial furrow; almost perpendicular to
sagittal line; proximal end separated by shallow axial furrow,
distal end separated from palpebral lobe by palpebral furrow.
Palpebral and postocular areas of fixigena downsloping adaxial-
ly. Baccula elliptical, moderately developed, length (exsag.)
equal to length of L1. Posterolateral projection of fixigena very
strongly downsloping, rapidly expanding abaxially. Posterior
border convex (exsag.), widens rapidly abaxially and is strongly
directed posterolaterally where the posterior border furrow
becomes shallow, and is then directed anteriorly at the distal
end of the posterolateral projection; posterior border furrow
deep, slightly directed posterolaterally, expanding abaxially, then
becomes shallow where posterior border becomes strongly
A
B
C
D E
TEXT -F IG . 10 . Penarosa retifera Opik, 1970. A–C, CPC7318, testate partial exoskeleton, holotype of Penarosa vittata Opik, 1970;
M124. A, dorsal view, stereo pair; · 2Æ5. B, enlargement of cranidium; · 3. C, enlargement of articulated thoracic segments; · 5. D,
CPC37116, testate cranidium, stereo pair; GF1; · 2Æ5. E, CPC37094, testate librigena; M180; · 4Æ5.
EXPLANATION OF PLATE 4
Figs 1–12. Penarosa elaticeps Opik, 1970. 1–4, CPC7311, holotype, testate cranidium. 1, dorsal view, stereo pair; · 2Æ5. 2, oblique lateral
view; · 3. 3, anterior view; · 3. 4, anterolateral view; · 2Æ5. 5, CPC7306, latex cast of external mould of partial exoskeleton;
· 3Æ5. 6–9, CPC7310, testate cranidium, holotype of Penarosa zeabunda Opik, 1970. 6, dorsal view, stereo pair; · 4Æ5. 7,
oblique lateral view; · 5. 8, anterior view; · 4Æ5. 9, anterolateral view; · 3Æ5. 10, CPC7309, testate cranidium, stereo pair;
· 6Æ5. 11, CPC7331, internal mould of meraspid cranidium; · 12Æ5. 12, CPC7332, internal moulds of meraspid cranidia;
· 11. All specimens from locality M179.
500 P A L A E O N T O L O G Y , V O L U M E 4 8
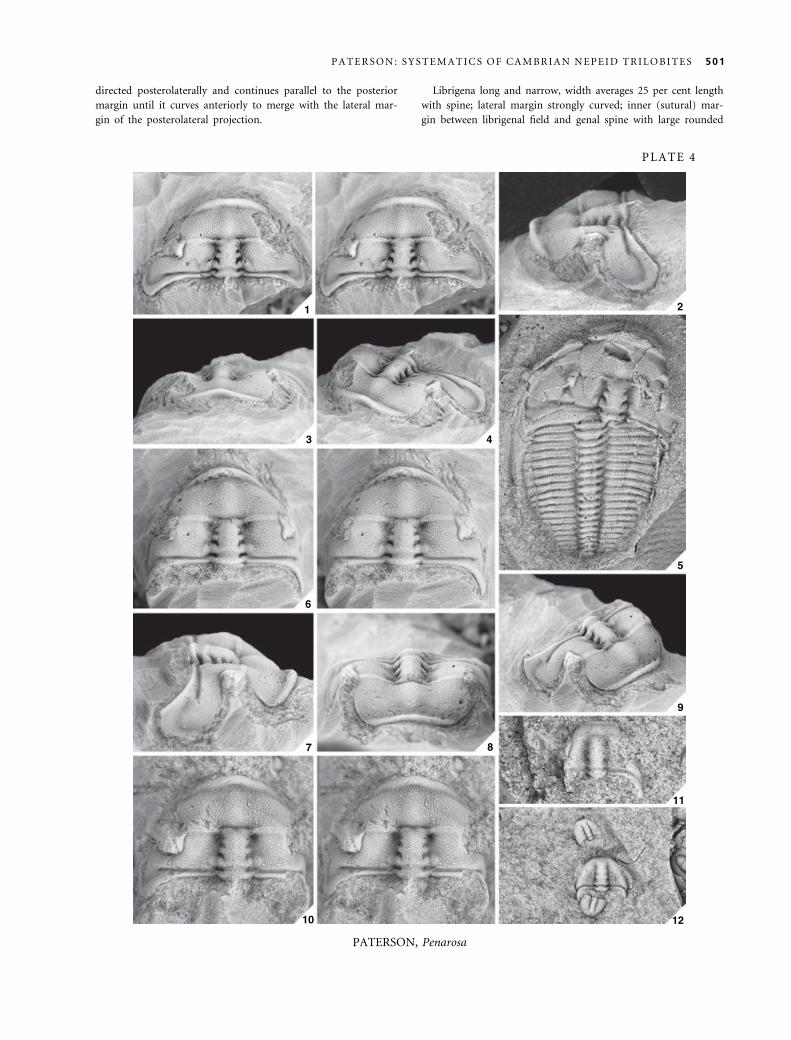
directed posterolaterally and continues parallel to the posterior
margin until it curves anteriorly to merge with the lateral mar-
gin of the posterolateral projection.
Librigena long and narrow, width averages 25 per cent length
with spine; lateral margin strongly curved; inner (sutural) mar-
gin between librigenal field and genal spine with large rounded
1 2
3 4
5
6
7 8
9
10
11
12
PLATE 4
PATERSON, Penarosa
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 501

embayment to accommodate the posterolateral projection of the
fixigena. Anterior part of dorsal surface developed as narrow
projection. Genal field trapezoidal, moderately convex, width
(tr.) averages 80 per cent librigenal width. Border flat to slightly
concave, width averages 20 per cent librigenal width; lateral bor-
der furrow very shallow and poorly defined. Genal spine with
narrow base, curved adaxially, occupying approximately 45 per
cent total length of librigena. Doublure wider anteriorly than
border.
Rostral plate and hypostome unknown.
Thorax with at least 26 thoracic segments preserved, moder-
ately decreasing in width posteriorly. Axial furrows well defined,
of moderate depth. Axial ring width (tr.) approximately 45 per
cent width of pleura on each thoracic segment, strongly convex
(tr.), straight medially to anterolaterally directed laterally; articu-
lating half ring relatively short (sag.), length slightly less than
that of axial ring, posterior margin straight; articulating furrow
well developed, wide (sag. and exsag.), lateral portion deepened
into an apodemal pit. Thoracic pleura projecting horizontally to
fulcrum, then downward and posterolaterally to distal end;
anterior pleural band of uniform width (exsag.) to fulcrum, then
expanding abaxially to pleural end; posterior pleural band of
uniform and equal width (exsag.) as anterior pleural band to ful-
crum, then expanding abaxially and becoming considerably
wider (exsag.) than the anterior pleural band at the pleural end;
pleural end bluntly pointed; pleural furrow moderately deep,
long (exsag.), uniform width to fulcrum, then tapering abaxially,
not quite reaching the pleural end.
Pygidium unknown.
Exoskeletal surface covered with granules of varying sizes;
palpebral lobes and posterior border furrow on the cranidium,
and the pleural furrows on the thoracic segments are smooth.
Glabella and occipital ring densely covered in small granules;
proximal portion of palpebral and postocular areas of fixigena
covered in moderately spaced, medium-sized granules, becoming
smaller and more closely spaced abaxially; posterolateral projec-
tions covered in small granules and genal caeca; frontal area
evenly covered in small and medium-sized granules and an anas-
tomosing network of genal caeca. Librigenal field densely cov-
ered in small and medium-sized granules and genal caeca;
librigenal border and spine covered with small, well-spaced gran-
ules. Axial ring and the anterior and posterior pleural bands of
the thoracic segments covered in small and medium-sized gran-
ules.
Remarks. Opik (1970) mentioned that Penarosa vittata
was closely related to P. retifera, but could be distin-
guished by its moderately vaulted cephalon, small
posterolateral projections, wide interocular cheeks,
absence of caeca on the anterior border, and the pres-
ence of a transverse ridge on the posterior part of the
anterior border. Specimens of P. vittata have been
slightly compressed, explaining why cranidia are moder-
ately vaulted. Moreover, compression of specimens also
provides an explanation for the transverse ridges in
P. vittata. Examination of specimens shows no evidence
of a ridge, but specimens display transverse kinks across
the anterior border as a result of compression (Text-
fig. 10B), creating the illusion of a ridge-like structure.
Furthermore, genal caeca are most certainly present on
the anterior border. Comparison of the posterolateral
projections of specimens of P. vittata with specimens of
equal size from locality M180 show no discernible differ-
ences in length or width. Bivariate analysis has demon-
strated that P. vittata and P. retifera cannot be
distinguished with regard to the relative width of the
‘interocular cheeks’ (i.e. post-palpebral cranidial width).
Lastly, P. vittata possesses a very short occipital spine,
moderately developed bacculae, lateral occipital lobes
separated from the posterior cranidial border by a shal-
low furrow, and has a pattern of ornamentation on the
cranidium identical to that of P. retifera. Therefore,
based on this evidence, P. vittata is herein considered a
junior subjective synonym of P. retifera.
Opik (1970) differentiated Penarosa petalifera from
P. retifera (and indeed the majority of other species of
Penarosa) by its ‘short’ frontal area. However, bivariate
analysis has demonstrated that species of Penarosa cannot
be distinguished on the relative length of the frontal area.
Opik also emphasized the ‘petaliform’ occipital spine as
a diagnostic character in P. petalifera. This unusual fea-
ture is obvious in the holotype of P. petalifera
(CPC7322); however, the ‘petaliform’ occipital spine is
less prominent in any other specimen of P. petalifera that
has the occipital ring preserved. Furthermore, the holo-
type of P. retifera exhibits a small occipital spine with a
broad (tr.) base, similar in shape to that of P. petalifera.
As the holotype of P. petalifera is less than half the size
of the holotype of P. retifera, it is possible that the for-
mer specimen represents a developmental stage in the
occipital spine, implying that the occipital spine displays
strong negative allometry during growth. Opik (1970)
EXPLANATION OF PLATE 5
Figs 1–12. Penarosa rhinodelphis Opik, 1970. 1–4, CPC7314, holotype, testate cranidium. 1, dorsal view, stereo pair; · 3Æ5. 2, lateral
view; · 4. 3, anterior view; · 5. 4, anterolateral view; · 4. 5, CPC37087, testate cranidium and partial exoskeleton; · 6Æ5. 6–9,
CPC7315, testate cranidium. 6, dorsal view, stereo pair; · 6. 7, oblique lateral view; · 6Æ5. 8, anterior view; · 8. 9,
anterolateral view; · 5Æ5. 10, CPC37083, testate cranidium; · 4Æ5. 11–12, CPC7313, testate cranidium, holotype of Penarosa
meniscops Opik, 1970. 11, dorsal view, stereo pair; · 3Æ5. 12, lateral view; · 4Æ5. All specimens from locality M179.
502 P A L A E O N T O L O G Y , V O L U M E 4 8
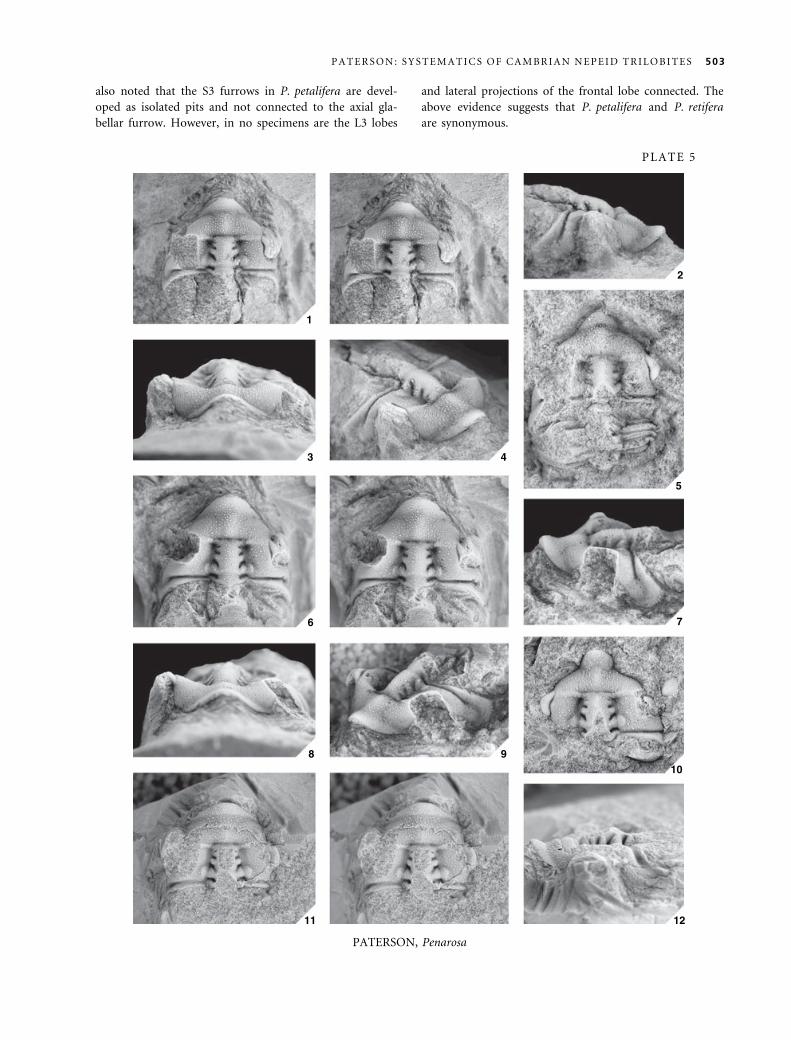
also noted that the S3 furrows in P. petalifera are devel-
oped as isolated pits and not connected to the axial gla-
bellar furrow. However, in no specimens are the L3 lobes
and lateral projections of the frontal lobe connected. The
above evidence suggests that P. petalifera and P. retifera
are synonymous.
1
2
3 4
5
6 7
8 910
11 12
PLATE 5
PATERSON, Penarosa
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 503

Opik (1970) used several characters to differentiate
Penarosa melota from other species of Penarosa. He
emphasized the narrow interocular cheeks, frontal area
length, anterior glabellar width, and occipital lobe width
as distinguishing features of the species; the bivariate ana-
lysis does not support this interpretation. Opik (1970,
p. 40) also used the ‘slender (not petaliform) occipital
spine’ as an additional diagnostic feature of P. melota.
However, examination of the holotype shows that the
occipital spine has a broad base, further supporting syn-
onymy with P. retifera.
Opik (1970) differentiated Penarosa sp. PA aff. reti-
fera, Penarosa sp. PB aff. vittata, Penarosa sp. nov. PD
aff. vittata and Penarosa sp. nov. PE based on linear
dimensions among other features. Inclusion of these spe-
cies in the bivariate and multivariate analyses demon-
strated that they cannot be distinguished by means of
the linear characters assessed. Opik (1970) also empha-
sized non-linear characters in these species; however,
many of these can be attributed to misinterpretation or
differences in preservation. For example, Opik noted
that Penarosa sp. PB aff. vittata and Penarosa sp. nov.
PD aff. vittata exhibit a transverse ridge on the anterior
border like that of P. vittata; however, these ridges rep-
resent kink structures resulting from compression, as is
the case for specimens of P. vittata. The only specimen
of Penarosa sp. PB aff. vittata (CPC7320) also displays
weakly impressed intercalated furrows on the lateral pro-
jections of the frontal lobe of the glabella; however, this
is common in some specimens of P. retifera. In the only
specimen of Penarosa sp. nov. PE (CPC7296), Opik only
recorded two pairs of glabellar furrows; however, close
examination of this specimen clearly shows three pairs
of glabellar furrows.
Opik (1970) observed four glabellar furrows in P. reti-
fera and the majority of other species of Penarosa;
however, examination of the holotype of P. retifera and
supplementary material from the type locality M180
shows that the majority of specimens do not exhibit
distinct S4 furrows or L4 lobes on the glabella. Some
specimens of Penarosa exhibit a diminutive depression
in one or both sides of the lateral projection of the
frontal lobe of the glabella (Pl. 3, figs 6, 11; Pl. 4,
figs 1, 6; Pl. 6, fig. 7), which may be interpreted as a
poorly developed intercalated furrow; however, this fea-
ture is not consistent in all specimens. Hughes (1993,
1994) has noted that intercalated furrows are common
in slightly compressed specimens; however, some speci-
mens of P. retifera that exhibit these intercalated fur-
rows show no obvious signs of compression. So,
because of their intermittent occurrence, it is unlikely
that the intercalated furrows in some specimens of
P. retifera and other species of Penarosa are of taxo-
nomic significance.
Jell (1977, p. 122) convincingly demonstrated that the
pygidium attributed to Penarosa retifera by Opik (1970,
p. 28, text-fig. 11; pl. 9, fig. 3) was incorrectly assigned,
and that it does not belong to the Nepeidae at all.
Comparison of the pygidium of P. netenta (QMF7062)
and that assigned to P. retifera by Opik (1970)
(CPC7301) shows that the overall morphology of the
two pygidia differ markedly. Firstly, the pygidium of
P. netenta is approximately 40 per cent as long as wide,
whereas specimen CPC7301 is around 75 per cent as
long as wide. Secondly, the pygidial axis of P. netenta is
60 per cent as wide as the pygidium at the midlength
of the axis, whereas in specimen CPC7301 this propor-
tion is only 33 per cent. The pygidium of P. netenta
can be differentiated further by its smooth pleural field
and absence of a border, whereas specimen CPC7301
possesses well-developed pleural ribs and a well-defined
border. Jell (1977) also discussed the similarity of the
pygidium of P. netenta to that of an undescribed species
of Nepea from Christmas Hills, north-west Tasmania; a
latex cast of the latter species was provided by Dr Jim
Jago to Dr Peter Jell for examination. Dr Jago has sub-
sequently provided me with photographs of a different
species from Native Track Tier, north-west Tasmania. I
concur with Jell (1977) that the pygidia of species of
Nepea are almost identical to that of P. netenta, repre-
senting very small simple plates, thus supporting the
conclusion that the pygidium illustrated by Opik
(1970) does not belong to P. retifera or any other
nepeid species.
The librigena assigned to Penarosa petalifera by Opik
(1970, pl. 16, fig. 2) is dubious. Opik (1970, p. 37)
noted in his description that ‘neither a rostral shield
nor a medial suture is present’. This is an unusual con-
dition for librigenae in Penarosa, because isolated librig-
enae of P. retifera and P. netenta exhibit sutural
terminations on the ventral doublure and anterior end
of the dorsal surface. Furthermore, it has been demon-
strated above that the cranidia of P. petalifera and
P. retifera are indistinguishable, and hence there is no
reason to believe the librigenae of these species should
differ in any way. Close examination of the librigenal
unit (CPC7323) shows no evidence of ventral sutures;
thus, it is unlikely that this specimen belongs to a spe-
cies of Penarosa.
Stratigraphical occurrence. Middle Cambrian (Late Templeto-
nian ⁄ Floran–Undillan). Acidusus atavus Zone, Arthur Creek
Formation, locality H4. Euagnostus opimus Zone, Age Creek
Formation, localities M157 and M160; Gowers Formation,
locality GF1; Currant Bush Limestone, localities M123,
M124, M180, M186, M393, UQL256, UQL463. Ptychagnostus
punctuosus Zone, Currant Bush Limestone, localities M28,
M130.
504 P A L A E O N T O L O G Y , V O L U M E 4 8

Penarosa elaticeps Opik, 1970
Plate 4, figures 1–2
v*1970 Penarosa elaticeps Opik, p. 34, pl. 11, fig. 3; pl. 12,
fig. 2a–d.
v1970 Penarosa zeabunda Opik, p. 31, pl. 10, figs 3–5; pl. 11,
figs 1a)3; pl. 12, fig. 1; pl. 17, figs 5–6; p. 33, text-
fig. 12.
Holotype. CPC7311, cranidium from the Currant Bush Lime-
stone, locality M179 (Pl. 4, figs 1–4).
Other material. Ten cranidia: CPC7307–7310, 7331–7332, 37080,
37082, 37088, 37090; one partial exoskeleton: CPC7306.
Revised diagnosis. Penarosa with trapeziform cranidium;
occipital spine absent; lateral occipital lobe separated
from posterior cranidial border by shallow furrow; baccu-
la subcircular, moderately well developed, length (exsag.)
slightly longer than L1; anterior border convex (sag. and
exsag.), moderately upturned, narrowing laterally; anterior
border furrow wide (sag.) and shallow; posterior border
furrow narrow (exsag.), less than width (sag.) of occipital
ring. Cranidial surface except for the palpebral lobes, pos-
terior border and border furrow covered in evenly spaced,
small and medium-sized granules; frontal area and post-
erolateral projection of fixigenae covered in genal caeca.
Remarks. Apart from linear dimensions that can be disre-
garded based on results of the morphometric analysis,
Opik (1970) differentiated P. zeabunda and P. elaticeps by
observing a ‘retral recess’ in the medial anterior border of
the latter species. However, examination of the holotype
specimen of P. elaticeps (Pl. 4, figs 1–4) shows no such
feature, as is the case for all other specimens from locality
M179. The ‘retral recess’ in the holotype of P. elaticeps
appears to be a misinterpretation of an illustration of the
specimen by Opik (1970, pl. 11, fig. 3). The oblique an-
terodorsal view at which the specimen was photographed
shows that the apparent ‘retral recess’, in fact, represents
the upturned nature of the cranidial border (Opik 1970,
pl. 12, fig. 2b–c). Opik (1970) also noted that the post-
erolateral projections of the fixigena are not strongly
downsloping in P. elaticeps as in other species of Penaro-
sa. This is easily explained by compaction of the individ-
ual as the posterolateral projections are asymmetrically
disposed and there are signs of breakage of the sclerite.
The meraspid cranidia of Penarosa zeabunda illustrated
by Opik (1970, p. 33, text-fig. 12; pl. 17, figs 5–6; see also
Pl. 4, figs 11–12) most likely belong to this species and
not to P. rhinodelphis (the other valid species from local-
ity M179), as they possess a relatively narrow (sag.) and
long (tr.) anterior border, like that of holaspid cranidia
of P. elaticeps. As shown in the bivariate analysis, small
holaspid cranidia of P. rhinodelphis display a very wide
(sag.) and short (tr.) anterior border.
Stratigraphical occurrence. Middle Cambrian (Late Templeto-
nian ⁄ Floran). Acidusus atavus Zone, Currant Bush Limestone,
locality M179.
Penarosa rhinodelphis Opik, 1970
Plate 5, figures 1–2
v*1970 Penarosa rhinodelphis Opik, p. 34, pl. 13, figs 1a)2c.
v1970 Penarosa meniscops Opik, p. 36 (partim), pl. 12,
fig. 3a–b (non pl. 14, fig. 2).
Holotype. CPC7314, cranidium from the Currant Bush Lime-
stone, locality M179 (Pl. 5, figs 1–4).
Other material. Six cranidia: CPC7313, 7315, 37083, 37085–
37087.
Revised diagnosis. Penarosa with subtriangular cranidium;
short posteriorly located occipital spine with broad (tr.)
base; lateral occipital lobe separated from posterior crani-
dial border by shallow furrow; baccula elliptical, large,
very well developed, longer than L1 (exsag.); anterior bor-
der subtriangular in outline, convex (sag.), strongly
upturned; anterior border furrow wide (sag.) and shallow;
posterior border furrow narrow (exsag.), less than width
(sag.) of occipital ring. Cranidial ornament identical to
P. elaticeps.
Remarks. The holotype specimen of Penarosa meniscops is
placed in synonymy with P. rhinodelphis primarily based
on possession of a convex (sag.), strongly upturned,
subtriangular anterior border and large, well-developed
bacculae. Opik (1970, p. 35) distinguished P. meniscops
from P. rhinodelphis by its ‘continuous frontal marginal
furrow, snoutless front and wide span of the frontal arc [i.e.
anterior border]’. Differences between these three features
most likely represent intraspecific ontogenetic variation. As
demonstrated in the bivariate analysis, dimensions of the
anterior border (i.e. length and width) in P. rhinodelphis
and P. meniscops (plus three other specimens from M179)
show continuous variation during growth (see Text-fig. 5).
This ontogenetic variation is also evident in the anterior
border furrow, with the furrow in some smaller specimens
being poorly developed medially (Pl. 5, figs 6, 10), and lar-
ger specimens displaying well-developed furrows along the
entire width of the border (Pl. 5, fig. 11).
Stratigraphical occurrence. Middle Cambrian (Late Templeto-
nian ⁄ Floran). Acidusus atavus Zone, Currant Bush Limestone,
locality M179.
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 505

Penarosa netenta Jell, 1977
Plate 6, figures 1–9
v1970 Penarosa meniscops Opik, p. 36 (partim), pl. 14, fig. 2
(non pl. 12, fig. 3a–b).
v1970 Penarosa sp. nov. PC Opik, p. 40, pl. 17, figs 3–4;
p. 41, text-fig. 14.
v*1977 Penarosa netenta Jell, p. 119, pl. 21, figs 1a)8.
Holotype. QMF7059, cranidium from locality QML152 (Pl. 6,
fig. 1).
Other material. Paratypes: QMF7060–7070; supplementary
material: QMF7371–7376; CPC7317, 7329–7330, 37112–37114.
Revised diagnosis. Penarosa with trapeziform cranidium;
slender, needle-like posteriorly located occipital spine
without broad (tr.) base; lateral occipital lobe separated
from posterior cranidial border by deep furrow; baccula
elliptical, moderately well developed, length (exsag.) equal
to length of L1; anterior border flat to moderately con-
cave (sag.), moderately upturned, moderately narrowing
laterally; anterior border furrow wide (sag.) and shallow;
posterior border furrow wide (exsag.), greater than or
equal to width (sag.) of occipital ring. Exoskeletal orna-
ment similar to P. retifera, but differs in displaying coar-
ser granules on the glabella, proximal portion of palpebral
and postocular areas of the fixigenae, and frontal area.
Remarks. Penarosa netenta can be clearly distinguished
from other species of Penarosa by its lateral occipital
lobes being clearly separated from the posterior cranidial
border by a deep furrow, in addition to its wide (exsag.)
posterior border furrow and coarse ornamentation.
Examination of a specimen of P. meniscops (CPC7317)
illustrated by Opik (1970, pl. 14, fig. 2) shows an
identical condition of the occipital lobes, in addition to a
similar anterior border, anterior border furrow, and pos-
terior border furrow to those in P. netenta. Furthermore,
specimen CPC7317 comes from locality M412 of Opik
(1970), which is in close proximity to Jell’s (1977) localit-
ies of the Chummy Bore Formation. Although Opik
(1970) referred to the stratigraphical unit at locality M412
as the Inca Formation, the lithologies in which P. netenta
and specimens from M412 occur are identical, and thus
the Chummy Bore Formation (sensu Jell 1977) and Inca
Formation (sensu Opik 1970) probably represent the same
stratigraphical unit. Jell (1977) noted that Penarosa sp.
nov. PC is probably conspecific with P. netenta, but that
Opik’s (1970) figured specimens did not allow complete
confidence in this assertion. Examination of Penarosa sp.
nov. PC specimen CPC7329 illustrated by Opik (1970,
pl. 17, fig. 3; p. 41, text-fig. 14) shows a similar anterior
border to P. netenta, in addition to its coarse ornamenta-
tion. Unfortunately the occipital ring of specimen
CPC7329 is not sufficiently preserved for comparison.
However, the occurrence of Penarosa sp. nov. PC at local-
ity M412 further supports synonymy with P. netenta.
Stratigraphical occurrence. Middle Cambrian (Late Templeto-
nian ⁄ Floran). Euagnostus opimus Zone, localities M412,
QML136, QML152.
Genus LOXONEPEA Opik, 1970
1970 Loxonepea; Opik, p. 21.
Type species. Loxonepea loxophrys Opik, 1970, p. 22.
Revised diagnosis. Cranidium trapeziform, length (sag.) 95
per cent post-palpebral cranidial width (tr.); glabella
moderately tapered, width (tr.) at anterior margin 50 per
cent occipital ring width, glabellar length (sag.) 50 per
cent cranidial length, S1–S2 directed posteriorly, S3 direc-
ted laterally; lateral occipital lobes separated from poster-
ior cranidial border by shallow furrow; preglabellar boss
clavate; eye ridge strongly arched anteriorly, directed
strongly anterolaterally from axial line.
Loxonepea loxophrys Opik, 1970
Text-figure 11A–C
v*1970 Loxonepea loxophrys Opik, p. 22, pl. 17, fig. 1; p. 22,
text-fig. 7.
v1970 Loxonepea sp. nov. ND Opik, p. 23, pl. 17, fig. 2.
Holotype. CPC7327, cranidium from the Currant Bush Lime-
stone, locality M124 (Text-fig. 11A–B).
Other material. One cranidium: CPC7328.
EXPLANATION OF PLATE 6
Figs 1–9. Penarosa netenta Jell, 1977. 1, QMF7059, holotype, internal mould of cranidium; QML152; · 4Æ5. 2, QMF7061, internal
mould of partial exoskeleton; QML152; · 4. 3)4, QMF7065, internal mould of partial exoskeleton; QML136. 3, dorsal view,
stereo pair; · 3Æ5. 4, oblique lateral view; · 3. 5–6, QMF7062, internal mould of partial exoskeleton; QML152. 5, dorsal view;
· 5. 6, enlargement of pygidium; · 8Æ5. 7, QMF7372, internal mould of partial cranidium; QML136; · 3. 8, QMF7060,
internal mould of librigena; QML136; · 4. 9, QMF7067, latex cast of external mould of librigena; QML136; · 3Æ5.
506 P A L A E O N T O L O G Y , V O L U M E 4 8
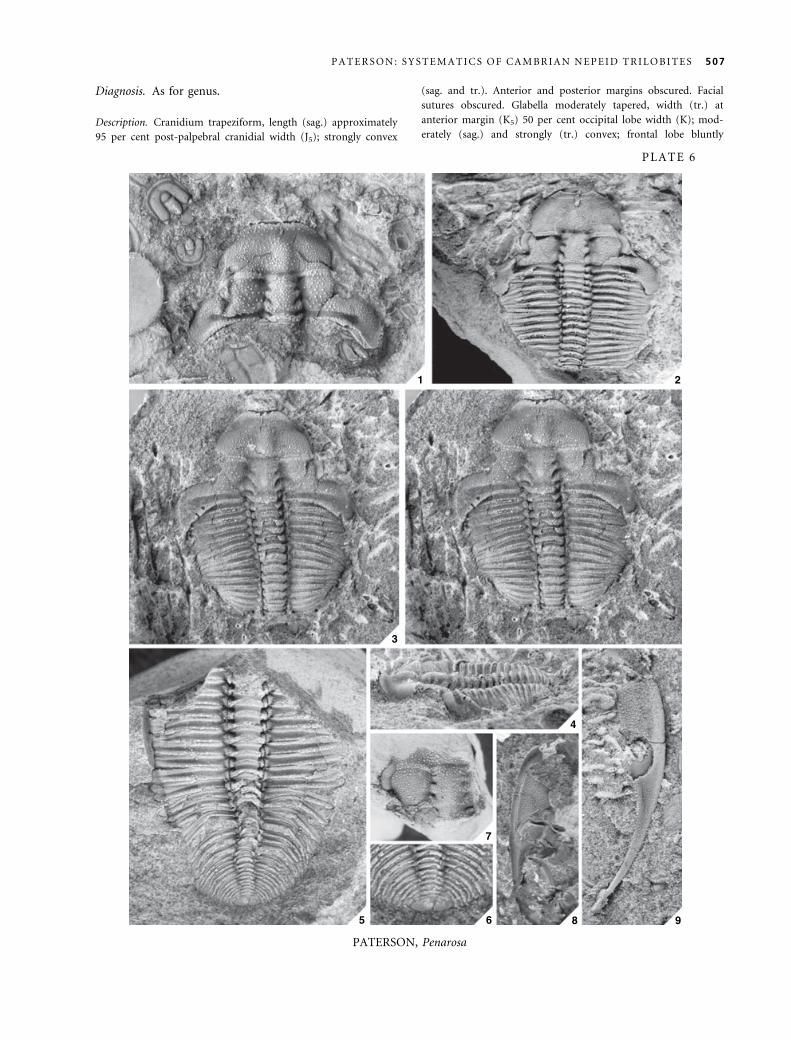
Diagnosis. As for genus.
Description. Cranidium trapeziform, length (sag.) approximately
95 per cent post-palpebral cranidial width (J5); strongly convex
(sag. and tr.). Anterior and posterior margins obscured. Facial
sutures obscured. Glabella moderately tapered, width (tr.) at
anterior margin (K5) 50 per cent occipital lobe width (K); mod-
erately (sag.) and strongly (tr.) convex; frontal lobe bluntly
1 2
3
4
5 6
7
8 9
PLATE 6
PATERSON, Penarosa
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 507
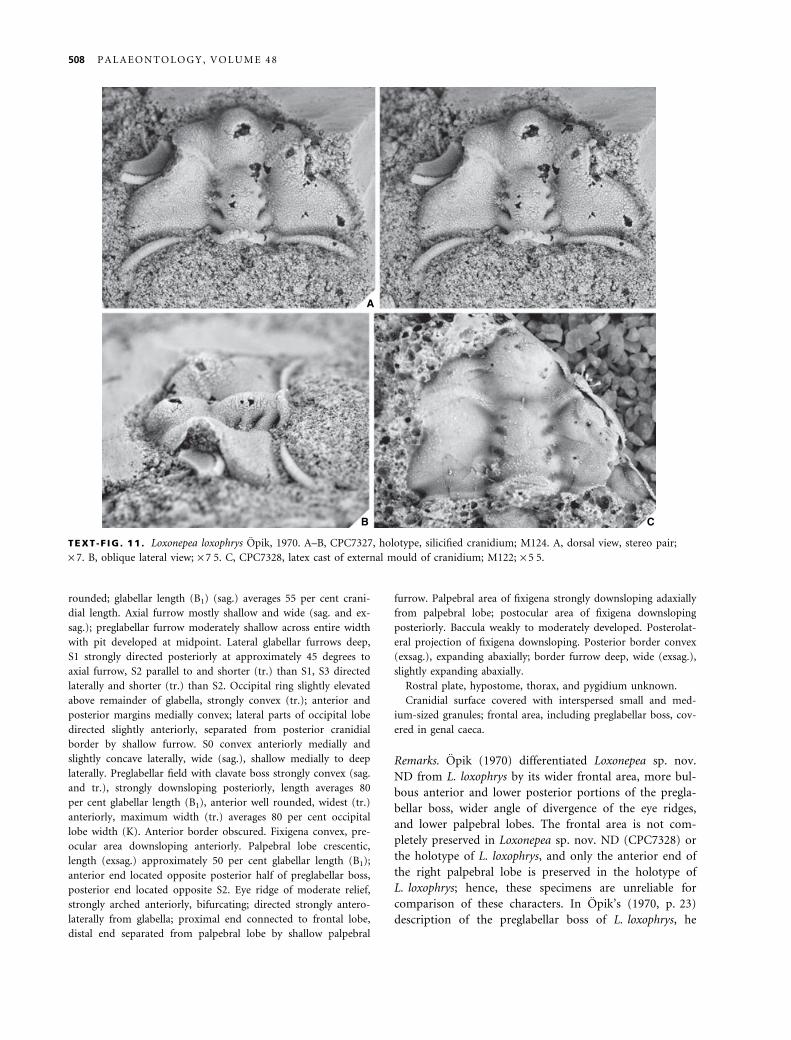
rounded; glabellar length (B1) (sag.) averages 55 per cent crani-
dial length. Axial furrow mostly shallow and wide (sag. and ex-
sag.); preglabellar furrow moderately shallow across entire width
with pit developed at midpoint. Lateral glabellar furrows deep,
S1 strongly directed posteriorly at approximately 45 degrees to
axial furrow, S2 parallel to and shorter (tr.) than S1, S3 directed
laterally and shorter (tr.) than S2. Occipital ring slightly elevated
above remainder of glabella, strongly convex (tr.); anterior and
posterior margins medially convex; lateral parts of occipital lobe
directed slightly anteriorly, separated from posterior cranidial
border by shallow furrow. S0 convex anteriorly medially and
slightly concave laterally, wide (sag.), shallow medially to deep
laterally. Preglabellar field with clavate boss strongly convex (sag.
and tr.), strongly downsloping posteriorly, length averages 80
per cent glabellar length (B1), anterior well rounded, widest (tr.)
anteriorly, maximum width (tr.) averages 80 per cent occipital
lobe width (K). Anterior border obscured. Fixigena convex, pre-
ocular area downsloping anteriorly. Palpebral lobe crescentic,
length (exsag.) approximately 50 per cent glabellar length (B1);
anterior end located opposite posterior half of preglabellar boss,
posterior end located opposite S2. Eye ridge of moderate relief,
strongly arched anteriorly, bifurcating; directed strongly antero-
laterally from glabella; proximal end connected to frontal lobe,
distal end separated from palpebral lobe by shallow palpebral
furrow. Palpebral area of fixigena strongly downsloping adaxially
from palpebral lobe; postocular area of fixigena downsloping
posteriorly. Baccula weakly to moderately developed. Posterolat-
eral projection of fixigena downsloping. Posterior border convex
(exsag.), expanding abaxially; border furrow deep, wide (exsag.),
slightly expanding abaxially.
Rostral plate, hypostome, thorax, and pygidium unknown.
Cranidial surface covered with interspersed small and med-
ium-sized granules; frontal area, including preglabellar boss, cov-
ered in genal caeca.
Remarks. Opik (1970) differentiated Loxonepea sp. nov.
ND from L. loxophrys by its wider frontal area, more bul-
bous anterior and lower posterior portions of the pregla-
bellar boss, wider angle of divergence of the eye ridges,
and lower palpebral lobes. The frontal area is not com-
pletely preserved in Loxonepea sp. nov. ND (CPC7328) or
the holotype of L. loxophrys, and only the anterior end of
the right palpebral lobe is preserved in the holotype of
L. loxophrys; hence, these specimens are unreliable for
comparison of these characters. In Opik’s (1970, p. 23)
description of the preglabellar boss of L. loxophrys, he
A
B C
TEXT -F IG . 11 . Loxonepea loxophrys Opik, 1970. A–B, CPC7327, holotype, silicified cranidium; M124. A, dorsal view, stereo pair;
· 7. B, oblique lateral view; · 7Æ5. C, CPC7328, latex cast of external mould of cranidium; M122; · 5Æ5.
508 P A L A E O N T O L O G Y , V O L U M E 4 8

noted that the boss ‘gains in tumidity forward’; however,
the same condition is observed in Loxonepea sp. nov. ND,
and therefore the differentiation of the two species using
this character is not possible. With regard to the angle of
divergence of the eye ridges, measurements reveal that the
angle for L. loxophrys is 80 degrees, whereas the angle for
Loxonepea sp. nov. ND is 110 degrees. The difference of
30 degrees is most likely due to the difference in size of
these specimens. The specimen of Loxonepea sp. nov. ND
is larger than the holotype of L. loxophrys: the glabellar
lengths are 4Æ5 mm and 2Æ8 mm, respectively. Therefore,
it is probable that the difference in the angle of diver-
gence in the eye ridges relates to the rearward migration
of the palpebral lobes during growth.
Stratigraphical occurrence. Middle Cambrian (Late Templeto-
nian ⁄ Floran). Acidusus atavus Zone, Age Creek Formation, local-
ity M122. Euagnostus opimus Zone, Currant Bush Limestone,
locality M124.
Genus FERENEPEA Opik, 1967
1967 Ferenepea Opik, p. 362.
1967 Ascionepea Opik, p. 365.
Type species. Ferenepea hispida Opik, 1967, p. 362.
Revised diagnosis. Cranidium trapeziform, length (sag.) 75
per cent post-palpebral cranidial width (tr.); glabella gently
tapered, width (tr.) at anterior margin 70 per cent occipital
lobe width, glabellar length (sag.) 70 per cent cranidial
length, S1 and S2 directed posteriorly, S3 poorly developed;
lateral occipital lobes connected to posterior cranidial bor-
der; preglabellar boss subquadrate; eye ridge slightly arched
anteriorly, directed slightly posteriorly abaxially.
Remarks. Opik (1967) distinguished Ascionepea from Fer-
enepea only by its flat and downsloping or obsolete anter-
ior border. Specimens of Ascionepea illustrated by Opik
(1967, pl. 40, figs 3–7; pl. 46, fig. 4) certainly show this
condition; however, these specimens also represent the
smallest of the Mindyallan species erected by Opik. Using
the glabellar length as a standard measure, type specimens
of Ascionepea range from 0Æ9 to 1Æ6 mm, with the holotype
of Ascionepea janitrix representing the largest specimen,
whereas type specimens of Ferenepea range from 1Æ9 to
2Æ1 mm. The most parsimonious view is that the four
Mindyallan species represent an ontogenetic series belong-
ing to a single species. The cranidia of Ascionepea anitys,
which lack an anterior border, represent the early holaspid
growth stages, A. janitrix represents an intermediate stage
in which the border has developed into a flat, downslop-
ing anterior border, while the cranidia of Ferenepea hispida
and F. pilaris represent the end members of the series and
have a well-developed anterior border. It is for this reason
that Ferenepea, and its type species F. hispida, were chosen
as the valid genus and species.
Ferenepea hispida Opik, 1967
Plate 7, figures 1–15
v*1967 Ferenepea hispida Opik, p. 362, pl. 39, figs 7a)8;
pl. 40, fig. 1a–b.
v1967 Ferenepea pilaris Opik, p. 364, pl. 40, fig. 2.
v1967 Ascionepea janitrix Opik, p. 365, pl. 40, fig. 3.
v1967 Ascionepea anitys Opik, p. 366, pl. 40, figs 4–6; pl. 46,
fig. 4.
v1967 Ascionepea sp. aff. anitys Opik, p. 366, pl. 40, fig. 7.
Holotype. CPC5666, cranidium from the O’Hara Shale, locality
D29 (Pl. 7, figs 1–4).
Other material. Eight cranidia: CPC5667, 5669–5674, 37064; one
librigena: CPC5668.
Diagnosis. As for genus.
Description. Cranidium trapeziform, length (sag.) 75 per cent
post-palpebral cranidial width (J5); convex (sag. and tr.); anter-
ior margin gently and evenly curved; posterior margin transverse
to distally strongly posterolaterally directed. Anterior branches of
facial sutures subparallel to about level with midlength of frontal
area (exsag.), then convergent to anterior margin; posterior
branches strongly divergent, curved to distal end of posterolater-
al projection. Glabella gently tapered, width (tr.) at anterior
margin (K5) averages 70 per cent occipital lobe width (K); mod-
erately (sag.), and strongly (tr.) convex; frontal lobe rounded,
projecting anteriorly beyond eye ridges; glabellar length (B1)
(sag.) averages 70 per cent cranidial length. Axial furrow of
moderate depth, and wide (tr.); preglabellar furrow moderately
deep across entire width, and wide (sag. and exsag.). Lateral gla-
bellar furrows deep, S1 directed posteriorly at approximately 45
degrees to axial furrow, S2 directed posteriorly and shorter (tr.)
than S1, S3 poorly developed to absent. Occipital ring not eleva-
ted above glabella, strongly convex (tr.), length (sag.) averages
25 per cent glabellar length; posterior margin convex posteriorly;
lateral parts of the occipital lobe directed slightly anteriorly, con-
nected to posterior cranidial border. S0 curved anteriorly, wide
(sag.), shallow medially to deep laterally. Frontal area length
(sag.) averages 35 per cent cranidial length. Preglabellar field
convex; boss subquadrate, convex (sag. and tr.), occupying entire
length of preglabellar field, length averages 25 per cent cranidial
length. Anterior border convex (sag. and exsag.), flattened dor-
sally, narrowing laterally. Anterior border furrow deep, wide
(sag. and exsag.). Fixigena convex, preocular area slightly down-
sloping anteriorly. Palpebral lobe small, length (exsag.) approxi-
mately 20 per cent glabellar length (B1). Eye ridge of moderate
relief, slightly arched anteriorly, bifurcates near axial furrow,
directed slightly posteriorly abaxially; proximal end separated
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 509

from frontal lobe by axial furrow, distal end abuts palpebral fur-
row. Palpebral and postocular areas of fixigena gently downslop-
ing adaxially. Baccula poorly developed. Posterolateral projection
of fixigena strongly downsloping, becoming narrower abaxially.
Posterior border widens slightly from axial furrow, then widens
more rapidly as it curves posteriorly, reaching its maximum
width (exsag.) at the posterior extremity of the fixigena. Poster-
ior border furrow widens slightly and evenly as it extends from
the axial furrow, at the same time curving slightly posteriorly;
near its distal extremity it abruptly curves forward, widens and
becomes less clearly defined.
Librigenae represented by one continuous unit, without a
median connective suture; lateral margin curved; inner (sutural)
margin between librigenal field and genal spine with embayment
to accommodate the posterolateral projection of the fixigena.
Border convex and continuous around entire unit, becoming
narrower at base of genal spine. Genal spine with a narrow base,
occupying approximately 50 per cent of the total librigenal
length.
Rostral plate, hypostome, thorax and pygidium unknown.
Exoskeletal surface covered in coarse granules, interspaced
with small, densely spaced granules. The frontal area and post-
erolateral projections of the fixigena of the cranidium are cov-
ered in genal caeca.
Remarks. Opik (1967, p. 364) differentiated Ferenepea pi-
laris from F. hispida by its ‘relatively large palpebral lobes,
overall pustulose boss, interocular cheeks about 1.1 of gla-
bellar width, and the trapezoidal outline of the crani-
dium’. Examination of specimens of F. hispida shows that
the palpebral lobes are rather poorly preserved, and
appear to be broken in some specimens (Pl. 7, figs 1, 7).
Comparison of surface ornamentation between species is
difficult because the holotype of F. pilaris (CPC5669) is
silicified and beekite rings obscure much of the surface
ornament (Pl. 7, fig. 10). The outline of the cranidium in
both species of Ferenepea is very similar. Based on this
evidence, there are no grounds on which to separate spec-
imens of F. hispida and F. pilaris. All of the characters
Opik (1967) used to differentiate Ascionepea anitys from
A. janitrix can be attributed to ontogenetic variation.
Such characters include the absence of an anterior border
furrow, minute palpebral lobes and the relative width of
the interocular cheeks. In specimens of A. anitys, which
are considerably smaller than the holotype of A. janitrix,
the anterior border has not yet developed, and hence the
absence of a border furrow (Pl. 7, figs 13–15). Moreover,
morphological characters such as palpebral lobes and the
relative width of interocular cheeks commonly display all-
ometry during growth, thus explaining why small speci-
mens of A. anitys possess smaller palpebral lobes and
wider interocular cheeks than those of A. janitrix.
The librigenal unit (CPC5668) is believed to have been
correctly assigned to Ferenepea hispida by Opik (1967)
because: (1) it is similar in size to associated cranidia; the
width (tr.) of the librigenal unit is 3Æ6 mm and the aver-
age width of the cranidium of F. hispida is around
4Æ5 mm; (2) the inner sutural margin exhibits an embay-
ment that conforms to the outline of the distal portion of
the posterolateral projection of the fixigenae; and (3) the
surface ornament of the librigenal unit is identical to that
of the cranidia of F. hispida.
Stratigraphical occurrence. Upper Middle–lower Upper Cambrian
(Boomerangian–Mindyallan). Lejopyge laevigata Zone, Steamboat
Sandstone, locality G103. Erediaspis eretes Zone, Mungerebar
Limestone, localities G8, G9, G114, G119, G417, G429. Acmarha-
chis quasivespa Zone, Mungerebar Limestone, localities G10,
G127, G150. Glyptagnostus stolidotus Zone, O’Hara Shale, localit-
ies D6, D28, D29; Georgina Limestone, locality W1.
Acknowledgements. I thank my supervisor Glenn A. Brock (Mac-
quarie University) for advice and support throughout the entire
period of this study, and for his assistance in the field. Ruth
Mawson and John A. Talent (Macquarie University) provided
much needed financial support for fieldwork. Thanks to John R.
Laurie (Geoscience Australia) for the loan of type material
housed in the Commonwealth Palaeontological Collection
(CPC), assistance with photography, commenting on an early
draft of the manuscript, and for encouraging me to work on the
nepeids. I am indebted to Nigel C. Hughes (University of Cali-
fornia, Riverside) for assistance with PCA. Thanks to Peter A.
Jell (Queensland Museum) for discussions, generous hospitality,
and permission to examine and photograph the type material of
N. narinosa and P. netenta during a visit to the Queensland
Museum in January 2003. I am grateful to Jim Jago (University
of South Australia) for providing images of a new species of Ne-
pea from Tasmania. Thanks also to Dave Mathieson for assist-
EXPLANATION OF PLATE 7
Figs 1–15. Ferenepea hispida Opik, 1967. 1)4, CPC5666, holotype, internal mould of cranidium; D29. 1, dorsal view, stereo pair; · 7.
2, oblique lateral view; · 7. 3, anterior view; · 8. 4, anterolateral view; · 5. 5–6, CPC5668, internal mould of librigenal unit; D29. 5,
lateral view; · 7Æ5. 6, anterior view; · 10. 7–9, CPC5667, internal mould of cranidium; D29. 7, dorsal view, stereo pair; · 7. 8, anterior
view; · 7. 9, anterolateral view; · 8Æ5. 10, CPC5669, silicified cranidium, holotype of Ferenepea pilaris Opik, 1967; G8; · 7. 11–12,
CPC5670, internal mould of cranidium, holotype of Ascionepea janitrix Opik, 1967; G103. 11, dorsal view; · 10. 12, anterolateral view;
· 7Æ5. 13, CPC5673, silicified cranidium, holotype of Ascionepea anitys Opik, 1967; G429; · 8. 14, CPC5671, silicified cranidium; G119;
· 9Æ5. 15, CPC5672, silicified cranidium; G8; · 16.
510 P A L A E O N T O L O G Y , V O L U M E 4 8
ance in the field, and Lloyd and Wendy Hick (owners of Thorn-
tonia Station) for access to the field area. Fred Sundberg and
Jim Jago provided helpful and informative reviews of the manu-
script. Partial funding for this research was provided by the
Betty Mayne Scientific Research Fund of the Linnean Society of
NSW.
PLATE 7
1
2
3 4
5
6
7
8
9
10 11 12
13 14 15
PATERSON, Ferenepea
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 511

REFERENCES
B R OC K , G. A., E N G E L B R E T S E N , M. J., J A G O , J. B.,
KR U S E , P. D., L A UR I E , J. R., S H E R G O L D, J. H., S H I ,
G. R. and S O R A UF , J. E. 2000. Palaeobiogeographic affinit-
ies of Australian Cambrian faunas. Association of Australasian
Palaeontologists, Memoir, 23, 1–61.
C OO P E R , R. A. and B R A D S H A W , M. A. 1985. Lower
Paleozoic of Nelson-Westland. Geological Society of New Zea-
land, Miscellaneous Publication, 33C, 1–42.
—— and S HE R G O L D, J. H. 1991. Palaeozoic invertebrates of
Antarctica. 455–486. In T I N G E Y , R. J. (ed.). The geology of
Antarctica. Clarendon Press, Oxford, 680 pp.
D A I L Y , B. and J A G O , J. B. 1975. The trilobite Lejopyge Hawle
and Corda and the middle-upper Cambrian boundary. Palae-
ontology, 18, 527–550.
E D W A R DS , J., W O H LT , K. E., S L A T E R , K. R., O L S H I N -
A , A. and H U TC H I N S O N , D. F. 1998. Heathcote and parts
of Woodend and Echuca, 1:100 000 map area geological
report. Geological Survey of Victoria, Report, 108, 1–212.
F O R TE Y , R. A. 1997. Classification. 289–302. In K A E S L E R ,
R. (ed.). Treatise on invertebrate paleontology, Part O (Arthro-
poda 1, Trilobita, Revised). Geological Society of America,
Boulder, and University of Kansas Press, Lawrence, 530 pp.
—— and H UG H E S , N. C. 1998. Brood pouches in trilobites.
Journal of Paleontology, 72, 638–649.
—— and OW E N S , R. M. 1997. Bubble-headed trilobites, and a
new olenid example. Palaeontology, 40, 451–459.
G E Y E R , G. and S H E R G O LD , J. H. 2000. The quest for inter-
nationally recognized divisions of Cambrian time. Episodes,
23, 188–195.
H A M M E R , Ø., H A R P E R , D. A. T. and R Y A N , P. D. 2001.
Past: Palaeontological statistics software package for education
and data analysis. Palaeontologia Electronica, 4, 9 pp. [http://
palaeo-electronica.org/2001_1/past/issue1_01.htm].
H A R R I N G T O N , H. J., H E N N I N G S M O E N , G., H O W E L L ,
B. F., J A A N U S S ON , V., L O C HM A N - B A L K, C.,
M O OR E , R. C., PO U LS E N , C., R A S E TT I , F., R I CH T E R ,
E., R I C H T E R , R., S C H M I D T , H., S D ZU Y , K., S T R U V E ,
W., S T UB B L E FI E L D, C. J., TR I PP , R., W E L L E R , J. M.
and W H I T T I N G T ON , H. B. 1959. Trilobita. O38–O560. In
M O OR E , R. C. (ed.). Treatise on invertebrate paleontology, Part
O, Arthropoda 1. Geological Society of America, Boulder, and
University of Kansas Press, Lawrence, 560 pp.
H A Y A M I , I. and M A T S U KU M A , A. 1970. Variation of
bivariate characters from the standpoint of allometry. Palaeon-
tology, 13, 588–605.
H UG H E S , N. C. 1993. Distribution, taphonomy, and func-
tional morphology of the Upper Cambrian trilobite Dikelo-
cephalus. Milwaukee Public Museum, Contributions in Biology
and Geology, 84, 1–49.
—— 1994. Ontogeny, intraspecific variation, and systematics of
the Late Cambrian trilobite Dikelocephalus. Smithsonian Con-
tributions to Paleobiology, 79, 1–89.
—— and J E L L , P. A. 1992. A statistical ⁄ computer-graphic tech-
nique for assessing variation in tectonically deformed fossils
and its application to Cambrian trilobites from Kashmir.
Lethaia, 25, 317–330.
—— and L A BA N DE I RA , C. C. 1995. The stability of species
in taxonomy. Paleobiology, 21, 401–403.
I M B R I E , J. 1956. Biometric methods in the study of inverteb-
rate fossils. Bulletin of the American Museum of Natural His-
tory, 108, 213–252.
J A GO , J. B., S U N , X. and Z A N G , W.L. 2002. Correlation
within early Palaeozoic basins of eastern South Australia.
Department of Primary Industries and Resources, South Austra-
lia, Report Book, 2002 ⁄ 33, 22 pp.
J E LL , P. A. 1977. Penarosa netenta, a new Middle Cambrian tri-
lobite from north-western Queensland. Memoirs of the Queens-
land Museum, 18, 119–123.
—— and R OB I S O N , R. A. 1978. Revision of a late Middle
Cambrian trilobite faunule from northwestern Queensland.
University of Kansas, Paleontological Contributions, 90, 1–21.
K E Y S E R , F. de and CO OK , P. J. 1972. Geology of the Middle
Cambrian phosphorites and associated sediments of north-
western Queensland. Bulletin of the Bureau of Mineral
Resources, Geology and Geophysics, Australia, 138, 1–79.
L A B A N DE I R A , C. C. and H U G HE S , N. C. 1994. Biometry
of the Late Cambrian trilobite genus Dikelocephalus and its
implications for trilobite systematics. Journal of Paleontology,
68, 492–517.
L A U R I E , J. R. 1988. Revision of some Australian Ptychagnosti-
nae (Agnostida, Cambrian). Alcheringa, 12, 169–205.
—— J A G O , J. B. and B A O , J.-S. 1995. Review of Tasmanian
Cambrian biostratigraphy. Australian Geological Survey Organ-
isation, Record, 1995 ⁄ 69, 30 pp.
M A TT H E W , G. F. 1887. Illustrations of the fauna of the St.
John Group, number 4, on the smaller-eyed trilobites of Divi-
sion I, with a few remarks on the species of the higher divi-
sions of the group. Canadian Record of Science, 2, 357–363.
M A TT H E W S , S. C. 1973. Notes on open nomenclature and
on synonymy lists. Palaeontology, 16, 713–719.
O PI K, A. A. 1961. The geology and palaeontology of the headwa-
ters of the Burke River, Queensland. Bulletin of the Bureau of
Mineral Resources, Geology and Geophysics, Australia, 53, 1–249.
—— 1963a. Nepea and the nepeids (Trilobites, Middle Cam-
brian, Australia). Journal of the Geological Society of Australia,
10, 313–316.
—— 1963b. Early Upper Cambrian fossils from Queensland.
Bulletin of the Bureau of Mineral Resources, Geology and Geo-
physics, Australia, 64, 1–133.
—— 1967. The Mindyallan fauna of north-western Queensland.
Bulletin of the Bureau of Mineral Resources, Geology and Geo-
physics, Australia, 74, 1–404, pls 1–51 [2 vols].
—— 1970. Nepeid trilobites of the Middle Cambrian of nor-
thern Australia. Bulletin of the Bureau of Mineral Resources,
Geology and Geophysics, Australia, 113, 1–48.
—— 1979. Middle Cambrian agnostids: systematics and biostra-
tigraphy. Bulletin of the Bureau of Mineral Resources, Geology
and Geophysics, Australia, 172, 1–188, pls 1–67 [2 vols].
P A L M E R , A. R. and G A T E HO US E , C. G. 1972. Early and
Middle Cambrian trilobites from Antarctica. US Geological
Survey, Professional Paper, 456D, 1–36.
P E N G , S. and R O B I S O N , R. A. 2000. Agnostoid biostratigra-
phy across the Middle-Upper Cambrian boundary in Hunan,
China. Paleontological Society, Memoir, 53, 1–104.
512 P A L A E O N T O L O G Y , V O L U M E 4 8

P R A T T , B. R. 1992. Trilobites of the Marjuman and Steptoean
stages (Upper Cambrian), Rabbitkettle Formation, southern
Mackenzie Mountains, northwest Canada. Palaeontographica
Canadiana, 9, 1–179.
R E Y M E N T , R. A., BL A C K I T H, R. E. and C A M P B E L L , N.
E. 1984. Multivariate morphometrics. Second edition. Academic
Press, London, 233 pp.
R I CH T E R , R. 1932. Crustacea (Palaontologie). 840–864. In
DI T TL E R , R., J O OS , G., K OR S C H E L T , E., L I N E K, G.,
OL T M A N N S , F. and S C HA U M , K. (eds). Handworten-
buch der Naturwissenschaften. Second edition. Gustav Fisher,
Jena.
R O B I S O N , R. A. 1978. Origin, taxonomy, and homeomorphs
of Doryagnostus (Cambrian Trilobita). University of Kansas,
Paleontological Contributions, 91, 1–10.
—— 1988. Trilobites of the Holm Dal Formation (late Middle
Cambrian), central North Greenland. Meddelelser om Grøn-
land, Geoscience, 20, 23–103.
S H A W , A. B. 1956. Quantitative trilobite studies I, the statisti-
cal description of trilobites. Journal of Paleontology, 30, 1209–
1224.
—— 1957. Quantitative trilobite studies II, measurement of the
dorsal shield of non-agnostidean trilobites. Journal of Paleon-
tology, 31, 193–207.
S H E R G O L D, J. H., J A G O, J. B., C O OP E R , R. A. and
L A UR I E , J. R. 1985. The Cambrian System in Australia, Ant-
arctica and New Zealand. International Union of Geological Sci-
ences, Publication, 19, 1–85.
—— and S OU T HG A T E , P. N. 1986. Middle Cambrian phos-
phatic and calcareous lithofacies along the eastern margin of the
Georgina Basin, western Queensland. Australian Sedimentolo-
gists Group, Field Guide Series, 2, Geological Society of Aus-
tralia, 89 pp.
S O U TH G A T E , P. N. 1986. The Gowers Formation and
Bronco Stromatolith Bed, two new stratigraphic units in the
Undilla portion of the Georgina Basin. Queensland Govern-
ment Mining Journal, 1986 (October), 407–411.
S W I N N E R T O N , H. H. 1915. Suggestions for a revised classi-
fication of trilobites. Geological Magazine, New Series, 2, 407–
496, 538–545.
T H OM A S , D. E. and S I N G L E T O N , O. P. 1956. The Cam-
brian stratigraphy of Victoria. 149–163. In R O D G E R S , J.
(ed.). El Sistema Cambrico, su Paleogeografia y el Problema de
su Base. Report of the 20th International Geological Congress,
Mexico.
W A L C H , J. E. I. 1771. Die Naturgeschichte der Versteinerungen
zur erlauterung der knorrischen Sammlung von merkwurkigkei-
ten der Natur. Volume 4, Part 3. Nurnberg, 235 pp.
W E S TE RG A R D , A. H. 1946. Agnostidea of the Middle Cam-
brian of Sweden. Sveriges Geologiska Undersøkning, Serie C,
477, 141 pp.
W H I TE H O US E , F. W. 1936. The Cambrian faunas of north-
eastern Australia. Part 1: stratigraphical outline. Part 2: Trilo-
bita (Miomera). Memoirs of the Queensland Museum, 11, 59–
112.
—— 1939. The Cambrian faunas of north-eastern Australia. Part
3: the polymerid trilobites (with supplement no. 1). Memoirs
of the Queensland Museum, 11, 179–282.
W H I TT I N GT O N , H. B., C HA T T E R T O N , B. D. E.,
S P E Y E R , S. E., F O RT E Y , R. A., OW E N S , R. M.,
CH A N G , W. T., DE A N , W. T., J E L L , P. A., L A U RI E ,
J. R., PA LM E R , A. R., R E PI N A , L. N., R US H T ON , A.
W. A., S H E R G OL D , J. H., C L A R KS ON , E. N. K.,
W I L M OT , N. V. and K E L L Y , S. R. A. 1997. Volume 1:
Introduction, Order Agnostida, Order Redlichiida. In
KA E S L E R , R. L. (ed.). Treatise on invertebrate paleontology,
Part O, Arthropoda 1, Trilobita, Revised. Geological Society
of America, Boulder, and University of Kansas Press, Law-
rence, 530 pp.
X I A N G LI-WEN and ZH A N G TAI-RONG 1985. Stratigraphy
and trilobite faunas of the Cambrian in the western part of
northern Tianshan, Xinjiang. Geological Memoirs of the Minis-
try of Geology and Mineral Resources, People’s Republic of
China, Series 2, 4, 243 pp. [In Chinese, English summary].
APPENDIX
1. Locality register
D6, Opik 1967; 21�25¢S 140�00¢E, ‘lower chert bed’ of the
O’Hara Shale, Selwyn Range, north-west Queensland; Glyptag-
nostus stolidotus Zone (Opik 1967).
D28, Opik 1967; 21�25¢S 139�58¢E, ‘lower chert bed’ of the
O’Hara Shale, Selwyn Range, north-west Queensland; Glyptag-
nostus stolidotus Zone (Opik 1967).
D29, Opik 1967; 21�26¢S 139�59Æ5¢E, ‘lower chert bed’ of the
O’Hara Shale, Selwyn Range, north-west Queensland; Glyptag-
nostus stolidotus Zone (Opik 1967).
D54, Opik 1970; 21�57¢S 139�04¢E, Steamboat Sandstone,
Quita Bore, on the bank of Quita Creek, Urandangi area, north-
west Queensland; Lejopyge laevigata Zone (Opik 1970, 1979).
D95, Opik 1970; 21�56¢S 139�00¢E, Steamboat Sandstone,
Quita Creek, Urandangi area, north-west Queensland; Lejopyge
laevigata Zone (Opik 1970).
D108, Opik 1970; 2Æ4 km south of locality D54, detrital lime-
stone layer at the base of the Steamboat Sandstone, Urandangi
area, north-west Queensland; Lejopyge laevigata Zone (Opik
1970).
G8, Opik 1967; 22�07¢S 139�12¢E, limestone horizon 7 m above
the base of the Mungerebar Limestone, Mungerebar-Mindyalla
area, north-west Queensland; Erediaspis eretes Zone (Opik 1967).
G9, Opik 1967; 22�08Æ5¢S 139�10¢E, light grey limestone of the
Mungerebar Limestone overlain by Cretaceous sandstone and
shale in east-facing escarpment, Mungerebar-Mindyalla area,
north-west Queensland; Erediaspis eretes Zone (Opik 1967).
G10, Opik 1967; 22�08¢S 139�15¢E, dark grey, banded lime-
stone of the Mungerebar Limestone, Mungerebar-Mindyalla area,
north-west Queensland; Acmarhachis quasivespa Zone (Opik
1967).
G103, Opik 1967; 22�08¢S 138�57¢E, chert biscuits weathered
out of shale from the top of the Steamboat Sandstone, Mungere-
bar-Mindyalla area, north-west Queensland; Lejopyge laevigata
Zone (Opik 1967).
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 513

G114, Opik 1967; 22�12¢S 138�57¢E, pale yellow limestone of
the Mungerebar Limestone, Mungerebar-Mindyalla area, north-
west Queensland; Erediaspis eretes Zone (Opik 1967).
G119, Opik 1967; 22�15¢S 139�00¢E, bituminous flaggy lime-
stone with chert and calcite pods of the Mungerebar Limestone,
Mungerebar-Mindyalla area, north-west Queensland; Erediaspis
eretes Zone (Opik 1967).
G127, Opik 1967; 22�17Æ5¢S 138�59Æ5¢E, bituminous and grey
sandy limestone of the Mungerebar Limestone, Mungerebar-
Mindyalla area, north-west Queensland; Acmarhachis quasivespa
Zone (Opik 1967).
G150, Opik 1967; 22�15Æ5¢S 138�57Æ5¢E, limestone with chert
and siliceous laminae of the Mungerebar Limestone, Mungere-
bar-Mindyalla area, north-west Queensland; Acmarhachis quasiv-
espa Zone (Opik 1967).
G417, Opik 1967; 22�09¢S 139�11¢E, flaggy bituminous lime-
stone of the Mungerebar Limestone, Mungerebar-Mindyalla area,
north-west Queensland; Erediaspis eretes Zone (Opik 1967).
G429, Opik 1967; 22�13¢S 139�01¢E, thin-bedded limestone,
chert and limestone breccia of the Mungerebar Limestone,
Mungerebar-Mindyalla area, north-west Queensland; Erediaspis
eretes Zone (Opik 1967).
GF1, new locality; 19�33¢8¢¢S 138�55¢16¢¢E, thin-bedded to
laminated grey limestone, 5 m above the base of the Gowers
Formation (type section), 5 km south-south-west of Thorntonia
homestead, north-west Queensland; Euagnostus opimus Zone
(Shergold and Southgate 1986).
H4, Opik 1970; coordinates unknown, black bituminous lime-
stone of the Arthur Creek Formation, 32 km due east from
Huckitta Station, close to the Arthur River, Northern Territory;
Acidusus atavus Zone (Opik 1970).
M28, Opik 1970 (see also Jell and Robison 1978); 19�27¢S138�38Æ5¢E, bituminous sandy limestone of the Currant Bush
Limestone, Camooweal area, north-west Queensland; Ptychagnos-
tus punctuosus Zone (Opik 1970).
M41, Opik 1970; 19�27Æ5¢S 138�37¢E, laminated grey sandy
limestone with chert pods of the V-Creek Limestone, Camoo-
weal area, north-west Queensland; Doryagnostus deltoides (¼punctuosus + nathorsti) Zone (Opik 1970, 1979; Laurie 1988).
M54, Opik 1970; 19�37Æ3¢S 138�38¢E, grey marly limestone of
the V-Creek Limestone, Douglas Creek stream bed, just south of
Undilla homestead, Camooweal area, north-west Queensland;
Ptychagnostus punctuosus Zone (Opik 1970, 1979; J. Laurie, pers.
comm. 2002).
M122, Opik 1970; 19�30Æ7¢S 138�47Æ6¢E, oolitic limestone and
chert interbed within the Age Creek Formation, Camooweal
area, north-west Queensland; Acidusus atavus Zone (Opik 1970).
M123, Opik 1970; 19�31¢S 138�47Æ6¢E, grey and pink lime-
stone and chert of the Currant Bush Limestone, collected from
the lowermost 3–5 m of a north-facing, low-angle cuesta (30 m
high), Camooweal area, north-west Queensland; Euagnostus opi-
mus Zone (Opik 1970, 1979; Laurie 1988).
M124, Opik 1970; 19�31Æ6¢S 138�47Æ4¢E, grey bituminous lime-
stone and chert of the Currant Bush Limestone, Camooweal
area, north-west Queensland; Euagnostus opimus Zone (Opik
1970, 1979).
M130, Opik 1970; 19�35Æ5¢S 138�48Æ5¢E, shale and chert at the
top of the Currant Bush Limestone, Camooweal area, north-west
Queensland; Ptychagnostus punctuosus Zone (Opik 1970, 1979; J.
Laurie, pers. comm. 2002).
M133, Opik 1970; 19�35¢S 138�41¢E, Split Rock Sandstone,
Camooweal area, north-west Queensland; Goniagnostus nathorsti
Zone (Opik 1970).
M141, Opik 1970; 19�39Æ5¢S 138�48¢E, red sandstone of the
Split Rock Sandstone, Camooweal area, north-west Queensland;
Goniagnostus nathorsti Zone (Opik 1970).
M157, Opik 1970; 19�25¢S 138�35¢E, bituminous limestone of
the Age Creek Formation, Morstone-Thorntonia area, north-
west Queensland; Euagnostus opimus Zone.
M160, Opik 1970; 19�23Æ5¢S 138�35Æ5¢E, chert interbed in
dolomite of the Age Creek Formation, Morstone-Thorntonia
area, north-west Queensland; Euagnostus opimus Zone (Opik
1970, 1979).
M179, Opik 1970; 18�38¢S 137�56Æ3¢E, grey nodular limestone
interbedded with shale layers in lower part of the Currant Bush
Limestone, southern bank of Lancewood Creek, Mount Drum-
mond area, Northern Territory; Acidusus atavus Zone (Opik
1970, 1979).
M180, Opik 1970; 18�38¢S 137�57Æ5¢E, grey marly limestone of
the Currant Bush Limestone, on Lancewood Creek, Mount
Drummond area, Northern Territory; Euagnostus opimus Zone
(Opik 1970, 1979; Laurie 1988).
M186, Opik 1970; 18�38Æ5¢S 137�58¢E, grey marly limestone of
the Currant Bush Limestone, approximately 0Æ8 km west of
locality M180 in the bed of Lancewood Creek, Mount Drum-
mond area, Northern Territory; Euagnostus opimus Zone (Opik
1970).
M226, Opik 1970; 20�09Æ5¢S 138�30¢E, sandstone and silicified
siltstone of the Split Rock Sandstone, west-facing scarp, Mount
Isa area, north-west Queensland; Goniagnostus nathorsti Zone
(Opik 1970).
M276, Opik 1970; 19�59Æ2¢S 138�30Æ4¢E, red sandstone in the
lower parts of the Split Rock Sandstone, overlying the thick-bed-
ded Mail Change Limestone exposed in a quarry along the Bar-
kly Highway, Camooweal area, north-west Queensland;
Goniagnostus nathorsti Zone (Opik 1970, 1979).
M344, Opik 1970; 19�57Æ5¢S 138�31¢E, white friable quartzose
sandstone in the uppermost part of the Split Rock Sandstone,
located at the face of an intermittent waterfall at the head of
Waroona Creek, Camooweal area, north-west Queensland; Goni-
agnostus nathorsti Zone (Opik 1970).
M393, Opik 1970; 18�38¢S 138�15¢E, silty siliceous limestone
of the Currant Bush Limestone, Lawn Hill area, north-west
Queensland; Euagnostus opimus Zone (Opik 1970).
M412, Opik 1970; 19�31¢S 138�53¢E, siliceous shale and chert
of the Inca Formation (¼ Chummy Bore Formation, sensu de
Keyser and Cook 1972 ¼Gowers Formation, sensu Southgate
1986), Chummy Bore, 7Æ5 km west-south-west of Thorntonia
homestead, north-west Queensland; Euagnostus opimus Zone
(Opik 1970, 1979).
M417, Opik 1970; 19�57¢S 138�29Æ5¢E, Split Rock Sandstone
(type locality), also the type locality for Nepea narinosa (see
Whitehouse 1939), Split Rock Waterhole on Waroona Creek,
approximately 3 km north of the Barkly Highway, Camooweal
area, north-west Queensland; Goniagnostus nathorsti Zone (Opik
1970).
514 P A L A E O N T O L O G Y , V O L U M E 4 8
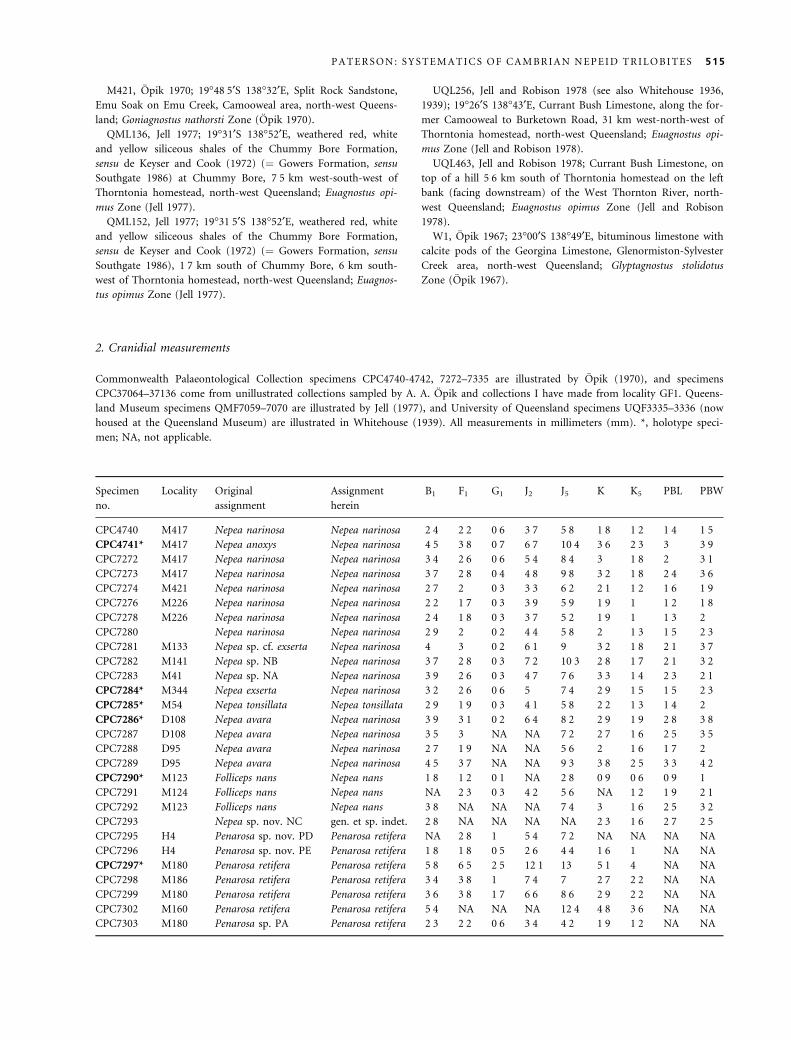
M421, Opik 1970; 19�48Æ5¢S 138�32¢E, Split Rock Sandstone,
Emu Soak on Emu Creek, Camooweal area, north-west Queens-
land; Goniagnostus nathorsti Zone (Opik 1970).
QML136, Jell 1977; 19�31¢S 138�52¢E, weathered red, white
and yellow siliceous shales of the Chummy Bore Formation,
sensu de Keyser and Cook (1972) (¼ Gowers Formation, sensu
Southgate 1986) at Chummy Bore, 7Æ5 km west-south-west of
Thorntonia homestead, north-west Queensland; Euagnostus opi-
mus Zone (Jell 1977).
QML152, Jell 1977; 19�31Æ5¢S 138�52¢E, weathered red, white
and yellow siliceous shales of the Chummy Bore Formation,
sensu de Keyser and Cook (1972) (¼ Gowers Formation, sensu
Southgate 1986), 1Æ7 km south of Chummy Bore, 6 km south-
west of Thorntonia homestead, north-west Queensland; Euagnos-
tus opimus Zone (Jell 1977).
UQL256, Jell and Robison 1978 (see also Whitehouse 1936,
1939); 19�26¢S 138�43¢E, Currant Bush Limestone, along the for-
mer Camooweal to Burketown Road, 31 km west-north-west of
Thorntonia homestead, north-west Queensland; Euagnostus opi-
mus Zone (Jell and Robison 1978).
UQL463, Jell and Robison 1978; Currant Bush Limestone, on
top of a hill 5Æ6 km south of Thorntonia homestead on the left
bank (facing downstream) of the West Thornton River, north-
west Queensland; Euagnostus opimus Zone (Jell and Robison
1978).
W1, Opik 1967; 23�00¢S 138�49¢E, bituminous limestone with
calcite pods of the Georgina Limestone, Glenormiston-Sylvester
Creek area, north-west Queensland; Glyptagnostus stolidotus
Zone (Opik 1967).
2. Cranidial measurements
Commonwealth Palaeontological Collection specimens CPC4740-4742, 7272–7335 are illustrated by Opik (1970), and specimens
CPC37064–37136 come from unillustrated collections sampled by A. A. Opik and collections I have made from locality GF1. Queens-
land Museum specimens QMF7059–7070 are illustrated by Jell (1977), and University of Queensland specimens UQF3335–3336 (now
housed at the Queensland Museum) are illustrated in Whitehouse (1939). All measurements in millimeters (mm). *, holotype speci-
men; NA, not applicable.
Specimen
no.
Locality Original
assignment
Assignment
herein
B1 F1 G1 J2 J5 K K5 PBL PBW
CPC4740 M417 Nepea narinosa Nepea narinosa 2Æ4 2Æ2 0Æ6 3Æ7 5Æ8 1Æ8 1Æ2 1Æ4 1Æ5CPC4741* M417 Nepea anoxys Nepea narinosa 4Æ5 3Æ8 0Æ7 6Æ7 10Æ4 3Æ6 2Æ3 3 3Æ9CPC7272 M417 Nepea narinosa Nepea narinosa 3Æ4 2Æ6 0Æ6 5Æ4 8Æ4 3 1Æ8 2 3Æ1CPC7273 M417 Nepea narinosa Nepea narinosa 3Æ7 2Æ8 0Æ4 4Æ8 9Æ8 3Æ2 1Æ8 2Æ4 3Æ6CPC7274 M421 Nepea narinosa Nepea narinosa 2Æ7 2 0Æ3 3Æ3 6Æ2 2Æ1 1Æ2 1Æ6 1Æ9CPC7276 M226 Nepea narinosa Nepea narinosa 2Æ2 1Æ7 0Æ3 3Æ9 5Æ9 1Æ9 1 1Æ2 1Æ8CPC7278 M226 Nepea narinosa Nepea narinosa 2Æ4 1Æ8 0Æ3 3Æ7 5Æ2 1Æ9 1 1Æ3 2
CPC7280 Nepea narinosa Nepea narinosa 2Æ9 2 0Æ2 4Æ4 5Æ8 2 1Æ3 1Æ5 2Æ3CPC7281 M133 Nepea sp. cf. exserta Nepea narinosa 4 3 0Æ2 6Æ1 9 3Æ2 1Æ8 2Æ1 3Æ7CPC7282 M141 Nepea sp. NB Nepea narinosa 3Æ7 2Æ8 0Æ3 7Æ2 10Æ3 2Æ8 1Æ7 2Æ1 3Æ2CPC7283 M41 Nepea sp. NA Nepea narinosa 3Æ9 2Æ6 0Æ3 4Æ7 7Æ6 3Æ3 1Æ4 2Æ3 2Æ1CPC7284* M344 Nepea exserta Nepea narinosa 3Æ2 2Æ6 0Æ6 5 7Æ4 2Æ9 1Æ5 1Æ5 2Æ3CPC7285* M54 Nepea tonsillata Nepea tonsillata 2Æ9 1Æ9 0Æ3 4Æ1 5Æ8 2Æ2 1Æ3 1Æ4 2
CPC7286* D108 Nepea avara Nepea narinosa 3Æ9 3Æ1 0Æ2 6Æ4 8Æ2 2Æ9 1Æ9 2Æ8 3Æ8CPC7287 D108 Nepea avara Nepea narinosa 3Æ5 3 NA NA 7Æ2 2Æ7 1Æ6 2Æ5 3Æ5CPC7288 D95 Nepea avara Nepea narinosa 2Æ7 1Æ9 NA NA 5Æ6 2 1Æ6 1Æ7 2
CPC7289 D95 Nepea avara Nepea narinosa 4Æ5 3Æ7 NA NA 9Æ3 3Æ8 2Æ5 3Æ3 4Æ2CPC7290* M123 Folliceps nans Nepea nans 1Æ8 1Æ2 0Æ1 NA 2Æ8 0Æ9 0Æ6 0Æ9 1
CPC7291 M124 Folliceps nans Nepea nans NA 2Æ3 0Æ3 4Æ2 5Æ6 NA 1Æ2 1Æ9 2Æ1CPC7292 M123 Folliceps nans Nepea nans 3Æ8 NA NA NA 7Æ4 3 1Æ6 2Æ5 3Æ2CPC7293 Nepea sp. nov. NC gen. et sp. indet. 2Æ8 NA NA NA NA 2Æ3 1Æ6 2Æ7 2Æ5CPC7295 H4 Penarosa sp. nov. PD Penarosa retifera NA 2Æ8 1 5Æ4 7Æ2 NA NA NA NA
CPC7296 H4 Penarosa sp. nov. PE Penarosa retifera 1Æ8 1Æ8 0Æ5 2Æ6 4Æ4 1Æ6 1 NA NA
CPC7297* M180 Penarosa retifera Penarosa retifera 5Æ8 6Æ5 2Æ5 12Æ1 13 5Æ1 4 NA NA
CPC7298 M186 Penarosa retifera Penarosa retifera 3Æ4 3Æ8 1 7Æ4 7 2Æ7 2Æ2 NA NA
CPC7299 M180 Penarosa retifera Penarosa retifera 3Æ6 3Æ8 1Æ7 6Æ6 8Æ6 2Æ9 2Æ2 NA NA
CPC7302 M160 Penarosa retifera Penarosa retifera 5Æ4 NA NA NA 12Æ4 4Æ8 3Æ6 NA NA
CPC7303 M180 Penarosa sp. PA Penarosa retifera 2Æ3 2Æ2 0Æ6 3Æ4 4Æ2 1Æ9 1Æ2 NA NA
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 515
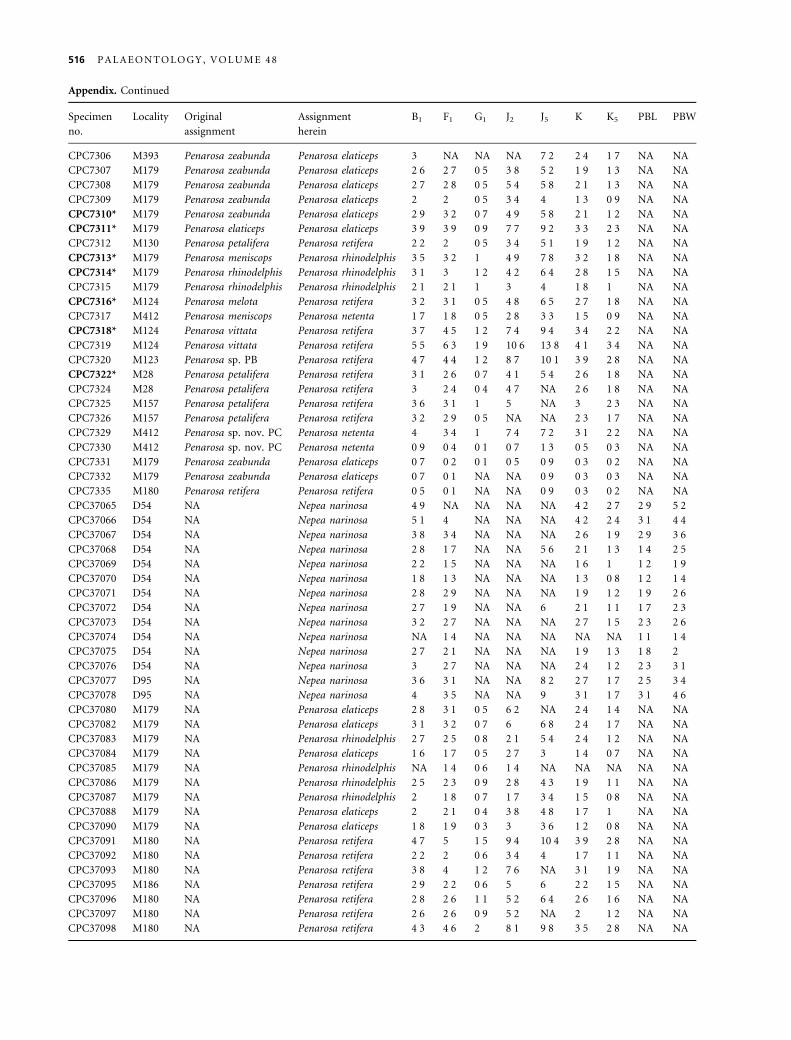
Appendix. Continued
Specimen
no.
Locality Original
assignment
Assignment
herein
B1 F1 G1 J2 J5 K K5 PBL PBW
CPC7306 M393 Penarosa zeabunda Penarosa elaticeps 3 NA NA NA 7Æ2 2Æ4 1Æ7 NA NA
CPC7307 M179 Penarosa zeabunda Penarosa elaticeps 2Æ6 2Æ7 0Æ5 3Æ8 5Æ2 1Æ9 1Æ3 NA NA
CPC7308 M179 Penarosa zeabunda Penarosa elaticeps 2Æ7 2Æ8 0Æ5 5Æ4 5Æ8 2Æ1 1Æ3 NA NA
CPC7309 M179 Penarosa zeabunda Penarosa elaticeps 2 2 0Æ5 3Æ4 4 1Æ3 0Æ9 NA NA
CPC7310* M179 Penarosa zeabunda Penarosa elaticeps 2Æ9 3Æ2 0Æ7 4Æ9 5Æ8 2Æ1 1Æ2 NA NA
CPC7311* M179 Penarosa elaticeps Penarosa elaticeps 3Æ9 3Æ9 0Æ9 7Æ7 9Æ2 3Æ3 2Æ3 NA NA
CPC7312 M130 Penarosa petalifera Penarosa retifera 2Æ2 2 0Æ5 3Æ4 5Æ1 1Æ9 1Æ2 NA NA
CPC7313* M179 Penarosa meniscops Penarosa rhinodelphis 3Æ5 3Æ2 1 4Æ9 7Æ8 3Æ2 1Æ8 NA NA
CPC7314* M179 Penarosa rhinodelphis Penarosa rhinodelphis 3Æ1 3 1Æ2 4Æ2 6Æ4 2Æ8 1Æ5 NA NA
CPC7315 M179 Penarosa rhinodelphis Penarosa rhinodelphis 2Æ1 2Æ1 1 3 4 1Æ8 1 NA NA
CPC7316* M124 Penarosa melota Penarosa retifera 3Æ2 3Æ1 0Æ5 4Æ8 6Æ5 2Æ7 1Æ8 NA NA
CPC7317 M412 Penarosa meniscops Penarosa netenta 1Æ7 1Æ8 0Æ5 2Æ8 3Æ3 1Æ5 0Æ9 NA NA
CPC7318* M124 Penarosa vittata Penarosa retifera 3Æ7 4Æ5 1Æ2 7Æ4 9Æ4 3Æ4 2Æ2 NA NA
CPC7319 M124 Penarosa vittata Penarosa retifera 5Æ5 6Æ3 1Æ9 10Æ6 13Æ8 4Æ1 3Æ4 NA NA
CPC7320 M123 Penarosa sp. PB Penarosa retifera 4Æ7 4Æ4 1Æ2 8Æ7 10Æ1 3Æ9 2Æ8 NA NA
CPC7322* M28 Penarosa petalifera Penarosa retifera 3Æ1 2Æ6 0Æ7 4Æ1 5Æ4 2Æ6 1Æ8 NA NA
CPC7324 M28 Penarosa petalifera Penarosa retifera 3 2Æ4 0Æ4 4Æ7 NA 2Æ6 1Æ8 NA NA
CPC7325 M157 Penarosa petalifera Penarosa retifera 3Æ6 3Æ1 1 5 NA 3 2Æ3 NA NA
CPC7326 M157 Penarosa petalifera Penarosa retifera 3Æ2 2Æ9 0Æ5 NA NA 2Æ3 1Æ7 NA NA
CPC7329 M412 Penarosa sp. nov. PC Penarosa netenta 4 3Æ4 1 7Æ4 7Æ2 3Æ1 2Æ2 NA NA
CPC7330 M412 Penarosa sp. nov. PC Penarosa netenta 0Æ9 0Æ4 0Æ1 0Æ7 1Æ3 0Æ5 0Æ3 NA NA
CPC7331 M179 Penarosa zeabunda Penarosa elaticeps 0Æ7 0Æ2 0Æ1 0Æ5 0Æ9 0Æ3 0Æ2 NA NA
CPC7332 M179 Penarosa zeabunda Penarosa elaticeps 0Æ7 0Æ1 NA NA 0Æ9 0Æ3 0Æ3 NA NA
CPC7335 M180 Penarosa retifera Penarosa retifera 0Æ5 0Æ1 NA NA 0Æ9 0Æ3 0Æ2 NA NA
CPC37065 D54 NA Nepea narinosa 4Æ9 NA NA NA NA 4Æ2 2Æ7 2Æ9 5Æ2CPC37066 D54 NA Nepea narinosa 5Æ1 4 NA NA NA 4Æ2 2Æ4 3Æ1 4Æ4CPC37067 D54 NA Nepea narinosa 3Æ8 3Æ4 NA NA NA 2Æ6 1Æ9 2Æ9 3Æ6CPC37068 D54 NA Nepea narinosa 2Æ8 1Æ7 NA NA 5Æ6 2Æ1 1Æ3 1Æ4 2Æ5CPC37069 D54 NA Nepea narinosa 2Æ2 1Æ5 NA NA NA 1Æ6 1 1Æ2 1Æ9CPC37070 D54 NA Nepea narinosa 1Æ8 1Æ3 NA NA NA 1Æ3 0Æ8 1Æ2 1Æ4CPC37071 D54 NA Nepea narinosa 2Æ8 2Æ9 NA NA NA 1Æ9 1Æ2 1Æ9 2Æ6CPC37072 D54 NA Nepea narinosa 2Æ7 1Æ9 NA NA 6 2Æ1 1Æ1 1Æ7 2Æ3CPC37073 D54 NA Nepea narinosa 3Æ2 2Æ7 NA NA NA 2Æ7 1Æ5 2Æ3 2Æ6CPC37074 D54 NA Nepea narinosa NA 1Æ4 NA NA NA NA NA 1Æ1 1Æ4CPC37075 D54 NA Nepea narinosa 2Æ7 2Æ1 NA NA NA 1Æ9 1Æ3 1Æ8 2
CPC37076 D54 NA Nepea narinosa 3 2Æ7 NA NA NA 2Æ4 1Æ2 2Æ3 3Æ1CPC37077 D95 NA Nepea narinosa 3Æ6 3Æ1 NA NA 8Æ2 2Æ7 1Æ7 2Æ5 3Æ4CPC37078 D95 NA Nepea narinosa 4 3Æ5 NA NA 9 3Æ1 1Æ7 3Æ1 4Æ6CPC37080 M179 NA Penarosa elaticeps 2Æ8 3Æ1 0Æ5 6Æ2 NA 2Æ4 1Æ4 NA NA
CPC37082 M179 NA Penarosa elaticeps 3Æ1 3Æ2 0Æ7 6 6Æ8 2Æ4 1Æ7 NA NA
CPC37083 M179 NA Penarosa rhinodelphis 2Æ7 2Æ5 0Æ8 2Æ1 5Æ4 2Æ4 1Æ2 NA NA
CPC37084 M179 NA Penarosa elaticeps 1Æ6 1Æ7 0Æ5 2Æ7 3 1Æ4 0Æ7 NA NA
CPC37085 M179 NA Penarosa rhinodelphis NA 1Æ4 0Æ6 1Æ4 NA NA NA NA NA
CPC37086 M179 NA Penarosa rhinodelphis 2Æ5 2Æ3 0Æ9 2Æ8 4Æ3 1Æ9 1Æ1 NA NA
CPC37087 M179 NA Penarosa rhinodelphis 2 1Æ8 0Æ7 1Æ7 3Æ4 1Æ5 0Æ8 NA NA
CPC37088 M179 NA Penarosa elaticeps 2 2Æ1 0Æ4 3Æ8 4Æ8 1Æ7 1 NA NA
CPC37090 M179 NA Penarosa elaticeps 1Æ8 1Æ9 0Æ3 3 3Æ6 1Æ2 0Æ8 NA NA
CPC37091 M180 NA Penarosa retifera 4Æ7 5 1Æ5 9Æ4 10Æ4 3Æ9 2Æ8 NA NA
CPC37092 M180 NA Penarosa retifera 2Æ2 2 0Æ6 3Æ4 4 1Æ7 1Æ1 NA NA
CPC37093 M180 NA Penarosa retifera 3Æ8 4 1Æ2 7Æ6 NA 3Æ1 1Æ9 NA NA
CPC37095 M186 NA Penarosa retifera 2Æ9 2Æ2 0Æ6 5 6 2Æ2 1Æ5 NA NA
CPC37096 M180 NA Penarosa retifera 2Æ8 2Æ6 1Æ1 5Æ2 6Æ4 2Æ6 1Æ6 NA NA
CPC37097 M180 NA Penarosa retifera 2Æ6 2Æ6 0Æ9 5Æ2 NA 2 1Æ2 NA NA
CPC37098 M180 NA Penarosa retifera 4Æ3 4Æ6 2 8Æ1 9Æ8 3Æ5 2Æ8 NA NA
516 P A L A E O N T O L O G Y , V O L U M E 4 8
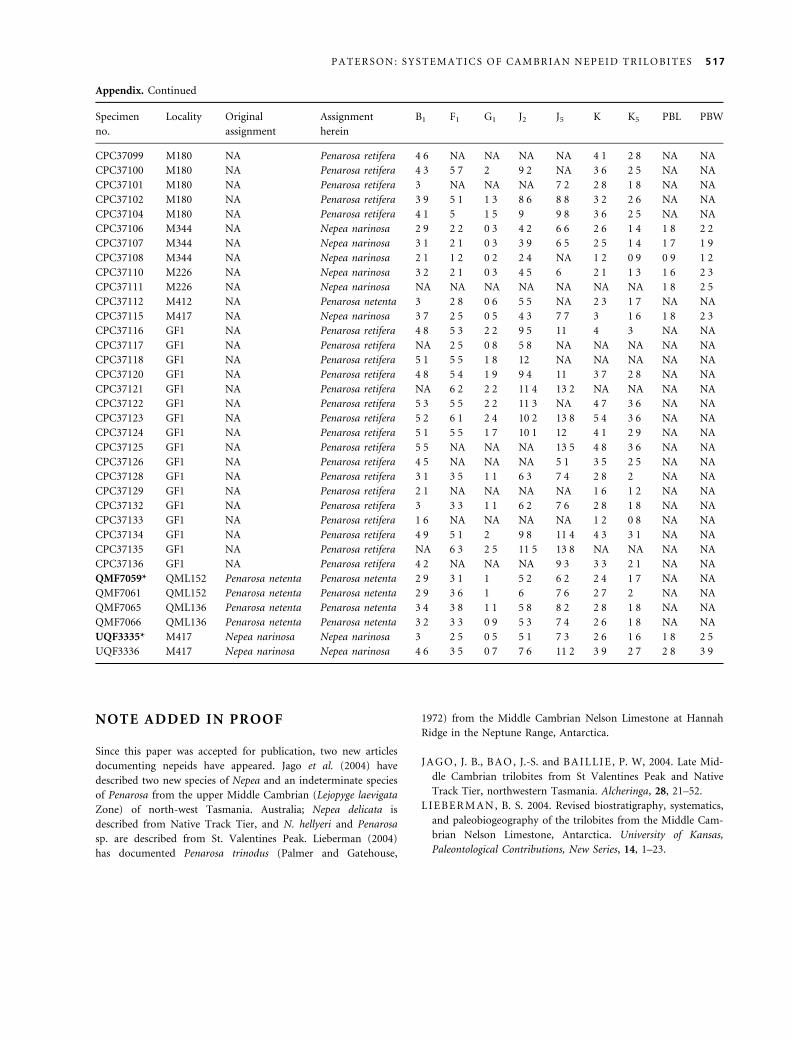
Appendix. Continued
Specimen
no.
Locality Original
assignment
Assignment
herein
B1 F1 G1 J2 J5 K K5 PBL PBW
CPC37099 M180 NA Penarosa retifera 4Æ6 NA NA NA NA 4Æ1 2Æ8 NA NA
CPC37100 M180 NA Penarosa retifera 4Æ3 5Æ7 2 9Æ2 NA 3Æ6 2Æ5 NA NA
CPC37101 M180 NA Penarosa retifera 3 NA NA NA 7Æ2 2Æ8 1Æ8 NA NA
CPC37102 M180 NA Penarosa retifera 3Æ9 5Æ1 1Æ3 8Æ6 8Æ8 3Æ2 2Æ6 NA NA
CPC37104 M180 NA Penarosa retifera 4Æ1 5 1Æ5 9 9Æ8 3Æ6 2Æ5 NA NA
CPC37106 M344 NA Nepea narinosa 2Æ9 2Æ2 0Æ3 4Æ2 6Æ6 2Æ6 1Æ4 1Æ8 2Æ2CPC37107 M344 NA Nepea narinosa 3Æ1 2Æ1 0Æ3 3Æ9 6Æ5 2Æ5 1Æ4 1Æ7 1Æ9CPC37108 M344 NA Nepea narinosa 2Æ1 1Æ2 0Æ2 2Æ4 NA 1Æ2 0Æ9 0Æ9 1Æ2CPC37110 M226 NA Nepea narinosa 3Æ2 2Æ1 0Æ3 4Æ5 6 2Æ1 1Æ3 1Æ6 2Æ3CPC37111 M226 NA Nepea narinosa NA NA NA NA NA NA NA 1Æ8 2Æ5CPC37112 M412 NA Penarosa netenta 3 2Æ8 0Æ6 5Æ5 NA 2Æ3 1Æ7 NA NA
CPC37115 M417 NA Nepea narinosa 3Æ7 2Æ5 0Æ5 4Æ3 7Æ7 3 1Æ6 1Æ8 2Æ3CPC37116 GF1 NA Penarosa retifera 4Æ8 5Æ3 2Æ2 9Æ5 11 4 3 NA NA
CPC37117 GF1 NA Penarosa retifera NA 2Æ5 0Æ8 5Æ8 NA NA NA NA NA
CPC37118 GF1 NA Penarosa retifera 5Æ1 5Æ5 1Æ8 12 NA NA NA NA NA
CPC37120 GF1 NA Penarosa retifera 4Æ8 5Æ4 1Æ9 9Æ4 11 3Æ7 2Æ8 NA NA
CPC37121 GF1 NA Penarosa retifera NA 6Æ2 2Æ2 11Æ4 13Æ2 NA NA NA NA
CPC37122 GF1 NA Penarosa retifera 5Æ3 5Æ5 2Æ2 11Æ3 NA 4Æ7 3Æ6 NA NA
CPC37123 GF1 NA Penarosa retifera 5Æ2 6Æ1 2Æ4 10Æ2 13Æ8 5Æ4 3Æ6 NA NA
CPC37124 GF1 NA Penarosa retifera 5Æ1 5Æ5 1Æ7 10Æ1 12 4Æ1 2Æ9 NA NA
CPC37125 GF1 NA Penarosa retifera 5Æ5 NA NA NA 13Æ5 4Æ8 3Æ6 NA NA
CPC37126 GF1 NA Penarosa retifera 4Æ5 NA NA NA 5Æ1 3Æ5 2Æ5 NA NA
CPC37128 GF1 NA Penarosa retifera 3Æ1 3Æ5 1Æ1 6Æ3 7Æ4 2Æ8 2 NA NA
CPC37129 GF1 NA Penarosa retifera 2Æ1 NA NA NA NA 1Æ6 1Æ2 NA NA
CPC37132 GF1 NA Penarosa retifera 3 3Æ3 1Æ1 6Æ2 7Æ6 2Æ8 1Æ8 NA NA
CPC37133 GF1 NA Penarosa retifera 1Æ6 NA NA NA NA 1Æ2 0Æ8 NA NA
CPC37134 GF1 NA Penarosa retifera 4Æ9 5Æ1 2 9Æ8 11Æ4 4Æ3 3Æ1 NA NA
CPC37135 GF1 NA Penarosa retifera NA 6Æ3 2Æ5 11Æ5 13Æ8 NA NA NA NA
CPC37136 GF1 NA Penarosa retifera 4Æ2 NA NA NA 9Æ3 3Æ3 2Æ1 NA NA
QMF7059* QML152 Penarosa netenta Penarosa netenta 2Æ9 3Æ1 1 5Æ2 6Æ2 2Æ4 1Æ7 NA NA
QMF7061 QML152 Penarosa netenta Penarosa netenta 2Æ9 3Æ6 1 6 7Æ6 2Æ7 2 NA NA
QMF7065 QML136 Penarosa netenta Penarosa netenta 3Æ4 3Æ8 1Æ1 5Æ8 8Æ2 2Æ8 1Æ8 NA NA
QMF7066 QML136 Penarosa netenta Penarosa netenta 3Æ2 3Æ3 0Æ9 5Æ3 7Æ4 2Æ6 1Æ8 NA NA
UQF3335* M417 Nepea narinosa Nepea narinosa 3 2Æ5 0Æ5 5Æ1 7Æ3 2Æ6 1Æ6 1Æ8 2Æ5UQF3336 M417 Nepea narinosa Nepea narinosa 4Æ6 3Æ5 0Æ7 7Æ6 11Æ2 3Æ9 2Æ7 2Æ8 3Æ9
NOTE ADDED IN PROOF
Since this paper was accepted for publication, two new articles
documenting nepeids have appeared. Jago et al. (2004) have
described two new species of Nepea and an indeterminate species
of Penarosa from the upper Middle Cambrian (Lejopyge laevigata
Zone) of north-west Tasmania. Australia; Nepea delicata is
described from Native Track Tier, and N. hellyeri and Penarosa
sp. are described from St. Valentines Peak. Lieberman (2004)
has documented Penarosa trinodus (Palmer and Gatehouse,
1972) from the Middle Cambrian Nelson Limestone at Hannah
Ridge in the Neptune Range, Antarctica.
J A GO , J. B., B A O , J.-S. and B A I L LI E , P. W, 2004. Late Mid-
dle Cambrian trilobites from St Valentines Peak and Native
Track Tier, northwestern Tasmania. Alcheringa, 28, 21–52.
L I E BE R M A N , B. S. 2004. Revised biostratigraphy, systematics,
and paleobiogeography of the trilobites from the Middle Cam-
brian Nelson Limestone, Antarctica. University of Kansas,
Paleontological Contributions, New Series, 14, 1–23.
P A T E R S O N : S Y S T E M A T I C S O F C A M B R I A N N E P E I D T R I L O B I T E S 517





























