Embed Size (px)
Citation preview

Developmental and Comparative Immunology, Vol. 19, No. 6, pp. $07-523, 1995 Copyright 0 1995 Elsevier Science Ltd Printed in the USA. All rights reserved
0145-305x/95 %9.50+0.00
ONTOGENY AND THYMUS-DEPENDENCE OF T CELL SURFACE ANTIGENS IN Xenopus: FLOW CYTOMETRIC
STUDIES ON MONOCLONAL ANTIBODY-STAINED THYMUS AND SPLEEN
Ian Gravenor, Trudy L. Horton, Pamela Ritchie, Emma Flint, and John D. Horton
Departrrlent of Biological Sciences, University of Durham, South Road, Durham DHl 3LE, U.K.
(Submitted June 1995; Accepted August 1995)
q Abstract--Recently generated anti- Xenopus T cell monoclonal antibodies (mAbs) to lthe 120 kDA XTLA-1 determi- nant and against the putative CD5 and CDS homologues, together with anti-IgM and anti-MHC class II mAbs, are used in dual colour flow cytometric experiments to characterize cell surface antigenic expression on lymphocytes in thymus and spleen of Xenopus laevis during larval and early adult life and also in metamorphosis-inhibited animals. Histological confirmation of T cell emergence early in larval ontogeny is sup- plied by cryostat sections stained for CDS. Five-day thymectomy, i.e. prior to T-lineage cell differentiation in the thymus, abolishes T cell marker expression in the spleen for up to 1 year. Moreover, late larval (20 days} or early adult (3 months) thymectomy (i.e. removal after peripheralization of T cells has occurred) a’lso leads to severe depletion of mAb-defineid T cells in the spleen.
q lKeywords-Flow cytometry; Monoclonal antibodies; T cell ontogeny; Thymectomy; Xenopus.
Introduction
The role of the thymus in T cell develop- ment is currently under scrutiny. The
Address correspondence to Dr. J. D. Horton, Depar~ent of Biological Sciences, University of Durham, Durham, DHl3LE, U.K.
possibility of an auxiliary, extra-thymic maturational pathway is indicated in experiments probing T cell marker expres- sion in athymic and euthymic rodents (1,2). In contrast, the thymus appears to be the unique source of monoclonal antibody (mAb)-define T cells in galli- naceous birds (3). Amongst ectothermic vertebrates the role of the thymus is being explored in depth in Xenopus (4). Removal of the thymus at a rudimentary stage of thymus histogenesis is readily feasible in this African clawed frog (5). Such thymectomized (TX) larvae and adults develop with a crippled thymus- dependent immune system, as judged by impaired alloimmune reactivity in vivo (5,6) and in vitro (7), loss of helper activity required for antibody production to thymus-dependent antigens (8), and by the ‘failure’ of lymphocytes from TX animals to respond to T cell mitogen stimulation by enhanced proliferation (7,9,10) and release of cytokines pro- moting lymphocyte proliferation (11). Nevertheless, the possibility that a popu- lation of T cells is generated in such ‘athymic’ Xenopus has been suggested in view of certain residual T cell-like functions in these animals (4,12,13).
Comparison of mAb-defined lympho- cyte populations in control and TX Xe~o~~ should help resolve the issue of whether extra-thymic T cell maturation
507

508 I. Gravenor et al.
occurs at this level of evolution. In this paper we employ mAbs against Xenopus T cell specific antigens WTLA-1 (14), CD5 (15) and CD8 (16,17)] to charac- terize ontogeny of T cell surface marker expression in thymus and spleen of euthymic Xenopus. Histological confirma- tion of T cell development suggested by flow cytometry is provided by immuno- histochemistry of CD8-stained sections. Dual colour flow cytometry confirms that the emergence of constitutive major his- tocompatibility complex (MHC) class II expression on T cells is metamorphosis- dependent (18,19) and reveals that the intensity of class II staining on T and B lymphocytes is distinct. Finally, thy- mectomy at various stages of develop- ment reveals a pronounced and prolonged influence of the thymus on splenic T cell marker expression.
Materials and Methods
Animals and Operations
Outbred Xenopus laevis were reared in the laboratory at 23 + 2°C as described elsewhere (5). Metamorphosis was com- pleted (stage 66 [20]) by approximately 8 weeks, except in those animals deliber- ately blocked from transforming by addi- tion of sodium perchlorate to aquaria (1 gL_‘), treatment begun at stage 53, as described elsewhere (19). Thymectomy of larvae at 5 or 20 days was by micro- cautery (5), whereas the thymus of 3-month-old Xenopus was removed surgi- cally (21). Controls were either non- operated or sham-thymectomized (5,21) siblings. Thymectomized animals remained in good health throughout the experimental period (up to 1 year).
Monoclonal Antibodies
Murine monoclonal antibodies (mAbs) used were XT-l (IgG2 with specificity for
a 120 kDA Xenopus T cell antigen [14], 2Bl (IgGi, anti-Xenopus CD5 [15]), F17 and AM22 (both IgM, anti-Xenopus CD8 [ 16,17]), D8 and 8E4 : 57 (both IgG, anti- Xenopus IgM [15,22]) and AM20 (IgG, anti-Xenopus class II MHC [17]). We gratefully acknowledge the generous gifts of mAbs or hybridoma cells from Dr. R. H. Clothier (8E4: 57), Dr. M. D. Cooper (2B1, F17, D8, CT3), Dr. L. Du Pasquier and Dr. M. Flajnik (AM22, AM20), and Dr. S. Nagata (XT-l).
Flow Cytometry
Lymphocytes were obtained from thymus and spleen as described elsewhere (11). Cells were washed in amphibian strength PBS (APBS with 0.1% NaNs and 0.1% BSA) before incubation with the primary mAb at optimal concentra- tion. After washing, cells were incubated with FITC-labelled, rabbit anti-mouse immunoglobulin (DAKO) that had pre- viously been adsorbed for 30min on ice with 5% Xenopus serum to remove non- specific staining. Cells were then washed with APBS containing 1% normal mouse serum and counter-stained with a phy- coerythrin (PE)-labelled mAb. Phycoery- thrin conjugation of mAbs (to CD5, CD8 and IgM) was performed by Southern Biotechnology Associates, Birmingham, Alabama. The control mouse antibody for indirect FITC stainings was CT3, an anti-chicken CD3 of IgGi isotype (23); the PE control was mouse IgG conjugated to phycoerythrin (DAKO). For each cell population markers were set to exclude 98% of cells stained with control reagents from positive analysis. Gates were set by forward and side scatter to exclude erythrocytes and dead cells from analysis, the latter being confirmed by propidium iodide staining. Five to ten thousand lymphocytes were analysed on a Coulter EPICS flow cytometer.

T cell ontogeny in Xenopus 509
Immunohistochemistry
Cryostat sections of liquid nitrogen- frozen larval thymus and spleen were cut at 5 pm and collected on to poly-l-lysine- coated slides. Slides were fixed for 2 min in acetone, then stored at -20°C prior to staining in a moist chamber at room temperature. Sections were first incubated for 30 min with blocking buffer (APBS containing 1% BSA) before incubation with primary mAb (AM22) for 1 h. AM22 alone was chosen to histologically monitor T cell development, in view of its excellent staining qualities compared with other anti-T cell mAbs. Following washing in APBS with 0.1% BSA, sections were incubated in biotinylated anti-mouse IgM (Vector Labs) for 30 min prior to removal of endogenous perox- idase by hydrogen peroxide/methanol treatment. An avidin-biotin-peroxidase complex (Vector) was then added to sections for 30 min and the bound peroxidase visualized with diamino benzi- dine substrate. Sections were counter- stained with haematoxylin, dehydrated and viewe’d under brightfield or darkfield illumination for CD8 + cells.
Results
Ontogeny of m/lb-defined T and B Cells in Thymus and Spleen
Absolute Lymphocyte Numbers. Flow cytometry was not feasible on animals at 5 days since at this time < 500 cells are found in the thymus and lymphopoiesis has not ye:t begun in the spleen (5,24,25). Thymuses were first taken for cytometry on day 7, and spleens on day 12, when a few thousand lymphocytes are first to be found in these organs (Fig. 1). After rapid expansion during weeks 2-4 of develop- ment, thymocyte and splenocyte numbers then tend to plateau until metamorphosis is completed (60 days). A striking increase
in lymphocytes is seen in both organs in early adult life.
T and B Cell Markers. Figure 2 illus- trates mean percentages of thymocytes and splenocytes stained for T (CD5, CD8, XTLA-1) and B (IgM) surface markers as judged by flow cytometry. Examples of staining properties of indi- vidual antibodies are shown in Figures 3 and 4.
CD5. The percentages of CDS+ thymo- cytes were difficult to quantify by flow cytometry, especially in larvae where fluorescence achieved by 2Bl staining was invariably of low intensity (Fig. 4). CD5 staining of thymocytes was first examined at 12 days, when approximately 40% CD5+ thymocytes were estimated to be CD5+‘““, on the basis that these displayed somewhat increased fluores- cence intensity compared to the same cells stained with the isotype matched control mAb (CT3). Some 5O-60% thy- mocytes were recorded as CD5 +‘Ow by 30 days and thereafter. Splenocytes stained distinctly with 2B1, the CD5 staining tending to be more intense in adults (Fig. 4). Some 3040% splenocytes from 12-37-day-old larvae expressed CD5, 50% were CDS+ at 45 days and some- what higher percentages emerged in adult life (Figs 24).
CD8, Flow Cytometry. Thymocytes and splenocytes stained distinctly with mAbs AM22 and F17. The overall staining pattern with these two mAbs was similar, but not identical (data not shown). Figures 2 and 3 reveal that at 7 days thymocytes were CD8-, whereas by 12 days about 70-80% had become CD8+. The level of CD8+ thymocytes was thereafter main- tained around 65-75%. The first time point that splenocytes were examined
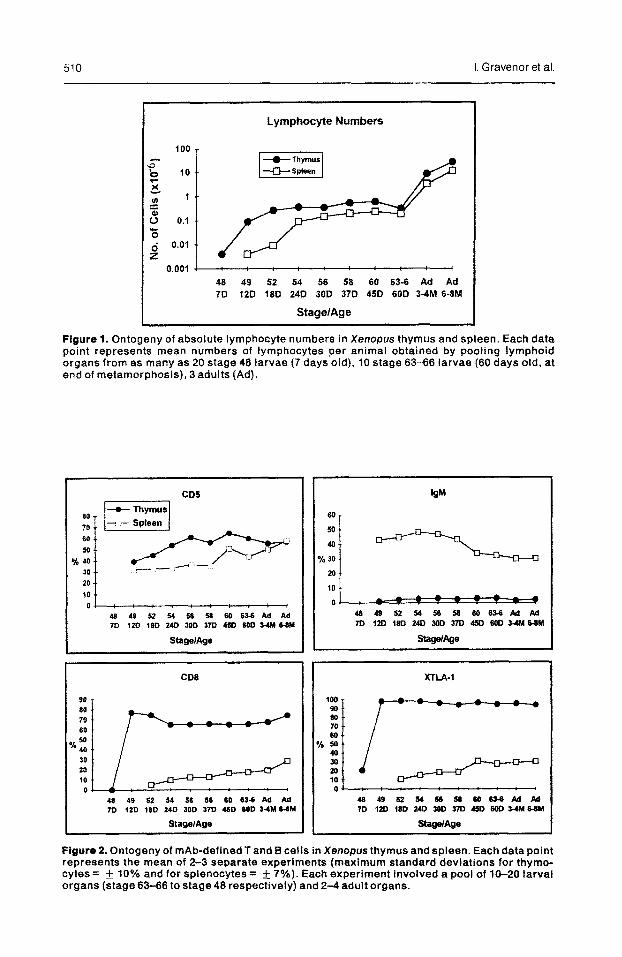
510 I. Gravenor et al.
100
53 ‘0 IO
5 ul 1
$ u 0.1
ij * P 0.01
0.001
Lymphocyte Numbers
!
48 49 52 54 56 58 60 63-s Ad Ad 70 120 180 24D 300 370 460 60D 3-4M 6-3M
Stagelhge
Figure 1. Ontogeny of absolute lymphocyte numbers in Xenopus thymus and spleen. Each data point represents mean numbers of lymphocytes per animal obtained by pooling lymphoid organs from as many as 20 stage 48 larvae (7 days old), 10 stage 63-66 larvae (60 days old, at end of metamorphosis), 3 adults (Ad).
48 49 52 54 56 33 60 63.6 Ad Ad 70 12D 180 240 300 37Il 4!iD SOD 3u1 MM
Stage/Age
XTLA-I
48 44 52 54 66 53 60 634 Ad Ad 484952s45sSllso63.sAdAd 10 12D 18D 240 JOD 370 46D 00D 34M 6-M 70 12D18D24LI3@D37D43DMw33-4MESM
Stage/Age Stage/Age
Figure 2. Ontogeny of mAb-defined T and B cells in Xenopus thymus and spleen. Each data point represents the mean of 2-3 separate experiments (maximum standard deviations for thymo- cytes = &- 10% and for splenocytes = f 7%). Each experiment involved a pool of lo-20 larval organs (stage 63-66 to stage 48 respectively) and 24 adult organs.
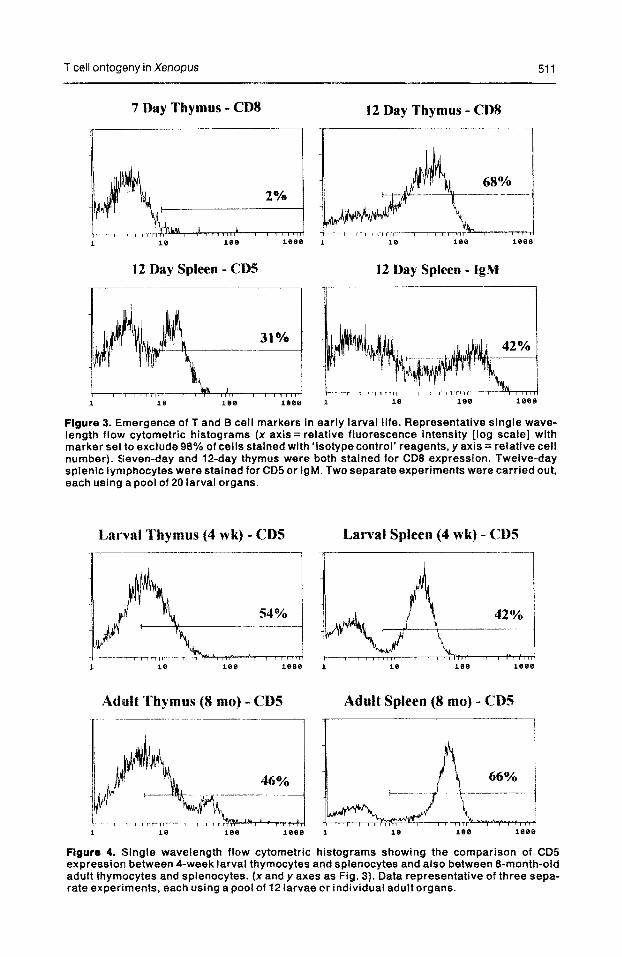
T cell ontogeny in Xenopus 511
7 Day Thymus - CDS
R1
12 Day Spleen - CDS
12 Day Thymus - CDS
,,I/, -wm, T
18 18.3 ieee
12 Day Spleen - IgM
Figure 3. Emergence of T and B cell markers in early larval life. Representative single wave- length flow cytometric histograms (x axis = relative fluorescence intensity [log scale] with marker set to exclude 98% of cells stained with ‘isotype control’ reagents, y axis = relative cell number). Seven-day and 12-day thymus were both stained for CD8 expression. Twelve-day splenic lymphocytes were stained for CD5 or IgM. Two separate experiments were carried out, each using a pool of 20 larval organs.
Larval Thymus (4 wk) - CDS Larva1 Spleen (4 wk) - CDS
Adult Tkymus (8 mo) - CD5
-1
Adult Spleen (8 mo) - CDS
Figure 4. Single wavelength flow cytometric histograms showing the comparison of CD5 expression between 4-week larval thymocytes and splenocytes and also between 8-month-old adult thymocytes and splenocytes. (x and y axes as Fig. 3). Data representative of three sepa- rate experiments, each using a pool of 12 larvae or individual adult organs.

512 I. Gravenor et al.
for CD8 expression by cytometry was at 18 days, when only 5% were positive. Percentages of CD8+ spleno- cytes then gradually rose during onto- geny to around 30% by 8 months.
Zmmunohistochemistry. AM22 staining of sections of larval thymus and spleen (beginning on days 6 and 11 respec- tively) provided histologic backup to the flow cytometric data on T cell develop- ment. Scattered CD8+ cells were first seen in the thymus of 8-day larvae and very occasional CD8+ cells were first identified in the 1Zday spleen. By 20 days the thymic cortex stained heavily for CD8, the medullary staining was far more scattered (Fig. 5A). In the spleen at 20 days CD8+ cells were prominent in a broad peripheral splenic zone (Fig. 5B), the central region at this stage being occupied largely by B cells (data not shown) marking the beginnings of white pulp formation.
XTLA-1. Just 20% thymocytes were XTLA-l+ at 7 days of age, whereas virtually all were stained with this anti- thymocyte mAb by 12 days and there- after (Fig. 2). In the spleen XTLA-l f expression was first assessed at 18 days, when some 10% splenocytes were posi- tive. Percentages of XTLA-l+ spleno- cytes then gradually increased to around 25% in young frogs.
ZgM. IgM+ cells were virtually un- detectable in the thymus. In contrast, the spleen was rich in B lymphocytes. About 40% IgM+ cells were found at 12 days (Figs 2, 3) and 50% were recorded in mid-larval life. Percentages of B cells gradually declined to around 30% in the adult spleen.
Use of Dual Staining to Reveal Emergence of a Peripheral T Cell-like Population in the Adult Thymus
After 12 days virtually all larval thymocytes are XTLA-1 + , including the CD8 + population (see Fig. 6A). In contrast, thymocytes from 8-month-old Xenopus adults contained a distinct, albeit small (< 5%) CD8+(high) population that was XTLA-l- (quadrant 1 [top left] of Fig. 6B). Moreover, dual staining of such thymocytes for CD8 and CD5 expression (Fig. 6C) revealed the CD8+(high) popula- tion also stains brightly for CD5 (quad- rant 2 [top right] of Fig. 6C): there is also a CDs- CD5+(high) thymocyte popula- tion (Fig. 6C, quadrant 4 [bottom right]). The distinct CD5 staining of these small thymocyte populations in the adult is more reminiscent of CD5 staining of splenocytes (Fig. 4). Moreover, the CD5f(high) populations of thymocytes also stain distinctly for MHC class II (quadrant 2 in Fig. 6D), another attribute of peripheral T cell staining seen in the adult spleen (see Figs 7, 8), MHC II molecules being ‘differentiation antigens’ in Xenopus (17).
Use of Dual Staining to Probe Ontogeny of MHC Class II Expression by Thymocytes and Splenic T Cells
Dual staining for class II and CD8 proved effective at illustrating the emer- gence of class II expression on a sizeable T-lineage population in adult Xenopus thymus. By 3-4 months of age > 25% CD8+ thymocytes (quadrant 2 of Fig. 7B) and some 20% CD8- thymocytes (quadrant 4 of Fig. 7B) are class II MHC+. Figure 7A indicates that some larval thymocytes (identified as T lineage cells by expression of CD8) also express some class II. In order to investigate MHC II expression on splenic T cells, the co-stain anti-CD5 rather than anti-CD8 is

T cell ontogeny in Xenopus
Figure 8. lmmunoperoxidase stained sections of 20”day-old larval thymus (A) and spleen (B), typical of that found in all three animals examined. In the thymus CD8 staining is heavy throughout the cortex (c), except for the lightly-stained subcapsular zone (s). Scattered CD8 cells are atso found in the medulla (m). Melanin pigment is noticeable in the cortex and is also associated with the thymus capsule (bottom right). Darkfield illumination of spieen highfights the distribution of brightly-stained CD8+cells. Unstained central region marks B cell-rich white pulp zone (wp). h-lag. (A) x 400, (B) x 100.
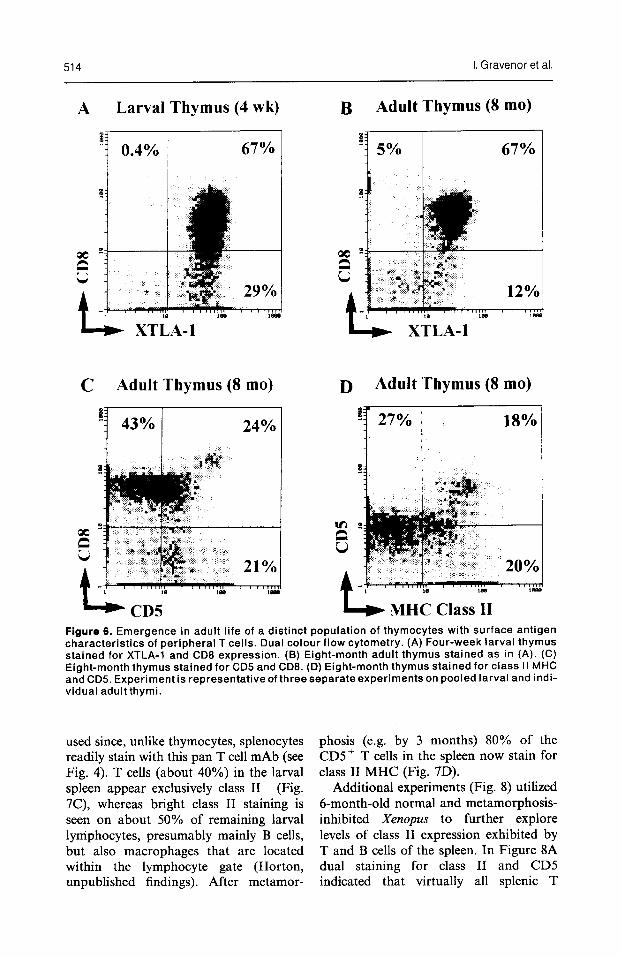
514 I. Gravenor et al.
A Larval Thymus (4 wk)
IL XTLA-1
C Adult Thymus (8 mo)
Figure 6. Emergence in adult life of a distinct population of thymocytes with surface antigen characteristics of peripheral T cells. Dual colour flow cytometry. (A) Four-week larval thymus stained for XTLA-1 and CD6 expression. (B) Eight-month adult thymus stained as in (A). (C) Eight-month thymus stained for CD5 and CDB. (D) Eight-month thymus stained for class II MHC and CD5. Exneriment is recresentative of threeseparate experiments on pooled larval and indi- vidual adult’thymi. ’
used since, unlike thymocytes, splenocytes readily stain with this pan T cell mAb (see Fig. 4). T cells (about 40%) in the larval spleen appear exclusively class II- (Fig. 7C), whereas bright class II staining is seen on about 50% of remaining larval lymphocytes, presumably mainly B cells, but also macrophages that are located within the lymphocyte gate (Horton, unpublished findings). After metamor-
B Adult Thymus (8 mo)
b XTLA-1
D Adult Thymus (8 mo)
k MHC Class II
phosis (e.g. by 3 months) 80% of the CD5+ T cells in the spleen now stain for class II MHC (Fig. 7D).
Additional experiments (Fig. 8) utilized 6-month-old normal and metamorphosis- inhibited Xenopus to further explore levels of class II expression exhibited by T and B cells of the spleen. In Figure 8A dual staining for class II and CD5 indicated that virtually all splenic T
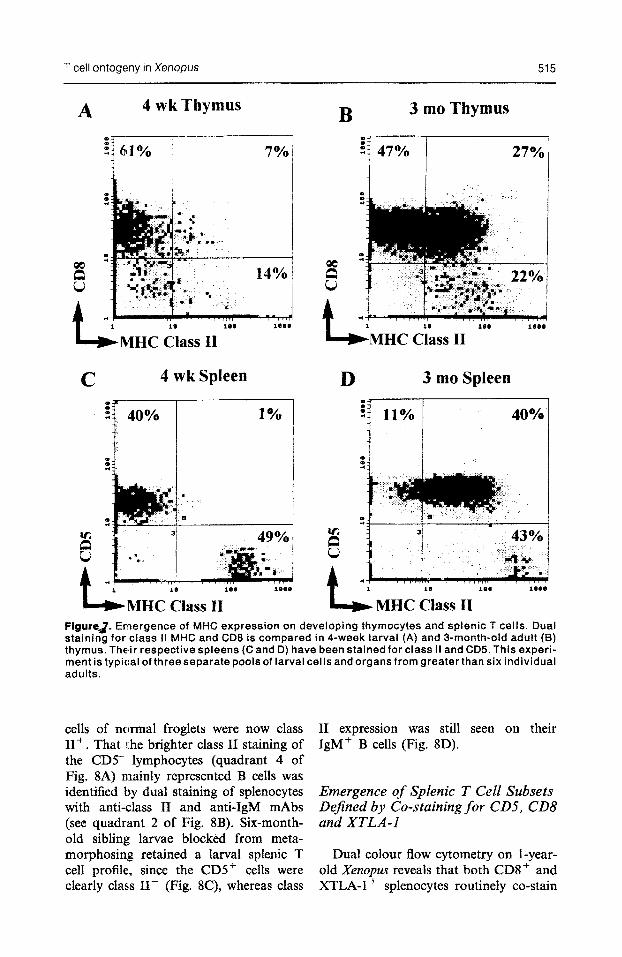
T cell ontogeny in Xenopus 515
A 4 wk Thymus
bMHC Class II
C 4 wk Spleen
! 40% r 1%
~MHC Class II
3 mo Thymus
27%
~MHC Class II
D 3 mo Spleen
: 11%
i
40%
MHC Class II FigureJ. Emergence of MHC expression on developing thymocytes and splenic T ceils. Dual staining for class II MHC and CD8 is compared in 4-week larval (A) and 3-month-old adult (8) thymus. Their respective spleens (C and D) have been stained for class II and CD5. This experi- ment is typical of three separate pools of larval cells and organs from greater than six individual adults.
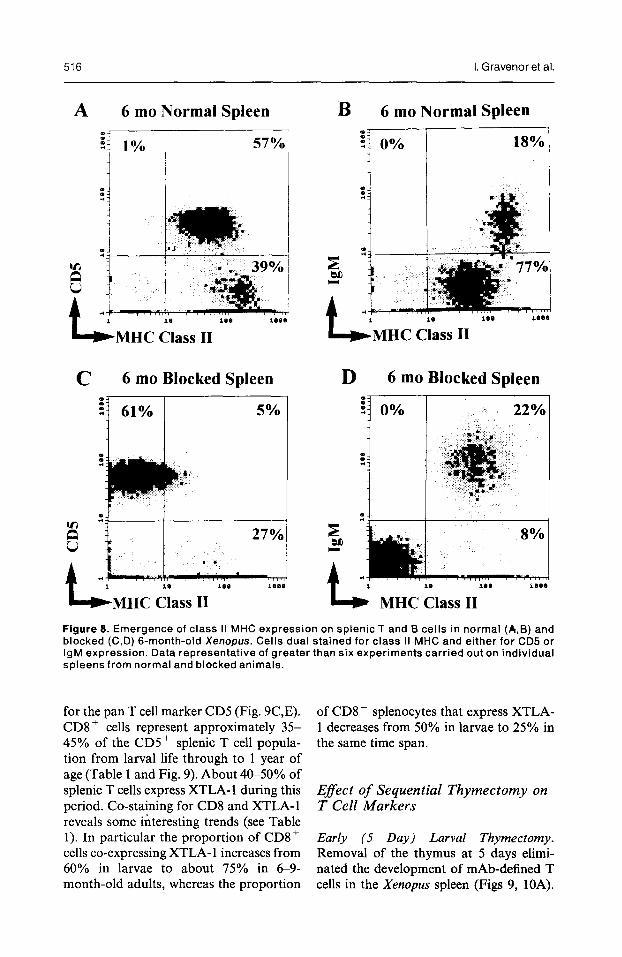
cells of normal froglets were now class II+. That the brighter class II staining of the CD5- lymphocytes (quadrant 4 of Fig. 8A) mainly represented B cells was identified by dual staining of splenocytes with anti-class II and anti-IgM mAbs (see quadcant 2 of Fig. 8B). Six-month- old sibling larvae blocked from meta- morphosing retained a larval splenic T cell profile, since the CD5+ cells were clearly class II- (Fig. 8C), whereas class
II expression was still seen on their IgMf B cells (Fig. 8D).
Emergence of Sphic T CelE Subsets Dejked by Co-staining for CD5, CD8 and XTLA-1
Dual colour flow cytometry on l-year- old ~e~opu~ reveals that both CD8+ and XTLA- 1 + splenocytes routinely co-stain
516 I. Gravenor et al.
A 6 mo Normal Spleen 4
‘i 1% 57% 1
WMHC Class II
c ii ::,,“‘“‘““““‘::
~MHC Class II
B 6 mo Normal Spleen
18%
~MHC Class II
D 6 mo Blocked Spleen
ii] 0% 1 : 22%
4 MHC Class II Figure 8. Emergence of class II MHC expression on splenic T and B cells in normal (A,B) and blocked (C,D) 6-month-old Xenopus. Cells dual stained for class II MHC and either for CD5 or IgM expression. Data representative of greater than six experiments carried out on individual spleens from normal and blocked animals.
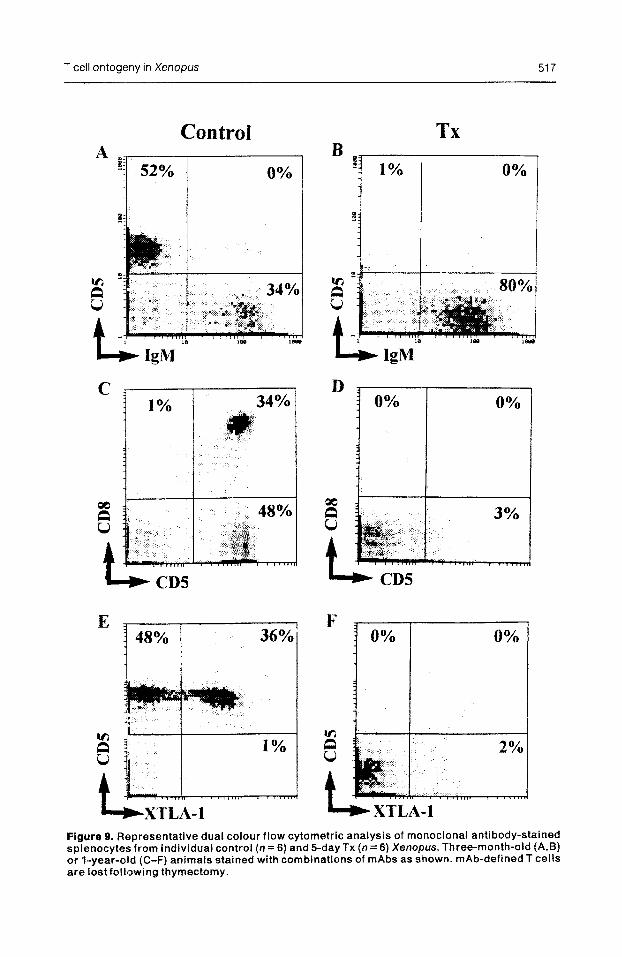
for the pan T cell marker CD5 (Fig. 9C,E). CD8 + cells represent approximately 35- 45% of the CD5 + splenic T cell popula- tion from larval life through to 1 year of age (Table 1 and Fig. 9). About 4&50% of splenic T cells express XTLA- 1 during this period. Co-staining for CD8 and XTLA-1 reveals some interesting trends (see Table 1). In particular the proportion of CD8+ cells co-expressing XTLA- 1 increases from 60% in larvae to about 75% in 69- month-old adults, whereas the proportion
of CD8- splenocytes that express XTLA- 1 decreases from 50% in larvae to 25% in the same time span.
Efect of Sequential Thymectomy on T Cell Markers
Early (5 Day) Larval Thymectomy. Removal of the thymus at 5 days elimi- nated the development of mAb-defined T cells in the Xenopus spleen (Figs 9, 1OA).
T cell ontogeny in Xenopus 517
Control
E 1 48% 36%
TX
0%
Figure 9. Representative dual colour flow cytometric analysis of monoclonal antibody-stained spienocytes from individual control (n = 6) and 5-day TX (n = 6) Xenopus. Three-month-old (A,B) or 1-year-o8d (C-F) animals stained with combinations of mAbs as shown. mAb-defined T cells are lostfolll~wing thymectomy.
518 I. Gravenor et al.
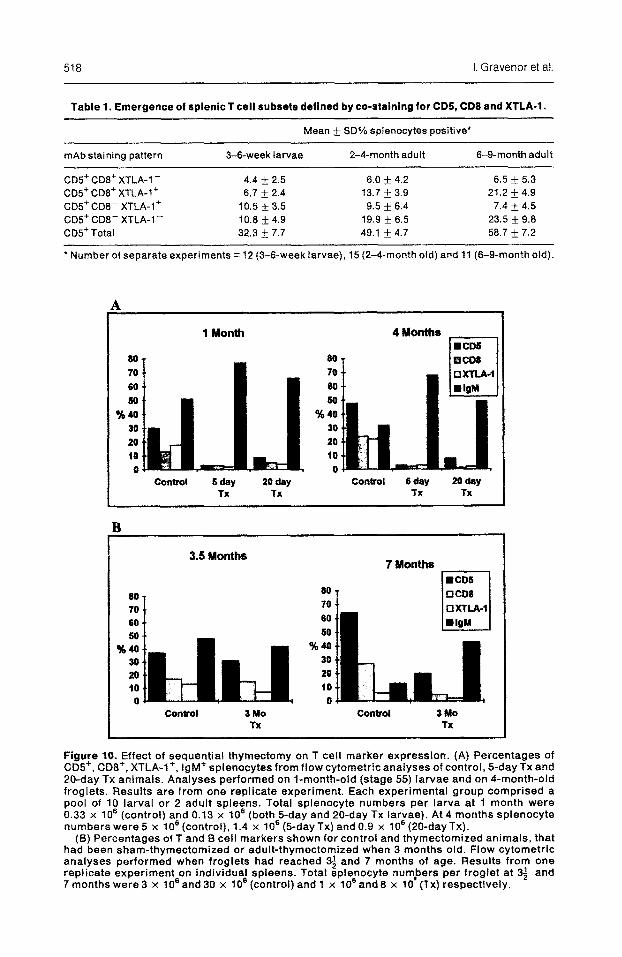
Table 1. Emergence of splenic T cell subsets defined by co-staining for CDS, CD8 and XTLA-1.
Mean f SD% splenocytes positive*
mAb staining pattern 3-&week larvae 2-4-month adult 6-g-month adult
CD5+ CD8+ XTLA-l- 4.4 f 2.5 6.0 + 4.2 6.5 + 5.3 C05+ CDS+ XTLA-1’ 6.7 & 2.4 13.7 + 3.9 21.2 * 4.9 CD!?+ CD8- XTLA-I+ 10.5 f 3.5 9.5 + 6.4 7.4 + 4.5 GD5+ CDB- XTLA-l- 10.6 k 4.9 19.9 + 6.5 23.5 k 9.6 CD!?Total 32.3 Ifr 7.7 49.1 + 4.7 58.7 + 7.2
l Number of separate experiments = 12 (3-f%week larvae), 15 (2-4-month old) and 11 (6-g-month old).
A
1 Month
80 80 70 70
80 60 so 50
%40 9440 30 30 20 20 IO 10 0 0
COYlWOl 6-y 20 day Control S&Y B&Y TX TX TX TX
3.5 Months 7 Months
Iiziiil
COntrol 3Nlo Contlol 3#ho TX TX
Figure 10. Effect of sequential thymectomy on T cell marker expression. (A) Percentages of CD!!+, CD8+, XTLA-l+, IgM+ sptenocytes from flow cytometric analyses of control, 5-day TX and 20-day TX animals. Analyses performed on l-month-old (stage 55) larvae and on 4-month-old froglets. Results are from one replicate experiment. Each experimental group comprised a pool of 10 larval or 2 adult spleens. Total splenocyte numbers per larva at 1 month were 0.33 x 10’ (control) and 0.13 x lo6 (both B-day and 20-day TX larvae). At 4 months spienocyte numbers were 5 x 706 (control), 1.4 x lo6 (5day Tx) and 0.9 x f06 (20-dayTx).
(5) Percentages of T and f3 cell markers shown for control and thymectomized animals, that had been sham-thymectomized or adult-thymectomized when 3 months old. Flow cytometric analyses performed when froglets had reached 3; and 7 months of age. Results from one replicate experiment on individual spleens. Total splenocyte numbers per froglet at 33 and 7 months were 3 x lo6 and 30 x lo8 (control) and 1 x lo6 and 8 x 10’ (TX) respectively.

T cell ontogeny in Xenopus 519
In young frogs (3 months old) CD5 expression was not evident in the TX spleen, whereas the percentage of IgM+ cells increa.sed from 34 to 80% after thymectomy (Fig. 9A,B). In spleens of l-year-old TX frogs all three T cell markers were ~detectable (Fig. 9D,F).
Late (20 Day) Larval Thymectomy. Animals thymectomized after T cell devel- opment in the spleen has begun also display dramatically reduced percentages of splenocytes expressing T cell markers (see Fig. lOA). Within just 1 week of 20- day thymectomy CD5+ percentages are reduced fro!m 30% to < loo/o and CD8 and XTLA-1 expression are only just above background levels. Larvae thymec- tomized at 5 days display no evidence of any T cell markers when 4 weeks old. By 4 months of age a small, but distinct, percentage (around 8% in the animals shown) of CD5+ IgM- splenocytes is maintained following 20-day thymectomy (Fig. lOA), whereas CD8 and XTLA-1 could no longer be detected. Thymectomy at 5 or 20 days reduced total splenocyte numbers by 66% or more (Fig. 10A legend).
Adult (3 faiths Thymectomy. The effect of adult thymectomy is illustrated in Fig. 10B. Two weeks post-thymectomy the percentages of splenocytes expressing CD5 and CD8 were comparable to control levels, whereas XTLA- 1 expres- sion was regularly reduced. Four months post adult thymectomy CD5+ levels in the spleen were about one-third the level seen in controls and all CDS+ cells were IgM-. Moreoever only 5% splenocytes of adult TX frogs now expressed CD8 (con- trast > 20% in controls) and XTLA-1 expression was no longer observed. Adult thymectomy causes a significant diminu- tion in spleen cell numbers, even within 2 weeks of the operation (see legend to Fig. 10B).
Discussion
Three anti-Xenopus T cell mAbs have been used here in concert to trace the emergence of T cell markers in Xenopus thymus and spleen from an early stage of lymphoid organ histogenesis. Dual colour flow cytometry has characterized the ontogeny of MHC class II expression on mAb-defined Xenopus T and B cells and has probed the emergence of T cell subset surface marker expression. The T cell specificity of the T cell mAbs has been confirmed by undertaking thymectomy experiments.
It is well documented that the Xenopus thymus buds from the pharyn- geal epitheli~ at about 3 days (25) and is then colonized by lymphoid stem cells at around 4 days (26,27). Here we show that in l-week-old, stage 48 larvae T- lineage di~erentiation in the thymus is still in its infancy, there being no detectable CDS’ lymphocytes and only 20% thymocytes express the T cell specific antigen XTLA- 1. Interestingly, XTLA-1 differentiation on thymocytes at 7 days (also revealed by Nagata [2X]) occurs coincidentally with the initial expression of MHC class II on the thymic epithelium (18). Thus MHC molecules may well be playing a crucial role in T cell education in Xe~opus from an early age, as they do in mammals (29). The flow cytometric experiments reported here indicate that XTLA-1 expression has developed on virtually all thymocytes by stage 49 (12 days of age) and this level of expression con- tinues thereafter. Flow cytometric obser- vations reveal a rather more rapid ontogeny of thymocyte expression of this T cell marker (during the second and third week of life) than Nagata was able to indicate from fluorescence micro- scopic observations (28). Emergence of CD8 expression on thymocytes has also occurred by 12 days, with some 80% now CD8’. Indeed immunohistochem- ical findings revealed that a few CD8 f

520 I. Gravenor et al.
cells can first be identified in the thymus at 8 days of age.
Our flow cytometric studies indicate that XT-l and the anti-CD8 mAbs stain thymocytes and peripheral splenic T cells (from larvae and adults) with similar fluorescence intensities. In contrast, we find that the 2Bl mAb (anti-CDS) stains larval and young adult thymus relatively poorly compared with its excellent staining properties on splenocytes. This may have led to our underestimating the percentage of CDS+ thymocytes from young animals, since others (15) have indicated that 8&90% Xenopus thymo- cytes are CD5+ from 10 days of age. Increased CD5 expression could well be an indication of thymocyte maturation, since this antigen is induced following in vitro culture of thymocytes (our unpub- lished observations) and shows aug- mented expression on thymus tumour cell lines (15,30). Moreover, use of the 2Bl mAb identified a small population of CD5+(high) cells in 8-month-old Xenopurr thymus that may represent ‘fully-differ- entiated’ T cells, since these stained for MHC II, but were XTLA-l-, surface properties expressed by many adult splenic T cells.
Combined use of B and T specific mAbs to stain larval and young adult Xenopus spleen revealed that B cell percentages gradually decline during ontogeny, whereas T cell percentages increase. Our findings concur with initial reports that CD5 is not constitutively expressed on Xenopus B cells, unlike the situation in many mammals (31). How- ever, it should be noted that this antigen can be induced by stimulation of B cells in vitro with phorbol ester provided that T cells are present in the culture (15,32).
Initial ontogenetic studies by Nagata with his XT-l mAb indicated that T cell differentiation in the Xenopus spleen had not begun by stage 50, but could be identified first at stage 52 (28). Although we confirm a low perce&ge of T cells (both XTLA-I+ and CD8+) exists in the
spleen at stage 52, T cells have in fact already emerged prior to this time. Thus 30% CD5+ cells are recorded in 12-day- old larvae and a scattering of CD8+ cells is found by immuno-histochemical analysis at this age (stage 49). T cells, therefore, begin to appear in the spleen within a few days of T-lineage cells first being detected in the thymus. This helps to explain earlier in vivo skin allograft experiments on young Xenopus larvae where lymphocytes can be seen invading skin allografts for the first time at stage 49 (33). Studies on in vitro proliferation responses of spleno- cytes to T cell mitogens were able to identify T cell activity by stage 51 (34).
Our studies on class II MHC expression confirm previous suggestions that both B and T cells of adult Xenopus express class II molecules, whereas amongst larval lymphocytes, only B cells are class II+ (18). Moreover, we confirm by perchlor- ate-blocking of metamorphosis, that class II expression on T cells is metamorphosis- dependent (19). Rollins-Smith and Blair (19) indirectly showed that adult Xenopus T cells were class II+ on the basis that there appeared only in the adult a major population of IgM+ class II+ cells. We have used dual staining with class II and anti-CD5 to directly identify that splenic T cells express this MHC type. However, co-staining for class II and either IgM or CD5 has led us to believe that B cells in Xenopus up to 6 months of age express more of the class II epitope recognized by AM20 than do T cells. This contrasts with previous findings based on single colour FACS analysis that indicate all adult Xenopus lymphocytes express equivalent levels of class II molecules (17). The constitutive expression of MHC class II by all adult Xenopus lymphocytes is unlike the situation in mice and humans, although class II molecules are found on T cells of various other mammalian species and on activated human T cells (see ref. 17, for discussion). The universal expression of class II by all Xenopus lymphocytes after metamorphosis is

T cell ontogeny in Xenopus 521
likely to increase the opportunity of adult T helper cells to undertake class II MHC- restricted antigen recognition, possibly beneficial in an animal that lacks specia- lized antigen presenting sites, such as lymph nodes and germinal centres (35). In the present study class II expression was also identified on many T-lineage cells developing within the adult thymus. Such MH’C expression on thymic lym- phocytes first became apparent in larval life, when, as noted elsewhere (19), a low percentage of thymocytes were recorded as class II’. Other authors (17,18) identi- fied class II on adult but not on larval Xenopus thymocytes.
Dual colour flow cytometry has revealed that throughout ontogeny XTLA-1 + and CD8+ splenocytes are overlapping, rather than distinct CD5+ T cell populations. When comparing larval splenocytes with splenocytes from 2-4-month or 6-9-month-old adults the proportion of the CD8+ T cell subset that co-express XTLA-1 increases whereas the proportion of CD8- T cells that are XTLA-1 + decreases. Presumably the CD8- T cells represent the helper T cell subset, although no antibody has been generated to date to define CD4+ Xenopus T cells. The ontogenetic changes occurring with respect to XTLA-1 expres- sion on CD8+ and CD8- T cell popula- tions suggest that XTLA-1, rather than being a lineage marker for T cells, may represent an activation antigen. Since larval cells express surface MHC class II but not class I (36), their class II-reactive lymphocytes (i.e. CD8- cells) are likely to be the predominant active T cell subset. With the emergence of cell surface MHC class I expression at metamorphosis (36), the adult CD8+ T cell subset would be expected to become centrally involved in immune reactivities. On the other hand it is’ possible that XTLA-1 represents a marker for ‘immature’ T cells, since a small population of thymocytes emerges in Xenopus adults that are CDjfhigh, XTLA-l-. This surface phenotype is
expressed by the majority of peripheral T cells, whereas thymic lymphocytes are predominantly CD5 +I”‘“, XTLA- 1 + .
The sequential thymectomy experi- ments outlined here indicate that when the thymus is removed at 20 days of larval life (after T cell peripheralization is under way) or even at 3 months of age, a dramatic loss of T cell markers, especially CD8 and XTLA-1 soon becomes evident in the spleen. The Xenopus thymus apparently continues to play a crucial role in establishing peripheral T cells both in late larval and early adult life. This prolonged thymic influence on the T cell compartment of Xenopus probably explains the findings of earlier functional studies following late thymectomy. For example, although larval thymectomy as late as 30 days has little if any influence on the establishment of alloimmune reac- tivity in vivo or in vitro, responsiveness to T cell mitogens is abrogated and antibody to thymus-dependent antigens, such as SRBC, is impaired, due to lack of helper T cells (9,10,21). Adult thymectomy has no apparent effect on antibody responses to SRBC or lymphocyte responsiveness to T cell mitogens (9,21), but does achieve removal of T cell subsets required in splenocyte reactivity to the hapten TNP presented on erythrocyte or Ficoll carriers (37,38). In view of the deficiencies in T cell marker expression noted in this paper, the cellular basis of the intact ‘thymus-depen- dent’ immune responses e.g. alloimmune responses) seen after late thymectomy would be interesting to determine.
The T cell marker ontogeny findings on euthymic Xenopus indicate that thy- mectomy at 1 week of age or earlier should provide a clean system for exploring whether T cells can develop extrathymically. Several laboratories, including our own, have previously indi- cated that some batches of such early-TX Xenopus can display immune responses indicative of T-like cells [e.g. chronic allograft rejection (5,13) and low level T cell mitogen reactivity (12)]. Further-

522 I, Gravenor et al.
more, in preliminary experiments some TX Xenopus at around 6 months of age began to display low levels of mAb- defined T cells markers in their spleen (4). This contrasts the experiments reported here where T cell surface anti- gens appear not to develop for at least 1
year in the spleen of 5-day TX animals. Absence of mAb-defined T cell develop- ment is not confined to the spleen, but is also found in blood and intestine of recent batches of Xenopus thymectomized at 5-7 days (39,40). Furthermore, func- tional studies on these same TX Xenopus
indicate a complete failure both to reject
allogeneic skin grafts and respond in vitro to the T cell mitogen Con A (32).
The overall conclusion to be drawn from
1.
2.
3.
4.
5.
6.
studies on TX Xenopus is that there is no substantial extra-thymic pathway of T cell development at the amphibian level of evolution. Indeed a complete failure of mhb-defined T cells is one scenario that can follow early thymic ablation.
Acknowledgements-This work was funded by
project grants from The Wellcome Trust, The Royal Society and The University of Durham (to J.D.H.), and through a BBSRC student- ship (to I.G.). We thank Dr. Alice Givan, Mr. Ian Dimmick and Mrs. S. Skinner for assis- tance with flow cytometry and Paul Loftus for
help with animal husbandry.
References
Rocha, B.; Vassalli, P.; Guy-Grand, D. The extrathymic T-cell development pathway. Immunol. Todav 13 : 449454: 1992. Lefrancois, L. -Extrathymic ‘differentiation of intraepithelial lymphocytes: generation of a 10. separate and unequal T-cell repertoire. Immunol. Today 12:436438; 1991. Dunon, D.; Cooper, M. D.; Imhof, B. A. Thymic origin and migration patterns of intest- inal yS T cells. J. exp. Med. 177 : 257-263; 1993. 11. Horton, J.D.; Horton, T. L.; Ritchie, P. Immune system of Xenopus: T cell biology. In: Tinsley, R. C.; Kobel, H. R., Eds. The biology of Xenopus. Symposium of the Zoological Society of 12. London, 68. Oxford: Oxford University Press; 279-299 (in press). Horton, J. D.; Manning, M. J. Response to skin allografts in Xenopus luevis following thy- mectomy at early stages of lymphoid organ maturation. Transplantation 14: 141-154; 1972. 13. Katagiri, C.; Tochinai, S. Ontogeny of thymus- dependent immune responses and lymphoid cell differentiation in Xenopus luevis. Devel. Growth Differ. 29 : 297-305; 1987.
leucocyte reactivity and phytohaemagglutinin responsiveness. In: Solomon, J. B.; Horton, J. D. Eds. Developmental immunology. Amsterdam: Elsevier; 1977; 283-290. Manning, M. J.; Collie, M. H. The ontogeny of thymic dependence in the amphibian Xenopus laevis. In: Solomon, J. B.; Horton, J. D. Eds. Developmental immunology. Amsterdam: Else- vier; 1977; 291-298. Turner, S. L.; Horton, T. L.; Ritchie, P.; Horton, J. D. Splenocyte response to T cell- derived cytokines in thymectomized Xenopus. Devel. Comp. Immunol. 15 : 319-328; 1991. Manning, M. J.; Donnelly, N.; Cohen, N. Thymus-dependent and thymus-independent components of the amphibian immune system. In: Wright, R. K.; Cooper, E. L. Eds. Phylogeny of thymus and bone marrow-bursa cells. Amsterdam: Elsevier; 1976; 123-132. Nagata, S., Cohen, N. Specific in vivo and non specific in vitro alloreactivities of adult frogs (Xenopus luevis) that were thymectomized during early larval life. Eur. J. Immunol. 13 : 541-545; 1983.
Du Pasquier, L.; Horton, J. D. The effect of 14. Nagata, S. T cell-specific antigen in Xenopus thymectomy on the mixed leucocyte reaction identified with a mouse monoclonal antibody: and phytohemagglutinin responsiveness in the biochemical characterization and species distri- clawed toad Xenopus laevis. Immunogenetics bution. Zool. Science 5 : 77-83; 1988. 3 : 105-112; 1976. 15. Jiirgens, J. B.; Gartland, L. A.; DuPasquier, L.; Du Pasquier, L.; Horton, J. D. Restoration of Horton, J. D.: CooDer. M. D. Identification of a antibody responsiveness in early thymectomized candidate CD5 homologue in the amphibian Xenopus by implantation of major histocompat- Xenopus laevis. J. Immunol. (in press). ibility complex-mismatched larval thymus. Eur. 16. Ibrahim, B.; Gartland, L. A.; Kishimoto, T.; J. Immunol. 12: 546551; 1982. Dzialo, R.; Kubagawa, H.; Bucy, R. P.; Cooper, Horton, J. D.; Sherif, N. E. H. S. Sequential M. D. Analysis of T cell development in thymectomy in the clawed toad: effect on mixed Xenopus. Fed. Proc. 5: 7651; 1991.

T cell ontogeny in Xenopus 523
17. Flajnik, M. F.; Ferrone, S.; Cohen, N.; DuPas- 30. quier, L. Evolution of the MHC: antigenicity and unusual tissue distribution of Xenopus (frog) class II molecules. Molec. Immunol. 27:451- 462; 1990. 31.
18. Du Pasquier, L.; Flainik, M. F. Expression of MHC class II antigens during Xenopis develop- ment. Devel. Immunol. 1 : 8595: 1990. 3’)
19. Rollins-Smith, L. A.; Blair, P.. Expression of JL’ class II major histocompatibility complex anti- gens on adult T cells in Xenopus is metamor- phosis-dependent. Devel. Immunol. 1: 97-104;
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
i990. - Nieuwkoop, P. D.; Faber, J. Normal table of Xenopus I’uevis (Daudin). Amsterdam: North Holland; 1967. Horton, J. D.; Rimmer, J. J.; Horton, T. L. Critical role of the thymus in establishing humoral immunity in amphibians: studies on Xenopus tbymectomized in larval and adult life. Devel. Coma. Immunol. 1: 119-130; 1977. Langeberg, L.; Ruben, L. N.; Clothier, R. H.; Shiigi, S. The characterization of the toad splenocytes which bind mouse anti-human IL-2 receptor antibody. Immun. Letters 16 : 43-48; 1987. Chen, C. H.; Ager, L. L.; Gartland, G. L.; Cooper, Id. D. Identification of a T3/T cell receptor complex in chickens. J. exp. Med. 164 : 375-380; 1986. Horton, J. D.; Horton, T. L. Development of transplant.ation immunity and restoration experiments in the thymectomized amphibian. Am. Zool. 15 : 73-84; 1975. Manning, M. J.; Horton, J. D. Histogenesis of lymphoid organs in larvae of the South African clawed toad, Xenopus laevis (Daudin). J. Embryol. ~exp. Morph. 22 : 265-277; 1969. Tochinai, S. Thymocyte stem cell inflow in Xenopus luevis, after grafting diploid thymic rudiments into triploid tadpoles. Devel. Comp. Immunol. 2 : 627635; 1978. Turpen, J. B.; Smith, P. B. Precursor immigra- tion and thymocyte succession during larval development and metamorphosis in Xenopus. J. Immunol. 142: 4147; 1989. Nagata, S. Development of T lymphocytes in Xenopus luevis: appearance of the antigen recognized by an anti-thymocyte mouse mono- clonal antibody. Devel. Biol. 114 : 389-394; 1986. Kruisbeek, A. M. Development of ap T cells. Current Opinion Immunol. 5 : 227-234; 1993.
33.
34.
35.
36.
37.
38.
39.
40.
Robert, J.; Guiet, C.; DuPasquier, L. Lymphoid tumours of Xenopus luevis with different capa- cities for growth in larvae and adults. Devel. Immunol. 3 : 297-307; 1994. Herzenberg, L. A.; Kantor, A. B. B cell lineages exist in the mouse. Immunol. Today 14 : 79-83; 1993. Horton, J. D.; Horton, T. L.; Ritchie, P.; Gravenor, I. & Horsham, K. Immune system development in Xenopus. In: Stolen, J. Ed. Modulators of immune responses: the evolu- tionary trail. New Haven: SOS Publications, in press. Horton, J. D. Ontogeny of the immune responses to skin allografts in relation to lymphoid organ development in the amphibian Xenonus laevis Daudin. J. EXP. Zool. 170 : 449- 466; i969. Rollins-Smith, L. A.; Parsons, S. C. V.; Cohen, N. During frog ontogeny, PHA and Con A responsiveness of splenocytes precedes that of thymocytes. Immunology 52 : 491-500; 1984. Horton, J. D.; Ratcliffe, N. A. Evolution of Immunity. In: Roitt, I., Male, D.; Brostoff, J. Eds. Immunology, Fourth Edn. London: Mosby; 15.1-15.22, 1995. Flajnik, M. F.; Hsu, E.; Kaufman, J. F.; Du Pasquier, L. Changes in the immune system during metamorphosis of Xenopus. Immunol. Today 8 : 58-64; 1987. Gruenwald, D. A.; Ruben, L. N. The effect of adult thymectomy upon helper function in Xenopus laevis, the South African clawed toad. Immunology 38: 191-194; 1979. Ruben, L. N.; Mette, S. A.; Horton, J. D.; Edwards, B. F.; Tournefier, A.; Stack, J. Carrier dependence in hapten responses, tolerance and memory in Xenopus luevis, the South African clawed toad. In: Manning, M. J. Ed. Phylogeny of immunological memory. Amsterdam: North Holland; 1980: 207-216. Horton, J. D.; Horton, T. L.; Ritchie, P.; Gravenor, I; Gartland, L.; Cooper, M. D. Use of monoclonal antibodies to demonstrate absence of T cell development in early-thymec- tomized Xenopus. Devel. Comp. Immunol. 18:S95; 1994. Horton, J. D.; Horton T. L.; Ritchie, P.; Dzialo, R.: Gartland, L.: Coooer, M. D. The thvmus is essential for ‘T cell development in an -amphi- bian. (In prep.).











![PROBLEMATYKA BADAŃ WITRAŻY ŚREDNIOWIECZNYCH [Issues in medieval stained glass research]](https://img.dokumen.tips/doc/110x75/63528c6f5c21d80fde0b135f/problematyka-badan-witrazy-sredniowiecznych-issues-in-medieval-stained-glass.jpg)

















