Embed Size (px)
Citation preview

1
1
Novel giant siphovirus from Bacillus anthracis features unusual 2
genome characteristics and relatedness to Spounavirinae 3
4
Holly H. Ganz1#, Christina Law1, Martina Schmuki2, Martin J. Loessner2, Richard Calendar3, Wayne M. 5
Getz1,4, Jonas Korlach5, and Jochen Klumpp2* 6
7
1 University of California, Berkeley, Department of Environmental Science, Policy & Management, 8
Berkeley, CA, USA 9
2 Institute of Food, Nutrition and Health, ETH Zurich, Switzerland 10
3 University of California, Berkeley, Department of Molecular and Cellular Biology, Berkeley, CA, USA 11
4 School of Mathematical Sciences, University of KwaZulu-Natal, Durban, South Africa 12
5Pacific Biosciences, Menlo Park, CA, USA 13
14
Running title: Giant siphovirus specific for Bacillus ACT group 15
16
Key words: Siphovirus, bacteriophage, Bacillus anthracis, genome, PacBio, SMRT sequencing 17
18
* Corresponding author: 19
Jochen Klumpp: Institute of Food, Nutrition and Health, ETH Zurich, Schmelzbergstrasse 7, 8092 20
Zurich, Switzerland. Phone: +41-44-6325378; Fax: +41-44-6321266; email: [email protected] 21
22
# Present address: 23
Holly Ganz: University of California, Davis, School of Veterinary Medicine, Davis, CA 95616 USA 24
25

2
Abstract 26
We present vB_BanS-Etosha, a novel temperate phage isolated from Bacillus 27
anthracis. Etosha is a giant siphovirus, featuring a long, flexible and non-contractile 28
tail of 440 nm and an isometric head of 82 nm. We induced Etosha phage from two 29
different isolates of a B. anthracis genotype responsible for many anthrax outbreaks 30
occurring in wildlife in Etosha National Park, Namibia. The phage genome is the 31
largest sequenced Bacillus siphovirus, containing 168,876 bp and 269 ORFs. 32

3
We present a novel virus species isolated from Bacillus anthracis, the agent 33
responsible for anthrax infections in wildlife, livestock and humans [1]. Along with 34
Bacillus cereus and Bacillus thuringiensis, B. anthracis is a member of the Bacillus 35
ACT group [2]. Genomic studies have identified a number of putative prophages in 36
the Bacillus ACT group (e.g. [3, 4]). Lysogeny occurs commonly in B. anthracis [5, 6] 37
and may play an essential role in its life cycle [7]. 38
We obtained isolates of the siphovirus from two zebra carcass sites in Etosha 39
National Park (ENP), Namibia, a 22,915 km2 wildlife reserve where anthrax infections 40
occur regularly (reviewed in [8]). We isolated bacteriophage from a B. anthracis 41
colony obtained from a swab from 2006 (GPS coordinates: -18.99736, 15.81584) and 42
from a soil sample collected in 2010 (GPS coordinates: -19.1731, 15.92603). Isolates 43
of B. anthracis from the two carcass sites belong to genotype 6 in the A3a group, 44
which has dominated outbreaks in ENP for more than 30 years [8]. We induced 45
phage from bacterial isolates by enrichment culture and mitomycin C induction [9, 46
10]. Both phages were morphologically similar and unusually large. It is intriguing that 47
such a large siphovirus is rarely isolated and yet appeared twice in spatially and 48
temporally separate samples. 49
Phage preparations were purified and concentrated using standard techniques 50
[10, 11]. Plaque assays were performed with a spore preparation of an avirulent B. 51
anthracis strain (6602 R1, [12]) by LB agar soft-agar overlay method [13]. Serial 52
dilutions of plaque-extracts were soft-agar plated with strain 6602 R1 [12]. Phage 53
lysates were PEG precipitated [14] and purified by cesium chloride density gradient 54
centrifugation [10]. 55
We used the spot-on-the-lawn method [15] for host-range testing (Table 1). 56
The phage infects the Bacillus ACT group and is particularly adapted to B. anthracis. 57

4
Etosha lysed 25% (6/24) of B. cereus strains, 40% (2/5) of B. thuringiensis strains 58
and 86% (6/7) of B. anthracis strains tested. The phage did not lyse any other 59
bacteria tested (Table 1). Thus, Etosha is a narrow host-range, species-specific virus, 60
with potential suitability for biocontrol approaches of B. anthracis. 61
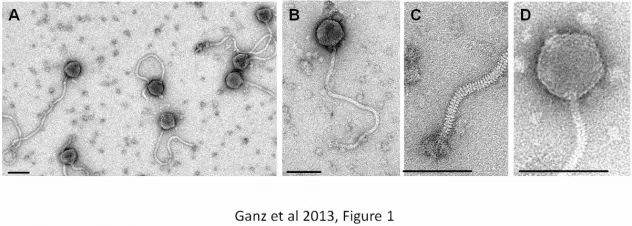
TEM images have been acquired from a preparation of pure phage particles 62
negatively stained with 2% uranyl acetate on carbon-coated copper grids (Quantifoil, 63
Jena, Germany) and observed using a Philips CM12 microscope at 120 kV 64
acceleration voltage with a Gatan Orius digital camera. Etosha exhibits typical 65
siphovirus morphology (order Caudovirales), and has a long, flexible and non-66
contractile tail of 440 nm (not including baseplate structure) and an isometric head of 67
82 nm in diameter (Figure 1 A, B). Individual tail striations and a baseplate structure 68
with appendages are visible (Figure 1C). The head features visible capsomers 69
(Figure 1D) similar to those observed in other large bacteriophages [16]. To our 70
knowledge, Etosha is the largest sequenced siphovirus infecting Bacillus. Two larger 71
Bacillus siphoviruses are known but not characterized, B. mycoides phage N5 and B. 72
thuringiensis phage II, which are speculated to be identical (H.-W. Ackermann, 73
personal communication). 74
We sequenced DNA from the isolates using a single-molecule approach 75
(Pacific Biosciences RS) with 10 kb and 800 bp insert libraries (C2 chemistry) and 76
used one SMRT Cell for each library. We used the standard error-correction workflow 77
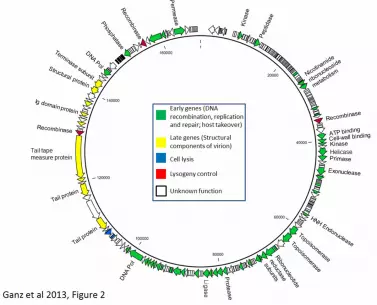
and SMRT portal software 1.3.1 for assembly of 36166 post-filter reads (2582 bp 78
average read length), resulting in one large contig with an average coverage of 550-79
fold (Figure 2). Both phage isolates were identical. A repeat structure of 284 bp at the 80
genome end was identified during assembly and confirmed in restriction profiles 81
(Figure 3). Methylome analysis revealed no base modifications in the genome. The 82
genome sequence is 168,876 bp in length. Open reading frames were predicted by 83

5
RAST [17] and edited manually. Etosha features 272 open reading frames, 17 tRNA 84
and 2 pseudo-tRNA genes (Figure 2). The GC content is 34%, similar to published 85
genome sequences of B. anthracis. The genome sequence was deposited at 86
GenBank under accession number KC481682. 87
Like many siphoviruses, the genome is structured in functional modules. The 88
early gene cluster (genes for DNA replication, modification and repair, host takeover 89
and nucleotide metabolism) spans roughly 70% of the genome, indicating active 90
participation of virus-encoded genes in the metabolic processes associated with 91
replication in the host cell. It is notable that the Etosha genome encodes three 92
tyrosine integrase/recombinase enzymes of the Cre/XERD type (gp94, gp227, 93
gp255; [18, 19]) which exhibit no nucleotide homology to each other. Etosha features 94
a temperate lifestyle and these three enzymes may serve as means to integrate into 95
different attB sites and ensure a large host range for lysogeny. Further work will 96
elucidate the specificity and activity of the three recombinases. We also note the 97
presence of two Ig-domain containing proteins, gp233 and gp213 [19, 20], which may 98
play accessory roles during infection [21]. 99
Interestingly, Etosha features distributed homologies in its structural proteins 100
to SPO1-related phages A511, A9, LP65 and SPO1 [16]. This finding is very unusual 101
because SPO1-related phages belong to an unrelated family of bacteriophages 102
(Myoviridae); and Etosha is a temperate phage, while SPO1-related phages are 103
strictly virulent. Etosha also displays individual capsomers thought to be a hallmark of 104
the SPO1-related phages (Figure 1) [16]. Virion proteins of Etosha were separated 105
on a 10-20% SDS gradient PAGE. Resulting bands were extracted and protein 106
content identified by mass spectrometry [22]. Six bands were allocated to gene 107
products. The tape measure protein is present in two protein bands of 280 and 100 108
kDa in size, presumably because of instability of the large protein. gp206 features an 109

6
estimated mass of 38 kDa and gp199 and 207 were identified in bands of 26.5 and 110
19 kDa, respectively. Etosha features an unusually long tail of 440 nm, which 111
corresponds with the large size of the tape measure protein (3123 aa) [23, 24] but the 112
protein is disproportionately large in comparison to other sequenced bacteriophages. 113
The large unknown gene 221 likely encodes for a tail fiber component, with the C-114
terminus featuring significant homologies to Cellobiosidase, S-layer associated 115
endoglucanase or glycoside hydrolase domains. 116
In conclusion, we present vB_BanS-Etosha, a novel temperate phage 117
obtained from B. anthracis that is specific to the Bacillus ACT group. To our 118
knowledge Etosha is the largest sequenced siphovirus infecting Bacillus organisms. 119
Its giant head holds a genome of 168 kb. In addition to potential effects on activity 120
and environmental survival of its host, Etosha may have utility in detection and 121
control of B. anthracis. 122
123

7
Acknowledgments 124
We thank Margaret Smith for help with recombinase identification, Matthew Boitano 125
and Tyson A. Clark for genome assembly and initial analysis, and Martina Kusters, 126
Wendy C. Turner and Wilferd Versfeld for field support. Field sampling was 127
authorized by the Namibian Ministry of Environment and Tourism (MET) under permit 128
number 1448/2009 to HHG. We are grateful to the scientific staff at the Etosha 129
Ecological Institute (EEI) for facility resources and logistical support. The isolates 130
studied here were obtained from EEI carcass numbers EB060318-01WV and 131
EB100228-01MK. This research was funded by NIH Grant GM083863 to WMG. CL 132
was supported by the University of California, Berkeley, College of Natural 133
Resources-Biology Scholar’s Program, a Rosberg-Geist Fellowship from the Center 134
for African Studies and an Undergraduate Merit Scholarship from the Institute of 135
International Studies. 136
137

8
References 138
1. Carter KC. 1988. The Koch-Pasteur dispute on establishing the cause of 139
anthrax. Bull Hist Med. 62: 42-57. 140
2. Okstad OA, Kolsto A-B. 2011. Genomics of Bacillus Species, p. 29-55. In 141
Wiedmann M and Zhang W (ed), Genomics of Foodborne Pathogens. 2011, 142
Springer, New York. 143
3. Stromsten NJ, Benson SD, Burnett RM, Bamford DH, Bamford JK. 2003. 144
The Bacillus thuringiensis linear double-stranded DNA phage Bam35, which is 145
highly similar to the Bacillus cereus linear plasmid pBClin15, has a prophage 146
state. J Bacteriol. 185: 6985-9. 147
4. Rasko DA, Altherr MR, Han CS, Ravel J. 2005. Genomics of the Bacillus 148
cereus group of organisms. FEMS Microbiol Rev. 29: 303-29. 149
5. Buck CA, Anacker RL, Newman FS, Eisenstark A. 1963. Phage Isolated 150
from Lysogenic Bacillus anthracis. J Bacteriol. 85: 1423-30. 151
6. Saile E, Koehler TM. 2006. Bacillus anthracis multiplication, persistence, and 152
genetic exchange in the rhizosphere of grass plants. Appl Environ Microbiol. 153
72: 3168-74. 154
7. Schuch R, Fischetti VA. 2009. The secret life of the anthrax agent Bacillus 155
anthracis: bacteriophage-mediated ecological adaptations. PLoS One. 4: 156
e6532. 157
8. Beyer W, Bellan S, Eberle G, Ganz HH, Getz WM, Haumacher R, Hilss KA, 158
Kilian W, Lazak J, Turner WC, Turnbull PC. 2012. Distribution and 159
molecular evolution of Bacillus anthracis genotypes in Namibia. PLoS Negl 160
Trop Dis. 6: e1534. 161
9. Van Twest R, Kropinski AM. 2009. Bacteriophage enrichment from water 162
and soil. Methods Mol Biol. 501: 15-21. 163
10. Sambrook J and Russell DW. 2001. Molecular Cloning - A Laboratory 164
Manual. 3rd ed., Vol. 1-3. Cold Spring Harbor Laboratory Press, New York. 165
11. Klumpp J, Dorscht J, Lurz R, Bielmann R, Wieland M, Zimmer M, 166
Calendar R, Loessner MJ. 2008. The Terminally Redundant, Nonpermuted 167
Genome of Listeria Bacteriophage A511: a Model for the SPO1-Like 168
Myoviruses of Gram-Positive Bacteria. J Bacteriol. 190: 5753-5765. 169
12. Green BD, Battisti L, Koehler TM, Thorne CB, Ivins BE. 1985. 170
Demonstration of a capsule plasmid in Bacillus anthracis. Infect Immun. 49: 171
291-7. 172
13. Adams MH. 1959. Methods of study of bacterial viruses, p. 443-457. In 173
Bacteriophages. Interscience publishers, Inc., New York. 174
14. Yamamoto KR, Alberts BM, Benzinger R, Lawhorne L, Treiber G. 1970. 175
Rapid bacteriophage sedimentation in the presence of polyethylene glycol and 176
its application to large-scale virus purification. Virology. 40: 734-44. 177
15. Loessner MJ. 1991. Improved procedure for bacteriophage typing of Listeria 178
strains and evaluation of new phages. Appl Environ Microbiol. 57: 882-4. 179

9
16. Klumpp J, Lavigne R, Loessner MJ, Ackermann HW. 2010. The SPO1-180
related bacteriophages. Arch Virol. 155: 1547-1561. 181
17. Aziz RK, Bartels D, Best AA, DeJongh M, Disz T, Edwards RA, Formsma 182
K, Gerdes S, Glass EM, Kubal M, Meyer F, Olsen GJ, Olson R, Osterman 183
AL, Overbeek RA, McNeil LK, Paarmann D, Paczian T, Parrello B, Pusch 184
GD, Reich C, Stevens R, Vassieva O, Vonstein V, Wilke A, Zagnitko O. 185
2008. The RAST Server: rapid annotations using subsystems technology. 186
BMC Genomics. 9: 75. 187
18. Smith MC Thorpe HM. 2002. Diversity in the serine recombinases. Mol 188
Microbiol. 44: 299-307. 189
19. Kilcher S, Loessner MJ, Klumpp J. 2010. Brochothrix thermosphacta 190
bacteriophages feature heterogeneous and highly mosaic genomes and utilize 191
unique prophage insertion sites. J Bacteriol. 192: 5441-5453. 192
20. Kelly G, Prasannan S, Daniell S, Fleming K, Frankel G, Dougan G, 193
Connerton I, Matthews S. 1999. Structure of the cell-adhesion fragment of 194
intimin from enteropathogenic Escherichia coli. Nat Struct Biol. 6: 313-8. 195
21. Fraser JS, Yu Z, Maxwell KL, Davidson AR. 2006. Ig-like domains on 196
bacteriophages: a tale of promiscuity and deceit. J Mol Biol. 359: 496-507. 197
22. Marti R, Zurfluh K, Hagens S, Pianezzi J, Klumpp J, Loessner MJ. 2013. 198
Long tail fibers of the novel broad host range T-even bacteriophage S16 199
specifically recognize Salmonella OmpC. Mol Microbiol. 87: 818-834. 200
23. Katsura I. 1987. Determination of bacteriophage lambda tail length by a 201
protein ruler. Nature. 327: 73-5. 202
24. Katsura I. 1990. Mechanism of length determination in bacteriophage lambda 203
tails. Adv Biophys. 26: 1-18. 204
205
206

10
Figure legends 207
Figure 1: Electron microscopy of Etosha phage. A. Preparation overview. B. Close-208
up of single phage particle. C. Details of the phage tail distal end. D. Details of the 209
phage head structure. Individual capsomers are visible, an observation previously 210
made for SPO1-related phages ([16]). Scale bars represent 100 nm. 211
212
Figure 2: Genome map of phage Etosha. Open reading frames are drawn to scale 213
and transcription direction is indicated by arrows. Selected proteins with putative 214
function are labeled. Genetic modules (i.e. structural genes, early genes) are 215
indicated by coloring. 216
217
Figure 3: Restriction profile of Etosha phage. Clear band separation up to 24 kb in 218
size could be achieved and the restriction profiles match with the sequenced genome 219
size. The terminal redundancy location and size was determined from the fragment 220
sizes as previously described [10, 11]. Enzymes used: 1: Alw44I (NEB); 2: Eco91I 221
(Fermentas); 3: NheI (NEB); 4: PacI (NEB); 5: SwaI (NEB); 6: Van91I (Fermentas); 7: 222
XcmI (NEB). M1: Lambda 19 Mix Size standard (Fermentas); M2: 1kb size standard 223
(Fermentas). Numbers to the left indicate band size in kb. 224

Table 1: Host range of Etosha on different Bacillus and non-Bacillus strains. The presence of plaques (successful infection of this strain) is indicated by + and an absence is indicated by -.
Strain name Organism Notes Source Infection6602 R1 Bacillus anthracis pXO1-pXO2 negative {Green, 1985 #228} +Sterne Bacillus anthracis pXO2 negative Institut Pasteur #7702 +Weybridge UM44 Bacillus anthracis pXO2 negative {Green, 1985 #228} +Ames-non reverting
Bacillus anthracis pXO2 negative U.S. Dept. of Agriculture,
Ames, Iowa +
Ames Bacillus anthracis U.S. Dept. of Agriculture,
Ames, Iowa +
Vollum 1b Bacillus anthracis Laboratory Strain +
PAK-1 Bacillus anthracis Pakistan isolate, M. Hugh-
Jones collection -
LA 925 Bacillus cereus CHUV -ATCC 14579 Bacillus cereus ATCC -ATCC 11778 Bacillus cereus ATCC +ATCC 10702 Bacillus cereus ATCC -ATCC 10876 Bacillus cereus ATCC +DSM 2302 Bacillus cereus DSM -BO 366 Bacillus cereus This study -BO 372 Bacillus cereus This study -BO 493 Bacillus cereus This study -DSM 4218 Bacillus cereus DSM -ATCC 33019 Bacillus cereus ATCC +ATCC 14737 Bacillus cereus ATCC +DSM1274 Bacillus cereus DSM -ATCC 27522 Bacillus cereus ATCC -NCTC 11143 Bacillus cereus NCTC -NCIMB 8705 Bacillus cereus NCIMB +ATCC 6464 Bacillus cereus ATCC -B346 Bacillus cereus Mouse isolate -DSM360 Bacillus cereus DSM -HER1399 Bacillus cereus HER +WSBC 10530 Bacillus cereus WSBC -WSBC 10556 Bacillus cereus WSBC -WSBC 10566 Bacillus cereus WSBC -WSBC 10583 Bacillus cereus WSBC -DSM4421 Bacillus thuringiensis DSM -WSBC 10204 Bacillus thuringiensis WSBC -HER1211 Bacillus thuringiensis HER +Kurstaki Bacillus thuringiensis R. Calendar, UC Berkeley -ATCC 10792 Bacillus thuringiensis ATCC +DSM168 Bacillus subtilis DSM -DSM675 Bacillus subtilis DSM -ATCC 23059 Bacillus subtilis ATCC -DSM395 Bacillus sphaericus DSM -DSM90 Bacillus megaterium DSM -WSBC 10550 Bacillus weihenstephanensis WSBC -WSLC 3009 Listeria ivanovii WSLC -ATCC BAA-679 Listeria monocytogenes ATCC -PSK Staphylococcus aureus Laboratory Stock -Twort Staphylococcus aureus Laboratory Stock -414 Staphylococcus epidermidis Laboratory Stock -100655 Staphylococcus epidermidis Laboratory Stock -602 Staphylococcus epidermidis Laboratory Stock -DT7155 Salmonella typhimurium Laboratory Stock -CGSC 4401 Escherichia coli CGSC -DSM 20560 Streptococcus salivarius DSM -NZ9000 Lactococcus lactis Laboratory Stock -ATCC 19433 Enterococcus faecalis ATCC -

CHUV: Strain collection of the Centre Hospitalier universitaire Vaudois, Switzerland; HER: Félix d'Hérelle Reference Center for bacterial viruses, Laval, Canada; ATCC: American Type Culture Collection; NCTC: National Collection of Type Cultures; DSM: Deutsche Sammlung von Mikroorganismen; NCIMB: National Collection of Industrial Bacteria; WSBC: Weihenstephan Bacillus Collection; WSLC= Weihenstephan Listeria Collection. CGSC: Coli Genetic Stock Center, Yale, USA