Embed Size (px)
Citation preview

Biswas, K. K. and Oono, Y.
− 45 −
New Insights to Auxin Signaling through The Use of Small MoleculesKamal Kanti Biswas and Yutaka Oono*Medical and Biotechnological Application Division, Japan Atomic Energy Agency (JAEA), Takasaki 370-1292, Japan
Abstract
Elucidating the perception mechanism of auxin has been a long-standing goal in plant biology. To understand the mechanism, studies on the chemical structure and activity relationships of auxin-related small molecules have been done for more than half a century. Today, aided mainly by Arabidopsis genetics, researchers have identified the core complexes for auxin perception, including the auxin receptors TIR1/AFBs, SKP2 and ABP1. Forward chemical screening using auxin response markers has also played an important role in identifying small molecules affecting the auxin-signaling pathways and led to the screening of additional auxin related mutants. This approach has helped in identifying several unique molecular components of the auxin-signaling pathways, and discovering that would have been impossible using traditional screening of mutants against auxin. This review is focused on recently acquired knowledge of multiple auxin signaling pathways and related small compounds that can be used to dissect auxin signal transduction pathways. In addition, the involvement of distinct auxin-signaling machineries during gravity response is discussed. ©2011 Jpn. Soc. Biol. Sci. Space; Article ID: 112502007
Introduction
The plant hormone auxin is an important growth regulator that controls many developmental processes, including tropisms. How auxin is perceived by plant cells has been one of the most fundamental questions for plant physiologists since auxin was discovered. Elucidation of the relationship between chemical structure and activity is a universal approach to understanding the mode of action of small molecules. A large number of artificial auxins and their chemical relatives were extensively studied
during the 1950s and 1960s. These studies identified several different types of core chemical structures that are required to elicit the auxin response in plants (reviewed in Thimann, 1977). At least two moieties of auxin, an aromatic ring and an anionic carboxylate at an appropriate distance from the aromatic ring, were thought to be important for mimicking auxin activity.
Biochemical approaches searching for the auxin binding fraction in plant extracts based on the auxin receptor hypothesis were undertaken in the 1970s. Several auxin binding proteins, including ABP1 (auxin-binding protein 1), were purified as a candidates of auxin receptors (reviewed in Venis & Napier, 1995). In the mid-1980s, the focus on identifying the components of auxin perception shifted to the molecular biology level, which resulted in identification of early auxin-induced genes, such as Aux/IAA (auxin/indole-3-acetic acid) and GH3, that are transcribed within a few minutes after auxin application (Theologis, 1986; Hagen & Guilfoyle, 1985). Over the decades, this molecular approach revealed auxin-responsive regulatory DNA sequences in the promoter regions of auxin responsive genes (reviewed in Abel & Theologis, 1996). These findings substantially advanced our current understanding of auxin action. Additionally, development of molecular markers for auxin response, such as DR5:GUS and BA:GUS, further enhanced our understanding of intracellular auxin perception mechanisms (Ulmasov et al., 1997; Oono et al., 1998).
In Arabidopsis, the first auxin resistant gene, AXR1 (auxin resistant 1), was identified using a positional cloning strategy (Leyser et al., 1993). Since then, identif ication of a number of auxin-related genes has helped greatly to construct a modern model of auxin perception. One of the best-characterized key components in the auxin signal transduction pathway is the SCFTIR1/AFBs ubiquitin E3 ligase, which regulates auxin-stimulated Aux/IAA protein degradation (reviewed in Leyser, 2002; Woodward & Bartel, 2005). Most of the Aux/IAA proteins share a highly conserved motif called domain II, which is required for the SCFTIR1/AFBs-dependent degradation of Aux/IAA proteins (Ramos et al., 2001; Dharmasiri et al., 2005a; Kepinski & Leyser, 2005; Tan et al., 2007). Aux/IAA proteins bind ARFs (auxin response factors), DNA-binding transcriptional factors that interact with the auxin-responsive element in the promoter regions of early auxin responsive genes, and suppress ARF activity. The ARFs act as transcriptional repressors or activators depending on their amino acid sequence in the middle region between N-terminal DNA binding and C-terminal Aux/IAA interacting regions (Tiwari et al., 2003). Thus, auxin dependent degradation of Aux/IAA proteins results in activation of ARFs and changes the pattern of downstream gene expression (reviewed in Rogg & Bartel, 2001).
In parallel, classical structure and activity analyses were expanded to chemical genetics and combined with studies of genetics and molecular markers. The screening for new small molecules having auxin activity or antagonistic activity from chemical libraries turned
Received: September 2, 2011; Accepted: December 12, 2011*To whom correspondence should be addressed:Tel: +81-(0)27-346-9537; Fax: +81-(0)27-346-9688E-mail: [email protected]
Special Issue: Auxin and Gravity Response in Plants
Biological Sciences in Space, Vol.25 No.2-4, 45-55, 2011

Auxin signaling and related small molecules
− 46 −
out to be a powerful tool to dissect auxin response mechanisms (reviewed in Blackwell & Zhao, 2003; Tóth & Hoorn, 2010). Further use of these new auxin-related molecules for genetic screening resulted in identification of novel signaling components in the auxin response pathway (Cheng et al., 2004; Rahman et al., 2006; Walsh et al., 2006; Biswas et al., 2007).
In this review, we provide overview of the recent progress in studies of auxin signaling components, following we focus on the role of small molecules for furthering our understanding of auxin signaling.
Auxin Receptors
To date, eight proteins have been recognized as auxin receptors in Arabidopsis. These are classified into three groups: 1) TIR1 (transport inhibitor response 1) and AFBs 1-5 (auxin-signaling F-box proteins) (Dharmasiri et al., 2005b; Parry et al., 2009), 2) SKP2A (S-phase kinase-associated protein 2A) (Jurado et al., 2010), and 3) ABP1 (auxin binding protein 1) (Woo et al., 2002). The TIR1 and
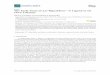
AFB group, all of which recognize Aux/IAA repressors as targets, is further separated into two subgroups based on the phylogenetic relationships and physiological activities observed in their mutants (Greenham et al., 2011). With the exception of ABP1, all seven proteins are F-box proteins, function in the nucleus, and are components of the SCF complex. ABP1 is functionally associated with plasma membrane and is structurally distinct from other auxin receptors (Figure 1).
Transport Inhibitor Response 1 (TIR1) and Auxin-Signaling F-Box Proteins (AFBs)
One of the most widely documented auxin receptor proteins, TIR1, was identified from a screen against an auxin transport inhibitor, CPD (2-carboxyphenyl-3-phenylpropane-1,2-dione), rather than against auxin itself (Ruegger et al., 1998). Mutations in TIR1 confer resistance to both 2,4-D (2,4-dichlorophenoxyacetic acid) and IAA (Ruegger et al., 1998). Genetic analysis revealed that TIR1 is an F-box protein that assembles
TIR1
ubiquitination
proteindegradation
gene expression
(1) SCFTIR1/AFBs
auxin response
Auxin
AFB4
?
gene expression
(2) SCFSKP2A
DPBAux/IAA Aux
/IAA
Nucleus
?
proteindegradation
ubiquitination
ROP
cytoskeleton
Plasmamembrane
clathrin-mediatedendocytosis
PIN1
clathrin
(3) ABP1
?
?
H+ATPase
cell wall
?
?
CUL1CUL1CUL1
SKP2AABP1
(a) (b)
Fig. 1. Auxin receptors and model of auxin signaling. To date, eight proteins, grouped into three different types of auxin receptors, have been recognized (objects with thick borders). (1) TIR1/AFBs are components of SCF-type E3 ubiquitin ligases. Auxin (indicated by red triangles) binds to TIR1 (or AFBs), promoting the binding of TIR1 (or AFBs) to Aux/IAA repressors. (a) Aux/IAA repressors that bind to TIR1 and AFB1-3 are then ubiquitinated and degraded. Degradation of the Aux/IAA repressors causes changes in downstream gene expression patterns and leads to physiological changes, the so-called auxin response (Dharmasiri et al., 2005b). (b) Mutant phenotypes of two AFB proteins, AFB4 and 5 have suggested that AFB4 and 5 are negative regulators of auxin signaling (Greenham et al., 2011). (2) SKP2A is also an F-box auxin binding protein and is a component of SCFSKP2A. The target proteins for SCFSKP2A ubiquitin E3 ligase are cell cycle transcription factors such as E2FC and DPB (Jurado et al., 2008). SKP2A itself is also degraded via ubiquitin-mediated proteolysis. The degradation of SKP2A is promoted by auxin binding to SKP2A, but it is not yet clear whether SKP2A is degraded through auto-ubiquitination or another E3-dependent process. (4) ABP1 is a membrane associated auxin receptor. Crystal structural analysis suggests that it forms a homodimer, although whether dimerization is necessary for its function is not known. Although many ABP1-dependent biological events have been reported, the biochemical activity of ABP1, its direct downstream signaling components, and cellular location where ABP1 acts are not clear (Sauer & Kleine-Vehn, 2011).

Biswas, K. K. and Oono, Y.
− 47 −
with SKP1 (S-phase kinase-associated protein 1, or ASK1 for Arabidopsis SKP1-like 1), CUL1 (cullin 1), and RING H2 finger protein, RBX1 (RING-box 1), to constitute a large ubiquitin E3 ligase complex, SCFTIR1 (Ruegger et al., 1998). The multi-subunit SCFTIR1 complex participates in ubiquitin-dependent proteolysis of many proteins. F-box proteins function as substrate-recognition subunits during the protein degradation events. Biochemical auxin binding studies indicated that auxin directly binds to TIR1 and stimulates the interaction of TIR1 with Aux/IAA proteins, which are subsequently ubiquitinated and degraded (Dharmasiri et al., 2005a; Kepinski & Leyser, 2005). Docking experiments with the crystal structure of TIR1 revealed that the carboxyl group of IAA interacts with an arginine residue at the bottom of the TIR1 pocket that harbors Aux/IAA proteins, and the indole ring of IAA stacks on top of the pocket floor (Tan et al., 2007). 1-NAA (1-naphthaleneacetic acid) and 2,4-D bind to TIR1 in a similar manner to IAA, despite having different ring structures (Tan et al., 2007). A binding assay suggested that natural auxin, IAA, has a 10-100 times higher affinity to the receptor than either 2,4-D or 1-NAA (Kepinski & Leyser, 2005).
Considering the absolute necessity of auxin for plant growth and development, one would expect severe morphological defects in auxin receptor mutants. Surprisingly, tir1 mutants showed only mild growth defects (Rugger et al., 1998). Further, the level of IAA binding in tir1 extracts was indistinguishable from the wild type (Dharmasiri et al., 2005a). This observation raised the possibility of the existence of other auxin receptors besides TIR1. Based on this idea, AFBs, homologous of TIR1, were examined and identified as additional auxin receptors (Dharmasiri et al., 2005b). Analyses of single and multiple mutants deficient in the TIR1, AFB1, AFB2 and AFB3 genes suggested that TIR1 and AFB genes act redundantly to control plant growth and development (Parry et al., 2009). Furthermore, saturable IAA binding was undetectable in the extract of the quadruple mutant, indicating that auxin binding is dependent on TIR1 as well as other AFB proteins (Dharmasiri et al., 2005a). Detailed analyses of various mutants further suggested that the AFBs contribute unequally to auxin response (Parry et al., 2009). For example, the tir1afb2 double mutant was more resistant to 2,4-D than tir1. In contrast, the responses of the tir1afb1, tir1afb3, and tir1afb1afb3 mutants were similar to that of tir1 seedlings. Thus, the loss of AFB1 and/or AFB3 does not affect the response to 2,4-D when the seedling already lacks TIR1. In addition, the responses of afb1afb2, afb1afb3, and afb2afb3 roots were similar to wild type, suggesting that TIR1 is sufficient to confer normal response to 2,4-D. Interestingly, AFB2 exhibits a higher affinity to several Aux/IAA proteins than AFB1 or AFB3 (Parry et al., 2009).
AFB4 and AFB5 are also TIR1-related proteins but belong to a phylogenetically distinct group (Parry et al., 2009); the AFB1, AFB2 and AFB3 proteins are closely related to TIR1 (61% to 72% amino acid identity), while the AFB4 and AFB5 appear to be 45% to 46% similar
with TIR1 at the amino acid level (Walsh et al., 2006). Both AFB4 and AFB5 proteins bind to ASK1, presumably to form an SCF complex, and interact with IAA3 protein in an auxin-dependent manner (Greenham et al., 2011). However, distinct from TIR1/AFB1-3, the loss of AFB4 resulted in an elongated hypocotyl and petiole and an increased number of lateral roots (Greenham et al., 2011). Furthermore, the afb4-2 mutant showed a hypersensitive response to IAA-mediated hypocotyl elongation and early auxin gene expression. These strikingly contrasting phenotypes and responses led the authors to categorize AFB4 and AFB5 as negat ive regulators of auxin responses, although the precise mechanism of how these proteins affect SCF E3 activity as well as the stability of Aux/IAA protein remains elusive (Greenham et al., 2011).
S-Phase Kinase-Associated Protein 2A (SKP2A)
SKP2A is an F-box protein that forms an SCF complex and regulates the proteolysis of cell cycle transcription factors such as E2FC (E2 promoter transcription factor C) and DPB (E2F dimerization partner B) (Jurado et al., 2008). SKP2A itself is posttranslationally regulated by ubiquitin-dependent degradation, which is promoted by auxin (Jurado et al., 2008). Cell-free experiments suggested that auxin directly and specifically binds to SKP2A and promotes SKP2A degradation in a TIR1-independent manner (Jurado et al., 2010). Mutations in predicted auxin binding sites reduced the ability of SKP2A to bind to auxin and generated stable SKP2A. These non-auxin-binding SKP2A variants were unable to promote E2FC/DPB degradation in vivo and conferred resistance to auxin in pr imary root growth. Moreover, auxin enhanced the interaction between SKP2A and DPB. Hence, SKP2A is an auxin receptor that predominantly functions in auxin-regulated cell division (Jurado et al., 2010). Dominant negative mutants in the SCFTIR1 pathway, such as axr2 and axr3, showed reduced levels of SKP2A protein (Jurado et al., 2008). The DR5:GUS expression pattern was affected by the level of SKP2A gene expression (Jurado et al., 2008). These results indicate the possible presence of crosstalk between the SCFTIR1/AFBs and SCFSKP2A pathways.
Auxin Binding Protein 1 (ABP1)
One of the popular methods of identifying receptor complexes is through affinity purification. This technique has long been used to identify auxin binding proteins and has resulted in the purification of a number of auxin binding proteins from both membrane and nuclear fractions (reviewed in Venis & Napier, 1995). The best-characterized of these auxin binding proteins is ABP1, which was discovered almost 40 years ago (Hertel et al., 1972). ABP1 was purified as a soluble protein from a maize coleoptile membrane fraction and consisted of 201 amino acid residues, including an N-terminal hydrophobic signal sequence for translocation to the endoplasmic reticulum and a C-terminal KDEL amino acid sequence

Auxin signaling and related small molecules
− 48 −
for endoplasmic reticulum retention (Hesse et al., 1989; Tillmann et al., 1989). A binding assay using radiolabeled auxin suggested that ABP1 effectively binds to 1-NAA rather than IAA and shows low affinity to 2,4-D (Löbler & Klämbt, 1985).
The crystal structure analysis of ABP1 suggested that ABP1 is a dimer and that auxin binds within a predominantly hydrophobic pocket with a zinc ion coordinated with three histidines and a glutamate in each subunit (Woo et al., 2002). The zinc ion is important for interaction with the carboxyl group of auxin, and no conformational rearrangement of ABP1 was observed when auxin was bound to the protein in the crystal (Woo et al., 2002). Numerous functional descriptions of ABP1 have suggested that it is a bona fide auxin receptor that triggers early modification of ion fluxes across the plasma membrane by activating H+-ATPase, mediating auxin-dependent cell expansion and cell division, and coordinating early embryonic development (Tromas et al., 2010; David et al., 2007; Chen et al., 2006; Braun et al., 2008). The loss of ABP1 leads to an embryonic lethal phenotype in Arabidopsis (Chen et al., 2001). Recently, it has been shown that activation of the ROP (Rho-of-plant)-GTPases by auxin during morphogenesis of pavement cells depends on ABP1 (Xu et al., 2010). These ROP-GTPases are localized in the plasma membrane, and ABP1/ROP dependent pathways regulate actin/microtubule organization (Xu et al., 2010). It has also been reported that ABP1 promotes clathrin recruitment to the plasma membrane and clathrin-dependent endocytosis in the absence of auxin, whereas auxin binding to ABP1 inhibits such processes (Robert et al., 2010). The regulation of clathrin-mediated endocytosis by ABP1 subsequently regulates PIN1 (pin-formed 1) localization in the plasma membrane (Robert et al., 2010). Collectively, these results indicate that ABP1 possibly acts as an auxin receptor involving early responses in the plasma membrane. However, the exact molecular function, the direct targets of ABP1 and cellular location where ABP1 acts are still unclear (Sauer & Kleine-Vehn, 2011).
Identification and Characterization of Chemical Modulators of Auxin Signaling
A large number of small molecules that show auxinic or anti-auxinic activity have been studied in order to understand auxin’s mode of action. Early research monitored auxin activity with induced physiological responses, such as the split pea stem test (reviewed in Thimann, 1977). However, it is difficult to identify the targets of such compounds in the auxin signaling pathway from physiological techniques alone. The development of specific molecular markers for auxin signaling such as BA:GUS, DR5:GUS and HS:AXR3NT-GUS enabled researchers to examine the effects of small molecules by focusing on early steps in auxin signaling. Both BA and DR5 promoters are derived from the promoters of auxin-responsive genes whose gene expression is enhanced by auxin through auxin-responsive cis elements, binding
sites of ARF transcription factors (Ulmasov et al., 1997; Oono et al., 1998). AXR3NT-GUS is a GUS fusion protein with the N-terminal half of the AXR3 (IAA17) protein that is degraded by auxin through SCFTIR1/AFBs ligase (Gray et al., 2001). Thus, these markers are tightly linked to the TIR1/AFBs-dependent auxin-signaling pathway.
These markers are useful to quickly distinguish chemicals act ing in auxin percept ion f rom other compounds such as auxin transport inhibitors (Oono et al., 2003). For example, PCIB (p-chlorophenoxyisobutyric acid) has been known for a long time to be an anti-auxin (MacRae & Bonner, 1953). Because the mechanism of action of PCIB at the molecular level was unknown until recently, the effect of PCIB has been attributed to the inhibition of auxin transport (Katekar & Geissler, 1980; Tsai & Arteca, 1984), the regulation of auxin metabolism (Frenkel & Haard, 1973), or to a nonspecific toxic effect (Yip & Yang, 1986). The use of transgenic Arabidopsis containing the auxin responsive marker BA:GUS together with membrane-permeable auxin NAA clearly showed that PCIB impairs auxin perception in Arabidopsis root, whereas other auxin-related compounds such as NPA(1-naphthylphthalamic acid), NOA(1-naphthoxyacetic acid), maleic hydrazide, or trans-cinnamic acid do not (Oono et al., 2003; Figure 2).
A
B
NAA(1)
PCIB(20) PCIB(50) PCIB(100)
NAA(1)+
NOA(20) NOA(50) NOA(100)
NPA(20) NPA(50) NPA(100)
Cont.
2,4-D PCIB
Fig. 2. PCIB impairs auxin perception.A. The auxin inhibitory compound PCIB (right) is structurally similar to the synthetic auxin 2,4-D (left). B. PCIB effectively inhibits NAA-induced BA:GUS expression in Arabidopsis root tips, whereas auxin influx (NOA) or efflux (NPA) inhibitors do not. The concentrations (µM) of the compounds are indicated in parentheses.Panal B has been modified from Oono et al., 2003 (Copyright American Society of Plant Biologists, www.plantphysiol.org).

Biswas, K. K. and Oono, Y.
− 49 −
The use of markers also provides a rapid and robust assay system for quick screening of a large number of small molecules that inhibit the auxin-signaling pathway with a chemical genetics approach. Armstrong et al. (2004) developed a high-throughput whole-seedling assay by using the BA:GUS reporter and identified four compounds (compounds A-D) as inhibitors of auxin signaling in Arabidopsis roots. The four compounds also reduced auxin-induced DR5:GUS expression levels. In an HS:AXR3NT-GUS assay, compounds A-C, but not D, showed inhibitory effects (Armstrong et al., 2004). Subsequent studies by Sungur et al. (2007) reported that 2-FAA (2-furylacrylic acid), the active portion of compound A, completely blocked IAA-induced BA:GUS expression in the distal cell elongation zone of Arabidopsis roots. Furthermore, 2-FAA inhibited root growth and strongly attenuated the auxin induction of IAA5 expression. The root growth inhibition by 2,4-D or NAA was restored with media containing 2-FAA, suggesting that 2-FAA does not block the uptake or efflux of auxin into the cell, but rather alters some aspect of the signaling machinery. However, 2-FAA failed to alter the interaction of IAA7/AXR2 with the TIR1 complex, indicating that TIR1 is not a direct target of 2-FAA.
Several inhibitors for auxin signaling were also identified from microbial extracts using the reporter lines described above. YkB (Yokonolide B), a spiroketal-macrolide, was identified as an inhibitor of BA:GUS expression in Arabidopsis (Hayashi et al., 2003). YkB had no effect on NAA efflux or on 2,4-D influx in BY2 cells, confirming that YkB is an auxin action inhibitor. YkB inhibits degradation of an Aux/IAA repressor and blocks auxin-dependent cell division and epinastic growth mediated by ABP1, suggesting that YkB seems to have negative effects on both the SCFTIR1/AFBs and ABP1 auxin signaling pathways. TrfA (Terfestatin A) and Toyocamycin were identified by a similar screening using the reporter constructs (Yamazoe et al., 2005; Hayashi et al., 2009). TrfA shares structural similarities with auxins. The positions of the aromatic ring and the carboxyl group in auxins such as IAA and 1-NAA closely match the position of the aromatic A ring and the phenolic hydroxyl group in TrfA, respectively (Hayashi et al., 2008b). However, TrfA did not inhibit the auxin-dependent interaction between TIR1 and Aux/IAA proteins, and no direct evidence of binding between TrfA and TIR1 has been found (Yamazoe et al., 2005). TrfA also inhibits SKP2A degradation without affecting IAA binding to SKP2A (Jurado et al., 2010).
The auxin signaling markers are also useful for identification of novel compounds that show auxin activity. DR5:GUS-based screening identified sirtinol as a compounds that induced many auxin-inducible genes (Zhao et al., 2003). Sirtinol also led to a rapid degradation of the AXR3NT-GUS fusion protein. Auxin signaling mutants (axr2, tir1, and nph4) were less sensitive to sirtinol than wild type. In contrast, auxin-transport mutants (aux1, pin2, and tir3) showed a wild-type response, suggesting that sirtinol bypasses auxin transport. Further characterization of Arabidopsis sir (sirtinol resistant) mutants revealed that sirtinol is
converted to active auxin HNC (2-hydroxy-1-naphthoic acid), which is structurally similar to NAA (Dai et al., 2005). Interestingly, HNC itself did not show auxin activity when Arabidopsis seedlings were grown on medium with HNC. HNC is not effectively transported to cells; rather it becomes active when it is generated from sirtinol in plant cells. The markers can also be used to characterize the compounds obtained in primary screening for auxin-like activity in physiological response. WH7 [2-(4-chloro-2-methylphenoxy)-N-(4-H-1,2,4-triazol-3-yl)acetamide] shares structural similarity with 2,4-D and was identified by screening a chemical library for inhibition activity of Arabidopsis root growth. Experiments with DR5:GUS and DR5rev:GFP confirmed the involvement of WH7 in auxin-signaling (Christian et al., 2008).
Endogenous metabolites in plants may function as auxin inhibitors in addition to exogenous chemicals. Recently, Staswick (2009) reported that JA-Trp (jasmonic acid-tryptophan conjugates) and IAA-Trp (IAA-tryptophan conjugates) inhibited IAA-induced DR5:GUS expression as well as auxin-stimulated physiological responses of Arabidopsis. Both JA-Trp and IAA-Trp are synthesized in Arabidopsis. Comparison of the effects of the Trp conjugates with auxin transport inhibitor NPA on lateral root development and sensitivity testing of the conjugate in the auxin transport mutant aux1-7 indicated that the conjugates are not likely to act on auxin transport. However, neither JA-Trp nor IAA-Trp interfered with IAA-dependent interaction between TIR1 and AuX/IAA7 in cell-free assays. Interestingly, IAA and JA share core subunits of SCF E3 ligase for their signaling (Schwechheimer et al., 2002), and both are conjugated with amino acids by similar enzymes belonging to the GH3 family (Staswick et al., 2005). Furthermore, both JA and IAA conjugates act as auxin inhibitors, indicating the presence of a tight functional link between the two plant hormones.
The availability of structural information about auxin receptors enables us to estimate the mode of action of small compounds more precisely. Hayashi et al. (2008a) systematically designed a series of small agonists and antagonists of TIR1 by introducing different alkyl chains to the α position of IAA. The IAAs with methyl to propyl chain substitutions in the α position retained auxin activity, whereas the IAAs with butyl or longer chain substitutions showed anti-auxinic activity. Similar activity changes were also observed in two different types of α-alkyl auxins, NAA and 2,4-D. Several lines of experiment, as well as analysis of crystal structure and molecular docking calculations, revealed that all of these α-alkyl IAAs could bind to TIR1, but the interaction between TIR1 and Aux/IAA proteins was prevented when the α position was substituted with butyl or longer chains. In contrast, molecular modeling analyses suggested that IAAs with bulky alkyl chains in the α position could not interact with another auxin receptor, ABP1. Thus, these bulky auxin inhibitors are considered to be specific inhibitors for TIR1 and AFB receptors. In this context, BH-IAA (α-tert-butoxycarbonylaminohexyl-indole-3-acetic acid) and PEO-IAA [α-(phenylethyl-2-oxo)-indoleacetic acid] were recently used to elucidate the significance of

Auxin signaling and related small molecules
− 50 −
the TIR1/AFBs-dependent auxin signaling pathway in several biological processes such as gravitropic curvature (Nishimura et al., 2009), transition of the cell cycle (Ishida et al., 2010), stress/ACC response (Tsang et al., 2011), and shade avoidance control (Keuskamp et al., 2010). Most recently, a more potent TIR1 antagonist, auxinole [α-(2,4-dimethylphenylethyl-2-oxo)-indoleacetic acid], was reported (Hayashi et al., 2012). The auxin signaling inhibitors are listed in Table 1.
Auxin-related Compounds, an Approach to Identify Novel Auxin Signaling Components
The genetic approach has been based mainly on screening mutants with altered responses to auxin. However, given the absolute necessity of auxin for embryogenesis, the eff icacy of screening mutant seedlings against auxin may be limited. An alternative approach is to screen mutants using compounds that modify or antagonize auxin responsiveness. The use of auxin-related small molecules would be suitable for this approach. The first published report using such a strategy demonstrated isolation of mutants against auxin transport inhibitor, CPD, followed by the identification of the TIR1 auxin receptor (Ruegger et al., 1998; Dharmasiri et al., 2005a; Kepinski & Leyser, 2005). Screening of mutants against sirtinol also revealed an uncharacterized mutant deficient in the CAND1 (cullin-associated and neddylation dissociated) gene that is involved in the regulation of SCF ubiquitin E3 ligase activity (Zhao et al., 2003; Cheng et al., 2004).
A series of mutants were obtained in our lab from a forward genetic screening against the auxin action inhibitor, PCIB. The screening assay was based on primary root growth elongation on media containing PCIB. Seedlings with longer roots were recovered as mutants and were named aar (anti-auxin resistant). In addition to PCIB resistance, the aar mutants show resistance to the synthetic auxin 2,4-D in root elongation
and lateral root induction assays (Rahman et al., 2006, Biswas et al., 2007, Nakasone et al., 2009). Among these aar mutants, aar1 and aar3 were defective for previously uncharacterized genes. The causal gene responsible for the aar1 phenotype encodes SMAP1 (small acidic protein) that functions upstream of the Aux/IAA-protein degradation step in auxin signaling (Rahman et al., 2006). SMAP1 contains phenylalanine and aspartic acid-rich domain (F/D-rich domain) that consists of 18 amino acids at the C-terminus. The putative genes encoding SMAP1-like protein with F/D-rich domain are conserved in divergent animal and plant species, implying that the protein plays an important role that is common in many species even though its function is unknown (Rahman et al., 2006; Nakasone et al., 2009). The AAR3 gene encodes a nuclear protein that had not previously been implicated in auxin signaling and that shares homology with the DCN-1 (defective in cullin neddylation-1) protein identified in yeast and mammals through the conserved PONY domain. The DCN-1 protein functions as an E3 ligase for NEDD8 modification (neddylation) of CUL (cullin) proteins and directly binds to CULs through the PONY domain (Kurtz et al., 2008). NEDD8, or RUB (related ubiquitin) in plant and yeast cells, is a small ubiquitin-like protein that covalently binds to CULs. Its association with and dissociation from CULs are significant for the activity of CUL-based ubiquitin E3 ligase (Dreher & Callis, 2007). Therefore, AAR3 may function in NEDD8/RUB modification of CULs and may affect SCF activity as well as auxin signaling, although the detailed molecular mechanism is not yet known. The same screening approach also identified another three mutations that are located in two known auxin-related loci, TIR1 and CUL1 (Biswas et al., 2007). Thus, the approach we have taken to isolate mutants against PCIB indeed facilitated the identification of new factors involved in auxin or auxin-related signaling (Figure 3).
Table 1. List of auxin signaling inhibitors
Name of compound Auxin signaling processes inhibited by the compound Source Reference
PCIB(p-chlorophenoxyisobutyric acid) 1, 2 synthetic compound Oono et al., 2003TFIBA(4,4,4,-trifluoro-3-indole-3-butyric acid) 2 synthetic compound Oono et al., 2003YkB(yokonolide B) 1, 2, ABP1-mediated cell expansion microbial extracts Hayashi et al., 2003compound A-C 1, 2 synthetic compound Armstrong et al., 20042-FAA(2-furylacrylic acid) 1, 2 synthetic compound Sungur et al., 2007
TrfA(terfestatin A) 1, 2, SKP2A degradation microbial extracts Yamazoe et al., 2005 Jurado et al., 2010
toyocamycin 1, 2, microbial extracts Hayashi et al., 2009JA-Trp / IAA-Trp 2 endogenous metabolites Staswick, 2009BH-IAA PEO-IAA auxinole
1,2, interaction between TIR1 and Aux/IAA proteins synthetic compound Hayashi et al., 2012
1. Inhibiting auxin-induced degradation of AuX/IAA protein or AXR3NT-GUS marker.2. Inhibiting auxin-induced gene expression including BA:GUS, DR5:GUS, or DR5rev:GFP markers.

Biswas, K. K. and Oono, Y.
− 51 −
In a similar screening approach, the picolinate class of synthetic auxins was used (Walsh et al., 2006). This screen identified seven alleles at two distinct genetic loci that conferred significant resistance to picolinate auxins such as picloram (4-amino-3,4,5-trichloropicolinic acid) yet had minimal cross-resistance to 2,4-D or IAA. The mutations are identified on AFB5 and SGT1b, both of which code proteins associated with SCF-mediated ubiquitination. Consistently, picloram stimulated elongation of wild-type hypocotyls, while the afb5-5 mutant showed resistance to this stimulation (Greenham et al., 2011). AFB5, as well as AFB4, interacted with the IAA3 protein in a picloram-dependent manner, whereas TIR1 did not, suggesting that AFB5 and AFB4 are the major targets of the picolinate herbicides in Arabidopsis and that a significant proportion of plant responses to picolinate auxins is mediated via the SCFAFB4/AFB5 ubiquitin ligase (Greenham et al., 2011). Considering the evidence that AFB4 and (probably) AFB5 are negative regulators of auxin signaling, some of the auxinic activity of picloram may be due to inhibiting AFB4/AFB5 by its binding to them. The afb5 mutant is also resistant to another auxin-like herbicide, dicamba (Gleason et al., 2011). Interestingly, the tir1-1 mutant also showed dicamba resistance, but other F-box mutants (afb1, afb2, and afb3) did not. The interaction between dicamba and F-box auxin receptors is not yet understood.
Role of Auxin Signaling Components in Gravity Response
Plant gravity response critically depends on auxin. The Cholodny-Went theory proposes that auxin plays a key role in several tropic responses (reviewed in Gilroy & Masson, 2008). Recent molecular genetic studies have strongly supported this hypothesis for gravitropism. Gravitropic curvature is primarily driven by an asymmetric distribution of auxin that induces differential growth and bending (reviewed in Muday & Rahman, 2008). Auxin signaling occurs downstream of auxin transport. Hence, auxin signaling is expected to also play an important role in regulating root gravity response. Indeed most auxin signaling inhibitors, such as PCIB (Oono et al., 2003), TrfA (Yamazoe et al., 2005) and JA-Trp (Staswick, 2009), are reported to inhibit gravitropic responses. In cucumber, seedlings form a specialized protuberance, the peg, on the transition zone between the hypocotyl and the root. Gravistimulation has been suggested to affect auxin accumulation and subsequent peg formation on the lower side of the transition zone (Takahashi et al., 2000). Application of PCIB to cucumber seedlings inhibited peg formation, decreased auxin-induced CsIAA1 mRNAs, and increased auxin-repressible CsGRP1 mRNA on the lower side of the transition zone (Shimizu et al., 2008). These results confirm the significant role of endogenous auxin in peg formation at the gravistimulated transition zone of cucumber seedlings.
The availability of mutants and inhibitors for specific auxin signaling pathways enables us to evaluate the significance of particular pathways in auxin-mediated gravity response. In Arabidopsis, multiple mutations of TIR1/AFBs receptors showed reduced gravity responses in roots (Dharmasiri et al., 2005b), suggesting that these receptors are redundantly involved in gravity responses. The involvement of the SCFTIR1/AFBs pathway in gravity responses was also demonstrated in maize coleoptiles by using an auxin signaling inhibitor (Nishimura et al., 2009). Horizontally tilted maize coleoptile segments bend upward upon gravistimulation. Corresponding to asymmetric IAA distribution, an auxin-responsive ZmSAUR2 gene is up-regulated on the lower side and down-regulated on the upper side in the coleoptile elongation zone. PEO-IAA, an SCFTIR1/AFBs inhibitor, reduced the expression level of the ZmSAUR2 gene as well as the bending of coleoptiles. The results indicate that the TIR1/AFBs-mediated auxin response is significant in this process (Nishimura et al., 2009)
Although ABP1 is known to be involved in auxin-mediated cell expansion and elongation (Jones et al., 1998; Chen et al., 2001), there had been no convincing report describing the specific role of ABP1 in gravity response until recently. In a recent analysis (Effendi et al., 2011), heterozygous abp1/ABP1 in Arabidopsis showed an agravitropic phenotype, specifically in shoot and roots. This result indicated that ABP1 might be involved in gravity responses. Thus, several distinct auxin signaling components, including TIR1/AFBs and ABP1, seem to act in controlling gravity responses in plants.
TIR1 AUX/IAA
CUL1
AXR1SCFTIR1
SMAP1
AAR3
ubiquitin
Nedd8/RUB
Auxin
degradation
Fig. 3. Factors that confer PCIB resistance of the aar mutants (indicated by red outline and bold letters).In addition to TIR1 and CUL1, SMAP1 and AAR3 were identified by the positional cloning of the aar1 and aar3 mutations, respectively. AAR3 encodes a DCN1-like protein and putatively functions as E3 ligase for the Nedd8/RUB protein (Biswas et al., 2007). The Nedd8/RUB modification of CUL1 is important for the regulation of SCF E3 activity. AXR1 is a component of an E1 activating enzyme for the Nedd8/RUB modification. The axr1 mutant is also resistant to PCIB (Oono et al., 2003). Although the biochemical function of SMAP1 is unknown, genetic analyses suggested that SMAP1 acts upstream of the Aux/IAA-protein degradation step in auxin signaling.

Auxin signaling and related small molecules
− 52 −
PIN proteins, which are auxin efflux transporters, generate directional auxin flow in plant cells by localizing at one side of the plasma membrane and contribute to the asymmetric distribution of auxin. PIN2 and PIN3, especially, are significantly involved in gravitropic responses (Chen et al., 1998; Lushing et al., 1998; Muller et al., 1998; Friml et al., 2002). For example, in gravistimulated Arabidopsis roots, PIN3 that is uniformly distributed in the plasma membrane of columella cells relocates to the lower side of those cells. PIN2, which localizes at the upper side of lateral root cap and epidermal cells to direct the auxin flow from the root tip into the elongation zone, is also redistributed upon gravistimulation to become more abundantly localized on the lower side of gravistimulated roots than the upper side (Abas et al., 2006). This re-distribution is attributed to differential intracellular localization and degradation of PIN2 in the upper and lower sides of the roots. The intracellular localization of PIN proteins in the plasma membrane is controlled by membrane trafficking, including endocytosis and vesicle recycling, which are also affected by auxin (Paciorek et al., 2005). In Arabidopsis root epidermis cells, PIN2 localization into the plasma membrane is inhibited by the recycling inhibitor brefeldin A (BFA), and PIN2 protein is accumulated in intracellular compartments termed BFA bodies. Interestingly auxin interferes with this BFA-induced PIN2 internalization by inhibiting endocytosis of PIN2 (Paciorek et al., 2005). Furthermore, in double or triple mutants of the TIR1/AFBs family of auxin receptors, the inhibitory effect of auxin on PIN2 internalization is strongly reduced, suggesting that the SCFTIR1/AFBs pathway is required for auxin-dependent inhibition of PIN2 endocytosis (Pan et al., 2009). In addition, experiments by other groups have suggested that auxin inhibits the clathrin-dependent endocytosis that regulates internalization of PIN proteins through ABP1-mediated process in Arabidopsis roots (Robert et al., 2010). Slower basipetal auxin transport was observed in young roots of heterozygous abp1/ABP1 Arabidopsis (Effendi et al., 2011). These lines of evidence indicate that both TIR/AFBs- and ABP1-mediated auxin signaling pathways are involved in regulating the intracellular local izat ion of PIN proteins and the consequent establishment of an asymmetric auxin distribution. Taken together, distinct auxin signaling pathways play roles in plant gravitropic responses at multiple levels, including the establishment of auxin gradients by affecting auxin transport and the control of cell elongation via auxin perception.
Future Prospects
To date, TIR1, AFBs1-5, SKP2 and ABP1 have been recognized as auxin receptors. Development of specific inhibitors and marker genes like DR5:GUS and BA:GUS, as well as the establishment of mutants, have helped immensely to improve our current understanding of the physiological role of the SCFTIR1/AFBs pathway. However, the modes of action of the auxin receptors ABP1 and SKP2 still remain largely unknown. Further efforts to
develop specific molecular markers for the pathway and to screen specific inhibitors or agonists are necessary to dissect the role of distinct auxin pathways in regulating the intracellular auxin response. In this context, recent reports have demonstrated the usefulness of auxin analogues. PEO-IAA inhibits auxin-inducible gene expression related to the TIR1-dependent signaling process but shows an agonistic effect on BFA-induced PIN internalization that is inhibited by auxin. In contrast, another auxin analog, 5-fluoroindole-3-acetic acid activated DR5rev:GFP but failed to inhibit PIN internalization (Robert et al., 2011), suggesting distinct auxin response pathways would be functionally distinguishable by using small molecules.
The significance of auxin to the root gravity response is well-known, but how auxin signaling machineries are involved in this gravity response has not yet been fully elucidated. Recent reports that characterize mutants defective in auxin receptors indicate that both the TIR1/AFBs and ABP1 pathways are involved, but the two pathways probably act differently on the intracellular localization of PIN proteins that control the auxin gradient during gravity responses (Pan et al., 2009; Roberts et al., 2010). However, only a few reports describe the involvement of particular auxin signaling machineries during gravity response. More detailed studies must be done on how and in which step each auxin-signaling pathway contributes to gravity responses.
There are reports that the expression pattern of Aux/IAA genes that include the DR5 promoter, which is downstream of TIR1/AFBs, is affected by the SKP2 or ABP1 pathways (Jurado et al., 2008; Braun et al., 2008; Tromas et al., 2009; Effendi et al., 2011). Furthermore, expression of the ABP1 transcript is regulated by auxin itself (Effendi et al., 2011). This evidence collectively indicates the presence of crosstalk among distinct auxin signaling pathways. Thus, understanding the auxin-induced physiological responses will require making a clear map including more than one auxin signaling pathway. Continuous efforts to find small auxin-related compounds that specifically interact with a single auxin signaling pathway will be necessary to obtain powerful tools to elucidate the issues mentioned above. Use of such specific auxin modulators combined with specific molecular markers, genetic mutants, and biochemical/biophysical analyses of receptors will allow a more precise dissection of auxin signaling and a more accurate understanding of the mode of auxin action as well as the mechanism of auxin-mediated plant growth/development.
Acknowledgments
We are very grateful to Drs. Ken-ichiro Hayashi and Abidur Rahman for reading the manuscript and providing useful comments. This work was supported in part by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan (Kakenhi; no. 22570056) to Y.O.

Biswas, K. K. and Oono, Y.
− 53 −
References
Abas, L., Benjamins, R., Malenica, N., Paciorek, T., Wišniewska, J., Moulinier-Anzola, J. C., Sieberer, T., Friml, J., Luschnig, C. (2006) Intracellular trafficking and proteolysis of the Arabidopsis auxin-eff lux facilitator PIN2 are involved in root gravitropism. Nat. Cell Biol., 8, 249-256.
Abel, S., Theologis, A. (1996) Early genes and auxin action. Plant Physiol., 111, 9-17.
Armstrong, J. I., Yuan, S., Dale, J. M., Tanner, V. N., Theologis, A. (2004) Identification of inhibitors of auxin transcriptional activation by means of chemical genetics in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A., 101, 14978-14983.
Biswas, K. K., Ooura, C., Higuchi, K., Miyazaki, Y., Van Nguyen, V., Rahman, A., Uchimiya, H., Kiyosue, T., Koshiba, T., Tanaka, A., Narumi, I., Oono, Y. (2007) Genetic characterization of mutants resistant to the antiauxin p-chlorophenoxyisobutyric acid reveals that AAR3, a gene encoding a DCN1-like protein, regulates responses to the synthetic auxin 2,4-dichlorophenoxyacetic acid in Arabidopsis roots. Plant Physiol., 145, 773-785.
Blackwell, H. E., Zhao, Y. (2003) Chemical genetic approaches to plant biology. Plant Physiol., 133, 448-455.
Braun, N., Wyrzykowska, J., Muller, P., David, K., Couch, D., Perrot-Rechenmann, C., Fleming, A. J. (2008) Conditional repression of AUXIN BINDING PROTEIN1 reveals that it coordinates cell division and cell expansion during postembryonic shoot development in Arabidopsis and Tobacco. Plant Cell, 20, 2746-2762.
Chen, J.-G., Ullah, H., Young, J. C., Sussman, M. R., Jones, A. M. (2001) ABP1 is required for organized cell elongation and division in Arabidopsis. Genes Dev., 15, 902-911.
Chen, J.-G., Wang, S., Lazarus, C. M., Napier, R. M., Jones, A. M. (2006) Altered expression of auxin-binding protein 1 affects cell expansion and auxin pool size in tobacco cells. J. Plant Growth Regul., 25, 69-78.
Chen, R., Hilson, P., Sedbrook, J., Rosen, E., Caspar, T., Masson, P. H. (1998) The Arabidopsis thaliana AGRAVITROPIC 1 gene encodes a component of the polar-auxin-transport efflux carrier. Proc. Natl. Acad. Sci. U.S.A., 95, 15112-15117.
Cheng, Y., Dai, X., Zhao, Y. (2004) AtCAND1, a HEAT-repeat protein that participates in auxin signaling in Arabidopsis. Plant Physiol., 135, 1020-1026.
Christian, M., Hannah, W. B., Lüthen, H., Jones, A. M. (2008) Identification of auxins by a chemical genomics approach. J. Exp. Bot., 59, 2757-2767.
Dai, X., Hayashi, K.-i., Nozaki, H., Cheng, Y., Zhao, Y. (2005) Genetic and chemical analyses of the action mechanisms of sirtinol in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A., 102, 3129-3134.
Dav id , K. M. , Couch, D. , Braun, N. , Brown, S. ,
Grosclaude, J., Perrot-Rechenmann, C. (2007) The auxin-binding protein 1 is essential for the control of cell cycle. Plant J., 50, 197-206.
Dharmasiri, N., Dharmasiri, S., Estelle, M. (2005a) The F-box protein TIR1 is an auxin receptor. Nature, 435, 441-445.
Dharmasiri, N., Dharmasiri, S., Weijers, D., Lechner, E., Yamada, M., Hobbie, L., Ehrismann, J. S., Jurgens, G., Estelle, M. (2005b) Plant development is regulated by a family of auxin receptor F Box proteins. Dev. Cell, 9, 109-119.
Dreher, K., Callis, J. (2007) Ubiquitin, hormones and biotic stress in plants. Annals Bot., 99, 787-822
Effendi, Y., Rietz, S., Fischer, U., Scherer, G. F. E. (2011) The heterozygous abp1/ABP1 insertional mutant has defects in functions requiring polar auxin transport and in regulation of early auxin-regulated genes. Plant J., 65, 282-294.
Frenkel, C., Haard, N. F. (1973) Initiation of ripening in Bartlett pear with an antiauxin α(p-chlorophenoxy)isobutyric acid. Plant Physiol., 52, 380-384.
Friml, J., Wisniewska, J., Benkova, E., Mendgen, K., Palme, K. (2002) Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature, 415, 806-809.
Gilroy, S., Masson, P. H., eds (2008) Plant Tropisms. Blackwell Publishing, Oxford, UK.
Gleason, C., Foley, R. C., Singh, K. B. (2011) Mutant analysis in Arabidopsis provides insight into the molecular mode of action of the auxinic herbicide dicamba. PLoS ONE, 6, e17245.
Gray, W. M., Kepinski, S., Rouse, D., Leyser, O., Estelle, M. (2001) Auxin regulates SCFTIR1-dependent degradation of AUX/IAA proteins. Nature, 414, 271-276.
Greenham, K., Santner, A., Castillejo, C., Mooney, S., Sairanen, I., Ljung, K., Estelle, M. (2011) The AFB4 auxin receptor is a negative regulator of auxin signaling in seedlings. Curr. Biol., 21, 520-525.
Hagen, G. and Guilfoyle, T. J. (1985) Rapid induction of selective transcription by auxins. Mol. Cell. Biol., 5, 1197-1203.
Hayashi, K.-i., Jones, A. M., Ogino, K., Yamazoe, A., Oono, Y., Inoguchi, M., Kondo, H., Nozaki, H. (2003) Yokonolide B, a novel inhibitor of auxin action, blocks degradation of AUX/IAA factors. J. Biol. Chem., 278, 23797-23806.
Hayashi, K.-i., Kamio, S., Oono, Y., Townsend, L. B., Nozaki, H. (2009) Toyocamycin specifically inhibits auxin s ignal ing mediated by SCFTIR1 pathway. Phytochemistry, 70, 190-197.
Hayashi, K.-i., Neve, J., Hirose, M., Kuboki, A., Shimada, Y., Kepinski, S., Nozaki, H. (2012) Rational Design of an Auxin Antagonist of the SCFTIR1 Auxin Receptor Complex. ACS Chem. Biol., 7, 590-598.
Hayashi, K.-i., Tan, X., Zheng, N., Hatate, T., Kimura, Y., Kepinski, S., Nozaki, H. (2008a) Small-molecule

Auxin signaling and related small molecules
− 54 −
agonists and antagonists of F-box protein–substrate interactions in auxin perception and signaling. Proc. Natl. Acad. Sci. U.S.A., 105, 5632-5637.
Hayashi, K.-i., Yamazoe, A., Ishibashi, Y., Kusaka, N., Oono, Y., Nozaki, H. (2008b) Active core structure of terfestatin A, a new specific inhibitor of auxin signaling. Bioorg. Med. Chem., 16, 5331-5344.
Hertel, R., Thomson, K.-S., Russo, V. E. A. (1972) In-vitro auxin binding to particulate cell fractions from corn coleoptiles. Planta, 107, 325-340.
Hesse, T., Feldwisch, J., Balshüsemann, D., Bauw, G., Puype, M., Vandekerckhove, J., Löbler, M., Klämbt, D., Schell, J., Palme, K. (1989) Molecular cloning and structural analysis of a gene from Zea mays (L.) coding for a putative receptor for the plant hormone auxin. EMBO J., 8, 2453-2461.
Ishida, T., Adachi, S., Yoshimura, M., Shimizu, K., Umeda, M., Sugimoto, K. (2010) Auxin modulates the transition from the mitotic cycle to the endocycle in Arabidopsis. Development, 137, 63-71.
Jones, A. M., Im, K.-H., Savka, M. A., Wu, M.-J., DeWitt, N. G., Shillito, R., Binns, A. N. (1998) Auxin-dependent cell expansion mediated by overexpressed auxin-binding protein 1. Science, 282, 1114-1117.
Jurado, S., Abraham, Z., Manzano, C., López-Torrejón, G., Pacios, L. F., Del Pozo, J. C. (2010) The Arabidopsis cell cycle F-box protein SKP2A binds to auxin. Plant Cell, 22, 3891-3904.
Jurado, S., Díaz-Triviño, S., Abraham, Z., Manzano, C., Gutierrez, C., Pozo, C. d. (2008) SKP2A, an F-box protein that regulates cell division, is degraded via the ubiquitin pathway. Plant J., 53, 828-841.
Katekar, G. F., Geissler, A. F. (1980) Auxin transport inhibitors. Plant Physiol., 66, 1190-1195.
Kepinski, S., Leyser, O. (2005) The Arabidopsis F-box protein TIR1 is an auxin receptor. Nature, 435, 446-451.
Keuskamp, D. H., Pollmann, S., Voesenek, L. A. C. J., Peeters, A. J. M., Pierik, R. (2010) Auxin transport through PIN-FORMED 3 (PIN3) controls shade avoidance and fitness during competition. Proc. Natl Acad. Sci. U.S.A., 107, 22740-22744.
Kurz, T., Chou, Y.-C., Willems, A. R., Meyer-Schaller, N., Hecht, M.-L., Tyers, M., Peter, M., Sicheri, F. (2008) Dcn1 functions as a scaffold-type E3 ligase for cullin neddylation. Mol. Cell, 29, 23-35.
Leyser, H. M. O., Lincoln, C. A., Timpte, C., Lammer, D., Turner, J., Estelle, M. (1993) Arabidopsis auxin-resistance gene AXR1 encodes a protein related to ubiquitin-activating enzyme E1. Nature, 364, 161-164.
Leyser, O. (2002) Molecular genetics of auxin signaling. Annu. Rev. Plant. Biol., 53, 377-398.
Löbler, M., Klämbt, D. (1985) Auxin-binding protein from coleopti le membranes of corn (Zea mays L.). I. Purification by immunological methods and characterization. J. Biol. Chem., 260, 9848-9853.
Luschnig, C., Gaxiola, R. A., Grisafi, P., Fink, G. R. (1998) EIR1, a root-specific protein involved in auxin transport, is required for gravitropism in Arabidopsis
thaliana. Genes Dev., 12, 2175-2187.MacRae, D. H., Bonner, J. (1953) Chemical structure and
antiauxin activity. Physiol. Plantarum, 6, 485-510.Muday, G. K., Rahman, A. (2008) Auxin transport and the
integration of gravitropic growth. In Plant Tropisms (eds. S. Gilroy, P. Masson) Chapter 3. pp 47-68, Blackwell Publishing, Oxford, UK.
Muller, A., Guan, C., Galweiler, L., Tanzler, P., Huijser, P., Marchant, A., Parry, G., Bennett, M., Wisman, E., Palme, K. (1998) AtPIN2 defines a locus of Arabidopsis for root gravitropism control. EMBO J., 17, 6903-6911.
Nakasone, A., Kawai-Yamada, M., Kiyosue, T., Narumi, I., Uchimiya, H., Oono, Y. (2009) A gene encoding SMALL ACIDIC PROTEIN 2 potentially mediates the response to synthetic auxin, 2,4-dichlorophenoxyacetic acid, in Arabidopsis thaliana. J. Plant Physiol., 166, 1307 - 1313.
Nishimura, T., Nakano, H., Hayashi, K.-i., Niwa, C., Koshiba, T. (2009) Differential downward stream of auxin synthesized at the tip has a key role in gravitropic curvature via TIR1/AFBs-mediated auxin signaling pathways. Plant Cell Physiol., 50, 1874-1885.
Oono, Y., Chen, Q. G., Overvoorde, P. J., Kohler, C., Theologis, A. (1998) age mutants of Arabidopsis exhibited altered auxin-regulated gene expression. Plant Cell, 10, 1649-1662.
Oono, Y., Ooura, C., Rahman, A., Aspuria, E. T., Hayashi, K.-i., Tanaka, A., Uchimiya, H. (2003) p-chlorophenoxyisobutyric acid impairs auxin response in Arabidopsis root. Plant Physiol., 133, 1135-1147.
Paciorek, T., Zazimalova, E., Ruthardt, N., Petrasek, J., Stierhof, Y.-D., Kleine-Vehn, J., Morris, D. A., Emans, N., Jürgens, G., Geldner, N., Friml, J. (2005) Auxin inhibits endocytosis and promotes its own efflux from cells. Nature, 435, 1251-1256.
Pan, J., Fujioka, S., Peng, J., Chen, J., Li, G., Chen, R. (2009) The E3 ubiquitin ligase SCFTIR1/AFB and membrane sterols play key roles in auxin regulation of endocytosis, recycling, and plasma membrane accumulation of the auxin efflux transporter PIN2 in Arabidopsis thaliana. Plant Cell, 21, 568-580.
Parry, G., Calderon-Villalobos, L. I., Prigge, M., Peret, B., Dharmasiri, S., Itoh, H., Lechner, E., Gray, W. M., Bennett, M., Estelle, M. (2009) Complex regulation of the TIR1/AFB family of auxin receptors. Proc. Natl. Acad. Sci. U.S.A., 106, 22540-22545.
Rahman, A., Nakasone, A., Chhun, T., Ooura, C., Biswas, K. K., Uchimiya, H., Tsurumi, S., Baskin, T. I., Tanaka, A., Oono, Y. (2006) A small acidic protein 1 (SMAP1) mediates responses of the Arabidopsis root to the synthetic auxin 2,4-dichlorophenoxyacetic acid. Plant J., 47, 788-801.
Ramos, J. A., Zenser, N., Leyser, O., Callis, J. (2001) Rapid degradation of auxin/indoleacetic acid proteins requires conserved amino acids of domain II and is proteasome dependent. Plant Cell, 13, 2349-2360.
Robert, S., Kleine-Vehn, J., Barbez, E., Sauer, M., Paciorek, T., Baster, P., Vanneste, S., Zhang, J.,

Biswas, K. K. and Oono, Y.
− 55 −
Simon, S., Čovanová, M., Hayashi, K., Dhonukshe, P., Yang, Z., Bednarek, S. Y., Jones, A. M., Luschnig, C., Aniento, F., Zažímalová, E., Friml, J. (2010) ABP1 mediates auxin inhibition of clathrin-dependent endocytosis in Arabidopsis. Cell, 143, 111-121.
Rogg , L .E . , Bar te l , B . (2001) Aux in s igna l ing : derepression through regulated proteolysis. Dev. Cell, 1, 595-604.
Ruegger, M., Dewey, E., Gray, W. M., Hobbie, L., Turner, J., Estelle, M. (1998) The TIR1 protein of Arabidopsis functions in auxin response and is related to human SKP2 and yeast Grr1p. Genes Dev., 12, 198-207.
Sauer, M., Kleine-Vehn, J. (2011) AUXIN BINDING PROTEIN1: the outsider. Plant Cell, 23, 2033-2043.
Schwechheimer, C., Serino, G., Deng, X.-W. (2002) Multiple ubiquitin ligase-mediated processes require COP9 signalosome. Plant Cell, 14, 2553-2563.
Shimizu, M., Miyazawa, Y., Fujii, N., Takahashi, H. (2008) p-Chlorophenoxyisobutyric acid impairs auxin response for gravity-regulated peg formation in cucumber ( Cucumis sativus ) seedlings. J. Plant Res., 121, 107-114.
Staswick, P. E. (2009) The tryptophan conjugates of jasmonic and indole-3-acetic acids are endogenous auxin inhibitors. Plant Physiol., 150, 1310-1321.
Staswick, P. E., Serban, B., Rowe, M., Tiryaki, I., Maldonado, M. T., Maldonado, M. C., Suza, W. (2005) Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant Cell, 17, 616-627.
Sungur, C., Miller, S., Bergholz, J., Hoye, R. C., Brisbois, R. G., Overvoorde, P. (2007) The small molecule 2-furylacrylic acid inhibits auxin-mediated responses in Arabidopsis thaliana. Plant Cell Physiol., 48, 1693-1701.
Takahashi, H., Kamada, M., Yamazaki, Y., Fujii, N., Higashitani, A., Aizawa, S., Yoshizaki, I., Kamigaichi, S . , Muka i , C . , Sh imazu, T. , Fuku i , K . (2000) Morphogenesis in cucumber seedlings is negatively controlled by gravity. Planta, 210, 515-518.
Tan, X. T., Calderon-Villalobos, L. I. A., Sharon, M., Zheng, C., Robinson, C. V., Estelle, M., Zheng, N. (2007) Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature, 446, 640-645.
Theologis, A. (1986) Rapid gene regulation by auxin. Annu. Rev. Plant Physiol., 27, 407-438.
Thimann, K. V. (1977) Hormone action in the whole life of plants. University of Massachusetts Press, Amherst, MA
Tillmann, U., Viola, G., Kayser, B., Siemeister, G., Hesse, T. , Pa lme, K. , Löb ler, M. , K lämbt , D. (1989) cDNA clones of the auxin-binding protein from corn coleoptiles (Zea mays L.): isolation and characterization by immunological methods. EMBO J., 8, 2463-2467.
Tiwari, S. B., Hagen, G., Guilfoyle, T. (2003) The roles of auxin response factor domains in auxin-responsive transcription. Plant Cell, 15, 533-543.
Tóth, R., van der Hoorn, R. (2010) Emerging principles in plant chemical genetics. Trends Plant Sci., 15, 81-88.
Tromas, A., Braun, N., Muller, P., Khodus, T., Paponov, I. A., Palme, K., Ljung, K., Lee, J.-Y., Benfey, P., Murray, J. A. H., Scheres, B., Perrot-Rechenmann, C. (2009) The AUXIN BINDING PROTEIN 1 is required for differential auxin responses mediating root growth. PLoS ONE, 4, e6648.
Tromas, A., Paponov, I . , Perrot-Rechenmann, C. (2010) AUXIN BINDING PROTEIN 1: functional and evolutionary aspects. Trends Plant Sci., 15, 436-446.
Tsai, D.-S., Arteca, R. N. (1984) Inhibition of IAA-induced ethylene production in etiolated mung bean hypocotyl segments by 2,3,5-triiodobenzoic acid and 2-(p-chlorophenoxy)-2-mehyl propionic acid. Physiol. Plant., 62, 448-452.
Tsang, D. L., Edmond, C., Harrington, J. L., Nühse, T. S. (2011) Cell wall integrity controls root elongation via a general 1-aminocyclopropane-1-carboxylic acid-dependent, ethylene-independent pathway. Plant Physiol., 156, 596-604.
Ulmasov, T., Murfett, J., Hagen, G., Guilfoyle, T. (1997) Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell, 9, 1963-1971.
Venis, N. A., Napier, R. M. (1995) Auxin receptors and auxin binding proteins. Crit. Rev. Plant Sci., 14, 27-47.
Walsh, T. A., Neal, R., Merlo, A. O., Honma, M., Hicks, G. R., Wolff, K., Matsumura, W., Davies, J. P. (2006) Mutations in an auxin receptor homolog AFB5 and in SGT1b confer resistance to synthetic picolinate auxins and not to 2,4-dichlorophenoxyacetic acid or indole-3-acetic acid in Arabidopsis. Plant Physiol., 142, 542-552.
Woo, E.-J., Marshall, J., Bauly, J., Chen, J.-G., Venis, M., Napier, R. M., Pickersgill, R. W. (2002) Crystal structure of auxin-binding protein 1 in complex with auxin. EMBO J., 21, 2877-2885.
Woodward, A. W., Bartel, B. (2005) Auxin: regulation, action, and interaction. Ann. Bot., 95, 707-735.
Xu, T., Wen, M., Nagawa, S., Fu, Y., Chen, J.-G., Wu, M.-J., Perrot-Rechenmann, C., Friml, J., Jones, A. M., Yang, Z. (2010) Cell surface- and Rho GTPase-based auxin signaling controls cellular interdigitation in Arabidopsis. Cell, 143, 99-110.
Yamazoe, A., Hayashi, K.-i., Kepinski, S., Leyser, O., Nozaki, H. (2005) Characterization of terfestatin A, a new specific inhibitor for auxin signaling. Plant Physiol., 139, 779-789.
Yip, W.-K., Yang, S. F. (1986) Effect of thidiazuron, a cytokinin-active urea derivative, in cytokinin-dependent ethylene production systems. Plant Physiol., 80, 515-519.
Zhao, Y., Dai, X., Blackwell, H. E., Schreiber, S. L., Chory, J. (2003) SIR1, an upstream component in auxin signaling identified by chemical genetics. Science, 301, 1107-1110.