Embed Size (px)
Citation preview

Molecular heterogeneity of the hemocyanin isolated from the king crabParalithodes camtschaticae
Annamaria Molon1, Paolo Di Muro1, Luigi Bubacco1, Vadim Vasilyev3, Benedetto Salvato1,2, Mariano Beltramini1,2,Werner Conze4, Nadja Hellmann4 and Heinz Decker4
1Department of Biology and 2CNR Center for Metalloproteins, University of Padova, Italy; 3Department of Molecular Genetics,
Institute for Experimental Medicine, St. Petersburg, Russia; 4Institute of Molecular Biophysics, University of Mainz, Germany
Native Paralithodes camtschaticae hemocyanin is found as a mixture of dodecamers (24S; 80%) and hexamers
(16S; 20%). Removal of Ca21 ions by dialysis against EDTA-containing buffer solution at neutral pH induces
complete dissociation of the 24S form into the 16S form. Under these conditions, a further increase in pH to
9.2 produces complete dissociation of the hexamers into monomers (5S). In both cases, the dissociation process
is reversible. The dodecamer (24S) is composed of two different hexamers which can be discriminated only by
ion-exchange chromatography in the presence of Ca21 ions. At alkaline pH and in the presence of EDTA, two
major monomeric fractions can be separated by ion-exchange chromatography: ParcI (60%) and ParcII (40%).
The reassociation properties of the two fractions were studied separately to define their ability to form hexamers
and dodecamers.
The oxygen-binding properties of the different aggregation states were investigated. Native hemocyanin binds
O2 co-operatively (nH � 3) and with low affinity (p50 < 103 Torr). The two monomeric fractions, ParcI and
ParcII, are not co-operative and the affinity is twice that of the native protein ( p50 < 65 and 52 Torr).
Oxygen-binding measurements of native hemocyanin carried out at different pH values indicate a strong
positive Bohr effect within the pH range 6.5±8.0 and an increase in oxygen affinity at pH below 6.5.
Keywords: arthropods; Bohr effect; quaternary structure; hemocyanin; oxygen binding.
Hemocyanins are extracellular respiratory copper proteinspresent at high concentrations in the hemolymph of manyarthropods and molluscs. Native arthropod hemocyanins aremultimeric proteins in which each subunit has a molecular massof < 75 kDa (5S) and contains one binuclear copper sitecapable of binding oxygen [1]. Different functional subunits arearranged as hexamers (16S corresponding to 1 � 6-mer orbuilding block) or multiples of hexamers (24S, 36S, 48S and62S corresponding to 2 � 6-mers, 4 � 6-mers, 6 � 6-mers and8 � 6-mers, respectively). The native aggregation level appearsto be characteristic for each species and may also be related tothe taxonomic groups [1±5]. In the hemolymph of mostcrustacea, e.g. Paralithodes camtschaticae, hexamers (16S) ordodecamers (24S) or both are found. However, the biologicalsignificance of the different aggregation states is stillcontroversial [6]. Although most arthropod hemocyaninsexhibit only one or two aggregation states in vivo, it is usuallypossible to obtain dissociation to the lower structure by anappropriate choice of nonphysiological solution conditions.In particular, removal of bivalent cations (Ca21, Mg21) bydialysis against EDTA and/or a change of pH from the neutralrange will cause dissociation, which in many cases is reversible[1,7].
An interesting feature of arthropod hemocyanins is theirmarked heterogeneity at the subunit level. Such heterogeneityderives from both differences in amino-acid composition andmolecular mass. These types of difference have been observed
in all arthropod hemocyanins [8,9] with the possible exceptionof the polypeptide chain composition of the 16S hexamericcomponents of some isopods [10]. The studies of Markl et al.[8,9] have suggested that, generally, the assembly of thehigher dihexameric, tetrahexameric, hexahexameric and octa-hexameric hierarchies found in various crustacean, chelicerateand centipedal hemocyanins requires a large number ofdifferent subunits. In fact, whereas a single type of subunitappears to be adequate for the correct reassembly of the basichexameric unit [11], certain key bridging or `linker' subunitsare necessary for the formation of higher assemblies, largerthan hexamers [8,9]. Representative examples are the hemo-cyanins from the lobster Homarus americanus, which has fourdifferent subunits, and the spider Eurypelma californicum andscorpion Androctonus australis, which are composed of sevenand eight different subunits, respectively. As shown byimmunoelectron microscopy [12], different subunits occupyspecific locations within the oligomeric structure and aregenerally present in constant proportions. Furthermore, the totalset of subunits is required to reorganize the original aggregatefrom a mixture of dissociated products, indicating that eachof them plays a specific role in the architecture of the protein.This is indicated by the fact that purified subunits often possessthe ability to assemble into homohexamers [5], but hetero-geneity is necessary for the correct assembly of the nativequaternary structure. In crustacean hemocyanins, between twoand eight different monomer types can be electrophoreticallyseparated on high-resolution polyacrylamide gels [13] whilefour to six differently migrating polypeptide chains are found inSDS/PAGE experiments [8,9].
Subunit heterogeneity has a functional role in the modulationof the oxygen-binding properties of hemocyanins; in particular,
Eur. J. Biochem. 267, 7046±7057 (2000) q FEBS 2000
Correspondence to M. Beltramini, Department of Biology, University of
Padova, Via Ugo Bassi 58/B, 35131 Padova, Italy. Fax: 1 39 49 8276344,
Tel.: 1 39 49 8276337, E-mail: [email protected]
(Received 16 May 2000, revised 20 July 2000, accepted 4 October 2000)

it seems to be required for adaptation of organisms in responseto changes in environmental conditions. There is evidencethat, in some crustacean hemocyanin, the subunit compositionvaries with season, oxygen level, or salinity, which results inmodulation of functional properties [14,15]. Furthermore, ithas been shown that the polymorphism is not genetically fixed,and, in relation to this, Mangum & Rainer [16] reported achange in structural and functional phenotypes of Callinectessapidus hemocyanin in response to specific environmentalstimuli.
Hemocyanins bind molecular oxygen as peroxide in am2±h2:h2 co-ordination mode between the two Cu(II) ions[17,18]. The oxygen-binding properties of arthropod hemo-cyanins show a significant dependence on pH, concentrationsof specific ions, and other allosteric effectors [7,19±21]. Thep50 values vary widely, from less than 1 to over 100 Torr.This range reflects the wide variety of environmental conditionsand behavioral patterns observed for different organisms. Thedegree of co-operativity for oxygen binding also shows a widerange of variation and is generally greater in hemocyaninscontaining a larger number of subunits. There is clear evidence,in E. californicum for example, that co-operativity progres-sively increases at higher association states; however, thecomplete assortment of the different subunits is required inreassociated samples to obtain the degree of co-operativityfound in the native molecule [4,22±24]. In contrast, Makino[25] suggested that, in the native hemocyanin of Tachypleusgigas, only some of the subunits that form the hexamers playa specific role in the allosteric control of the co-operativity.As mentioned for the quaternary structure, the oxygen-bindingproperties of hemocyanin also show strong pH-dependence.Most arthropod hemocyanins exhibit a normal Bohr effectexcept for Limulus and other xiphosurans, in which the O2
affinity increases at low pH (reverse Bohr effect) [20,26].Whereas the reverse Bohr shift in Limulus is clearly adaptive,raising O2 affinity and enhancing O2 uptake either in hypoxicwater or during air exposure [27], the significance of thelarge normal Bohr shift in crustaceans remains unclear.
In this study the purification and characterization of hemo-cyanin from the hemolymph of the arthropod P. camtschaticaeis described; this is the largest arthropod, having a crab-likemorphology and a strongly calcified exoskeleton [28]. It livesalong the coasts of the Alaska and Camchatka peninsulasalways within the temperature range 3±7 8C, and, to stay withinthe corresponding isotherm, it undertakes seasonal migrations.Therefore, during the summer, P. camtschaticae migrates todeep water (about 2100 m) [29]. Against this background, wehave studied the molecular heterogeneity of P. camtschaticaehemocyanin in order to evaluate its role in the physiology ofthe organism.
M A T E R I A L S A N D M E T H O D S
Isolation and purification of hemocyanin
Living specimens of P. camtschaticae were caught off thePacific coast of the Gulf of Peter the Great (the Japanese sea).The hemolymph was then collected from living animals bypuncturing the proximal±medial leg joint; typically, about500±1000 mL hemolymph could be collected from a crab of3.5±5 kg. The hemolymph was stored at 220 8C in thepresence of sucrose (20%, w/v) as cryoprotecting agent untilused. The thawed hemolymph was dialysed overnight against50 mm Tris/HCl, containing 20 mm CaCl2 at pH 7.5 afteraddition of a suitable amount of 1 m NaH2PO4 to induce the
precipitation of Ca3(PO4)2. Carotenoids present in the hemo-lymph were adsorbed on hydroxyapatite and removed with thepellet by centrifugation for 10 min at 45 000 g, while the lipidswere eliminated in the same step by phase separation. Thesupernatant was then brought to 30 mm NaH2PO4 and 30 mmCaCl2 and centrifuged for 15 min at 45 000 g. A solution of1 m NaH2PO4 was added to the supernatant obtained from thelast centrifugation to a final concentration of 50 mm. Thesolution was dialysed overnight against 50 mm Tris/HCl/20 mmCaCl2 at pH 7.5 and then centrifuged as described above.Finally, hemocyanin was isolated by double sedimentation in aBeckman ultracentrifuge (model J2-21) at 296 000 g for 4 h at4 8C. The pellet was resuspended in 50 mm Tris/HCl containing20 mm CaCl2 at pH 7.5. Hemocyanin was stored at 220 8Cin the presence of 20% (w/v) sucrose until used. Thehemocyanin concentration was determined spectrophoto-metrically using the molar absorption coefficient at 280 nm:280 � 98 771 m21´cm21 at pH 7.5 and 20 8C (see below).
Determination of the absorption coefficient
The A280 of P. camtschaticae hemocyanin was determined bycorrelating the copper content of a hemocyanin solution with itsabsorbance, assuming a copper-to-protein stoichiometry of2.0 g-atoms copper per mol protein and a molecular mass of75 kDa for the minimal subunit containing one active site. Tothis end, absorption spectra of different hemocyanin solutionswere recorded and corrected for the contribution of lightscattering using the function:
DA�lx� � A0�l0/lx�4 �1�where DA(lx) represents the absorbance of the solution due tolight scattering at a given lx wavelength (nm), A0 is theabsorbance of the solution when there is no contribution fromthe protein (450 nm); at the wavelength l0 no absorbance isexpected from the protein (450 nm). The absorption spectrawere recorded on a Hewlett±Packard HP 8452 spectro-photometer. The copper content of the solutions was deter-mined by atomic absorption using the internal standard method[30].
Characterization of the aggregation states
The different aggregation states of P. camtschaticae hemo-cyanin were studied and characterized using a combination ofgel-filtration and ion-exchange chromatography using FPLCapparatus (Pharmacia). For gel-filtration experiments, aSuperdexw 200 10/30 analytical column and a Superdex200 26/60 preparative column were used. For ion-exchangechromatography, a Mono Qw HR 5/5 analytical column and aSepharose Qw 10/26 preparative column were used with theapplication of a 0±1.0 m NaCl gradient to elute the protein. Theflow rate was 0.5 mL´min21 throughout. The gel-filtrationcolumns were calibrated using a Pharmacia kit of proteinstandards ranging in molecular mass from 13.7 kDa (ribo-nulease) to 669 kDa (thyroglobulin). The void volume of thecolumn was determined using Rapana thomasiana hemocyanin(9000 kDa).
Dissociation and reassociation of hemocyanin
The aggregation state of P. camtschaticae hemocyanin dependson the pH and the presence of bivalent cations (Ca21 or Mg21).In 50 mm Tris/HCl, pH 7.5, and in the presence of 20 mmCaCl2, the protein is found in its native aggregation state. Under
q FEBS 2000 Properties of P. camtschaticae hemocyanin (Eur. J. Biochem. 267) 7047

these conditions, removal of Ca21 ions, by dialysis against10 mm EDTA (volume ratio 1 : 500), induces the completedissociation of dodecameric (24S) hemocyanin into thehexameric (16S) form. In the absence of Ca21 ions, increasingthe pH to 9.2 results in complete dissociation of the protein intomonomers (5S). Reassociation of the monomeric species wascarried out by dialysis of the samples against pH 7.5 buffercontaining either 10 mm EDTA, to obtain the hexamericspecies, or 20 mm CaCl2, to obtain the native form. If nototherwise indicated, the hemocyanin samples were dialysed for24 h at 4 8C.
PAGE
PAGE was performed using a 6% polyacrylamide gel with acontinuous buffer system [8,9] at pH 8.0 (in 50 mm Tris/HCl)or 9.2 (in glycine/NaOH 50 mm, EDTA 1 mm). SDS/PAGE wascarried out on 8% polyacrylamide gel (6% stacking gel) bythe method of Laemmli et al. [31]. A Hoeffer ScientificInstruments SE 600 electrophoresis unit was used throughout.The samples (7.5±10 mg) were run at 4.0 8C and 20 mA. Thegels were stained with Coomassie Brilliant Blue.
Preparation of copper-free hemocyanin
Apohemocyanin was prepared by dialysis of the nativehemocyanin against 50 mm Tris/HCl buffer, containing25 mm KCN, at pH 7.5 for 48 h. The two monomeric fractions,ParcI and ParcII, were analysed at both pH 7.5 and 9.2 tomaintain the protein in the aggregated state and the dissociatedform, respectively. The samples were then dialysed againstCN2-free buffer containing 10 mm EDTA and, finally, against50 mm Tris/HCl buffer at pH 7.5 or 9.2.
Emission spectroscopic measurements
Fluorescence emission spectra were recorded on a Perkin±Elmer LS50 spectrofluorimeter equipped with a thermostatedcell compartment. The two monomeric fractions, ParcI andParcII in the apo form, were diluted in 50 mm Tris/HCl buffer,pH 9.2, containing 10 mm EDTA in order to obtain a finalconcentration of 0.03 mg´mL21. Protein solutions had anabsorbance at the excitation wavelength (294 nm) lower than0.05 to minimize the inner-filter or self-absorption effects.Spectra were recorded in the `ratio' mode to reduce variationsdue to fluctuation of the source of light. In all cases, thesamples were allowed to reach thermal equilibrium for atleast 10 min before the measurements. Fluorescence quantumyields (Q) were calculated using N-acetyl-l-tryptophanamide(Ac-TrpCONH2) as standard with a Q of 0.13 (lecc � 294 nm)at 25 8C. The quantum yield of the protein was analysed as afunction of the temperature increase using the Arrheniusequation:
Q21 2 1 � ke2E a /RT �2�and its logarithmic form:
ln �1=Q 2 1� � 2Ea/RT 1 lnk �3�where T is the temperature expressed in Kelvin, Ea is theactivation energy of the nonradiative deactivation process ofthe excited singlet state (deduced from the slope of thelinear part of the plot in Fig. 7A), R is the gas constant(8.35 J´mol 21´K 21), and k is the rate constant.
Fluorescence emission measurements were carried outbetween 7 and 77 8C.
Oxygen-binding experiments
Two different methods were used to study the oxygen-bindingbehavior of P. camtschaticae hemocyanin.
Tonometric method. Oxygen-binding experiments were per-formed at 23 8C with 0.5±0.8 mg´mL21 hemocyanin solutionsin the appropriate buffers to obtain the aggregation statesrequired in the experiments (see above). The hemocyaninsolution (5 mL) was placed in a tonometer (246 mL) equippedwith a quartz cuvette (d � 1 cm). The degree of oxygenation ofthe protein was followed by measuring the variation inA338 with the tonometer in a Hewlett±Packard HP 8452spectrophotometer. The sample was deoxygenated with a flowof buffer-saturated argon until the A338 decreased to a constantminimum value. Known volumes (0.1±5 mL) of water-saturated oxygen were introduced into the tonometer using agas-tight Hamilton syringe, and the A338 was then recorded.The partial pressure of oxygen pO2 (Torr) was calculated usingthe equation:
p O2 � v/V� p �air�2 p �water�� �4�where v is the volume of oxygen injected, and V is thetonometer volume.
Fluorimetric polarographic method. Continuous oxygen-binding curves, following the intrinsic fluorescence ofhemocyanin, were obtained at different temperatures anddifferent pH values as described by Loewe [32]. Thehemocyanin concentration was below 0.15 mg´mL21 to avoidany inner-filter effect. The 0.1 m Tris/HCl buffer contained20 mm CaCl2 and 20 mm MgCl2. The buffer was adjusted topH 7.5 at 20 8C.
The experimental data resulting from both approaches wereanalysed using the following equation [24,33,34]:
A�x� � u�x��A�1�2 A�0�� � A�0� �5�u(x) is the fractional saturation of the hemocyanin. A(0), A(1)and A(x) are the values of the experimental parameter(absorbance or quantum yield, respectively) considered in theabsence of oxygen, at complete oxygen saturation and at thedifferent oxygen partial pressures, respectively.
The oxygen-binding data were plotted according to the Hillequation [35]:
log Y/�1 2 Y� � n log �pO2�2 n log �p50� �6�where Y is the fractional saturation [also defined as u(x) inEqn (5)], pO2 (Torr) is the partial pressure of oxygen, n is theHill coefficient, and p50 (Torr) is the oxygen partial pressure athalf-saturation.
R E S U LT S
Quaternary structure of P. camtschaticae hemocyanin andcharacterization of heterogeneity at oligomeric level
The first approach to the characterization of the quaternarystructure of P. camtschaticae hemocyanin was gel-filtrationchromatography. The elution profile of P. camtschaticae nativehemocyanin, in 50 mm Tris/HCl buffer, pH 7.5, containing20 mm CaCl2, showed a major peak corresponding to thedodecameric 24S hemocyanin (accounting for about 80% ofthe total hemocyanin) and a shoulder corresponding to thehexameric 16S hemocyanin (accounting for about 20%) (Fig. 1,solid line). This assignment is based on the molecular mass
7048 A. Molon et al. (Eur. J. Biochem. 267) q FEBS 2000
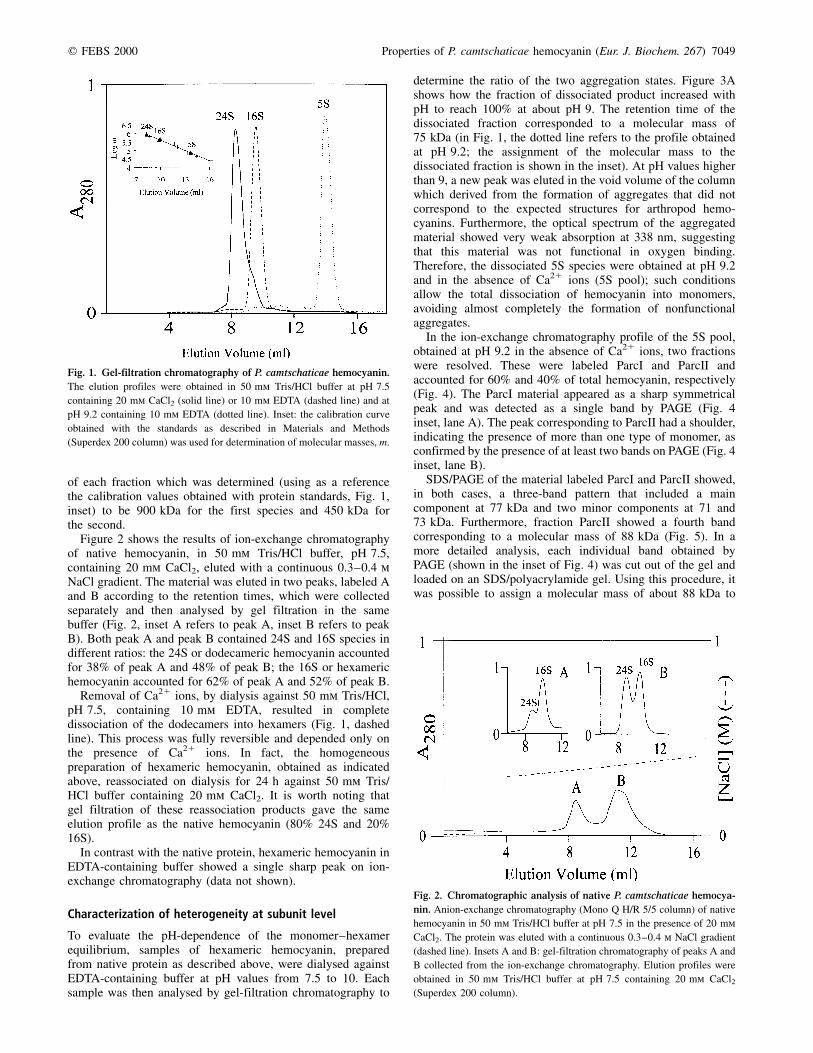
of each fraction which was determined (using as a referencethe calibration values obtained with protein standards, Fig. 1,inset) to be 900 kDa for the first species and 450 kDa forthe second.
Figure 2 shows the results of ion-exchange chromatographyof native hemocyanin, in 50 mm Tris/HCl buffer, pH 7.5,containing 20 mm CaCl2, eluted with a continuous 0.3±0.4 mNaCl gradient. The material was eluted in two peaks, labeled Aand B according to the retention times, which were collectedseparately and then analysed by gel filtration in the samebuffer (Fig. 2, inset A refers to peak A, inset B refers to peakB). Both peak A and peak B contained 24S and 16S species indifferent ratios: the 24S or dodecameric hemocyanin accountedfor 38% of peak A and 48% of peak B; the 16S or hexamerichemocyanin accounted for 62% of peak A and 52% of peak B.
Removal of Ca21 ions, by dialysis against 50 mm Tris/HCl,pH 7.5, containing 10 mm EDTA, resulted in completedissociation of the dodecamers into hexamers (Fig. 1, dashedline). This process was fully reversible and depended only onthe presence of Ca21 ions. In fact, the homogeneouspreparation of hexameric hemocyanin, obtained as indicatedabove, reassociated on dialysis for 24 h against 50 mm Tris/HCl buffer containing 20 mm CaCl2. It is worth noting thatgel filtration of these reassociation products gave the sameelution profile as the native hemocyanin (80% 24S and 20%16S).
In contrast with the native protein, hexameric hemocyanin inEDTA-containing buffer showed a single sharp peak on ion-exchange chromatography (data not shown).
Characterization of heterogeneity at subunit level
To evaluate the pH-dependence of the monomer±hexamerequilibrium, samples of hexameric hemocyanin, preparedfrom native protein as described above, were dialysed againstEDTA-containing buffer at pH values from 7.5 to 10. Eachsample was then analysed by gel-filtration chromatography to
determine the ratio of the two aggregation states. Figure 3Ashows how the fraction of dissociated product increased withpH to reach 100% at about pH 9. The retention time of thedissociated fraction corresponded to a molecular mass of75 kDa (in Fig. 1, the dotted line refers to the profile obtainedat pH 9.2; the assignment of the molecular mass to thedissociated fraction is shown in the inset). At pH values higherthan 9, a new peak was eluted in the void volume of the columnwhich derived from the formation of aggregates that did notcorrespond to the expected structures for arthropod hemo-cyanins. Furthermore, the optical spectrum of the aggregatedmaterial showed very weak absorption at 338 nm, suggestingthat this material was not functional in oxygen binding.Therefore, the dissociated 5S species were obtained at pH 9.2and in the absence of Ca21 ions (5S pool); such conditionsallow the total dissociation of hemocyanin into monomers,avoiding almost completely the formation of nonfunctionalaggregates.
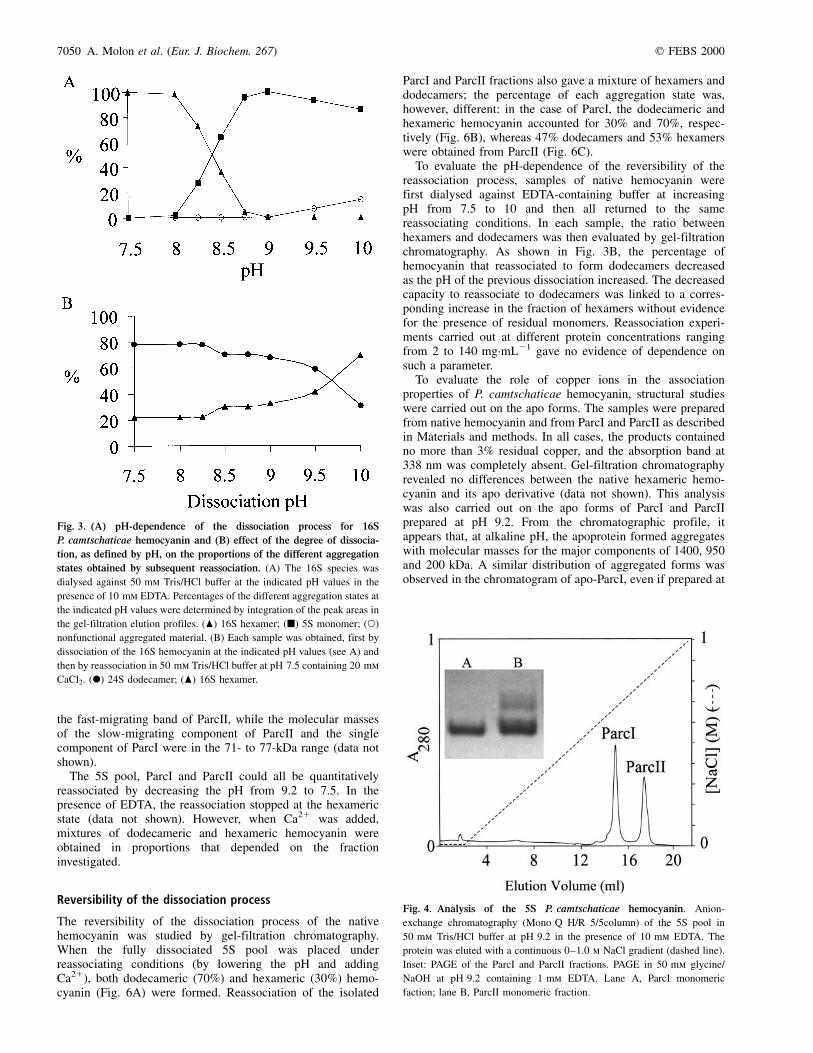
In the ion-exchange chromatography profile of the 5S pool,obtained at pH 9.2 in the absence of Ca21 ions, two fractionswere resolved. These were labeled ParcI and ParcII andaccounted for 60% and 40% of total hemocyanin, respectively(Fig. 4). The ParcI material appeared as a sharp symmetricalpeak and was detected as a single band by PAGE (Fig. 4inset, lane A). The peak corresponding to ParcII had a shoulder,indicating the presence of more than one type of monomer, asconfirmed by the presence of at least two bands on PAGE (Fig. 4inset, lane B).
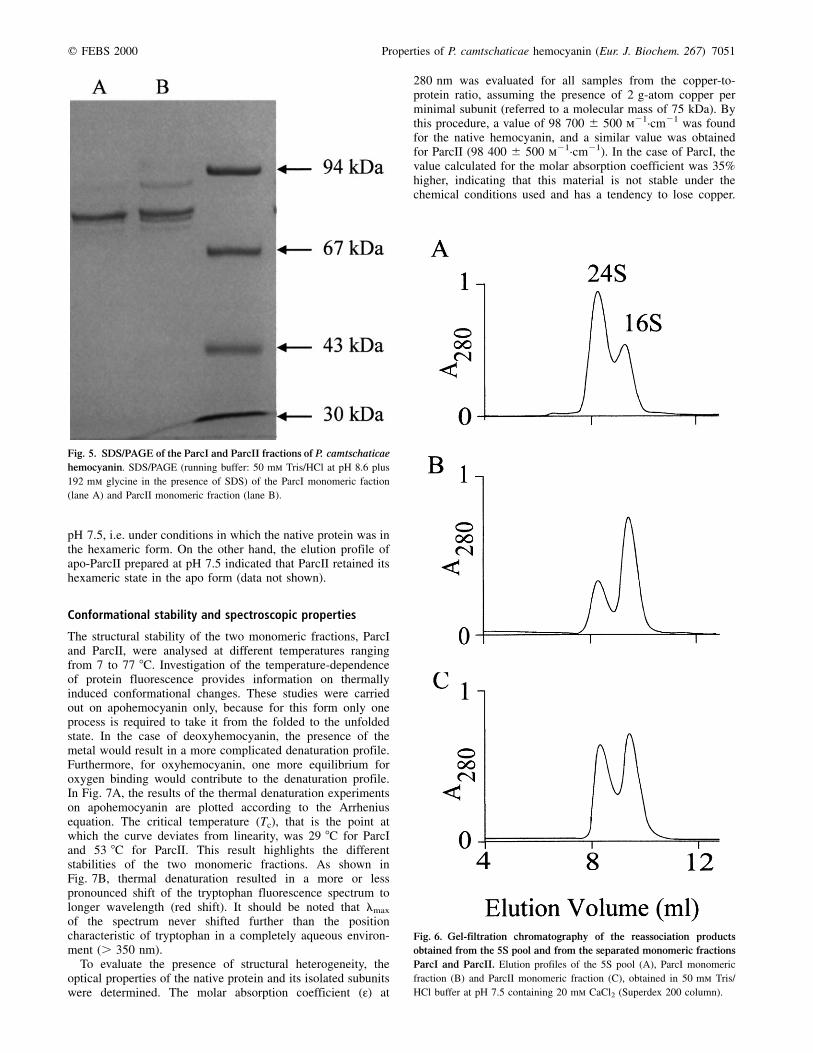
SDS/PAGE of the material labeled ParcI and ParcII showed,in both cases, a three-band pattern that included a maincomponent at 77 kDa and two minor components at 71 and73 kDa. Furthermore, fraction ParcII showed a fourth bandcorresponding to a molecular mass of 88 kDa (Fig. 5). In amore detailed analysis, each individual band obtained byPAGE (shown in the inset of Fig. 4) was cut out of the gel andloaded on an SDS/polyacrylamide gel. Using this procedure, itwas possible to assign a molecular mass of about 88 kDa to
Fig. 1. Gel-filtration chromatography of P. camtschaticae hemocyanin.
The elution profiles were obtained in 50 mm Tris/HCl buffer at pH 7.5
containing 20 mm CaCl2 (solid line) or 10 mm EDTA (dashed line) and at
pH 9.2 containing 10 mm EDTA (dotted line). Inset: the calibration curve
obtained with the standards as described in Materials and Methods
(Superdex 200 column) was used for determination of molecular masses, m.
Fig. 2. Chromatographic analysis of native P. camtschaticae hemocya-
nin. Anion-exchange chromatography (Mono Q H/R 5/5 column) of native
hemocyanin in 50 mm Tris/HCl buffer at pH 7.5 in the presence of 20 mm
CaCl2. The protein was eluted with a continuous 0.3±0.4 m NaCl gradient
(dashed line). Insets A and B: gel-filtration chromatography of peaks A and
B collected from the ion-exchange chromatography. Elution profiles were
obtained in 50 mm Tris/HCl buffer at pH 7.5 containing 20 mm CaCl2(Superdex 200 column).
q FEBS 2000 Properties of P. camtschaticae hemocyanin (Eur. J. Biochem. 267) 7049
the fast-migrating band of ParcII, while the molecular massesof the slow-migrating component of ParcII and the singlecomponent of ParcI were in the 71- to 77-kDa range (data notshown).
The 5S pool, ParcI and ParcII could all be quantitativelyreassociated by decreasing the pH from 9.2 to 7.5. In thepresence of EDTA, the reassociation stopped at the hexamericstate (data not shown). However, when Ca21 was added,mixtures of dodecameric and hexameric hemocyanin wereobtained in proportions that depended on the fractioninvestigated.
Reversibility of the dissociation process
The reversibility of the dissociation process of the nativehemocyanin was studied by gel-filtration chromatography.When the fully dissociated 5S pool was placed underreassociating conditions (by lowering the pH and addingCa21), both dodecameric (70%) and hexameric (30%) hemo-cyanin (Fig. 6A) were formed. Reassociation of the isolated
ParcI and ParcII fractions also gave a mixture of hexamers anddodecamers; the percentage of each aggregation state was,however, different: in the case of ParcI, the dodecameric andhexameric hemocyanin accounted for 30% and 70%, respec-tively (Fig. 6B), whereas 47% dodecamers and 53% hexamerswere obtained from ParcII (Fig. 6C).
To evaluate the pH-dependence of the reversibility of thereassociation process, samples of native hemocyanin werefirst dialysed against EDTA-containing buffer at increasingpH from 7.5 to 10 and then all returned to the samereassociating conditions. In each sample, the ratio betweenhexamers and dodecamers was then evaluated by gel-filtrationchromatography. As shown in Fig. 3B, the percentage ofhemocyanin that reassociated to form dodecamers decreasedas the pH of the previous dissociation increased. The decreasedcapacity to reassociate to dodecamers was linked to a corres-ponding increase in the fraction of hexamers without evidencefor the presence of residual monomers. Reassociation experi-ments carried out at different protein concentrations rangingfrom 2 to 140 mg´mL21 gave no evidence of dependence onsuch a parameter.
To evaluate the role of copper ions in the associationproperties of P. camtschaticae hemocyanin, structural studieswere carried out on the apo forms. The samples were preparedfrom native hemocyanin and from ParcI and ParcII as describedin Materials and methods. In all cases, the products containedno more than 3% residual copper, and the absorption band at338 nm was completely absent. Gel-filtration chromatographyrevealed no differences between the native hexameric hemo-cyanin and its apo derivative (data not shown). This analysiswas also carried out on the apo forms of ParcI and ParcIIprepared at pH 9.2. From the chromatographic profile, itappears that, at alkaline pH, the apoprotein formed aggregateswith molecular masses for the major components of 1400, 950and 200 kDa. A similar distribution of aggregated forms wasobserved in the chromatogram of apo-ParcI, even if prepared at
Fig. 4. Analysis of the 5S P. camtschaticae hemocyanin. Anion-
exchange chromatography (Mono Q H/R 5/5column) of the 5S pool in
50 mm Tris/HCl buffer at pH 9.2 in the presence of 10 mm EDTA. The
protein was eluted with a continuous 0±1.0 m NaCl gradient (dashed line).
Inset: PAGE of the ParcI and ParcII fractions. PAGE in 50 mm glycine/
NaOH at pH 9.2 containing 1 mm EDTA. Lane A, ParcI monomeric
faction; lane B, ParcII monomeric fraction.
Fig. 3. (A) pH-dependence of the dissociation process for 16S
P. camtschaticae hemocyanin and (B) effect of the degree of dissocia-
tion, as defined by pH, on the proportions of the different aggregation
states obtained by subsequent reassociation. (A) The 16S species was
dialysed against 50 mm Tris/HCl buffer at the indicated pH values in the
presence of 10 mm EDTA. Percentages of the different aggregation states at
the indicated pH values were determined by integration of the peak areas in
the gel-filtration elution profiles. (O) 16S hexamer; (B) 5S monomer; (W)
nonfunctional aggregated material. (B) Each sample was obtained, first by
dissociation of the 16S hemocyanin at the indicated pH values (see A) and
then by reassociation in 50 mm Tris/HCl buffer at pH 7.5 containing 20 mm
CaCl2. (X) 24S dodecamer; (O) 16S hexamer.
7050 A. Molon et al. (Eur. J. Biochem. 267) q FEBS 2000
pH 7.5, i.e. under conditions in which the native protein was inthe hexameric form. On the other hand, the elution profile ofapo-ParcII prepared at pH 7.5 indicated that ParcII retained itshexameric state in the apo form (data not shown).
Conformational stability and spectroscopic properties
The structural stability of the two monomeric fractions, ParcIand ParcII, were analysed at different temperatures rangingfrom 7 to 77 8C. Investigation of the temperature-dependenceof protein fluorescence provides information on thermallyinduced conformational changes. These studies were carriedout on apohemocyanin only, because for this form only oneprocess is required to take it from the folded to the unfoldedstate. In the case of deoxyhemocyanin, the presence of themetal would result in a more complicated denaturation profile.Furthermore, for oxyhemocyanin, one more equilibrium foroxygen binding would contribute to the denaturation profile.In Fig. 7A, the results of the thermal denaturation experimentson apohemocyanin are plotted according to the Arrheniusequation. The critical temperature (Tc), that is the point atwhich the curve deviates from linearity, was 29 8C for ParcIand 53 8C for ParcII. This result highlights the differentstabilities of the two monomeric fractions. As shown inFig. 7B, thermal denaturation resulted in a more or lesspronounced shift of the tryptophan fluorescence spectrum tolonger wavelength (red shift). It should be noted that lmax
of the spectrum never shifted further than the positioncharacteristic of tryptophan in a completely aqueous environ-ment (. 350 nm).
To evaluate the presence of structural heterogeneity, theoptical properties of the native protein and its isolated subunitswere determined. The molar absorption coefficient (:) at
280 nm was evaluated for all samples from the copper-to-protein ratio, assuming the presence of 2 g-atom copper perminimal subunit (referred to a molecular mass of 75 kDa). Bythis procedure, a value of 98 700 ^ 500 m21´cm21 was foundfor the native hemocyanin, and a similar value was obtainedfor ParcII (98 400 ^ 500 m21´cm21). In the case of ParcI, thevalue calculated for the molar absorption coefficient was 35%higher, indicating that this material is not stable under thechemical conditions used and has a tendency to lose copper.
Fig. 6. Gel-filtration chromatography of the reassociation products
obtained from the 5S pool and from the separated monomeric fractions
ParcI and ParcII. Elution profiles of the 5S pool (A), ParcI monomeric
fraction (B) and ParcII monomeric fraction (C), obtained in 50 mm Tris/
HCl buffer at pH 7.5 containing 20 mm CaCl2 (Superdex 200 column).
Fig. 5. SDS/PAGE of the ParcI and ParcII fractions of P. camtschaticae
hemocyanin. SDS/PAGE (running buffer: 50 mm Tris/HCl at pH 8.6 plus
192 mm glycine in the presence of SDS) of the ParcI monomeric faction
(lane A) and ParcII monomeric fraction (lane B).
q FEBS 2000 Properties of P. camtschaticae hemocyanin (Eur. J. Biochem. 267) 7051
The second spectroscopic feature analysed was the band at336 nm previously assigned to a charge-transfer transition ofthe copper±peroxide complex [36]. In this case, the : value wasobtained by dividing the absorption difference at 336 nmbetween the oxygenated and deoxygenated samples by theconcentration of protein determined from the absorption at280 nm. The :336 for the fully oxygenated native protein is19 800 ^ 500 m21´cm21. The values for monomeric andhexameric ParcI could not be calculated directly because boththe impossibility of determining :280 and the partial copperdepletion prevented an accurate determination.
Oxygen-binding properties
The equilibria with oxygen for the different aggregation stateswere studied by following the intensity of the charge-transfertransition of the copper±oxygen complex as a function ofligand concentration. Different experiments were carried outtonometrically at 23 8C for the various aggregation states of the
protein. The oxygen-binding curve for native P. camtschaticaehemocyanin, at pH 7.5 and in the presence of 20 mm Ca21, hada sigmoidal profile. Under these conditions, gel filtration of thehemocyanin sample indicated the presence of 80% 24S and20% 16S and oxygen binding with high co-operativity(nH < 3.0) and low affinity ( p50 < 103 Torr) (Table 1). Inthe presence of 10 mm EDTA, the hemocyanin sampleconsisted of two different 16S hemocyanins. At pH 7.5, theybound oxygen in a very similar way with respect toco-operativity and affinity. In comparison with the nativehemocyanin sample, the co-operativity (nH � 1.5) was lowerand the affinity (p50 < 93.5 Torr) slightly higher. The twohomohexamers obtained by reassociating ParcI and ParcII hadsimilar co-operativity and oxygen affinity. However, the twohomohexamers of ParcI and ParcII had a lower affinity than theheterohexamers (Table 1). As expected, ParcI and ParcIImonomers no longer bound oxygen co-operatively. The O2
affinities of the monomers were higher than those of thehomohexameric molecules (Table 1). It should be mentionedthat exposure to alkaline pH (not higher than 9), as required
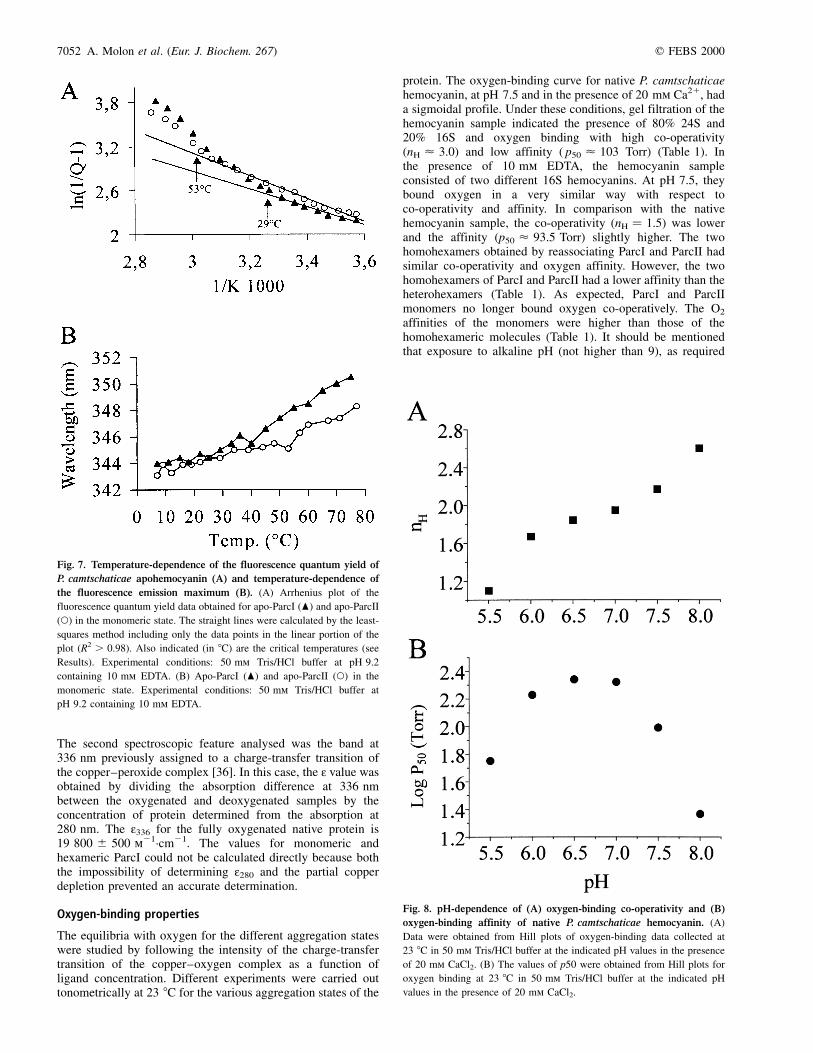
Fig. 8. pH-dependence of (A) oxygen-binding co-operativity and (B)
oxygen-binding affinity of native P. camtschaticae hemocyanin. (A)
Data were obtained from Hill plots of oxygen-binding data collected at
23 8C in 50 mm Tris/HCl buffer at the indicated pH values in the presence
of 20 mm CaCl2. (B) The values of p50 were obtained from Hill plots for
oxygen binding at 23 8C in 50 mm Tris/HCl buffer at the indicated pH
values in the presence of 20 mm CaCl2.
Fig. 7. Temperature-dependence of the fluorescence quantum yield of
P. camtschaticae apohemocyanin (A) and temperature-dependence of
the fluorescence emission maximum (B). (A) Arrhenius plot of the
fluorescence quantum yield data obtained for apo-ParcI (O) and apo-ParcII
(W) in the monomeric state. The straight lines were calculated by the least-
squares method including only the data points in the linear portion of the
plot (R2 . 0.98). Also indicated (in 8C) are the critical temperatures (see
Results). Experimental conditions: 50 mm Tris/HCl buffer at pH 9.2
containing 10 mm EDTA. (B) Apo-ParcI (O) and apo-ParcII (W) in the
monomeric state. Experimental conditions: 50 mm Tris/HCl buffer at
pH 9.2 containing 10 mm EDTA.
7052 A. Molon et al. (Eur. J. Biochem. 267) q FEBS 2000

to obtain dissociated subunits, did not modify the co-operativityor the affinity of the subsequently reassociated species (Table 1).
The pH-dependence of the oxygen-binding curves of nativehemocyanin were studied between pH 5.5 and pH 8.0. A strongbut different pH-dependence was observed for affinity andco-operativity. The Hill coefficient nH decreased with pH from2.6 at pH 8.0 to 1.1 at pH 5.5 (Fig. 8A). The pH-dependenceof the affinity was more complex. Between pH 6.5 and pH 8.0,an increase in pH caused a decrease in p50, corresponding to a
normal Bohr effect (Dlogp50/DpH) of about 20.7. BelowpH 6.5, an inverse Bohr effect was observed, with a valueof about 10.6, obtained by the fluorimetric polarographicmethod. In a second approach, the effect of the temperature andthe temperature-induced changes in pH on oxygen-bindingbehavior were studied for purified 24S hemocyanin. The pHof the 0.1 m Tris/HCl buffer containing 0.15 mg´mL21 hemo-cyanin was adjusted to 7.5 at 20 8C. In the first series ofexperiments, oxygen-binding curves were obtained at threedifferent temperatures (10, 20 and 30 8C). Four pH values (7.0,7.5, 8.0 and 8.5) were investigated. No variation in oxygenaffinity was observed at the four pH values studied when theexperiments were carried out at different temperatures(Fig. 9A). In addition, between pH 7.0 and 8.5, the Bohrcoefficient at 10 8C, 20 8C and 30 8C did not differ signifi-cantly, with a mean ^ SEM value of 20.60 ^ 0.05 (Fig. 10),in agreement with the results obtained by the tonometricapproach. In the same temperature range, the decrease inco-operativity (nH) with increasing temperature was directly
Fig. 9. Oxygen-binding affinity (A) and co-operativity (B) of native
P. camtschaticae hemocyanin in dependence of pH and temperature.
Four different pH values were investigated at three different temperatures.
The pH remained independent from temperature. The binding experiments
were performed in 0.1 m Tris/HCl buffer containing 20 mm CaCl2 and
20 mm MgCl2. The hemocyanin concentration was 0.15 mg´mL21. The
data were obtained from the Hill plot of each binding curve.
Table 1. Oxygen equilibrium parameters of P. camtschaticae hemocyanin under the indicated buffer conditions using the tonometric method.
Samples Buffer conditions n p50 (Torr)
Native pH 7�.5, CaCl2 3�.03 103�.04
Nativea pH 7�.5, CaCl2 2�.98 108�.64
16S pH 7�.5, EDTA 1�.48 93�.00
16Sa pH 7�.5, EDTA 1�.52 93�.97
ParcI pH 9�.2, EDTA 0�.83 64�.62
ParcII pH 9�.2, EDTA 0�.80 51�.67
16S(I) pH 7�.5, EDTA 1�.29 126�.00
16S(II) pH 7�.5, EDTA 1�.31 120�.00
a The samples were first dissociated at pH 9.2 in the absence of Ca21 ions and then brought to the indicated buffer conditions.
Fig. 10. Bohr effect between pH 7.0 and 8.5 of native P. camtschaticae
hemocyanin at 10 8C, 20 8C and 30 8C. The binding curves were obtained
in 0.1 m Tris/HCl buffer containing 20 mm CaCl2 and 20 mm MgCl2. The
hemocyanin concentration was 0.15 mg´mL21.
q FEBS 2000 Properties of P. camtschaticae hemocyanin (Eur. J. Biochem. 267) 7053
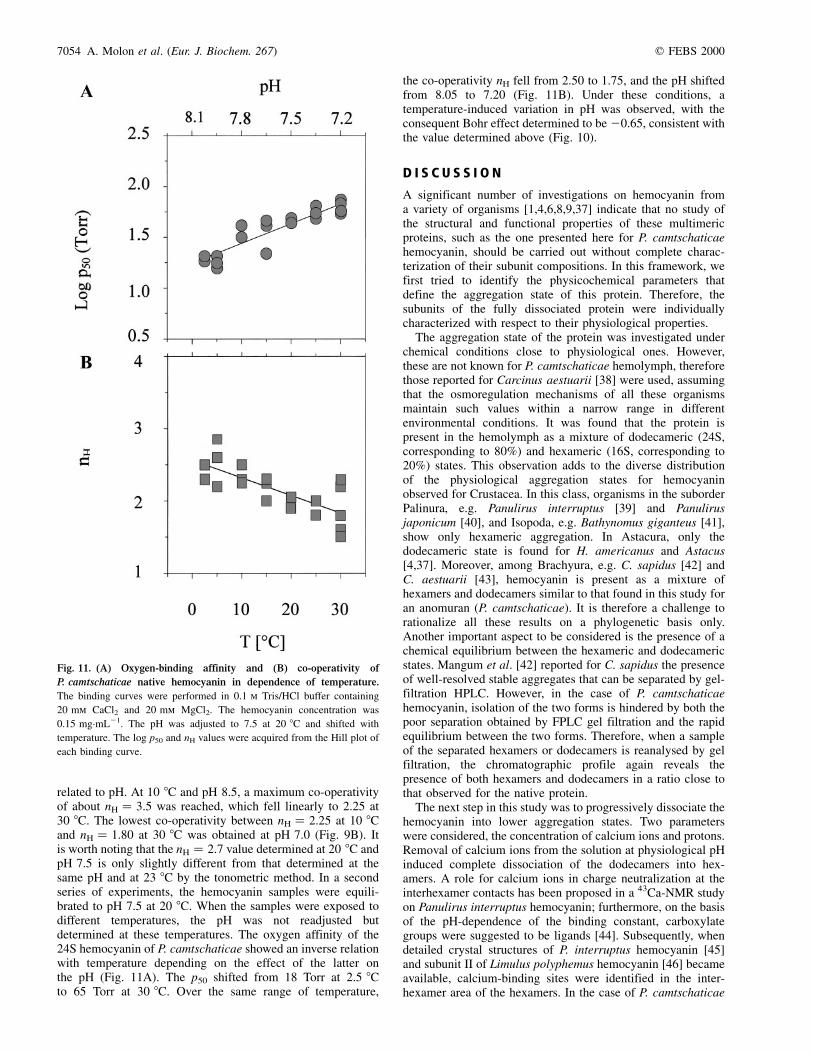
related to pH. At 10 8C and pH 8.5, a maximum co-operativityof about nH � 3.5 was reached, which fell linearly to 2.25 at30 8C. The lowest co-operativity between nH � 2.25 at 10 8Cand nH � 1.80 at 30 8C was obtained at pH 7.0 (Fig. 9B). Itis worth noting that the nH � 2.7 value determined at 20 8C andpH 7.5 is only slightly different from that determined at thesame pH and at 23 8C by the tonometric method. In a secondseries of experiments, the hemocyanin samples were equili-brated to pH 7.5 at 20 8C. When the samples were exposed todifferent temperatures, the pH was not readjusted butdetermined at these temperatures. The oxygen affinity of the24S hemocyanin of P. camtschaticae showed an inverse relationwith temperature depending on the effect of the latter onthe pH (Fig. 11A). The p50 shifted from 18 Torr at 2.5 8Cto 65 Torr at 30 8C. Over the same range of temperature,
the co-operativity nH fell from 2.50 to 1.75, and the pH shiftedfrom 8.05 to 7.20 (Fig. 11B). Under these conditions, atemperature-induced variation in pH was observed, with theconsequent Bohr effect determined to be 20.65, consistent withthe value determined above (Fig. 10).
D I S C U S S I O N
A significant number of investigations on hemocyanin froma variety of organisms [1,4,6,8,9,37] indicate that no study ofthe structural and functional properties of these multimericproteins, such as the one presented here for P. camtschaticaehemocyanin, should be carried out without complete charac-terization of their subunit compositions. In this framework, wefirst tried to identify the physicochemical parameters thatdefine the aggregation state of this protein. Therefore, thesubunits of the fully dissociated protein were individuallycharacterized with respect to their physiological properties.
The aggregation state of the protein was investigated underchemical conditions close to physiological ones. However,these are not known for P. camtschaticae hemolymph, thereforethose reported for Carcinus aestuarii [38] were used, assumingthat the osmoregulation mechanisms of all these organismsmaintain such values within a narrow range in differentenvironmental conditions. It was found that the protein ispresent in the hemolymph as a mixture of dodecameric (24S,corresponding to 80%) and hexameric (16S, corresponding to20%) states. This observation adds to the diverse distributionof the physiological aggregation states for hemocyaninobserved for Crustacea. In this class, organisms in the suborderPalinura, e.g. Panulirus interruptus [39] and Panulirusjaponicum [40], and Isopoda, e.g. Bathynomus giganteus [41],show only hexameric aggregation. In Astacura, only thedodecameric state is found for H. americanus and Astacus[4,37]. Moreover, among Brachyura, e.g. C. sapidus [42] andC. aestuarii [43], hemocyanin is present as a mixture ofhexamers and dodecamers similar to that found in this study foran anomuran (P. camtschaticae). It is therefore a challenge torationalize all these results on a phylogenetic basis only.Another important aspect to be considered is the presence of achemical equilibrium between the hexameric and dodecamericstates. Mangum et al. [42] reported for C. sapidus the presenceof well-resolved stable aggregates that can be separated by gel-filtration HPLC. However, in the case of P. camtschaticaehemocyanin, isolation of the two forms is hindered by both thepoor separation obtained by FPLC gel filtration and the rapidequilibrium between the two forms. Therefore, when a sampleof the separated hexamers or dodecamers is reanalysed by gelfiltration, the chromatographic profile again reveals thepresence of both hexamers and dodecamers in a ratio close tothat observed for the native protein.
The next step in this study was to progressively dissociate thehemocyanin into lower aggregation states. Two parameterswere considered, the concentration of calcium ions and protons.Removal of calcium ions from the solution at physiological pHinduced complete dissociation of the dodecamers into hex-amers. A role for calcium ions in charge neutralization at theinterhexamer contacts has been proposed in a 43Ca-NMR studyon Panulirus interruptus hemocyanin; furthermore, on the basisof the pH-dependence of the binding constant, carboxylategroups were suggested to be ligands [44]. Subsequently, whendetailed crystal structures of P. interruptus hemocyanin [45]and subunit II of Limulus polyphemus hemocyanin [46] becameavailable, calcium-binding sites were identified in the inter-hexamer area of the hexamers. In the case of P. camtschaticae
Fig. 11. (A) Oxygen-binding affinity and (B) co-operativity of
P. camtschaticae native hemocyanin in dependence of temperature.
The binding curves were performed in 0.1 m Tris/HCl buffer containing
20 mm CaCl2 and 20 mm MgCl2. The hemocyanin concentration was
0.15 mg´mL21. The pH was adjusted to 7.5 at 20 8C and shifted with
temperature. The log p50 and nH values were acquired from the Hill plot of
each binding curve.
7054 A. Molon et al. (Eur. J. Biochem. 267) q FEBS 2000

hemocyanin, our results suggest the presence of analogouscalcium-binding sites either in the interhexamer contact area orwith a charge neutralization effect at an intrahexamer site toexplain the dissociation of dodecamers on removal of theseions. The presence of a second class of weaker calcium-bindingsites, also proposed in the 43Ca-NMR study on P. interruptushemocyanin [44], is suggested by the observation that thehexamers appear to be a homogeneous species in the absence ofcalcium, but can be separated according to their overall charge,e.g. by ion-exchange chromatography, in the presence ofcalcium ions. This implies the presence of at least two hexamertypes which differ with respect to the number and/or type ofcalcium-binding sites.
The dissociation proceeds further from hexamers to mono-mers as the pH is increased, resulting in a homogeneousproduct above pH 9.0. The monomers obtained can beseparated by ion-exchange chromatography into two fractions(ParcI and ParcII). The denaturation profile of the twofractions was obtained from the temperature-dependence ofboth the fluorescence emission maximum and the fluorescencequantum yield of the tryptophans. The profiles for thedeoxygenated forms of the two fractions are analogous;however, a significant difference is observed between theapo forms of ParcI and ParcII. The observed behavior of theapo forms with respect to stability may derive from a differencein primary structure, which could also explain the overallcharge difference responsible for the ionic-exchange chromato-graphic separation. Furthermore, PAGE reveals the presence oftwo subunits in the ParcII fraction. An even more complicatedpattern is obtained on SDS/PAGE of the dissociated monomers,both fractions showing a distribution of apparent molecularmasses. These differences in molecular mass may notcorrespond to a real difference in size among the monomers,as Jekel et al. [47] reported, in a study on P. interruptushemocyanin, that the observed differences in mobility onSDS/PAGE between subunits a and b, which have the samemolecular size, derives from a difference in the number ofnegative charges between these subunits. As no sequence dataare available on the different monomers of P. camtschaticaehemocyanin, it is not possible to discriminate between anactual difference in size and a difference in the primarystructure of equally sized subunits. In conclusion, the electro-phoretic analysis provides a clear indication of heterogeneityamong the monomers that constitute native P. camtschaticaehemocyanin.
An important aspect to be considered is the reversibility ofthe dissociation process described above. The dodecamer/hexamer ratio of 80 and 20% observed for the native proteincan be obtained from a mixture of hexamers and monomers ina concentration-independent manner. Furthermore, when theprotein is fully dissociated into monomers, both dodecamersand hexamers are reassociated in a ratio (70 and 30%) not toodifferent from that of the native protein. This observationsuggests that the reassociation process for P. camtschaticaehemocyanin does not require any additional components, butonly the constituent subunits. This should be compared with theself-reassembly properties of other hemocyanins, such as thosefrom T. gigas, E. californicum, A. australis and L. polyphemus[1,6,48,49], where full reassociation could not be achievedstarting from the individual isolated subunits [25], suggestingthat different mechanisms govern the reassociation processof hemocyanin in different organisms. Reassociation to thedodecameric form could also be achieved starting fromthe isolated ParcI and ParcII fractions, indicating that, forP. camtschaticae hemocyanin, either no key or `linker' subunits
[6,37,50,51] are required for the formation of the dodecamersor at least two types are present, one in each fraction, to givethe fully aggregated form. The observed heterogeneity in thesubunit composition has been interpreted as being one methodof adapting to sudden changes in the environment. This hasbeen elegantly shown by Mangum & Rainer [15] for C. sapidus.We suggest that, in P. camtschaticae hemocyanin also, theobserved subunit heterogeneity plays a role in the physiologicalresponse to environmental changes.
The functional characterization of P. camtschaticae hemo-cyanin indicates that, under conditions similar to thosepreviously described for the hemolymph of arthropods, itexhibits co-operative oxygen binding. Comparable co-operativebehavior is observed for hemocyanin reassociated frompreviously dissociated monomers. Therefore, the reversibilityof the dissociation process is not only observed for thequaternary structure as described above, but also for thefunctional properties of the protein. As expected, a sharpdecrease in the degree of co-operativity is observed ondissociation of the dodecamer into hexamers. The comparisonof the oxygen-binding properties of the reassociated hexamersobtained from the separated ParcI and ParcII monomericfraction (nH � 1.3 ^ 0.1; p50 � 123 ^ 3 Torr) indicates thatboth the nH value and the p50 are equal in the two differenthomohexamers. These variables, however, are significantlydifferent from those measured for the native heterohexamer,indicating that subunits from both fractions are required toobtain the fully functional aggregated state.
The cold water crab P. camtschaticae belongs to the familyLithodidae. Its geographical setting is the muddy and sandyfloor of the littoral in the arctic sea of the north Pacific. Inwinter, it lives as deep as 150 m. During spring, female crabsmigrate to the shallower waters of the coast. Male crabs followa few weeks later for reproduction. In summer, when thetemperature of the water of the littoral increases by more than4 8C, the male crabs retreat to deeper levels. The range oftemperature to which this animal is exposed is 3±10 8C. Athigher latitudes, the seawater contains an almost constantamounts of oxygen (6±7 mL´L21) down to 3000 m [52].Therefore, the oxygen gradient between the surroundings andthe interior tissue of the crab is always large enough to ensure asufficient supply of oxygen for aerobic metabolism. The courseof temperature with depth can also be considered to be almostconstant. Thus, these animals experience very stable environ-mental conditions. This sufficient supply of oxygen fits wellwith the functional properties of the animal's hemocyaninand the relatively high p50 value (32 Torr) at 10 8C at pH 7.5.There is no requirement for the crab to increase the oxygenaffinity of its hemocyanin because it is not going to need torespond to decreased oxygen in its environment. A moderateBohr coefficient of 20.57 at 10 8C also indicates a hemocyaninthat is adjusted to a normoxic habitat. Therefore a sharpdepletion in hemocyanin, triggered by a strong Bohr coefficientto deliver enough O2 to the tissues, is not necessary. A Bohreffect of the same order of magnitude was reported for Cancerpagurus (20.70) and Carcinus maenas (20.83) by Truchot [53]and Lallier & Truchot [54], respectively. In contrast, crusta-ceans living under hypoxic conditions, such as the deep-seahydrothermal vent shrimp, have hemocyanin with a large Bohreffect of 21.80 [55]. The low co-operativity (nH � 2.3; pH 7.5;10 8C) also indicates adequate oxygen concentration in thenatural environment of P. camtsacticae. The O2 gradient(DpO2) between the pO2 of hemolymph and tissue is largeenough to allow the hemocyanin to release its oxygen, not in asmall fall in pO2, but slowly in a moderate decrease in pO2.
q FEBS 2000 Properties of P. camtschaticae hemocyanin (Eur. J. Biochem. 267) 7055

Obviously all binding parameters at low temperature andpH 7.5 indicate a hemocyanin that is adapted to a normoxicenvironment. Only the constant p50 values at differenttemperatures but fixed pH values is contrary to the aboveexplanation. This temperature independence of the p50 is oftenfound in crustaceans living under hypoxic conditions, asreported by Sanders & Childress [56] for the mysid Gnatho-phausia ingens. For such a `hypoxic' animal, a drifting p50 withtemperature cannot be tolerated, because of the small pO2
difference between the environment and the tissue. If the p50
increased with increasing temperature, the hemocyanin wouldnot transport enough O2 to the tissue to ensure sufficientaerobic metabolism. However, this cannot be the reason for thep50 of P. camtschaticae hemocyanin being independent oftemperature. It is not clear why it keeps its p50 almost constantover a range of temperatures far away from its nativeenvironmental conditions and therefore providing no evolu-tionary pressure.
A C K N O W L E D G E M E N T S
This work was partially supported by the Russian Foundation for
Fundamental Research Grant N97-04-48888 and by CNR and MURST
(prot. 9805192993-001/-002) grants and the NMFZ Mainz. The research
was carried out in the framework of the CNR-RAMS program for
collaborative research `Copper proteins and copper metabolism'.
R E F E R E N C E S
1. van Holde, K.E. & Miller, K.I. (1995) Hemocyanins. Adv. Protein
Chem. 47, 1±81.
2. Mangum, C.P., Scott, J.L., Black, R.E., Miller, K.I. & Van Holde, K.E.
(1985) Centipedal hemocyanin. Its structure and its implications for
arthropod phylogeny. Proc. Natl Acad. Sci. USA 82, 3721±3725.
3. Morse, M.P., Meyhofer, E., Otto, J.J. & Kuzirian, A.M. (1986)
Hemocyanin respiratory pigment in bivalve molluscs. Science 321,
1302±1304.
4. Markl, J. (1986) Evolution and function of structurally diverse subunits
in the respiratory protein hemocyanin from arthropods. Biol. Bull.
171, 90±115.
5. Magnus, K.A., Lattman, E.E., Volbeda, A. & Hol, W.G.J. (1991)
Hexamers of subunit II from Limulus hemocyanin (a 48-mer) have
the same quaternary structure as whole Panulirus hemocyanin
molecules. Proteins Struct. Funct. 9, 240±254.
6. Markl, J. & Decker, H. (1992) Molecular structure of arthropod
hemocyanins. Adv. Comp. Environ. Physiol. 13, 325±376.
7. Ellerton, H.D., Ellerton, N.F. & Robinson, H.A. (1983) Hemocyanin: a
current perspective. Prog. Biophys. Mol. Biol. 41, 143±248.
8. Markl, J., Markl, A., Schartau, W. & Linzen, B. (1979) Subunit
heterogeneity in arthropod hemocyanin: chelicerata. J. Comp.
Physiol. 130, 283±292.
9. Markl, J., Hofer, A., Bauer, G., Markl, A., Kempter, B., Brenzinger, M.
& Linzen, B. (1979) Subunit heterogeneity in arthropod hemocyanin:
crustacea. J. Comp. Physiol. 133, 167±175.
10. Ramos, R.A., Hamilton, M.G., Herskovits, T.T. & Witkus, E.R. (1987)
Ultracentrifugal characterisation of hemolymph proteins in isopods.
Fed. Proc. 46, 2023±2028.
11. Brenowitz, M., Bonaventura, C. & Bonaventura, J. (1983) Assembly
and calcium-induced cooperativity of Limulus IV hemocyanin: a
model system for analysis of structure-function relationships in the
absence of subunit heterogeneity. Biochemistry 22, 4707±4713.
12. Lamy, J., Bijholt, M.M.C., Sizaret, P.Y., Lamy, J. & van Bruggen, E.F.J.
(1981) Quaternary structure of scorpion (Androctonus australis)
hemocyanin. Localization of subunits with immunological methods
and electron microscopy. Biochemistry 20, 1849±1856.
13. Brenowitz, M., Bonaventura, C., Bonaventura, J. & Gianazza, E.
(1981) Subunit composition of a high molecular weight oligomer:
Limulus polyphemus hemocyanin. Arch. Biochem. Biophys. 210,
748±761.
14. Bellelli, A., Giardina, B., Corda, M., Pellegrini, M.G., Cau, A., Condo,
S.G. & Brunori, M. (1988) Sexual and seasonal variability of lobster
hemocyanin. Comp. Biochem. Physiol. 91A, 445±449.
15. Mangum, C.P. & Rainer, J.S. (1988) The relationship between subunit
composition and O2 binding of blue crab hemocyanin. Biol. Bull. 174,
77±82.
16. Mangum, C.P. (1994) Subunit composition of hemocyanin of
Callinectes sapidus: phenotypes from naturally hypoxic waters and
isolated oligomers. Comp. Biochem. Physiol. 108B, 537±541.
17. Magnus, K.A., Hazes, B., Ton-That, H., Bonaventura, C., Bonaventura,
J. & Hol, W.G.J. (1994) Crystallographic analysis of oxygenated and
deoxygenated states of arthropod hemocyanin shows unusual
differences. Proteins Struct. Funct. Genet. 19, 302±309.
18. Magnus, K.A., Ton-That, H. & Carpenter, J.E. (1994) Recent structural
work on the oxygen transport protein hemocyanin. Chem. Rev. 94,
727±735.
19. Brouwer, M., Bonaventura, C. & Bonaventura, J. (1978) Analysis of
the effect of three different allosteric ligands on oxygen binding by
hemocyanin of the shrimp, Penaeus setiferus. Biochemistry 17,
2148±2154.
20. Diefenbach, C. & Mangum, C.P. (1983) The effect of inorganic ions
and acclimation salinity on oxygen binding of the hemocyanin of
the horseshoe crab, Limulus polyphemus. Mol. Physiol. 4, 197±206.
21. Mason, R.P., Mangum, C.P. & Godette, G. (1983) The influence
of inorganic ions and acclimation salinity on hemocyanin-oxygen
binding in the blue crab Callinectes sapidus. Bioll. Bull. 164,
104±123.
22. Savel-Niemann, A., Markl, J. & Linzen, B. (1988) Hemocyanins in
spiders. XXII. Range of allosteric interaction in a four-hexamer
hemocyanin. Co-operativity and Bohr effect in dissociation inter-
mediates. J. Mol. Biol. 204, 385±395.
23. Decker, H., Savel-Niemann, A., Korschenhausen, D., Ekerskorn, E. &
Markl, J. (1989) Allosteric oxygen-binding properties of reassembled
tarentula (Eurypelma californicum) hemocyanin with incorporated
Apo- or Met- subunits. Biol. Chem. Hoppe-Seyler 370, 511±523.
24. Decker, H. & Sterner, R. (1990) Nested allostery of arthropodan
hemocyanin (Eurypelma californicum and Homarus americanus).
The role of protons. J. Mol. Biol. 211, 281±293.
25. Makino, N. (1989) Hemocyanin from Tachypleus gigas. II. Cooperative
interaction of the subunits. J. Biochem.(Tokyo) 106, 423±429.
26. Truchot, J.P. (1992) Respiratory function of arthropod hemocyanins.
Adv. Comp. Environ. Physiol. 13, 385±386.
27. Mangum, C.P. (1983) The effect of hypoxia on the hemocyanin±
oxygen binding in the horseshoe crab Limulus polyphemus. Mol.
Physiol. 3, 217±224.
28. Cunningham, C.V., Blackstone, N.W. & Buss, L.W. (1992) Evolution
of king crabs from hermit crab ancestors. Nature (London) 355,
539±542.
29. Stone, R.P., O'Clair, C.E. & Shirley, T.C. (1992) Seasonal migration
and distribution of female red king crabs in a Southeast alaskan
estuary. J. Crust. Biol. 12, 546±560.
30. Bubacco, L., Magliozzo, R.S., Beltramini, M., Salvato, B. & Peisach, J.
(1992) Preparation and spectroscopic characterisation of a coupled
binuclear center in cobalt(II)-substituted hemocyanin. Biochemistry
31, 9294±9303.
31. Laemmli, U.K. (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature (London) 227,
680±685.
32. Loewe, R. (1978) Hemocyanins in spiders. V. Fluorimetric recording of
oxygen binding curves, and its application to the analysis of allosteric
interactions in Eurypelma californicum hemocyanin. J. Comp.
Physiol. 128, 161±168.
33. Gill, S.J., Di Cera, E., Doyle, M.L., Bishop, G.A. & Robert, C.H.
(1987) Oxygen binding constants for human haemoglobin tetramers.
Biochemistry 26, 3995±4002.
34. Gill, S.J., Connelly, P.R., Di Cera, E. & Robert, C.H. (1988) Analysis
7056 A. Molon et al. (Eur. J. Biochem. 267) q FEBS 2000

and parameter resolution in highly cooperative. Biophys. Chem. 30,
133±141.
35. Hill, A.V. (1910) The possible effect of the aggregation of the
molecule of haemoglobin on its dissociation curve. J. Physiol.
London 40, 4±7.
36. Solomon, E.I. (1981) Binuclear copper active site. Copper Proteins
(Spiro, T.G., ed), pp. 41±108. Academic Press, New York.
37. Herskovits, T.T. (1988) Recent aspects of the subunit organisation
and dissociation of hemocyanins. Comp. Biochem. Physiol. 91B,
597±611.
38. Angelini, E. (1997) Sistemi coinvolti nel trasporto dell'ossigeno:
adattamento alle basse temperature nell'ambiente marino. PhD
Thesis, University of Padua.
39. Kuiper, H.A., Gaastra, W., Beintema, J.J., van Bruggen, E.F.J.,
Schepman, A.M.H. & drenth, J. (1975) Subunit composition, X-ray
difraction, amino acid analysis and oxygen binding behaviour of
Panulirus interruptus hemocyanin. J. Mol. Biol. 99, 619±629.
40. Makino, N. (1986) Analysis of oxygen binding to Panulirus japonicus
hemocyanin. The effect of divalent cations on the allosteric transition.
Eur. J. Biochem. 154, 49±55.
41. van Holde, K.E. & Brenowitz, M. (1981) Subunit structure and physical
properties of the hemocyanin of the giant Isopod Bathynomus
giganteus. Biochemistry 20, 5232±5239.
42. Mangum, C.P., Greaves, J. & Rainer, J.S. (1991) Oligomer composition
and oxygen binding of the hemocyanin of the blue crab Callinectes
sapidus. Biol. Bull. 181, 453±458.
43. Dainese, E., Di Muro, P., Beltramini, M., Salvato, B. & Decker, H.
(1998) Subunits composition and allosteric control in Carcinus
aestuarii hemocyanin. Eur. J. Biochem. 256, 350±358.
44. Andersson, T., Chiancone, E. & Forsen, S. (1982) Characterisation of
cation-binding sites on Panulirus interruptus hemocyanin by 43Ca
and 23Na NMR. Eur. J. Biochem. 125, 103±108.
45. Volbeda, A. & Hol, W.G.J. (1989) Crystal structure of hexameric
hemocyanin from Panulirus interruptus refined at 3.2 AÊ resolution.
J. Mol. Biol. 209, 249±279.
46. Hazes, B., Magnus, K.A., Kalk, K.H., Bonaventura, C. & Hol, W.G.J.
(1996) Nitrate binding to Limulus polyphemus subunit type II hemo-
cyanin and its functional complications. J. Mol. Biol. 262, 532±542.
47. Jekel, P.A., Bak, H.J., Soeter, N.M., Vereijken, J.M. & Beintema, J.J.
(1988) Panulirus interruptus hemocyanin; the amino acid sequence
of subunit b and anomalous behaviour a and b on polyacrylamide
gel electrophoresis in the presence of SDS. Eur. J. Biochem. 178,
403±412.
48. Markl, J., Decker, H., Linzen, B., Schtter, W.G. & van Bruggen, E.F.J.
(1982) Hemocyanins in spiders. XV. The role of individual subunits in
the assembly of Eurypelma hemocyanin. Hoppe-Seyler's Z. Physiol.
Chem. 363, 73±87.
49. van Holde, K.E. & Miller, K.J. (1982) Hemocyanins. Q. Rev. Biophys.
15, 1±129.
50. StoÈcker, W., Raeder, U., Bijolt, M.M., Wichertjes, T., van Bruggen
E.F.J. & Markl, J. (1988) The quaternary structure of four crustacean
two-hexameric hemocyanins: immunocorrelation, stoichiometry,
reassembly and topology of individual subunits. J. Comp. Physiol.
B 158, 271±289.
51. Markl, J., Decker, H., StoÈcker, W., Savel, A., Linzen, B., Schutter W.G.
& van Bruggen E.J.F. (1981) On the role of dimeric subunits in
the quaternary structure of arthropod hemocyanins. Hoppe-Seylers's
Z. Physiol. Chem. 362, 185±188.
52. Tardent, P. (1979) Meeresbiologie, 1st edn, p. 233. Thieme-Verlag,
Stuttgart.
53. Truchot, J.P. (1980) Lactate increases the oxygen affinity of crab
hemocyanin. J. Exp. Zool. 214, 205±208.
54. Lallier, F.H. & Truchot, J.-P. (1989) Hemolymph oxygen transport
during environmental hypoxia in the shore crab, Carcinus maenas.
Respir. Physiol. 77, 323±336.
55. Lallier, F.H. & Truchot, J.-P. (1997) Hemocyanin oxygen-binding
properties of a deep-sea hydrothermal vent shrimp: evidence for a
novel cofactor. J. Exp. Zool. 277, 357±364.
56. Sanders, N.K. & Childress, J.J. (1990) Adaptations to the deep-sea
oxygen minimum layer: oxygen binding by the hemocyanin of the
bathypelagic mysid, Gnathophausia ingens Dohrn. Biol. Bull. 178,
286±294.
q FEBS 2000 Properties of P. camtschaticae hemocyanin (Eur. J. Biochem. 267) 7057





























