Embed Size (px)
Citation preview
Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/23764220
ModularControlofPointingbeyondArm'sLength
ARTICLEinTHEJOURNALOFNEUROSCIENCE:THEOFFICIALJOURNALOFTHESOCIETYFORNEUROSCIENCE·FEBRUARY2009
ImpactFactor:6.34·DOI:10.1523/JNEUROSCI.3426-08.2009·Source:PubMed
CITATIONS
30
READS
35
4AUTHORS,INCLUDING:
BastienBerret
UniversitéParis-Sud11
32PUBLICATIONS302CITATIONS
SEEPROFILE
ThierryPozzo
INSERMDijon
182PUBLICATIONS3,120CITATIONS
SEEPROFILE
Availablefrom:BastienBerret
Retrievedon:03February2016

Behavioral/Systems/Cognitive
Modular Control of Pointing beyond Arm’s Length
Bastien Berret,1,2 Francois Bonnetblanc,1,2 Charalambos Papaxanthis,1,2 and Thierry Pozzo1,2,3
1Universite de Bourgogne, Dijon, Campus Universitaire, Unite de Formation et de Recherche en Sciences et Techniques des Activites Physiques et Sportives,and 2Institut National de la Sante et de la Recherche Medicale, Unite 887, Motricite-Plasticite, F-21078 Dijon, France, and 3Italian Institute of Technology,16163 Genoa, Italy
Hand reaching and bipedal equilibrium are two important functions of the human motor behavior. However, how the brain plansgoal-oriented actions combining target reaching with equilibrium regulation is not yet clearly understood. An important question iswhether postural control and reaching are integrated in one single module or controlled separately. Here, we show that postural controland reaching motor commands are processed by means of a modular and flexible organization. Principal component and correlationanalyses between pairs of angles were used to extract global and local coupling during a whole-body pointing beyond arm’s length. Alow-dimensional organization of the redundant kinematic chain allowing simultaneous target reaching and regulation of the center ofmass (CoM) displacement in extrinsic space emerged from the first analysis. In follow-up experiments, both the CoM and finger trajec-tories were constrained by asking participants to reach from a reduced base of support with or without knee flexion, or by moving theendpoint along a predefined trajectory (straight or semicircular trajectories). Whereas joint covaried during free conditions and underequilibrium restrictions, it was decomposed in two task-dependent and task-independent modules, corresponding to a dissociation ofarm versus legs, trunk, and head coordination, respectively, under imposed finger path conditions. A numerical simulation supported theidea that both postural and focal subtasks are basically integrated into the same motor command and that the CNS is able to combine orto separate the movement into autonomous functional synergies according to the task requirements.
Key words: whole-body pointing; equilibrium; center of mass; finger trajectory; joint coupling; modular control
IntroductionMost of our daily life activities involve the entire body. However,the two traditional ways to investigate human motor control ei-ther focus on the equilibrium mechanisms (for review, see Horakand McPherson, 1996) without considering hand trajectory for-mation or, conversely, on reaching movements by artificially re-stricting the motion of the lower body parts (for review, see Shad-mehr and Wise, 2005). Nevertheless, the picture of how the CNSplans goal-oriented action may probably be clarified by askinghow all body parts are coordinated during a pointing task. Forinstance, when reaching beyond arm’s length from a standingposture, the trunk-plus-arm action causes a displacement of thewhole body with respect to the direction of gravity. Thus, inaddition to arm trajectory formation and target reaching, theCNS must anticipate the potential loss of balance.
An open question is whether these two subtasks are controlledas one single central command or separately (Hess, 1954; Belenkiiet al., 1967; Cordo and Nashner, 1982; Schepens and Drew,2003). A classical approach is to consider the two mechanisms ina hierarchical manner: equilibrium maintenance would be sub-ordinated to reaching and lower limbs would offset the displace-
ment of the center of mass (CoM) accompanying voluntary armmovement (Massion, 1992). According to this interpretation,lower and upper body parts would ensure equilibrium preserva-tion and finger trajectory formation, respectively. Alternatively, agrouping of body angles was proposed as a kinematic solution toreduce the effective number of degrees of freedom and simplifythe motor command (Bernstein, 1967; Alexandrov et al., 1998,2001a,b; Lacquaniti et al., 1999; Thomas et al., 2005). In this case,equilibrium and reaching subtasks would be integrated in thesame module, a term that we use to denote the covariation of aspecific set of angles. To check whether the CNS would controlone or several functional modules regrouping or dissociating fo-cal and equilibrium task objectives, we tested the hypothesis that,according to the task demand, the CNS controls the whole bodyeither as one single or two distinct modules. The finding of bothglobal and local kinematic synergies would support a flexiblemodular organization (Bizzi et al., 2008) able to integrate or dis-sociate, if necessary, equilibrium maintenance and target reach-ing rather than a subordinate mode of control.
Our approach consisted of applying equilibrium and spatialconstraints on a natural unstable hand reaching and analyze thekinematics of the resulting movements to infer the mechanismsunderlying the coordination of the two subtasks. In one condi-tion, reaching was performed from a reduced base of support orwithout knee flexion to limit the capacity of producing horizontalor vertical CoM displacements. In another condition, subjectswere asked to reach the target following a straight or semicircularfinger trajectory. The priority given to one of the subtasks mayclarify how the different body parts enter into coordination. Fig-
Received July 22, 2008; revised Nov. 24, 2008; accepted Nov. 26, 2008.This work was supported by Centre National d’Etudes Spatiales and by Conseil Regional de Bourgogne. We thank
C. Darlot, P. Morasso, and P. J. Stapley for their useful comments.Correspondence should be addressed to Thierry Pozzo, Institut National de la Sante et de la Recherche Medicale,
Unite 887, Campus Universitaire, Motricite-Plasticite, F-21078 Dijon, France. E-mail: [email protected]:10.1523/JNEUROSCI.3426-08.2009
Copyright © 2009 Society for Neuroscience 0270-6474/09/290191-15$15.00/0
The Journal of Neuroscience, January 7, 2009 • 29(1):191–205 • 191

ure 1 sums up the possible motor strategiesin response to these experimental restric-tions. Reaching a target under postural orfocal constraints could produce a globaleffect in which all body parts are integratedinto one module, supporting the idea of asynergic control of postural and goal-oriented aspects of the action. Alterna-tively, the motor planning could be split intwo distinct modules, one driving armreaching and one maintaining equilibri-um; the two modules could be new or bederived from the basic motor strategy ini-tially present in the free condition. The re-sults show that, when priority is given tohand trajectory, a second module origi-nates from the basic kinematic strategysuggesting a flexible coordination of pos-ture and movement.
Materials and MethodsParticipants. Twenty-six adults (all males,18 –29 years; mean height, 1.76 � 0.08 m; meanweight, 68 � 9 kg) agreed voluntarily to partic-ipate in the experiments. All were healthy withno previous neurological disease and normal orcorrected-to-normal vision. The experimentsconformed to the Declaration of Helsinki andinformed consent was obtained from all theparticipants according to the protocol of the Ethics Committee of theUniversite de Bourgogne. The experimental protocol is illustrated inFigure 2. The participants were divided in two groups: the first one per-formed the “basic” and “equilibrium” experimental tasks (Fig. 2a,b,b�),whereas the second group performed the “basic” and “spatial” experi-mental tasks (Fig. 2a,c,c�).
Basic motor task. All the participants performed the basic motor task (Bcondition) (Fig. 2a) and the recorded results served as reference to com-pare the results of the other experimental tasks. A similar protocol wasalready used in recent studies (Pozzo et al., 2002; Schmid et al., 2006).Participants were asked to point with the two arms simultaneously to-ward a pair of targets placed at two different distances [lateral coordi-nates: Z � �0.2 m; vertical coordinates: Y � 15% of each participant’sheight; anteroposterior (A-P) coordinates: X, depending on the targetdistance]. The first target distance corresponded to 5% (short distance,D1), and the second to 30% (long distance, D2) of the participants’height. Both distances were measured from the distal end of the partici-pants’ great toe. Participants started from an upright standing positionand their hands were located initially at the external side of the thighs andthus induced hand-pointing movements in a semipronated position. Thewhole movement was performed in the sagittal plane, with each side ofthe body moving together. Target accuracy was not the primary con-straint imposed on the participants during the experiments, and no in-struction was given to them regarding the movements to be effected bythe body segments. Each participant achieved, at a natural (self-selected)speed, a block of 10 pointing movements for each condition of distance.
Equilibrium constraint. Twelve participants took part in the experi-ment in which equilibrium constraints were added to the basic motortask (Fig. 2b,b�). Equilibrium constraints were introduced in two differ-ent ways. Participants were asked to point toward the targets (1) byfreezing their knee joints, a condition that imposed a high inverted pen-dulum configuration reducing stability (K condition, extended kneecondition) (Fig. 2b), and (2) from a reduced base of support (R condi-tion) defined by a 40 cm large horizontal square (wooden board) fixed ona thin piece of wood (5 cm high, 5 cm large, and 40 cm long) (Fig. 2b�). Inboth conditions, they were asked to perform the motor task withoutfalling from the small support or loosing equilibrium control. Partici-pants stayed on the wooden board with the vertical projection of the
malleolus of each foot at the backward limit of the thin piece of wood.Both motor tasks (K and R) were executed at a natural pace. Within eachmotor task, the number of trials and the order of execution of the twodistances were similar to those of the basic motor task. One or twopreliminary trials allowed participants to familiarize themselves with theexperimental restrictions.
Spatial constraint. Fourteen participants took part in the experiment(Fig. 2c,c�) in which spatial constraints were added to the basic motortask. Participants were asked to point to the targets by using a straightfinger trajectory (S condition) (Fig. 2c). Participants initially performedthree nonrecorded trials by following a straight wire connecting the ini-tial finger position to the target. After this short period, they were askedto perform the task without wire and to keep the same speed as in previ-ous experiments. Participants were also requested to reach the targetswith large finger path curvatures (C condition; semicircular finger tra-jectory) (Fig. 2c�). The imposed path was concave in the sagittal plane(semicircle whose diameter was equal to the distance between the initialfinger position and the target). Again, the participants performed threenonrecorded trials by tracking a curved wire connecting the initial fingerposition and the targets. Then, they were asked to perform the task with-out wire. Both tasks were performed at a natural pace. Within each motortask, the number of trials and the order of execution of the two distanceswere similar to those of the basic motor task. The participants wereallowed to rest 1 min between each condition and the total duration ofthe whole experiment was 1.5 h.
Data collection and processing. Movement kinematics was capturedusing an optoelectronic device (SMART-BTS). Nine cameras were usedto measure the position of 11 retro-reflective markers (15 mm in diam-eter), which were placed at various anatomical locations on the right sideof the body (external cantus of the eye, auditory meatus, acromial pro-cess, humeral lateral condyle, ulnar styloid process, apex of the indexfinger, D1 vertebra, greater trochanter, knee interstitial joint space, ex-ternal malleolus, and fifth metatarsal head of the foot).
All analyses were performed with custom software written in Matlab(Mathworks) from the recorded three-dimensional position of the 11markers (sampling frequency, 120 Hz). Recorded signals were low-passfiltered using a digital fifth-order Butterworth filter at a cutoff frequencyof 7.5 Hz (Matlab filtfilt function). The whole-body motion was per-
Figure 1. Schema illustrating the framework hypotheses. On the left, the rectangle represents the basic motor strategy (A)composed of a single module. Because of experimental task restrictions (spatial and equilibrium constraints) applied to A, twopossible cases can be considered: a new motor strategy composed of one module, different from A or similar to A, can beelaborated by the CNS (B); alternatively, a new motor strategy (C) composed of two or more modules (ci) can be elaborated.However, two possibilities can be considered in such a case: C can be composed of new modules, task dependent and differentfrom any submodule (a submodule corresponds to the covariation of some body parts initially included in a module) of A, or someof these modules can be task independent and come from the initial motor strategy A.
192 • J. Neurosci., January 7, 2009 • 29(1):191–205 Berret et al. • Modular Motor Control
formed in the sagittal plane. To verify the planarity hypothesis of themovement, we used a principal component analysis (PCA) (Jolliffe,1986) on the whole three-dimensional dataset of each participant(grouping all markers as individual observations). The variance ac-counted for (VAF) by the two first principal components (PCs) was�98% for all participants, indicating that most markers trajectories wereapproximately lying on one plane. In particular, this plane was nearly theone defined by the reference frame of the acquisition system (XY, whereX was the A-P axis and Y was the vertical axis). The angle between normalvectors of the planes was �4 � 1.5°. Therefore, we projected the three-dimensional data on the plane XY (X, anteroposterior axis; Y, verticalaxis) defined by our recording system.
Movement analysis. An estimation of CoM displacements was used tocharacterize the equilibrium performance. We estimated the position ofthe CoM using an eight-segment mathematical model consisting of thefollowing rigid segments: head, trunk, thigh, shank, foot, upper arm,forearm, and hand. Using this model, the position of the CoM was cal-culated via standard procedures and using documented anthropometricparameters (Winter, 1990). The model used to determine the whole-body CoM position has previously been validated for similar whole-bodyreaching movements (Stapley et al., 1999). The final whole-body mo-mentum (WBM) was also evaluated using an inverse pendulum model inwhich the whole-body mass was concentrated in its CoM. It was evalu-ated using the standard formula mgxA-P, where m was the subject mass, gis the gravity acceleration (�9.81 m � s �2), and xA-P was the distancebetween the ankle (fixed point) and the projection of the CoM on the A-Paxis.
The finger kinematics was used to analyze the pointing performance.Finger movement onset time was defined as the instant at which thelinear tangential velocity of the index fingertip exceeded 5% of its peakand the end of movement as the point at which the same velocity dropped
below the 5% threshold. Standard kinematic parameters usually de-scribed in arm-pointing studies (Papaxanthis et al., 2005) were calculatedas follows: movement duration (MD), peak velocity (PV), mean velocity(MV), relative time to peak velocity (TPV) defined as the ratio of theduration of acceleration and MD, index of finger path curvature (IPC �Dev/LD) defined as the ratio of the maximum path deviation (Dev) froma straight line connecting the initial and final finger positions [lineardistance (LD)], and curvilinear distance of the finger (CD) defined by theintegral over time from 0 to MD of the norm of the finger velocity vector(in plane XY ). To evaluate the consistency of the final finger position, wecalculated the 95% confidence ellipses in plane XY. Constant errors weredefined as the distance between the finger position and the target in threedimensions.
Eight elevation angles (with respect to the vertical axis Y; Sk, shank; Th,thigh; Pe, pelvis; Tr, trunk; He, head; Hu, humerus; Fo, forearm; Ha,hand) were defined (Fig. 2, inset). Arm angles will be referred to as“upper limb” angles in opposition to the angles of leg, trunk, and head,which will be referred to as “lower limb” angles, for the sake of simplicity.The temporal window used for analyzing the angular displacement wasbased on the finger onset and ending times defined above. We visuallyverified that the finger marker was the first and the last to move com-pared with the other markers placed on the body. The angular time seriesare depicted in Figure 3. These time series were then used to compute (1)the amplitude of each angular displacement (defined as the absolutevalue of difference between the initial and final angle) and (2) an index ofthe whole-body coordination using PCA (see below).
All time series were time-normalized to 200 points by using Matlabroutines of interpolation (Matlab spline function).
Principal component analysis. PCA was applied to the angular displace-ments. Each sample of time was considered as a single observation lying
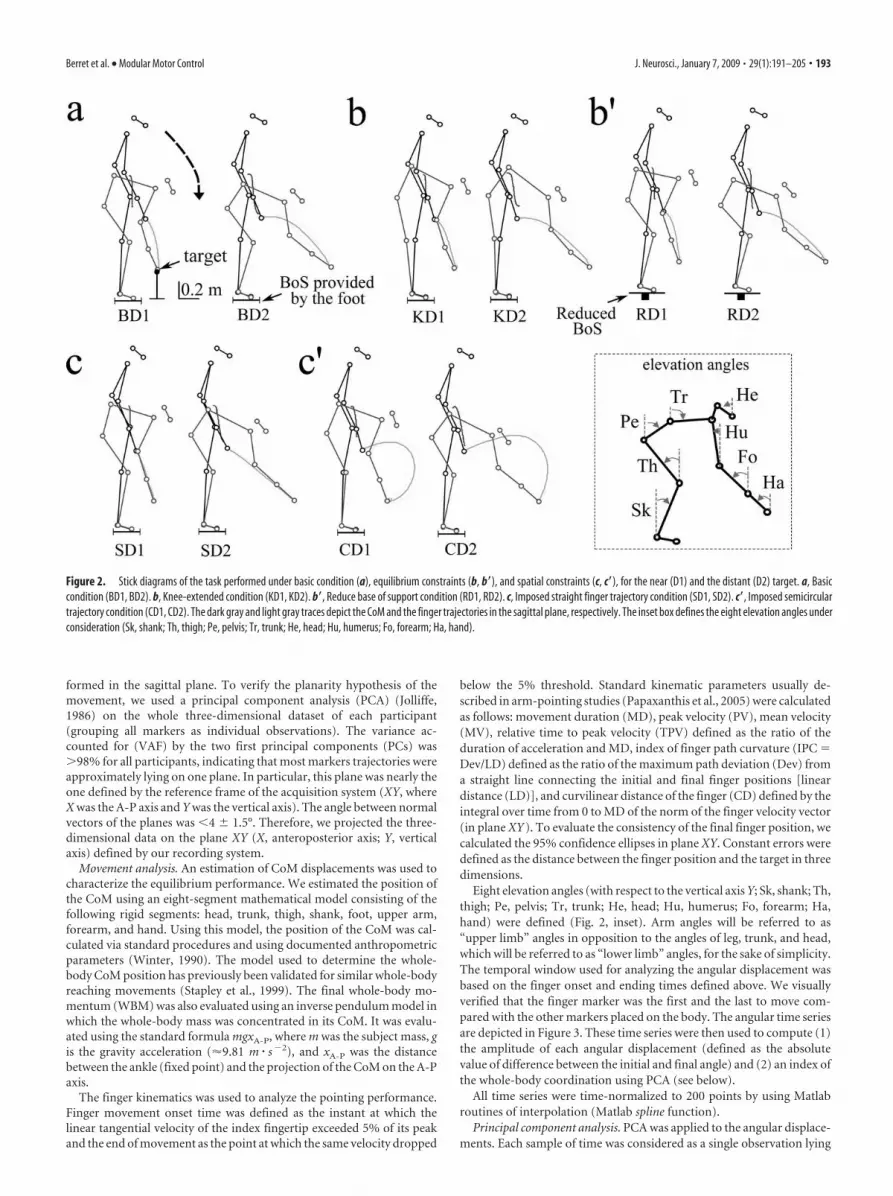
Figure 2. Stick diagrams of the task performed under basic condition (a), equilibrium constraints (b, b�), and spatial constraints (c, c�), for the near (D1) and the distant (D2) target. a, Basiccondition (BD1, BD2). b, Knee-extended condition (KD1, KD2). b�, Reduce base of support condition (RD1, RD2). c, Imposed straight finger trajectory condition (SD1, SD2). c�, Imposed semicirculartrajectory condition (CD1, CD2). The dark gray and light gray traces depict the CoM and the finger trajectories in the sagittal plane, respectively. The inset box defines the eight elevation angles underconsideration (Sk, shank; Th, thigh; Pe, pelvis; Tr, trunk; He, head; Hu, humerus; Fo, forearm; Ha, hand).
Berret et al. • Modular Motor Control J. Neurosci., January 7, 2009 • 29(1):191–205 • 193
in an ambient vector space whose dimension was given by the number oftime series included in the analysis.
For instance, consider a simple input dataset composed of eight col-umns (the angular displacements recorded during one trial) and 200rows (normalized time). PCA could be thought as a generalization of acorrelation analysis in a high dimensional space. In this case, PCA ex-tracted the commonality between the angular displacements, which wassometimes referred to as “waveforms” because it especially focused onthe shape of the angular time series.
Let � be the matrix of time-normalized angular displacements (200rows 8 columns). For each i �{1, . . . , 8}, the ith column of � (denotedby �i) was centered and normalized in amplitude using the formula �i �(�i � mi)/si, where mi and si were the mean and SD of �i, respectively,and thus, we got a new matrix �. Then, the covariance matrix of � wascomputed (i.e., the correlation matrix of �) and decomposed on thebasis of eigenvectors (by orthonormal diagonalization). We preferen-tially used the correlation matrix to take into account the different rangesof motion of each segment. After reordering the eigenvalues by decreas-ing order (and thus rearranging the corresponding eigenvectors), thematrix of eigenvectors was denoted by W � (wij)1 � i,j � 8. This matrix
contained the weighting coefficients, i.e., the loadings of PCA. Then, theprincipal components (denoted by , and referred to as PCs), whichrepresented the most common waveforms contained in the input dataset,
were defined as the following linear combination: � �W.For instance, the first PC (also referred to as PC1) was obtained by the
following formula:
1 � �i�1
i�8
wi1�i
[the first column of (i.e., a vector with 200 rows corresponding to thetime)].
When performing PCA, it was useful to examine the loadings to un-cover how angular displacements were captured by each PC. To this aim,we thus analyzed each column of W (i.e., each eigenvector) that defineda one-dimensional vector subspace (i.e., a certain direction in the eight-dimensional vector space). These eigenvectors represented a welladapted basis of the eight-dimensional vector space, characterizing themost important directions (in the sense of the VAF), and, in this setting,
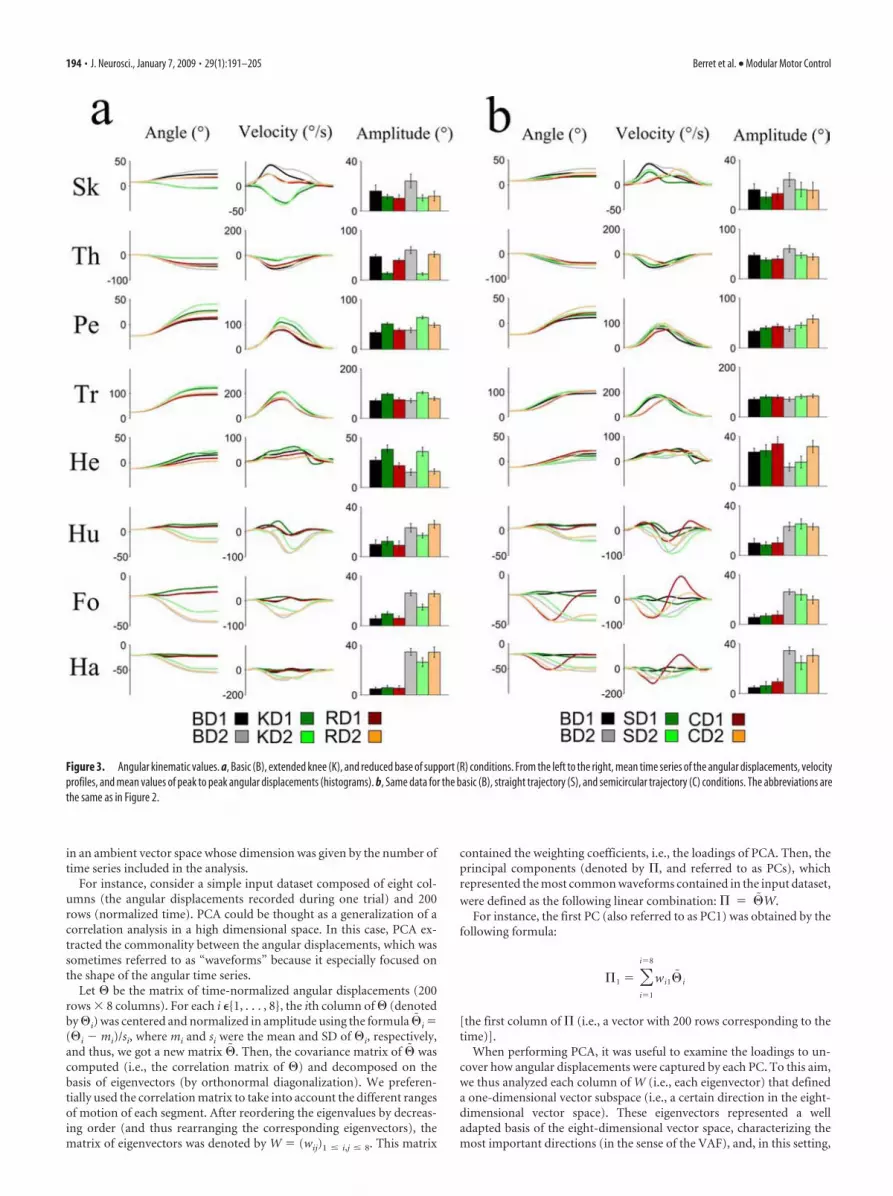
Figure 3. Angular kinematic values. a, Basic (B), extended knee (K), and reduced base of support (R) conditions. From the left to the right, mean time series of the angular displacements, velocityprofiles, and mean values of peak to peak angular displacements (histograms). b, Same data for the basic (B), straight trajectory (S), and semicircular trajectory (C) conditions. The abbreviations arethe same as in Figure 2.
194 • J. Neurosci., January 7, 2009 • 29(1):191–205 Berret et al. • Modular Motor Control

a principal component was simply the projection of the data onto asubspace spanned by a certain eigenvector. The ratio between the firsteigenvalue and the sum of all eigenvalues could be viewed as an index ofthe whole-body coordination (this value is commonly called the VAF bythe PC1 and is referred to as PC1%). A PC1% value equal to 100% meantthat the trajectory in the space of angles was a straight line (i.e., all angleswere linearly correlated together). However, a low PC1% value indicatedthat only one principal component could not describe precisely thewhole-body movement (i.e., the observations lay in a subspace whosedimension was �2, and thus, nonlinear relationships between anglesexisted). Therefore, we also reported the second PC whose VAF is de-noted by PC2%. Large PC2% values expressed that at least one angulardisplacement differed from the most common waveform given by PC1.
Here, instead of using 1560 independent PCAs (a PCA for each trial,each condition, and each participant), we used a PCA whose input data-set consisted of 800 or 960 columns (8 angles 10 trials 10 or 12participants) and 200 rows (normalized time) for each experimental con-dition. The analysis was exactly the same as the one depicted above,except that the number of column of the input dataset changed. In thismanner, the PCA automatically extracted the commonality between theshapes of the angular displacements [as in the study by Thomas et al.(2005)]. Nevertheless, individual PCA were also performed when neces-sary to seek intertrials adaptation, especially during the experiments withequilibrium or spatial constraints.
Reconstruction from PCs. A useful feature of PCA is the possibility toreconstruct an approximated whole-body movement from the data pro-jected on the vector subspace spanned by the first eigenvector (or bymore eigenvectors), using the inverse (or equivalently the transpose inour case) of the matrix W. Here, the reconstructed centered and normal-ized angular displacements were simply obtained by the following for-
mula: �Rec � W�1. For instance, the reconstruction from PC1 onlywas obtained by using the first column of and the first row of W �1 ofthe above formula. Then, the reconstructed angular displacements � Rec
were given, for each column i, by the following equation:
�iRec�si�i
Rec�mi. The same procedure was used to reconstruct move-ments from both PC1 and PC2 (by taking the appropriate rows andcolumns of and W �1, respectively). Notice that the reconstructedangular displacements � Rec were exactly equal to � when all PCs wereused. Otherwise, we just got an approximation of the original angulardisplacements.
Once the reconstructed angular waveforms were obtained, the posi-tion of markers in plane XY were then recalculated. To reconstruct themarkers kinematics, we assumed that the foot was fixed (which wasapproximately the case in practice). Head movement was reconstructedassuming that the trajectory of the auditory meatus marker was the onerecorded in practice. Notice that this approximation had no influence onthe finger trajectory and little influence on the CoM position.
Finally, the same analyses were performed using the covariance matrixrather than the correlation matrix. In particular, this did not change themain results presented below. Here, we considered that the correlationmatrix was better suited because angular displacements had significantdifferent amplitudes.
Correlation analyses. Covariation between angles was also estimated bymeans of a pairwise correlation analysis. Although loadings gave infor-mation concerning the joint coupling between pairs of angles (indeed, ifloadings were the same for two angles, one could expect that these angleswere highly correlated), a more direct method to analyze the couplingbetween pairs of angles was to compute the correlation coefficients be-tween each pair of angles. In the two-dimensional case (i.e., if we observeonly two angles), PC1% value was correlated to the coefficient of corre-lation. Thus, this analysis could be viewed as a local analysis comparedwith the global joint coupling given by the PC1% value. For a total ofeight angles, 28 correlations were computed on all pairs of angles, whichcorresponded to the correlation matrix coefficients analysis, for eachsingle movement. Statistically significant correlation coefficients ( p �0.05) were only retained by means of Matlab corrcoef function.
Numerical simulation. To test the feasibility of a strategy combining anangular covariation with the production of a desired finger trajectory in
the Cartesian space (S and C conditions), we conducted a numericalsimulation. To this aim, we hypothesized that the movement was optimalaccording to the minimum angle jerk criterion in the space of elevationangles (Wada et al., 2001). Under this assumption, the angles covariationwas perfect (i.e., the VAF by PC1 was exactly equal to 100%). This meantthat all body angles were driven by one single principal component or, inother words, that one single module ensured both spatial and equilib-rium subtasks. This was a plausible assumption according to previousresults (Alexandrov et al., 1998; Thomas et al., 2005), which indicatedthat a single PC captured similar whole-body reaching tasks. In such atask, there was an infinity of final postures allowing to put the fingertipon the target and to preserve equilibrium because there were 8 degrees offreedom. In this simulation, the initial posture was fixed and we used anumerical optimization procedure aimed at finding the final posturesuch that the resulting finger trajectory fitted at best to the imposed fingerpath in task space (either a straight line or a circular arc, as in the S and Cconditions). This was possible because, once a final posture was selected,the minimum angle jerk criterion allowed us to determine all limb dis-placements and, therefore, the finger path. The detailed procedure isgiven in Appendix.
Statistical analysis. All variables were normally distributed (Shapiro–Wilk’s W test) and their variance was equivalent (Levene’s test). Statisti-cal effects were tested by performing ANOVAs (repeated measures)when appropriate [either a 5 (B, K, R, S, C) 2 (D1, D2), a 3 (B, K, R) 2 (D1, D2), or a 3 (B, S, C) 2 (D1, D2) analysis]. Post hoc analyses wereconducted with Scheffe’s test and Student’s t tests were also used.
ResultsExperimental task achievementEndpoint accuracyEndpoint accuracy of the fingertip was first measured by meansof 95% confidence ellipse analysis, in the sagittal plane. The areaof the ellipses was equal to 6 � 2.5 cm 2 on average, and theirmajor and minor axes had similar lengths. The inspection of theareas of all individual ellipses across all experimental conditionsdid not reveal significant differences (area of ellipses; ANOVA,p � 0.82). Moreover, the mean endpoint error was 3 � 2 cm inthree dimensions, and the LD between the initial finger positionand the target were consistent among the different experimentalconditions ( p � 0.61) (Table 1).
Angular variabilityWe verified, for each participant and each angle, the consistencyof angular displacements, in each experimental condition. Themaximum intrasubject SD of amplitudes was �4°.
Movement durationWe found similar movement durations for all the experimentalconditions (ANOVA, p � 0.2 for all conditions), except for thesemicircular finger path condition in which the movement dura-tion was significantly longer (40%; ANOVA, p � 0.05) comparedwith the other experimental conditions (Table 1). This result islogical because the finger path imposed in this condition wasobviously longer. However, the average velocities recorded in thiscondition were similar to those of all the other conditions ( p �0.23) (Table 1).
Specific experimental constraintsAll participants succeeded in performing the constrained motortasks (extended-knee, K; reduced base of support, R; straighttrajectory, S; and semicircular trajectory, C).
Concerning the equilibrium constraints, as expected, the av-erage knee joint amplitudes recorded in the extended knee con-dition (5 � 2°) were significantly lower (t tests, p � 0.001, forboth distances) than those recorded during the basic motor task(B condition, 50 � 20°). Furthermore, the CoM position re-mained significantly higher (�0.1 m), compared with the basic
Berret et al. • Modular Motor Control J. Neurosci., January 7, 2009 • 29(1):191–205 • 195

motor task, when participants were re-quested to keep their knees extended, as aconsequence of the lack of knee flexion.The extended knee condition was charac-terized by an important forward trunkbending (Fig. 3, green histograms for Peand Tr). Moreover, all participants signif-icantly decreased the A-P CoM displace-ments in the reduced base of support con-dition (t tests, p � 0.01, for bothdistances). The average decrease was 25%for the short distance and 50% for the longdistance (Table 2, Fig. 4).
Concerning the spatial constraints, theindex of finger path curvature increasedsignificantly (t tests, p � 0.001) by �400%in the imposed semicircular trajectorycondition compared with the basic oneand remained concave in the sagittal plane(Table 1, Fig. 4). Analysis of index of fingerpath curvature in the imposed straight fin-ger path condition confirmed that all par-ticipants produced approximately straightpaths. Path curvature significantly de-creased by 75% compared with those mea-sured in the unconstrained condition (ttests, p � 0.001).
Finally, no statistical difference be-tween the two groups of participants thatperformed the basic motor task was found.
Effect of equilibrium constraintsFigure 2, a, b, and b�, qualitatively depictsthe initial and final whole-body configura-tions, as well as the CoM and finger trajec-tories, in the basic (B) and equilibriumconstraints conditions (K and R) for bothtarget distances.
Finger paths in the sagittal plane were strictly concave andapproximately similar between the different experimental condi-tions (ANOVA, B K R 2 distances; p � 0.52) (Table 1, Fig.4), although a slight decrease of the index of path curvature wasnoticed in the reduced base of support condition. Tangentialfinger velocity profiles were bell-shaped in these conditions, asdepicted in Figure 4. The velocity profiles recorded in the basicmotor task were strongly asymmetric: the acceleration durationwas shorter than the deceleration duration [average relative timeto peak of velocity (i.e., TPV value) was equal to 0.45 � 0.05 in thebasic condition] (Table 1). A significant difference was foundbetween the basic and equilibrium conditions (ANOVA, p �0.05), and a post hoc analysis showed that TPV values significantlytended to 0.5 in the extended knee condition ( p � 0.05), whereasthey remained invariant in the reduced base of support condition( p � 0.57).
Forward CoM displacements in the basic and extended kneeconditions were �40 and 70% of the foot length for the short andlong distances, respectively. In contrast, as already noticed, theparticipants limited the forward A-P CoM displacement [�4 cm(i.e., �30% of the foot length)] in the reduced base of supportcondition (R), because of the experimental restriction; significantstatistical differences ( p � 0.01) were obtained when comparingthe reduced base of support condition with the basic and ex-tended knee conditions. Despite these modifications in ampli-
tude, CoM path was task independent and kept the same charac-teristic two components: an initial curvilinear and goal-orientedforward displacement followed by a straight downward move-ment (Stapley et al., 1999; Pozzo et al., 2002). Downward dis-placements of CoM were larger in the basic and reduced base ofsupport conditions compared with the knee extended condition( p � 0.01) (Table 2). Notice that forward and downward dis-placements of the CoM significantly increased for the long dis-tance ( p � 0.01) as a consequence of a greater contribution ofshank, pelvis, and humerus segments, necessary to reach the tar-get (Fig. 3, histograms). The velocity profiles of the CoM weresingle-peaked in all conditions (Fig. 4) and exhibited the sametrend as finger TPV values.
Interestingly, participants were able to keep their equilibriumin both conditions. The changes in angular displacements werenot detrimental to the covariation of elevation angles. Indeed,PCA showed that the VAF by PC1 was constantly �85% for allthe conditions ( p � 0.45) (Fig. 5a, black histograms), which iscomparable with the values calculated in previous similar motortasks (Alexandrov et al., 1998; Thomas et al., 2005). The PC1%value increased significantly (�3%) at the long distance com-pared with the short distance ( p � 0.01). Moreover, the principalcomponent waveforms did not change significantly as reflectedby the large correlation coefficients computed when comparingPC1 and PC2 between conditions (Fig. 5b). The PC1 captured themonotonic angular displacements with bell-shaped velocity pro-
Table 1. Mean values and SD of finger kinematics parameters for both short (D1) and long (D2) distances, and inall experimental conditions (basic, knee extended, reduced base of support, straight, and semicircular fingerpaths)
B K R S C
D1PV (m � s�1) 1.51 � 0.30 1.36 � 0.19 1.41 � 0.31 1.36 � 0.22 1.49 � 0.16TPV 0.46 � 0.04 0.50 � 0.04 0.47 � 0.03 0.49 � 0.04 0.62 � 0.05MV (m � s�1) 0.79 � 0.15 0.68 � 0.08 0.74 � 0.13 0.72 � 0.12 0.79 � 0.24MD (s) 0.64 � 0.13 0.71 � 0.11 0.70 � 0.14 0.70 � 0.12 0.96 � 0.13CD (m) 0.49 � 0.04 0.48 � 0.04 0.50 � 0.05 0.48 � 0.04 0.76 � 0.09LD (m) 0.45 � 0.03 0.44 � 0.03 0.46 � 0.04 0.47 � 0.04 0.47 � 0.03IPC 0.12 � 0.05 0.11 � 0.05 0.10 � 0.04 0.03 � 0.01 0.44 � 0.11
D2PV (m � s�1) 2.02 � 0.39 1.74 � 0.23 1.84 � 0.30 1.91 � 0.34 1.92 � 0.52TPV 0.44 � 0.06 0.49 � 0.04 0.44 � 0.06 0.49 � 0.03 0.57 � 0.15MV (m � s�1) 1.09 � 0.17 0.96 � 0.11 1.00 � 0.12 1.02 � 0.20 1.18 � 0.29MD (s) 0.71 � 0.13 0.79 � 0.14 0.75 � 0.10 0.71 � 0.09 1.01 � 0.19CD (m) 0.76 � 0.05 0.76 � 0.05 0.75 � 0.06 0.73 � 0.04 1.18 � 0.20LD (m) 0.71 � 0.04 0.71 � 0.04 0.70 � 0.05 0.72 � 0.04 0.73 � 0.04IPC 0.11 � 0.05 0.11 � 0.06 0.10 � 0.05 0.03 � 0.01 0.49 � 0.21
The parameters included are as follows: peak velocity (PV), relative time to peak velocity (TPV), mean velocity (MV), movement duration (MD), curvilineardistance (CD), linear distance (LD), and index of path curvature (IPC).
Table 2. CoM displacements along A-P and vertical axes and final WBM
B K R S C
D1A-P (m) 0.05 � 0.03 0.04 � 0.02 0.03 � 0.01 0.05 � 0.02 0.06 � 0.04Vertical (m) 0.30 � 0.03 0.24 � 0.01 0.28 � 0.02 0.26 � 0.03 0.29 � 0.04TPV 0.45 � 0.03 0.47 � 0.04 0.46 � 0.04 0.47 � 0.04 0.53 � 0.06WBM (N � m) 31 � 20 29 � 7 23 � 13 31 � 20 38 � 27
D2A-P (m) 0.08 � 0.02 0.09 � 0.02 0.04 � 0.01 0.10 � 0.02 0.11 � 0.03Vertical (m) 0.42 � 0.05 0.28 � 0.02 0.39 � 0.05 0.34 � 0.05 0.38 � 0.06TPV 0.45 � 0.04 0.50 � 0.03 0.45 � 0.04 0.50 � 0.03 0.55 � 0.05WBM (N � m) 52 � 13 57 � 13 27 � 8 63 � 12 72 � 19
196 • J. Neurosci., January 7, 2009 • 29(1):191–205 Berret et al. • Modular Motor Control

files, whereas the second PC captured nonmonotonic angulardisplacements with double-peaked speed profiles (Fig. 3a,b, mid-dle column). In addition, an inspection of the loadings did notreveal significant changes in the reduced base of support condi-tion for both distances (average r 2 � 0.97). At variance, in theextended knee condition, the angles were differently distributedamong PC1 and PC2, and showed changes mainly in the legangles (e.g., shank and thigh), because of the joint restrictionimposed at the knee (Fig. 5c). However, the PC1% value re-mained high despite these changes.
Effect of spatial constraintsFigure 2, c and c�, qualitatively illustrates the initial and finalwhole-body configurations, as well as the CoM and finger trajec-tories, under spatial constraints (S and C) for both targetdistances.
As already noticed, participants performed approximatelystraight and semicircular finger paths in these conditions. Thevelocity time courses tended to become more symmetric in theimposed straight finger trajectory condition (TPV value was onaverage 0.49 � 0.04; p � 0.06), being close to what would bepredicted by a minimum Cartesian jerk model (straight trajec-tory and symmetric speed profile) (Flash and Hogan, 1985). Incontrast, the velocity profiles recorded in the semicircular condi-tion were double-peaked (Table 1, Fig. 4) similar to those ob-tained during via point movement (Viviani and Flash, 1995).
The analysis of CoM displacements revealed significantchanges across the experimental conditions (Table 2, Fig. 4). Spe-cifically, in the imposed finger trajectory conditions, A-P andvertical CoM displacements increased (�22%; p � 0.05) anddecreased (�12%; p � 0.01), respectively, compared with thebasic condition. These changes led the participants to adopt a
more risky inverted pendulum configuration because of (1) theA-P CoM displacement close to the forward limit of the BoS and(2) a longer level arm inducing a larger potential energy and agreater whole-body momentum (approximately �22%) (Table2) compared with the basic experiment. In contrast to fingertrajectory, the CoM velocity remained single-peaked in Ccondition.
Remarkably, the covariation of elevation angles was stronglyaffected by spatial constraints onto the finger path. The statisticalanalysis revealed a significant decrease of the PC1% value in theseconditions (�8 and �13%, respectively; p � 0.01) comparedwith the basic condition. This was compensated by a drastic in-crease of the PC2% value that was about twice larger than thevalues computed in the basic condition (Fig. 5, gray histograms).Similarly to the basic motor task, the covariation of elevationangles was greater (�6%) for the long distance compared withthe short distance ( p � 0.05) in the motor tasks with spatialconstraints. The inspection of principal components revealedthat their waveforms were unchanged compared with the basiccondition (Fig. 5b, large correlation coefficients). Nevertheless,the loadings analysis revealed changes in their distributionamong the PC1 and especially the PC2. A noticeable effect wasan increase (in absolute value) of the loadings for the armangles (humerus, forearm, and hand): the upper limb angleswere more strongly captured by the PC2 in the imposed fingertrajectory conditions compared with the basic one. This resultwas quite striking because these changes were accompanied bya significant decrease and increase of the PC1% and PC2%values, respectively. Thus, spatial but not equilibrium con-straints modified significantly the relative importance of PC1and PC2.
In particular, an interesting result was the decrease of the
Figure 4. Average CoM and finger trajectories and tangential velocity profiles, in the sagittal plane, for all experimental conditions: basic (B), extended knee (K), reduced base of support (R),straight finger trajectory (S), and semicircular finger trajectory (C). SDs are not depicted for the sake of clarity.
Berret et al. • Modular Motor Control J. Neurosci., January 7, 2009 • 29(1):191–205 • 197
PC1% and the increase of PC2% values(14 of 14 participants for the short dis-tance; 12 of 14 participants for the longdistance) consecutively to the straight fin-ger path constraint. To evaluate a possibleeffect of a learning process on this changein the covariation of elevation angles, weasked 4 among the 14 participants to per-form 50 successive trials in the straightCartesian trajectory condition for the longdistance. To avoid the potential influenceof fatigue, participants took a rest of 1 minafter each block of 10 trials (Schmid et al.,2006). Figure 6 shows average results forthe PC1% value. A linear regression anal-ysis on the average PC1% values amongthe 50 repetitions showed slope and r 2 co-efficients �10�3 indicating a lack of adap-tation among the 50 trials in the imposedstraight finger trajectory condition, for thelong distance (SD2). Furthermore, abetween-participants t test comparisonamong the PC1% average values of the 10first and 10 last trials did not give signifi-cant differences ( p � 0.29).
Motion reconstruction from PCsTo better elucidate the respective roles ofthe PC1 and PC2 on the movement, wereconstructed the angular displacementsand then the kinematic features of thebody markers, from either the first or boththe first and the second principal compo-nents (see Materials and Methods).
First, the whole movement was recon-structed based on the data contained in thefirst principal component. Figure 7 showsthat the reconstructed movement was verysimilar to the one recorded in the basiccondition (B), for a typical participant.The characteristic features of the basicmotor task were captured well by thePC1 (e.g., the forward CoM projectionand the concave finger trajectory) (Table3) (Pozzo et al., 2002). In particular, theCoM and finger trajectories were close tothe recorded ones (Fig. 7a), the angulardisplacements were also similar as indi-cated by the large correlation coeffi-cients (Fig. 7b), and the velocity profilesof both the finger and the CoM weresingle-peaked (Fig. 7c). Second, whenthe motion was reconstructed using thedata contained in the PC1 and PC2, thewhole-body movements became ex-tremely close to the recorded ones, whatever the experimentalconditions (as proved by the high correlation coefficients re-ported in Fig. 7b,c).
However, the main result lies in the differences observed whenreconstructing motions from PC1 in the motor tasks with spatialconstraints imposed on finger trajectory. Movements recon-structed in these conditions showed that the requested Cartesianconstraints were not captured by the PC1 (i.e., the straight and
semicircular finger paths constraints were not encoded in thePC1) (Fig. 7a, second and third columns). Table 3 shows, onspecific parameters, that the A-P CoM displacement was wellreconstructed by the use of PC1 in all experimental conditions, incontrast to the finger trajectory. Indeed, the index of finger pathcurvature did not fit well with those recorded experimentally (or,equivalently, obtained when reconstructing the motion from allPCs). Whatever the condition, the reconstructed finger path was
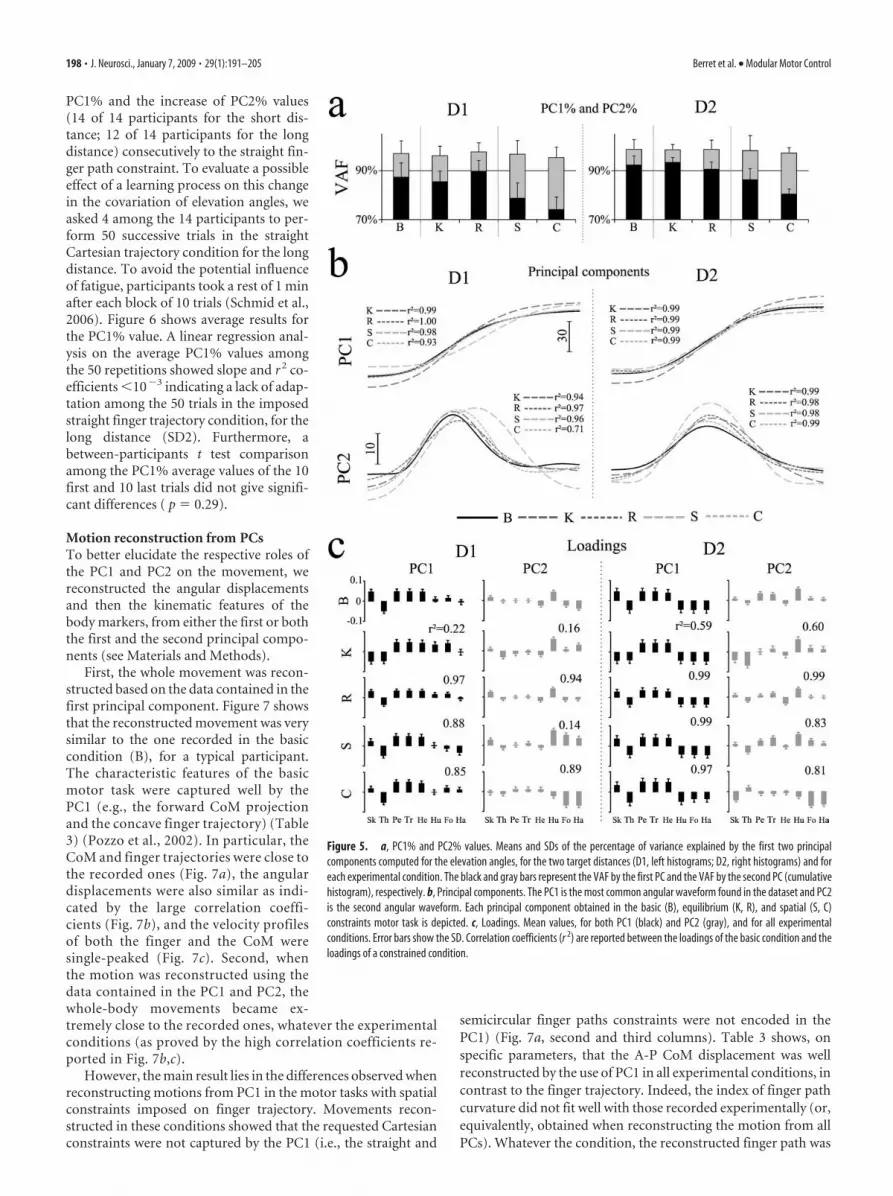
Figure 5. a, PC1% and PC2% values. Means and SDs of the percentage of variance explained by the first two principalcomponents computed for the elevation angles, for the two target distances (D1, left histograms; D2, right histograms) and foreach experimental condition. The black and gray bars represent the VAF by the first PC and the VAF by the second PC (cumulativehistogram), respectively. b, Principal components. The PC1 is the most common angular waveform found in the dataset and PC2is the second angular waveform. Each principal component obtained in the basic (B), equilibrium (K, R), and spatial (S, C)constraints motor task is depicted. c, Loadings. Mean values, for both PC1 (black) and PC2 (gray), and for all experimentalconditions. Error bars show the SD. Correlation coefficients (r 2) are reported between the loadings of the basic condition and theloadings of a constrained condition.
198 • J. Neurosci., January 7, 2009 • 29(1):191–205 Berret et al. • Modular Motor Control

constantly slightly concave and, in particular, did not fit with thespatial constraints. The addition of PC2 in the reconstructionprocess was necessary to recover the spatial requirements im-posed to the motor task (Table 3). Therefore, the PC2 appeared tobe strongly implied in a fine control of the fingertip path, whereasthe PC1 encoded the main characteristics of the whole-body mo-tion performed in the basic condition, including the forwardCoM displacement and the slightly concave finger path. Correla-tion analyses between the recorded and reconstructed angulardisplacements were also performed to quantify in which propor-tion each angular displacement was reconstructed. Lower corre-lation coefficients were measured between angular displacementsfor the angles of upper limbs, especially in the semicircular con-dition. This result suggests that PC1 primarily described the equi-librium and free reaching subtasks, whereas PC2 (for whichPC2% values increased significantly in the spatial constraintsconditions) adjusted the finger trajectory to the task requirement(S and C conditions). Such a reorganization was mainly achievedby means of the hand, forearm, and humerus angular displace-ments. These findings were consistent for both distances andacross participants, even for those who maintained a relativelyhigh joint coupling during the straight finger trajectorycondition.
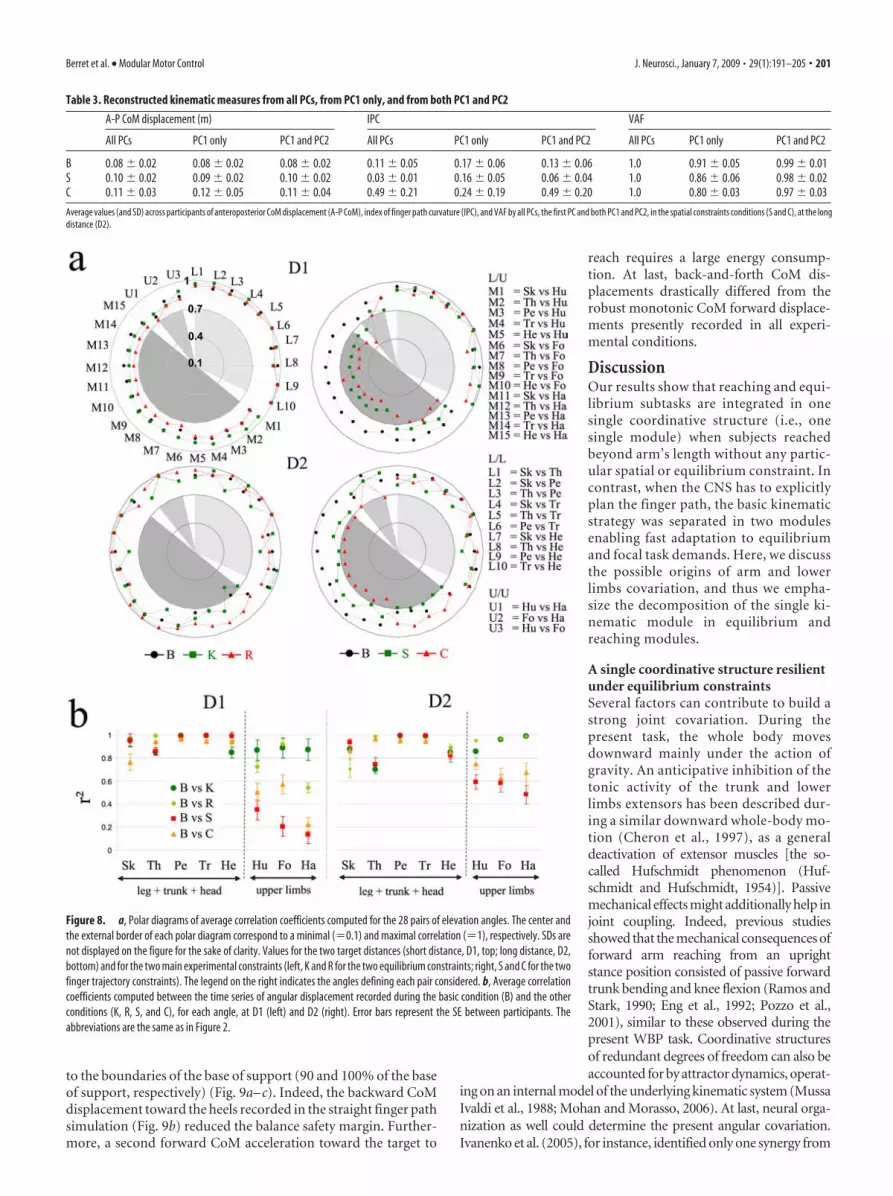
Local couplingTo uncover specific angles covariations at different levels of thewhole body, we performed a correlation analysis between eachpair of angles (see Materials and Methods) (Fig. 8a). Becauseeight angles were measured, a total of 28 correlation coefficientswere analyzed. In most cases, the results showed that all correla-tion coefficients were �0.7 in the basic and equilibrium con-straint motor tasks (see top left polar graph), as expected from theresults given by PCA in these conditions (Fig. 5). An ANOVAperformed on the 28 coefficients (28 factors), for each conditionseparately, showed a significant main effect ( p � 0.001) (i.e., atleast two correlation coefficients were significantly differentamong the 28 comparisons). Despite the fact that all correlationcoefficients were �0.7, it was noticeable that, when calculatingcorrelations between angles of the upper limbs (U) with angles ofthe lower limbs (L), the correlation coefficients were slightlylower (Fig. 8, mixed pairs of angles, L/U, M1 to M15) than thecoefficients corresponding to L1 to L10 (L/L, pairs of angleswithin the lower limbs group). Nevertheless, a post hoc analysisdid not reveal important differences (in general, p � 0.05) for thebasic and equilibrium constraints conditions.
At variance, in the spatial constraints conditions (Fig. 8a, topright polar graph), the correlation coefficients significantly de-creased for the mixed pairs of angles (L/U, M1 to M15, �0.7).
Post hoc analysis proved that all coeffi-cients of this group (L/U, M1 to M15) weresignificantly different from both the upperlimbs (U/U, U1 to U3) and the lower limbs(L/L, L1 to L10) comparisons ( p � 0.05 atD1), illustrating a change in the whole-body motor strategy (a significant separa-tion between upper and lower limbs). Forthe long distance (Fig. 8a, bottom rightpart), correlation coefficient values in-creased compared with the short distancecondition ( p � 0.01), indicating a stron-ger coupling of upper and lower limbswhen the task required a fine equilibriumcontrol (long distance, D2). It should be
noted that the decrease of the covariation of elevation angles,observed in the straight (S) and semicircular (C), was less pro-nounced at D2 compared with the short distance (D1), remain-ing, however, below the values obtained in the basic and equilib-rium constraints motor tasks (Fig. 8a, left).
To verify the idea of a dissociation of reaching limbs (armsegments) from a robust equilibrium module during spatial con-straint, we computed the average correlation coefficients betweenthe waveforms measured in the basic motor task (B) and theother experimental conditions, for each of the eight angles (Fig.8b). In this way, we quantified the changes in the angular dis-placements induced by the experimental conditions with respectto the basic condition. A repeated-measure ANOVA showed thatthese changes were significantly concentrated in the hand, fore-arm, and humerus angles compared with the leg, trunk, and headangles ( p � 0.001 and values of r 2 � 0.92 and 0.72 for lower limbsand upper limbs, respectively). Thus, in addition to the drasticreorganization of upper limbs in response to the spatial con-straints, the lower limbs displacements remained invariant (i.e.,highly correlated to the ones observed in the basic condition).
Overall, these analyses revealed one PC that captured lowerlimbs and CoM displacements and resisted to spatial constraintsexerted on the finger, and another one, task-dependent, thatemerged when focal subtask became a priority.
SimulationThe previous results showed that, in the basic condition, equilib-rium and focal subtasks were integrated in the same module fromwhich emerged an additional module when specific finger pathhad to be planned. Because priority was given to finger trajectorywithout requirement concerning the joint strategy to be used(i.e., one could have asked the participants to produce a straightfinger path and to move all body segments at the same time), thedissociation of equilibrium and focal task goals in two modulescould be inherent to the spatial constraints. Thus, to test theplausibility of one single module to generate simultaneously astraight path in angle space and a specific finger trajectory inCartesian space, we performed a simulation (for details, see Ma-terials and Methods and Appendix).
Therefore, we simulated the integration of both the equilib-rium and reaching task objectives in one single module duringwhole-body pointing (WBP) under spatial restriction. The resultof this simulation showed that the production of a perfectlystraight trajectory in the elevation angles space, combined withan almost straight or semicircular finger path in Cartesian space is(1) possible with respect to the physiological ranges of joint an-gular displacement but (2) detrimental to equilibrium control. Inparticular, the spatial constraints led to a CoM displacement close
Figure 6. Typical PC1% values of one typical subject for 50 successive trials. The continuous black line corresponds to the PC1%baseline measured during the basic condition. The dashed line indicates the linear regression calculated on the 50 recordsperformed in the straight finger trajectory condition at long distance (SD2).
Berret et al. • Modular Motor Control J. Neurosci., January 7, 2009 • 29(1):191–205 • 199
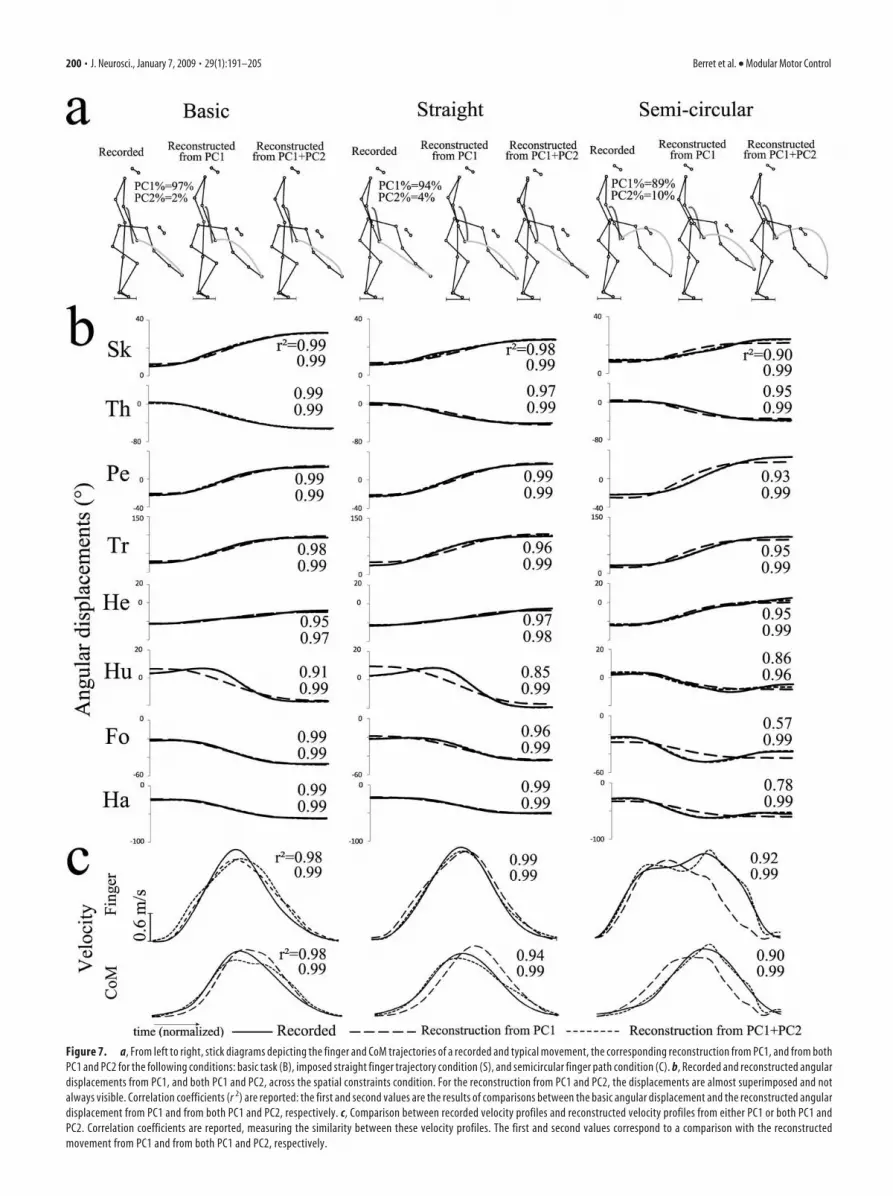
Figure 7. a, From left to right, stick diagrams depicting the finger and CoM trajectories of a recorded and typical movement, the corresponding reconstruction from PC1, and from bothPC1 and PC2 for the following conditions: basic task (B), imposed straight finger trajectory condition (S), and semicircular finger path condition (C). b, Recorded and reconstructed angulardisplacements from PC1, and both PC1 and PC2, across the spatial constraints condition. For the reconstruction from PC1 and PC2, the displacements are almost superimposed and notalways visible. Correlation coefficients (r 2) are reported: the first and second values are the results of comparisons between the basic angular displacement and the reconstructed angulardisplacement from PC1 and from both PC1 and PC2, respectively. c, Comparison between recorded velocity profiles and reconstructed velocity profiles from either PC1 or both PC1 andPC2. Correlation coefficients are reported, measuring the similarity between these velocity profiles. The first and second values correspond to a comparison with the reconstructedmovement from PC1 and from both PC1 and PC2, respectively.
200 • J. Neurosci., January 7, 2009 • 29(1):191–205 Berret et al. • Modular Motor Control
to the boundaries of the base of support (90 and 100% of the baseof support, respectively) (Fig. 9a– c). Indeed, the backward CoMdisplacement toward the heels recorded in the straight finger pathsimulation (Fig. 9b) reduced the balance safety margin. Further-more, a second forward CoM acceleration toward the target to
reach requires a large energy consump-tion. At last, back-and-forth CoM dis-placements drastically differed from therobust monotonic CoM forward displace-ments presently recorded in all experi-mental conditions.
DiscussionOur results show that reaching and equi-librium subtasks are integrated in onesingle coordinative structure (i.e., onesingle module) when subjects reachedbeyond arm’s length without any partic-ular spatial or equilibrium constraint. Incontrast, when the CNS has to explicitlyplan the finger path, the basic kinematicstrategy was separated in two modulesenabling fast adaptation to equilibriumand focal task demands. Here, we discussthe possible origins of arm and lowerlimbs covariation, and thus we empha-size the decomposition of the single ki-nematic module in equilibrium andreaching modules.
A single coordinative structure resilientunder equilibrium constraintsSeveral factors can contribute to build astrong joint covariation. During thepresent task, the whole body movesdownward mainly under the action ofgravity. An anticipative inhibition of thetonic activity of the trunk and lowerlimbs extensors has been described dur-ing a similar downward whole-body mo-tion (Cheron et al., 1997), as a generaldeactivation of extensor muscles [the so-called Hufschmidt phenomenon (Huf-schmidt and Hufschmidt, 1954)]. Passivemechanical effects might additionally help injoint coupling. Indeed, previous studiesshowed that the mechanical consequences offorward arm reaching from an uprightstance position consisted of passive forwardtrunk bending and knee flexion (Ramos andStark, 1990; Eng et al., 1992; Pozzo et al.,2001), similar to these observed during thepresent WBP task. Coordinative structuresof redundant degrees of freedom can also beaccounted for by attractor dynamics, operat-
ing on an internal model of the underlying kinematic system (MussaIvaldi et al., 1988; Mohan and Morasso, 2006). At last, neural orga-nization as well could determine the present angular covariation.Ivanenko et al. (2005), for instance, identified only one synergy from
Table 3. Reconstructed kinematic measures from all PCs, from PC1 only, and from both PC1 and PC2
A-P CoM displacement (m) IPC VAF
All PCs PC1 only PC1 and PC2 All PCs PC1 only PC1 and PC2 All PCs PC1 only PC1 and PC2
B 0.08 � 0.02 0.08 � 0.02 0.08 � 0.02 0.11 � 0.05 0.17 � 0.06 0.13 � 0.06 1.0 0.91 � 0.05 0.99 � 0.01S 0.10 � 0.02 0.09 � 0.02 0.10 � 0.02 0.03 � 0.01 0.16 � 0.05 0.06 � 0.04 1.0 0.86 � 0.06 0.98 � 0.02C 0.11 � 0.03 0.12 � 0.05 0.11 � 0.04 0.49 � 0.21 0.24 � 0.19 0.49 � 0.20 1.0 0.80 � 0.03 0.97 � 0.03
Average values (and SD) across participants of anteroposterior CoM displacement (A-P CoM), index of finger path curvature (IPC), and VAF by all PCs, the first PC and both PC1 and PC2, in the spatial constraints conditions (S and C), at the longdistance (D2).
Figure 8. a, Polar diagrams of average correlation coefficients computed for the 28 pairs of elevation angles. The center andthe external border of each polar diagram correspond to a minimal (�0.1) and maximal correlation (�1), respectively. SDs arenot displayed on the figure for the sake of clarity. Values for the two target distances (short distance, D1, top; long distance, D2,bottom) and for the two main experimental constraints (left, K and R for the two equilibrium constraints; right, S and C for the twofinger trajectory constraints). The legend on the right indicates the angles defining each pair considered. b, Average correlationcoefficients computed between the time series of angular displacement recorded during the basic condition (B) and the otherconditions (K, R, S, and C), for each angle, at D1 (left) and D2 (right). Error bars represent the SE between participants. Theabbreviations are the same as in Figure 2.
Berret et al. • Modular Motor Control J. Neurosci., January 7, 2009 • 29(1):191–205 • 201

the recorded activity of 32 muscles during a stoop in place task.Moreover, it has been shown that trunk bending (as an importantaspect of the present task) was under the control of “axial synergies”(i.e., a coordinated and opposing displacements of trunk and lowerbody segments) (Babinski, 1899; Alexandrov et al., 1998, 2001b).The present joint coupling is compatible with the functional mech-anism previously proposed by which descending neural commandrelated to the desired CoM trajectories and final finger position aretransformed into a specific joint angle covariation (Ivanenko et al.,2005; d’Avella et al., 2006; Ting, 2007). For instance, when kneeextension was imposed (K condition), the pointing was mainly at-tributable to forward trunk bending to compensate for the lack ofknee flexion. This new joint configuration induced a higher level ofthe position of the CoM as a consequence of the lack of knee flexion,and represented an additional source of postural instability. Surpris-ingly, these changes did not affect joint coupling compared with thebasic condition (B), revealing a significant flexibility of the motorcommand to keep joint covariation and consecutive similar fingerand CoM trajectories in the sagittal plane. Furthermore, whenplaced on a thin support (R condition), subjects still exhibited strongcovariation of elevation angles. Thus, the well preserved joint co-variation, the associated stereotypical CoM, and finger trajectoriesare compatible with a process that would map low-dimensional taskvariables (the final finger position and the CoM trajectory) ontohigh-dimensional anatomical details (individual limbs) (Ting,2007). In the same vein, covariation of limb rotation in extrinsicspace points to a simple organization previously identified for thelower limbs (Lacquaniti et al., 1999) or the head segment (Pozzo etal., 1990) during locomotion that both reduces the high dimension-ality of the kinematic chain and allows the control of the CoM dis-placement with respect to gravity. This strategy likely facilitates theintegration of reaching and equilibrium subtasks in the same motorplan. Furthermore, because of the mechanical link existing betweenupper and lower body parts, separate controllers seem not function-ally appropriate to predict and compensate the interaction torquesinherent to multijoint movements. Pigeon et al. (2003) also demon-strated that, during turn and reach movement involving the trunk,
the different body parts are integrated in a single frame of referenceto predict appropriate joint torque and prevent disturbing effect onhand path. Finally, the strong temporal coupling between times topeak velocity of the finger and the whole-body center of mass, re-gardless of drastic change in sensory and mechanical context (earthversus microgravity) previously described for the same task (Patronet al., 2005) (Fig. 7; Tables 1, 2), support the idea that the two taskobjectives may to some degree share common neural commands.
Decomposition of the coordinative structure in two modulesunder spatial constraintsAn important result is that, when subjects were forced to movethe finger along a straight (S condition) or a semicircular (Ccondition) path, a second coordinative structure dedicated toarm segment coordination emerged from the initial kinematicstrategy. For the exaggerated C condition, one could have ex-pected such a dissociation because of the large discrepancybetween natural and imposed finger trajectories. In contrast, itwas more unlikely for the S condition that imposed a straightline close to the basic slightly curved finger path. This resultdemonstrates that the curved finger path recorded in the freecondition was not the result of an error in trajectory planning.Thus, if desired, participants could execute the WBP task byadopting another reaching movement, a straight hand path forinstance that is not the one chosen in the free condition.
In S and C conditions, priority was given to the control ofhand movement, as during object manipulation or objectavoidance while reaching performed from a standing posture.Thus, inefficiency in controlling the finger path and the cur-rent highly redundant body joint system at the same timerather than a voluntary strategy adopted by the participants,could explain arm and lower limbs decoupling. Several argu-ments contradict with such a hypothesis. First, the unstablehigh inverted pendulum posture adopted by the participantswhen spatial restrictions were present required an efficientequilibrium control. Thus, a precise prediction of the mechan-ical effect of the CoM acceleration must be incorporated in the
Figure 9. Recorded data. a, Typical stick diagram of a whole-body pointing task performed in the basic condition, at long distance (BD2). The black and gray traces depict the CoM and the fingertrajectory in the sagittal plane, respectively. Simulated data. b, Motor strategy obtained by imposing both a perfect covariation of the elevation angles and a straight finger path. c, Motor strategyobtained by imposing both a perfect covariation of the elevation angles and an exaggerated curved finger path. d, Angular displacements of elevation angles obtained for the (dark line) recordeddata, and simulated data of the straight (S) and semicircular (C) conditions (dotted line). The abbreviations are the same as in Figure 2.
202 • J. Neurosci., January 7, 2009 • 29(1):191–205 Berret et al. • Modular Motor Control

inverse model of the motor plan to produce a continuoussmooth and linear finger path. Because participants succeededafter only two or three trials, without increasing movementduration compared with the free condition or any adaptationafter several repetitions (Fig. 6), we conclude that kinematicdecoupling was actively performed. Second, simulation pro-cedures have clearly shown that the two subtasks could beintegrated in the same motor plan with only few changes inexperimentally recorded body geometry, supporting the bio-mechanical feasibility of keeping both joint covariation andstraight finger path. However, CoM analysis revealed that sucha strategy was detrimental to equilibrium control, a likely rea-son for the CNS not to adopt this way. Additionally, this pos-tural solution contrasts with the stereotyped trajectory re-corded whatever the condition (Fig. 4) and characterized by aninitial forward CoM acceleration. The robustness of this pat-tern, existing among a wide range of voluntary movements(Crenna and Frigo, 1991; Stapley et al., 1998), could representa neural constraint contributing to preserve the equilibriummodule invariant when reaching under finger path restriction.Consequently, rather than revealing inefficient coordinationof the multijoint system because of the experimental condi-tion, we propose that the dissociation of the basic coordinativestructure in two modules would correspond to an intendedstrategy.
Finally, the spatial constraints did not destroy the localcoupling of lower segments indicating that postural modulewas task independent. Instead, a new module used to producean optional finger trajectory (S or C) was added up to themodule found in the basic whole-body reaching condition aspredicted in the top right part of Figure 1.
The present addition of a new kinematic module to anexisting one is consistent with the proposal that compoundmovements are generated by means of a combination of motorprograms (Bizzi et al., 2008). Similarly, other studies havedemonstrated modularity of muscle use in cat (Ting andMacpherson, 2005; Krouchev et al., 2006), in frogs (Hart andGiszter, 2004; d’Avella and Bizzi, 2005), and in humans (Iv-anenko et al., 2005; d’Avella et al., 2006). Basic synergies builtduring development are probably slowly adjusted and differslightly their expression to tolerate a wide range of tasks in-ducing various CoM and endpoint trajectories. However, inan unusual condition, it seems that they can adapt very rapidlyor can be completed with additional ones to provide morebehavioral flexibility. At which stage of the CNS the motorsynergies are adapted to environmental demands or combinedremains an open question. Even far from the scope of thisstudy, we would like to discuss briefly some points accordingto the literature. Capaday (2004) has emphasized the func-tional linking of motor cortical points based on disinhibitionprocess. Thus, synergies could emerge from the recruitment ofvarious motor cortical points by selected excitation as well asby selected release from inhibition. At the level of the spinalcord, central descending pathways could influence and mod-ulate reflex pathways depending on the context within whichwe must act (for review, see Dietz, 1992). Various descendingpathways can then modulate the spinal cord for greater flexi-bility of basic reflexes. For instance, recent data recorded in catshowed that neurons in pontomedullary reticular formationcan both ensure integrated or independent control of postureand reaching (Schepens and Drew, 2004; Schepens et al.,2008). Together, one can speculate that the basic coordinativestructure could first emerge at the level of M1 depending on
task demands. To provide flexibility, the motor commandscould be adapted at the level of spinal cord via excitation orinhibition processes channeling appropriate contexts to thekinematic strategy and thus constructing the reaching andequilibrium subtasks by using one or two modules. Neverthe-less, without a detailed analysis of patterns of focal and pos-tural muscle activity, definite conclusions relating to the orga-nization of descending neural commands are difficult tomake. An upcoming study of muscle activation patternsshould reveal how many muscle synergies are required for taskperformance and their robustness to different contexts.
AppendixThis appendix gives details concerning the kinematic simulationthat we briefly described in Materials and Methods.
Let us consider a rigid nine-segment model of the body anddefine eight elevation angles (Fig. 2, inset) as follows: �Sk,�Th,�Pe,�Tr,�He,�Hu,�Fo,�Ha)��.The foot angle was neglected because ankle marker coordinatesremained constant.
First, we assumed that the trajectory in angle space resultedfrom the minimum angle jerk criterion, which minimized thefollowing integral cost (Wada et al., 2001):
1
2�0
MD�k�1
k�8�d3�k
dt3 �2
dt,
where �k was the kth component of �, and t denoted the time.In the following, the movement duration was normalized
(MD � 1). It is well known that the optimal solutions are straightlines in the eight-dimensional vector space connecting the initialconfiguration of segments (�i) to the final one (�f) with thefollowing formula:
� t� � �i � t3 10 � 15t � 6t2� �f � �i�. (1)
Notice that, therefore, the PC1% value was equal to 100%.Then, the finger trajectory was given by an equation of the
form f(x, y) � 0. Because the initial position (x0, y0) and the finalposition (xf, yf) of the finger were known, we defined the follow-ing two functions:
For the straight finger path case, we set
fS x,y� � y � y0� �yf � y0
xf � x0 x � x0�
.
For the semicircular finger path case, we set
fC x,y� � x � xc�2 � y � yc�
2 � �LD
2 �2
and kept only the part of the curve in the semiplane defined byfs(x, y) � 0. The latter equation described an arc of a circle. Thepoint (xcyc) was the middle of the segment of length LD that wasdefined in Materials and Methods.
The relationship between the elevation angles and the fingerposition was given by the following:
Berret et al. • Modular Motor Control J. Neurosci., January 7, 2009 • 29(1):191–205 • 203

�x � xAk�lSksin�Sk�lThsin�Th
�lPesin�Pe�lTrsin�Tr
�lHusin�Hu�lFosin�Fo
�lHasin�Ha
y � yAk�lSkcos�Sk�lThcos�Th
�lPecos�Pe�lTrcos�Tr
�lHucos�Hu�lFocos�Fo
�lHacos�Ha
. (2)
The li parameter corresponded to the segment length of eachsegment, and (xAk, yAk) corresponded to the ankle marker posi-tion. Notice that Equation 2 did not depend on the head angle.
The optimal problem was to find a final configuration �f
minimizing the error between the imposed finger path [givenby equations f(x, y) � 0] and the path resulting from thechoice of �f, given by Equation 2. Notice that the initial limbconfiguration was already known and fixed in this procedure.
For the imposed straight finger path, the cost under minimi-zation was defined as the straightness of the finger trajectory,which was measured with the correlation coefficient of the fingercoordinates (x, y).
For the imposed semicircular finger path, the cost was definedas the IPC parameter that had to tend to 0.5 if the finger trajectorywas a circular arc.
For both conditions, we also tested to directly minimize theroot mean squared error between the simulated and the imposedfinger path, and this did not change significantly the kinematicstrategy found by the optimal process.
To conserve a degree of biological validity, the following non-linear constraints were added to the optimal problem: (1) TheA-P position of the CoM was forced to remain in a positionwithin the points of support bounded by the heels and toes. Thislength was evaluated as the distance between the foot markersplus 6 cm in both forward and backward directions. (2) The finalbody configuration was requested to be compatible with thereach of the target (using the finger marker position) and suchthat biological articular limits were respected (using the biologi-cal ranges of joint angles).
This problem was solved numerically by using the optimi-zation Matlab toolbox. A SQP (sequential quadratic program-ming) method was used ( fmincon Matlab function). The min-imization of the above costs was subject to nonlinearinequalities (position of the CoM and articular limits) andequalities (position of the finger on the target) defined in auser-supplied function.
Basically, the procedure was initialized to the final limb con-figuration recorded during the basic motor task (Fig. 9a). We alsoinitialized the process with 10 other admissible final limbs con-figurations selected randomly to limit the problem of local min-ima, and the resulting kinematic solutions were strongly similarwhatever the chosen initial limb configuration. The solutionsthat we got are depicted in Figure 9b– d.
ReferencesAlexandrov A, Frolov A, Massion J (1998) Axial synergies during human
upper trunk bending. Exp Brain Res 118:210 –220.Alexandrov AV, Frolov AA, Massion J (2001a) Biomechanical analysis of
movement strategies in human forward trunk bending. I. Modeling. BiolCybern 84:425– 434.
Alexandrov AV, Frolov AA, Massion J (2001b) Biomechanical analysis of
movement strategies in human forward trunk bending. II. Experimentalstudy. Biol Cybern 84:435– 443.
Babinski J (1899) De l’asynergie cerebelleuse. Rev Neurol 7:806 – 816.Belenkii V, Gurfinkel VS, Paltsev Y (1967) Elements of control of voluntary
movements. Biofizika 12:135–141.Bernstein N (1967) The coordination and regulation of movements. Ox-
ford: Pergamon.Bizzi E, Cheung VC, d’Avella A, Saltiel P, Tresch M (2008) Combining mod-
ules for movement. Brain Res Rev 57:125–133.Capaday C (2004) The integrated nature of motor cortical function. Neu-
roscientist 10:207–220.Cheron G, Bengoetxea A, Pozzo T, Bourgeois M, Draye JP (1997) Evidence
of a preprogrammed deactivation of the hamstring muscles for triggeringrapid changes of posture in humans. Electroencephalogr Clin Neuro-physiol 105:58 –71.
Cordo PJ, Nashner LM (1982) Properties of postural adjustments associ-ated with rapid arm movements. J Neurophysiol 47:287–302.
Crenna P, Frigo C (1991) A motor programme for the initiation of forward-oriented movements in humans. J Physiol 437:635– 653.
d’Avella A, Bizzi E (2005) Shared and specific muscle synergies in naturalmotor behaviors. Proc Natl Acad Sci U S A 102:3076 –3081.
d’Avella A, Portone A, Fernandez L, Lacquaniti F (2006) Control of fast-reaching movements by muscle synergy combinations. J Neurosci26:7791–7810.
Dietz V (1992) Human neuronal control of automatic functional move-ments: interaction between central programs and afferent input. PhysiolRev 72:33– 69.
Eng JJ, Winter DA, MacKinnon CD, Patla AE (1992) Interaction of thereactive moments and center of mass displacement for postural controlduring voluntary arm movements. Neurosci Res Commun 11:73– 80.
Flash T, Hogan N (1985) The coordination of arm movements: an experi-mentally confirmed mathematical model. J Neurosci 5:1688 –1703.
Hart CB, Giszter SF (2004) Modular premotor drives and unit bursts asprimitives for frog motor behaviors. J Neurosci 24:5269 –5282.
Hess WR (1954) Teleokinetisches und ereismatisches kraftesystem in derbiomotorik. Helv Physiol Pharmacol Acta 1:C62–C63.
Horak F, McPherson J (1996) Postural orientation and equilibrium. In:Handbook of physiology (Rowell LB, Shepherd JT, eds). New York: Ox-ford UP.
Hufschmidt HJ, Hufschmidt T (1954) Antagonist inhibition as the earliestsign of a sensory-motor reaction. Nature 174:607.
Ivanenko YP, Cappellini G, Dominici N, Poppele RE, Lacquaniti F (2005)Coordination of locomotion with voluntary movements in humans.J Neurosci 25:7238 –7253.
Jolliffe I (1986) Principal components analysis. New York: Springer.Krouchev N, Kalaska JF, Drew T (2006) Sequential activation of muscle
synergies during locomotion in the intact cat as revealed by cluster anal-ysis and direct decomposition. J Neurophysiol 96:1991–2010.
Lacquaniti F, Grasso R, Zago M (1999) Motor patterns in walking. NewsPhysiol Sci 14:168 –174.
Massion J (1992) Movement, posture and equilibrium: interaction and co-ordination. Prog Neurobiol 38:35–56.
Mohan V, Morasso P (2006) A forward/inverse motor controller for cogni-tive robotics. Lect Notes Comput Sci 4131:602– 611.
Mussa Ivaldi FA, Morasso P, Zaccaria R (1988) Kinematic networks. A dis-tributed model for representing and regularizing motor redundancy. BiolCybern 60:1–16.
Papaxanthis C, Pozzo T, McIntyre J (2005) Kinematic and dynamic pro-cesses for the control of pointing movements in humans revealed byshort-term exposure to microgravity. Neuroscience 135:371–383.
Patron J, Stapley P, Pozzo T (2005) Human whole-body reaching in normalgravity and microgravity reveals a strong temporal coordination betweenpostural and focal task components. Exp Brain Res 165:84 –96.
Pigeon P, Bortolami SB, DiZio P, Lackner JR (2003) Coordinated turn-and-reach movements. I. Anticipatory compensation for self-generated corio-lis and interaction torques. J Neurophysiol 89:276 –289.
Pozzo T, Berthoz A, Lefort L (1990) Head stabilization during various loco-motor tasks in humans. I. Normal subjects. Exp Brain Res 82:97–106.
Pozzo T, Ouamer M, Gentil C (2001) Simulating mechanical consequencesof voluntary movement upon whole-body equilibrium: the arm-raisingparadigm revisited. Biol Cybern 85:39 – 49.
Pozzo T, Stapley PJ, Papaxanthis C (2002) Coordination between equilib-
204 • J. Neurosci., January 7, 2009 • 29(1):191–205 Berret et al. • Modular Motor Control

rium and hand trajectories during whole body pointing movements. ExpBrain Res 144:343–350.
Ramos CF, Stark LW (1990) Postural maintenance during fast forwardbending: a model simulation experiment determines the “reduced trajec-tory.” Exp Brain Res 82:651– 657.
Schepens B, Drew T (2003) Strategies for the integration of posture andmovement during reaching in the cat. J Neurophysiol 90:3066 –3086.
Schepens B, Drew T (2004) Independent and convergent signals from thepontomedullary reticular formation contribute to the control of postureand movement during reaching in the cat. J Neurophysiol 92:2217–2238.
Schepens B, Stapley P, Drew T (2008) Neurons in the pontomedullary re-ticular formation signal posture and movement both as an integratedbehavior and independently. J Neurophysiol 100:2235–2253.
Schmid M, Schieppati M, Pozzo T (2006) Effect of fatigue on the precisionof a whole-body pointing task. Neuroscience 139:909 –920.
Shadmehr R, Wise SP (2005) The computational neurobiology of reachingand pointing: a foundation for motor learning. Cambridge, MA: MIT.
Stapley P, Pozzo T, Grishin A (1998) The role of anticipatory postural ad-justments during whole body forward reaching movements. Neuroreport9:395– 401.
Stapley PJ, Pozzo T, Cheron G, Grishin A (1999) Does the coordinationbetween posture and movement during human whole-body reaching en-sure center of mass stabilization? Exp Brain Res 129:134 –146.
Thomas JS, Corcos DM, Hasan Z (2005) Kinematic and kinetic constraintson arm, trunk, and leg segments in target-reaching movements. J Neuro-physiol 93:352–364.
Ting LH (2007) Dimensional reduction in sensorimotor systems: a frame-work for understanding muscle coordination of posture. Prog Brain Res165:299 –321.
Ting LH, Macpherson JM (2005) A limited set of muscle synergies forforce control during a postural task. J Neurophysiol 93:609 – 613.
Viviani P, Flash T (1995) Minimum-jerk, two-thirds power law, and isoch-rony: converging approaches to movement planning. J Exp Psychol HumPercept Perform 21:32–53.
Wada Y, Kaneko Y, Nakano E, Osu R, Kawato M (2001) Quantitative exam-inations for multi joint arm trajectory planning-using a robust calculationalgorithm of the minimum commanded torque change trajectory. NeuralNetw 14:381–393.
Winter D (1990) Biomechanics and motor control of human movement.New York: Wiley.
Berret et al. • Modular Motor Control J. Neurosci., January 7, 2009 • 29(1):191–205 • 205




























