Embed Size (px)
Citation preview

G
A
M
Aa
b
c
a
ARRA
KBBCTDP
1
utcscgpTvgi
AT
0d
ARTICLE IN PRESSModel
GMET-4236; No. of Pages 9
Agricultural and Forest Meteorology xxx (2010) xxx–xxx
Contents lists available at ScienceDirect
Agricultural and Forest Meteorology
journa l homepage: www.e lsev ier .com/ locate /agr formet
odelling the canopy development of bambara groundnut
.S. Karunaratnea,∗, S.N. Azam-Ali a, I. Al-Shareefa, A. Sesayb, S.T. Jørgensenc, N.M.J. Crouta
School of Biosciences, University Park Campus, University of Nottingham, NG7 2RD, United KingdomBotswana College of Agriculture, Private Bag 0027, Gaborone, BotswanaDepartment of Agriculture and Ecology, Faculty of Life Sciences, University of Copenhagen, Højbakkegård Allé 13, DK-2630, Denmark
r t i c l e i n f o
rticle history:eceived 7 October 2009eceived in revised form 26 February 2010ccepted 21 March 2010
eywords:ambara groundnutAMGRO modelanopyemperature stressrought stresshotoperiod
a b s t r a c t
Canopy development of bambara groundnut (Vigna subterranea (L.) Verdc) is affected by temperaturestress, drought stress and photoperiod. The quantification of these documented effects by means of asuitable crop model, BAMGRO is presented in this paper. Data on canopy development from five growthchamber, four glasshouse and three field experiments were analyzed to calibrate and validate the BAM-GRO model to produce simulations for temperature stress, drought stress and photoperiodic effect ontwo contrasting landraces; Uniswa Red (Swaziland) and S19-3 (Namibia). The daily initiation rate of newleaves is calculated by means of a Gaussian function and is altered by temperature stress, drought stress,photoperiod and plant density. The rate in dead leaf number is dependent upon the maximum senes-cence fraction which can be explained by physiological maturity, mutual shading, temperature stress anddrought stress. The leaf area expansion is calculated as a function of leaf number and individual leaf size.The canopy development of bambara groundnut is modelled (and tested) in BAMGRO model by meansof leaf initiation and leaf area expansion and branching and stem production are not considered. Nashand Sutcliffe value (N–S) is used as the measure of the mean square error to the observed variance.
The simulated leaf number per plant correlated well with the glasshouse measurements for UniswaRed for the two considered years (2007 and 2008) with high N–S value (Nash and Sutcliffe; from 0.76 to0.92), low mean absolute error (MAE; from ±2.91 to 3.98) and Botswana field grown in January 18 andFebruary 1, 2007 (N–S, 0.88 and 0.74; MAE, ±0.59 and ±6.18). Overall, simulated LAI correlated well withmeasured values for the two tested landraces; S19-3 (N–S value, 0.84) and Uniswa Red (N–S value, 0.80)with maximum MAE less than ±0.50 thus explaining canopy development under temperature stress,
le ph
drought stress and variab. Introduction
Bambara groundnut (Vigna subterranea (L.) Verdc) is a relativelynder researched indigenous legume that is cultivated in subsis-ence agricultural systems in sub-Saharan Africa. Its nutritionalomposition (protein content is 16-25%) is highly comparable oruperior to other legumes, providing an important supplement toereal-based diets (Linnemann and Azam-Ali, 1993). It is mainlyrown for its seeds and is eaten fresh when unripe and as aulse when ripened and mature (Linnemann and Azam-Ali, 1993).
Please cite this article in press as: Karunaratne, A.S., et al., Modelling the(2010), doi:10.1016/j.agrformet.2010.03.006
he crop is cultivated from local landraces and there are no truearieties of the species bred for specific traits or areas. Bambararoundnut exhibits a considerable degree of phenotypic diversityn morphology, growth habit and crop duration between landraces
∗ Corresponding author at: University of Sabaragamuwa, Department of Exportgriculture, Faculty of Agricultural Sciences, Belihuloya, Sri Lanka.el.: +94 45 2280046.
E-mail address: [email protected] (A.S. Karunaratne).
168-1923/$ – see front matter © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.agrformet.2010.03.006
otoperiods.© 2010 Elsevier B.V. All rights reserved.
including canopy development (Linnemann and Azam-Ali, 1993;Collinson et al., 1996, 1997). Even though canopy development is afundamental process for crop growth there has been little researcheffort focussing on modelling the canopy development for bam-bara groundnut, especially under abiotic stress conditions. Earlyefforts to model bambara groundnut simulated canopy expansionas a function of thermal time and senescence through phenologi-cal stages and water limitation (Bannayan, 2001) with leaf biomasscomputed by way of shoot to root ratio calculated from experimen-tal evidence. Leaf area was derived by a fixed ratio between leaf areato dry mass (specific leaf area – SLA; cm2 g−1).
In later work Cornelissen (2005), simulated the canopy devel-opment of bambara groundnut by relating leaf number to thermaldevelopment (Matthews and Stephens, 1998a), in conjunction withrelationships between leaf number and potential leaf area taking
canopy development of bambara groundnut. Agric. Forest Meteorol.
account of senescence due to shading, soil moisture and tempera-ture. However, field testing of the model using data from Swazilandfound that the model greatly overestimated LAI during the initialgrowth stages and this was attributed to the poor representationof stress responses (Cornelissen, 2005). In the present study the

ARTICLE ING Model
AGMET-4236; No. of Pages 9
2 A.S. Karunaratne et al. / Agricultural and Fo
Table 1Summary of experiments used for model data sets.
Experiment Data sets used for Location and year
Growth room Calibration University of Copenhagen,Denmark (2006–2008)
Glasshouse Calibration University of Nottingham,UK (2006)
Glasshouse Validation University of Nottingham,UK (2007)
Glasshouse Validation University of Nottingham,UK (2008)
ratai(ssrv
2
2
egEcNe
pNii
2
gUtasTvttlraese
ttacs
Field Validation Botswana (2006–2008)Field Validation Swaziland (2002–2003)Previous Calibration University of Nottingham,
UK
elationships are developed to simulate the rate of leaf appearances a Gaussian function of cumulative thermal time (modified byemperature stress, drought stress, photoperiod and plant density)nd thereby leaf area expansion through a further relationship forndividual leaf area. The objectives of the current study were to1) obtain experimental data on the time course of canopy expan-ion of bambara groundnut landraces at different air temperatures,oil moistures and photoperiods; (2) develop new functions for theate of new leaf production, leaf expansion and senescence underariable temperature regimes.
. Model data sets
.1. Experiments
Data for model calibration were derived from the glasshousexperiments in University of Nottingham, in 2002 and 2006 androwth room experiments in University of Copenhagen, Denmark.xperimental details of experimental design, plant sampling pro-edures, irrigation treatments and standard measurements forottingham experiments have been previously explained in Mwalet al. (2007) so only outline details are presented here.
The model was tested using data from six experiments com-rising 2 years of glasshouse experiments (Nottingham-2007,ottingham-2008), two field experiments in Botswana (two sow-
ng dates; January 18 and February 1, Notwane) and two field trialsn Swaziland (Malkerns and Luve) (Table 1).
.1.1. Nottingham calibration and validation experimentsThe three main experiments were conducted at the five
lasshouses of the Tropical Crops Research Unit (TCRU) at theniversity of Nottingham, School of Biosciences, Sutton Boning-
on Campus, United Kingdom (52◦50′ north, 1◦15′ west; 45 mltitude). They are aligned in a North–South direction to reducetructural shading and 15 m apart to avoid mutual shading. TheCRU glasshouse (Cambridge Glasshouse Company, UK) are of con-entional aluminium and glass construction erected as part of longerm research on agro-physiology of tropical crops under con-rolled conditions. The dimensions of each glasshouse are: 10.1 mong, 4.7 m wide, 2.3 m high at the eave and 3.5 m to the centralidge (Monteith et al., 1983). These dimensions provide a croppingrea of 32 m2 and a pathway of 0.2 m a around the perimeter ofach house. A gravelly/stony sand subsoil was overlaid by 0.3 m ofandy loam soil is. The soil PH was monitored at the beginning ofach season and remained at a mean of 6.7 ± 0.2.
Butyl sheeting has been used to line each house by digging out
Please cite this article in press as: Karunaratne, A.S., et al., Modelling the(2010), doi:10.1016/j.agrformet.2010.03.006
he soil to a depth of 1.25 m. This prevents horizontal and ver-ical water seepage from and to the external environment andlso separates the plots within the house. This facilitates an effi-ient control of water treatments within each plot. The excavatedoil was replaced carefully to restore the soil profile with 0.3 m
PRESSrest Meteorology xxx (2010) xxx–xxx
loamy “plough soil” overlying a gravel loam subsoil (bulk densityof 1.41 g cm−3). Four “dip wells” (two in each plot) of 1 m depthhave been installed to pump out excess water from the houses.Four aluminium access tubes are installed in each plot to monitorthe soil water content by capacitance probes (PR2; Delta-T Device).The access tubes allow soil water content to be measured to a depthof 100 cm.
A trickle irrigation system with plastic pipes (seep hose) whichcan be directed to each crop row is fitted to each house. A portablebuilding erected on the same site of the glasshouses acts as theoffice of the TCRU and coordinates a central control computer sys-tem used to control temperature, ventilation, humidity and CO2concentration in each house separately (Monteith et al., 1983).A gas-fired heater of 18 kW capacity (Powermatic Ltd., UK) con-trols air temperature in each glasshouse. Two directional bafflespositioned at the top of each heater injects hot air to increasethe temperature and it is lowered by automatically opening roofvents which run the full length of glasshouse on each side ofthe central apex. Air is continuously blown through the baf-fles, whether the heater is on or off to ensure adequate mixingof the air. The spinning disc humidifiers (Mellor-Bromley, UK),mounted at a height of 2.4 m above ground level and adjacentto the gas heaters increases humidity of the air in each houseand saturation deficits are achieved by heating and ventilation.Tube solarimeters are installed in each house above and belowthe canopy to record the incident, reflected and transmitted radi-ation by each crop stands. Readings were recorded every 30 s ona data logger (Campbell Scientific CR 10) and averaged for everyhour.
Over the summer months of 2006, 2007 and 2008 (Aprilto September), two contrasting bambara groundnut landraces;Uniswa Red (Swaziland) and S19-3 (Namibia) were grown in fiveglasshouses (TCRU) with each house having a Uniswa Red plot anda S19-3 plot under controlled temperature regimes. Two temper-atures 23 ± 5 ◦C (LT) and 33 ± 5 ◦C (HT) were imposed in the fiveglasshouses. Soil moisture in each house was non-limiting in 2006(total irrigation 381 mm for LT and 437 for HT) which is the maincalibration experiment and irrigation was stopped at 77 DAS (totalirrigation 260 mm for LT and 305 for HT) and 33 DAS (total irrigation107 mm for LT and 160 mm for HT) in 2007 and 2008, respectively.The average solar radiation varies from 9 to 12 MJ day−1 withinthe glass houses. The treatments were allocated according to split-plot design that combined two bambara groundnut landraces andtwo different temperature regimes with each treatment and wasreplicated twice and thrice at low and high temperature, respec-tively. As bambara groundnut is a short-day plant, the day lengthwas controlled to 12 h per day from 21 to 113 DAS. This was doneby covering the crop stand with a black polythene screen, fittedover a metal frame above the crop, everyday between 20:00 and08:00 h.
2.1.2. Copenhagen calibration experimentsTwo bambara groundnut landraces (Uniswa Red and S19-3)
were tested for phenological development under controlled envi-ronment (C-E) conditions at five day lengths (10, 11, 12, 13 and 14 h)at University of Copenhagen, Denmark. Plants were grown in pots(20 cm diameter, 3.5 L) using sphagnum moss as a growing mediaand temperature was 26 ◦C at night and 30 ◦C at daytime. Pots wereirrigated daily with a drip irrigation system which contained fullnutrient solution especially nitrogen (N) to avoid variation betweengenotypes in N-fixing ability. The experimental setup was split-plot
canopy development of bambara groundnut. Agric. Forest Meteorol.
with four replicates. Six plants per landrace were used as subplotsto compensate for the lack of uniformity of the landraces tested.This protocol provided data from 24 plants per treatment. Throughthe growth cycle non-destructive growth and developmental mea-surements were made.

IN PRESSG
A
nd Forest Meteorology xxx (2010) xxx–xxx 3
2
taa2ts2udadJ
2
2st3afBgpsss
2
tgt3
ailoaw(
en(
umb
N
wmc
soto
Table 2Landrace specific cardinal temperatures and parameters used for the calculation ofleaf number and leaf area for two landraces (Uniswa Red and S19-3) (values based onTCRU glasshouse experiment 2002, 2006 and growth room experiment in Universityof Copenhagen, Denmark). The units for p1 and LA1 are leaf number per hour andleaf area cm−2 per leaf, respectively. Coefficients a, b and c are unit less.
Landrace coefficient Description Uniswa Red S19-3
Tbase Base temperature 8.5 ◦C 12 ◦CTopt Optimum temperature 28 ◦C 30 ◦CThigh Ceiling temperature 38 ◦C 45 ◦CTlower Lower thresh hold 17 ◦C 24 ◦CTupper Upper thresh hold 35 ◦C 38 ◦Ca 0.075 0.13b 1100 900
ARTICLEModel
GMET-4236; No. of Pages 9
A.S. Karunaratne et al. / Agricultural a
.1.3. Botswana validation experimentsThe experimental farm, Notwane (Botswana College of Agricul-
ure – 24◦33′South, 25◦54′East, 994 m altitude and 400–600 mmverage rain fall) carried out field experiments for a set of landracest three sowing dates: December 21, January 18 and February 1 in006/2007 growing season where a range of environmental condi-ions were considered including photoperiodic effect. The averageolar radiation in the experimental site varies between 22 and6 MJ day−1 while the day length ranges from 10 to 13 h during Jan-ary to April. The experiments were conducted in a single split-plotesign with three sowing dates as the main plots and the landracess the sub plots, replicated four times. This paper explains theataset for model validation for Uniswa Red at two sowing dates;
anuary 18 and February 1.
.1.4. Swaziland validation experimentsAs described by Cornelissen (2005), from December 2002 to May
003 a set of Swaziland and Botswana landraces were grown in fieldites in Swaziland (Malkerns; 26◦30′South, 31◦13′East, 700 m alti-ude and 850–1000 mm average rain fall and Luve; 26◦20′South,1◦14′East, 580 m altitude and 700–850 mm average rain fall). Onverage the experimental sites reported a day length variationrom 10 to 13 h. Uniswa Red was selected for the validation ofAMGRO as it was common to one of the landraces grown in thelasshouse conditions, in Nottingham. During the experimentaleriod at Malkerns it was reported a well-distributed rainfall, thusuggesting no drought stress for the crop during the cropping sea-on. In the Luve field site the crop experienced a severe droughttress.
.2. Measurements
In the Nottingham experiments, leaf number was recordedwice a week on 5 previously tagged plants in each plot from emer-ence to final harvesting. Sequential growth analysis recorded theotal leaf area of individual plants using leaf area meter-model LI-100 (LI-COR, Inc. Lincoln, Nebraska, USA).
In the Botswana field experiments, sequential leaf countingnd leaf area measurements were conducted at regular 2 weeksntervals during the growing period. Similarly, both sites in Swazi-and, leaf counts were carried out twice a week for the durationf the experiment on 10 pre-selected plants which were taggedfter emergence. The leaf area per plant could not be measuredith a leaf area meter and was estimated from length and width
Cornelissen et al., 2002).In Copenhagen the leaf number and leaf size were recorded
very 20 days from sowing until 120 days after sowing (DAS) usingon-destructive measurements as described by Cornelissen et al.2002).
The performance of the model was evaluated by comparing sim-lated and measured values. Performance was expressed as theean absolute error (MAE) and model efficiency measure proposed
y Nash and Sutcliffe (1970) hereafter referred to as N–S:
–S = 1 −∑n
i=1(Mi − Si)2∑n
i=1(Mi − M)2
(1)
here Mi is the ith measured value and Si is the correspondingodel simulated value, and n is the number of measured values
onsidered.
Please cite this article in press as: Karunaratne, A.S., et al., Modelling the(2010), doi:10.1016/j.agrformet.2010.03.006
Essentially N–S is the proportion of the variation in the mea-ured data accounted for by the model and is analogous to the usef r2 in regression analysis. However, unlike r2, N–S can be nega-ive, indicating that the error about the model is greater than thebserved variation about the mean (i.e. a very poorly fitting model).
c 260 200p1 0.27 0.25LA1 33 35
Mean absolute error (MAE) is simply the mean absolute devia-tion between the simulated (Si) and measured (Mi) values:
MAE =∑n
i=1
∣∣Mi − Si
∣∣n
(2)
3. Model development and parameterisation
3.1. Rate of leaf appearance
Bambara groundnut is an indeterminate crop so the leaf appear-ance can occur from emergence until maturity, depending upon thesupply of assimilates available for leaf growth, photoperiod andthe effect of stress due to drought and temperature. Senescencealso occurs dependent upon the phenological stage and the abioticstress factors. Therefore the actual leaf number production rate canbe explained as a function between the rate of new leaf initiationand the rate of leaf senescence (Eq. (3)):
dLN
dt= (LNnew) − (LNdead) (3)
where LN is the number of leaves per plant; LNnew, is the rate ofnew leaf production (leaves per plant per day; Eq. (4)) and, LNdeadis the rate of leaf loss (leaves per plant per day; Eq. (5)).
The rate of new leaf production (LNnew) is affected by photope-riod, abiotic stress factors (drought, heat and cold) and the geneticbackground of the plant population. The proposed relationship is
LNnew ={
a × Ks1 × Densityfac × exp
[− (TT − b)2
(c)2
]}× �TTi × DLfac (4)
where TT is cumulative thermal time (degree days); �TTi, dailythermal time (degree hours day−1); Ks1, stress effect due to cold anddrought; Densityfac, the effect of density relative to TCRU density;DLfac, day length factor and a, b, c, landrace coefficients (Table 2).
A logistic function was fitted for the glasshouse data (TCRUexperiment 2002; 28 ± 5 ◦C, TCRU experiment 2006; 23 ± 5 ◦C and23 ± 5 ◦C, University of Nottingham, UK) using Genstat 8.1 todescribe the new leaves produced per cumulative thermal units.This relationship was used to calculate the daily rate of leaf produc-tion at each measurement point, yielding a data set which was usedto fit a Gaussian function (Eq. (4)) (Fig. 1). The values for parametersa, b and c were taken from the fitted equation in Gaussian function(Table 2).
Thermal time (�TTi) is calculated hourly and integrated over theday (24 h). The accumulation of thermal time (TT) over the growing
canopy development of bambara groundnut. Agric. Forest Meteorol.
period is calculated according to Eq. (5):
Td(i) = Tmean + 0.5 × (Tmax − Tmin) × cos (0.2618 × (i − 14))dTT/dt = Min[(Td(i) − Tbase/24), 0] for i = 1, 24
}(5)

ARTICLE IN PRESSG Model
AGMET-4236; No. of Pages 9
4 A.S. Karunaratne et al. / Agricultural and Forest Meteorology xxx (2010) xxx–xxx
Fig. 1. Corresponding measured (symbols; calculated from fitted logistic function)and simulated (lines) for the rate of new leaf production with cumulative ther-mal time in landraces, Uniswa Red (a) and S19-3 (b) grown at low temperature(23 ± 5 ◦C), ‘optimum’ temperature (28 ± 5 ◦C) and high temperature (33 ± 5 ◦C):T(nf
wtb
ats(m
wta
wa(wprcsWt
Fig. 2. Regression of rate of leaf appearance against day lengths for two bambaragroundnut landraces grown under day lengths (10, 11, 12, 13, and 14 h) at growthchambers: 40 DAS and 60 DAS: (a) Uniswa Red and (b) S19-3. The regression
CRU glasshouse experiment 2002 and 2006. Measured data are the average of 1028 ± 5 ◦C), 20 (23 ± 5 ◦C) and 30 (33 ± 5 ◦C) plants per landrace (standard deviationsot shown to improve the clarity). Simulated lines were obtained using Gaussian
unction (Eq. (4)) with parameters and correlation coefficients in Table 2.
here Tmean is daily mean temperature (◦C); Tmin, daily minimumemperature (◦C); Tmax, daily maximum temperature (◦C); Tbase,ase temperature (◦C); Td(i), hourly temperature in hour i (◦C).
In addition, the effect of major abiotic stress on the rate of leafppearance is explained by a modifier Ks1 that varies from zeroo one with one representing no stress and zero is for maximumtress (Eq. (6)). This is calculated considering the temperature stresslow temperature) and water stress as a novel approach in BAMGRO
odel:
Ks1 = Min(WSTRESS, 1) for Tmean > Tupper
Ks1 = Min(TSTRESS, WSTRESS) for Tmean ≤ Tlower
}(6)
here WSTRESS is drought stress; Tlower, lower threshold level ofemperature (◦C); Tupper, upper threshold level of temperature (◦C)nd TSTRESS, temperature stress index.
If the crop is exposed to a restriction of water supply and theater uptake by roots is insufficient to replenish transpiration
t potential growth rates the plant is exposed to moisture stressKarunaratne et al., 2009). The basis of calculation of WSTRESSas derived from BAMnut (Bannayan, 2001) and modified for theresent model BAMGRO considering the minimum value of the
Please cite this article in press as: Karunaratne, A.S., et al., Modelling the(2010), doi:10.1016/j.agrformet.2010.03.006
atio between two potential growth rates, one determined by inter-epted radiation (light limited growth – LLG), and the other byoil water supply (water limited growth – WLG) compared to one.
hen the crop is exposed to a range of temperatures within thewo threshold levels, the model calculates TSTRESS (cold stress with
equations for Uniswa Red are Y = 0.096 × DL − 0.19, R2 = 0.70; Y = 0.27 × DL − 1.84,R2 = 0.97 at 40 DAS and 60 DAS, respectively. The regression equations for S19-3 areY = 0.13 × DL − 0.63, R2 = 0.80; Y = 0.25 × DL − 1.55, R2 = 0.95 at 40 DAS and 60 DAS,respectively.
Tmean lower than the Tlower while temperatures above Tupper heatstress). Both WSTRESS and TSTRESS range between zero (maximumstress) and one (no stress).
A positive correlation between day length and the rate of newleaf production was observed in the controlled environmentalexperiments (growth room) in University of Copenhagen, Denmark(Fig. 2). This relationship was used to calculate the DLfac. There-fore BAMGRO calculates the day length factor to adjust the dailyrate of change in new leaf production (Eq. (7)) when the crop isgrown above 12 h day length. The gradient of the linear function ofrate of leaf production with different day lengths is simply used tocalculate (DLfac) as a new approach in BAMGRO model:
DLfac = 1 for DL ≤ 12hDLfac = p1 × DL for DL > 12h
}(7)
where DL is day length (h); DLfac, day length factor and p1, landraceparameter (Table 2).
Planting density and the increase in competition for resourceshas an effect on leaf number (Sesay and Yarmah, 1996). For mod-elling purposes, it is important to quantify this influence. A detailedwork was done at Malkerns, Swaziland with three landraces(spreading, intermediate spreading and compact growth habit) for
canopy development of bambara groundnut. Agric. Forest Meteorol.
three spacing levels (22 × 22 cm, 20.7 plants m−2; 45 cm × 45 cm,4.9 plants m−2 and 60 cm × 60 cm, 2.8 plants m−2) (Cornelissen,2005). Statistical analysis (ANOVA, p < 0.05) of the results in aboveexperiment showed a significant influence of plant spacing on leafnumber per plant, when the 22 cm × 22 cm spacing was compared

IN PRESSG
A
nd Forest Meteorology xxx (2010) xxx–xxx 5
whsasbppldlddide
3
rsfsbtmiTCdsatpsigss
Tma
TtbiwaUTnwtcu
Fig. 3. Model calibration for leaf production with cumulative thermal time in lan-draces, Uniswa Red (a) and S19-3 (b) grown at low temperature (23 ± 5 ◦C) and hightemperature (33 ± 5 ◦C): TCRU glasshouse experiment 2006. Measured data are the
◦ ◦
ARTICLEModel
GMET-4236; No. of Pages 9
A.S. Karunaratne et al. / Agricultural a
ith the 45 cm × 45 cm spacing. The 45 cm × 45 cm spacing alwaysad a higher number of leaves per plant, than the 22 cm × 22 cmpacing. However, there was no significant effect of spacing onverage leaf number per plant (p < 0.05) when the 45 cm × 45 cmpacing was compared to the 60 cm × 60 cm spacing. The possi-le reason for this result is extremely dense plants competing forhysical space light and nutrients while lower densities allow thelants to spread successfully. Also there’s a saturation point of
eaf production beyond which leaf number is not affected by theensity. According to the available information, a negative corre-
ation was derived for the rate of leaf production and the plantensity under field condition in semi-arid Africa (Botswana fieldata; Cornelissen, 2005). The planting density in the glasshouse
s 15 plants m−2, while in the field it is 10 plants m−2. Thereforeensity factor was calculated relative to the density of controllednvironment in TCRU experiments (15 plants m−2), Nottingham.
.2. Leaf senescence
BAMGRO calculates a base level of senescence due to the envi-onmental factors which is subsequently adapted according to thetress level on a daily basis. The model calculates a senescenceraction due to physiological maturity, shading (low light inten-ity), temperature (heat and cold stress), and drought stress. Theasis of senescence due to shading, temperature and soil mois-ure is similar to BAMFOOD project model (Cornelissen, 2005) and
odified for the present model BAMGRO. Due to the unavailabil-ty of data on senescence the parameter values were derived fromCRU-2006 experiment data sets by means of fitting the values inornelissen (2005) experiments under irrigated and drought con-itions and optimising for present study using Model Maker 3.oftware. When phenological stage is at flowering, 1% of leavesre assumed to senescence each day (SenPHY). In the absence ofemperature and water stress, leaf senescence is exclusively due tohysiological maturity and mutual shading. The model calculatesenescence fraction due to mutual shading (SenL) as 5% of LAI whent exceeds three under low temperatures and 10% of LAI for the croprown at temperatures above optimum with LAI greater than 5. Theenescence fractions are 1 and 10% under maximum temperaturetress (SenT) and water stress (SenW) respectively (Eq. (8)):
SenPHY = 0.01SenL = Max(Min((0.05 × (LAI − 3)), 1), 0) for Tmean ≤ Topt
SenL = Max(Min((0.1 × (LAI − 5)), 1), 0) for Tmean > Topt
SenT = Max(Min((TSTRESS × 0.01), 1), 0)SenW = Max(Min((WSTRESS × 0.1), 1), 0) for 0 ≤ WSTRESS ≤ 1
⎫⎪⎬⎪⎭ (8)
he overall fraction of leaf senescence is Ks2 and taken as a maxi-um value of SenPHY, SenL, SenT and SenW and this fraction of leaves
re removed daily from the plants as dead leaves (Eq. (9)):
Ks2 = Max(SenPHY , SenL, SenT , SenW )LNdead = LNnew × Ks2
}(9)
he model calibration results for number of leaves per plant forhe tested two landraces (Uniswa Red and S19) that are calculatedy Eq. (3) are shown in Fig. 3. The accuracy and proper function-
ng of BAMGO model for leaf production under temperature stressas tested with temperature stress function switch off (TSRESS = 1)
nd simulation results are shown in Fig. 3. Swaziland landraceniswa Red explains the significantly lower leaf production whenSTRESS = 1 (no heat stress) whereas in Namibian landrace, S19-3,
Please cite this article in press as: Karunaratne, A.S., et al., Modelling the(2010), doi:10.1016/j.agrformet.2010.03.006
o reduction. Also both landraces reported higher leaf productionhen the model assumes no TSRESS (no cold stress) and it was rela-
ively higher in S19-3 compared to Uniswa Red (Fig. 3). The resultslearly show the behaviour of BAMGRO model for leaf productionnder temperature stress.
average of 20 (23 ± 5 C) and thirty (33 ± 5 C) plants per landrace. The simulationresults with no stress (TSRESS = 1) are shown with calibration results for LT and HT.Vertical bars are standard error (±SE). Simulated lines were obtained by Eq. (4) withparameters and correlation coefficients in Table 2.
3.3. Leaf area expansion
The potential total canopy leaf area is calculated using the leafnumber (Eq. (3)) together with a fitted relationship for the areaof individual leaves. The slope of the regression line in leaf num-ber per plant with individual leaf area fitted for TCRU-2006 datasets provides the parameter values for leaf area in individual leaf(LA1; leaf area cm2 per leaf). The major assumption in calculatingthe leaf area is uniform size in individual leaf irrespective of theage of the leaf. Therefore, BAMGRO uses a single coefficient LA1 toestimate leaf area expansion and finally leaf area index (LAI) as inEq. (10):
LA = LA1 × LN
LAI = LA × density
104
}(10)
where LA is the potential leaf area (cm2); LAI, leaf area index of thecanopy and LA1 (Table 2), landrace coefficient for leaf area expan-sion.
canopy development of bambara groundnut. Agric. Forest Meteorol.
3.4. Model validation
The canopy development model was validated using the val-ues for leaf number per plant (Eq. (3)) and LAI (Eq. (10)) fortwo landraces (Uniswa Red and S19-3) grown under glasshouse

ARTICLE IN PRESSG Model
AGMET-4236; No. of Pages 9
6 A.S. Karunaratne et al. / Agricultural and Forest Meteorology xxx (2010) xxx–xxx
Fig. 4. Validation of leaf number per plant with cumulative thermal time grownunder glasshouse conditions for two bambara groundnut landraces Uniswa Red (a1
aD((
(s
3
mdT2o(icTa
Table 3Comparison of leaf number model predictions with experimental data for glasshouse(TCRU), Sutton Bonington Campus, University of Nottingham, UK and field sites inNotwane, Botswana.
Location/experimentTreatment
Period Number ofobservations
N–S MAE
TCRU glasshouseUniswa Red: 23 ± 5 ◦C 2007 36 0.89 3.09Uniswa Red: 33 ± 5 ◦C 2007 36 0.42 20.25S19-3: 23 ± 5 ◦C 2007 36 −0.10 3.77S19-3: 33 ± 5 ◦C 2007 36 0.82 7.34
TCRU glasshouseUniswa Red: 23 ± 5 ◦C 2008 32 0.76 2.91Uniswa Red: 33 ± 5 ◦C 2008 32 0.92 3.98S19-3: 23 ± 5 ◦C 2008 32 0.52 4.96S19-3: 33 ± 5 ◦C 2008 32 0.49 10.11
Botswana
the N–S values were 0.88 and 0.74 with lower MAE (±0.59 and±6.18)(Fig. 5).
nd a2), S19-3 (b1 and b2) with drought at 77 DAS (TCRU 2007 experiment) and 33AS (TCRU-2008 experiment), respectively. Measured data are the average of 20
23 ± 5 ◦C) and 30 (33 ± 5 ◦C) plants per landrace. Vertical bars are standard error±SE).
Nottingham-2007 and Nottingham-2008) and Uniswa Red in fieldites in Botswana and Swaziland.
.5. Leaf appearance
Experimentally, the rate of leaf production varied with the dailyean temperature and was reduced with drought in both lan-
races and these abotic stresses are well described by the model.he model predictions for the glasshouse experiments (2007 and008), showed that both Uniswa Red and S19-3 reduced the ratef leaf production significantly with decreasing temperature LT
◦
Please cite this article in press as: Karunaratne, A.S., et al., Modelling the(2010), doi:10.1016/j.agrformet.2010.03.006
23 ± 5 C) exhibiting the adjustments to the rate of new leafnitiation (Eq. (4)) and the rate of senescence (Eq. (9)) throughumulative stress coefficients Ks1 and Ks2, respectively (Fig. 4).he statistical results (N–S and MAE) for the model comparisonre summarised in Table 3. Simulated leaf number per plant cor-
Uniswa Red: January 18 2007 06 0.88 5.9Uniswa Red: February 1 2007 06 0.74 6.18
related well with glasshouse measurements for Uniswa Red forthe two considered years (2007 and 2008) with high N–S valuesthat ranges from 0.76 to 0.92 and low MAE (±2.91–3.98) exceptfor 2007. Here, the simulation results for Uniswa Red under HT(33 ± 5 ◦C) in 2007 reported poor correlation with low N–S (0.42)and high MAE (±20.25) during the middle part of the growth cycle(Fig. 4). The model comparison of leaf number for S19-3 reportedrelatively low N–S that ranges from 0.49 to 0.82 and higher MAE(±4.96–10.11) compared to Uniswa Red except for 2007. Similarto Uniswa Red, the weakest correlation was reported for S19-3 in2007 experiment but under LT (Table 3) for S19-3 (Fig. 4). How-ever, the results for the simulation of leaf number followed thegeneral trend of the measured values throughout the study periodfor both landraces with minor deviations coinciding with peak val-ues.
BAMGRO was further tested against the field data in semi-aridAfrica (Botswana and Swaziland). The leaf production varied withthe sowing date and the model comparison with measured val-ues reported that January 18 and February 1, 2007 in Botswana,
canopy development of bambara groundnut. Agric. Forest Meteorol.
Fig. 5. Validation of leaf number per plant measured at field sites in Botswana fortwo sowing dates: January 18 and February 1 in growing season 2006/2007 forUniswa Red. Measured data are the average of six plants per landrace. Vertical barsare standard error (±SE).
ARTICLE IN PRESSG Model
AGMET-4236; No. of Pages 9
A.S. Karunaratne et al. / Agricultural and Forest Meteorology xxx (2010) xxx–xxx 7
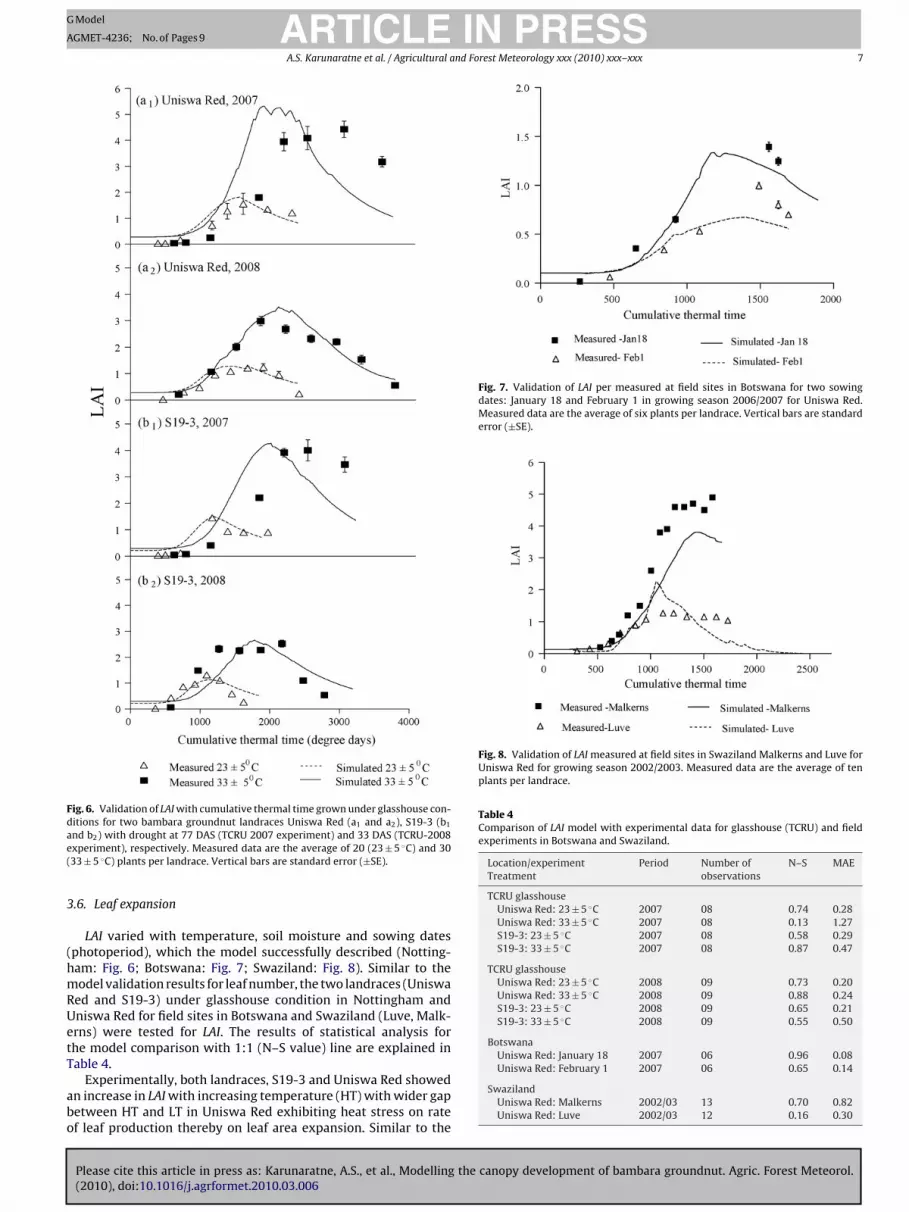
Fig. 6. Validation of LAI with cumulative thermal time grown under glasshouse con-ditions for two bambara groundnut landraces Uniswa Red (a1 and a2), S19-3 (b1
ae(
3
(hmRUetT
abo
Fig. 7. Validation of LAI per measured at field sites in Botswana for two sowingdates: January 18 and February 1 in growing season 2006/2007 for Uniswa Red.Measured data are the average of six plants per landrace. Vertical bars are standarderror (±SE).
Fig. 8. Validation of LAI measured at field sites in Swaziland Malkerns and Luve forUniswa Red for growing season 2002/2003. Measured data are the average of tenplants per landrace.
Table 4Comparison of LAI model with experimental data for glasshouse (TCRU) and fieldexperiments in Botswana and Swaziland.
Location/experimentTreatment
Period Number ofobservations
N–S MAE
TCRU glasshouseUniswa Red: 23 ± 5 ◦C 2007 08 0.74 0.28Uniswa Red: 33 ± 5 ◦C 2007 08 0.13 1.27S19-3: 23 ± 5 ◦C 2007 08 0.58 0.29S19-3: 33 ± 5 ◦C 2007 08 0.87 0.47
TCRU glasshouseUniswa Red: 23 ± 5 ◦C 2008 09 0.73 0.20Uniswa Red: 33 ± 5 ◦C 2008 09 0.88 0.24S19-3: 23 ± 5 ◦C 2008 09 0.65 0.21S19-3: 33 ± 5 ◦C 2008 09 0.55 0.50
BotswanaUniswa Red: January 18 2007 06 0.96 0.08
nd b2) with drought at 77 DAS (TCRU 2007 experiment) and 33 DAS (TCRU-2008xperiment), respectively. Measured data are the average of 20 (23 ± 5 ◦C) and 3033 ± 5 ◦C) plants per landrace. Vertical bars are standard error (±SE).
.6. Leaf expansion
LAI varied with temperature, soil moisture and sowing datesphotoperiod), which the model successfully described (Notting-am: Fig. 6; Botswana: Fig. 7; Swaziland: Fig. 8). Similar to theodel validation results for leaf number, the two landraces (Uniswa
ed and S19-3) under glasshouse condition in Nottingham andniswa Red for field sites in Botswana and Swaziland (Luve, Malk-rns) were tested for LAI. The results of statistical analysis forhe model comparison with 1:1 (N–S value) line are explained inable 4.
Please cite this article in press as: Karunaratne, A.S., et al., Modelling the canopy development of bambara groundnut. Agric. Forest Meteorol.(2010), doi:10.1016/j.agrformet.2010.03.006
Experimentally, both landraces, S19-3 and Uniswa Red showedn increase in LAI with increasing temperature (HT) with wider gapetween HT and LT in Uniswa Red exhibiting heat stress on ratef leaf production thereby on leaf area expansion. Similar to the
Uniswa Red: February 1 2007 06 0.65 0.14
SwazilandUniswa Red: Malkerns 2002/03 13 0.70 0.82Uniswa Red: Luve 2002/03 12 0.16 0.30

ARTICLE ING Model
AGMET-4236; No. of Pages 9
8 A.S. Karunaratne et al. / Agricultural and Fo
Fg(
rTBmvTsMoN
ff(R
(2wrsR((aBi
dvi(atmw1
ig. 9. Comparison between measured and simulated LAI for tested four landraces inlasshouse experiments, Nottingham, UK and field sites in Botswana and Swazilanda) Uniswa Red (b) S19-3 1:1 line (—) and regression line (—).
ate of leaf production, drought reduced the LAI in both landraces.his variation in canopy development is simulated successfully byAMGRO (Table 4). LAI correlated well with glasshouse measure-ents for Uniswa Red for 2 years (2007 and 2008) with high N–S
alues (ranges from 0.74 to 0.87) and low MAE (±0.20 to 0.28).he poor correlation in rate of leaf appearance in HT during 2007eason was consistent with simulation results of LAI (N–S, 0.13 andAE, ±1.27). S19-3 reported a good fit between the predicted to the
bserved values in all the treatments (2007 and 2008) with higher–S (ranges from 0.55 to 0.87) and lower MAE (±0.20–0.50).
LAI varied with date of sowing in field experiments as it is aunction of leaf number and the model simulates the variation of LAIor two sowing dates (January 18, February 1) tested in BotswanaFig. 7). BAMGRO simulates LAI for Botswana field grown Uniswaed with N–S values 0.96 and 0.65 (Table 4).
BAMGRO was finally tested for two field trials in SwazilandMalkerns and Luve). During the experimental period (December002 to May 2003), Malkerns reported non-water limited conditionhereas the Luve experiment was regarded as a water limited envi-
onment. The experimental observations reported that LAI showedtrong reduction with drought stress in Luve (Fig. 8). For Uniswaed the model simulation for LAI was well correlated with high N–S0.70) and low MAE (±0.82) in Malkerns but it poorly predictedN–S, 0.16; MAE, ±0.30) for the Luve experiment thus indicatingn under estimation under drought conditions (Table 4). However,AMGRO is successful in capturing the drought effect in Luve exper-
ment, thus indicating severe reduction in LAI.The BAMGRO model is intended to predict the performance of
ifferent genotypes under variable climates. Therefore the indi-idual model comparison for LAI from glasshouse experimentsn Nottingham, field trials in Botswana (Notwane) and SwazilandMalkerns and Luve) were pooled for each landrace (Fig. 9). Over-
Please cite this article in press as: Karunaratne, A.S., et al., Modelling the(2010), doi:10.1016/j.agrformet.2010.03.006
ll, simulated LAI correlated well with measured values for the twoested landraces; S19-3 (N–S, 0.84) and Uniswa Red (N–S, 0.80) with
aximum MAE less than ±0.50. The intercept of the regression lineas not significantly (p > 0.001) different to the intercept (zero) of
:1 line and the simulations starts through the origin. However, the
PRESSrest Meteorology xxx (2010) xxx–xxx
slope of regression line was significantly (p < 0.001) lower to theslope (one) of 1:1 line explaining the observed underestimation.
4. Discussion
The modified model of canopy development accounting for abi-otic stress described in this study presents a more mechanisticplatform than previous models (BAMFOOD project – Cornelissen,2005; BamNut – Bannayan, 2001) in incorporating temperature,soil moisture and photoperiod responses into a more compre-hensive canopy development model. Bambara groundnut is aphotoperiod sensitive crop therefore canopy development is mod-elled in response to the combined effect of temperature andphotoperiod.
Canopy development of bambara groundnut varied primarilydue to differences in air temperature, soil moisture and pho-toperiod. This large variation in canopy development throughappearance initiation, expansion and senescence of leaves wasaccurately reproduced in model simulations. Cold and droughtstress are simulated as a factor that reduces the rate of leaf pro-duction while heat stress and increase in photoperiod simulateda higher rate of leaf production resulted in better correlation withmeasurements for different genotypes in glasshouse, UK, Botswana(Notwane) and Swaziland (Malkern and Luve).
The new function developed in BAMGRO for the rate of new leafinitiation which is explained by a Gaussian function (Eq. (4)) suc-cessful captures the major abiotic stress (heat, cold and drought)by means of the stress index (Ks1). However, the over estima-tion of leaf number for both Uniswa Red (HT) and S19-3 (LT) in2007 season under glasshouse condition are not possible to explaindue to the unavailability of physiological information of the crop.Those two situations coincided with late season drought (77 DAS),heat and cold stress. According to the experimental evidence fromthe present study (TCRU-2006, 2007, 2008) and previous studies(Mwale et al., 2007) the Namibian landrace, S19-3 showed a fasterrate of development, which led to earlier maturity and also reportedrelatively better economy of water use compared to Uniswa Red. Incontrast, Uniswa Red was slow growing and reduces the pod for-mation under higher temperatures above upper thresh hold level.According to the detailed evaluation of responses of Uniswa Redand S19-3 for drought, Mwale et al. (2007) reported that, S19-3short phenology and fast development may reflects its adaptationto low rainfall (365 mm mean annual rainfall) and warm conditionswith short growing period. Whereas Uniswa Red showed its agro-ecological adaptation to relatively cooler, high rainfall (1390 mmmean annual rainfall) conditions having longer growing period.Considering the agro-ecological adaptations of the landraces, thisoverestimation may be due to the weak link in the model to cal-culate interaction between extreme temperatures and late seasondrought specific for each landrace.
The incorporation of day length factor (DLfac) within the rateof new leaf production function provides pathways to simulateincreased rate of leaf production when the crop is grown in theday lengths above 12 h. Moreover, the model for new leaf produc-tion accounts the differences in planting densities through densityfactor. Since LAI (Eq. (10)) is dependent on rate of leaf production,BAMGRO was capable of predicting the variation of LAI successfullyover the growing season under heat, cold and drought stress fortested landraces: Uniswa Red (TCRU-2007, TCRU 2008, SwazilandMalkerns and Luve); S19-3 (TCRU-2007 and TCRU-2008). However,
canopy development of bambara groundnut. Agric. Forest Meteorol.
there is a follow up effect of leaf number on LAI with over esti-mation especially for Uniswa Red (HT) in 2007. The main reasonfor poor model simulations in the field sites in Botswana mightbe due to the fact that the differences in growing environment inglasshouse conditions, Nottingham, UK from which the parameters

ING
A
nd Fo
wdtIB
arbfebTsa1fntsbobimbGaTaar
eoefitatrrSfifcup
5
apusfdS
the International Bambara Groundnut Symposium, Nottingham, UK, pp. 45–59.
ARTICLEModel
GMET-4236; No. of Pages 9
A.S. Karunaratne et al. / Agricultural a
ere derived. Environmental conditions in Swaziland (Malkerns)uring the growing season (2002–2003) were mild and were closero UK summer (2002, 2006) than the experiments in Botswana.n addition, the intra-landrace variability that is not considered inAMGRO model and this may have contributed to this discrepancy.
There is limited published information on the role of temper-ture for the rate of leaf appearance (Massawe et al., 2003) andole of photoperiod on canopy development (Brink, 1997) of bam-ara groundnut landraces. However the present model BAMGROollows the approaches as adapted in classical crop simulation mod-ls (Wheeler et al., 1999), in which the leaf production is simulatedy thermal time approach in the absence of photoperiodic effects.he simulation of leaf area expansion is derived from the relation-hip between temperature and rate of leaf appearance, leaf numbernd individual leaf area as in most crop growth models (Bonnet,998). Canopy expansion model was developed for pigeonpea withunctional relationships between thermal time and main stemode appearance, between main stem nodes and leaves per plant,he distribution of individual leaf area by node, and between leafenescence and thermal time (Ranganathan et al., 2001). Althoughambara groundnut is an indeterminate crop the canopy devel-pment is modelled through the leaf appearance and expansionut branching and stem node appearance are not considered as
n pigeonpea. The functions and relationships for canopy develop-ent have formulated for most of the annual crops with a common
asis of thermal time approach were successfully applied in BAM-RO. Most potato models estimate the leaf area expansion rate asn exponential function of cumulative thermal time (Fleisher andimlin, 2006). The CERES-Sorghum model estimates potential leafrea expansion as a function of leaf tip position on the main stemnd genotypic specific maximum expansion rate using a Gompertzelationship (Thornley and Johnson, 1990).
The unavailability of physiological information on interactionffect of drought and temperature results under estimations andverestimations in some occasions (TCRU-2007 experiment). Theffects of differences in the quality/quantity, of solar radiation ineld sites in semi-arid Africa and glasshouse environments in Not-ingham, UK are not well understood and no approach has beenttempted to quantify this aspect. Therefore one practical approacho understand and overcome the effect of variability due to solaradiation is to conduct detailed field experiments in contrastingadiation environments to calibrate BAMGRO for Uniswa Red and19-3. Meanwhile BAMGRO can precede with two categories foreld and glasshouse experiments in future validations. Overall, the
unctions and relationships developed within the present studyan be further improved more comprehensively with thoroughnderstanding of physiological mechanisms of temperature andhotoperiod by means of suitable experiments.
. Conclusions
A model for simulation for leaf appearance, leaf area expansionnd senescence is developed by this study as a modified approach ofrevious bambara groundnut models and other classical crop sim-
Please cite this article in press as: Karunaratne, A.S., et al., Modelling the(2010), doi:10.1016/j.agrformet.2010.03.006
lation models. The simulation results for leaf number reportedtrong correlation with observations in glasshouse experimentsrom Nottingham and in field trials from Botswana for the sowingate February 1, but an over estimation for Uniswa Red (HT) and19-3 (LT) due to the unavailability of information on physiologi-
PRESSrest Meteorology xxx (2010) xxx–xxx 9
cal mechanism of interactions effect of drought and temperature.LAI showed an excellent fit to the experimental data from theglasshouse experiments and a field trial from Swaziland (Malkerns)compared to Botswana field site mainly because the climatic con-ditions in Swaziland (Malkerns) are closer to UK summer wherethe calibration experiments were conducted. Overall the combinedresults of LAI for individual landrace are well correlated with 1:1line, but with a significant (p < 0.001) underestimation.
Acknowledgements
The authors express their gratitude to Dr. S. Mwale and Dr. R.Cornelissen, for providing data from their PhD research to calibrateand validate BAMGRO model. Special thanks to BAMLINK projectpartners for various assistance given during the study.
References
Bannayan, M., 2001. BAMnut: a crop simulation model for bambara groundnut.Agric. Sci. Technol. 15, 101–110.
Bonnet, G.D., 1998. Rate of leaf appearance in sugarcane, including a comparison ofrange of varieties. Aust. J. Plant Physiol. 25, 829–834.
Brink, M., 1997. Rates of progress towards flowering and podding in bambaragroundnut (Vigna subterranea) as a function of temperature and photoperiod.Ann. Bot. 80, 505–513.
Collinson, S.T., Azam-Ali, S.N., Chavula, K.M., Hodson, D.A., 1996. Growth, develop-ment and yield of bambara groundnut (Vigna (Vigna subterranea) in response tosoil moisture. J. Agric. Sci. (Cam.) 126, 307–318.
Collinson, S.T., Clawson, E.J., Azam-Ali, S.N., Black, C.R., 1997. Effect of soil moisturedeficits on the water relations of bambara groundnut (Vigna subterranean (L)Verdc.). J. Exp. Bot. 48, 877–884.
Cornelissen, R., Deaswarte, J.C., Azam-Ali, S.N., 2002. A preliminary stage for modeldevelopment in bambara groundnut. In: Sesay, A., Edje, O.T., Cornelissen, R.(Eds.), Increasing the Productivity of Bambara Groundnut (Vigna subterranea) forSustainable Food Production in Semi-arid Africa. Proceedings of Mid workshopUniversity of Swaziland, Kwaluseni Campus, pp. 167–176.
Cornelissen, R.L.E.J., 2005. Modelling variation in the physiology of bambara ground-nut (Vigna subterranea (L) Verdc.). PhD Thesis, School of Biosciences, Universityof Nottingham, UK.
Fleisher, D.H., Timlin, D., 2006. Modelling expansion of individual leaves in potatocanopy. Agric. Forest Meteorol. 139, 84–93.
Karunaratne, A.S., Crout, N., Azam-Ali, S.N., Mayes, S., Steduto, P., Izzi, G., 2009.Modelling the response of bambara groundnut: a key underutilized crop in agri-cultural systems. In: Proceedings of 23rd European Conference on Modelling andSimulation: ECMS, Madrid, Spain, pp. 841–847.
Linnemann, A.R., Azam-Ali, S.N., 1993. Bambara groundnut (Vigna subteranea). In:Underutilised Crops, Pulses and Vegetables. Chapman and Hall, London.
Massawe, F.J., Azam-Ali, S.N., Roberts, J.A., 2003. The impact of temperatureon leaf appearance in bambara groundnut landraces. Crop Sci. 43, 1375–1379.
Matthews, R.B., Stephens, W., 1998a. CUPPA TEA: a simulation model describingseasonal yield variation and potential production of tea. 1. Shoot developmentand extension. Exp. Agric. 34, 345–367.
Monteith, J.L., Marshall, B., Saffell, R.A., Clarke, D., Gallangher, N., Gregory, P.J., Ong,C.K., Squire, G.R., Terry, A., 1983. Environmental control of a glasshouse suit forcrop physiology. J. Exp. Bot. 34, 309, 321.
Mwale, S.S., Azam-Ali, S.N., Massawe, F.J., 2007. Growth and development of bam-bara groundnut (Vigna subterranea) in response to soil moisture. 1. Dry matterand yield. Eur. J. Agron. 26, 345–353.
Nash, J.E., Sutcliffe, J.V., 1970. River flow forecasting through conceptual models.Part 1. A discussion of principles. J. Hydrol. 227, 282–290.
Ranganathan, R., Chahuan, Y.S., Flower, D.J., Robertson, M.J., Sanetra, C., Silim, S.N.,2001. Predicting growth and development of pigeonpea: leaf area development.Field Crops Res. 69, 163–172.
Sesay, A., Yarmah, A., 1996. Field studies of bambara groundnut. In: Proceedings of
canopy development of bambara groundnut. Agric. Forest Meteorol.
Thornley, J.H.M., Johnson, J.R., 1990. Plant and crop modelling. In: A MathematicalApproach to Plant and Crop Physiology. Clarendon Press, Oxford.
Wheeler, T.R., Keatinge, J.D.H., Ellis, R.H., Summerfield, R.J., Craufurd, P.Q., 1999.Modelling the effect of temperature on the rates of seedling emergence and leafappearance in legume cover crops. Exp. Agric. 35, 327–344.





























