Embed Size (px)
Citation preview

MR
SKa
m2b
Sc
HT
AahunpcmibrcpccGcL
Kdc
Gttato(Gc
*UfEAeiacocd
Neuroscience 137 (2006) 607–617
0d
IXED ANTAGONISTIC EFFECTS OF BILOBALIDE AT �1 GABAC
ECEPTORbRt(2
(te�PcwPesrG1Crb1ra(
saGiM2oslaaZietsabS
tl
. H. HUANG,a R. K. DUKE,a,b* M. CHEBIB,b K. SASAKI,c
. WADAc AND G. A. R. JOHNSTONa
Adrien Albert Laboratory of Medicinal Chemistry, Department of Phar-acology D06, Faculty of Medicine, The University of Sydney, NSW006, Australia
Pharmaceutical Chemistry, Faculty of Pharmacy, The University ofydney, NSW 2006, Australia
Department of Hygienic Chemistry, Faculty of Pharmaceutical Sciences,ealth Sciences University of Hokkaido, Kanazawa 1757, Ishikari-obetsu, Hokkaido 061-0293, Japan
bstract—Bilobalide was found to be a moderately potentntagonist with a weak use-dependent effect at recombinantuman �1 GABAC receptors expressed in Xenopus oocytessing two-electrode voltage clamp methodology. Antago-ism of bilobalide at homomeric �1 GABAC receptors ap-eared to be mixed. At low concentration, bilobalide (3 �M)aused a parallel right shift and surmountable GABA maxi-al response of the GABA dose-response curve character-
stic of a competitive antagonist. At high concentrations,ilobalide (10–100 �M) caused nonparallel right shifts andeduced maximal GABA responses of GABA dose-responseurves characteristic of a noncompetitive antagonist. Theotency of bilobalide appears to be dependent on the con-entrations of GABA and was more potent at lower GABAoncentrations. The mechanism of action of bilobalide at �1
ABAC receptors appears to be similar to that of the chloridehannel blocker picrotoxinin. © 2005 Published by Elseviertd on behalf of IBRO.
ey words: picrotoxinin, bilobalide, mixed antagonist, use-ependent action, GABAC receptors, two-electrode voltagelamp.
ABA, the most ubiquitously distributed inhibitory neuro-ransmitter in the mammalian CNS, mediates fast neuro-ransmission via ionotropic GABA receptors, termed GABAA
nd GABAC receptors. GABAC receptors were identified fromheir insensitivity to the classical GABAA competitive antag-nist bicuculline, as well as the GABAB agonist baclofenDrew et al., 1984). GABAC receptors also differ fromABAA receptors in that they are relatively insensitive tolassical GABAA modulators such as benzodiazepines,
Correspondence to: R. Duke, Department of Pharmacology D06, Theniversity of Sydney, NSW 2006, Australia. Tel: �61-2-9351-6204;
ax: �61-2-9351-3868.-mail address: [email protected] (R. Duke).bbreviations: DMSO, dimethyl sulfoxide; ECn, effective doses thatvoked n% of maximal or control current generated by GABA for each
ndividual cell; I, peak amplitude of current at a given dose of agonist orgonist/antagonist; IC50, effective doses that inhibited 50% of maximal orontrol current generated by GABA for each individual cell; Imax, maximalr control current generated by GABA for each individual cell; nH, Hill
eoefficient; S.E.M., standard error of the mean; TPMPA, (1,2,5,6-tetrahy-ropyridine-4-yl)-methyl-phosphinic acid.
306-4522/06$30.00�0.00 © 2005 Published by Elsevier Ltd on behalf of IBRO.oi:10.1016/j.neuroscience.2005.08.071
607
arbiturates and neurosteroids (Johnston, 1996; Qian andipps, 2001) and are competitively inhibited by (1,2,5,6-
etrahydropyridine-4-yl)-methyl-phosphinic acid (TPMPA)Muratra et al., 1996; Ragozzino et al., 1996; Duke et al.,000; Johnston, 2002).
GABAC receptors are generally made up of � subunitsKusama et al., 1993a; Zhang et al., 1995) forming func-ional homomeric receptors with �1 or �2 subunits (Kusamat al., 1993a,b), or heteromeric receptors with a mixture of
1 and �2 subunits (Enz and Cutting, 1999; Zhang andan, 2001; Johnston, 2002). More recent evidence indi-ates that the GABAC �1 subunit can also co-assembleith GABAA �1 and �2 subunits (Qian and Ripps, 1999;an et al., 2000; Milligan et al., 2004). Furthermore, het-romeric receptors coexpressed by the �1 and the �2
ubunits in Xenopus oocytes have been shown to displayesponse properties very similar to those of the nativeABAC receptors on retinal bipolar cells (Qian and Ripps,999). GABAC receptors are distributed throughout theNS and are enriched mainly on neurons of the vertebrate
etina (Qian and Ripps, 2001) and less abundant in cere-ellum (Drew et al., 1984), spinal cord (Johnston et al.,975) and hippocampus (Strata and Cherubini, 1994). Aange of CNS functions including vision, sleep, cognitionnd memory is believed to involve GABAC receptorsJohnston et al., 2003).
Like the GABAA receptor, the GABAC receptor is con-idered to be made up of five protein subunits arrangedround a central channel axis that is directly gated byABA. The channel of both GABAA and GABAC receptors
s blocked by the plant convulsant picrotoxinin (Fig. 1,acdonald and Olsen, 1994; Zhorov and Bregestovski,000). Picrotoxinin is a potent noncompetitive antagonistf GABAA receptors (Newland and Cull-Candy, 1992; Ca-ida, 1993; Yoon et al., 1993), and a mixed antagonist of
obster muscle GABA receptors (Constanti, 1978; Smartnd Constanti, 1986) and GABAC receptors (Woodward etl., 1992; Qian and Dowling, 1994; Wang et al., 1995;hang et al., 1995; Qian et al., 2005). Picrotoxinin exhib-
ted characteristic features of noncompetitive and (appar-nt) competitive antagonism at GABAC receptors. Picro-oxinin competitive antagonism at GABAC receptors is con-idered to be allosteric (Qian and Dowling, 1994; Qian etl., 2005) as its site of action is distinct from the agonistinding site of GABAC receptors (Wang et al., 1995;edelnikova et al., 2005).
Bilobalide (Fig. 1) is one of the active constituents ofhe Ginkgo biloba leaf extract. The ginkgo leaves haveong been used in traditional Chinese remedies and the
xtracts of its leaves are currently being utilized widely for
tcnd2r5c2
tTlecitva2nGabacv
M
HDImCoTocd(Ci
dab27w
EX
TopvauwawcH(slp5
u�wkipvmbc
E
RtwIsaTrFmoGriaG8nicp1ort
A
T
FtPqmrbiu
S. H. Huang et al. / Neuroscience 137 (2006) 607–617608
he treatment of cerebral and peripheral vascular insuffi-iency, symptoms associated with cognitive decline andeurosensory impairments that may be associated withementia, ageing and senility (American Botany Council,000). The extract is prepared to a 35–67:1 (average 50:1)atio of dried leaves to final extract. The extract contains–7% terpene lactones, of which approximately 2.6–3.2%onsist of the sesquiterpene bilobalide, and approximately.8–3.4% of the diterpene ginkgolides.
Bilobalide is a remarkable compound with many activi-ies. It is a potent anticonvulsant (Sasaki et al., 1995, 1997).he anticonvulsant activity of bilobalide may be explained, at
east in part, in terms of its ability to inhibit the release ofxcitatory amino acids glutamate and aspartate from mouseortical slices (Jones et al., 2002; Davies et al., 2003) andncrease GABA levels in the hippocampus and cerebral cor-ex in mice (Sasaki et al., 1999). In contrast to its anticon-ulsant activity, bilobalide has been shown to be a potentntagonist of cortical GABAA and glycine receptors (Ivic et al.,003). We have also shown that bilobalide exerted largelyoncompetitive antagonism at recombinant human �1�2�2L
ABAA receptors (Huang et al., 2003). This paper reportsnd compares the effect and possible mechanism of action ofilobalide with those of TPMPA and picrotoxinin on GABA-ctivated currents from recombinant human �1 GABAC re-eptors expressed in Xenopus oocytes using two-electrodeoltage clamp methodology.
EXPERIMENTAL PROCEDURES
aterials
uman �1 cDNA subcloned into pcDNA 1.1 (Invitrogen, Saniego, CA, USA) was kindly provided by Dr. George Uhl (National
nstitute for Drug Abuse, Baltimore, MD, USA). GABA and di-ethyl sulfoxide (DMSO) were purchased from Sigma Chemicalo. (St. Louis, MO, USA). Bilobalide was isolated from the extractf Ginkgo biloba leaves provided by Schwabe Greenwave Ltd.,okyo, Japan and purified following the method described previ-usly by Wada and coworkers (1993). Picrotoxinin was purified byhromatographic separation from picrotoxin following the methodescribed by Jarboe et al. (1968). 1H-nuclear magnetic resonance
1H-NMR) spectra in CDCl3 and 13C-NMR spectra in CDCl3/D OD of picrotoxinin were consistent with the published spectra
ig. 1. The structures of picrotoxinin and bilobalide illustrating struc-ural similarities in the hydrophilic cavity and lipophilic side chain.icrotoxinin is a sesquiterpene epoxydilactone and bilobalide a ses-uiterpene trilactone. These compounds have cavity-like structuresade up of a highly oxygenated carbon skeleton, including two lactone
ings and an epoxy group in picrotoxinin, and three lactone rings inilobalide. The lipophilic side chain, the isopropenyl group in picrotox-
nin and t-butyl group in bilobalide is attached to the cavity from thenderside.
3
n (CD3)2CO (Perry et al., 2001). Drug solutions were prepared by o
iluting 100 mM aqueous stock solution of GABA and TPMPA,nd 100 mM or 200 mM DMSO stock solutions of picrotoxinin andilobalide in frog Ringer’s solution (ND96 buffer: 96 mM NaCl,mM KCl, 1 mM MgCl2.6H2O, 1.8 mM CaCl2, 5 mM HEPES, pH
.5). The highest concentration of DMSO superfusing the oocytesas 0.8% at which concentration DMSO had no effects.
xpression of �1 GABAC receptors inenopus laevis oocytes
he procedures involved the use of Xenopus laevis. The numberf animals used, and their suffering, was minimized. All surgicalrocedures and post-operative care were approved by the Uni-ersity of Sydney Animal Ethics committee and conducted inccordance with the Australian code of practice for the care andse of animals for scientific purposes. Female Xenopus laevisere anesthetized with 0.17% ethyl 3-aminobenzoate in salinend a lobe of the ovaries surgically removed. The lobe of ovariesas rinsed with a low Cl� Ringer’s solution (OR-2 buffer) thatontained 82.5 mM NaCl, 2 mM KCl, 1 mM MgCl2.6H2O, 5 mMEPES, pH 7.5, and suspended in a solution of Collagenase A
2 mg/ml in OR-2, Boehringer Mannheim, Germany) for 2 h toeparate oocytes from connective tissues and follicular cells. Re-eased oocytes were then thoroughly rinsed in ND96 buffer sup-lemented with 2.5 mM sodium pyruvate, 0.5 mM theophylline and0 �g/ml gentamycin, and stage V to VI oocytes were collected.
Human �1 cDNA subcloned in pcDNA 1.1 was linearizedsing the restriction enzyme XbaI. Linearized plasmids containing1 cDNA were transcribed using T7 RNA Polymerase and cappedith 5,7-methylguanosine using the “mMESSAGE mMACHINE”it (Ambion Inc., Austin, TX, USA). �1 cRNA (10 ng/50 nl) was
njected using a 15–20 �m diameter tip micropipette (micropipetteuller, Sutter Instruments, Inc., USA) into the cytoplasm of indi-idual defolliculated oocytes by using a Nanoject injector (Drum-ond Scientific Co., Broomali, PA, USA). The oocytes were incu-ated for 2–7 days at 18 °C in ND96 buffer with a twice-dailyhange of buffer.
lectrophysiological recording
eceptor activity was measured with two-electrode voltage clampechniques 2–7 days after injection. Recording microelectrodesere fabricated with a micropipette puller (Narishige Scientific
nstrument Laboratory, Tokyo, Japan) and filled with 3 M KClolution. Oocytes were placed in a cell bath and voltage clampedt �60 mV. Cells were continuously superfused with ND96 buffer.he currents elicited in response to the application of drugs wereecorded using a Geneclamp 500 amplifier (Axon Instrument Inc.,oster City, CA, USA), a Mac Laboratory 2e recorder (AD Instru-ents, Sydney, NSW, Australia), and Chart version 3.5.2 programn a Macintosh Quadra 605 computer. A test dose of 10 �MABA was applied to each oocyte to confirm there was sufficient
eceptor expression for recording prior to the start of each exper-ment. For measurements of inhibitory action of drugs on receptorctivation, drugs were added to the buffer solution containingABA at the concentration producing approximately 15%, 50%,0% and 100% of the effect (GABA effective doses that evoked% of maximal or control current generated by GABA for each
ndividual cell (EC15, EC50, EC80 and EC100) at the receptors foronstructing GABA inhibition dose-response curves. The samerocedure, but with a fixed concentration of antagonists (3 �M,0 �M, 30 �M, 50 �M and 100 �M) and increasing concentrationsf GABA (0.01–100 �M), was applied to construct GABA dose-esponse curves. A washout period of 3–5 min was allowed be-ween each drug application to prevent receptor desensitization.
nalysis of data
he peak amplitude of current in response to each concentration
f drug was recorded and standardized by calculating the ratio
%giDtd((tt(
cGna(tmtfGP
FX
Cowv1mwtaG
I
Tcemoop2at
wrtd03sb
ta
0G
GgoG9G3dsG
cwlpwpr1od1
c(a0
Mp
Tlc5tGoc
ppGT23
pr(pddpG
S. H. Huang et al. / Neuroscience 137 (2006) 607–617 609
Imax�I/Imax100, where I is the peak amplitude of current at aiven dose of agonist or agonist/antagonist, and Imax is the max-
mal or control current generated by GABA for each individual cell.ata were expressed as the averaged % Imaxstandard error of
he mean (S.E.M.). The EC50 were calculated from dose-responseata constructed with % Imax as a function of agonist concentration[A]) by least square method to the Hill equation I�Imax [A]nH/EC50
nH�[A]nH), where nH is the Hill coefficient. The effective doseshat inhibited 50% of Imax (IC50) were calculated in a similar mannero EC50 values from the inverse Hill equation I�Imax�{Imax[Ant]nH/IC50
nH�[Ant]nH)}, where [Ant] is the concentration of the antagonist.EC50, IC50 and nH numbers were estimated by fitting the
oncentration-response relationships to the logistic equation usingraphPad Prism v3.02 (GraphPad Software). Unless otherwiseoted, parameters were calculated for individual cells and thenveraged. These parameters are reported as meanS.E.M.n�4–15 oocytes). The statistical significance of differences be-ween GABA responses with and without antagonists was deter-ined by two-way ANOVA method, whereas the differences be-
ween IC50 values at different GABA concentrations and the dif-erences in levels of inhibition between peak and steady-stateABA responses, by Student t-test at the significance level of�0.05.
RESULTS
unctional properties of �1 GABAC receptors in. laevis oocytes
ytoplasmic injection of human wild-type �1 cRNA intoocytes formed functional homomeric GABA receptorsith pharmacological profiles consistent with reports pre-iously described for GABAC receptors (Kusama et al.,993a,b; Chebib et al., 1998; Duke et al., 2000). Theagnitude of inward whole-cell currents of 200–3000 nAas recorded at �60 mV. GABA EC50 and nH values were in
he range of 0.90–1.24 �M and 1.78–2.36. There was little ifny desensitization to GABA upon continuing application ofABA observed for �1 GABAC receptors.
nhibition of direct GABA-mediated currents
PMPA, picrotoxinin and bilobalide inhibited the chlorideurrents generated by 1.2 �M GABA (EC50) from oocytesxpressing functional �1 receptors in a dose-dependentanner (Fig. 2A–C) but had no effect when tested on theirwn at 100 �M. The GABAC receptor competitive antag-nist, TPMPA, at high concentrations (10–100 �M) com-letely blocked GABA-mediated chloride currents (Fig.A). In contrast, the maximum percentage inhibitionchieved by picrotoxinin and bilobalide was always lesshan 100% (Fig. 2B and C).
The potency of TPMPA, picrotoxinin and bilobalideas calculated from the inhibition dose-response curves
epresenting the effects of a range of antagonist concen-rations on a fixed concentration of GABA. The inhibitionose-response curves of picrotoxinin and bilobalide at.5 �M, 1.2 �M, 3 �M and 10 �M GABA are shown in Fig.A–D and TPMPA at 1.2 �M GABA in Fig. 3B. The re-pective IC50 and nH values for TPMPA, picrotoxinin andilobalide are tabulated in Table 1.
TPMPA (IC50�1.070.12 �M) was approximately 1.8imes less potent than picrotoxinin (IC �0.590.06 �M) and
50bout 1.5 times more potent than bilobalide (IC50�1.6 0
.12 �M) at inhibiting the response from 1.2 �M GABA at �1
ABAC receptors.Picrotoxinin was most potent at inhibiting 0.5 �M
ABA (the lowest GABA concentration examined) andradually became less potent with increased concentrationsf GABA (Table 1). The potency of picrotoxinin at 0.5 �MABA (IC50�0.340.03 �M) was approximately 1.7, 14 and3 times higher than its potency at 1.2 �M, 3 �M and 10 �MABA (IC50�0.590.06 �M, 4.830.51 �M and1.554.59 �M) respectively. IC50 values for picrotoxininetermined at 1.2 �M, 3 �M and 10 �M GABA wereignificantly different from the value determined at 0.5 �MABA (P�0.0037, �0.0001 and �0.0001).
Similar to picrotoxinin, the potency of bilobalide was alsolearly dependent on the concentrations of GABA. Bilobalideas also most potent at inhibiting 0.5 �M GABA and became
ess potent as the concentrations of GABA increased. Theotency of bilobalide at 0.5 �M GABA (IC50�0.510.05 �M)as approximately 3, 17 and 286 times higher than itsotency determined at 1.2 �M, 3 �M and 10 �M GABA,espectively (IC50�1.600.12 �M, 8.641.03 �M and46.0811.89 �M, respectively). The potency of bilobalidebtained at higher GABA concentrations was significantlyifferent from that of 0.5 �M GABA (P�0.0001 for 1.2–0 �M GABA).
Bilobalide was less potent than picrotoxinin at all GABAoncentrations examined. It was approximately 1.5 timesP�0.0150), 2.7 times (P�0.0001), 1.8 times (P�0.0028)nd 4.6 times (P�0.0001) less potent than picrotoxinin at.5 �M, 1.2 �M, 3 �M and 10 �M GABA, respectively.
echanism of action of TPMPA,icrotoxinin and bilobalide
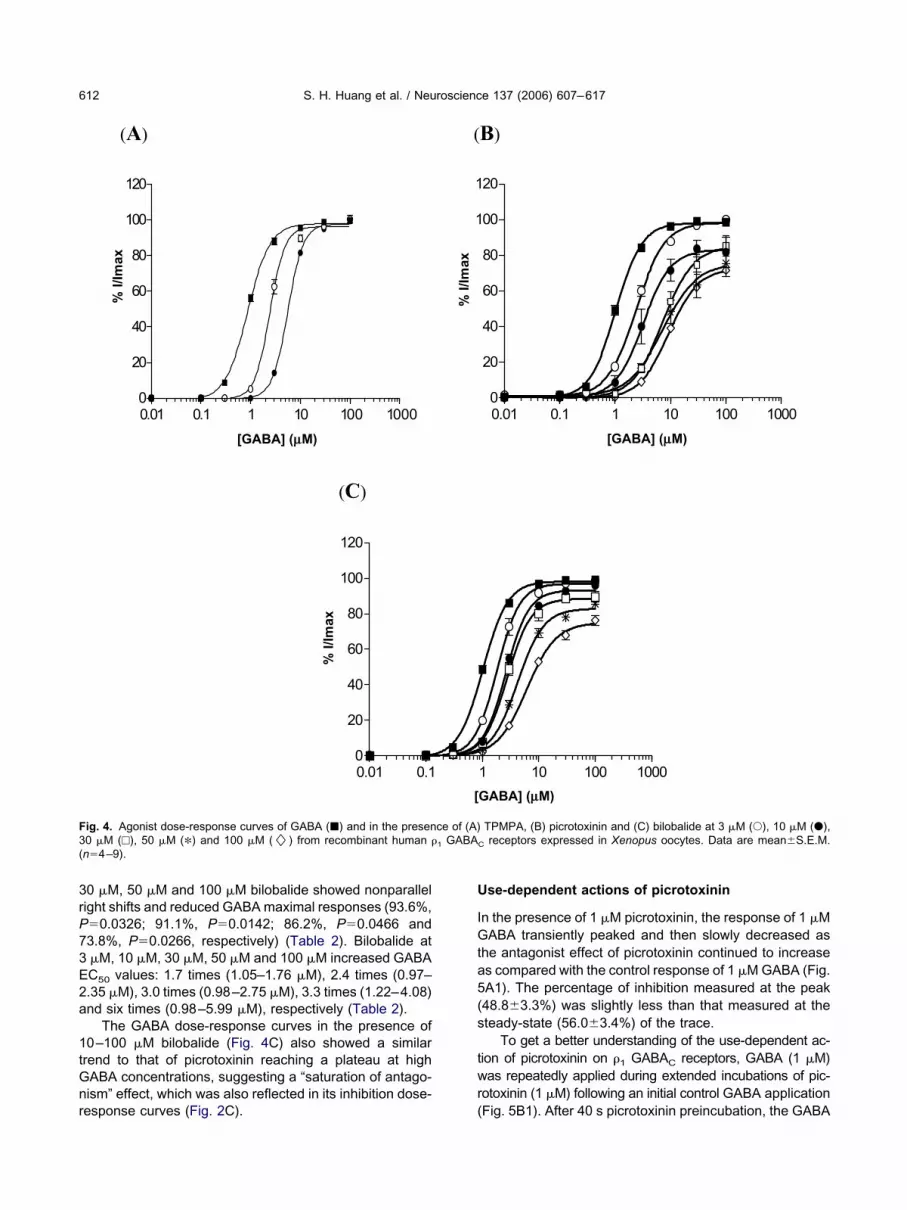
he mechanism of action of TPMPA, picrotoxinin and bi-obalide was determined by evaluating the effect of eachompound at a fixed concentration (3 �M, 10 �M, 30 �M,0 �M and 100 �M) in the presence of increasing concen-rations of GABA and comparing with that of GABA alone.ABA dose-response curves in the absence and presencef TPMPA, picrotoxinin and bilobalide at different fixedoncentrations were constructed.
TPMPA. The GABA dose-response curves in theresence of 3 �M and 10 �M TPMPA (Fig. 4A) displayed aarallel right shift and attained the maximal response ofABA (100.7%, P�0.4523 and 99.4%, P�0.7406) (Table 2).he GABA EC50 values were right shifted 2.7 times (0.92–.52 �M) and 6.4 times (0.90–5.78 �M) in the presence ofand 10 �M TPMPA (Table 2).
Picrotoxinin. The GABA dose-response curves in theresence of 3 �M picrotoxinin (Fig. 4B) displayed a parallelight shift and attained the maximal response of GABA99.5%, P�0.6377) (Table 2). At higher concentrations oficrotoxinin (10 �M, 30 �M, 50 �M and 100 �M) the GABAose-response curves all showed nonparallel right shifts andid not reach the maximal response of GABA (Fig. 4B). In theresence of 10 �M, 30 �M, 50 �M and 100 �M picrotoxinin,ABA responses were 81.0% (P�0.0344), 83.3% (P�
.0454), 76.3% (P�0.0308) and 76.9% (P�0.0342) of its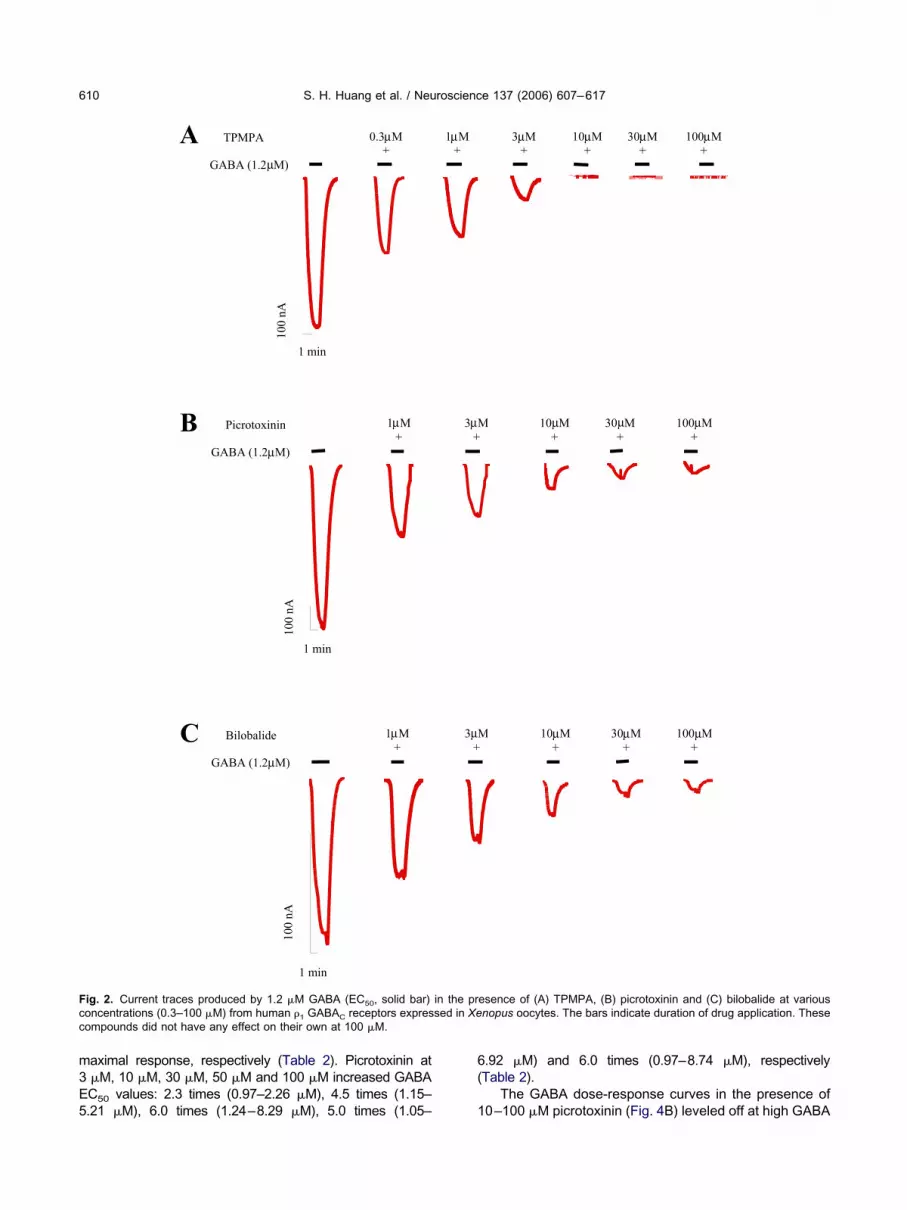
m3E5
6(
Fcc
S. H. Huang et al. / Neuroscience 137 (2006) 607–617610
aximal response, respectively (Table 2). Picrotoxinin at�M, 10 �M, 30 �M, 50 �M and 100 �M increased GABAC values: 2.3 times (0.97–2.26 �M), 4.5 times (1.15–
ig. 2. Current traces produced by 1.2 �M GABA (EC50, solid bar)oncentrations (0.3–100 �M) from human �1 GABAC receptors expresompounds did not have any effect on their own at 100 �M.
50
.21 �M), 6.0 times (1.24–8.29 �M), 5.0 times (1.05– 1
.92 �M) and 6.0 times (0.97–8.74 �M), respectivelyTable 2).
The GABA dose-response curves in the presence of
esence of (A) TPMPA, (B) picrotoxinin and (C) bilobalide at variousnopus oocytes. The bars indicate duration of drug application. These
in the prsed in Xe
0–100 �M picrotoxinin (Fig. 4B) leveled off at high GABA

dsmm1
r4mt
Fa ressed in
T
C
*PB
a
*
S. H. Huang et al. / Neuroscience 137 (2006) 607–617 611
oses, indicating a “saturation of antagonism” effect (Con-tanti, 1978; Smart and Constanti, 1986). This effect wasirrored in the inhibition dose-response curves where theaximum inhibition by picrotoxinin was always less than00% (Fig. 2B).
ig. 3. Inhibition dose-response curves of (A) 0.5 �M, (B) 1.2 �M, (Cnd bilobalide (�) from recombinant human �1 GABAC receptors exp
able 1. IC50 and nH values for TPMPA, picrotoxinin and bilobalide in
ompounds 0.5 �M GABA 1.2 �M GABA
IC50a (�M) nH
a IC50a (�M) nH
a
TPMPA – – 1.070.12 �1.3icrotoxinin 0.340.03 �1.210.07 0.590.06 �0.8ilobalide 0.510.05 �0.850.04 1.600.12 �0.8
Data are the meanS.E.M. (n�4–9).
TPMPA KB�2.0–2.3 �M (Chebib et al., 1998; Carland et al., 2004).Bilobalide. Similar to picrotoxinin, the GABA dose-esponse curves in the presence of 3 �M bilobalide (Fig.C) displayed a parallel shift to the right and attained theaximal response of GABA (98.4%, P�0.0615). In con-
rast, the dose-response curves in the presence of 10 �M,
d (D) 10 �M GABA in the presence of TPMPA (’), picrotoxinin (□)Xenopus oocytes. Data are meanS.E.M. (n�4–9).
ence of 0.5, 1.2, 3 and 10 �M GABA at �1 GABAC receptors
3 �M GABA 10 �M GABA
IC50a (�M) nH
a IC50a (�M) nH
a
– – – –4.830.51 �0.730.03 31.554.59 �0.570.048.641.03 �1.030.11 146.0811.89 �0.640.03
) 3 �M an
the pres
10.0250.0600.03
3rP73E2a
1tGnr
U
IGta5(s
twr
F3 1
(
S. H. Huang et al. / Neuroscience 137 (2006) 607–617612
0 �M, 50 �M and 100 �M bilobalide showed nonparallelight shifts and reduced GABA maximal responses (93.6%,�0.0326; 91.1%, P�0.0142; 86.2%, P�0.0466 and3.8%, P�0.0266, respectively) (Table 2). Bilobalide at�M, 10 �M, 30 �M, 50 �M and 100 �M increased GABAC50 values: 1.7 times (1.05–1.76 �M), 2.4 times (0.97–.35 �M), 3.0 times (0.98–2.75 �M), 3.3 times (1.22–4.08)nd six times (0.98–5.99 �M), respectively (Table 2).
The GABA dose-response curves in the presence of0–100 �M bilobalide (Fig. 4C) also showed a similar
rend to that of picrotoxinin reaching a plateau at highABA concentrations, suggesting a “saturation of antago-ism” effect, which was also reflected in its inhibition dose-
ig. 4. Agonist dose-response curves of GABA (�) and in the presen0 �M (□), 50 �M (�) and 100 �M (�) from recombinant human �n�4–9).
esponse curves (Fig. 2C). (
se-dependent actions of picrotoxinin
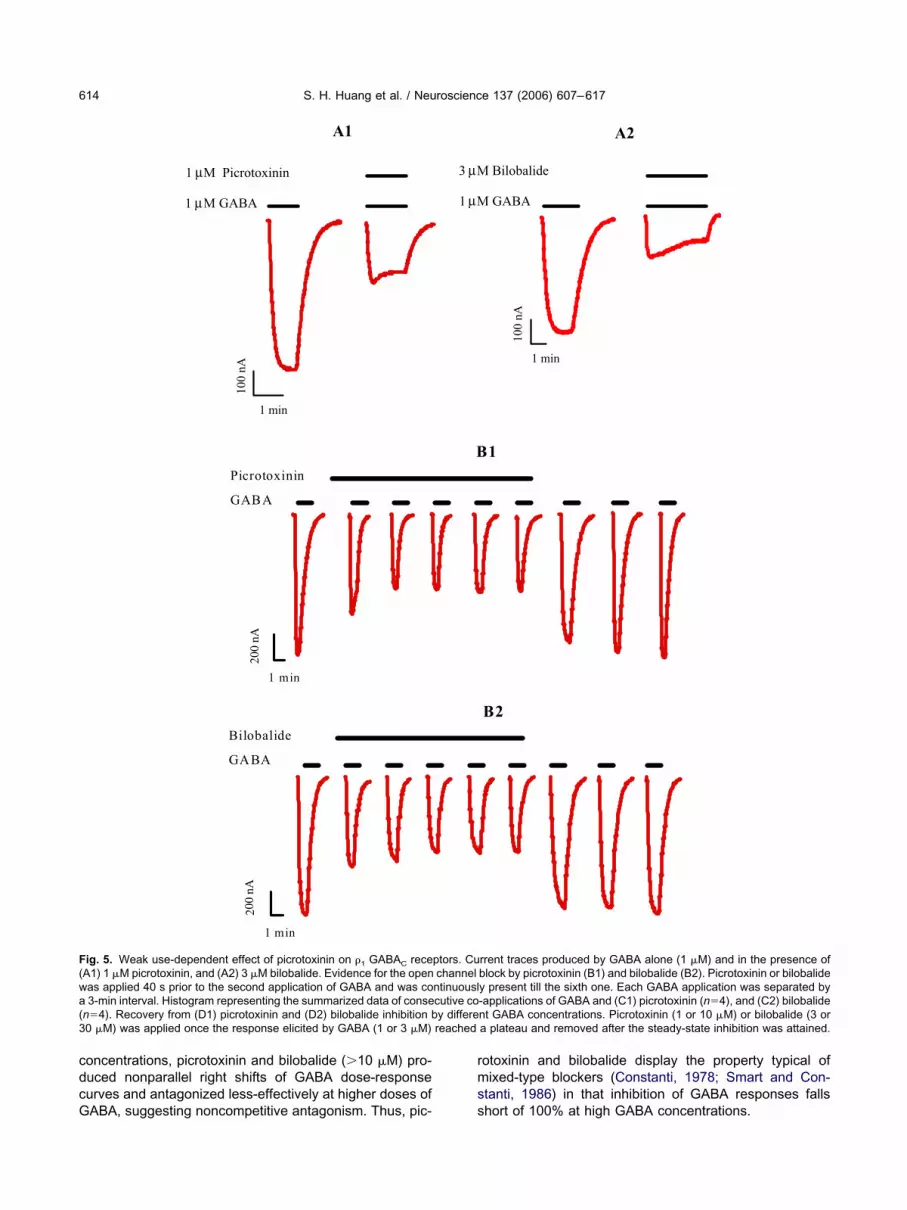
n the presence of 1 �M picrotoxinin, the response of 1 �MABA transiently peaked and then slowly decreased as
he antagonist effect of picrotoxinin continued to increases compared with the control response of 1 �M GABA (Fig.A1). The percentage of inhibition measured at the peak48.83.3%) was slightly less than that measured at theteady-state (56.03.4%) of the trace.
To get a better understanding of the use-dependent ac-ion of picrotoxinin on �1 GABAC receptors, GABA (1 �M)as repeatedly applied during extended incubations of pic-
otoxinin (1 �M) following an initial control GABA application
TPMPA, (B) picrotoxinin and (C) bilobalide at 3 �M (Œ), 10 �M (�),
C receptors expressed in Xenopus oocytes. Data are meanS.E.M.
ce of (A)GABA
Fig. 5B1). After 40 s picrotoxinin preincubation, the GABA

rbG(aiatf(
r�otd5iG511t
U
BorctdN(m
bbwAr
pTaAoosa
bnhA9a5i(b(c
PlatWTbGdtrGbt(o
TG
A
1
1
a
S. H. Huang et al. / Neuroscience 137 (2006) 607–617 613
esponse was reduced considerably to 55.48.1% andriefly reached a plateau (44.86.0%) after three–fiveABA applications. There was a significant difference
P�0.0394) in the levels of inhibition between the firstpplication of GABA and the steady-state. The levels of
nhibition obtained in experiments illustrated in Fig. 5A1nd 5B1 were comparable. The antagonist effect of picro-oxinin appeared to be reversible as demonstrated by theull recovery of GABA responses after a washout periodFig. 5B1 and 5C1).
In accordance with Goutman and Calvo (2004), theecovery of GABA responses from picrotoxinin inhibition at
1 GABAC receptors in this study also showed dependencyn the agonist concentrations. Picrotoxinin was applied to
he receptors once the chloride currents generated byifferent concentrations of GABA reached a plateau (Fig.D1). The recovery time (measured as the 10–90% rise
nterval) from the effect caused by 1 �M picrotoxinin at 1 �MABA was 46.76.5 s and the inhibition produced was3.72.8%. Whereas at 3 �M GABA, the recovery time by
�M picrotoxinin was approximately three times faster:4.33.6 s with 25.21.9% inhibition, and by 10 �M picro-oxinin also faster (18.04.4 s) with 54.22.1% inhibition.
se-dependent actions of bilobalide
ilobalide was also examined for its use-dependent actionn �1 GABAC receptors and Fig. 5A2 illustrates the chlo-ide currents generated by 1 �M GABA and by co-appli-ation of 1 �M GABA and 3 �M bilobalide. Unlike picro-oxinin, co-application of 1 �M GABA and 3 �M bilobalideid not produce a sharp peaked response (43.73.7%).evertheless, the response to GABA decreased gradually
to 35.35.3%) as the effect of bilobalide steadily accu-ulated.
Similar to picrotoxinin, the use-dependent action ofilobalide on �1 GABAC receptors was also demonstratedy successive applications of 1 �M GABA on receptorsith extended bilobalide (3 �M) incubations (Fig. 5B2).fter 40 s bilobalide preincubation, the GABA response was
able 2. EC50 and nH values and maximal responses of GABA for GABAC receptors
ntagonists (Ant) GABA EC50a (�M) GABA�A
3 �M TPMPA 0.920.04 2.520.110 �M TPMPA 0.900.02 5.780.03 �M Picrotoxinin 0.970.06 2.260.1
10 �M Picrotoxinin 1.150.09 5.210.230 �M Picrotoxinin 1.240.08 8.290.850 �M Picrotoxinin 1.050.16 6.920.800 �M Picrotoxinin 0.970.04 8.740.33 �M Bilobalide 1.050.10 1.760.1
10 �M Bilobalide 0.970.07 2.350.230 �M Bilobalide 0.980.10 2.750.150 �M Bilobalide 1.220.13 4.080.400 �M Bilobalide 0.980.06 5.990.1
Data are the meanS.E.M. (n�4–9).
educed dramatically to 51.35.7% and shortly reached a r
lateau (42.85.3%) after three–five GABA applications.he levels of inhibition between the first GABA applicationnd the steady-state were significantly different (P�0.0276).gain, these inhibition values were comparable to thosebtained in experiments illustrated in Fig. 5A2. The antag-nist effect of bilobalide appeared to be reversible ashown by the complete recovery of GABA responses afterwashout period (Fig. 5B2 and 5C2).
Interestingly, the recovery of GABA responses fromilobalide inhibition also showed dependency on the ago-ist concentrations (Fig. 5D2). Bilobalide was shown toave faster washout rates at higher GABA concentrations.t 1 �M GABA, the recovery time (measured as the 10–0% rise interval) from the effect caused by 3 �M bilob-lide was 59.83.6 s and the inhibition produced was7.72.5%. Whereas at 3 �M GABA, the 10–90% rise
nterval was found to be approximately six times faster10.31.4 s) with an inhibition level of 20.11.4% by 3 �Milobalide, and was approximately four times faster15.03.7 s) with 54.64.5% inhibition by a higher con-entration of bilobalide (30 �M).
DISCUSSION
icrotoxinin was first shown to exert mixed antagonism onobster muscle GABA receptors (Constanti, 1978; Smartnd Constanti, 1986) and subsequently on GABAC recep-
ors (Woodward et al., 1992; Qian and Dowling, 1994;ang et al., 1995; Zhang et al., 1995; Qian et al., 2005).
his study shows that bilobalide is also a mixed antagonistut less potent than picrotoxinin at recombinant human �1
ABAC receptors. At human �1 GABAC receptors, TPMPAisplayed the general properties of a competitive-type an-agonist, producing a parallel right shift of GABA dose-esponse curves without affecting the maximal response ofABA. In contrast, picrotoxinin and bilobalide exhibitedoth competitive and noncompetitive antagonisms charac-eristic of the mixed antagonist. At a low concentration3 �M), picrotoxinin and bilobalide caused a parallel right shiftf the GABA dose-response curve with maximal GABA
e and in the presence of TPMPA, picrotoxinin and bilobalide at �1
(�M) GABA nHa GABA�Ant nH
a Max. GABAresponse (%)
1.850.21 2.860.43 100.71.782.060.10 2.830.01 99.430.301.940.11 1.670.02 99.480.562.010.08 1.920.13 81.022.441.780.10 1.520.05 83.322.872.300.32 1.510.17 76.284.401.800.12 1.650.03 76.912.502.330.17 2.280.09 98.400.342.250.23 2.070.18 93.592.402.160.23 1.800.12 91.121.342.360.04 2.110.17 86.161.092.300.21 1.790.11 73.833.25
ABA alon
nt EC50a
497543952739
esponse, indicating competitive-type antagonism. At high
cdcG
rms
F(wa(3 reached
S. H. Huang et al. / Neuroscience 137 (2006) 607–617614
oncentrations, picrotoxinin and bilobalide (�10 �M) pro-uced nonparallel right shifts of GABA dose-responseurves and antagonized less-effectively at higher doses of
ig. 5. Weak use-dependent effect of picrotoxinin on �1 GABAC receA1) 1 �M picrotoxinin, and (A2) 3 �M bilobalide. Evidence for the openas applied 40 s prior to the second application of GABA and was co3-min interval. Histogram representing the summarized data of conse
n�4). Recovery from (D1) picrotoxinin and (D2) bilobalide inhibition b0 �M) was applied once the response elicited by GABA (1 or 3 �M)
ABA, suggesting noncompetitive antagonism. Thus, pic- s
otoxinin and bilobalide display the property typical ofixed-type blockers (Constanti, 1978; Smart and Con-
tanti, 1986) in that inhibition of GABA responses falls
rrent traces produced by GABA alone (1 �M) and in the presence ofblock by picrotoxinin (B1) and bilobalide (B2). Picrotoxinin or bilobalidey present till the sixth one. Each GABA application was separated by-applications of GABA and (C1) picrotoxinin (n�4), and (C2) bilobalident GABA concentrations. Picrotoxinin (1 or 10 �M) or bilobalide (3 ora plateau and removed after the steady-state inhibition was attained.
ptors. Cuchannelntinuouslcutive coy differe
hort of 100% at high GABA concentrations.

tcGatcotwacsaots
rca
sdanreiptbdrs(
ttaCf
S. H. Huang et al. / Neuroscience 137 (2006) 607–617 615
Competitive-type antagonism by bilobalide and picro-oxinin was also evident in the inhibition dose-responseurves in that their potencies were clearly dependent onABA concentrations. Both compounds were most potentt the lowest GABA concentration tested (0.5 �M) andheir potency decreased as GABA concentrations in-reased (1.2–10 �M). Since a true noncompetitive antag-nist would produce a constant IC50 value regardless of
he agonist concentrations, the variation in their potencyith different GABA concentrations shows that picrotoxininnd bilobalide are neither classical competitive nor classi-al noncompetitive antagonists thus confirming conclu-ions made earlier from GABA dose-response curves. Inddition, these compounds showed a “saturation of antag-nism” effect at high antagonist concentrations, which ishe characteristic feature of mixed-type blockers (Con-tanti, 1978; Smart and Constanti, 1986).
We also examined the use-dependent actions of pic-otoxinin and bilobalide on �1 GABAC receptors using pro-edures described by Goutman and Calvo (2004). Co-
Fig. 5
pplication of GABA and picrotoxinin produced a steady- p
tate inhibition as the action of picrotoxinin graduallyeveloped (Fig. 5A1) similar to the findings of Goutmannd Calvo (2004). Under the same condition, the antago-istic effect of bilobalide also increased slightly over timeeaching a steady-state (Fig. 5A2). In the prolonged pres-nce of either bilobalide or picrotoxinin, a similar trend of
ncreasing levels of inhibition was observed during re-eated applications of GABA (Fig. 5B1 and 5B2 respec-ively). These findings clearly indicate that picrotoxinin andilobalide antagonism at �1 receptors involves use-depen-ent actions suggesting an “agonist-bound state of theeceptor” (and subsequent stabilization of agonist-boundhut state) being a prerequisite for inhibitory interactionsSmart and Constanti, 1986).
In addition, a clear dependency of agonist concentra-ion for �1 receptors recovery from picrotoxinin block duringhe washout by GABA was observed. This finding is ingreement with what was observed by Goutman andalvo (2004). Recovery of the responses was found to be
acilitated by high GABA concentrations independent of
ued)
. (Continicrotoxinin concentrations. This was also the case for

bfbcfnb
prctaatirat
cccn1ats3pt
ratcpp(saosonaphtaaroaa
vtc
eescwh
Itcarm(uwG
ADTmac
A
C
C
C
C
D
D
D
E
G
S. H. Huang et al. / Neuroscience 137 (2006) 607–617616
ilobalide. Recovery of GABA responses was found to beacilitated by high GABA concentrations, independent ofilobalide concentrations. These results show that the re-overy of the receptor from bilobalide antagonism wasaster at high GABA concentrations suggesting that chan-el activation by GABA is required for the dissociation ofilobalide from �1 GABAC receptors.
Bilobalide exhibits similar but distinct pharmacologicalrofile to picrotoxinin at �1�2�2L GABAA and �1 GABAC
eceptors. It was less potent than picrotoxinin at both re-eptors. Both compounds largely exert noncompetitive an-agonism at �1�2�2L GABAA receptors (Huang et al., 2003)nd mixed antagonism with a weak use-dependent actiont �1 GABAC receptors. To date, they appear to differ inheir use-dependent action at GABAA receptors. Picrotox-nin showed use-dependent action on GABAA receptors ofat sympathetic neurons (Newland and Cull-Candy, 1992)nd bilobalide use-independent action on GABAA recep-
ors of rat cortical slices (Ivic et al., 2003).At �1 GABAC receptors, the potency of bilobalide de-
reased significantly as the concentrations of GABA in-reased (0.5 �M, 1.2 �M, 3 �M and 10 �M GABA). Inontrast, at �1�2�2L GABAA receptors, there was no sig-ificant difference in the potency of bilobalide between0 �M and 40 �M GABA. However, the potency of bilob-lide decreased approximately two times as the concen-rations of GABA increased to 100 �M and above (noignificant difference in its potency between 100 �M,00 �M and 1 mM GABA). The discrepancy in the potencyrofiles suggests that bilobalide interacts differently with
he subunits of �1�2�2L GABAA and �1 GABAC receptors.A recent study showed that picrotoxinin accelerated
elaxation of GABAC receptors (Qian et al., 2005). Qiannd coworkers adopted a model of picrotoxinin inhibitionhat incorporates its effect on both agonist binding andhannel gating of the GABAC receptors. In this modelicrotoxinin interacts with the ligand-bound receptor com-lex in both channel closed and channel open statesSmart and Constanti, 1986). The results from the modeluggested that the affinity of picrotoxinin for the receptor isltered by receptor channel gating inferring different affinityf picrotoxinin for the receptor in the open and closedtates. For example, high picrotoxinin affinity for channelpen states of GABAA receptors will generate predomi-antly noncompetitive inhibition, whereas high picrotoxininffinity for channel closed states will exhibit mostly com-etitive inhibition (Qian et al., 2005). Bilobalide may have aigher affinity for channel closed states of GABA receptorshan picrotoxinin as it exhibited a significantly greater vari-tion in the potency with respect to GABA concentrationst both �1 GABAC receptors (Table 1) and �2�2�2L GABAA
eceptors (Huang et al., 2003). The difference in the affinityf bilobalide and picrotoxinin for the channel states may bereflection of the structural variation that determines howcompound binds to the receptor.
It is noteworthy that Smart and Constanti (1986) de-eloped the model to explain mixed antagonism of picro-oxinin on lobster muscle GABA receptors and recently,
ultured lobster thoracic neurons have been shown toxhibit similar pharmacology to GABAC receptors (Jackelt al., 1994a,b). Although, it has yet to be cloned, it noweems likely that the crustacean muscle GABA receptorsharacterized pharmacologically 30 years ago and onhich the “mixed” model for picrotoxinin was formulatedave a high sequence homology with GABAC receptors.
CONCLUSION
n conclusion, the antagonism profile of bilobalide is similaro picrotoxinin at ionotropic GABA receptors including non-ompetitive antagonism at GABAA receptors and mixedntagonism with an use-dependent action at �1 GABAC
eceptors. With many pharmacological features in com-on with picrotoxinin at ionotropic GABA receptors
Huang et al., 2003; Ivic et al., 2003), bilobalide could besed as an alternative pharmacological tool or togetherith picrotoxinin to identify and differentiate betweenABA receptor subtypes.
cknowledgment—We thank Dr. George Uhl, National Institute forrug Abuse, Baltimore, MD, USA for human �1 cDNA. Dr. Hueran, Kong Li, Dr. Erica Campbell and Suzanne Habjan, Depart-ent of Pharmacology, the University of Sydney for technicalssistance, and the National Health and Medical Research Coun-il of Australia for financial support.
REFERENCES
merican Botany Council (2000). Ginkgo biloba leaf extract. In: Herbalmedicine: expanded Commission E monograph: herb mono-graphs, based on those created by a special expert committee ofthe German Federal Institute for Drugs and Medical Devices (Blu-menthal M, Goldberg A, Brinckman J, eds), pp 160–169. NewtonMA, USA: Integrative medicines communications.
arland JE, Moore AM, Hanrahan JR, Mewett KN, Duke RK, JohnstonGAR, Chebib M (2004) Mutations of the 2= praline in the M2domain of the human GABAC �1 subunit alter agonist responses.Neuropharmacology 46:770–781.
asida JE (1993) Insecticide action at the GABA-gated chloridechannel: recognition, progress, and prospects. Arch Insect Bio-chem 22:13–23.
hebib M, Mewett KN, Johnston GAR (1998) GABAC receptor antag-onists differentiate between human �1 and �2 receptors expressedin Xenopus oocytes. Eur J Pharmacol 357:227–234.
onstanti A (1978) The “mixed” effect of picrotoxin on the GABAdose/conductance relation recorded from lobster muscle. Neuro-pharmacology 17:159–167.
avies JA, Johns L, Jones FA (2003) Effects of bilobalide oncerebral amino acid neurotransmission. Pharmacopsychiatry36(Suppl 1):S84 –S88.
rew CA, Johnston GAR, Weatherby RP (1984) Bicuculline-insensi-tive GABA receptors: studies on the binding of (-)-baclofen to ratcerebellar membranes. Neurosci Lett 52:317–321.
uke RK, Chebib M, Allan RD, Mewett KN, Johnston GAR (2000) (�)-And (-)-CAMP (cis-2-aminomethylcyclopropanecarboxylic acid),show opposite pharmacology at recombinant �1 and �2 GABAC
receptors. J Neurochem 75:2602–2610.nz R, Cutting GR (1999) GABAC receptor � subunits are heteroge-
neously expressed in the human CNS and form homo- and heteroo-ligomers with distinct physical properties. Eur J Neurosci 11:41–50.
outman JD, Calvo DJ (2004) Studies on the mechanisms of action ofpicrotoxin, quercetin and pregnanolone at the GABA� receptor.
1Br J Pharmacol 141:717–727.

H
I
J
J
J
J
J
J
J
J
K
K
M
M
M
N
P
P
Q
Q
Q
Q
R
S
S
S
S
S
S
W
W
W
Y
Z
Z
Z
S. H. Huang et al. / Neuroscience 137 (2006) 607–617 617
uang SH, Duke RK, Chebib M, Sasaki K, Wada K, Johnston GAR(2003) Bilobalide, a sesquiterpene trilactone from Ginkgo biloba, is anantagonist at recombinant �1�2�2L GABAA receptors. Eur J Pharma-col 464:1–8.
vic L, Sands TTJ, Fishkin N, Nakanishi K, Kriegstein AR, Strømgaard K(2003) Terpene trilactones from Ginkgo biloba are antagonists ofcortical glycine and GABAA receptors. J Biol Chem 278:49279–49285.
ackel C, Krenz WD, Nagy F (1994a) Bicuculline/baclofen-insensitiveGABA response in crustacean neurons in culture. J Exp Biol 191:167–193.
ackel C, Krenz WD, Nagy F (1994b) A receptor with GABAC-like phar-macology in invertebrate neurons in culture. Neuroreport 5:1097–1101.
arboe CH, Porter LA, Buckler RT (1968) Structural aspects of picro-toxinin action. J Med Chem 2:729–731.
ohnston GAR (1996) GABAA receptors pharmacology. PharmacolTher 69:173–198.
ohnston GAR (2002) Medicinal chemistry and molecular pharmacol-ogy of GABAC receptors. Curr Top Med Chem 2:903–913.
ohnston GAR, Chebib M, Hanrahan JR, Mewett KN (2003) GABAC
receptors as drug targets. Curr Drug Targets CNS Neurol Dis 2:260–268.
ohnston GAR, Curtis DR, Beart PM, Game CJA, Mcculloch RM,Twitchin B (1975) Cis- and trans-4-aminocrotonic acid as GABAanalogues of restricted conformation. J Neurochem 24:157–160.
ones FA, Chatterjee SS, Davies JA (2002) Effects of bilobalide onamino acid release and electrophysiology of cortical slices. AminoAcids 22:369–379.
usama T, Spivak DE, Whiting PJ, Dawson VL, Schaeffer JC, Uhl GR(1993a) Pharmacology of GABA�1 and GABA�/� receptors expressedin Xenopus oocytes and Cos cells. Br J Pharmacol 109:200–206.
usama T, Wang TL, Guggino WB, Cutting GR, Uhl GR (1993b)GABA �2 receptor pharmacological profile: GABA recognition sitesimilarities to �1. Eur J Pharmacol Mol Pharmaco Sect 245:83–84.
acdonald RL, Olsen RW (1994) GABAA receptor channels. AnnuRev Neurosci 17:569–602.
uratra Y, Woodward RM, Miledi R, Overman LE (1996) The firstselective antagonist for a GABAC receptor. Bioorg Med Chem Lett6:2071–2076.
illigan CJ, Buckley NJ, Garret M, Deuchars J, Deuchars SA (2004)Evidence for inhibition mediated by coassembly of GABAA andGABAC receptor subunits in native central neurons. J Neurosci24:7241–7250.
ewland CF, Cull-Candy SG (1992) On the mechanism of action ofpicrotoxin on GABA receptor channels in dissociated sympatheticneurons of the rat. J Physiol (Lond) 447:191–213.
an ZH, Zhang D, Zhang X, Lipton SA (2000) Evidence for coassem-bly of mutant GABAC rho 1 with GABAA gamma2S, glycine alpha1and glycine alpha2 receptor subunits in vitro. Eur J Neurosci12:3137–3145.
erry NB, Aiyaz M, Kerr DS, Lake RJ, Leach MT (2001) NOESY onneurotoxins: NMR and conformational assignments of picrotoxins.Phytochem Anal 12:69–72.
ian H, Dowling JE (1994) Pharmacology of novel GABA receptorsfound on rod horizontal cells of the white perch retina. J Neurosci
14:4299–4307.ian H, Pan Y, Zhu Y, Khalili P (2005) Picrotoxin accelerates relax-ation of GABAC receptors. Mol Pharmacol 67:470–479.
ian H, Ripps H (1999) Response kinetics and pharmacological prop-erties of heteromeric receptors formed by coassembly of GABA�- and �2-subunits. Proc R Soc Lond B Biol 266:2419–2425.
ian H, Ripps H (2001) The GABAC receptors of retinal neurons. ProgBrain Res 131:295–308.
agozzino D, Woodward RM, Murata F, Eusebi F, Overman LE, MilediR (1996) Design and in vitro pharmacology of a selective �-ami-nobutyric acidC receptor antagonist. Mol Pharmacol 50:1024–1030.
asaki K, Hatta S, Wada K, Ohshika H, Haga M (1995) Anticonvulsantactivity of bilobalide, a sesquiterpene in Ginkgo biloba L. leavesagainst chemical-induced and electroshock-induced convulsionsin mice. Res Commun Biol Psychol Psychiatry 20:145–156.
asaki K, Hatta S, Wada K, Ohshika H, Haga M (1997) Bilobalide, aconstituent of Ginkgo biloba L., potentiates drug-metabolizing en-zyme activities in mice: possible mechanism for anticonvulsantactivity against 4-O-methylpyridoxine-induced convulsions. ResCommun Mol Pathol Pharmacol 96:45–56.
asaki K, Hatta S, Wada K, Haga M, Ohshika H (1999) Effects ofbilobalide on �-aminobutyric acid levels and glutamic acid decar-boxylase in mouse brain. Eur J Pharmacol 367:165–173.
edelnikova A, Smith CD, Zakharkin SO, Davis D, Weiss DS, ChangY (2005) Mapping the �1 GABAC receptor agonist binding pocket:constructing a complete model. J Biol Chem 280:1535–1542.
mart TG, Constanti A (1986) Studies on the mechanism of action ofpicrotoxinin and other convulsants at the crustacean muscle GABAreceptors. Proc R Soc Lond B Biol 227:191–216.
trata R, Cherubini E (1994) Transient expression of a novel type ofGABA response in rat CA3 hippocampal neurons during develop-ment. J Physiol (Lond) 480:493–503.
ada K, Sasaki K, Miura K, Yagi M, Kubabta Y, Matumoto T, Haga M(1993) Bilobalide and ginkgolide A, isolated from Ginkgo biloba L.shortened the sleeping time induced in mice by anesthetics. BiolPharm Bull 16:210–212.
ang T-L, Hackam AS, Guggino WA, Cutting GR (1995) A singleamino acid in �-aminobutyric acid �1 receptors affects competitiveand noncompetitive components of picrotoxin inhibition. Proc NatlAcad Sci USA 92:11751–11755.
oodward RM, Polenzani L, Miledi P (1992) Characterization of bicu-culline/baclofen-insensitive �-aminobutyric acid receptors ex-pressed in Xenopus oocytes. 1. Effects of Cl- channel inhibitors.Mol Pharmacol 42:165–173.
oon K-W, Covey DF, Rothman SM (1993) Multiple mechanisms ofpicrotoxin block of GABA induced currents in rat hippocampalneurons. J Physiol 464:423–439.
hang D, Pan ZH, Awobuluyi M, Lipton SA (2001) Structure andfunction of GABAC receptors: a comparison of native versus re-combinant receptors. Trends Pharmacol Sci 22:121–131.
hang D, Pan ZH, Zhang X, Brideau AD, Lipton SA (1995) Cloning of a�-aminobutyric acid type C receptor subunit in rat retina with a me-thionine residue critical for picrotoxinin channel block. Proc Natl AcadSci USA 92:11756–11760.
horov BS, Bregestovski PD (2000) Chloride channels of glycine andGABA receptors with blockers: Monte Carlo minimization and
structure-activity relationships. Biophys J 78:1786–1803.(Accepted 26 August 2005)(Available online 21 November 2005)





























