Embed Size (px)
Citation preview

Microb Ecol (1992)23:279-301 MICROBIAL ECOLOGY © Springer-Verlag New York Inc. 1992
Microbiology of Vadose Zone Paleosols in South-Central Washington State
Fred J. Brockman, 1 Thomas L. Kieft, 2 James K. Fredrickson, ~ Bruce N. Bjornstad, 1 Shu-mei W. Li, ~ Walt Spangenburg, 3 and Philip E. Long ~
~Pacific Northwest Laboratory, Richland, Washington 99352; 2New Mexico Institute of Mining and Technology, Socorro, New Mexico 87801; and 3Spokane Falls Community College, Spokane, Washington 99635, USA
Received: September 10, 1991; Revised, January 8, 1992
Abstract. Three unsaturated subsurface paleosols influenced by moisture recharge, including a highly developed calcic paleosol, were studied to investigate the microbiology of paleosols. Two near-surface paleosols, one impacted by moisture recharge and the other beyond the influence of re- charge, were also sampled to directly assess the effect of moisture recharge on the activity and composition of the microbial community associated with paleosols. The highly developed paleosol had a higher population of culturable heterotrophs, a greater glucose mineralization potential, a higher microbial diversity based on colony morphology, and a more than 20-fold higher concentration of ATP than the two weakly developed paleosols. The recharged near-surface paleosol, as compared to the near-surface paleosol unaffected by recharge, had a lower population of culturable heterotrophs, smaller mineralization rate constant, and lower richness based on colony morphology. The recharged paleosols contained predominantly gram-neg- ative isolates, whereas the paleosol unaffected by recharge contained pre- dominantly gram-positive isolates. Storage at 4°C of subsurface and near- surface paleosol samples containing high water potential increased the population ofculturable aerobic heterotrophs, decreased diversity in colony morphology, and increased first-order rate constants and decreased lag times for glucose mineralization. These results indicate that aerobic het- erotrophs are present in deep vadose zone paleosols and that there is po- tential for stimulation of their in situ growth and activity.
Introduction
Recent results have indicated that diverse and metabolically active microbial populations are present within deep saturated zones of the southeastern Atlantic coastal plain [2, 3, 13, 14, 19-21, 29, 32, 38, 39, 44, 49]. Because the climate,
Offprint requests to: F. J. Brockman.
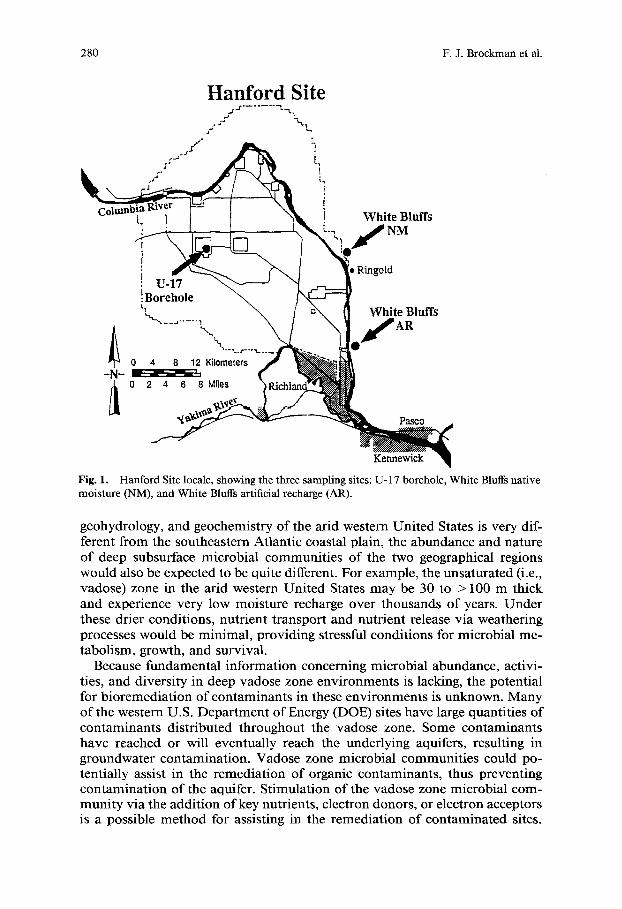
280 F.J. Broekman et al.
Colur~
Hanford Site ..r .,f . . . . . . . - L . x
,.a 1 -,-'" \L ,
. a
_ j r " e-a j"
:l
t, I
1
I
i
J U-17
IBorehole
"~'k~,..~.~_
0 4 8 12 Ki lomete rs - N - B E E ~ =
0 2 4 6 8 Miles
White Bluffs
Ringold
White Bluffs . ~ A R
Pasco
Kennewick •
Fig. 1. Hanford Site locale, showing the three sampling sites: U-17 borehole, White Bluffs native moisture (NM), and White Bluffs artificial recharge (AR).
geohydrology, and geochemistry of the arid western United States is very dif- ferent from the southeastern Atlantic coastal plain, the abundance and nature of deep subsurface microbial communities of the two geographical regions would also be expected to be quite different. For example, the unsaturated (i.e., vadose) zone in the arid western United States may be 30 to > 100 m thick and experience very low moisture recharge over thousands of years. Under these drier conditions, nutrient transport and nutrient release via weathering processes would be minimal, providing stressful conditions for microbial me- tabolism, growth, and survival.
Because fundamental information concerning microbial abundance, activi- ties, and diversity in deep vadose zone environments is lacking, the potential for bioremediation of contaminants in these environments is unknown. Many of the western U.S. Department of Energy (DOE) sites have large quantities of contaminants distributed throughout the vadose zone. Some contaminants have reached or will eventually reach the underlying aquifers, resulting in groundwater contamination. Vadose zone microbial communities could po- tentially assist in the remediation of organic contaminants, thus preventing contamination of the aquifer. Stimulation of the vadose zone microbial com- munity via the addition of key nutrients, electron donors, or electron acceptors is a possible method for assisting in the remediation of contaminated sites.

Microbiology of Vadose Zone Paleosols 281
H o w e v e r , t he p r e s e n c e o f v i a b l e m i c r o o r g a n i s m s in, a n d the effect o f n u t r i e n t a d d i t i o n s on , d e e p v a d o s e zone m i c r o b i a l c o m m u n i t i e s has n o t been s tud ied .
T h e o b j e c t i v e s o f t h i s s t u d y were to i n v e s t i g a t e t he m i c r o b i o l o g y o f unsa t - u r a t e d subsu r f ace s e d i m e n t s , focus ing on pa leoso l s , a n d to assess t he effect o f m o i s t u r e r echa rge on v a d o s e z o n e m i c r o b i a l c o m m u n i t i e s . Pa l eoso l s a re sed- i m e n t s t ha t were s u b j e c t e d to soi l d e v e l o p m e n t p roces ses a t the sur face p r i o r to b u r i a l and , as a resul t , m a y c o n t a i n e l e v a t e d c o n c e n t r a t i o n s o f o rga n i c c a r b o n o r c a l c i u m c a r b o n a t e a n d m o r e a d v a n c e d s t ruc tu ra l d e v e l o p m e n t . W e h y p o t h - e s i zed t h a t p a l e o s o l s m a y be subsu r face zones w h e r e m i c r o b i a l s u r v i v a l o r a c t i v i t y is e n h a n c e d b y e l e v a t e d c o n c e n t r a t i o n s o f o rgan ic nu t r i en t s . T h e s e n u t r i e n t s m a y be u t i l i z e d for g r o w t h a n d m a i n t e n a n c e o v e r geo log ica l t i m e p e r i o d s , p a r t i c u l a r l y d u r i n g e p i s o d i c m o i s t u r e r echarge even t s , b y d e s i c c a t i o n - t o l e r a n t m i c r o b e s . T h e in f luence o f m o i s t u r e r echarge in the v a d o s e zone is an i m p o r t a n t c o n s i d e r a t i o n at D O E ' s a r i d H a n f o r d Si te in s o u t h - c e n t r a l W a s h - i ng ton State , b e c a u s e ac t i v i t i e s t he re h a v e d i s c h a r g e d m o r e t h a n 8.6 x 1011 l i ters o f w a t e r f r o m the C o l u m b i a R i v e r to the u p p e r v a d o s e zone [1 1]. M o v e - m e n t o f w a t e r i n to d e e p v a d o s e zone s e d i m e n t s w o u l d i nc rea se t he b i o a v a i l - a b i l i t y a n d t r a n s p o r t o f s e d i m e n t - a s s o c i a t e d nu t r i en t s , i n t r o d u c e d i s s o l v e d nu- t r i en t s a n d c o n t a m i n a n t s , a n d p o t e n t i a l l y i n t r o d u c e sur face m i c r o o r g a n i s m s i n to d e e p s e d i m e n t s . T o d e t e r m i n e the effects o f m o i s t u r e r echarge on the m i c r o b i o l o g y o f d e e p l y b u r i e d s e d i m e n t s , a n e a r - s u r f a c e p a l e o s o l i n f luenced b y r echa rge was c o m p a r e d to the i d e n t i c a l p a l e o s o l a t a l o c a t i o n unaf fec ted b y recharge .
Mater ia l s and M e t h o d s
Geologic Setting and Sampling Rationale
Samples were collected at three depths from a borehole designated U-17 on the Hanford Site, and at two near-surface sites along the White Bluffs, located east of the Hanford Site (Fig. 1). All sediment samples that were collected in this study were identified as paleosols based on soil development characteristics. Hereafter, the generic term "sediment" will include these paleosols.
Vadose zone strata sampled at the U-17 borehole were selected on the basis of sediment de- scriptions in nearby boreholes. These strata, from oldest to youngest, included a fine-textured fluvial sediment (upper Ringold unit; sampled at 63.7-m depth; hereafter termed the 64-m sedi- ment), a buried calcic paleosol sediment (Plio-Pleistocene unit; sampled at 56.7-m depth; hereafter termed the 57-m sediment), and an eolian (wind-deposited) sediment (early "Palouse" unit; sam- pled at 53.6-m depth; hereafter termed the 54-m sediment) (Fig. 2). The ages of these sediments varied from >- 4 million years for the Ringold Formation to 1-4 million years for the Plio-Pleistocene and early "Palouse" units [17]. These sediments are overlain by Pleistocene cataclysmic flood deposits, informally referred to as the Hanford formation [35, 52].
The upper Ringold unit at U- 17 consisted of a moderately calcareous, yellowish brown, fine sandy mud (mud: a field description for combined silt and clay of undetermined composition). Very weak soil development was indicated by the presence of a few relict root traces toward the top of this unit; however, most primary sedimentary stratification was preserved. The fine-grained upper Ringold sequence graded downward into coarser-grained, gravelly sediments of the middle Ringold unit (Fig. 2).
The Plio-Pleistocene unit [5, 6] consisted of a mixture of light-colored, cemented sand and mud, about 6 m thick, which represents a highly weathered paleosol [9, 12]. Pedogenic CaCO3 cement, commonly associated with near-surface soil development in arid environments [25, 40], often

282 F .J . Brockman et al.
Depth Stratigraphy (m below
surface)
Natural Gamma Log (cps)
Lithology sand
gr . . . . , mud
-- 36.6
HANFORD FORMATION
-- 42.7
EARLY "PALOUSE"
SOIL
PLIO- PLEISTOCENE
UNIT
- 48.8
- 54.9
Upper - 67.1
Unit I Ringold
Middle - 73.2 Ringold ~ Unit
w a t e r ~ , t a b l e
'edogenesis 1 Weak soil development Advanced soil develop. (calcic)
Root traces
53.6
56.7
- 63.7
Fig. 2. Stratigraphy and lithology of the sampled intervals at the U-17 borehole.
exceeds 30% (w/w) within the Plio-Pleistocene unit. In places, carbonate cement completely fills the voids between sedimentary particles, forming a relatively impermeable calcrete. Other features indicative of soil development include platy to prismatic structure, animal burrows, and numerous root traces. The abundance of root traces is significant in that they represent areas of extensive microbial colonization in the geologic past.
Overlying the Plio-Pleistocene unit is a sequence of unconsolidated silty sand to fine sandy silt sediments, approximately 6 m thick, which belongs to a unit referred to as the early "Palouse" soil [52]. Aggradation of this unit apparently occurred relatively quickly, probably from physical reworking of the underlying sediments [ 10], as no well-developed soil horizonation is present. Core samples of the early "Palouse" soil have the texture, structure, and color of modern loess (windblown silt and sand) deposits, which locally blanket the surface of the region. This unit therefore is believed to represent an early Pleistocene eolian deposit. Very weak soil development is indicated by the presence of a few relict root traces.
Samples were also collected from near-surface sediments of the Ringold Formation along the White Bluffs, located 30 km east of the U-17 borehole on the east side of the Columbia River (Fig. 1). The average annual precipitation at White Bluffs is 17 cm. The White Bluffs sediments are geologically similar to the 64-m sediment at U-17 in that both contain upper Ringold parent

Microbiology of Vadose Zone Paleosols 283
material. However, based on regional stratigraphic correlations, the White Bluffs sediments are probably younger than the upper Ringold unit at U-17. Near-surface sediments known to be influenced by artificial recharge (AR) and others not influenced by artificial recharge (i.e., under native moisture, NM) were collected from the same paleosol unit, as indicated by stratigraphic position with respect to a distinctive tuff/diatomite sequence (Fig. 3). The sampling of the same paleosol in the presence and absence of artificial recharge allowed for a direct comparison of the effect of moisture recharge on the microbial community. The paleosol was weakly developed and consisted of an olive-colored clayey silt to silty clay with root traces and a well-developed blocky to prismatic (i.e., pedogenic) texture. AR samples were collected from a recently exposed, vertical, landslide scarp induced by over 40 years of lateral seepage of irrigation water [41]. NM samples were collected from the identical paleosol at a site not influenced by drainage of irrigation water, approximately 20 km to the north (Fig. 1).
Sample Acquisition
Sediment samples were collected during the installation of groundwater monitoring well U-17. Sediment was monitored during the process of drilling for the presence of organic vapors and alpha and beta radiation. Sediment was also analyzed for total alpha and beta radiation by liquid scin- tillation counting (model LS7000; Beckman Instruments, Fullerton, CA) prior to physical, chemical, and microbiological analysis. Drilling was accomplished using reverse air-circulation rotary coring (Becker Industries, Denver, CO). At the designated depths, samples were collected via percussion hammer by driving a 10-cm inside diameter split-barrel sampler 25-30 cm into the formation. Stainless steel liners used inside the sampler were wrapped in foil and autoclaved prior to use. The split barrel, core catcher, and drive shoe were alcohol-flamed for disinfection in the field. The apparatus was assembled and the end covered with sterile foil prior to sampling. The retrieved cores were immediately covered and sealed with sterile plastic bags and transported to the laboratory on ice.
At the White Bluffs sites, cores were sampled from a paleosol 1.0-1.2 m below a distinctive tuff/ diatomite sequence. At the NM site, approximately 0.7 m of talus was excavated from directly below the exposed tuff/diatomite sequence and a fresh face exposed by digging into the face approximately 0.3 m. An autoclaved aluminum liner (4.75 cm inside diameter, l0 cm long) was placed inside the shoe of a manually operated percussion device (Forestry Suppliers Inc., Jackson, MS) and driven into the face. At the AR site, a fresh face was exposed by prying offlarge prismatic blocks of sediment, digging into the face approximately 0.3 m, and sampling in the manner described above. Sediment samples were collected from the NM and AR sites on two (AR) or three (NM) different occasions. At each occasion, a cluster of three cores within centimeters of each other, and a second cluster of three cores within centimeters of each other from the same stratum one lateral meter away, were collected to examine spatial variability of the microbial community. Samples for physical and chemical analysis (BB90-10, BB90-15, and BB90-16; Fig. 3) were collected adjacent to the microbiological samples. All samples were placed in sterile plastic bags, sealed, and placed in a cooler containing ice for transport to the laboratory. Sediment temperatures were measured by driving a nail of appropriate size 10 cm into the sediment face, removing the nail, and inserting the probe end of a digital thermometer into the bottom of the hole.
Physical and Chemical Analysis
Detailed sample descriptions were recorded by a geologist at the drill site or sampling site. Sediment samples collected for moisture content were placed in airtight containers such that little or no headspace was present. Moisture content was determined gravimetrically by drying approximately 450 g of sediment at 105°C for 48 hours. Particle size was analyzed by a combination of dry sieving and hydrometer methods [23]. Sediments were analyzed for total organic carbon (TOC) (model CR-12 Carbon Determinator, Leco Corp., St. Joseph, MI), total Kjeldahl nitrogen (method 351.2;

284 F.J . Brockman et al.
ref. 18), total phosphorus (method 365.1; ref. 18), and carbonates (method 91-4; ref. 7) by Twin City Testing Corp. (St. Paul, MN). Water potential was measured with a thermocouple psychrometer (model SC-10A, Decagon Devices, Pullman, WA).
Sample Preparation, Plate Counts, and Direct Counts
Metal liners containing the cores were placed in a laminar flow hood and sediment was removed from the center of the cores (inner 6-cm diameter, U-17 cores; inner 3-cm diameter, White Bluffs cores) with a sterile spoon and/or screwdriver. Removal of sediment from the metal liners resulted in a large amount of fracturing and disturbance. For this reason, sediment removed from the center of the core was thoroughly mixed prior to microbiological analyses. The single exception was that one-gram portions of intact White Bluffs sediments were used for the initial enumerations of culturable bacteria (to allow greater sensitivity in examining spatial variability). Homogenized sediment was double-bagged and sealed in sterile plastic bags and stored at 4°C. Two clusters of cores from the NM and AR sites (i.e., 6 cores from each site) were analyzed for microbiological characteristics. Plate count enumeration of microorganisms was also performed on the other six NM and 12 AR cores that were sampled.
Culturable heterotrophs were enumerated by adding 10 g of sediment to 95 ml 0.1% sodium pyrophosphate (pH 7.0) in a sterile blender container, subjecting the sediment to two 30-sec bursts in the blender, placing the suspension in a cup and shaking at 120 rpm for 15 rain, and performing standard dilution series in 0.1% pyrophosphate. Heterotrophs were cultured on 1% peptone-tryp- tone-yeast-glucose (PTYG) medium [4]. The U-17 sediments were also plated on Lit tman Oxgall and Actinomycetes Isolation media (Difco) for selection of fungi and actinomycetes. For White Bluffs sediments, one g of sediment was placed in 9.5 ml 0.1% pyrophosphate and subjected to vigorous vortexing for 1 rain before further dilution. All sediment samples were plated in duplicate, beginning with a 10 -1 dilution of the sediment, and plates were incubated at 28°C for 30-40 days before colonies were enumerated. Populations of culturable heterotrophs were log-transformed for statistical analysis. Microorganisms were also enumerated on 1% PTYG after storing sediment at 4°C for various lengths of time.
Diversity indices were based on colony morphology characteristics and were calculated from 1% PTYG plates containing 30-300 colonies, except for a few instances where fewer than 30 colonies were present at the lowest dilution. Richness was used as a measure of the number of different colony morphologies, whereas equitability was used as a measure of how individual colony mor- phologies were proportioned among the total number of colonies, and the Shannon-Weaver index was used as an indicator of both richness and equitability [1 ]. Larger values of the index indicate greater diversity.
Direct microscopic counts of total bacteria were performed by the acridine-orange (AO) method [24], with modifications [49]. The numbers of actively respiring microorganisms were estimated as the number of AO-stained cells capable of reducing INT [2-(p-iodophenyl)-3-(p-nitrophenyl)- 5-phenyl tetrazolium chloride] [57], as modified by Webster et al. [54]. For both direct enumeration methods, 50 microscopic fields from sample and control slides were counted at 630 × magnification, yielding a min imum detection limit of approximately log 4.6 cells g-i moist sediment. Samples were prepared for direct microscopic counts at New Mexico Institute of Mining and Technology within 24 hours of receipt by overnight express mail, and after sediments were stored at 4°C for various lengths of time.
Nutrient Addition Experiments
Upon sample acquisition, 20-g portions of U-17 54-m and 64-m sediments were incubated at room temperature under native and saturated moisture regimes with four nutrient amendment treatments to assess potential microbial activity and the ability to stimulate activity by nutrient addition, Ambient moisture regime treatments consisted of 0.4 ml of water containing either no amendment (control); 200 ~g acetate; inorganics (one-tenth strength Shelton's mineral salts medium

Microbiology of Vadose Zone Paleosols 285
containing 40 ug nitrate; ref. 48); and acetate plus inorganics. Moisture release curves indicated that the experimental procedure resulted in a water potential that was -<-0.1 MPa less negative. An autoclaved sediment control with acetate plus inorganics was also employed. The amendment was dripped evenly over the sediment in a 175-ml sterile milk dilution bottle and the bottle agitated to mix thoroughly. Treatments for the saturated moisture regime were identical except that 4.0 ml of amendments at one-tenth strength were added, so that the final amounts of nutrients added were the same for both moisture regimes. The amendment increased the water content of the sediments by approximately 16.7%, saturating the sediments. A sterile scintillation vial containing 10 ml of water was hung in each bottle (to prevent soil drying) with Teflon tape. The bottles were sealed with a sterile rubber stopper wrapped with Teflon tape, and incubated at 22°C.
Adenosine 5'-triphosphate (ATP) was measured as an estimate of microbial biomass at sample acquisition, in one set of treatments after 3 days of incubation, and in a second set of treatments after 42 days of incubation. Triplicate 1-g aliquots of moist sediment were sampled from each treatment. ATP was extracted by adding 5 ml 0.5 M NaHCO3 (pH 8.5) and 1.5 ml chloroform to each aliquot of sediment, vortexing the mixture at high speed for 15 sec, adding 7.5 ml 0.5 M NaHCO3, and vortexing an additional 15 see. Eight ml was centrifuged at 2°C at 3,500 g, and 5 ml of the aqueous phase transferred to a tube. Traces of chloroform were removed by connecting tubes to a vacuum and incubating for 2 rain in a 60°C water bath. Tubes were capped, sealed with parafilm, and stored at -40°C until analysis. ATP was quantitated via the luciferin-luciferase assay (Sigma Chemical, St. Louis, MO) using a luminometer (model 1251, LKB Instruments, Gaithers- burg, MD). The detection limit was 110-150 ng ATP g-~ moist sediment. Culturable heterotrophs from the 42-day incubations were enumerated on 1% PTYG.
Effect o f Storage Time on Radiotracer Mineralization and Microbial Populations and Diversity
At sample acquisition, duplicate 1.5-g portions of sediment from U- 17 54-, 57-, and 64-m sediments were added to 8-ml serum vials, 10 #1 [14C(U)]-glucose (3.0 mCi mmo1-1, 99% pure; Sigma Chemical Co.) was added evenly to the sediment, and the sediment was agitated to mix thoroughly. The final concentration of glucose was 1.8 Izg g-~ moist sediment. Moisture release curves indicated that the experimental procedure resulted in a water potential that was <0.1 MPa less negative. A sterile polypropylene center well (Kontes, Vineland, N J) containing a strip of Whatman # 1 chro- matography paper and 0.3 ml of 0.4 M NaOH as a ]4CO2 trap was attached to the underside of a rubber stopper which served to seal the vial. The vials were incubated at 22°C for four weeks. Autoclaved sediment controls also received ~4C-glucose. NaOH was removed at 7-day intervals for scintillation counting (model 460C; Packard Instrument Co., Downers Grove, IL) and replaced with fresh 0.4 M NaOH.
After 97 days of storage at 4°C, 10-g portions of the same sediments were placed in 175-ml milk dilution bottles, 200 lz114C-glucose solution (total of 0.5 lzCi) was dripped evenly onto the sediment, and the bottle was agitated to mix thoroughly. The final concentration of glucose was 3.0 #g g moist sediment. Moisture release curves indicated that the experimental procedure resulted in a water potential that was -<0.1 MPa less negative. Autoclaved sediments were included as controls. A 7-ml sterile scintillation vial containing 1 ml of 0.3 M KOH was used to trap ~4CO2. Bottles were sealed and drying was prevented as described earlier, and the sediments were incubated at 22°C. KOH was removed periodically for scintillation counting (model LS7000; Beckman Instru- ments, Fullerton, CA) and replaced with fresh 0.3 M KOH.
Microbial characteristics of White Bluffs near-surface sediments were examined at sample ac- quisition and after 15, 42, and 75 days of storage at 4°C, using the protocol described for the U-17 sediments at 97 days. Moisture release curves indicated that the experimental procedure resulted in a water potential that was ca. 0.3 MPa and -<0.1 MPa less negative, respectively, for the NM and AR sediments.
First-order nonlinear regression of percent CO2 evolved [34] was modeled using a statistical analysis program (Systat, Evanston, IL) to obtain values for first-order mineralization rate constants
286 F .J . Brockman et al.
8
1::
, ~~ , ~
~ , ~
~ o
~ .,=
©
~ 0
~ - ~ v ~
d ~ N d ~ N d
M N M ~ d
d d N N £ d d ~ o i ~ ~ V ~
-'2"
0
0
©
o
0
o
e~
<
E 0
~ 0 ~, .~
oE~=

Microbiology of Vadose Zone Paleosols 287
White Bluffs NM Site decreasing particle size
BB90-10
White Bluffs AR Site decreasing Elevation particle size (m above
sea level)
i ~ 182.9
°t l I ~ BB90-15 BB90-16
152.4
No horizontal scale
sample no. - - • correlation line
[ ] diatomite 1 tuff
P e ~ e n i c Features blocky texture carbonate b b stringers
o carbonate Q nodules
q q root traces
Fig. 3. Stratigraphy and lithology of the near-surface White Bluffs native moisture and White Bluffs artificial recharge samples. The BB90- prefix indicates samples collected for physical and chemical analyses. Samples for microbiological analysis were collected at the locations indicated by samples BB90-10 and BB90-16.
and lag times. Culturable heterotrophs were enumerated on 1% PTYG and diversity of colony morphology was examined at the initiation of each radiotracer mineralization experiment.
Results
Physical and Chemical Characteristics
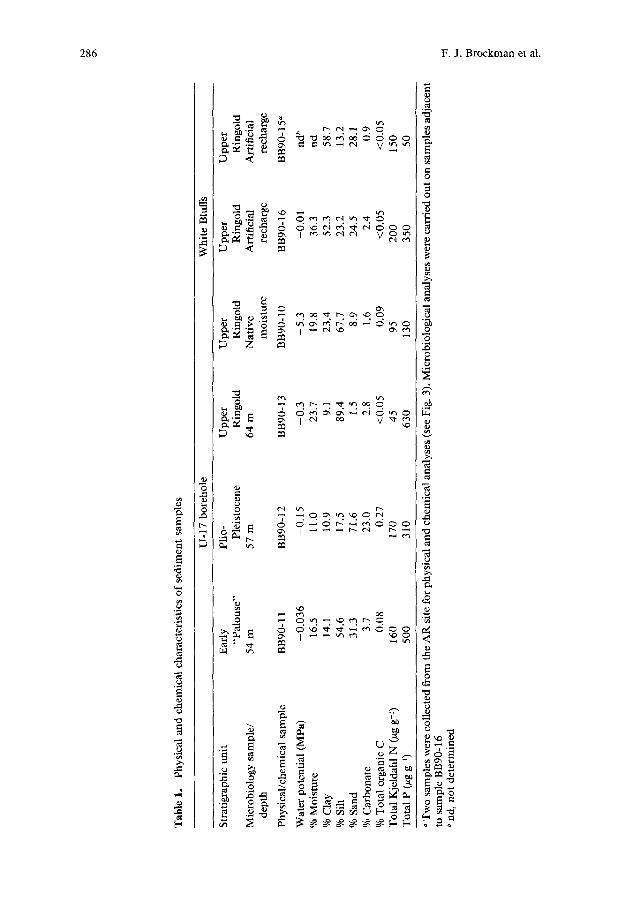
Strat igraphic and lithologic relat ionships o f the U- 17 sample intervals and near- surface sampl ing sites are shown in Figs. 2 and 3, respectively. Nei ther organic vapors nor radia t ion were detected during drilling, and liquid scintillation count ing indicated that none o f the sediments were con tamina ted with radio- act ive wastes. The results o f physical and chemical analyses are presented in Table 1. Water contents were highest for the upper Ringold sediments, es- pecially the White Bluffs AR, and the U-17 57-m sediment (highly developed paleosol) had the lowest water content. The lowest water potentials were as- sociated with the upper Ringold sediments in U-17 64-m and White Bluffs N M sediments , as a result o f their finer textures (i.e., higher silt/clay content). Sediment tempera tures at the t ime o f sampling at White Bluffs N M and A R sites were 21 and 19°C, respectively.

288 F.J. Brockman et al.
g
0 ©
e~
0
0
o
0
0
~o
©
e4
M d ~ N N
~ . ~ . ~
d d d d d
c 5 o ~ c 5 c ~
v-: .v-: .o. . .
v v v
<
<
.= .~ ~:
2 o Z ~
" c O II ~2
- ~
'~ :~ ~ o ~
~ o ~'-~
= II ~ ~ ~--,.=-,
, ~ o ~ ~

Microbiology of Vadose Zone Paleosols 289
Particle-size analysis of U-17 sediments revealed that the 54-m sediment was predominantly a mixture of sand and silt, and the 64-m sediment was predominantly silt. The 57-m sediment contained significantly more sand. However, the sand particles in this sample may have actually been aggregates of finer-grained silt and clay, cemented together with CaCO3. In the White Bluffs upper Ringold sediments, the AR sediment was predominantly clay with equal amounts of sand and silt, whereas the NM sediment was mostly silt. Percent carbonate was about an order of magnitude greater in the 57-m sedi- ment than in the other sediments, and is indicative of a highly developed paleosol. The 57-m sediment also contained the highest concentration of TOC. The White Bluffs NM sediment contained more TOC than the AR sediment. Total nitrogen and phosphorus did not vary systematically among the samples collected.
Plate Counts and Direct Counts
In U-17 borehole sediments, culturable heterotrophs in the 57-m sediment were more abundant than in the 54- and 64-m sediments (Table 2). In all 3 sediments, colony-forming units (CFU) were present at <log 1.0-1.5 g-1 moist sediment on Actinomycetes Isolation medium and at ---log 1.0 g-1 moist sediment on Littman Oxgall medium. Plating of 1-g portions of intact White Bluffs AR and NM sediments allowed examination of variability in the spatial distribution of culturable heterotrophs. For a particular sediment, variability both within a cluster of cores and among clusters was high, with standard deviations generally greater than the mean values, indicating a spatially het- erogeneous distribution ofculturable heterotrophs. Variability in the number of culturable heterotrophs was greater in the NM cores (log 2.0 to 4.7 CFU g-i dry sediment) than in the AR cores (log 1.9 to 2.9 CFU g-1 dry sediment). Culturable heterotrophs in the White Bluffs NM and AR cores that were fully characterized microbiologically were not statistically different (Table 2). How- ever, using data from all sampled cores, culturable heterotrophs were statis- tically greater in the NM sediments (log 3.2) than in the AR sediments (log 2.4) (data not shown).
Total microbial populations, as determined by acridine orange direct counts, were similar among all U-17 and White Bluffs sediments. Total microbial populations were approximately one order of magnitude greater than the num- ber of respiring cells (AODC-INT) in the U-17 sediments, and 1.5 to ->2.5 orders of magnitude greater than the number of respiring cells in the White Bluffs sediments (Table 2). Respiring cells were two to four orders of magnitude greater than plate counts, indicating that most of the metabolically active mi- croorganisms were nonculturable under the growth conditions used.
Gram Reaction and Diversity in Colony Morphology
Colony morphologies indicated that actinomycetes were the dominant micro- organisms in the NM sediment, whereas non-mycelial, non-sporeforming bac-
290 F.J. Brockman et al.
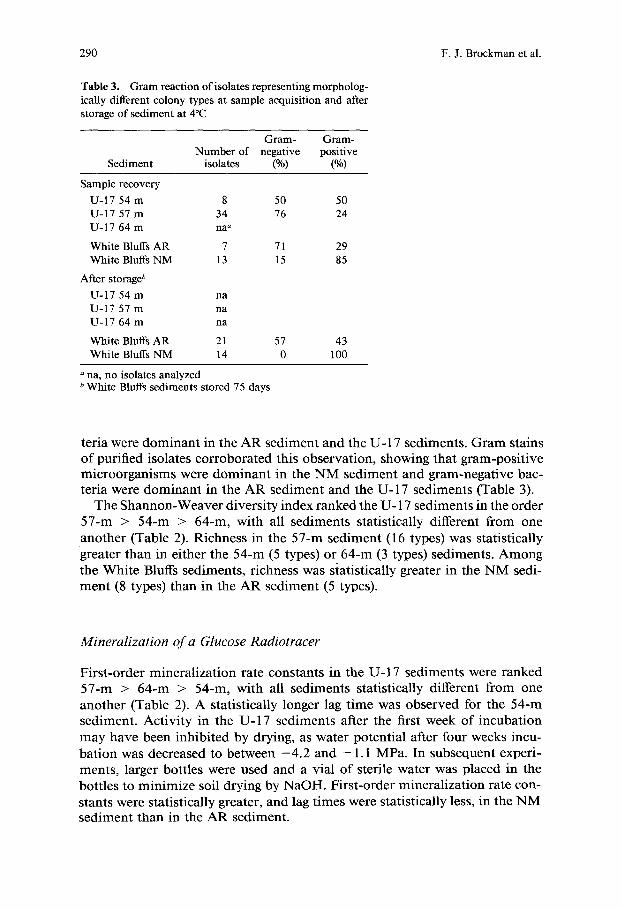
Table 3. Gram reaction of isolates representing morpholog- ically different colony types at sample acquisition and after storage of sediment at 4°C
Gram- Gram- Number of negative positive
Sediment isolates (%) (%)
Sample recovery U-17 54 m 8 50 50 U-17 57 m 34 76 24 U-17 64 m na ~
White Bluffs AR 7 71 29 White Bluffs NM 13 15 85
After storage b U-17 54 m na U-17 57 m na U-17 64 m na
White Bluffs AR 21 57 43 White Bluffs NM 14 0 100
ana, no isolates analyzed b White Bluffs sediments stored 75 days
teria were dominan t in the A R sediment and the U- 17 sediments. Gram stains o f purified isolates corroborated this observation, showing that gram-posit ive microorganisms were dominan t in the N M sediment and gram-negative bac- teria were dominan t in the AR sediment and the U-17 sediments (Table 3).
The Shannon-Weaver diversity index ranked the U- 17 sediments in the order 57-m > 54-m > 64-m, with all sediments statistically different from one another (Table 2). Richness in the 57-m sediment (16 types) was statistically greater than in either the 54-m (5 types) or 64-m (3 types) sediments. Among the White Bluffs sediments, richness was statistically greater in the N M sedi- ment (8 types) than in the A R sediment (5 types).
Mineralization of a Glucose Radiotracer
First-order mineralization rate constants in the U-17 sediments were ranked 57-m > 64-m > 54-m, with all sediments statistically different f rom one another (Table 2). A statistically longer lag t ime was observed for the 54-m sediment. Activity in the U-17 sediments after the first week of incubation may have been inhibited by drying, as water potential after four weeks incu- bation was decreased to between - 4 . 2 and - 1 . 1 MPa. In subsequent experi- ments, larger bottles were used and a vial o f sterile water was placed in the bottles to minimize soil drying by NaOH. First-order mineralization rate con- stants were statistically greater, and lag times were statistically less, in the N M sediment than in the A R sediment.
Microbiology of Vadose Zone Paleosols
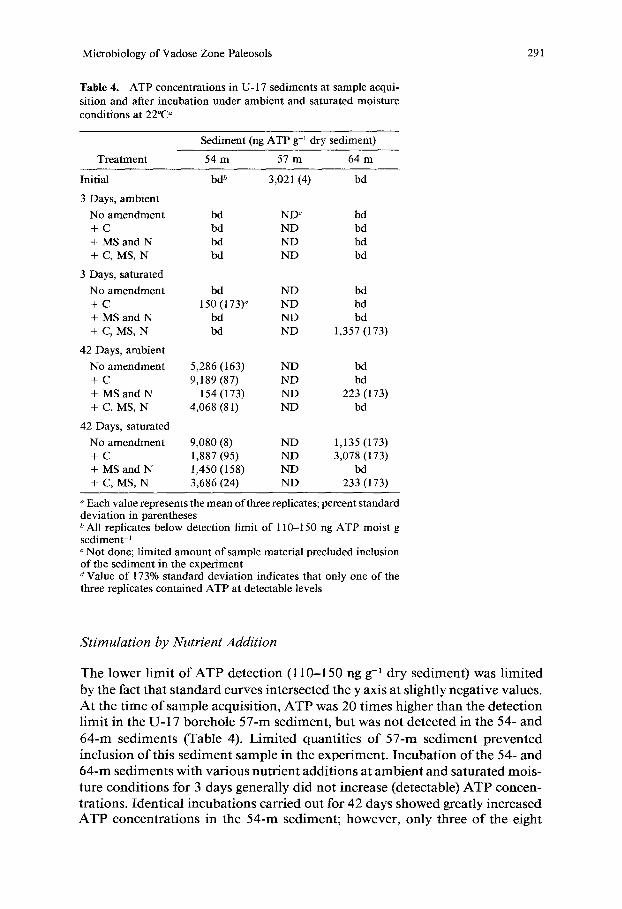
Table 4. ATP concentrations in U- 17 sediments at sample acqui- sition and after incubation under ambient and saturated moisture conditions at 22°C a
Sediment (ng ATP g-i dry sediment)
Treatment 54 m 57 m 64 m
Initial
3 Days, ambient
No amendment + C + MS and N + C, MS, N
3 Days, saturated
No amendment + C + MS and N + C, MS, N
42 Days, ambient
No amendment + C + MS and N + C, MS, N
42 Days, saturated
No amendment + C + MS and N + C, MS, N
b& 3,021 (4) bd
bd ND c bd bd ND bd bd ND bd bd ND bd
bd ND bd 150 (173) d ND bd
bd ND bd bd ND 1,357 (173)
5,286 (163) ND bd 9,189 (87) ND bd
154 (173) ND 223 (173) 4,068 (81) ND bd
9,080 (8) ND 1,135 (173) 1,887 (95) ND 3,078 (173) 1,450 (158) ND bd 3,686 (24) ND 233 (173)
Each value represents the mean of three replicates; percent standard deviation in parentheses b All replicates below detection limit of 110-150 ng ATP moist g sediment -~ c Not done; limited amount of sample material precluded inclusion of the sediment in the experiment a Value of 173% standard deviation indicates that only one of the three replicates contained ATP at detectable levels
291
Stimulation by Nutrient Addition
T h e lower l im i t o f A T P de tec t ion ( 1 1 0 - 1 5 0 ng g-i d ry sed imen t ) was l i m i t e d
by the fact tha t s t a n d a r d curves in te rsec ted the y axis at sl ightly nega t ive values.
A t the t i m e o f sample acqu i s i t ion , A T P was 20 t imes h igher t h a n the de tec t ion l im i t in the U - 1 7 bo reho le 5 7 - m sed imen t , b u t was n o t de tec ted in the 54- a n d
6 4 - m s e d i m e n t s (Table 4). L i m i t e d quan t i t i e s o f 5 7 - m s e d i m e n t p r e v e n t e d
i n c l u s i o n o f this s e d i m e n t s ample in the expe r imen t . I n c u b a t i o n o f the 54- a n d 6 4 - m s e d i m e n t s wi th va r ious n u t r i e n t add i t i o ns at a m b i e n t a n d sa tu ra ted mo i s -
ture c o n d i t i o n s for 3 days genera l ly d id n o t increase (detectable) A T P concen -
t ra t ions . Iden t i ca l i n c u b a t i o n s carr ied ou t for 42 days showed great ly inc reased A T P c o n c e n t r a t i o n s in the 5 4 - m sed imen t ; however , on ly three o f the eight

292 F.J. Brockman et al.
treatments contained detectable ATP in all three replicates. Incubation of the 64-m sediment for 42 days had a small effect on ATP concentrations, with one replicate in four of the eight treatments containing detectable ATP. ATP con- centrations in unamended (water only) controls were similar to those in the nutrient-amended treatments, and there were no obvious differences between the ambient and saturated moisture conditions. ATP concentrations generally varied widely among replicates of the same treatment, indicating that microbial activity and/or growth was probably localized to microsites that were distrib- uted heterogeneously.
Culturable heterotroph populations in the 54-m sediment after 42 days in- cubation ranged from log 5.2 to 6.5 CFU g-~ dry sediment in all treatments, with no noticeable differences due to amendments (including the unamended control) or different moisture conditions (data not shown). Based on morpho- logical characteristics, all treatments were dominated (84-100% of the colonies) by the same (or very similar) bacterium. The 64-m sediment, in general, did not contain greater populations of culturable heterotrophs after 42 days in- cubation. Populations as determined by plate count methods were below log 1.8 CFU g-' dry sediment in seven of the eight treatments; the remaining treatment (ambient moisture, unamended control) contained log 4.2 CFU g-~ dry sediment. Culturable microorganisms were below log 1.0 CFU g-~ moist sediment in autoclaved controls containing organic plus inorganic amendments after 42 days.
Effect of Storage Time on Microbial Numbers, Diversity in Colony Morphology, and Radiotracer Mineralization
Culturable heterotroph populations in the U-17 sediments were statistically different from each other after 83 days storage at 4°C (Table 2). Each of the sediments had statistically larger means after 83 days storage than at the time of sample acquisition (comparisons not shown). Increased heterotroph popu- lations were confirmed in sediment samples stored for 161 days at 4°C in New Mexico. In these samples, culturable heterotroph populations were log 3.23, 7.49, and 4.99 in the 54-, 57-, and 64-m sediments, respectively. The decrease in culturable heterotrophs at 161 days vs 83 days in the 54-m sediment may have been due to spatial variability or may represent a decline phase of the culturable microbial population. AO direct counts were approximately the same at sample acquisition and after 210 days storage (Table 2). AO-INT direct counts after 210 days storage had declined to below detection (log 4.6 CFU g-i dry sediment) in two of the three sediments. AO-INT direct counts at 210 days in the 54- and 57-m sediments were one to two orders of magnitude less than plate counts at 83 days. In addition, the numbers of AO cells and culturable heterotrophs were nearly equal in the 54- and 57-m sediments after storage as compared to a log 4.5 (54-m) and log 4.0 (57-m) greater number of AO cells vs culturable heterotrophs at sample acquisition. These data show that the physiological state of the community was very different after 83 days storage and after 210 days storage, and at sample acquisition and after storage.
After 83 days storage, richness remained higher in the 57-m sediment than

Microbiology of Vadose Zone Paleosols 293
in the 54- and 64-m sediments (Table 2). Each of the sediments had statistically smaller means after 83 days storage than at the time of sample acquisition (comparisons not shown). The pattern of statistical significance at sample ac- quisition and after 83 days storage remained the same. The same relationships were present for the Shannon-Weaver index. Equitability indices in each of the U-17 sediments were also statistically lower after 83 days storage than at the time of sample acquisition (comparisons not shown). The 64-m sediment con- tained only one microbial type, producing diversity indices of zero. The relative decrease in colonial morphology diversity with storage was smallest in the 57-m sediment. Decreases in colonial morphology diversity represent a rapid response to changed conditions by one or a few opportunistic species.
Storage of U-17 sediments for 97 days at 4°C resulted in statistically similar glucose mineralization rate constants and lag times in all three sediments (Table 2). The autoclaved sediment controls did not mineralize glucose. Comparisons of mineralization parameters for a particular U-17 sediment at sample acqui- sition and after 97 days storage were not possible due to different experimental conditions at the two times.
Enumerations of culturable heterotrophs from White Bluffs sediments after 15, 42, and 75 days storage at 4°C showed increasing numbers of culturable heterotrophs (data for 15 days and 42 days not shown). Populations in the AR and NM sediments were statistically different from each other after 75 days storage, whereas they were not statistically different at sample acquisition (Table 2). Populations in the AR sediment were statistically larger than at sample acquisition after 15, 42, and 75 days storage, whereas populations in the NM sediment became statistically larger than at sample acquisition only after 75 days storage (data not shown). AO and AO-INT direct counts were approxi- mately the same at 103 days storage as they were at the time of sample ac- quisition (Table 2). However, the numbers of AO cells and culturable hetero- trophs were nearly equal in the AR sediment after storage as compared to a log 4.5 greater number of AO cells vs culturable heterotrophs at sample ac- quisition, showing that the physiological state of the community changed dra- matically with storage.
Diversity in colony morphology was measured after sediments had been stored for 15 and 75 days. Richness in the NM sediment remained statistically greater than in the AR sediment following 75 days storage (Table 2). All three diversity indices for the AR sediment showed decreasing means with increased storage time, with statistically smaller means after 75 days storage than at sample acquisition (comparisons not shown). Equitability for the AR sediment was also statistically lower after 15 days storage than at sample acquisition (comparisons not shown). In contrast, diversity indices for stored NM sediment were not statistically different than at sample acquisition (comparisons not shown).
The glucose mineralization rate constant was statistically greater and the lag time was statistically less in the AR sediment than in the NM sediment after 75 days storage at 4°C (Table 2). Results after 75 days storage were the reverse of those at sample acquisition because the AR sediment had increased min- eralization rate constants and decreased lag times with increasing storage, whereas mineralization in the NM sediment remained relatively constant with storage

294 F.J. Brockman et al.
time. The AR sediment also had statistically different mineralization rate con- stants and lag times after 15 and 42 days storage than at sample acquisition (comparisons not shown). Mineralization parameters in the AR sediment after 42 days storage were similar to those obtained with the U-17 sediments after 97 days storage. The autoclaved sediment controls did not mineralize glucose.
Discussion
Two key findings have been made from this study regarding the microbiology of vadose zone sediments. The first is that a more extensive and responsive microbial community was present in the highly developed, TOC-rich paleosol (57-m) at the U-17 borehole, as indicated by the higher number of culturable heterotrophs, greater glucose mineralization potential, higher microbial diver- sity based on colony morphology, and higher ATP concentration, than in the two weakly developed paleosols. Second, storage at 4°C of deeply buried and near-surface sediments with high water potential resulted in large increases in culturable microbes, increased first-order rate constants and decreased lag times for glucose mineralization, and decreased microbial diversity based on colony morphology.
Effect of Moisture Recharge on the Microbiology o f Vadose Zone Paleosols
Moisture in the deep vadose zone at the U-17 borehole could result from artificial recharge from Hanford Site operations, residual moisture from drain- age of the sediment profile following the last glacial flood approximately 13,000 years ago (see below), and possibly small amounts of natural recharge [22, 46]. The amount of natural recharge varies with local topography, sediment texture, and vegetation. For example, recent unpublished data based on soil moisture chloride profiles suggest that in areas of active vegetation, natural recharge to below the root zone may be less than 0.02 mm year -1 (E. Murphy, personal communication). In contrast, between 1952 and 1960, a total of 3.8 × 108 liters of wastewater were disposed to the U-8 crib (a crib is an underground structure designed to allow wastewater to seep into the subsurface) 210 m away from the U-17 borehole [ 11 ]. A pipeline that has carried wastewater to the U-8 crib from 1952 to the present passes 60 m away from the borehole. This pipeline has been damaged near the borehole, but the history and amount of leakage from the pipeline is unknown (M. Brown, personal communication). The U-17 crib is located 150 m from the borehole and has received approxi- mately 1 × 106 liters of wastewater annually since 1989. In addition, three other cribs (U-l/2, U-12, U-16) 365-425 m from the borehole have received a total of 6.1 x 108 liters ofwastewater from 1951 to the present [11].
Water contents in the U-17 borehole samples (11-24%) were high relative to other deep vadose zone sediments at the Hanford Site. For example, sediment samples obtained from a second borehole, located approximately 15 km to the southwest, that was hydrologically upgradient from regions potentially affected

Microbiology of Vadose Zone Paleosols 295
by recharge from Hanford Site operations and at similar distances above the water table, had water contents ranging from 2 to 5% [37]. Therefore, it is likely that the U- 17 borehole sediments have been influenced by recharge from Han- ford Site operations. Although it is possible that moisture recharge may have delivered contaminants to the sediments, the lack of detection of radioactivity and volatile organic carbon during drilling operations and in sediment samples suggests that only water reached the material encountered in the borehole.
The gram reaction of the culturable microorganisms suggests that moisture recharge increases the number of gram-negative micoorganisms. The U-17 sediments were dominated by gram-negative isolates (71%), whereas sediments from depths between 30 and 100 m at the second, upgradient Hanford Site borehole contained only ~32% gram-negative isolates [36]. A similar pattern was observed in the near-surface White Bluffs sediments, which were studied to directly assess the effect of moisture recharge on the microbial community of stratigraphically identical paleosols. The wetter White Bluffs AR sediments ( -0 .01 MPa) contained five times more gram-negative isolates (71% vs 15%) than did the much drier White Bluffs NM sediments ( -5 .3 MPa). Because gram-positive microorganisms are more resistant to desiccation [27], drier sediments would select for their presence and/or greater ease of culturing. An increased proportion of culturable gram-negative isolates in the moist U-17 and White Bluffs AR sediments could have resulted from transport of surface microorganisms with recharge, or from moisture-induced stimulation of in- digenous dormant and/or previously nonculturable microorganisms that are more competitive than gram-positive bacteria at the higher water potential.
In addition to a higher proportion of gram-negative isolates, the White Bluffs AR sediment also had lower numbers ofculturable heterotrophs, a lower min- eralization rate constant, and lower richness based on colony morphology than the White Bluffs NM sediment. These effects suggest that environmental change (continuous high water availability in an initially moisture-limited environ- ment) was responsible for altering the composition of the original microbial community. Chemical and physical factors other than increased moisture may also contribute to the microbiological differences between the two sediments, but were not examined in detail in this study.
The survival and growth of microorganisms in deep vadose zones over geo- logic time periods may be assisted by episodic recharge events that introduce nutrients from surface sources, aid in bioavailability and transport of sediment- associated nutrients, redistribute nutrients between microsites of microbial activity, and promote weathering by increasing chemical gradients at surfaces. During the Pleistocene (~ 1 million to 13,000 years ago), glaciation of northern North America resulted in multiple cataclysmic floods in eastern Washington State [8, 53]. Episodes of flooding and subsequent drainage of the sediment profile could revive microbial activity and promote growth of entrained mi- croorganisms. Moisture recharge from Hanford Site operations could have a similar stimulating effect.
Microorganisms in deep vadose zone sediments may originate from entrain- ment of microorganisms during deposition, colonization during soil develop- ment processes, transport of microorganisms to the sediment during artificial recharge (e.g., Hanford Site operations) or geologic moisture recharge events,

296 F.J. Brockman et al.
or a combination of these processes. It is not known to what extent these processes contributed to the presence of microorganisms in the U-17 borehole samples. However, a variety of deep vadose zone nonpaleosol materials from the arid western United States that have not been influenced by artificial mois- ture recharge typically contain populations ofculturable heterotrophs that range from below detection to log 2.7 CFU g-1 dry material. These materials include non-welded and welded tuff in New Mexico [28]; sedimentary interbeds be- tween basalts, basalt fracture infills, and basalt fractures at a high desert in Idaho [15, 36]; and a variety of mixed clay, silt, sand, and gravel deposits sampled from another borehole at the Hanford Site in southeastern Washington State [36]. It is therefore likely that at least some of the microorganisms in the U-17 sediments were present prior to moisture recharge from Hanford Site operations.
Effect of Paleosol Organic Matter on the Microbiology of Vadose Zone Paleosols
Survival of microorganisms in deep vadose zones over geologic time periods could also be assisted by characteristics of the sediment organic matter. The TOC measured in paleosols is likely to be only a small fraction of the TOC originally present in the soil [45]. In fact, many paleosols that lack elevated TOC can be defined by other features of soil development. Utilization of the available carbon component of paleosol organic matter by microorganisms over geologic time periods is one explanation for the depletion of TOC in paleosols [51]. We suggest that the nature and concentration of TOC in vadose zone sediments is an important control on the survival of heterotrophic mi- croorganisms for geologic time periods in deep vadose zone sediments.
We studied three deeply buried paleosols to determine whether microbial survival or activity was enhanced by elevated concentrations of organic matter. The rank order of TOC and the rank orders of culturable heterotrophs, richness index, and the Shannon-Weaver index in the U-17 sediments were identical: 57-m > 54-m > 64-m. The 57-m sediment had statistically higher values for these parameters and for the glucose mineralization rate constant than did the 54- and 64-m sediments. In addition, ATP concentrations were, at minimum, 20 times as high in the 57-m sediment. Diversity indices have been used as indicators of environmental stress on microbial communities [1]. Microbial diversity tends to be low in ecosystems that are dominated by a strong, uni- directional physical stress because adaptations to the prevailing stress are highly selected. Therefore, the greater microbial diversity in the 57-m sediment could be consistent with greater nutrient availability at the present time. Because it is likely that the 57-m sediment had more TOC than the other two sediments prior to burial, initially higher microbial populations could also contribute to the greater activity and diversity observed in the 57-m sediment.
The survival of microorganisms over geologic time periods would be de- pendent on their ability to prevent cell death imposed by desiccation and concomitant nutrient limitation. Log-linear relationships between matric water potential and microbial respiration [42, 55], and between matric water potential

Microbiology of Vadose Zone Paleosols 297
and decomposition of organic matter [50], have been observed in soils. Lim- itations on both solute diffusion (i.e., nutrient availability) and bacterial move- ment are considered to be responsible for the decrease in microbial activity that occurs with decreasing matric water potential [27, 43, 55, 56]. It is possible that deep vadose zone microorganisms have evolved nutrient uptake systems that enable slow, steady uptake of organic carbon at matric potentials lower than uptake systems that have been studied to date. A second adaptive strategy for long-term survival is the ability to enter a resting state characterized by a lack of metabolism or very slow utilization of endogenous energy reserves at low matric water potentials. "Bursts" of nutrient uptake and metabolism during and following episodic moisture recharge events (e.g., precipitation, glacial floods, or anthropogenic sources) could be responsible for most utilization of organic matter in paleosols and other vadose zone sediments over geologic time periods. Such increased metabolism could cause spore germination, re- suscitation of inactive cells, an increase in cell size and/or cell numbers, and storage of nutrients as endogenous energy reserves.
The relatively impermeable, calcic 57-m sediment could have acted to filter out microorganisms that were transported with percolating water following glacial floods, resulting in a higher concentration of microorganisms in this sediment than in overlying or underlying sediments. A calcic paleosol (< 0.05% TOC) from a depth of 72 m at another borehole sampled for microbiological studies on the Hanford Site possessed the highest activity and had the highest population of culturable bacteria among 12 vadose zone sediment samples ranging in depth from 30-100 m [36, 37]. All sediments sampled from this borehole contained < 0.05% TOC and were not impacted by artificial recharge, suggesting that the calcic nature of the sediment may have been responsible for the higher concentration of microorganisms. Future studies will focus on other vadose zone calcic paleosols and on paleosols in the saturated zone to further investigate the effect of paleosol organic matter on characteristics of the microbial community.
Effect of Storage on Characteristics of the Microbial Community
Results from this study and others [26, 36] indicate that the microbiology of vadose zone sediments can change dramatically with sample storage. These changes were noted in incubations designed to investigate the ability to stim- ulate microbial activity/biomass with different nutrient treatments. Surpris- ingly, after a 42-day incubation at room temperature, large increases in ATP and culturable microorganisms were noted in all treatments of the 54-m sed- iment. These results suggest that the indigenous microorganisms were able to efficiently metabolize indigenous organic matter to support cell activity and culturability and/or to support cell growth. This metabolism occurred in the absence of exogenous organic or inorganic nutrients and in the presence of small increases in sediment moisture (-0.1 MPa less negative). As confirmatory evidence, examination of the U-17 sediments stored for 83 and 161 days at 4°C at PNL and at New Mexico showed similar increases in culturable hetero- trophs that were consistent with the increases in the 42-day incubation at 22°C.

298 F.J. Brockman et al.
The effects of storage on U-17 sediments and on near-surface White Bluffs sediments that were geologically similar to the U-17 upper Ringold sediment (64 m) revealed that moisture was an important controlling factor. In all U-17 sediments and in the White Bluffs AR sediment, which had water potentials of - 0 . 3 MPa or above, storage resulted in higher populations of culturable heterotrophs, increased glucose mineralization, and decreased microbial di- versity. In contrast, there was little or no change in these parameters with storage of the moisture-limited White Bluffs NM sediment ( -5 .0 MPa). The change from a predominantly nonculturable state (i.e., respiting cells >> plate counts) at sample acquisition to a subsequent predominantly culturable state (i.e., total AODC ~ plate counts) in three of the four moist sediments suggests that diffusion-limited availability of other nutrients is a likely controlling factor on microbial activity and culturability.
Although the processes responsible for the increases in microbial activity and culturability in vadose zone sediments with increasing storage time remain to be determined, physical disturbance of sediment during sampling and lab- oratory procedures probably contributes to increased nutrient availability. Physical disturbance of the sediment could stimulate biomass turnover by redistributing moisture and other nutrients, by increasing gaseous diffusion, and by relocating viable cells to substrate. Deep vadose zone sediments may contain high numbers of moribund cells, and intact or partially decomposed dead cells [33] whose biomass could be recycled by actively respiring cells. The changes in sediment microbiology with storage may be similar to the flush of microbial activity that is observed after soil fumigation or after physical mixing of soil by cultivation. The flush of activity is caused by the decomposition of organisms killed during fumigation [30] and by the disruption of soil aggregates that exposes hitherto inaccessible substrate to microbial attack [16, 31, 47]. Future research needs include identification of the relative roles of cell growth vs resuscitation of dormant cells, and of cell biomass recycling vs use of non- biomass sediment organic matter.
A basic understanding of the process(es) underlying the changes in sediment microbiology during the storage of sediment could be exploited for bioreme- diating contaminated deep vadose zones, by stimulating the process(es) in situ using appropriate engineering applications. With the application of methods for in situ stimulation of indigenous microorganisms, paleosols could provide inocula for the colonization of, and development of genetic diversity in, ad- jacent sediments containing smaller and less diverse microbial communities.
Conclusions
To the best of our knowledge, this research is the first examination of the microbiology of paleosols and represents a new contribution to the character- ization of the microbial ecology of deep subsurface environments. This study suggests that TOC concentrations are an important control on the culturability, activity, and diversity of deep vadose zone sediments, and that high water potentials are required for the storage-associated increases in microbial activity and culturability observed in the sediments. The synthesis of ATP and the

Microbiology of Vadose Zone Paleosols 299
utilization of sediment-associated nutrients in the presence of small increases in water potential demonstrate that large increases in microbial growth and activity may be achieved (in the laboratory) in some Hanford Site vadose zones without the addition of other nutrients. Stress-tolerant microorganisms indig- enous to the Hanford Site deep vadose zone may also possess other useful traits that could be exploited for the bioremediation of contaminants in these en- vironments.
Acknowledgments. This research was supported by the Subsurface Science Program, Office of Health and Environmental Research, U.S. Department of Energy (DOE). The continued support of Dr. F. J. Wobber is greatly appreciated. Pacific Northwest Laboratory is operated for the DOE by Battelle Memorial Institute under Contract DE-AC06-76RLO 1830. TLK is supported by DOE grants DE-FG04-88ER-60711 and DE-FG04-90ER-60992. We also acknowledge the technical assistance of Laurie Rosacker and Kady Christ.
References
1. Atlas RM, Bartha R (1981) Microbial ecology. Addison-Wesley, Menlo Park CA, pp 241-244 2. Balkwill DL (1989) Numbers, diversity, and morphological characteristics of aerobic, che-
moheterotrophic bacteria in deep subsurface sediments from a site in South Carolina. Geomi- crobiol J 7:33-52
3. Balkwill DL, Fredrickson JK, Thomas JM (1989) Vertical and horizontal variations in the physiological diversity of the aerobic chemoheterotrophic bacterial microflora in deep south- east coastal plain subsurface sediments. Appl Environ Microbiol 55:1058-1065
4. Balkwill DL, Ghiorse WC (1985) Characterization of subsurface bacteria associated with two shallow aquifers in Oklahoma. Appl Environ Microbiol 50:580-588
5. Bjornstad BN (1984) Suprabasalt stratigraphy within and adjacent to the reference repository location, SD-BWI-DP-039. Rockwell Hanford Operations, Richland, WA
6. Bjornstad BN (1985) Late Cenozoic stratigraphy and tectonic evolution within a subsiding basin, south-central Washington. Geol Soc Am Abstr 17:524
7. Black CA (1965) Methods of soil analysis. American Society of Agronomy Publication 90-3, Madison, WI
8. Bretz JH (1959) Washington's channeled scabland, bulletin 45. Washington Division of Mines and Geology, Olympia, WA
9. Brown DJ (1959) Subsurface geology of the Hanford separations areas, HW-61780. General Electric Company, Richland, WA
10. Brown DJ (1960) An eolian deposit beneath the 200-West Area, HW-67549. General Electric Company, Richland, WA
11. Brown M J, P'Pool PK, Thomas SP (1990) Westinghouse Hanford Company effluent discharges and solid waste management report for calendar year 1989:200/600 areas, WHC-ER-0141- 2. Westinghouse Hanford Company, Richland, WA
12. Brown RE (1960) An introduction to the surface ofthe Ringold formation beneath the Hanford works area, HW-66289. General Electric Company, Richland, WA
13. Chappelle FH, Lovely DR (1989) Rates ofmicrobial metabolism in deep coastal plain aquifers. Appl Environ Microbiol 56:1865-1874
14. Chappelle FH, Morris JT, McMahon PB, Zelibor JR (1988) Bacterial metabolism and the delta-13C composition of groundwater, Floridan aquifer system, South Carolina. Geology 16: 117-121
15. Colwell FS (1989) Microbiological comparison of surface soil and unsaturated subsurface soil from a semiarid high desert. Appl Environ Microbiol 55:2420-2423
16. Craswell ET, Waring SA (1972) Effect of grinding on the decomposition of soil organic matter: Oxygen uptake and nitrogen mineralization in virgin and cultivated cracking clay soils. Soil Biol Biochem 4:435--442

300 F.J. Brockman et al.
17. Department of Energy (1988) Consultation draft, site characterization plan, reference repository location, Hanford Site, Washington, DOE/RW-0164 Vol 1. U.S. Department of Energy, Wash- ington, DC
18. Environmental Protection Agency (1983) Methods for chemical analysis of waters and wastes, EPA-600/4-79-020. U.S. Environmental Protection Agency, Washington, DC
19. Francis A J, Slater J M, Dodge CJ (1989) Denitrification in deep subsurface sediments. Geomi- crobiol J 7:103-116
20. Fredrickson JK, Balkwill DL, Zachara JM, Li SW, Brockman FJ, Simmons MA (1991) Phys- iological diversity and distributions of heterotrophic bacteria in deep Cretaceous sediments of the Atlantic coastal plain. Appl Environ Microbiol 57:402-411
21. Fredrickson JK, Garland TR, Hicks RJ, Thomas JM, Li SW, McFadden KM (1989) Litho- trophic and heterotrophic bacteria in deep subsurface sediments and their relation to sediment properties. Geomicrobiol J 7:53-66
22. Gee GW (1987) Recharge at the Hanford Site: Status report, PNL-6403. Pacific Northwest Laboratory, Richland, WA
23. Gee GW, Bauder JW (1986) Particle size analysis. In: Klute A (ed) Methods of soil analysis, part 1. American Society of Agronomy, Madison, WI, pp 383-409
24. Ghiorse WC, Balkwill DL (1983) Enumeration and morphological characterization of bacteria indigenous to subsurface environments. Dev Ind Microbiol 24:213-224
25. Gile LH, Peterson FF, Grossman RB (1966) Morphological and genetic sequences of carbonate accumulation in desert soils. Soil Sci 101:347-360
26. Haldeman DL, Amy PS (1991) Storage and growth conditions for optimal recovery of deep subsurface microbes at the Nevada Test Site. Abstr Ann Meet Am Soc Microbiol, American Society for Microbiology, Washington, DC, pp 287
27. Harris RF (1981) Effect of water potential on microbial growth and activity. In: Parr JF, Gardner WR, Elliott LF (eds) Water potential relations in soil microbiology. Soil Science Society of America, Madison, WI, pp 23-96
28. Hersman L, Purtymun W, Sinclair J (1988) Preliminary microbial analysis of the vadose zone, Pajarito Plateau, New Mexico. Abstr Ann Meet Am Soc Microbiol, American Society for Microbiology, Washington, DC, pp 252
29. Hicks RJ, Fredrickson JK (1989) Aerobic metabolic potential of microbial populations in- digenous to deep subsurface environments. Geomicrobiol J 7:67-78
30. Jenkinson DS (1966) Studies on the decomposition ofplant material in soil: Partial sterilization of soil and the soil biomass. J Soil Sci 17:280-302
31. Jenkinson DS, Powlson DS (1976) The effects of biocidal treatments on metabolism in soil: A method for measuring soil biomass. Soil Biol Biochem 8:209-213
32. Jones RE, Beeman RE, Suflita JM (1989) Anaerobic metabolic processes in the deep terrestrial subsurface. Geornicrobiol J 7:117-130
33. Kieft TL, Rosacker LL (1991) Application of respiration- and adenylate-based soil microbi- ological assays to deep subsurface terrestrial sediments. Soil Biol Biochem 23:563-568
34. Larson RJ (1984) Kinetic and ecological approaches for predicting biodegradation rates of xenobiotic organic chemicals in natural ecosystems. In: Klug MJ, Reddy CA (eds) Current perspectives in microbial ecology. American Society for Microbiology, Washington, DC, pp 677-686
35. Last GV, Bjornstad BN, Bergeron MP, Wallace RW, Newcomer DR, Schramke JA, Chamness MA, Cline CS, Airhart SP, Wilbur JS (1989) Hydrogeology of the 200 areas low-level burial grounds--An interim report, PNL-6820. Pacific Northwest Laboratory, Richland, WA
36. Li SW, Fredrickson JK, Spangenburg W, Workman DW, Brockman FJ, Stevens TO, Long PE (1991) Microbial activities in two dedicated deep subsurface boreholes. Abstr Ann Meet Amer Soc Microbiol, American Society for Microbiology, Washington, DC, pp 246
37. Long PE, Fredrickson JK, Murphy EM, Rawson SA, Brockman FJ, McKinley JP, Bjornstad BN (1991) Geohydrologic and geochemical controls on subsurface microorganisms in late Cenozoic sediments, south-central Washington. Geol Soc Am Abstr 23:A377
38. Lovely DR, Chappelle FH, Phillips EJP (1990) Fe(III)-reducing bacteria in deeply buried sediments of the Atlantic coastal plain. Geology 55:954-597

Microbiology of Vadose Zone Paleosols 301
39. Machette MN (1985) Calcic soils of the southwestern United States. In: Weide D (ed) Soils and Quaternary geology of the southwestern United States. Geological Society of America, Boulder, CO, pp 1-21
40. Madsen EL, Bollag JM (1989) Aerobic and anaerobic microbial activity in deep subsurface sediments from the Savannah River Plant. Geomicrobiol J 7:93-102
41. Marratt WJ (1988) Study of landslides along the Columbia River in the block 15 area of Franklin County, Washington. Franklin Conservation District, Pasco, WA
42. Orchard VA, Cook FJ (1983) Relationship between soil respiration and soil moisture. Soil Biol Biochem 15:447-453
43. Papendick RI, Campbell GS (1981) Theory and measurement of water potential. In: Parr JF, Gardner WR, Elliott LF (eds) Water potential relations in soil microbiology. Soil Science Society of America, Madison, WI, pp 1-22
44. Phelps TJ, Raione EG, White DC, Fliermans CB (1989) Microbial activities in deep subsurface environments. Geomicrobiol J 7:79-91
45. Rettalack GJ (1990) Soils of the past. Unwin Hyman, Winchester, MA, pp 520 46. Rockhold ML, Fayer MJ, Gee GW, Kanyid MJ (1990) Natural groundwater recharge and
water balance at the Hanford Site, PNL-7215. Pacific Northwest Laboratory, Richland, WA 47. Rovira AD, Greacen EL (1957) The effect of aggregate disruption on the activity of micro-
organisms in the soil. Aust J Agric Res 8:659-673 48. Shelton DR, Tiedje JM (1984) General method for determining anaerobic biodegradation
potential. Appl Environ Microbiol 47:850-857 49. Sinclair JL, Ghiorse WC (1989) Distribution of aerobic and anaerobic bacteria, protozoa,
algae, and fungi in deep subsurface sediments. Geomicrobiol J 7:15-31 50. Sommers LE, Gilmour CM, Wildung RE, Beck SM (1981) The effect of water potential on
decomposition processes in soils. In: Parr JF, Gardner WR, Elliott LF (eds) Water potential relations in soil microbiology. Soil Science Society of America, Madison, WI, pp 97-118
51. Stevenson FJ (1986) Cycles of soil. Wiley and Sons, New York 52. Tallman AM, Fecht KR, Marratt MC, Last GV (1979) Geology of the separations areas,
Hanford Site, south-central Washington, RHO-ST-23. Rockwell Hanford Operations, Rich- land, WA
53. Waitt RB Jr (1985) Case for periodic, colossal jokulhlaups from Pleistocene glacial Lake Missoula. Geol Soc Am Bull 96:1271-1286
54. Webster J J, Hampton GJ, Wilson JT, Ghiorse WC, Leach FR (1985) Determination of mi- crobial cell numbers in subsurface samples. Ground Water 23:17-25
55. Wilson JM, Griffin DM (1975) Water potential and the respiration of microorganisms in soil. Soil Biol Biochem 7:199-204
56. Wong PTW, Griffin DM (1976) Bacterial movement at high matric potentials: In artificial and natural soils. Soil Biol Biochem 8:215-218
57. Zimmerman RC, Iturriaga R, Becker-Birck J (1978) Simultaneous determination of the total number of aquatic bacteria and the number thereof involved in respiration. Appl Environ Microbiol 36:926-935





























