Embed Size (px)
Citation preview

Microbial dolomite crusts from the carbonate platformoff western India
V. PURNACHANDRA RAO*, PRATIMA M. KESSARKAR*, W. E. KRUMBEIN� ,K. P. KRAJEWSKI� and ROBERT J. SCHNEIDER§*National Institute of Oceanography, Dona Paula-403 004, Goa, India (E-mail: [email protected])�Geomicrobiology, ICBM, Carl von Ossietzky-University of Oldenburg, Oldenburg, Germany�Institute of Geological Sciences, Polish Academy of Sciences, Twarda 51/55, 00-818 Warsaw, Poland§Woods Hole Oceanographic Institution, Woods Hole, MA 02543-1539, USA
ABSTRACT
The occurrence of Late Pleistocene dolomite crusts that occur at 64 m depth on
the carbonate platform off western India is documented. Dolomite is the most
predominant mineral in the crusts. In thin section, the crust consists of
dolomitized microlaminae interspersed with detrital particles. Under scanning
electron microscopy, these laminae are made up of tubular filaments or cellular
structures of probable cyanobacterial origin. Dolomite crystals encrust or
overgrow the surfaces of the microbial filaments and/or cells; progressive
mineralization obliterates their morphology. Well-preserved microbial mats,
sulphide minerals (pyrrhotite and marcasite) and the stable isotope
composition of dolomite in the crusts indicate hypersaline and anoxic
conditions during dolomite formation. The crusts are similar to dolomite
stromatolites, and biogeochemical processes related to decaying microbial mats
under anoxic conditions probably played an important role in dolomite
precipitation. The dolomite is therefore primary and/or very early diagenetic in
origin. The dolomite crusts are interpreted to be a composite of microbial
dolomite overprinted by early burial organic dolomite. The results of this study
suggest that a microbial model for dolomite formation may be relevant for the
origin of ancient massive dolomites in marine successions characterized by
cryptalgal laminites. The age of the crusts further suggests that the platform was
situated at shallow subtidal depths during the Last Glacial Maximum.
Keywords Carbonate platform, dolomite stromatolites, late Pleistocene,microbial processes, western India.
INTRODUCTION
Dolomite is found in far greater abundance inEarth’s early history than during Quaternary/modern times, in which its occurrence is smallscale and disparate. Although numerous modelshave been proposed for dolomite formation(Machel & Mountjoy, 1986; Hardie, 1987), theexplanation for its unequal distribution throughtime still remains an enigma – known as the‘dolomite problem’. Modern dolomite occurs ina wide variety of settings from lacustrine tosubtidal to deep-sea environments. Most of thehydrological models proposed for the formationof shallow-water dolomites involve pumping of
large volumes of normal sea water or brinesthrough the sediments (Adams & Rhodes, 1960;Carballo et al., 1987; Vahrenkamp et al., 1991;Swart & Melim, 2000; Melim & Scholle, 2002) orits production in a mixing zone of marine andmeteoric waters (Badiozamani, 1973; Humphery& Quinn, 1990; Humphery & Radjef, 1991;Humphrey, 2000). These models, however, donot convincingly explain how dolomite precipi-tation has overcome the kinetic inhibiting fac-tors, such as the high hydration energy of Mg2+
(Lippmann, 1973), the extremely low activity ofCO3
2– anions (Garrels & Thompson, 1962) andthe presence of even very low concentrations ofsulphate (Baker & Kastner, 1981; Slaughter &
Sedimentology (2003) 50, 819–830 doi: 10.1046/j.1365-3091.2003.00574.x
� 2003 International Association of Sedimentologists 819

Hill, 1991). The organogenic model for deep-seadolomites in organic-rich sediments proposesdolomite precipitation by simultaneous increasein carbonate alkalinity and decrease in sulphateconcentrations to near zero by sulphate-reducingbacterial degradation of organic matter (Irwin,1980; Kelts & McKenzie, 1982; Compton, 1988).This model, however, has not been invoked forthe origin of massive dolomites in ancientPrecambrian and Phanerozoic carbonate plat-forms.
Modern primary or penecontemporaneousdolomites occur in several lacustrine and shal-low-marine regions, namely South Bonaire, theBahamas, the Persian Gulf, the Florida keys andthe ephemeral lakes of the Coorong region ofAustralia (Deffeyes et al., 1965; Illings et al.,1965; Shinn et al., 1965; Shinn, 1968; Von derBorch & Jones, 1976; Von der Borch & Lock, 1979;Patterson & Kinsman, 1982; Carballo et al., 1987;Rosen et al., 1988; Rosen & Coshell, 1992; Wright,2000). In these regions, dolomites are associatedwith organic-rich sediments or with buried algalmats that occur at the same level with, or inproximity to, dolomite formation. A review ofsome of these modern dolomite occurrencessuggests a plausible involvement of benthicmicrobial communities in the precipitation ofdolomite (see Wright, 2000). Microbial mediationof dolomite was suggested for the ephemeral lakesof the Coorong region (Wright, 2000). AlthoughNadson (1928) and Neher (1959) produced dolo-mite under laboratory conditions using sulphate-reducing bacteria, the microbial link in dolomiteformation was first documented by Vasconceloset al. (1995) and Vasconcelos & McKenzie (1997).Modern primary dolomites occur in the tropicalhypersaline lagoon Lagoa, Vermelha, Brazil.Vasconcelos et al. (1995) and Warthmann et al.(2000) used sulphate-reducing bacteria from theselagoonal sediments and successfully achieveddolomite precipitation under laboratory condi-tions by simulating the anoxic depositional envi-ronment of the lagoon. Many ancient dolomitedeposits are associated with stromatolitic lime-stones or organic-rich sediments (Valdiya, 1972;Gebelian & Hoffman, 1973; Montanez & Read,1992; Montanez & Osleger, 1993; Wright, 1997),suggesting a possible mechanistic link to micro-bial mediation of dolomite. In this paper, weadvance the microbial dolomite model by report-ing on late Pleistocene dolomite crusts from thecarbonate platform off western India and suggestthat dolomitization by benthic microbial commu-nities occurred under anoxic conditions.
GEOLOGICAL SETTING
The studied carbonate platform lies on the outercontinental shelf off the north-western margin ofIndia largely at depths between 60 and 90 m(Fig. 1). However, the depressions within theplatform reach up to a water depth of 110 m.The inner shelf is a clastic depocentre – theDahanu depression – in which Narmada and TaptiRiver-derived sediments have been depositedsince the Eocene. The sediments on the platformare largely faecal pellet-dominated aragonitesands, subordinate oolites and a few skeletalfragments (Rao et al., 1994). Twenty-eight radio-carbon ages on surficial carbonate sediments andlimestones (including nine AMS ages on oolites)from the platform range between 21 000 and6730 years BP (Rao et al., 2003). These indicatecarbonate deposition on the platform throughoutthe glacio-eustatic sea-level low of 18 ka and thesubsequent sea-level rise of ‡ 100 m during theHolocene. Relict fringing reefs occur on the north-eastern edge of the platform (see Fig. 1). Aragonitefaecal pellets, fragments of corals, bivalves andbryozoans compose the coarse fraction of thesediments (Rao et al., 1994). Dark-grey clayeysands were recovered at 64 m water depth in aseaward depression of the fringing reef. Dark-grey
Fig. 1. Location of dolomite crusts (sample no. 1466)on the carbonate platform.
820 V. P. Rao et al.
� 2003 International Association of Sedimentologists, Sedimentology, 50, 819–830

dolomite crusts (2 · 2Æ5 cm to 3 · 0Æ5 cm) andpebbles occur within these sediments.
METHODS
Thin sections of the crusts were studied petro-graphically. Several freshly broken surfaces of thecrust were examined by scanning electron micro-scope (SEM). Powders of the crusts were analysedon a Philips X-ray diffractometer using nickel-filtered Cu Ka radiation and scanned from 8� to60� 2h at 1� 2h min)1. The stable isotope compo-sition of the crust was determined in duplicate atthe laboratory of the Polish Academy of Sciences,Warsaw, Poland. A modified procedure of CO2
extraction from co-existing calcite and dolomitemixtures was used (Ray & Ramesh, 1998). Thesamples were treated with anhydrous phosphoricacid at 25 �C in a vacuum using a ‘progressiveleach’ procedure. Spectrometric gas was collectedafter 15 min, with an in-between, 4 and 20 h, gas-rejection step, and then 72 h after reaction. Gassamples collected after 15 min and 72 h representcalcite and dolomite phases respectively. Gassamples were purified by cryogenic distillationand analysed using a Finnigan Mat Delta Plusmass spectrometer working in dual inlet modewith universal triple collector. 18O/16O and13C/12C ratios were measured with respect to aprecalibrated internal laboratory standard (Carr-ara marble). The results are expressed as d13C andd18O notations relative to VPDB. Fractionationfactors (a) used for calcite and dolomite were1Æ01025 and 1Æ01178 respectively (Rosenbaum &Sheppard, 1986). The 13C and 18O were correctedfor 17O using the procedure of Craig (1957).Analytical reproducibility for the in-house stan-dard (pure calcite) was better than ± 0Æ05& and± 0Æ1& for d13C and d18O respectively. Crust ageswere determined by accelerator mass spectro-meter (AMS) analysis at Woods Hole Oceanogra-phic Institution (WHOI), USA (reference receiptno. at WHOI ¼ 28795).
RESULTS
Petrology
The dolomite crusts are hard and irregular withsurface borings and bryozoan encrustations. Thebroken surface of the crusts exhibits dark-greycompact peripheral portions and light-greyloosely cemented central portions (Fig. 2A). Thin
sections show 50 to 70 lm thick dolomitizedlaminae (Fig. 2B) or 15 lm thin irregular laminaeinterspersed with bioclastic detritus (Fig. 2C).Thin filamentous structures < 15 lm in diameter
Fig. 2. (A) Broken surface of part of the dolomite crustshowing dark-grey rim at the peripheral portions.(B and C) Thin-section photomicrographs of the crust.(B) Thick dolomite microlaminae interspersed withdetrital particles. (C) Thin strands of microfilaments inthe matrix.
Microbial dolomite crusts off western India 821
� 2003 International Association of Sedimentologists, Sedimentology, 50, 819–830
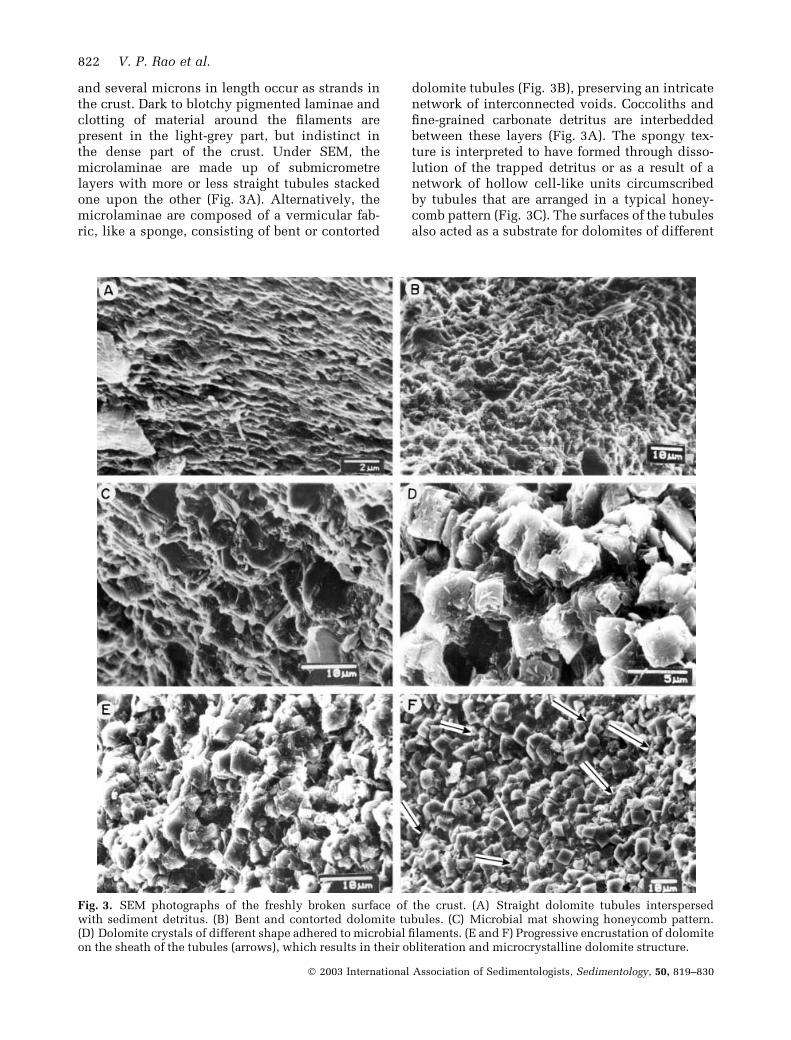
and several microns in length occur as strands inthe crust. Dark to blotchy pigmented laminae andclotting of material around the filaments arepresent in the light-grey part, but indistinct inthe dense part of the crust. Under SEM, themicrolaminae are made up of submicrometrelayers with more or less straight tubules stackedone upon the other (Fig. 3A). Alternatively, themicrolaminae are composed of a vermicular fab-ric, like a sponge, consisting of bent or contorted
dolomite tubules (Fig. 3B), preserving an intricatenetwork of interconnected voids. Coccoliths andfine-grained carbonate detritus are interbeddedbetween these layers (Fig. 3A). The spongy tex-ture is interpreted to have formed through disso-lution of the trapped detritus or as a result of anetwork of hollow cell-like units circumscribedby tubules that are arranged in a typical honey-comb pattern (Fig. 3C). The surfaces of the tubulesalso acted as a substrate for dolomites of different
Fig. 3. SEM photographs of the freshly broken surface of the crust. (A) Straight dolomite tubules interspersedwith sediment detritus. (B) Bent and contorted dolomite tubules. (C) Microbial mat showing honeycomb pattern.(D) Dolomite crystals of different shape adhered to microbial filaments. (E and F) Progressive encrustation of dolomiteon the sheath of the tubules (arrows), which results in their obliteration and microcrystalline dolomite structure.
822 V. P. Rao et al.
� 2003 International Association of Sedimentologists, Sedimentology, 50, 819–830
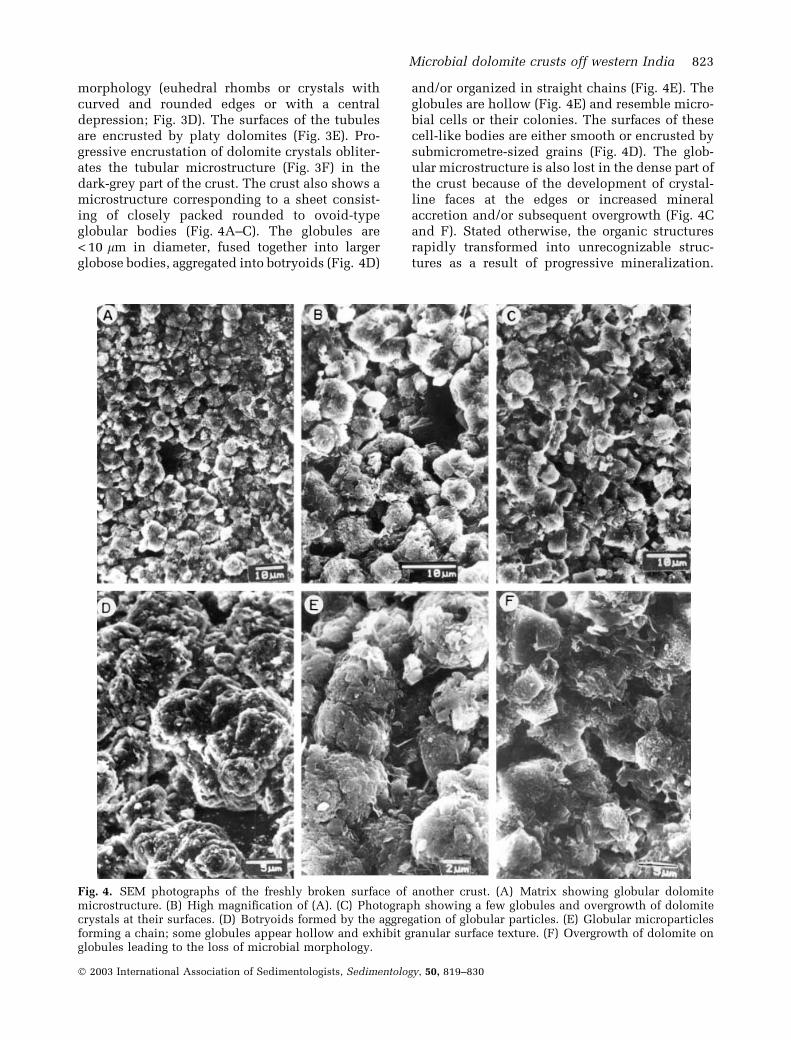
morphology (euhedral rhombs or crystals withcurved and rounded edges or with a centraldepression; Fig. 3D). The surfaces of the tubulesare encrusted by platy dolomites (Fig. 3E). Pro-gressive encrustation of dolomite crystals obliter-ates the tubular microstructure (Fig. 3F) in thedark-grey part of the crust. The crust also shows amicrostructure corresponding to a sheet consist-ing of closely packed rounded to ovoid-typeglobular bodies (Fig. 4A–C). The globules are< 10 lm in diameter, fused together into largerglobose bodies, aggregated into botryoids (Fig. 4D)
and/or organized in straight chains (Fig. 4E). Theglobules are hollow (Fig. 4E) and resemble micro-bial cells or their colonies. The surfaces of thesecell-like bodies are either smooth or encrusted bysubmicrometre-sized grains (Fig. 4D). The glob-ular microstructure is also lost in the dense part ofthe crust because of the development of crystal-line faces at the edges or increased mineralaccretion and/or subsequent overgrowth (Fig. 4Cand F). Stated otherwise, the organic structuresrapidly transformed into unrecognizable struc-tures as a result of progressive mineralization.
Fig. 4. SEM photographs of the freshly broken surface of another crust. (A) Matrix showing globular dolomitemicrostructure. (B) High magnification of (A). (C) Photograph showing a few globules and overgrowth of dolomitecrystals at their surfaces. (D) Botryoids formed by the aggregation of globular particles. (E) Globular microparticlesforming a chain; some globules appear hollow and exhibit granular surface texture. (F) Overgrowth of dolomite onglobules leading to the loss of microbial morphology.
Microbial dolomite crusts off western India 823
� 2003 International Association of Sedimentologists, Sedimentology, 50, 819–830
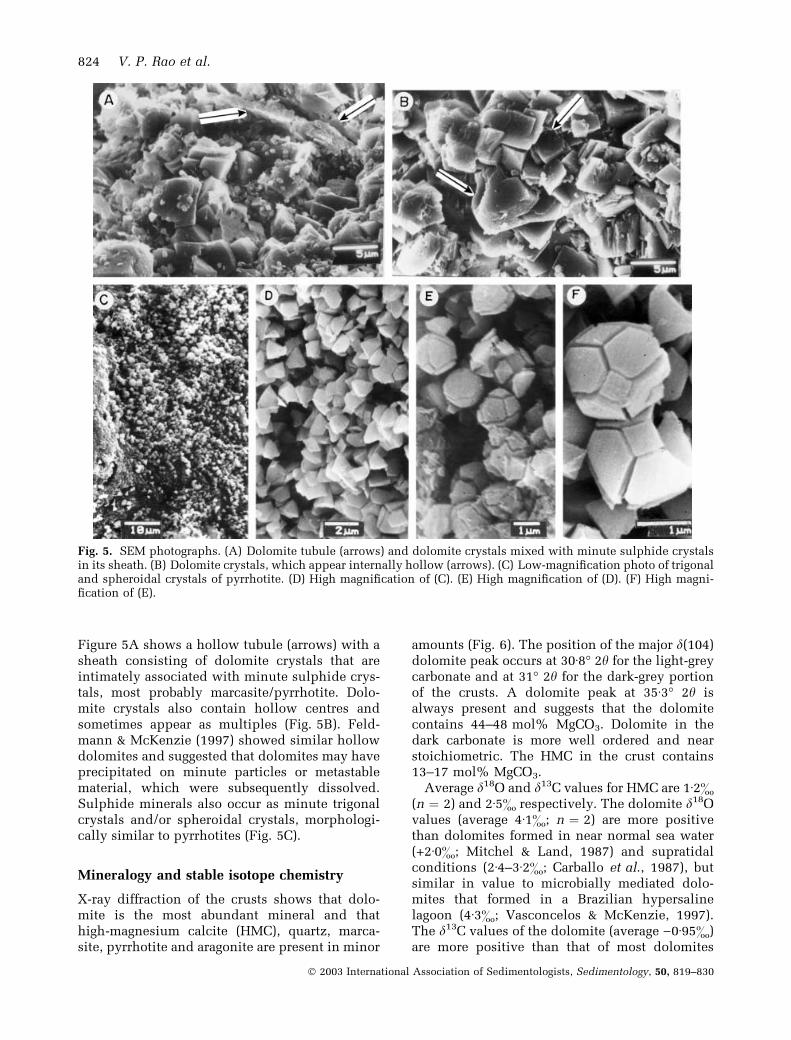
Figure 5A shows a hollow tubule (arrows) with asheath consisting of dolomite crystals that areintimately associated with minute sulphide crys-tals, most probably marcasite/pyrrhotite. Dolo-mite crystals also contain hollow centres andsometimes appear as multiples (Fig. 5B). Feld-mann & McKenzie (1997) showed similar hollowdolomites and suggested that dolomites may haveprecipitated on minute particles or metastablematerial, which were subsequently dissolved.Sulphide minerals also occur as minute trigonalcrystals and/or spheroidal crystals, morphologi-cally similar to pyrrhotites (Fig. 5C).
Mineralogy and stable isotope chemistry
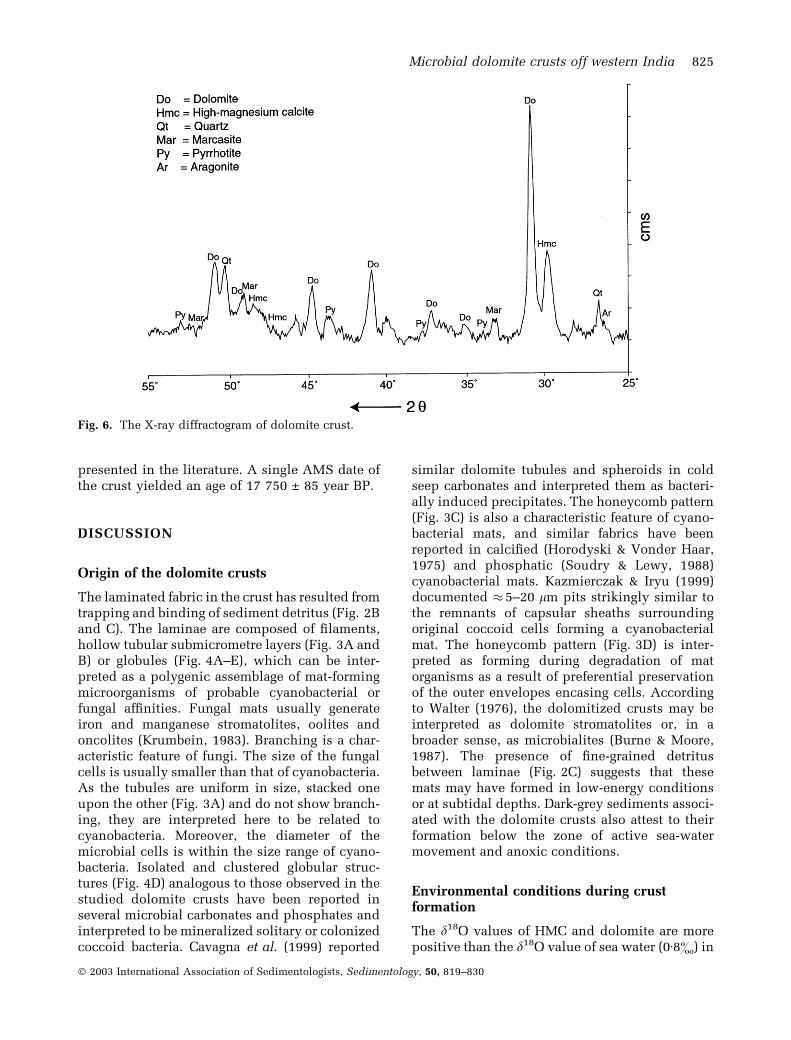
X-ray diffraction of the crusts shows that dolo-mite is the most abundant mineral and thathigh-magnesium calcite (HMC), quartz, marca-site, pyrrhotite and aragonite are present in minor
amounts (Fig. 6). The position of the major d(104)dolomite peak occurs at 30Æ8� 2h for the light-greycarbonate and at 31� 2h for the dark-grey portionof the crusts. A dolomite peak at 35Æ3� 2h isalways present and suggests that the dolomitecontains 44–48 mol% MgCO3. Dolomite in thedark carbonate is more well ordered and nearstoichiometric. The HMC in the crust contains13–17 mol% MgCO3.
Average d18O and d13C values for HMC are 1Æ2&(n ¼ 2) and 2Æ5& respectively. The dolomite d18Ovalues (average 4Æ1&; n ¼ 2) are more positivethan dolomites formed in near normal sea water(+2Æ0&; Mitchel & Land, 1987) and supratidalconditions (2Æ4–3Æ2&; Carballo et al., 1987), butsimilar in value to microbially mediated dolo-mites that formed in a Brazilian hypersalinelagoon (4Æ3&; Vasconcelos & McKenzie, 1997).The d13C values of the dolomite (average )0Æ95&)are more positive than that of most dolomites
Fig. 5. SEM photographs. (A) Dolomite tubule (arrows) and dolomite crystals mixed with minute sulphide crystalsin its sheath. (B) Dolomite crystals, which appear internally hollow (arrows). (C) Low-magnification photo of trigonaland spheroidal crystals of pyrrhotite. (D) High magnification of (C). (E) High magnification of (D). (F) High magni-fication of (E).
824 V. P. Rao et al.
� 2003 International Association of Sedimentologists, Sedimentology, 50, 819–830
presented in the literature. A single AMS date ofthe crust yielded an age of 17 750 ± 85 year BP.
DISCUSSION
Origin of the dolomite crusts
The laminated fabric in the crust has resulted fromtrapping and binding of sediment detritus (Fig. 2Band C). The laminae are composed of filaments,hollow tubular submicrometre layers (Fig. 3A andB) or globules (Fig. 4A–E), which can be inter-preted as a polygenic assemblage of mat-formingmicroorganisms of probable cyanobacterial orfungal affinities. Fungal mats usually generateiron and manganese stromatolites, oolites andoncolites (Krumbein, 1983). Branching is a char-acteristic feature of fungi. The size of the fungalcells is usually smaller than that of cyanobacteria.As the tubules are uniform in size, stacked oneupon the other (Fig. 3A) and do not show branch-ing, they are interpreted here to be related tocyanobacteria. Moreover, the diameter of themicrobial cells is within the size range of cyano-bacteria. Isolated and clustered globular struc-tures (Fig. 4D) analogous to those observed in thestudied dolomite crusts have been reported inseveral microbial carbonates and phosphates andinterpreted to be mineralized solitary or colonizedcoccoid bacteria. Cavagna et al. (1999) reported
similar dolomite tubules and spheroids in coldseep carbonates and interpreted them as bacteri-ally induced precipitates. The honeycomb pattern(Fig. 3C) is also a characteristic feature of cyano-bacterial mats, and similar fabrics have beenreported in calcified (Horodyski & Vonder Haar,1975) and phosphatic (Soudry & Lewy, 1988)cyanobacterial mats. Kazmierczak & Iryu (1999)documented � 5–20 lm pits strikingly similar tothe remnants of capsular sheaths surroundingoriginal coccoid cells forming a cyanobacterialmat. The honeycomb pattern (Fig. 3D) is inter-preted as forming during degradation of matorganisms as a result of preferential preservationof the outer envelopes encasing cells. Accordingto Walter (1976), the dolomitized crusts may beinterpreted as dolomite stromatolites or, in abroader sense, as microbialites (Burne & Moore,1987). The presence of fine-grained detritusbetween laminae (Fig. 2C) suggests that thesemats may have formed in low-energy conditionsor at subtidal depths. Dark-grey sediments associ-ated with the dolomite crusts also attest to theirformation below the zone of active sea-watermovement and anoxic conditions.
Environmental conditions during crustformation
The d18O values of HMC and dolomite are morepositive than the d18O value of sea water (0Æ8&) in
Fig. 6. The X-ray diffractogram of dolomite crust.
Microbial dolomite crusts off western India 825
� 2003 International Association of Sedimentologists, Sedimentology, 50, 819–830

the offshore regions of the study area (Sarkaret al., 2000). This indicates that the HMC in thecrust and the dolomite probably precipitated fromhypersaline waters. The average d18O value(4Æ3&) of modern dolomites that precipitated inthe tropical hypersaline lagoon of Lagoa, Brazil(Vasconcelos & McKenzie, 1997), is similar tothose of the studied dolomites (average 4Æ1&).Sulphide minerals such as marcasite and pyrrho-tite (Figs 5A–C and 6) indicate sulphate-reducingconditions. As these minerals are intimatelyassociated with dolomite crystals (Fig. 5A andB), they most probably developed in the crustcontemporary with dolomite formation. Well-preserved dolomite microbial mats and cellularstructures (Figs 3–5) imply that these cyanobac-terial remains may have passed quickly into theanaerobic zone as the mat community builtupwards, or were preserved by the coincidenceof hypersaline and anaerobic conditions duringmineralization (see Krumbein & Cohen, 1977).Dark-grey organic-rich sediments associated withthe crust support the assertion of oxygen-deficientconditions. HMC d13C values (average 2Æ5&) aresimilar to those of shallow-marine limestones(average 2Æ5&; Milliman, 1974). However, dolo-mite d13C values (average )1Æ0&) are relativelydepleted. If organic carbon diagenesis was pre-dominant, one would expect much lower d13Cvalues. For example, the average d13C value ofmicrobial dolomites of the Vermelha Lagoon is)9Æ0&. More positive values suggest that thecarbonate pool during dolomitization for thestudied dolomites may have been derived fromtwo different sources and admixed: (1) chemo-trophic oxidation of organic matter and/or per-haps from atmospheric CO2; and (2) highproportions of bicarbonate-containing ‘heavy’carbon derived from the dissolution of carbonatedetritus during sulphate-reducing conditionsand/or dissolved inorganic carbon diffusedinto the pores from the overlying waters. Thelatter would have diluted the ‘light’, organicallyderived carbon in the porewaters resulting inmore positive porewater d13C values.
Modern microbial dolomites occur in the eph-emeral lakes of the Coorong region, Australia.Benthic microbial communities associated withbacterial sulphate reduction are prevalent in theupper layers of the lake sediments (Wright, 2000).Although one would expect much more negatived13C values for these dolomites because of organicdiagenesis, Borg & Von der Borch (1984) reporteda range of d13C values from )2& to +4&. Theseimply that the d13C values of dolomite depend on
the amount of dilution of organic-derived carbonby carbon released by dissolution of precursorcarbonates. Sulphate-reducing conditions in thesediments and hypersalinity of the overlyingwaters at the time of dolomite formation aretherefore interpreted.
Mode of dolomite formation
The geochemical signatures and petrolographicrelationships of the dolomite crusts suggest twosimultaneous processes involved in dolomiteformation: (1) primary dolomite formation; and(2) dolomitization of the mat during early organicmatter diagenesis. The d18O value of dolomite(average 4Æ1&) is more positive than that of HMC(average 1Æ2&) in the crust, and closer to that ofthe dolomite–calcite fractionation value (3Æ8&;Land, 1980). This indicates that the dolomite isprimary and co-precipitated with the calcite.Montanez & Read (1992) suggested that initialdolomite d18O values are commonly not retainedeven if the dolomites are least altered diagenet-ically. Dolomite stromatolites in this study aresimilar to those of stromatolites in which themineral precipitation is a metabolic product ofthe mat-forming microorganisms, mostly bycyanobacteria. The well-preserved dolomite tubu-lar laminae (Fig. 3A–C) are microbially precipita-ted. Cyanobacteria generally have a gelatinoussheath 2–10 lm thick (Pentecost & Riding, 1986;Burne & Moore, 1987). Several workers havesuggested that cyanobacterial calcification is onlypartly within the influence of the microorgan-isms. It requires the presence of a sheath toprovide a site for nucleation and suitable envir-onmental conditions favouring precipitation ofcalcium carbonate (Krumbein & Cohen, 1977;Pentecost & Riding, 1986; Riding, 2000). Dolomiteencrustations on tubules (Fig. 3D–F) and over-growth of dolomite on cell-like particles (Fig. 4Cand F) suggest that dolomite formation continuedon the sheath during early diagenesis. As dis-cussed previously, the dolomite d13C valuessupport an early diagenetic origin influenced bymatrix carbonate dissolution. As mats growupwards, the mat-forming organisms in the oldermats die off. In turn, the bacteria associated withthe decaying polysaccharide sheath of the micro-organisms may play a major role in mineraliza-tion. Mineral precipitation nucleates at varioussites in the sheath with characteristics that arespecies specific (Riding, 2000). The carbonatesaturation state and the viscosity of the mediumand the sheath influence the mineralogy and
826 V. P. Rao et al.
� 2003 International Association of Sedimentologists, Sedimentology, 50, 819–830

morphology of the precipitates (Pentecost &Riding, 1986; Buczynski & Chaftez, 1991; Riding,2000).
By analogy, we presume that similar biogeo-chemical factors operated on the sheaths of thefossil mats to form dolomite stromatolites. Theconditions of dolomitization, however, were sul-phate reducing and hypersaline. Laboratory cul-tures have shown that microbes use sulphate fortheir metabolism under sulphate-reducing condi-tions, which may in turn help to overcome theinhibiting effect of dissolved sulphate on dolomiteprecipitation (Warthmann et al., 2000). The role ofbacteria during degradation of organic sheaths isto increase the pH and alkalinity in the microen-vironment proximal to the sheaths and to providethe necessary conditions for dolomite precipita-tion (Slaughter & Hill, 1991). The more positived13C values of the crust than expected via micro-bial mediation may indicate high proportions inporewaters of carbonate dissolved from precursorcalcites (see aforementioned discussion).
The importance of sheaths and microenviron-ments around bacterial cells and tubules, inwhich microbes maintain anoxic conditions andthe ion concentrations required to overcome thekinetic inhibiting factors for dolomite precipita-tion is emphasized. Cyanobacterial sheaths areknown to contain high contents of unhydratedMg (Gebelian & Hoffman, 1973), which may havebeen available in the pore microenvironmentsupon release from the sheaths. Variations in themorphology of the dolomites, rhombohedralcrystals (Fig. 3E and F) or submicron-sized grains(Fig. 4D and E) may reflect differences in thesheath material and associated environmen-tal conditions. Submicron-sized grains on micro-bial cells (Fig. 4D) may well be mineralizednannobacteria, as proposed by Folk (1993).Spherular dolomites from the Coorong area arecomposed of 100 nm diameter spherical particles(Von der Borch & Jones, 1976). Well-ordered andnear-stoichiometric dolomite and abundant mi-crocrystalline dolomite with few relics of micro-organisms (Figs 3F and 4F) in the dark-greyportions of the crust are interpreted to recordprogressive and continued mineralization oforganic sheaths (see Fig. 4C). Microcrystallinedolomite fabric, which preserves no clues to itsorigin (Fig. 3F), is probably the end-product ofprogressive dolomitization. It is proposed that thedolomite should be considered as a biomineral.
It has been suggested that amorphous mineralsmay predominate in the early stages of biominer-alization reactions, after which these minerals
may transform into less hydrated, more crystal-line phases depending on the interaction betweenlocalized chemical processes and microbial cel-lular organisms. Various chemical and biochemi-cal factors can influence the crystallo-chemicalproperties, crystal size, structure, morphologyand composition of the mineral (see Krumbein,1986). The shift in the observed XRD peak ofdolomite from 30Æ8� 2h to 31� 2h from the light-grey to dark-grey regions of the crusts may be anindication of transformation of the primarypoorly ordered and non-stoichiometric mineralphase towards a well-ordered near-stoichiometricdolomite.
Implications of microbially mediateddolomite
Sea level was at )120 m during the Last GlacialMaximum (LGM) at about 18 000 years BP. Atpresent, water depths on the carbonate platformrange between 60 and 90 m but, in depressions ofthe platform, the depth reaches up to 110 m. Thus,the platform should have been subaeriallyexposed, and marine carbonate deposits wouldnot have formed during the LGM. The studieddolomite crusts formed at shallow subtidal depths,but yield a radiocarbon age of 17 750 years BP.Aragonite oolites obtained at 88 m depth yield aradiocarbon age of 21 000 years BP (J. D. Milliman,pers. comm., 2002). These radiocarbon ages implythat the platform was not exposed to subaerialconditions, but was at shallow water depths duringthe LGM. Other radiocarbon ages of coralline algallimestones, corals, aragonite faecal pellets andoolites from the carbonate platform range between12 440 years and 6730 years BP. Corals collectedfrom 25 m water depth in the Gulf of Kachchh,located north of the platform, are dated at10 130 years BP. Thus, the ages and estimateddepth of formation of these deposits do not corres-pond with that predicted from the glacio-eustaticsea-level curve (Fairbanks, 1989). These resultsnecessitate late Quaternary neotectonic activityalong the north-western margin of India, especiallyon the carbonate platform, during the late Pleisto-cene and Holocene as previously proposed by Raoet al. (1996, 2003).
The dolomite crusts in the sediments of thelate Quaternary carbonate platform off westernIndia are interpreted to be of primary and earlydiagenetic origin, and to have formed by micro-bial processes in shallow, hypersaline andanoxic porewaters. Subtidal massive dolomitesuccessions have been reported extensively on
Microbial dolomite crusts off western India 827
� 2003 International Association of Sedimentologists, Sedimentology, 50, 819–830

ancient carbonate platforms (Machel & Mountjoy,1986). The majority of these dolomitized succes-sions contain evidence of fossil cyanobacteria,suggesting that the ‘seed’ for ancient dolomites inshallow-water carbonate successions may bemicrobial dolomites. For example, Wright (1997,2000) reported dolomitization of Archean andPalaeozoic ooids and carbonates, which he inter-preted to have formed in response to the decay ofcyanobacteria. Gasiewicz et al. (1987) suggesteddolomite formation in Upper Permian sequencesby chemoheterotrophic bacteria under anaerobicconditions. However, as documented in thisstudy, progressive mineralization (Figs 3D–Fand 4C and F) can obliterate the signatures ofmicrobial activity (see also Brehm et al., 2002).Ancient massive dolomites have been shown tobe susceptible to multiple episodes of recrystal-lization during burial (Gao & Land, 1991; Kupecz& Land, 1991; Montanez & Read, 1992; Montanez,1994) that may mask the origins of the earliestformed dolomite. The results of this study arguefor a re-evaluation of the possible influence ofmicrobes and their decomposition on dolomiteprecipitation in ancient carbonate successions.
CONCLUSIONS
Late Pleistocene dolomite crusts are shown tohave formed by trapping and binding of sedimentdetritus by microbial mats. These dolomite stro-matolites are interpreted to have formed underhypersaline and anoxic sulphate-reducing condi-tions. Sulphate-reducing bacteria associated withthe decaying organic sheaths promoted dolomiteformation. This early formed dolomite is there-fore considered a biomineral. Progressive miner-alization of sheaths resulted in microcrystallinedolomite that does not preserve original microbialfabrics. We suggest that microbial dolomites mayhave provided the ‘seed’ for the origin of dolomitein many ancient carbonate successions. The ageof the dolomite crusts further suggests that theplatform may not have been exposed during theLast Glacial Maximum.
ACKNOWLEDGEMENTS
Dr Rao wishes to thank Dr E. Desa, Director,National Institute of Oceanography, Goa, India,for facilities and encouragement. This work wasfinanced by Research Project No. ESS/CA/A1-25/97, Department of Science and Technology, New
Delhi (V. P. Rao). This work is NIO contributionno. 3815.
REFERENCES
Adams, J.E. and Rhodes, M.L. (1960) Dolomitization by
seepage reflux. AAPG Bull., 44, 1912–1921.
Badiozamani, K. (1973) The Dorag dolomitization model –
application to the Middle Ordovician of Wisconsin. J. Sed.
Petrol., 43, 965–984.
Baker, P.A. and Kastner, M. (1981) Constraints on the for-
mation of sedimentary dolomite. Science, 213, 214–216.
Borg, R.W. and von der Borch, C.C. (1984) Stable isotope
study of carbonate sediments from the Coorong area, south
Australia. Sedimentology, 31, 837–849.
Brehm, U., Gasiewicz, A., Gerdes, G. and Krumbein, W.E.(2002) Biolaminoid facies in a peritidal sabkha: Permian
platy dolomite of Northern Poland. Int. J. Earth Sci., 91,260–271.
Buczynski, C. and Chaftez, H.S. (1991) Habit of bacterially
inducedprecipitatesofcalciumcarbonateandtheinfluenceof
medium viscosity on mineralogy. J. Sed. Petrol., 61, 226–233.
Burne, R.V. and Moore, L.S. (1987) Microbialites: organosed-
imentary deposits of benthic microbial communities. Pala-
ios, 2, 241–254.
Carballo, J.D., Land, L.S. and Miser, D.E. (1987) Holocene
dolomitization of supratidal sediments by active tidal
pumping, Sugarloaf Key, Florida. J. Sed. Petrol., 57, 153–165.
Cavagna, S., Clari, P. and Martire, L. (1999) The role of bac-
teria in the formation of cold seep carbonates: geological
evidence from Monferrato (Tertiary, NW Italy). Sed. Geol.,
126, 253–270.
Compton, J.S. (1988) Degree of supersaturation and precipi-
tation of organogenic dolomite. Geology, 16, 318–321.
Craig, H. (1957) Isotopic standards for carbon and oxygen and
correction factors for mass spectrometric analysis of carbon
dioxide. Geochim. Cosmochim. Acta, 12, 133–149.
Deffeyes, K.S., Lucia, F.J. and Weyl, P.K. (1965) Dolomitiza-
tion of recent and Plio-Pleistocene sediments by marine
evaporite waters on Bonaire, Netherlands Antilles. In: Dol-
omitization and Limestone Diagenesis (Eds L.C. Pray and
R.C. Murray), SEPM Spec. Publ., 13, 71–88.
Fairbanks, R.G. (1989) A 17,000 year glacio-eustatic sea level
record: influence of glacial melting rates on the Younger
Dryas event and deep ocean circulation. Nature, 342, 637–
642.
Feldmann, M. and McKenzie, J.A. (1997) Messinian stromat-
olite–thrombolite associations, Santa Pola, SE Spain: an
analogue for Palaeozoic. Sedimentology, 44, 893–914.
Folk, R.L. (1993) SEM imaging of bacteria and nanobacteria in
carbonate sediments and rocks. J. Sed. Petrol., 63, 990–999.
Gao, G. and Land, L.S. (1991) Early Ordovician Cool Creek
dolomite, Middle Arbuckle Group, Slick hills, SW Okla-
homa, USA: origin and modification. J. Sed. Petrol., 61, 161–
173.
Garrels, R.M. and Thompson, M.E. (1962) A chemical model
for sea water at 25�C and one atmospheric total pressure.
Am. J. Sci., 260, 57–66.
Gasiewicz, A., Gerdes, G. and Krumbein, W.E. (1987) The
peritidal sabkha type stromatolites of the platy dolomite
(Ca3) of the Leba elevation (northern Poland). In: The
Zechstein Facies in Europe. Lecture Notes in Earth Sciences
(Ed. T.M. Peryt), 10, pp. 253–272. Springer, Berlin.
828 V. P. Rao et al.
� 2003 International Association of Sedimentologists, Sedimentology, 50, 819–830

Gebelian, C.D. and Hoffman, P. (1973) Algal origin of dolomite
laminations in stromatolitic limestone. J. Sed. Petrol., 43,603–613.
Hardie, L.A. (1987) Dolomitization: a critical view of some
current views. J. Sed. Petrol., 57, 166–183.
Horodyski, R.J. and Vonder Haar, S. (1975) Recent calcareous
stromatolites from Laguna Mormona (Baja California). J. Sed.
Petrol., 45, 894–906.
Humphery, J.D. and Quinn, T.M. (1990) Coastal mixing zone
dolomite, forward modelling and massive dolomitization of
platform margin carbonates – reply. J. Sed. Petrol., 60, 1013–
1016.
Humphery, J.D. and Radjef, E.M. (1991) Dolomite stochio-
metric variability resulting from changing aquifer condi-
tions, Barbados, West Indies. Sed. Geol., 71, 129–136.
Humphrey, J.D. (2000) New geochemical support for mixing-
zone dolomitization at Golden Grove, Barbados. J. Sed. Res.,
70, 1160–1170.
Illings, L.V., Wells, A.J. and Taylor, J.C.M. (1965) Penecon-
temporary dolomite in the Persian Gulf. In: Dolomitization
and Limestone Diagenesis (Eds L.C. Pray and R.C. Murray),
SEPM Spec. Publ., 13, 89–111.
Irwin, H. (1980) Early diagenetic carbonate precipitation and
pore fluid migration in the Kimmeridge Clay of Dorset,
England. Sedimentology, 27, 577–591.
Kazmierczak, J. and Iryu, Y. (1999) Cyanobacterial origin of
microcrystalline cements from Pleistocene rhodoliths and
coralline algal crusts of Okierabu-jima, Japan. Acta Palae-
ontol. Polonica, 44, 117–130.
Kelts, K. and McKenzie, J.A. (1982) Diagenetic dolomite for-
mation in Quaternary anoxic diatomaceous muds of DSDP
Leg 64, Gulf of California. In: Initial Reports of the Deep Sea
Drilling Project (Eds J.R. Curray, D.G. Moore et al.), 64, part
2, pp. 553–569. US Government Printing Office, Washing-
ton, DC.
Krumbein, W.E. (1983) Introduction. In: Microbial Geochem-
istry (Ed. W.E. Krumbein), pp. 1–4. Blackwell Scientific,
Oxford.
Krumbein, W.E. (1986) Biotransfer of minerals by microbes
and microbial mats. In: Biomineralization in Lower Plants
and Animals (Eds B.S.C. Leadbeater and R. Riding), pp. 54–
72. Oxford University Press, Oxford.
Krumbein, W.E. and Cohen, Y. (1977) Primary production,
mat formation and lithification: contribution of oxygenic
and facultative anoxygenic cyanobacteria. In: Fossil Algae(Ed. E. Flugel), pp. 37–56. Springer-Verlag, Berlin.
Kupecz, J.A. and Land, L.S. (1991) Late stage dolomitization of
Lower Ordovician Ellenburger Group, West Texas. J. Sed.Petrol., 61, 551–574.
Land, L.S. (1980) The isotopic and trace element geochemistry
of dolomite: the state of the art. SEPM Spec. Publ., 28, 87–110.
Lippmann, F. (1973) Sedimentary Carbonate Minerals.Springer-Verlag, Berlin, 230 pp.
Machel, H.G. and Mountjoy, E.W. (1986) Chemistry and
environment of dolomitization – a reappraisal. Earth-Sci.
Rev., 23, 175–222.
Melim, L.A. and Scholle, P.A. (2002) Dolomitization of the
Capitan Formation forereef facies (Permian, west Texas and
New Mexico): seepage reflux revisited. Sedimentology, 49,1207–1227.
Milliman, J.D. (1974) Marine Carbonates. Springer-Verlag,
Berlin, 375 pp.
Mitchel, J.T. and Land, L.S. (1987) Modern marine dolomite
cement in a north Jamaican fringing reef. Geology, 15, 557–
560.
Montanez, I.P. (1994) Late diagenetic dolomitization of Lower
Ordovician Upper Knox Carbonates: a record of the hydro-
dynamic evolution of the southern Appalachian Basin.
AAPG Bull., 78, 1210–1239.
Montanez, I.P. and Osleger, D.A. (1993) Parasequence stacking
patterns, third-order accommodation events and sequence
stratigraphy of Middle to upper Cambrian platform car-
bonates, Bonanza King Formation, southern Great Basin. In:
Carbonate Sequence Stratigraphy: Recent Developments
and Applications (Eds R.G. Loucks and F.R. Sarg), AAPG
Mem., 57, 305–326.
Montanez, I.P. and Read, J.F. (1992) Fluid–rock interaction
history during stabilization of early dolomites, Upper Knox
Group (Lower Ordovician), US Appalachians. J. Sed. Petrol.,
62, 753–778.
Nadson, G.A. (1928) Beitrag zur Kenntnis der Bakteriogenen
Kalkablagerungen. Arch. Hydrobiol., 19, 154–164.
Neher, J. (1959) Bakterien in Tieferliegenden Gesteinslagen.
Eclogae Geol. Helv., 52, 619–625.
Patterson, R.J. and Kinsman, D.J. (1982) Formation of dia-
genetic dolomite in coastal sabkha along Persian Gulf.
AAPG Bull., 66, 28–43.
Pentecost, A. and Riding, R. (1986) Calcification in cyano-
bacteria. In: Biomineralization in Lower Plants and Animals
(Eds B.S.C. Leadbeater and R. Riding), pp. 73–90. Oxford
University Press, Oxford.
Rao, V.P., Veerayya, M., Nair, R.R., Dupeuble, P.A. and
Lamboy, M. (1994) Late Quaternary Halimeda bioherms and
aragonitic faecal pellet-dominated sediments on the car-
bonate platform of the western continental shelf of India.
Mar. Geol., 121, 294–315.
Rao, V.P., Veerayya, M., Thamban, M. and Wagle, B.G. (1996)
Evidences of late Quaternary neotectonic activity and sea
level changes along the western margin of India. Curr. Sci.,
71, 213–219.
Rao, V.P., Rajagopalan, G., Vora, K.H. and Almeida, F. (2003)
Late Quaternary sea level and environmental changes from
relic carbonate deposits of the western margin of India.
Proc. Indian Acad. Sci., 112, 1–25.
Ray, J.S. and Ramesh, R. (1998) Stable carbon and oxygen iso-
tope analysis of natural calcite and dolomite mixtures using
selective acid extraction. J. Geol. Soc. India, 52, 323–332.
Riding, R. (2000) Microbial carbonates: the geological record
of calcified bacterial algal mats and biofilms. Sedimentolo-
gy, 47, 179–214.
Rosen, M.R. and Coshell, L. (1992) A new location of Holocene
dolomite formation, Lake Hayward, Western Australia.
Sedimentology, 39, 1062–1074.
Rosen, M.R., Misser, D.E. and Warren, J.K. (1988) Sedimen-
tation, mineralogy and isotopic analysis of Pellet Lake,
Coorong region, South Australia. Sedimentology, 35, 105–
122.
Rosenbaum, J. and Sheppard, S.M.F. (1986) An isotopic study
of siderites, dolomites and ankerites at high temperatures.
Geochim. Cosmochim. Acta, 50, 1147–1150.
Sarkar, A., Ramesh, R., Somayajulu, B.L.K., Agnihotri, R.,Jull, A.J.T. and Burr, G.S. (2000) High-resolution Holocene
monsoon record from the eastern Arabian Sea. Earth Planet.
Sci. Lett., 177, 209–218.
Shinn, E.A. (1968) Selective dolomitization of recent sedi-
mentary structures. J. Sed. Petrol., 38, 212–216.
Shinn, E.A., Ginsburg, R.N. and Lloyd, R.M. (1965) Recent
supratidal dolomite from Andros Island, Bahamas. In:
Dolomitization and Limestone Diagenesis (Eds L.C. Pray
and R.C. Murray), SEPM Spec. Publ., 13, 112–123.
Microbial dolomite crusts off western India 829
� 2003 International Association of Sedimentologists, Sedimentology, 50, 819–830

Slaughter, M. and Hill, R.J. (1991) The influence of organic
matter in organogenic dolomitization. J. Sed. Petrol., 61,296–303.
Soudry, D. and Lewy, Z. (1988) Microbially influenced for-
mation of phosphate nodules and Mega fossil molds (Negev,
southern Israel). Palaeogeogr. Palaeoclimatol. Palaeoecol.,
84, 15–34.
Swart, P.K. and Melim, L.A. (2000) The origin of dolomites in
Tertiary sediments from the margin of Great Bahama Bank.
J. Sed. Res., 70, 738–748.
Vahrenkamp, V.C., Swart, P.K. and Ruiz, J. (1991) Episodic
dolomitization of late Cenozoic carbonates in the Bahamas –
evidence from strontium isotopes. J. Sed. Petrol., 61, 1002–
1014.
Valdiya, K.S. (1972) Origin of phosphorite of the Late Pre-
cambrian Gangolihat dolomite of Pithoragarh, Kumaun
Himalaya. Sedimentology, 19, 115–128.
Vasconcelos, C. and McKenzie, J.A. (1997) Microbial medi-
ation of modern dolomite precipitation and diagenesis
under anoxic conditions (Lagoa Vermelha, Rio de Janeiro,
Brazil). J. Sed. Res., 67, 378–390.
Vasconcelos, C., McKenzie, J.A., Bernasconi, S., Grujic, D.and Tien, A.J. (1995) Microbial mediation as a possible
mechanism for dolomite formation. Nature, 377, 220–222.
Von der Borch, C.C. and Jones, J.B. (1976) Spherular modern
dolomite from the Coorong area, South Australia. Sedi-
mentology, 23, 587–591.
Von der Borch, C.C. and Lock, D. (1979) Geological signifi-
cance of Coorong dolomites. Sedimentology, 26, 813–824.
Walter, M.R. (1976) Stromatolites. Developments in Sedi-
mentology 20. Elsevier, Amsterdam, 790 pp.
Warthmannn, R., Lith, Y.V., Vasconcelos, C., McKenzie, J.A.and Karpoff, A.M. (2000) Bacterially induced dolomite
precipitation in anoxic culture experiments. Geology, 12,1091–1094.
Wright, D.T. (1997) An organogenic origin of widespread
dolomite in the Cambrian Eilean Dubh formation, north-
western Scotland. J. Sed. Petrol., 67, 54–64.
Wright, D.T. (2000) Benthic microbial communities and
dolomite formation in marine and lacustrine environments
– a new dolomite model. In: Marine Authigenesis from
Global to Microbial (Eds C.R. Glenn, J. Lucas and L. Prevot),
SEPM Spec. Publ., 66, 7–20.
Manuscript received 28 January 2002;revision accepted 26 March 2003.
830 V. P. Rao et al.
� 2003 International Association of Sedimentologists, Sedimentology, 50, 819–830





























