Embed Size (px)
Citation preview

Melatonin improves cardiovascular function and ameliorates renal,cardiac and cerebral damage in rats with renovascularhypertension
Introduction
The renin angiotensin system is crucial for the maintenanceof blood pressure (BP), fluid, and sodium homeostasis andthus, plays a major role in the pathogenesis of hyperten-
sion. Renovascular hypertension, a relatively rare form ofsecondary hypertension, is associated with the activation ofrenin angiotensin system as a result of reduced renal blood
flow and perfusion pressure due to renal artery stenosis [1].The immediate hypertensive effects of angiotensin II (AngII) occur as a result of vasoconstriction and antinatriuresis
[2]. However, in the long-term Ang II causes remodeling ofthe arterial vasculature by vascular remodeling, acceleratedatherogenesis, extracellular matrix deposition and glomer-ulosclerosis [3], all of which may contribute to the
progression of cardiovascular and renal damage beyondthe effects of high BP alone [4]. Experimental evidencesuggests that reactive oxygen species (ROS), such as
superoxide anion and hydrogen peroxide (H2O2), whichoccur as a consequence of renal artery stenosis, play asignificant role in the hypertensive and hypertrophic
responses to Ang II [5–8].In physiological conditions, ROS are produced at low
concentrations and function as signaling molecules to
maintain vascular integrity by regulating endothelial func-tion and vascular contraction/relaxation [9]. However,under pathological conditions, increased ROS bioactivity
leads to endothelial dysfunction, increased contractility,vascular smooth muscle cell (VSMC) growth, monocyteinvasion, lipid peroxidation, inflammation, and increased
deposition of extracellular matrix proteins, importantfactors in hypertensive vascular damage [10]. It is wellknown that both endothelial cells and VSMC are capable of
producing superoxide anions (O2–•) and other oxygen-
derived radical metabolites [7]. Increased vascular O2)•
production in Ang II-induced hypertension was shown toreduce the biological activity of endothelium-derived nitric
oxide (NO) [11]. On the other hand, increased productionof H2O2 in response to Ang II also plays a critical role inthe development of vascular smooth muscle hypertrophy
[6]. Structural alterations in the microcirculation, which areconsidered as the cause of hypertension, are also acceptedas the basis of hypertension-related target organ damages in
the brain, heart, or kidneys [12].Melatonin, the major product of the vertebral pineal
gland, functions as a modulator of sleep, sexual behavior,
immune function, and circadian rhythms. Following theidentification of melatonin (N-acetyl-5-methoxytryptamine)
Abstract: The effect of melatonin was investigated in an angiotensin
II-dependent renovascular hypertension model in Wistar albino rats by
placing a renal artery clip (two-kidney, one-clip; 2K1C), while sham rats did
not have clip placement. Starting either on the operation day or 3 wk after
the operation, the rats received melatonin (10 mg/kg/day) or vehicle for the
following 6 wk. At the end of the nineth week, after blood pressure (BP) and
echocardiographic recordings were obtained, plasma samples were obtained
to assay lactate dehydrogenase (LDH), creatine kinase (CK), antioxidant
capacity (AOC), asymmetric dimethylarginine (ADMA), and nitric oxide
(NOx) levels. In the kidney, heart and brain tissues, malondialdehyde
(MDA) and glutathione (GSH) levels, superoxide dismutase (SOD), catalase
(CAT), myeloperoxidase (MPO) and Na+-K+ ATPase activities were
determined. 2K1C caused an increase in BP and left ventricular (LV)
dysfunction. In hypertensive animals LDH, CK, ADMA levels were
increased in plasma with a concomitant reduction in AOC and NOx.
Moreover, hypertension caused a significant decrease in tissue SOD, CAT,
and Na+, K+-ATPase activities and glutathione content, while MDA levels
and MPO activity were increased in all studied tissues. On the other hand,
both melatonin regimens significantly reduced BP, alleviated oxidative injury
and improved LV function. In conclusion, melatonin protected against
renovascular hypertension-induced tissue damage and improved cardiac
function presumably due to both its direct antioxidant and receptor-
dependent actions, suggesting that melatonin may be of therapeutic use in
preventing oxidative stress due to hypertension.
Mehmet Ersahin1, Ozer Sehirli2,Hale Z. Toklu2, SelamiSuleymanoglu3, Ebru Emekli-Alturfan4, Aysen Yarat4, ElifTatlıdede2, Berrak C. Yegen5 andGoksel Sener2
1Department of Neurosurgery, Haydarpasa
Numune Education and Research Hospital;2Department of Pharmacology, Marmara
University School of Pharmacy; 3Department
of Pediatric Cardiology, Gulhane Military
Medical Academy; 4Department of
Biochemistry, Marmara University School of
Dentistry; 5Department of Physiology,
Marmara University School of Medicine,
Istanbul, Turkey
Key words: asymmetric dimethylarginine,
hypertension, melatonin, oxidative stress
Address reprint requests to Goksel Sener, PhD,
Marmara University, School of Pharmacy,
Tıbbiye Cad. 34668 _Istanbul, Turkey.
E-mail: [email protected],
Received April 8, 2009;
accepted May 8, 2009.
J. Pineal Res. 2009; 47:97–106Doi:10.1111/j.1600-079X.2009.00693.x
� 2009 The AuthorsJournal compilation � 2009 John Wiley & Sons A/S
Journal of Pineal Research
97
Mo
lecu
lar,
Bio
log
ical
,Ph
ysio
log
ical
an
d C
lin
ical
Asp
ects
of
Mel
ato
nin

by Lerner et al. [13], it has been shown that melatonin isinvolved in the regulation of many physiological systemsincluding cardiovascular system [14, 15]. Melatonin reduces
BP [16–19] and has an anti-adrenergic action on myocardialcontractility [20], which are mediated by its receptors in theheart [21] and arteries [22]. On the other hand, actingdirectly as an electron donor, melatonin scavenges free
radicals, stimulates antioxidant enzyme systems [23, 24] andalong with its metabolites it has powerful anti-inflamma-tory properties proven to be highly effective in reducing
oxidative stress and inflammation [25, 26].In the light of above findings, we tested the hypothesis
that melatonin could alleviate renovascular hypertension
through its antioxidant effects, as well as by its modulatoryrole on cardiovascular functions. Therefore, in an angio-tensin II-dependent renovascular hypertension model, ther-apeutic effects of melatonin on cardiovascular functions, as
well as its protective effects against oxidative damage intarget organs were evaluated using various physiologicaland biochemical parameters.
Materials and methods
Animals
All experimental protocols were approved by the Marmara
University Animal Care and Use Committee. Male Wistaralbino rats (200–250 g) were kept at a constant temperature(22 ± 1�C) with 12 hr light and dark cycles and fed astandard rat chow.
Surgery and experimental design
Two-kidney, one-clip (2K1C) has been studied as an AngII-dependent model of renovascular hypertension withelevated circulating levels of Ang II and high Ang II
concentration in the cortical tissue of the clipped and non-clipped kidneys [27]. Clipping of the left renal artery andsham surgery were performed as previously described [28].Briefly, a silver clip (internal diameter 0.25 mm) was placed
around the left renal artery (2K1C group; n = 24) of therats that were anesthetized with ketamine (100 mg/kg) andchlorpromazine (0.75 mg/kg) given intraperitoneally (i.p.).
In the sham-operated control group (n = 8), animals hadsimilar surgical procedures without clip placement. Startingon the day of surgery, rats received either melatonin
(10 mg/kg/day; i.p.) or vehicle (1 mL/kg/day saline with1% alcohol) for 9 wk (early treatment), while in a subgroupof 2K1C rats melatonin treatment was started at the end of
the third week following the surgery and continued for theremaining 6 wk (late treatment).To obtain basal readings, systolic BP and heart rate (HR)
recordings were obtained in all rats before the surgical
procedures, and these measurements were repeated at theend of the third and nineth weeks after the surgeries. At theend of the nineth week, echocardiographic measurements
were done in all rats before they were decapitated. Trunkblood was collected and immediately centrifuged at 3000 gfor 10 min to assay the levels of lactate dehydrogenase
(LDH), creatine kinase (CK), antioxidant capacity (AOC),asymmetric dimethylarginine (ADMA) and nitric oxide
(NO) metabolites in the plasma. In order to evaluate targetorgan injury, renal, cardiac and cerebral tissue sampleswere taken and stored at )80�C to determining the
superoxide dismutase (SOD), catalase (CAT), malondial-dehyde (MDA) and glutathione (GSH) levels, along withmyelopreoxidase and Na+, K+-ATPase activities.
Measurement of blood pressure
Indirect BP measurement was made by the tail cuff method
(Bipoac Systems, Goleta, CA, USA) before the surgery andat the end of third and nineth weeks following surgery.Initially, the rats were placed for 10 min in a chamber
heated to 35�C. Then the rats were placed in individualplastic restrainers and a cuff with a pneumatic pulse sensorwas wrapped around their tails. Blood pressure recordedduring each measurement period was averaged from at least
three consecutive readings on that occasion obtained fromeach rat.
Echocardiography
Echocardiographic imaging and calculations were done
according to the guidelines published by the AmericanSociety of Echocardiography [29] using a 12 MHz lineartransducer and 5–8 MHz sector transducer (Vivid 3,Gen-
eral Electric Medical Systems Ultrasound, Tirat Carmel,Israel). Under ketamine (50 mg/kg, ip) anesthesia, mea-surements were made from M-mode and two-dimensionalimages obtained in the parasternal long and short axes at
the level of the papillary muscles after observation of atleast 6 cardiac cycles. Interventricular septal thickness(IVS), left ventricular diameter (LVD) and left ventricular
posterior wall thickness (LVPW) were measured duringsystole (s) and diastole (d). Ejection fraction (EF), frac-tional shortening (FS) and left ventricular mass (LVM) and
relative wall thickness (RWT) were calculated fromthe M-mode images using the following formulas:%EF ¼ ½ðLVDdÞ3 � ðLVDsÞ3=ðLVDdÞ3 � 100�; %FS ¼½LVDd-LVDs/LVDd� 100�; LVM ¼ ½1:04� ððLVDdþLVPWd+IVSdÞ3 � ðLVDdÞ3Þ � 0:8þ 0:14�; RWT ¼ ½2�ðLVPWd/LVDd)�.
Plasma assays
Plasma levels of LDH and CK were determined spectro-
photometrically using an automated analyzer (Bayer OperaBiochemical Analyzer, Leverkusen, Germany), while totalAOC was measured by using a colorimetric test system
(ImAnOx, Immunodiagnostic AG, Bensheim, Germany),according to the instructions provided by the manufacturer.NO metabolites (nitrates and nitrites) were assayed inplasma by the colorimetric method of Griess after enzy-
matic conversion of nitrates to nitrites by nitrate reductaseusing a colorimetric assay kit (Cayman Chemical, AnnAr-bor, MI, USA). ADMA concentration in plasma was
measured with a highly sensitive ELISA kit (Immunodiag-nostic AG). The intensity of the color reaction, measuredby reading the optical density at 450 nm with a microtiter
plate reader, is known to be inversely proportional with theamount of ADMA in the sample.
Ersahin et al.
98

Measurement of tissue superoxide dismutase andcatalase activities
Superoxide dismutase activity in the heart, kidney andbrain samples was measured according to the previouslydescribed method [30]. Briefly, measurements were per-formed in cuvettes containing 2.8 mL 50 mm potassium
phosphate (pH = 7.8) with 0.1 mm EDTA, 0.1 mm
0.39 mm riboflavin in 10 mm potasium phosphate (pH7.5), 0.1 mL of 6 mm O-dianisidin.2 HCl in deionized
water, and tissue extract (50, 100 lL). Cuvettes with alltheir components were illuminated with 20-W SlylvaniaGrow Lux fluorecent tubes that were placed 5 cm above
and to one side of cuvettes maintaining a temperature of37�C. Absorbance were measured at 460 nm with Schi-madzu UV-02 model spectrophotometer. A standard curvewas prepared routinely with bovine SOD (Sigma Chemical
Co, Catalog Number: S-2515, 3000 Unit). Absorbancereadings were taken at 0 and 8 min of illumination and thenet absorbance were calculated.
The method for the measurement of CAT activity isbased on the catalytic activity of the enzyme whichcatalyses the decomposition reaction of H2O2 to give H2O
and O2 [31]. Briefly, the absorbances of the tissue samplescontaining 0.4 mL homogenate and 0.2 mL H2O2 wereread at 240 nm and 20�C against a blank containing 0.2 mL
phosphate buffer and 0.4 mL homogenate for about 1 min.
Measurement of tissue malondialdehyde andglutathione levels
Heart, kidney and brain samples were homogenized withice-cold 150 mm KCl for the determination of MDA and
GSH levels. The MDA levels were assayed for products oflipid peroxidation by monitoring thiobarbituric acid reac-tive substance formation as described previously [32]. Lipid
peroxidation was expressed in terms of MDA equivalentsusing an extinction coefficient of 1.56 · 105/m/cm andresults are expressed as nmol MDA/g tissue. GSH mea-surements were performed using a modification of the
Ellman procedure [33]. Briefly, after centrifugation at1500 g for 10 min, 0.5 mL of supernatant was added to2 mL of 0.3 mol/L Na2HPO4.2H2O solution. A 0.2 mL
solution of dithiobisnitrobenzoate (0.4 mg/mL 1% sodiumcitrate) was added and the absorbance at 412 nm wasmeasured immediately after mixing. GSH levels were calcu-
lated using an extinction coefficient of 1.36 · 104/m/cm.Results are expressed in lmol GSH/g tissue.
Measurement of tissue myeloperoxidase activity
Since the activity of myeloperoxidase (MPO), an enzymethat is found predominantly in the azurophilic granules of
polymorphonuclear leukocytes (PMN), correlates with thehistochemically determined PMN in tissues, MPO activityis frequently utilized to estimate PMN accumulation in the
inflamed tissues. MPO activity was measured in tissues in aprocedure similar to that documented by Hillegass et al.[34]. Tissue samples were homogenized in 50 mm potassium
phosphate buffer (PB, pH 6.0), and centrifuged at 41,400 g(10 min); pellets were suspended in 50 mm PB containing
0.5% hexadecyltrimethylammonium bromide (HETAB).After three freeze and thaw cycles, with sonication betweencycles, the samples were centrifuged at 41,400 g for 10 min.
Aliquots (0.3 mL) were added to 2.3 mL of reactionmixture containing 50 mm PB, o-dianisidine, and 20 mm
H2O2 solution. One unit of enzyme activity was defined asthe amount of MPO present that caused a change in
absorbance measured at 460 nm for 3 min. MPO activitywas expressed as U/g tissue.
Measurement of Na+, K+-ATPase activity
The activity of Na+, K+-ATPase, a membrane-bound
enzyme required for cellular transport, is very sensitive tofree radical reactions and lipid peroxidation. Accordingly, areduction in Na+, K+-ATPase activity indirectly indicatesmembrane damage. Measurement of Na+, K+-ATPase
activity is based on the measurement of inorganic phos-phate released by ATP hydrolysis during incubation ofhomogenates with an appropriate medium. The total
ATPase activity was determined in the presence of100 mm NaCl, 5 mm KCl, 6 mm MgCl2, 0.1 mm EDTA,30 mm Tris–HCl (pH 7.4), while the Mg2+-ATPase activity
was determined in the presence of 1 mm ouabain. Thedifference between the total and the Mg2+-ATPase activ-ities was taken as a measure of the Na+, K+-ATPase
activity [35, 36]. The reaction was initiated with theaddition of the homogenate (0.1 mL) and a 5-min pre-incubation period at 37�C was allowed. Following theaddition of 3 mm Na2ATP and a 10-min re-incubation
period, the reaction was terminated by the addition of ice-cold 6% perchloric acid. The mixture was then centrifugedat 3500 g, and Pi in the supernatant fraction was deter-
mined by the method of Fiske and Subarrow [37]. Thespecific activity of the enzyme was expressed as nmol Pi/mgprotein/h. The protein concentration of the supernatant
was measured by the Lowry method [38].
Statistics
Statistical analysis was carried out using GraphPad Prism3.0 (GraphPad Software, San Diego, CA, USA). Eachgroup consisted of 8 animals. All data were expressed as
mean ± S.E.M. Groups of data were compared with ananalysis of variance (ANOVA) followed by Tukey�s multi-ple comparison tests. Values of P < 0.05 were regarded as
significant.
Results
The basal blood pressures that were recorded before thesurgery were not different among the groups (Fig. 1). In thevehicle-treated group with 2K1C, the BP was significantly
elevated at the third- (172 ± 6.2 mmHg; P < 0.001) andninth- (190 ± 9.3 mmHg; P < 0.001) week recordingswith respect to basal values. Similarly, at the third week
measurement of the 2K1C late treatment group that has notreceived melatonin yet, BP was elevated (169 ± 7.5 mmHg;P < 0.001) to the levels of vehicle-treated 2K1C group.
However, at the nineth week measurement of this 2K1Clate treatment group, the BP was reduced significantly
Melatonin treatment improves renovascular hypertension
99
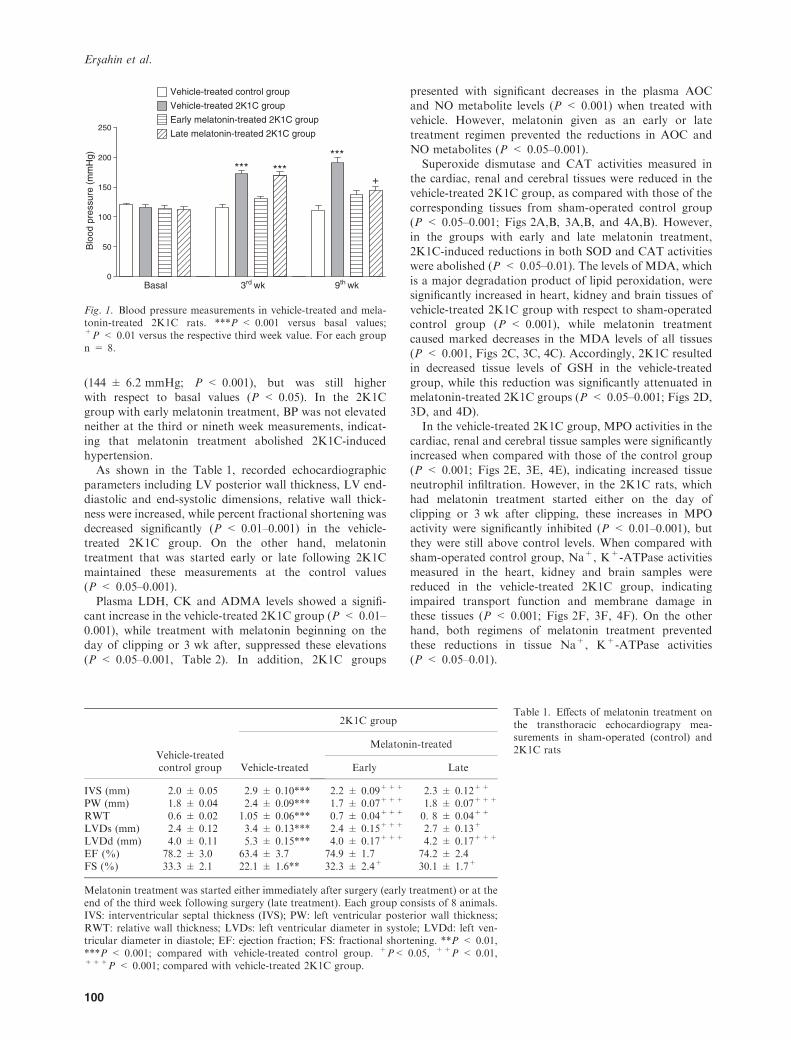
(144 ± 6.2 mmHg; P < 0.001), but was still higherwith respect to basal values (P < 0.05). In the 2K1Cgroup with early melatonin treatment, BP was not elevatedneither at the third or nineth week measurements, indicat-
ing that melatonin treatment abolished 2K1C-inducedhypertension.As shown in the Table 1, recorded echocardiographic
parameters including LV posterior wall thickness, LV end-diastolic and end-systolic dimensions, relative wall thick-ness were increased, while percent fractional shortening was
decreased significantly (P < 0.01–0.001) in the vehicle-treated 2K1C group. On the other hand, melatonintreatment that was started early or late following 2K1C
maintained these measurements at the control values(P < 0.05–0.001).Plasma LDH, CK and ADMA levels showed a signifi-
cant increase in the vehicle-treated 2K1C group (P < 0.01–
0.001), while treatment with melatonin beginning on theday of clipping or 3 wk after, suppressed these elevations(P < 0.05–0.001, Table 2). In addition, 2K1C groups
presented with significant decreases in the plasma AOCand NO metabolite levels (P < 0.001) when treated withvehicle. However, melatonin given as an early or late
treatment regimen prevented the reductions in AOC andNO metabolites (P < 0.05–0.001).Superoxide dismutase and CAT activities measured in
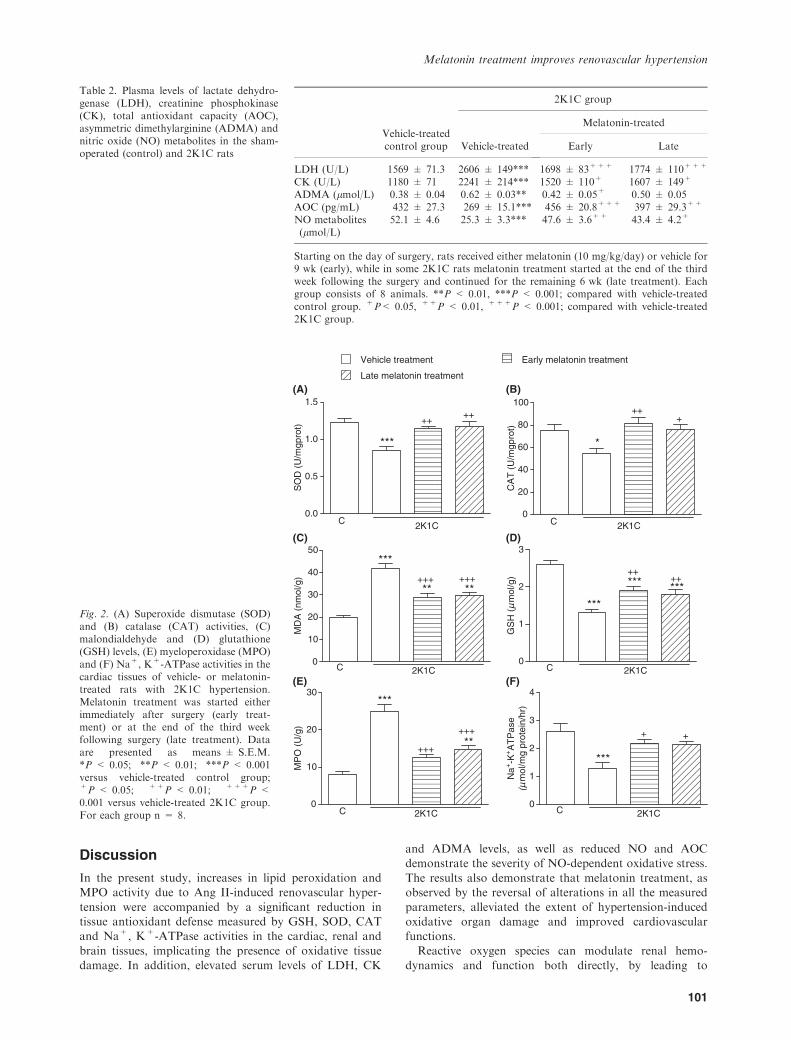
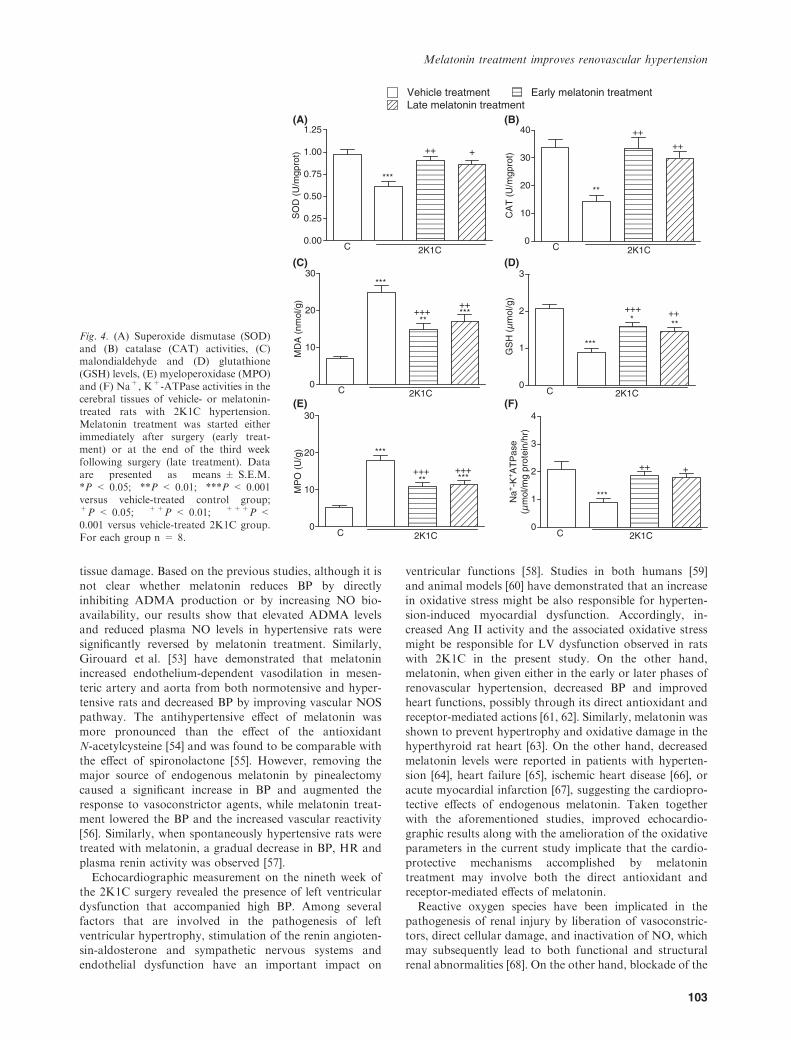
the cardiac, renal and cerebral tissues were reduced in the
vehicle-treated 2K1C group, as compared with those of thecorresponding tissues from sham-operated control group(P < 0.05–0.001; Figs 2A,B, 3A,B, and 4A,B). However,
in the groups with early and late melatonin treatment,2K1C-induced reductions in both SOD and CAT activitieswere abolished (P < 0.05–0.01). The levels of MDA, which
is a major degradation product of lipid peroxidation, weresignificantly increased in heart, kidney and brain tissues ofvehicle-treated 2K1C group with respect to sham-operatedcontrol group (P < 0.001), while melatonin treatment
caused marked decreases in the MDA levels of all tissues(P < 0.001, Figs 2C, 3C, 4C). Accordingly, 2K1C resultedin decreased tissue levels of GSH in the vehicle-treated
group, while this reduction was significantly attenuated inmelatonin-treated 2K1C groups (P < 0.05–0.001; Figs 2D,3D, and 4D).
In the vehicle-treated 2K1C group, MPO activities in thecardiac, renal and cerebral tissue samples were significantlyincreased when compared with those of the control group
(P < 0.001; Figs 2E, 3E, 4E), indicating increased tissueneutrophil infiltration. However, in the 2K1C rats, whichhad melatonin treatment started either on the day ofclipping or 3 wk after clipping, these increases in MPO
activity were significantly inhibited (P < 0.01–0.001), butthey were still above control levels. When compared withsham-operated control group, Na+, K+-ATPase activities
measured in the heart, kidney and brain samples werereduced in the vehicle-treated 2K1C group, indicatingimpaired transport function and membrane damage in
these tissues (P < 0.001; Figs 2F, 3F, 4F). On the otherhand, both regimens of melatonin treatment preventedthese reductions in tissue Na+, K+-ATPase activities
(P < 0.05–0.01).
0
50
100
150
200
250
Basal 3rd wk 9th wk
Vehicle-treated control group
Vehicle-treated 2K1C group
Early melatonin-treated 2K1C group
Late melatonin-treated 2K1C group
*** ******
+
Blo
od p
ress
ure
(mm
Hg)
Fig. 1. Blood pressure measurements in vehicle-treated and mela-tonin-treated 2K1C rats. ***P < 0.001 versus basal values;+P < 0.01 versus the respective third week value. For each groupn = 8.
Table 1. Effects of melatonin treatment onthe transthoracic echocardiograpy mea-surements in sham-operated (control) and2K1C rats
Vehicle-treatedcontrol group
2K1C group
Vehicle-treated
Melatonin-treated
Early Late
IVS (mm) 2.0 ± 0.05 2.9 ± 0.10*** 2.2 ± 0.09+++ 2.3 ± 0.12++
PW (mm) 1.8 ± 0.04 2.4 ± 0.09*** 1.7 ± 0.07+++ 1.8 ± 0.07+++
RWT 0.6 ± 0.02 1.05 ± 0.06*** 0.7 ± 0.04+++ 0. 8 ± 0.04++
LVDs (mm) 2.4 ± 0.12 3.4 ± 0.13*** 2.4 ± 0.15+++ 2.7 ± 0.13+
LVDd (mm) 4.0 ± 0.11 5.3 ± 0.15*** 4.0 ± 0.17+++ 4.2 ± 0.17+++
EF (%) 78.2 ± 3.0 63.4 ± 3.7 74.9 ± 1.7 74.2 ± 2.4FS (%) 33.3 ± 2.1 22.1 ± 1.6** 32.3 ± 2.4+ 30.1 ± 1.7+
Melatonin treatment was started either immediately after surgery (early treatment) or at theend of the third week following surgery (late treatment). Each group consists of 8 animals.IVS: interventricular septal thickness (IVS); PW: left ventricular posterior wall thickness;RWT: relative wall thickness; LVDs: left ventricular diameter in systole; LVDd: left ven-tricular diameter in diastole; EF: ejection fraction; FS: fractional shortening. **P < 0.01,***P < 0.001; compared with vehicle-treated control group. +P< 0.05, ++P < 0.01,+++P < 0.001; compared with vehicle-treated 2K1C group.
Ersahin et al.
100
Discussion
In the present study, increases in lipid peroxidation andMPO activity due to Ang II-induced renovascular hyper-
tension were accompanied by a significant reduction intissue antioxidant defense measured by GSH, SOD, CATand Na+, K+-ATPase activities in the cardiac, renal and
brain tissues, implicating the presence of oxidative tissuedamage. In addition, elevated serum levels of LDH, CK
and ADMA levels, as well as reduced NO and AOCdemonstrate the severity of NO-dependent oxidative stress.
The results also demonstrate that melatonin treatment, asobserved by the reversal of alterations in all the measuredparameters, alleviated the extent of hypertension-induced
oxidative organ damage and improved cardiovascularfunctions.Reactive oxygen species can modulate renal hemo-
dynamics and function both directly, by leading to
Table 2. Plasma levels of lactate dehydro-genase (LDH), creatinine phosphokinase(CK), total antioxidant capacity (AOC),asymmetric dimethylarginine (ADMA) andnitric oxide (NO) metabolites in the sham-operated (control) and 2K1C rats
Vehicle-treatedcontrol group
2K1C group
Vehicle-treated
Melatonin-treated
Early Late
LDH (U/L) 1569 ± 71.3 2606 ± 149*** 1698 ± 83+++ 1774 ± 110+++
CK (U/L) 1180 ± 71 2241 ± 214*** 1520 ± 110+ 1607 ± 149+
ADMA (lmol/L) 0.38 ± 0.04 0.62 ± 0.03** 0.42 ± 0.05+ 0.50 ± 0.05AOC (pg/mL) 432 ± 27.3 269 ± 15.1*** 456 ± 20.8+++ 397 ± 29.3++
NO metabolites(lmol/L)
52.1 ± 4.6 25.3 ± 3.3*** 47.6 ± 3.6++ 43.4 ± 4.2+
Starting on the day of surgery, rats received either melatonin (10 mg/kg/day) or vehicle for9 wk (early), while in some 2K1C rats melatonin treatment started at the end of the thirdweek following the surgery and continued for the remaining 6 wk (late treatment). Eachgroup consists of 8 animals. **P < 0.01, ***P < 0.001; compared with vehicle-treatedcontrol group. +P< 0.05, ++P < 0.01, +++P < 0.001; compared with vehicle-treated2K1C group.
Vehicle treatment Early melatonin treatment
Late melatonin treatment
(A)
0.0
0.5
1.0
1.5
C
***
++++
2K1C
SO
D (
U/m
gpro
t)
(B)
(C)
0
10
20
30
40
50***
****++++++
C 2K1C
MD
A (
nmol
/g)
(D)
0
1
2
3
***
++++
******
C 2K1C
GS
H (
µm
ol/g
)
(E)
0
10
20
30***
**+++
+++
C 2K1C
MP
O (
U/g
)
(F)
0
1
2
3
4
***
++
C 2K1C
Na+
-K+A
TP
ase
(µm
ol/m
g pr
otei
n/hr
)0
20
40
60
80
100
*
+++
C 2K1C
CA
T (
U/m
gpro
t)
Fig. 2. (A) Superoxide dismutase (SOD)and (B) catalase (CAT) activities, (C)malondialdehyde and (D) glutathione(GSH) levels, (E) myeloperoxidase (MPO)and (F) Na+, K+-ATPase activities in thecardiac tissues of vehicle- or melatonin-treated rats with 2K1C hypertension.Melatonin treatment was started eitherimmediately after surgery (early treat-ment) or at the end of the third weekfollowing surgery (late treatment). Dataare presented as means ± S.E.M.*P < 0.05; **P < 0.01; ***P < 0.001versus vehicle-treated control group;+P < 0.05; ++P < 0.01; +++P <0.001 versus vehicle-treated 2K1C group.For each group n = 8.
Melatonin treatment improves renovascular hypertension
101

vasoconstriction, and indirectly, by inducing renal inflam-
mation. Evidence obtained from humans with essentialhypertension [39] and from animal models [7, 40, 41]support the hypothesis that oxidative stress and inflamma-
tion are associated with the development of hypertensionand hypertension-induced damage in target organs [12].Oxidative stress may contribute to the generation andmaintenance of hypertension via the inactivation of NO
[42], which antagonizes the vasoconstrictive and pro-atherosclerotic effects of Ang II in physiological conditions.On the other hand, Ang II also decreases NO bioavailabil-
ity by promoting oxidative stress. O2–•, which is the main
free radical involved in the pathology of different forms ofhypertension, can either cause vasoconstriction or, under
certain circumstances, can interact rapidly with NO toreduce its bioavailability and further increase vasoconstric-tion and BP [43]. In the 2K1C hypertension model,
presenting with increased plasma levels of angiotensin,high BP is usually established within 3 to 4 wk afterclipping [44]. In this renovascular hypertension model, thebalance in the endothelium between NO, Ang II and ROS,
which is necessary for maintaining the homeostasis of thevascular wall, is altered. In turn, endothelial dysfunctiondue to the loss of NO bioactivity increases vascular
reactivity and initiates cardiovascular inflammatorychanges leading to elevation of BP [45]. It has beensuggested that ADMA is an endogenous regulator of
arteriolar tone through the inhibition of constitutive nitric
oxide synthase (cNOS), that may be used as a significantpredictor of renal blood flow and resistance [7, 46]. ADMAactivates renin angiotensin system and NAD(P)H oxidase
leading to increased production of O2–•, which then
interferes with the bioavailability of NO, resulting indiminished dilation and increased arteriolar tone [47].Therefore, increased ADMA is known as a potential
cardiovascular risk factor indicating reduced renal plasmaflow and increased renovascular resistance [48, 49]. Inagreement with previous observations, 2K1C procedure in
the present study resulted in elevated BP along withincreased plasma levels of ADMA and increased MDAlevels in various tissues, indicating the presence of endo-
thelial dysfunction associated with NOS inhibition as wellas increased lipid peroxidation. Similarly, in a group ofhypertensive patients, elevated levels of ADMA and
increased lipid peroxidation products along with enhancedrelease of ROS were suggested to be contributing to theassociated microvascular endothelial dysfunction and ele-vated BP [50]. The causal association of oxidative stress and
hypertension has also been supported by the observationthat antioxidant therapy ameliorates hypertension in exper-imental models [51, 52].
Treatment of 2K1C rats with the antioxidant melatoninin the current study improved cardiovascular dysfunction,reversed the ADMA levels and ameliorated the oxidative
Vehicle treatment Early melatonin treatment
Late melatonin treatment
0.00
0.25
0.50
0.75
1.00
C
**
+ +
2K1C
(A)
SO
D (
U/m
gpro
t)
0
10
20
30
C
**
++ ++
2K1C
(B)
CA
T (
U/m
gpro
t)
0
25
50
75
***
+++**
+++
C 2K1C
(C)
MD
A (
nmol
/g)
0
1
2
3
***
+++***
C 2K1C
(D)
GS
H (
µm
ol/g
)
0
10
20
30
***
+++++**
***
C 2K1C
(E)
MP
O (
U/g
)
0
1
2
3
4
***
+++*
C 2K1C
(F)
Na+
-K+A
TP
ase
(µm
ol/m
g pr
otei
n/hr
)
Fig. 3. (A) Superoxide dismutase (SOD)and (B) catalase (CAT) activities, (C)malondialdehyde and (D) glutathione(GSH) levels, (E) myeloperoxidase (MPO)and (F) Na+, K+-ATPase activities in therenal tissues of vehicle- or melatonin-treated rats with 2K1C hypertension.Melatonin treatment was started eitherimmediately after surgery (early treat-ment) or at the end of the third weekfollowing surgery (late treatment). Dataare presented as means ± S.E.M.*P < 0.05; **P < 0.01; ***P < 0.001versus vehicle-treated control group;+P < 0.05; ++P < 0.01; +++P <0.001 versus vehicle-treated 2K1C group.For each group n = 8.
Ersahin et al.
102
tissue damage. Based on the previous studies, although it is
not clear whether melatonin reduces BP by directlyinhibiting ADMA production or by increasing NO bio-availability, our results show that elevated ADMA levelsand reduced plasma NO levels in hypertensive rats were
significantly reversed by melatonin treatment. Similarly,Girouard et al. [53] have demonstrated that melatoninincreased endothelium-dependent vasodilation in mesen-
teric artery and aorta from both normotensive and hyper-tensive rats and decreased BP by improving vascular NOSpathway. The antihypertensive effect of melatonin was
more pronounced than the effect of the antioxidantN-acetylcysteine [54] and was found to be comparable withthe effect of spironolactone [55]. However, removing themajor source of endogenous melatonin by pinealectomy
caused a significant increase in BP and augmented theresponse to vasoconstrictor agents, while melatonin treat-ment lowered the BP and the increased vascular reactivity
[56]. Similarly, when spontaneously hypertensive rats weretreated with melatonin, a gradual decrease in BP, HR andplasma renin activity was observed [57].
Echocardiographic measurement on the nineth week ofthe 2K1C surgery revealed the presence of left ventriculardysfunction that accompanied high BP. Among several
factors that are involved in the pathogenesis of leftventricular hypertrophy, stimulation of the renin angioten-sin-aldosterone and sympathetic nervous systems andendothelial dysfunction have an important impact on
ventricular functions [58]. Studies in both humans [59]
and animal models [60] have demonstrated that an increasein oxidative stress might be also responsible for hyperten-sion-induced myocardial dysfunction. Accordingly, in-creased Ang II activity and the associated oxidative stress
might be responsible for LV dysfunction observed in ratswith 2K1C in the present study. On the other hand,melatonin, when given either in the early or later phases of
renovascular hypertension, decreased BP and improvedheart functions, possibly through its direct antioxidant andreceptor-mediated actions [61, 62]. Similarly, melatonin was
shown to prevent hypertrophy and oxidative damage in thehyperthyroid rat heart [63]. On the other hand, decreasedmelatonin levels were reported in patients with hyperten-sion [64], heart failure [65], ischemic heart disease [66], or
acute myocardial infarction [67], suggesting the cardiopro-tective effects of endogenous melatonin. Taken togetherwith the aforementioned studies, improved echocardio-
graphic results along with the amelioration of the oxidativeparameters in the current study implicate that the cardio-protective mechanisms accomplished by melatonin
treatment may involve both the direct antioxidant andreceptor-mediated effects of melatonin.Reactive oxygen species have been implicated in the
pathogenesis of renal injury by liberation of vasoconstric-tors, direct cellular damage, and inactivation of NO, whichmay subsequently lead to both functional and structuralrenal abnormalities [68]. On the other hand, blockade of the
Vehicle treatment Early melatonin treatment
0.00
0.25
0.50
0.75
1.00
1.25
C
***
++ +
2K1C
SO
D (
U/m
gpro
t)
0
10
20
30
40
C
**
++
++
2K1C
CA
T (
U/m
gpro
t)
0
10
20
30***
+++++
*****
C 2K1C
MD
A (
nmol
/g)
0
1
2
3
***
+++ ++***
C 2K1C
GS
H (
µm
ol/g
)
0
10
20
30
***
+++** ***
+++
C 2K1C
MP
O (
U/g
)
+
0
1
2
3
4
***
++
C 2K1C
Na+
-K+A
TP
ase
(µm
ol/m
g pr
otei
n/hr
)
(A) (B)
(C) (D)
(E) (F)
Late melatonin treatment
Fig. 4. (A) Superoxide dismutase (SOD)and (B) catalase (CAT) activities, (C)malondialdehyde and (D) glutathione(GSH) levels, (E) myeloperoxidase (MPO)and (F) Na+, K+-ATPase activities in thecerebral tissues of vehicle- or melatonin-treated rats with 2K1C hypertension.Melatonin treatment was started eitherimmediately after surgery (early treat-ment) or at the end of the third weekfollowing surgery (late treatment). Dataare presented as means ± S.E.M.*P < 0.05; **P < 0.01; ***P < 0.001versus vehicle-treated control group;+P < 0.05; ++P < 0.01; +++P <0.001 versus vehicle-treated 2K1C group.For each group n = 8.
Melatonin treatment improves renovascular hypertension
103

oxidative stress pathway in an experimental model ofrenovascular disease was shown to improve renal bloodperfusion and to prevent inflammation and fibrosis in the
stenotic kidney [69]. In the present study, MDA levels in thekidneys, as well as in the cardiac and cerebral tissues, weresignificantly increased, indicating that oxidative stressgenerated in the kidneys, which contributes to the gener-
ation and maintenance of hypertension, also causes oxida-tive damage in multiple organs remote to the kidneys.Moreover, increased activity of Na+/K+ ATP-ase, which is
a membrane-bound enzyme that requires phospholipids forits activity, also indicates lipid peroxidation-induced injuryin the studied tissues [70]. As it is observed in the present
study, oxidative stress reduces tissue levels of GSH and theactivities of superoxide dismutase and catalase, which areessential compounds for maintaining cell integrity. It isknown that deficiency or depletion of GSH and other
antioxidant enzymes causes damage to macromolecules orto membrane lipids when there is consistent formation ofoxygen free radicals. The results demonstrate that treat-
ment with the antioxidant melatonin reversed the altera-tions in tissue MDA and GSH levels, SOD, catalase andNa+/K+ ATP-ase activities. Thus, in accordance with the
previous results, the current study demonstrates that theprotective effects of melatonin in hypertension-inducedoxidative damage may be due to an augmentation of the
endogenous antioxidants and the inhibition of lipid perox-idation by maintaining a balance in oxidant-antioxidantstatus. It is well established in numerous studies thatmelatonin displays significant beneficial actions against
oxidative cellular toxicity through its capability of scav-enging both oxygen- and nitrogen-based reactants and byblocking transcriptional factors, which induce pro-inflam-
matory cytokines [71].Since MPO plays a fundamental role in oxidant
production by neutrophils and is used as an index for
the neutrophil infiltration, the increased MPO activityobserved in the current study clearly demonstrates that2K1C-induced damage in the kidneys and the two target
organs is neutrophil-dependent. Activated neutrophilsrelease MPO, causing production of large amounts ofHOCl, which oxidizes and damages macromolecules,including proteins, lipids, carbohydrates and nucleic
acids. Increasing evidence also suggests that neutrophilsrelease chemotactic substances, which further promoteneutrophil migration to the tissue, activate neutrophils
and increase the damage [72]. Previously, melatonin wasshown to reverse L-NAME-induced BP elevation andprotect against cardiac oxidative injury, while depressing
MPO activity [19]. In parallel to these results, currentdata also show that neutrophil migration plays animportant role in the development of oxidative injury inhypertensive rats. Recently, it was demonstrated that
melatonin directly modulates the formation of MPOintermediates and their decay rates, and thereby mayserve as a potent inhibitor of MPO and its downstream
inflammatory pathways [73].In an experimental renovascular hypertension model,
melatonin significantly reduced BP, improved left ventric-
ular function and alleviated oxidative injury of the kidney,heart and brain by its ability to inhibit neutrophil infiltra-
tion, to balance the oxidant-antioxidant status, and toregulate the generation of inflammatory mediators. Inconclusion, the findings of the present study support the
critical pathogenic contribution of increased oxidativestress in renovascular hypertension, and suggests a rolefor melatonin in preserving the kidney and the targetorgans against hypertension-induced tissue damage.
References
1. Textor SC, Wilcox CS. Renal artery stenosis: a common,
treatable cause of renal failure? Annu Rev Med 2001; 52:421–
442.
2. Magrini F, Reggiani P, Roberts N, Meazza R, Ciulla M,
Zanchetti A. Effects of angiotensin and angiotensin blockade
on coronary circulation and coronary reserve. Am J Med 1988;
84:55–60.
3. Kim S, Iwao H. Molecular and cellular mechanisms of
angiotensin-II mediated cardiovascular and renal diseases.
Pharmacol Rev 2000; 52:11–34.
4. Minuz P, Patrignani P, Gaino S et al. Increased oxidative
stress and platelet activation in patients with hypertension and
renovascular disease. Circulation 2002; 106:2800–2805.
5. Ushio-Fukai M, Zafari AM, Fukui T, Ishizaka N,
Griendling KK. p22phox is a critical component of the
superoxide-generating NADH/NADPH oxidase system and
regulates angiotensin II-induced hypertrophy in vascular
smooth muscle cells. J Biol Chem 1996; 271:23317–23321.
6. Zafari AM, Ushio-Fukai M, Akers M et al. Role of NADH/
NADPH oxidase-derived H2O2 in angiotensin II–induced
vascular hypertrophy. Hypertension 1998; 32:488–495.
7. Laursen JB, Rajagopalan S, Galis Z, Tarpey M, Freeman
BA, Harrison DG. Role of superoxide in angiotensin-II-in-
duced but not catecholamine-induced hypertension. Circula-
tion 1997; 95:588–593.
8. Somers MJ, Mavromatis K, Galis ZS, Harrison DG.
Vascular superoxide production and vasomotor function in
hypertension induced by deoxycorticosterone acetate-salt.
Circulation 2000; 101:1722–1728.
9. Touyz RM, Tabet F, Schiffrin EL. Redox-dependent sig-
nalling by angiotensin II and vascular remodelling in hyper-
tension. Clin Exp Pharmacol Physiol 2003; 30:860–866.
10. Diep QN, Amiri F, Touyz RM et al. PPAR alpha activator
effects on Ang II-induced vascular oxidative stress and
inflammation. Hypertension 2002; 40:866–871.
11. Rajagopalan S, Kurz S, Munzel T et al. Angiotensin
II-mediated hypertension in the rat increases vascular super-
oxide production via membrane NADH/NADPH oxidase
activation: contribution to alterations of vasomotor tone.
J Clin Invest 1996; 97:1916–1923.
12. Cohuet G, Struijker-Boudier H. Mechanisms of target
organ damage caused by hypertension: therapeutic potential.
Pharm Ther 2006; 111:81–98.
13. Lerner AB, Case JD, Takahashi Y et al. Isolation of mel-
atonin, a pineal factor that lightens melanocytes. J Am Chem
Soc 1958; 80:2587–2589.
14. Tengattini S, Reiter RJ, Tan DX, Terron MP, Rodella
LF, Rezzani R. Cardiovascular diseases: protective effects of
melatonin. J Pineal Res 2008; 44:16–25.
15. Simko F, Paulis L. Melatonin as a potential antihypertensive
treatment. J Pineal Res 2007; 42:319–322.
16. Arangino S, Cagnacci A, Angiolucci M et al. Effects of
melatonin on vascular reactivity, catecholamine levels, and
Ersahin et al.
104

blood pressure in healthy men. Am J Cardiol 1999; 83:1417–
1419.
17. Pechanova O, Zicha J, Paulis L et al. The effect of N-ace-
tylcysteine and melatonin in adult spontaneously hypertensive
rats with established hypertension. Eur J Pharmacol 2007;
561:129–136.
18. Girouard H, Denault C, Chulak C, De Champlain J.
Treatment by n-acetylcysteine and melatonin increases cardiac
baroreflex and improves antioxidant reserve. Am J Hypertens
2004; 17:947–954.
19. Sahna E, Deniz E, Bay-Karabulut A, Burma O. Melatonin
protects myocardium from ischemia-reperfusion injury in
hypertensive rats: role of myeloperoxidase activity. Clin Exp
Hypertens 2008; 30:673–681.
20. Abete P, Bianco S, Calabrese C et al. Effects of melatonin in
isolated rat papillary muscle. FEBS Lett 1997; 412:79–85.
21. Pang CS, Xi SC, Brown GM, Pang SF, Shiu SY. 2[125I]
Iodomelatonin binding and interaction with beta-adrenergic
signaling in chick heart/coronary artery physiology. J Pineal
Res 2002; 32:243–252.
22. Masana MI, Doolen S, Ersahin C et al. MT(2) melatonin
receptors are present and functional in rat caudal artery.
J Pharmacol Exp Ther 2002; 302:1295–1302.
23. PeyrotF, DucrocqC. Potential role of tryptophan derivatives
in stress responses characterized by the generation of reactive
oxygen and nitrogen species. J Pineal Res 2008; 45:235–246.
24. ReiterRJ,TanDX,PilarTerronM,FloresLJ,Czarnocki
Z. Melatonin and its metabolites: new findings regarding their
production and their radical scavenging actions. Acta Biochim
Pol 2007; 54:1–9.
25. Mayo JC, Sainz RM, Tan DX et al. Anti-inflammatory
actions of melatonin and its metabolites, N1-acetyl-N2-formyl-
5-methoxykynuramine (AFMK) and N1-acetyl-5-methoxyky-
nuramine(AMK), in macrophages. J Neuroimmunol 2005;
165:139–149.
26. Carrillo-Vico A, Guerrero JM, Lardone PJ, Reiter RJ.
A review of the multiple actions of melatonin on the immune
system. Endocr 2005; 27:189–200.
27. Navar LG, Harrison-Bernard LM, Nishiyama A, Kobori
H. Regulation of intrarenal angiotensin II in hypertension.
Hypertension 2002; 39:316–322.
28. Goldblatt H, Lynch J, Hanzal RF, Summerville MD.
Studies on Experimental Hypertension I. The production of
persistent elevation of the systolic blood pressure by means of
renal ischemia. J Exper Med 1934; 59:347–379.
29. Schiller NB, Shah PM, Crawford M et al. Recommenda-
tions for quantitation of the left ventricle by two-dimensional
echocardiography. American Society of Echocardiography
Committee on Standards, Subcommittee on Quantitation of
Two-Dimensional Echocardiograms. J Am Soc Echocardiogr
1989; 2:358–367.
30. Mylorie AA, Collins H, Umbles C, Kyle J. Erythrocyte
superoxide dismutase activity and other parameters of cupper
status in rats ingesting lead acetate. Toxicol Appl Pharmacol
1986; 82:512–520.
31. Aebi H. Catalase in vitro. In: Methods of Enzymatic Analysis,
2nd edn, Vol. 2. Bergmeyer HU (ed), FL, 1974; pp. 121–
126.
32. Beuge JA, Aust SD. Microsomal lipid peroxidation. Meth
Enzymol 1978; 52:302–311.
33. Beutler E. Glutathione in red blood cell metabolism. A
Manuel of Biochemical Methods. Grune & Stratton, New
York, 1975, pp. 112–114.
34. Hillegass LM, Griswold DE, Brickson B, Albrightson-
Winslow C. Assessment of myeloperoxidase activity in whole
rat kidney. J Pharmacol Meth 1990; 24:285–295.
35. Kim YK, Lee SH, Goldinger JM, Hong SK. Effect of eth-
anol on organic ion transport in rabbit kidney. Toxicol Appl
Pharmacol 1986; 86:411–420.
36. Reading HW, Isbir T. The role of cation activated ATPase in
transmitter release from the rat iris. Q J Exp Physiol 1980;
65:105–116.
37. Fiske CH, Subbarow Y. The colorimetric determination of
phosphorus. J Biol Chem 1925; 66:375–400.
38. Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ.
Protein measurements with the folin phenol reagent. J Biol
Chem 1951; 193:265–275.
39. Russo C, Olivieri O, Girelli D et al. Anti-oxidant status and
lipid peroxidation in patients with essential hypertension.
J Hypertens 1998; 16:1267–1271.
40. Schnackenberg CG, Welch WJ, Wilcox CS. Normalization
of blood pressure and renal vascular resistance in SHR with a
membranepermeable superoxide dismutase mimetic: role of
nitric oxide. Hypertension 1998; 32:59–64.
41. Swei A, Lacy F, Delano FA, Parks DA, Schmid-Schonbein
GW. A mechanism of oxygen free radical production in the
Dahl hypertensive rat. Microcirculation 1999; 6:179–187.
42. Vaziri ND, Wang XQ, Oveisi F, Rad B. Induction of oxi-
dative stress by glutathione depletion causes severe hyperten-
sion in normal rats. Hypertension 2000; 36:142–146.
43. Cosentino F, Sill JC, Katusic ZS. Role of superoxide anions
in the mediation of endothelium-dependent contractions.
Hypertension 1994; 23:229–235.
44. Martinez ML, Castro MM, Rizzi E et al. Lercanidipine
reduces matrix metalloproteinase-2 activity and reverses vas-
cular dysfunction in renovascular hypertensive rats. Eur J
Pharmacol 2008; 591:224–230.
45. Reckelhoff JF, Romero JC. Role of oxidative stress in
angiotensin-induced hypertension. Am J Physiol Regul Integr
Comp Physiol 2003; 284:R893–912.
46. Kielstein JT, Bode-Boger SM, Frolich JC, Ritz E, Hal-
ler H, Fliser D. Asymmetric dimethylarginine, blood pres-
sure, and renal perfusion in elderly subjects. Circulation 2003;
107:1891–1895.
47. Veresh Z, Racz A, Lotz G, Koller A. ADMA impairs nitric
oxide-mediated arteriolar function due to increased superoxide
production by angiotensin II-NAD(P)H oxidase pathway.
Hypertension 2008; 52:960–966.
48. Miyazaki H, Matsuoka H, Cooke JP et al. Endogenous
nitric oxide synthase inhibitor: a novel marker of atheroscle-
rosis. Circulation 1999; 99:1141–1146.
49. Jacobi J, Maas R, Cordasic N et al. Role of asymmetric
dimethylarginine for angiotensin II-induced target organ
damage in mice. Am J Physiol Heart Circ Physiol 2008;
294:H1058–1066.
50. Wang D, Strandgaard S, Iversen J, Wilcox CS. Asym-
metric dimethylarginine, oxidative stress, and vascular nitric
oxide synthase in essential hypertension. Am J Physiol Regul
Integr Comp Physiol 2009; 296:R195–200.
51. Schnackenberg CG, Wilcox CS. Two-week administration
of tempol attenuates both hypertension and renal excretion of
8-Iso prostaglandin F2 alpha. Hypertension 1999; 33:424–
428.
52. Atarashi K, Ishiyama A, Takagi M et al. Vitamin E
ameliorates the renal injury of Dahl salt-sensitive rats. Am J
Hypertens 1997; 10:116S–119S.
Melatonin treatment improves renovascular hypertension
105

53. Girouard H, Chulak C, Lejossec M, Lamontagne D,
De Champlain J. Vasorelaxant effects of the chronic treat-
ment with melatonin on mesenteric artery and aorta of
spontaneously hypertensive rats. J Hypertens 2001; 19:1369–
1377.
54. Kojsova S, Jendekova L, Zicha J, Kunes J, Andriantsi-
tohaina R, Pechanova O. The effect of different antioxidants
on nitric oxide production in hypertensive rats. Physiol Res
2006; 55(Suppl 1):S3–16.
55. Paulis L, Matuskova J, Adamcova M et al. Regression of
left ventricular hypertrophy and aortic remodelling in
NO-deficient hypertensive rats: effect of L-arginine and spir-
onolactone. Acta Physiol (Oxf) 2008; 194:45–55.
56. Cunnane SC, Manku MS, Oka M, Horrobin DF. Enhanced
vascular reactivity to various vasoconstrictor agents following
pinealectomy in the rat: role of melatonin. Can J Physiol
Pharmacol 1980; 58:287–293.
57. Kawashima K, Miwa Y, Fujimoto K, Oohata H, Nishino H,
Koike H. Antihypertensive action of melatonin in the sponta-
neously hypertensive rat. Clin Exp Hypertens A 1987; 9:1121–
1131.
58. Gradman AH, Alfayoumi F. From left ventricular hyper-
trophy to congestive heart failure: management of hypertensive
heart disease. Prog Cardiovasc Dis 2006; 48:326–341.
59. Higashi Y, Sasaki S, Nakagawa K, MatsuuraH, Oshima T,
Chayama K. Endothelial function and oxidative stress in
renovascular hypertension. N Engl J Med 2002; 346:1954–
1962.
60. De Nigris F, Lerman LO, Condorelli M, Lerman A,
Napoli C. Oxidationsensitive transcription factors and
molecular mechanisms in the arterial wall. Antioxid Redox
Signal 2001; 3:1119–1130.
61. Dubocovich M, Markowska M. Functional MT1 and MT2
melatonin receptors in mammals. Endocr 2005; 27:101–110.
62. TanDX,ManchesterLC,TerronMP,FloresLJ,ReiterRJ.
One molecule, many derivatives: a never-ending interaction of
melatoninwithreactiveoxygenandnitrogenspecies?JPinealRes
2007; 42:28–42.
63. Ghosh G, De K, Maity S et al. Melatonin protects against
oxidative damage and restores expression of GLUT4 gene in
the hyperthyroid rat heart. J Pineal Res 2007; 42:71–82.
64. JonasM, Garfinkel D, ZisapelN, Laudon M,GrossmanE.
Impaired nocturnal melatonin secretion in non-dipper hyper-
tensive patients. Blood Press 2003; 12:19–24.
65. Girotti L, Lago M, Ianovsky O et al. Low urinary 6-sul-
fatoxymelatonin levels in patients with severe congestive heart
failure. Endocr 2003; 22:245–248.
66. Brugger P, Marktl W, Herold M. Impaired nocturnal
secretion of melatonin in coronary heart disease. Lancet 1995;
345:1408.
67. DomInguez-RodrIguez A, Abreu-Gonzalez P, GarcIa MJ,
Sanchez J, Marrero F, De Armas-Trujillo D. Decreased
nocturnal melatonin levels during acute myocardial infarction.
J Pineal Res 2002; 33:248–252.
68. Lerman L, Textor SC. Pathophysiology of ischemic
nephropathy. Urol Clin North Am 2001; 28:793–803.
69. Chade AR, Rodriguez-Porcel M, Herrmann J et al.
Antioxidant Intervention Blunts Renal Injury in Experimental
Renovascular Disease. J Am Soc Nephrol 2004; 15:958–966.
70. Jain SK, Lim G. Lipoic acid decreases lipid peroxidation and
protein glycosylation and increases (Na(+) + K(+))- and
Ca(++)-ATPase activities in high glucose-treated human ery-
throcytes. Free Radic Biol Med 2000; 29:1122–1128.
71. Korkmaz A, Reiter RJ, Topal T, Manchester LC, Oter S,
Tan DX. Melatonin: an established antioxidant worthy of use
in clinical trials. Mol Med 2009; 15:43–50.
72. Kettle AJ, Winterbourn CC. Myeloperoxidase: a key reg-
ulator of neutrophil oxidant production. Redox Rep 1997;
3:3–15.
73. Galijasevic S, Abdulhamid I, Abu-Soud HM. Melatonin is
a potent inhibitor for myeloperoxidase. Biochemistry 2008;
47:2668–2677.
Ersahin et al.
106





























