Embed Size (px)
Citation preview

0014-2980/98/1111-3605$17.50+.50/0© WILEY-VCH Verlag GmbH, D-69451 Weinheim, 1998
MAdCAM-1 costimulates T cell proliferationexclusively through integrin > 4 I 7, whereas VCAM-1and CS-1 peptide use > 4 I 1: evidence for “remote”costimulation and induction ofhyperresponsiveness to B7 molecules
Klaus Lehnert, Cristin G. Print, Yi Yang and Geoffrey W. Krissansen
Department of Molecular Medicine, School of Medicine and Health Sciences, University ofAuckland, Auckland, New Zealand
We have analyzed the effects of the § 4 integrin ligands mucosal addressin cell adhesionmolecule-1 (MAdCAM-1), vascular cell adhesion molecule-1 (VCAM-1), and the fibronectinCS-1 splice variant on T cell activation. Immobilized MAdCAM-1 and VCAM-1 IgG-Fc chi-meras and a fibronectin CS-1 peptide efficiently costimulate T cell proliferation when antigenpresentation is mimicked by anti-CD3 antibody. VCAM-1-Fc and fibronectin CS-1, which areadhesive ligands for both the § 4 g 1 and § 4 g 7 integrins, medicate T cell costimulation exclu-sively through integrin § 4 g 1, but not through § 4 g 7. The inability of VCAM-1-Fc to costimulatevia § 4 g 7 suggests that cell adhesion per se is insufficient, and that exquisite recognition andactivation events must be triggered. MAdCAM-1-Fc mediates costimulation exclusively via§4g 7, and can both synergize with and induce hyperresponsiveness to the classical costimu-
lator B7-2. MAdCAM-1-Fc and VCAM-1-Fc, but not B7-2, effectively costimulate whenimmobilized on sites spatially distant from the anti-CD3 antibody (“remote” costimulation). Invitro, the relative potencies of the CAM were VCAM-1-Fc G ICAM-1-Fc G MAdCAM-1-Fc G B7-Fc, except at high concentrations where ICAM-1 was the most potent. Features ofcostimulatory CAM revealed by this study have important implications for the design ofimmunotherapeutic vaccine strategies to combat cancer and infection.
Key words: MAdCAM-1 / Costimulation / Integrin / T cell / B7 molecule
Received 2/2/98Revised 7/8/98Accepted 11/8/98
[I 18042]
Abbreviations: MAdCAM-1: Mucosal addressin celladhesion molecule-1 VCAM-1: Vascular cell adhesionmolecule-1 ICAM-1: Intercellular cell adhesion molecule-1EC: Endothelial cell CHO: Chinese hamster ovary
1 Introduction
The antigen-specific activation of CD4+ T lymphocytes isinitiated with the presentation by APC of MHC class II-bound antigenic peptides to the CD3/TCR complex.Engagement of accessory T cell surface molecules bytheir counterreceptors on APC provides a separate,MHC- and antigen-independent “costimulatory” signalthat is required for optimal T cell activation. Both signalsare integrated to induce the transcription of genes forcytokines and their receptors and ultimately lead to cellproliferation and differentiation [1, 2].
Antigen presentation to T cells in the absence of costim-ulation can result in T cell anergy and clonal deletion[2–4]. Efficient T cell activation requires direct physicalcontacts between T cells and APC [5]. Consequently,many costimulatory molecules have been shown to beinvolved in cell adhesion [6]. The integrin superfamily isthe largest group of cell adhesion molecules, comprisingat least 16 § subunits that noncovalently associate withone or more of 8 g subunits to form heterodimeric type-Imembrane glycoproteins. The recognition by integrins ofcellular and extracellular ligands is crucial for interac-tions between cells and between cells and the extracel-lular matrix [7–9].
In addition to strengthening the adhesion between Tcells and APC, binding of integrins to their ligands itselfcan deliver costimulatory signals. Engagement of theintegrin § L g 2 by its major ligand ICAM-1 induces the pro-liferation of antigen-stimulated T lymphocytes in vitro[10]. In simplified experimental systems using antibodies
Eur. J. Immunol. 1998. 28: 3605–3615 T cell costimulation by § 4-integrin ligands and B7-molecules 3605

against the TCR/CD3 complex to provide an antigen-independent first signal, binding of integrin g 1 familymembers to extracellular matrix proteins has also beenshown to costimulate T cell proliferation [11, 12].
The integrin § 4 subunit can associate with the g 7 subunitor the g 1 subunit to form two integrin heterodimers thatexhibit cell type-dependent and overlapping adhesiveproperties. The §
4g 7 integrin [13], restricted to leuko-
cytes, is the major homing receptor for mucosal addres-sin cell adhesion molecule-1 (MAdCAM-1), a cell surfaceglycoprotein normally expressed almost exclusively onhigh endothelial venules of gut-associated lymphoid tis-sues [14–17]. MAdCAM-1 expression by other endothe-lial cells has only been demonstrated in nonobese dia-betic mice, in chronic relapsing experimental autoim-mune encephalomyelitis and in inflammatory bowel dis-ease [18–20]. § 4 g 7 also binds to vascular cell adhesionmolecule-1 (VCAM-1), expressed on activated vascularendothelium and on lymphoid dendritic cells [15, 21].The alternatively spliced domain CS-1 within the type IIIconnecting segment of the extracellular matrix proteinfibronectin is a third ligand for the § 4 g 7 integrin [21, 22].VCAM-1 and fibronectin CS-1 are also ligands for the§4g 1 integrin, expressed by myeloid and lymphoid cells
[23, 24]. Only activated peripheral blood monocytes, butnot T lymphocytes have been shown to use VLA-4 foradhesion to MAdCAM-1 [19, 20, 25].
While the costimulatory effects of VLA-4 integrinengagement are well documented [11, 12, 26, 27], T cellcostimulation through § 4 g 7 integrin by its physiologicalligands has not been investigated. Here, we use pep-tides and recombinant soluble IgG chimeras containingthe ligand-binding domains of MAdCAM-1, VCAM-1 andfibronectin to characterize the costimulatory propertiesof § 4 integrins, and to identify the integrin through whichthey trigger T cell activation. In addition, we investigatewhether § 4 integrin ligands influence T cell activation bythe classical costimulatory molecules B7-1 and B7-2.
2 Results
2.1 >4 integrin ligands costimulate T cell
proliferation and IL-2 secretion
Recombinant § 4 integrin ligands MAdCAM-1-Fc, VCAM-1-Fc and CS-1 peptide were co-immobilized in microtiterplates with a submitogenic dose of the anti-CD3 mAbOKT3, which delivers an antigen-independent primarysignal through the CD3/TCR complex. Fig. 1 shows theproliferative response of purified human CD4+ T cells tothe three § 4 integrin ligands. Both MAdCAM-1-Fc andVCAM-1-Fc efficiently costimulated the proliferation of
human T cells (Fig. 1A). While VCAM-1-Fc induced maxi-mal stimulation at only 50 ng/well, approximately 750 ngof MAdCAM-1-Fc was required to achieve a similar levelof cell proliferation. No thymidine incorporation wasobserved when cells were costimulated with OKT3 plusIgG1, or when the chimeric cell adhesion molecules wereimmobilized without OKT3. The CS-1 peptide alsocostimulated proliferation of CD4+ T cells, although notas efficiently as MAdCAM-1-Fc or VCAM-1-Fc. Noresponse to the peptide was observed without OKT3, orwhen the scrambled control peptide was co-immobilizedwith OKT3 (Fig. 1B).
As shown in Fig. 2, 7-12-fold increased steady-state lev-els of IL-2 were detected in culture supernatants aftercostimulation with MAdCAM-1-Fc or VCAM-1-Fc.Costimulation by MAdCAM-1-Fc resulted in 1.5–2.5-foldhigher IL-2 levels than stimulation by VCAM-1-Fc plusOKT3. The lower IL-2 levels in VCAM-1-Fc-costimulatedcultures are likely the result of increased IL-2 consump-tion, as T cells responded to VCAM-1-Fc costimulationwith stronger cell proliferation than to MAdCAM-1-Fccostimulation (Fig. 1A).
2.2 Costimulation by > 4 integrin ligands isreceptor specific
Adhesion-blocking mAb were employed to identifywhich of the two § 4 integrins mediates T cell costimula-tion by MAdCAM-1-Fc, VCAM-1-Fc and fibronectinCS-1. Fig. 3A shows that antibodies against the integring 7 subunit or the integrin § 4 subunit, as well as mAb Act-
1, which recognizes a combinatorial epitope on the § 4 g 7heterodimer, could inhibit costimulation by MAdCAM-1-Fc. In contrast, an antibody against the integrin g 1 sub-unit or control antibodies had no effect. Conversely, onlymAb 4B4, directed against the integrin g 1 subunit, andthe anti- § 4 integrin subunit mAb PS/2, but none of theanti- g 7 antibodies, were able to block VCAM-1-Fc-mediated costimulation (Fig. 3B). Likewise, T cell prolifer-ation costimulated by immobilized CS-1-peptide wasinhibited only by antibodies against the § 4 and g 1 integrinsubunits, whereas anti- g 7 integrin subunit antibodies andcontrol antibodies had no effect (Fig. 3C). Interestingly,preincubation of CD4+ T lymphocytes with solubleVCAM-1-Fc could partially inhibit costimulation byMAdCAM-1-Fc, while preincubation with solubleMAdCAM-1-Fc had no effect on costimulation byVCAM-1-Fc (Fig. 3A,B). Blockade of immobilizedMAdCAM-1-Fc or VCAM-1-Fc resulted in the completeinhibition of T cell proliferation (Fig. 3A, B), demonstrat-ing that the T cell costimulation observed in these exper-iments was specifically mediated by the recombinant celladhesion molecules.
3606 K. Lehnert et al. Eur. J. Immunol. 1998. 28: 3605–3615
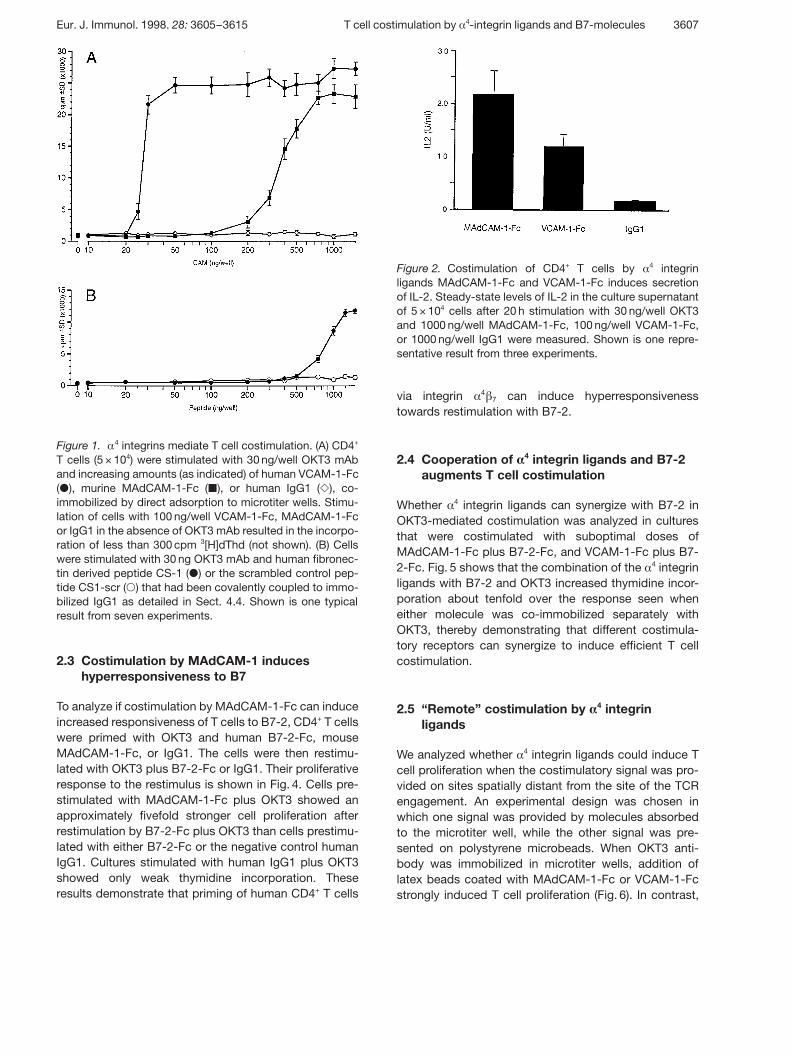
Figure 1. § 4 integrins mediate T cell costimulation. (A) CD4+
T cells (5 × 104) were stimulated with 30 ng/well OKT3 mAband increasing amounts (as indicated) of human VCAM-1-Fc( Æ ), murine MAdCAM-1-Fc ( | ), or human IgG1 ( Y ), co-immobilized by direct adsorption to microtiter wells. Stimu-lation of cells with 100 ng/well VCAM-1-Fc, MAdCAM-1-Fcor IgG1 in the absence of OKT3 mAb resulted in the incorpo-ration of less than 300 cpm 3[H]dThd (not shown). (B) Cellswere stimulated with 30 ng OKT3 mAb and human fibronec-tin derived peptide CS-1 ( Æ ) or the scrambled control pep-tide CS1-scr ( 1 ) that had been covalently coupled to immo-bilized IgG1 as detailed in Sect. 4.4. Shown is one typicalresult from seven experiments.
Figure 2. Costimulation of CD4+ T cells by §4 integrin
ligands MAdCAM-1-Fc and VCAM-1-Fc induces secretionof IL-2. Steady-state levels of IL-2 in the culture supernatantof 5 × 104 cells after 20 h stimulation with 30 ng/well OKT3and 1000 ng/well MAdCAM-1-Fc, 100 ng/well VCAM-1-Fc,or 1000 ng/well IgG1 were measured. Shown is one repre-sentative result from three experiments.
2.3 Costimulation by MAdCAM-1 induceshyperresponsiveness to B7
To analyze if costimulation by MAdCAM-1-Fc can induceincreased responsiveness of T cells to B7-2, CD4+ T cellswere primed with OKT3 and human B7-2-Fc, mouseMAdCAM-1-Fc, or IgG1. The cells were then restimu-lated with OKT3 plus B7-2-Fc or IgG1. Their proliferativeresponse to the restimulus is shown in Fig. 4. Cells pre-stimulated with MAdCAM-1-Fc plus OKT3 showed anapproximately fivefold stronger cell proliferation afterrestimulation by B7-2-Fc plus OKT3 than cells prestimu-lated with either B7-2-Fc or the negative control humanIgG1. Cultures stimulated with human IgG1 plus OKT3showed only weak thymidine incorporation. Theseresults demonstrate that priming of human CD4+ T cells
via integrin §4g 7 can induce hyperresponsiveness
towards restimulation with B7-2.
2.4 Cooperation of > 4 integrin ligands and B7-2augments T cell costimulation
Whether § 4 integrin ligands can synergize with B7-2 inOKT3-mediated costimulation was analyzed in culturesthat were costimulated with suboptimal doses ofMAdCAM-1-Fc plus B7-2-Fc, and VCAM-1-Fc plus B7-2-Fc. Fig. 5 shows that the combination of the § 4 integrinligands with B7-2 and OKT3 increased thymidine incor-poration about tenfold over the response seen wheneither molecule was co-immobilized separately withOKT3, thereby demonstrating that different costimula-tory receptors can synergize to induce efficient T cellcostimulation.
2.5 “Remote” costimulation by > 4 integrinligands
We analyzed whether § 4 integrin ligands could induce Tcell proliferation when the costimulatory signal was pro-vided on sites spatially distant from the site of the TCRengagement. An experimental design was chosen inwhich one signal was provided by molecules absorbedto the microtiter well, while the other signal was pre-sented on polystyrene microbeads. When OKT3 anti-body was immobilized in microtiter wells, addition oflatex beads coated with MAdCAM-1-Fc or VCAM-1-Fcstrongly induced T cell proliferation (Fig. 6). In contrast,
Eur. J. Immunol. 1998. 28: 3605–3615 T cell costimulation by § 4-integrin ligands and B7-molecules 3607
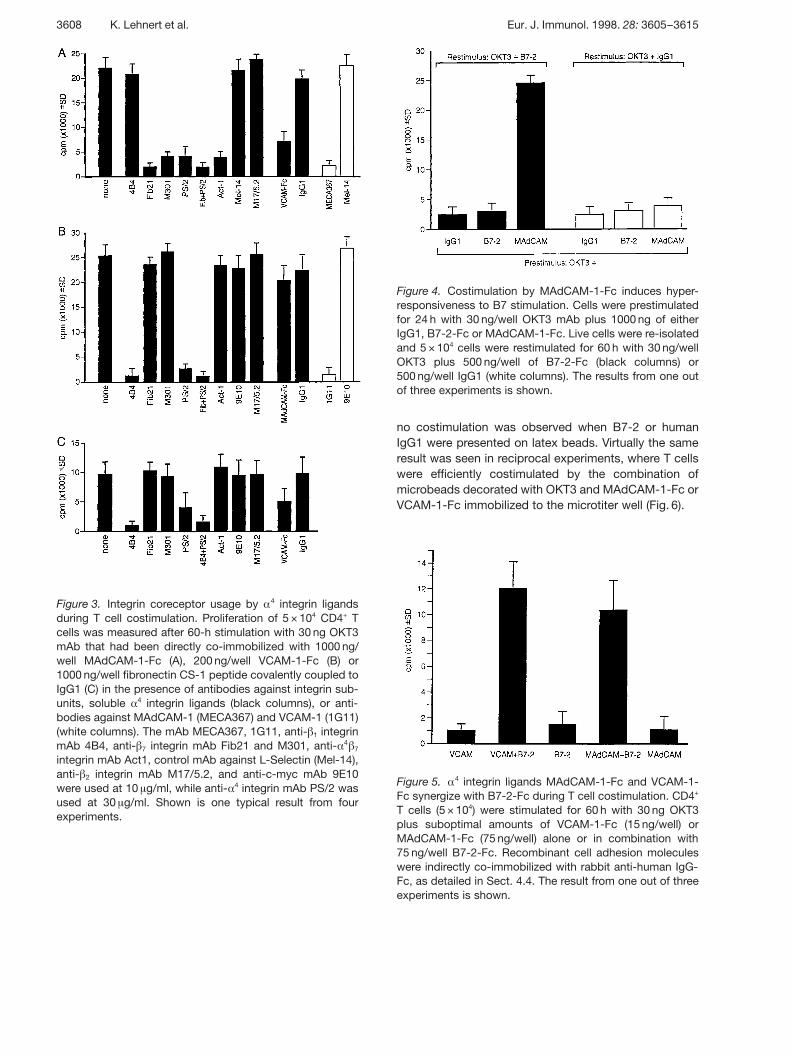
Figure 3. Integrin coreceptor usage by § 4 integrin ligandsduring T cell costimulation. Proliferation of 5 × 104 CD4+ Tcells was measured after 60-h stimulation with 30 ng OKT3mAb that had been directly co-immobilized with 1000 ng/well MAdCAM-1-Fc (A), 200 ng/well VCAM-1-Fc (B) or1000 ng/well fibronectin CS-1 peptide covalently coupled toIgG1 (C) in the presence of antibodies against integrin sub-units, soluble § 4 integrin ligands (black columns), or anti-bodies against MAdCAM-1 (MECA367) and VCAM-1 (1G11)(white columns). The mAb MECA367, 1G11, anti- g 1 integrinmAb 4B4, anti- g 7 integrin mAb Fib21 and M301, anti- § 4 g 7integrin mAb Act1, control mAb against L-Selectin (Mel-14),anti- g 2 integrin mAb M17/5.2, and anti-c-myc mAb 9E10were used at 10 ? g/ml, while anti- § 4 integrin mAb PS/2 wasused at 30 ? g/ml. Shown is one typical result from fourexperiments.
Figure 4. Costimulation by MAdCAM-1-Fc induces hyper-responsiveness to B7 stimulation. Cells were prestimulatedfor 24 h with 30 ng/well OKT3 mAb plus 1000 ng of eitherIgG1, B7-2-Fc or MAdCAM-1-Fc. Live cells were re-isolatedand 5 × 104 cells were restimulated for 60 h with 30 ng/wellOKT3 plus 500 ng/well of B7-2-Fc (black columns) or500 ng/well IgG1 (white columns). The results from one outof three experiments is shown.
Figure 5. § 4 integrin ligands MAdCAM-1-Fc and VCAM-1-Fc synergize with B7-2-Fc during T cell costimulation. CD4+
T cells (5 × 104) were stimulated for 60 h with 30 ng OKT3plus suboptimal amounts of VCAM-1-Fc (15 ng/well) orMAdCAM-1-Fc (75 ng/well) alone or in combination with75 ng/well B7-2-Fc. Recombinant cell adhesion moleculeswere indirectly co-immobilized with rabbit anti-human IgG-Fc, as detailed in Sect. 4.4. The result from one out of threeexperiments is shown.
no costimulation was observed when B7-2 or humanIgG1 were presented on latex beads. Virtually the sameresult was seen in reciprocal experiments, where T cellswere efficiently costimulated by the combination ofmicrobeads decorated with OKT3 and MAdCAM-1-Fc orVCAM-1-Fc immobilized to the microtiter well (Fig. 6).
3608 K. Lehnert et al. Eur. J. Immunol. 1998. 28: 3605–3615
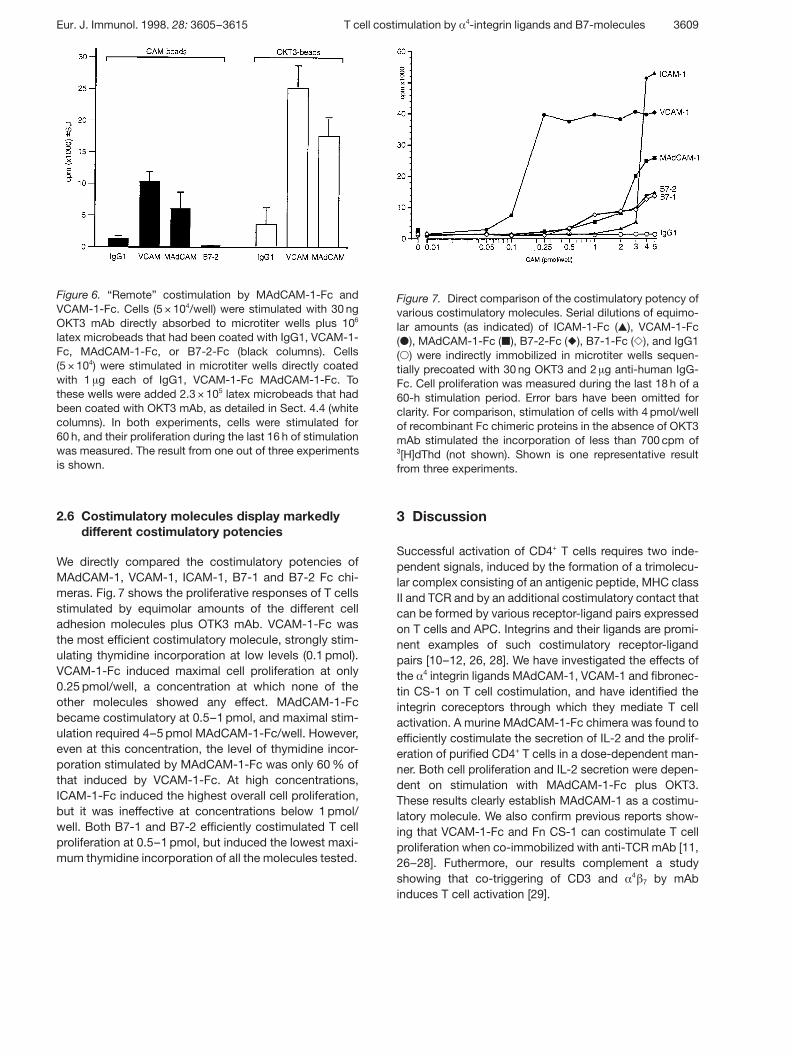
Figure 6. “Remote” costimulation by MAdCAM-1-Fc andVCAM-1-Fc. Cells (5 × 104/well) were stimulated with 30 ngOKT3 mAb directly absorbed to microtiter wells plus 106
latex microbeads that had been coated with IgG1, VCAM-1-Fc, MAdCAM-1-Fc, or B7-2-Fc (black columns). Cells(5 × 104) were stimulated in microtiter wells directly coatedwith 1 ? g each of IgG1, VCAM-1-Fc MAdCAM-1-Fc. Tothese wells were added 2.3 × 105 latex microbeads that hadbeen coated with OKT3 mAb, as detailed in Sect. 4.4 (whitecolumns). In both experiments, cells were stimulated for60 h, and their proliferation during the last 16 h of stimulationwas measured. The result from one out of three experimentsis shown.
Figure 7. Direct comparison of the costimulatory potency ofvarious costimulatory molecules. Serial dilutions of equimo-lar amounts (as indicated) of ICAM-1-Fc ( , ), VCAM-1-Fc( Æ ), MAdCAM-1-Fc ( | ), B7-2-Fc ( ! ), B7-1-Fc ( Y ), and IgG1( 1 ) were indirectly immobilized in microtiter wells sequen-tially precoated with 30 ng OKT3 and 2 ? g anti-human IgG-Fc. Cell proliferation was measured during the last 18 h of a60-h stimulation period. Error bars have been omitted forclarity. For comparison, stimulation of cells with 4 pmol/wellof recombinant Fc chimeric proteins in the absence of OKT3mAb stimulated the incorporation of less than 700 cpm of3[H]dThd (not shown). Shown is one representative resultfrom three experiments.
2.6 Costimulatory molecules display markedlydifferent costimulatory potencies
We directly compared the costimulatory potencies ofMAdCAM-1, VCAM-1, ICAM-1, B7-1 and B7-2 Fc chi-meras. Fig. 7 shows the proliferative responses of T cellsstimulated by equimolar amounts of the different celladhesion molecules plus OTK3 mAb. VCAM-1-Fc wasthe most efficient costimulatory molecule, strongly stim-ulating thymidine incorporation at low levels (0.1 pmol).VCAM-1-Fc induced maximal cell proliferation at only0.25 pmol/well, a concentration at which none of theother molecules showed any effect. MAdCAM-1-Fcbecame costimulatory at 0.5–1 pmol, and maximal stim-ulation required 4–5 pmol MAdCAM-1-Fc/well. However,even at this concentration, the level of thymidine incor-poration stimulated by MAdCAM-1-Fc was only 60 % ofthat induced by VCAM-1-Fc. At high concentrations,ICAM-1-Fc induced the highest overall cell proliferation,but it was ineffective at concentrations below 1 pmol/well. Both B7-1 and B7-2 efficiently costimulated T cellproliferation at 0.5–1 pmol, but induced the lowest maxi-mum thymidine incorporation of all the molecules tested.
3 Discussion
Successful activation of CD4+ T cells requires two inde-pendent signals, induced by the formation of a trimolecu-lar complex consisting of an antigenic peptide, MHC classII and TCR and by an additional costimulatory contact thatcan be formed by various receptor-ligand pairs expressedon T cells and APC. Integrins and their ligands are promi-nent examples of such costimulatory receptor-ligandpairs [10–12, 26, 28]. We have investigated the effects ofthe § 4 integrin ligands MAdCAM-1, VCAM-1 and fibronec-tin CS-1 on T cell costimulation, and have identified theintegrin coreceptors through which they mediate T cellactivation. A murine MAdCAM-1-Fc chimera was found toefficiently costimulate the secretion of IL-2 and the prolif-eration of purified CD4+ T cells in a dose-dependent man-ner. Both cell proliferation and IL-2 secretion were depen-dent on stimulation with MAdCAM-1-Fc plus OKT3.These results clearly establish MAdCAM-1 as a costimu-latory molecule. We also confirm previous reports show-ing that VCAM-1-Fc and Fn CS-1 can costimulate T cellproliferation when co-immobilized with anti-TCR mAb [11,26–28]. Futhermore, our results complement a studyshowing that co-triggering of CD3 and §
4g 7 by mAb
induces T cell activation [29].
Eur. J. Immunol. 1998. 28: 3605–3615 T cell costimulation by § 4-integrin ligands and B7-molecules 3609

The results of our study demonstrate that T cell costimu-lation by MAdCAM-1-Fc is exclusively mediated byintegrin § 4 g 7, as an antibody against the g 1 subunit failedto inhibit MAdCAM-1-Fc induced costimulation,whereas antibodies against the g 7 and the § 4 subunits,as well as the mAb against the §
4g 7 heterodimer,
completely inhibited T cell costimulation.
Since VCAM-1 and fibronectin CS-1 are adhesiveligands for both §
4g 1 and §
4g 7 [22–24], we sought to
identify the integrin receptor through which they mediatecostimulation. TCR-dependent costimulation byVCAM-1-Fc or CS-1 was inhibited by antibodies againstthe g 1 or § 4 integrin subunits, but not by mAb against theg 7 subunit or the §
4g 7 heterodimer, suggesting that
VCAM-1-Fc and CS-1 mediate costimulation exclusivelythrough integrin §
4g 1. We also show that soluble
VCAM-1-Fc can spatially inhibit costimulation byMAdCAM-1-Fc. This inhibition is most likely caused bybinding of VCAM-1-Fc to § 4 g 7, thereby blocking its inter-action with MAdCAM-1-Fc, as the binding sites forVCAM-1 and MAdCAM-1 on § 4 g 7 overlap [15]. SolubleMAdCAM-1-Fc did not affect costimulation by VCAM-1-Fc, which is in agreement with the finding that VCAM-1stimulates exclusively through integrin §
4g 1. We con-
clude from these results that CD4+ T lymphocytes canbind to VCAM-1-Fc via integrin § 4 g 7, but this interactiondoes not result in successful T cell costimulation. To ourknowledge, this represents the first case in which thesuccessful interaction of a pair of intrinsically costimula-tory molecules fails to trigger T cell proliferation. Evi-dence was presented that § 4 g 1 and §
4g 7 integrins can
costimulate T cells when engaged at contact sites dis-tinct from the site of CD3/TCR engagement. This“remote” costimulation was as effective as local costim-ulation, where both signals are provided on the samesurface.
At present, the physiological relevance of costimulationby MAdCAM-1 and VCAM-1 is unclear. While VCAM-1has been reported to be expressed by dendritic cells [30]and could therefore be involved in the activation of Tlymphocytes by dendritic cells, MAdCAM-1 is not nor-mally expressed by professional APC. Both MAdCAM-1and VCAM-1 are expressed by activated EC, which canacquire MHC class II and productively present Ag to Tcells [31]. It is possible that both cell adhesion moleculesparticipate in T cell activation by EC. This “local” costim-ulation may be particularly relevant at sites of T cellextravasation, where potential antigen-presenting ECand antigen-specific T cells interact. Alternatively, extra-vasating T cells may be “remotely” costimulated in ascenario where the EC provides a costimulatory integrinligand, and a professional “bystander” APC presents thespecific antigen.
Various reports have established that resting unactivatedT cells cannot be efficiently costimulated by anti-TCRand B7 [28, 32–34]. However, these cells respondstrongly to costimulation by anti-TCR and VCAM-1 orICAM-1 with cell proliferation, increased expression ofB7-ligands CD28 and CTLA-4, and cell adhesion to B7[10, 26, 32, 33]. We show that priming of CD4+ T cellswith MAdCAM-1-Fc and anti-TCR mAb augments theproliferative response of T cells upon restimulation withB7 and OKT3. It is therefore likely that priming byMAdCAM-1-Fc plus OKT3 also induces up-regulation ofCD28 and/or CTLA-4, which may allow the primed Tcells to bind more efficiently to B7, ultimately resulting inthe increased proliferative response observed afterrestimulation with B7-2-Fc and OKT3. In light of recentstudies showing that engagement of CTLA-4 inducespredominantly negative signals that block cell cycle pro-gression of activated T cells [35, 36], the increased T cellproliferation after restimulation with B7-2-Fc and OKT3shown here may seem unexpected. However, it has alsobeen shown that CTLA-4 costimulation by mAb can syn-ergize with suboptimal CD28 costimulation [33]. Theeffects of CTLA-4 engagement on T cell activation arepoorly understood. In the most advanced model, thenegative regulatory effects of CTLA-4 on T cell costimu-lation are explained by competition between CTLA-4 andCD28 for B7-molecules [37]. This competition, andtherefore the negative effects of CTLA-4, are likely to beovercome when sufficient amounts of B7-molecules areavailable. It is likely that the experimental conditions inthis study represent such a situation.
Our results demonstrate that both MAdCAM-1-Fc andVCAM-1-Fc can synergize with B7-2-Fc to induce maxi-mal CD3-dependent T cell costimulation. Since the com-bination of § 4 integrin ligands with B7 induced had a syn-ergistic effect, it cannot be attributed to the additiveresponses of two separate cell subsets. Alternatively, thesimultaneous engagement of integrins and CD28/CTLA-4 could trigger separate or partially overlapping signaltransduction pathways. It is also possible that the syner-gism between costimulatory molecules is an indirectresult of increased cell adhesion, which may promotemore efficient TCR-ligand interaction and increased sig-naling through the TCR complex. Several observationsargue against this possibility. First, an elegant study byUdagawa et al. [38] has shown that VCAM-1-Fc medi-ated costimulation through § 4 g 1 does not merely aug-ment signaling through the CD3/TCR complex but pro-vides an independent second signal. Second, ICAM-1has been shown to costimulate T cell proliferation evenwhen a TCR-dependent signal was induced in anadhesion-independent manner [10]. Third, it has beenshown that immobilized OKT3 alone mediates almostcomplete T cell adhesion [39]. Finally, our results show-
3610 K. Lehnert et al. Eur. J. Immunol. 1998. 28: 3605–3615

ing that § 4 g 7 integrin-mediated T cell adhesion to VCAM-1-Fc does not result in costimulation, and that bothMAdCAM-1-Fc and VCAM-1-Fc can “remotely” costi-mulate T cell proliferation clearly demonstrate that celladhesion per se, even in the presence of TCR stimula-tion, is insufficient to generate a costimulatory signal.
Our study demonstrates significant differences in thecostimulatory potencies of integrin ligands and B7molecules. The observed differences are largely consis-tent with results published by other investigators, whereboth B7 molecules have been shown weakly to costimul-ate the proliferation of naive T cells, resulting in interme-diate levels of thymidine incorporation by mixed cellpopulations consisting of both naive and memory T cells[28]. In contrast, both ICAM-1 and VCAM-1 have beenshown to induce proliferation of both T cell subsets [28,32]. In this study, costimulation by ICAM-1-Fc inducedthe strongest proliferative response, although only at rel-atively high concentrations. It appears that VCAM-1 caninduce T cell costimulation more efficiently thanMAdCAM-1-Fc. This is surprising, as their counterrecep-tors VLA-4 and LPAM-1 are expressed by similar subsetsof peripheral T cells [40]. At present, the reasons for thehigh potency of VCAM-1 in T cell proliferation remainunclear.
In conclusion, we have established MAdCAM-1 as anovel costimulatory molecule that mediates T cellcostimulation through interaction with integrin § 4 g 7, andwe present evidence that multiple costimulatory mole-cules can synergize to provide more efficient T cell stim-ulation. However, the successful interaction of costimu-latory molecules did not always result in effective cellactivation, suggesting the existence of exquisite recog-nition, and finely tuned cellular regulatory mechanismsthat discriminate between adhesive events and therebyregulate cell activation.
4 Materials and methods
4.1 Cell lines and antibodies
CTLL-2 cells [41], obtained from the American Type CultureCollection (ATCC, Rockville, MD), were maintained in RPMI1640 supplemented with 10 % (vol:vol) FCS (Gibco BRL,Auckland, New Zealand), 2 mM L-glutamine, 50 U/ml penicil-lin, 50 ? g/ml streptomycin, 100 ? g/ml neomycin, and 2 U/mlrat IL-2 (Sigma, Castle Hill, Australia).
Murine hybridoma secreting mAb against CD3 4 (OKT3 [42]),CD4 (OKT4 [42]), CD11b (OKM1 [42] and MY904 [43]), CD14(63D3 [44]), HLA-DR (IVA12 [45]) and c-myc (9E10 [46]) werefrom the ATCC, as were the rat hybridoma cell lines PS/2
(anti-murine § 4 integrin subunit [47]), M17/5.2 (anti-murineCD11a [48]) and Mel-14 (anti-murine CD62L [49]). The rathybridoma cell lines Fib21 [15] and MECA-367 [14] secreteantibody against the murine g 7 integrin subunit und murineMAdCAM-1, respectively, and were kindly provided by Dr. E.Butcher (Stanford University, Stanford, CA). The rat hybrid-oma M301 (anti-murine g 7 integrin subunit [50]) was donatedby Dr. P. J. Kilshaw (AFRC Institute of Animal Physiology andGenetics Research, Cambridge, GB). Monoclonal anti-bodies were purified on protein G-Sepharose (Sigma). ThemAb Act-1 (anti-integrin § 4 g 7 [51]) was provided by Dr. A.Lazarovits (University of Western Ontario, London, Canada).The mAb 4B4 (anti-integrin g 1 subunit [52]) was purchasedfrom Coulter Electronics (Sydney, Australia), and the mAbagainst human VCAM-1 (1G11 [53]) was obtained fromImmunotech (Haem Pty Ltd., Camberwell, Australia). Puri-fied human IgG1 was from Sigma.
4.2 Cell isolation
PBMC from adult healthy donors were isolated by Ficoll-Paque (AMRAD Pharmacia Biotech, Auckland, NZ) densitygradient centrifugation, resuspended at 1 × 106/ml in RPMI1640 supplemented with 10 % FCS and 2 mM L-glutamine,and incubated for 1 h at 37 °C. CD4+ T lymphocytes wereisolated by negative immunoaffinity chromatography (Iso-CellTM CD4; Pierce, Auckland, NZ). For maximal depletion ofmonocytes and activated T cells, the selection mixture wassupplemented with 50 ? g each of mAb OKM1 and MY904,and 10 ? g of mAb IVA12. The isolated cells were resus-pended at 5 × 105 cells/ml in RPMI 1640 supplemented with10 % FCS, 2 mM L-glutamine, 50 U/ml penicillin, 50 ? g/mlstreptomycin and 100 ? g/ml neomycin. The resulting cellpopulations were typically G 96 % CD3+ CD4+, containedless than 0.1 % CD14+ cells (as determined by direct immu-nofluorescence) and did not respond to optimal concentra-tions (1:200 dilution [54]) of PHA (Gibco BRL) or to solubleOKT3 (200 ng/ml).
4.3 Peptides and Fc fusion proteins
Two peptides derived from the CS-1 domain of the type III-connecting segment of human fibronectin were synthesizedby Chiron Mimotope Peptide Systems (Clayton, Australia).The CS-1 peptide (H2N-CGGGYDELPQLVTLPHPNLHG-PEILDVPST-COOH) contains an EILDVPST motif that hasbeen shown to mediate binding of fibronectin to VLA-4 andLPAM-1 [34]. In peptide CS-1scr (H2N-CGGGYDELPQLV-TLPHPNLHGPPVTSELID-COOH) the EILDVPST motif hasbeen scrambled to prevent integrin binding [34]. cDNAencoding the extracellular regions of VCAM-1 ([55]; cDNApurchased from R&D Systems, Abingdon, GB), B7-2 ([56];cDNA kindly provided by Dr. G. Freeman, Dana Farber Can-cer Institute, Boston, MA), ICAM-1 ([57]; cDNA provided byDr. J. Ni, Human Genome Sciences Inc., Rockville, MD), andmurine B7-1 ([58], cDNA donated by Dr. P. Linsley, Bristol-
Eur. J. Immunol. 1998. 28: 3605–3615 T cell costimulation by § 4-integrin ligands and B7-molecules 3611

Myers-Squibb, Seattle, WA) fused to the hinge, CH2 andCH3 domains of IgG1 were constructed as described previ-ously for the murine MAdCAM-1-Fc chimera [59], insertedinto the pEE14 expression vector ([60]; kindly provided byDr. C. Bebbington, Celltech Therapeutics Ltd, Berkshire, GB)and transfected into CHO-K1 cells (ATCC). Cells secretingfusion proteins were selected in in culture medium supple-mented with L-methionine sulfoximine as described [61] andcloned twice. The Fc fusion proteins were purified on immo-bilized protein A (PerSeptive Biosystems, Cambridge, MA).All fusion proteins were secreted into the supernatant as sol-uble, N-glycosylated, disulfide-linked dimers (not shown).
4.4 Proliferation assays
Flat-bottom 96-well microtiter plates were coated overnightat 4 °C with 30 ng/well of OKT3 in 50 ? l PBS. After washingtwice with cold PBS, recombinant cell adhesion moleculesor human IgG1 were absorbed for 20 h at 4 °C. Wells wereblocked with 10 % FCS in PBS for 8 h at 4 °C. Alternatively,the recombinant Fc-chimeric proteins were immobilized inwells that had been precoated with 2 ? g of rabbit anti-human Fc antibody (Dako, MED-BIO Enterprises, Christ-church, NZ). For immobilization of fibronectin-derived syn-thetic peptides, human IgG1 was derivatized with N-succinimidyl-3(2-pyridyldithio) propionate (SPDP, Pierce) ata molar ratio of 1:20 (IgG:SPDP) in 100 mM NaCl/50 mMsodium phosphate, pH 7.5. Briefly, microtiter wells, pre-coated with 30 ng OKT3, were coated overnight at 4 °C with1 ? g SPDP-activated human IgG1 in 50 ? l PBS. UnboundIgG was removed, and fibronectin-derived peptides(0–1500 ng in 50 ? l PBS) were incubated overnight at roomtemperature before wells were washed and blocked with10 % FCS. In antibody blocking experiments, cells wereincubated on ice for 30 min in 200 ? l culture medium con-taining 10 ? g/ml mAb (30 ? g/ml for PS/2) before stimulation.Immobilized cell adhesion molecules were incubated with10 ? g/ml mAb for 60 min at 37 °C. Polystyrene microbeads(1.5 × 107; diameter 2.5 ? m; Bangs Laboratories, Fishers, IN)were coated overnight at 4 °C with 20 ? g MAdCAM-1-Fc,VCAM-1-Fc, B7-2-Fc or human IgG1 in 100 ? l, or with 3 ? gOKT3 mAb (total volume 50 ? l), washed, and blocked with10 % FCS for 4 h at 37 °C. Triplicate cultures of 5 × 104 puri-fied CD4+ T cells per well were stimulated for 60 h at 37 °C ina humidified 5 % CO2/95 % air atmosphere and pulsed with0.25 ? Ci (9.25 kBq) of [3H]thymidine (35 Ci/mmol; ICN, Irvine,CA) 14 h before harvesting the cells onto glass fiber filters.Radiolabel incorporation into newly synthesized DNA wasmeasured by liquid scintillation counting and is expressedas arithmetic mean counts per minute ± standard deviation.
4.5 Analysis of IL-2 expression
Cell-free culture supernatants were collected after 20 h ofstimulation and assayed for IL-2 content using IL-2-dependent CTLL-2 cells [62]. The IL-2 concentration in the
samples was estimated by reference to a standard curveobtained with rIL-2 (National Institute for Biological Stan-dards and Control, Hertfordshire, GB).
Acknowledgments: This work was supported in part bygrants from the Cancer Society of New Zealand, the CancerSociety (Wellington Division), the Health Research Council ofNew Zealand, the Wellcome Trust, the Lottery Grants Boardof New Zealand, the Marsden Fund, the Auckland MedicalResearch Foundation, and the Royal Society of New Zea-land. G. W. Krissansen is a former recipient of a WellcomeSenior Research Fellowship in Medical Science in New Zea-land, and more recently of a James Cook Research Fellow-ship from the Royal Society of New Zealand.
Addendum: After the first submission of this manuscript,the expression of MAdCAM-1 by mucosal follicular dendriticcells has been reported (M.C. Szabo et al., J. Immunol.1997. 158: 5584–5588).
5 References
1 Weiss, A. and Littman, D. R., Signal transduction bylymphocyte antigen receptors. Cell 1994. 76: 263–274.
2 Wange, R. L. and Samelson, L. E., Complex com-plexes: signaling at the TCR. Immunity 1996. 5:197–205.
3 Harding, F. A., McArthur, J. G., Gross, J. A., Raulet, D.H. and Allison, J. P., CD28-mediated signalling co-stimulates murine T cells and prevents induction ofanergy in T-cell clones. Nature 1992. 356: 607–609.
4 Schwartz, R. H., A cell culture model for T lymphocyteanergy. Science 1990. 248: 1349–1356.
5 Clark, E. A. and Ledbetter, J. A., How B and T cells talkto each other. Nature 1994. 367: 425–428.
6 Van Seventer, G. A., Shimizu, Y. and Shaw, S., Roles ofmultiple accessory molecules in T cell activation. Curr.Opin. Immunol. 1991. 3: 294–303.
7 Hynes, R. O., Integrins: versatility, modulation, and sig-nalling in cell adhesion. Cell 1992. 69: 11–25.
8 Butcher, E. C. and Picker, L. J., Lymphocyte homingand homeostasis. Science 1996. 272: 60–66.
9 Sonnenberg, A., Integrins and their ligands. Curr. Top.Microbiol. Immunol. 1993. 184: 7–35.
10 Van Seventer, G. A., Shimizu, Y., Horgan, K. J. andShaw, S., The LFA-1 ligand ICAM-1 provides an impor-tant costimulatory signal for T cell receptor-mediatedactivation of resting T cells. J. Immunol. 1990. 144:4579–4586.
3612 K. Lehnert et al. Eur. J. Immunol. 1998. 28: 3605–3615

11 Nojima, Y., Humphries, M. J., Mould, A. P., Komoriya,A., Yamada, K. M., Schlossman, S. F. and Morimoto,C., VLA-4 mediates CD3-dependent CD4+ T cell activa-tion cia the CS-1 alternatively spliced domain of fibro-nectin. J. Exp. Med. 1990. 172: 1185–1192.
12 Shimizu, Y., van Seventer, G. A. and Shaw, S., Costim-ulation of proliferative responses of resting CD4+ T cellsby the interaction of VLA-4 and VLA-5 with fibronectin orVLA-6 with laminin. J. Immunol. 1990. 145: 59–67.
13 Erle, D. J., Briskin, M. J., Butcher, E. C., Garcia Pardo,A., Lazarovits, A. I. and Tidswell, M., Expression andfunction of the MAdCAM-1 receptor, integrin §
4g 7 on
human leukocytes. J. Immunol. 1994. 153: 517–528.
14 Berlin, C., Berg, E. L., Briskin, M. J., Andrew, D. P., Kil-shaw, P. J., Holzmann, B., Weissman, I. L., Hamann,A. and Butcher, E. C., § 4 g 7 integrin mediates lympho-cyte binding to the mucosal vascular addressinMAdCAM-1. Cell 1993. 74: 185–195.
15 Andrew, D. P., Berlin, C., Honda, S., Yoshino, T.,Hamann, A., Holzmann, B., Kilshaw, P. J. andButcher, E. C., Distinct but overlapping epitopes areinvolved in §
4g 7-mediated adhesion to vascular cell
adhesion molecule-1, mucosal addressin-1, fibronectin,and lymphocyte aggregation. J. Immunol. 1994. 153:3847–3861.
16 Briskin, M. J., McEvoy, L. M. and Butcher, E. C.,MAdCAM-1 has homology to immunoglobulin andmucin-like adhesion receptors and to IgA1. Nature 1993.363: 461–463.
17 Leung, E., Greene, J., Ni, J., Raymond, L. G., Lehnert,K., Langley, R. and Krissansen, G. W., Cloning of themucosal addressin MAdCAM-1 from human brain. Iden-tification of novel alternatively spliced transcripts. Immu-nol. Cell Biol. 1996. 74: 490–496.
18 Shyjan, A. M., Bertagnolli, M., Kenney, C. J. and Bris-kin, M. J. Human mucosal addressin cell adhesionmolecule-1 (MAdCAM-1) demonstrates structural andfunctional similarities to the §
4g 7-integrin binding
domains of murine MAdCAM-1, but extreme divergenceof mucin-like sequences. J. Immunol. 1996. 156:2851–2857.
19 Hanninen, A., Taylor, C., Streeter, P. R., Stark, L. S.,Sarte, J. M., Shizuru, J. A., Simell, O. and Michie, S.A., Vascular addressins are induced on islet vessels dur-ing insulitis in nonobese diabetic mice, and are involvedin lymphoid cell binding to islet endothelium. J. Clin.Invest. 1993. 92: 2509–2515.
20 O’Neill, J. K., Butter, C., Baker, D., Gschmeisser, S. E.,Kraal, G., Butcher, E. C. and Turk, J. L., Expression ofvascular addressins and ICAM-1 by endothelial cells inthe spinal cord during chronic relapsing experimentalallergic encephalomyelitis in the Biozzi AB/H mouse.Immunology 1991. 72: 520–525.
21 Postigo, A. A., Sanchez Mateos, P., Lazarovits, A. I.,Sanchez-Madrid, F. and de Landazuri, M. O., § 4 g 7integrin mediates B cell binding to fibronectin and vas-cular cell adhesion molecule-1. Expression and functionof § 4 integrins on human B lymphocytes. J. Immunol.1993. 151: 2471–2483.
22 Ruegg, C., Postigo, A. A., Sikorski, E. E., Butcher, E. C.,Pytela, R. and Erle, D. J., Role of integrin § 4 g 7/ § 4 g p in lym-phocyte adherence to fibronectin and VCAM-1 and inhomotypic cell clustering. J. Cell Biol. 1992. 117: 179–189.
23 Guan, J. L. and Hynes, R. O., Lymphoid cells recognizean alternatively spliced segment of fibronectin via theintegrin receptor § 4 g 1. Cell 1990. 60: 53–61.
24 Elices, M. J., Osborn, L., Takada, Y., Crouse, C.,Luhowskyj, S., Hemler, M. E. and Lobb, R. R., VCAM-1on activated endothelium interacts with the leukocyteintegrin VLA-4 at a site distant from the VLA-4/fibronec-tin binding site. Cell 1990. 60: 577–548.
25 Yang, Y., Harrison, J. E. B., Print, C. G., Lehnert, K.,Sammar, M., Lazarovits, A. I. and Krissansen, G. W.,Interaction of monocytoid cells with the mucosaladdressin MAdCAM-1 via the integrins VLA-4 andLPAM-1. Immunol. Cell Biol. 1996. 74: 383–393.
26 Damle, N. K. and Aruffo, A., Vascular cell adhesionmolecule-1 induces T-cell antigen receptor-dependentactivation of CD4+ T lymphocytes. Proc. Natl. Acad. Sci.USA 1991. 88: 6403–6407.
27 Burkly, L. C., Jakubowski, A., Newman, B. M., Rosa,M. D., Chi Rosso, G. and Lobb, R. R., Signaling by vas-cular cell adhesion molecule-1 (VCAM-1) through VLA-4promotes CD3-dependent T cell proliferation. Eur. J.Immunol. 1991. 21: 2871–2875.
28 Damle, N. K., Klussman, K., Linsley, P. S. and Aruffo,A., Differential costimulatory effects of adhesion mole-cules B7, ICAM-1, LFA-3, and VCAM-1 on resting andantigen-primed CD4+ T lymphocytes. J. Immunol. 1992.148: 1985–1992.
29 Teague, T. K., Lazarovits, A. I. and McIntyre, B. W.,Integrin § 4 g 7 co-stimulation of human peripheral blood Tcell proliferation. Cell Adhes. Commun. 1994. 2:539–547.
30 Bobryshev, Y. V., Lord, R. S., Rainer, S. P. and Munro,V. F., VCAM-1 expression and network of VCAM-1 posi-tive vascular dendritic cells in advanced atheroscleroticlesions of carotid arteries and aortas. Acta Histochem.1996. 98: 185–194.
31 Hughes, C. C. W., Savage, C. O. S. and Pober, J. S.,The endothelial cell as a regulator of T-cell function.Immunol. Rev. 1990. 117: 85–102.
Eur. J. Immunol. 1998. 28: 3605–3615 T cell costimulation by § 4-integrin ligands and B7-molecules 3613

32 Damle, N. K., Klussman, K., Linsley, P. S., Aruffo, A.and Ledbetter, J. A., Differential regulatory effects ofintercellular adhesion molecule-1 on costimulation bythe CD28 counter-receptor B7. J. Immunol. 1992. 149:2541–2548.
33 Damle, N. K., Klussman, K., Leytze, G., Myrdal, S.,Aruffo, A., Ledbetter, J. A. and Linsley, P. S., Costimu-lation of T lymphocytes with integrin ligands intercellularadhesion molecule-1 or vascular cell adhesionmolecule-1 induces functional expression of CTLA-4, asecond receptor for B7. J. Immunol. 1994. 152:2686–2697.
34 Linsley, P. S., Greene, J. L., Tan, P., Bradshaw, J., Led-better, J. A., Anasetti, C. and Damle, N. K., Coexpres-sion and functional cooperation of CTLA-4 and CD28 onactivated T lymphocytes. J. Exp. Med. 1992. 176:1595–1604.
35 Krummel, M. F. and Allison, J. P., CD28 and CTLA-4have opposing effects on the response of T cells to stim-ulation. J. Exp. Med. 1995. 182: 459–465.
36 Krummel, M. F. and Allison, J. P., CTLA-4 engagementinhibits IL-2 accumulation and cell cycle progressionupon activation of resting T cells. J. Exp. Med. 1996.183: 2533–2540.
37 Bluestone, J., Is CTLA-4 a master switch for peripheralT cell tolerance. J. Immunol. 1997. 158: 1989–1993.
38 Udagawa, T., Woodside, D. G. and McIntyre, B. W.,§4g 1 (CD94d/CD29) integrin costimulation of human T
cells enhances transcription factor and cytokine induc-tion in the absence of altered sensitivity to anti-CD3stimulation. J. Immunol. 1996. 157: 1965–1972.
39 Shimizu, Y., van Seventer, G. A., Horgan, K. J. andShaw, S., Roles of adhesion molecules in T-cell recogni-tion. Fundamental similarities between four integrins onresting human T cells (LFA-1, VLA-4, VLA-5, VLA-6) inexpression, binding and costimulation. Immunol. Rev.1990. 114: 109–143.
40 Rott, L. S., Briskin, M. J., Andrew, D. P., Berg, E. L.and Butcher, E. C., A fundamental subdivision of circu-lating lymphocytes defined by adhesion to mucosaladdressin cell adhesion molecule-1. J. Immunol. 1996.156: 3727–3736.
41 Gillis, S. and Smith, K. A., Long term culture of tumor-specific cytotoxic T cells. Nature 1977. 268: 154–156.
42 Hoffman, R. A., Kung, P. C., Hansen, W. P. and Gold-stein, G., Simple and rapid measurement of human Tlymphocytes and their subclasses in peripheral blood.Proc. Natl. Acad. Sci. USA 1990. 77: 4919–4917.
43 Dana, N., Styrt, B., Griffin, J.D., Todd, R. F., IIIrd,Klempner, M. S. and Arnaout, M. A., Two functional
domains in the phagocyte membrane glycoprotein Mo1identified with monoclonal antibodies. J. Immunol. 1986.137: 3259–3263.
44 Ugolini, V., Nunez, G., Smith, R. G., Stasny, P. andCapra, J. D. Initial characterization of monoclonal anti-bodies against human monocytes. Proc. Natl. Acad. Sci.USA 1980. 77: 6764–6768.
45 Shaw, S., Ziegler, A. and DeMars, R., Specificity ofmonoclonal antibodies directed against human andmurine class II histocompatibility antigens as analyzedby binding to HLA-deletion mutant cell lines. Hum.Immunol. 1985. 12: 191–211.
46 Evans, G. I., Lewis, G. K., Ramsay, G. and Bishop, J.M., Isolation of monoclonal antibodies specific forhuman c-myc proto-oncogene product. Mol. Cell. Biol.1985. 5: 3610–3616.
47 Miyake, K., Medina, K., Ishihara, K., Kimoto, M.,Auerbach, R. and Kincade, P. W., A VCAM-like adhe-sion molecule in murine bone-marrow stromal cellsmediates binding of lymphocyte precursors in culture. J.Cell Biol. 1991. 114: 557–565.
48 Sanchez-Madrid, F., Davignon, D., Martz, E. andSpringer, T. A., Antigens involved in mouse cytolytic T-lymphocyte (CTL)-mediated killing. Functional screeningand topographic relationship. Cell. Immunol. 1982. 73:1–11.
49 Gallatin, W. M., Weissman, I. L. and Butcher, E. C., Acell-surface molecule involved in organ-specific homingof lymphocytes. Nature 1983. 304: 30–34.
50 Kilshaw, P. J. and Murant, S. J., Expression and regula-tion of § 7( g p) integrins on mouse lymphocytes: relevanceto the mucosal immune system. Eur. J. Immunol. 1991.21: 2591–2597.
51 Lazarovits, A. I., Moscicki, R. A., Kurnick, J. T., Came-rini, D., Bhan, A. K., Baird, L. G., Eriksson, M. and Col-vin, R. B., Lymphocyte activation antigens. I. A mono-clonal antibody, anti-Act-1, defines a new late lympho-cyte activation antigen. J. Immunol. 1984. 133: 1875.
52 Yamada, A., Najima, Y., Sugita, K., Dang, N. H.,Schlossman, S. F. and Morimoto, C., Cross-linking ofVLA/CD29 molecule has a co-mitogenic effect with anti-CD3 on CD4 cell activation in serum-free culture system.Eur. J. Immunol. 1991. 21: 319–325.
53 Thornhill, M., Wellcome, S. M., Mahiouz, D. L.,Lanchbury, J. S. S., Kyan-Aung, U. and Haskard, D.O., TNF combines with IL-4 or IFN- + to selectivelyenhance endothelial cell adhesiveness for T cells. Thecontribution of VCAM-1 dependent and independentbinding mechanisms. J. Immunol. 1991. 146: 592–598.
3614 K. Lehnert et al. Eur. J. Immunol. 1998. 28: 3605–3615

54 Davies, L. and Lipsky, P., Signals involved in T cell acti-vation. J. Immunol. 1986. 136: 3588–3596.
55 Osborn, L., Hession, C., Tizard, R., Vassallo, C.,Luhowskyi, S., Chi-Rosso, G. and Lobb, R., Directexpression cloning of vascular cell adhesion molecule-1,a cytokine-induced endothelial protein that binds to lym-phocytes. Cell 1989. 59: 1203–1211.
56 Freeman, G. J., Gribben, J. G., Boussiotis, V. A., Ng, J.W., Restivo, V. A., Jr., Lombard, L. A., Gray, G. S. andNadler, L. M., Cloning of B7-2: A CTLA-4 counter recep-tor that costimulates human T cell proliferation. Science1993. 262: 909–911.
57 Staunton, D. E., Marlin, S. E., Stratowa, C., Dustin, M.L. and Springer, T. A., Primary structure of ICAM-1demonstrates interaction between members of theimmunoglobulin and integrin supergene families. Cell1988. 52: 925–933.
58 Freeman, G. J., Gray, G. S., Gimmi, C. D., Lombard, D.B., Zhou, L.-J., White, M., Fingeroth, J. D., Gribben, J.G. and Nadler, L. M., Structure, expression, and T cellcostimulatory activity of the murine homologue of thehuman B lymphocyte activation antigen B7. J. Exp. Med.1991. 174: 625–631.
59 Yang, Y., Sammar, M., Harrison, J. E. B., Lehnert, K.,Print, C. G., Leung, E., Prestidge, R. and Krissansen,G. W., Construction and adhesive properties of solubleMAdCAM-1-Fc chimera expressed in a baculovirus sys-tem. Phylogenetic conservation of receptor-ligand inter-action. Scand. J. Immunol. 1995. 42: 235–247.
60 Bebbington, C. R. and Hentschel, C. C. G., The use ofvectors based on gene amplification of cloned genes inmammalian cells. In Glover, D. (Ed) DNA cloning, Vol. III.Academic Press, San Diego 1987, pp 163–188.
61 Kingston, R. E., Kaufman, R. J., Bebbington, C. R.and Rolfe, M. R., Amplification using glutamine synthe-tase. In Ausubel, F. A., Brent, R., Kingston, R. E.,Moore, D. D., Seidman, J. G., Smith, J. A. and Struhl,K. (Eds) Current Protocols in Molecular Biology. JohnWiley & Sons, New York 1992, pp 16.14.7–16.14.13.
62 Davis, L. S. and Lipsky, P. E., Measurement of humanand murine interleukin 2 and interleukin 4. In Coligan J.E., Kruisbeek, A. M., Marguiles, D. H., Shevach, E. M.and Strober, W. (Eds) Current Protocols in Immunology.John Wiley & Sons, New York 1995, pp 6.3.1–6.3.13.
Correspondence: Geoffrey W. Krissansen, University ofAuckland, School of Medicine, Park Rd., Grafton, Auckland,New ZealandFax: +64-9-373-7492e-mail: gw.krissansen — auckland.ac.nz.
C. G. Print’s present address: The Walter and Eliza HallInstitute, Royal Melbourne Hospital, Melbourne, Australia
Eur. J. Immunol. 1998. 28: 3605–3615 T cell costimulation by § 4-integrin ligands and B7-molecules 3615















